 Jorge García-Márquez1†
Jorge García-Márquez1† Marta Domínguez-Maqueda1†
Marta Domínguez-Maqueda1† Inmaculada Llamas2,3
Inmaculada Llamas2,3 Silvana Teresa Tapia-Paniagua1
Silvana Teresa Tapia-Paniagua1 Salvador Arijo1
Salvador Arijo1 Miguel Ángel Moriñigo1*
Miguel Ángel Moriñigo1* María Carmen Balebona1
María Carmen Balebona1- 1Department of Microbiology, Faculty of Science, Instituto Andaluz de Biotecnología y Desarrollo Azul, University of Málaga, Málaga, Spain
- 2Department of Microbiology, Faculty of Pharmacy, University of Granada, Granada, Spain
- 3Institute of Biotechnology, Biomedical Research Center (CIBM), University of Granada, Granada, Spain
The consequences of the overuse of antibiotics in aquaculture have intensified the search for alternative pathogen control strategies, including postbiotics and antimicrobial proteins. In this study, we evaluated the potential of extracellular products (ECPs) from Bacillus pumilus UMA 169 and UMA 216, cultivated in media supplemented with microalgal and cyanobacterial biomass, to inhibit biofilm formation, engage in quorum quenching (QQ), and modulate virulence in key aquaculture pathogens. Our findings show that most of the tested ECPs, with the exception of MICRO 216, significantly inhibited biofilm formation by Aeromonas hydrophila and Vibrio anguillarum. However, all ECPs downregulated the expression of aip56, a key virulence gene in Photobacterium damselae subsp. piscicida. They also modulated the transcription of iron acquisition genes (hutB, hutD, irp1, irp2), suggesting a broader impact on bacterial virulence. These findings highlight the potential of B. pumilus-derived ECPs as both biofilm inhibitors and virulence modulators for aquaculture pathogens. Further studies should explore their application as postbiotic agents in pathogen control strategies aiming to reduce antibiotic overuse and improve fish health in aquaculture systems.
1 Introduction
The aquaculture industry is a vital source of protein, supplying nearly half of the world’s fish products (FAO, 2022). However, its rapid expansion has led to an increased prevalence of bacterial diseases (Austin, 2019). The pathogenicity of these bacteria is closely linked to virulence factors and strategies such as biofilm formation, which can be regulated by quorum sensing (QS) mechanisms (Priya et al., 2023; Li et al., 2025). Biofilms facilitate bacterial adhesion to surfaces and provide protection against host immune and antimicrobial agents, contributing to persistent infections (Ramírez-Larrota and Eckhard, 2022; Rahman et al., 2024). QS is a bacterial cell-to-cell communication process that enables bacteria to coordinate gene expression based on population density (Juszczuk-Kubiak, 2024). By using signal molecules like N-acyl homoserine lactones (AHLs) in Gram-negative bacteria (Porras et al., 2025), bacteria can collectively activate pathways that lead to increased virulence, including the production of toxins, hydrolytic enzyme production, the expression of virulence-associated genes, and the formation of biofilms (Whiteley et al., 2017; Gonzales et al., 2024). Furthermore, it has been reported that pathogens utilize QS systems to evade early detection by host defenses (Ganesh et al., 2020). Thus, given the role of QS in regulating virulence, disrupting bacterial communication by interfering with QS (a process known as quorum quenching, QQ) is a potential approach for infection treatment or prevention (Torres et al., 2016; Ghanei-Motlagh et al., 2020).
The reliance on conventional antimicrobials in aquaculture has accelerated the global rise of antimicrobial resistance (Pereira et al., 2022), making the development of sustainable alternatives, including probiotics, a top priority for achieving a sustainable and environmentally friendly aquaculture industry (Yousuf et al., 2023). Probiotics are defined as live microorganisms that, when administered in adequate amounts, confer health benefits to the host (Hill et al., 2014). In a previous study, we identified two Bacillus pumilus isolates (UMA 169 and UMA 216) from the intestinal tract of healthy seabream (Sparus aurata) fed with a diet partially supplemented with microalgae, exhibiting potential probiotic properties. These isolates exhibited antagonistic activity against fish pathogens, including Tenacibaculum maritimum, Photobacterium damselae subsp. piscicida, and Vibrio anguillarum (García-Márquez et al., 2023). Although Bacillus species are well-known for their probiotic effects in aquaculture (Yi et al., 2018; Santos et al., 2021a; Thakur et al., 2025), and authors such as Guo et al. (2025) have shown benefits in fish fed with diets containing a B. pumilus strain, their potential as a source of postbiotics remains largely underexplored (Lu et al., 2022; Guo et al., 2025).
Postbiotics are non-viable bacterial products or metabolic by-products that exert beneficial biological effects on the host (Aguilar-Toalá et al., 2018). They include a diverse range of compounds such as bacteriocins, organic acids, enzymes, peptides with antimicrobial and immunomodulatory properties, and extracellular products (ECPs) (Yan et al., 2024). Compared to probiotics, postbiotics offer advantages such as greater stability, a well-defined composition, involvement in more specific mechanisms of action, and a reduced risk of infection (Barros et al., 2020; Cerezo et al., 2025). ECPs, in particular, have shown great promise in interfering with bacterial biofilms and other virulence mechanisms, making them promising candidates for disease control in aquaculture (Moradi et al., 2021; Domínguez-Maqueda et al., 2024a).
The composition of the culture medium, and therefore the carbon source, is a critical factor influencing the bioactivity and metabolic profile of bacterial products (Pokorzynski and Groisman, 2023). Microalgae and cyanobacteria are gaining attention as sustainable, nutrient-rich alternatives to conventional bacterial growth media (Ma and Hu, 2023; Ricós-Muñoz et al., 2023). Their biomass contains bioactive compounds that can modulate bacterial metabolism and enhance the production of antimicrobial ECPs (Borges Lopes et al., 2025). However, the specific relationship between microalgae-supplemented media and the bioactivity of Bacillus-derived postbiotics remains poorly understood, highlighting the need for further investigation.
Based on our previous work, in which ECPs from two B. pumilus isolates (UMA 169 and UMA 216) were produced using various microalgae and cyanobacteria (García-Márquez et al., 2025), this study evaluates the potential of select ECP preparations as novel anti-virulence agents. Among the ECPs obtained from B. pumilus cultured on different microalgal- and cyanobacterial-supplemented media, five conditions stood out for their hydrolytic activity and antagonism against several fish pathogens (García-Márquez et al., 2025): MICRO 169 (ECPs from B. pumilus UMA 169 grown with 5% Microchloropsis gaditana); CHL 216, MICRO 216, and ATH 216 (ECPs from B. pumilus UMA 216 grown with 5% Chlorella vulgaris, 5% M. gaditana, and 5% Arthrospira platensis, respectively) and TSA 216 (ECPs from B. pumilus UMA 216 cultured on a standard tryptone soy agar medium).
We hypothesize that these ECPs will effectively inhibit key virulence factors in certain important bacterial fish pathogens. Specifically, we assessed their antibiofilm and quorum quenching activities and their effect on the in vitro transcription of the gene encoding the AIP56 toxin in P. damselae subsp. piscicida Lg41/01 (Phdp-Lg41/01).
Briefly, the purpose of this study is to thoroughly evaluate the anti-virulence potential of these five selected ECP preparations derived from B. pumilus under different culturing conditions. By focusing on postbiotics, this research explores sustainable tools for disease control in aquaculture by exploring alternatives to conventional antibiotics and addressing the antimicrobial resistance crisis. In addition, this work delves into the potential of Bacillus as a source of postbiotics, a field that remains largely unexplored in this bacterial species. This study helps bridge the gap in understanding the specific relationship between microalgae-supplemented media and the bioactivity of Bacillus-derived postbiotics. Finally, the study utilizes ECPs produced using microalgae and cyanobacteria as nutrient-rich alternatives to conventional growth media. This is relevant as their biomass contains bioactive compounds that can modulate bacterial metabolism and enhance the production of antimicrobial ECPs.
2 Materials and methods
2.1 Bacterial strains and culture conditions
B. pumilus strains UMA 169 and UMA 216 were isolated from the gastrointestinal tract of healthy gilthead seabream specimens (García-Márquez et al., 2023). Both isolates were cultured on tryptic soy agar (Oxoid Ltd.) supplemented with 1.5% (w/v) NaCl (TSAs) at 23°C for 24 h. Then, one to two colonies were transferred to 50 mL-flasks containing tryptic soy broth (Oxoid Ltd) supplemented with 1.5% (w/v) NaCl (TSBs) and incubated at 23°C for 36 h (until 109 colony-forming units (CFU) mL-1, early stationary phase) on shaking at 80 rpm.
Agrobacterium tumefaciens NTL4 (pZLR4) (Shaw et al., 1997) was cultured at 30°C in Luria Bertani (LB) medium (10 g tryptone, 5 g yeast extract and 10 g NaCl per liter of distilled water) supplemented with 2.5 mmol L-1 CaCl2 x 2H2O and 2.5 mmol L-1 MgSO4 × 7 H2O (LB/MC) containing 50 mg gentamycin mL-1 and in AB medium (Chilton et al., 1974). A. tumefaciens NTL4 allows the detection of medium- to long-chain HSLs (C6–C12).
Strains Chromobacterium subtsugae CV026 (Harrison and Soby, 2020), formerly C. violaceum CV026 (McClean et al., 1997) and C. violaceum VIR07 (Morohoshi et al., 2008) were used to detect the quorum quenching mechanism. These strains detect the presence of AHL in the medium but are unable to produce it. The strains were grown at 30°C in LB medium supplemented with kanamycin (50 mg mL-1). C. violaceum CV026 and VIR07 were used for the detection of short-chain HSLs (C4–C8) and HSLs with an oxo group, respectively.
Phdp-Lg 41/01 strain, originally isolated from cultured diseased Senegalese sole (Díaz-Rosales et al., 2003), was cultured on TSAs plates at 23°C for 48 h. Then, one or two colonies were transferred to TSBs 10 mL tubes and incubated at 23°C for 18 h, under shaking conditions (80 rpm) until reaching mid-exponential phase (OD600nm = 0.8 ~ 104 CFU mL-1) (Núñez-Díaz et al., 2018). T. maritimum (CECT 4276) was cultured on Flexibacter maritimus agar (FMM) plates (Pazos et al., 1996) at 28°C for 48 h. V. anguillarum (CECT 522) and A. hydrophila (CECT 5734) were grown on TSAs plates at 23°C for 24 h.
2.2 Preparation of media for ECP production
Microalgal (C. vulgaris and M. gaditana) and cyanobacterial (A. platensis) biomass was kindly provided by LifeBioencapsulation S.L. (Spin-off; Universidad de Almería, Almería, Spain). After cultivation, harvesting, freeze-drying, and milling, the resulting powder was stored at –20°C until use.
ECPs were obtained from B. pumilus UMA 169 and UMA 216, following a modified cellophane plate technique described by Liu (1957). Briefly, 1 mL of UMA 169 bacterial suspension obtained as described above was spread over sterile cellophane sheets placed on solid medium plates (1.5% w/v agar) supplemented with 5% (w/v) M. gaditana (MICRO 169 condition). Additionally, 1 mL of UMA 216 bacterial suspension was spread over sterile cellophane sheets placed on solid medium plates (1.5% w/v agar) supplemented with the following: i) 5% (w/v) C. vulgaris (CHL 216 condition), ii) 5% (w/v) M. gaditana (MICRO 216 condition), iii) 5% (w/v) A. platensis (ATH 216 condition), and iv) TSAs plates (TSA 216 condition) that were used as a control medium. To determine the potential background from the media, cellophane sheets were placed on plates containing all media but without inoculating B. pumilus UMA 169 and UMA 216 isolates, serving as internal controls (ICs), and no microbial growth was observed. Incubation of all plates was carried out at 23°C for 24 h.
After incubation, bacterial cells from the various growth conditions and internal controls were collected in 2 mL sterile phosphate-buffered saline (pH 7.2) and centrifuged at 10,000 × g for 20 min at 4°C. The supernatants were filtered sequentially first through 0.45 and then 0.2 µm pore-size membrane filters (Merck Millipore, USA) to obtain the ECPs without cells. The protein concentration was determined using the Qubit Protein assay kits and the Qubit 2.0 fluorometer (Thermo Fisher Scientific, USA). Aliquots of the ECP samples were cultured on TSAs plates and incubated for 24 h at 23°C to confirm the absence of growth. The ECPs were stored at –80°C until further use.
2.3 Biofilm inhibition assay
The ability of ECPs to inhibit biofilm formation was assessed using a 96-well microtiter plate crystal violet assay (CV) described by Vivas et al. (2008), and optimized by Domínguez-Maqueda et al. (2024a). Overnight bacterial cultures of T. maritimum, A. hydrophila, and V. anguillarum species were selected based on the important role of biofilm formation in their virulence, as reported for strains of these species (Croxatto et al., 2007; Rasmussen-Ivey et al., 2016; Mabrok et al., 2023). Bacterial suspensions were adjusted to OD595nm ~ 0.1, and 20 µL were transferred into 96-well plates, followed by the addition of the pathogenic bacterial suspensions alone (to measure growth performance), with ECP samples or ICs. The biofilm development was assessed after incubation at 28°C for 48 h for T. maritimum and 23°C for 24 h for V. anguillarum and A. hydrophila.
Following incubation, wells were washed three times with sterile phosphate-buffered saline (PBS, pH 7.4) to remove non-adherent cells. Biofilms were then stained with 200 μL of 0.1% crystal violet for 15 min at room temperature. Excess dye was removed by rinsing wells with sterile distilled water, and the plates were air-dried. To quantify biofilm biomass, stained biofilms were solubilized with 200 μL of 95% ethanol, and absorbance was measured at OD595nm using a microplate reader (Multiskan FC, Thermo Fisher). Biofilm inhibition was calculated as the percentage reduction in biofilm formation relative to the control wells. All assays were performed in triplicate, with three independent biological replicates per condition.
2.4 DNase activity
DNase activity in the ECPs was determined using DNase agar test plates (Oxoid, UK). Wells of 6 mm in diameter were created in the agar plates, and 50 μL of each ECP sample (0.5 μg protein μL-¹) was inoculated into the wells. ICs were included in all assays. Plates were incubated at 23°C for 24–48 h, after which 1M HCl was added to flood the plates. The presence of a clear halo around the wells indicated DNase activity, whereas its absence was interpreted as a lack of DNase activity. The lowest ECP concentration that produced a clear zone was recorded as the minimum concentration required for DNase activity. A 50 μL volume of phosphate-buffered saline (PBS, pH 7.4) was used as a negative control. Each ECP condition was tested in triplicate, and experiments were repeated twice.
2.5 Study of quorum quenching activity
Quorum quenching (QQ) activity was assessed by detecting N-acyl homoserine lactone (AHL) degradation, following the method described by Torres et al. (2016). The synthetic AHLs tested included short-chain AHLs: C4-HSL (N-butyryl-DL-homoserine lactone), C6-HSL (N-hexanoyl- DL-homoserine lactone); and long-chain AHLs: C8-HSL (N-octanoyl-DL-homoserine lactone), C10-HSL (N-decanoyl-DL-homoserine lactone), C12- HSL (N-dodecanoyl-DL-homoserine lactone), 3-oxo-C8-HSL (N-3-oxo-octanoyl-DL-homoserine lactone), 3-oxo-C10-HSL (N-3-oxo-decanoyl-DL-homoserine lactone) and 3-oxo-C12-HSL (N-3-oxo-dodecanoyl-DL-homoserine lactone). All AHLs were obtained from Sigma-Aldrich (USA).
B. pumilus isolates UMA 169 and UMA 216 were cultured on TSAs at 23°C for 24 h. One or two colonies were then transferred to 2 mL TSB tubes and incubated at 23°C for 24 h under shaking (120 rpm). Simultaneously and under the same culture conditions, 60–80 µL (30 µg of protein) from different ECP conditions or ICs were added to separate 2 mL tubes of TSB to evaluate the ability of ECPs to degrade synthetic AHLs. After 24 h, a stock solution of synthetic AHLs was added to all the tubes to reach a final concentration of 10 µM, followed by an additional incubation of 24 h at 23°C and shaking at 120 rpm. Control tubes of TSB containing the same concentration of each AHL were incubated under identical conditions.
To detect AHL degradation, overnight cultures of biosensor strains CV026 and VIR07 were diluted 1:10 in 5 mL of LB soft agar (0.7% w/v) and plated onto LB agar plates (for CV026 and VIR07); AB medium supplemented with 80 μg mL-1 of 5-bromo-4-chloro-3-indolyl-ß-D-galactopyranoside (X-gal) for NTL4. Six millimeter-diameter wells were created in the medium using a sterile glass Pasteur pipette, and 100 μL of B. pumilus cell-free supernatant and ECP and IC-treated media were loaded into the wells. TSB tubes containing AHLs without ECPs served as positive controls.
Plates were incubated at 28°C for 24 h to determine the presence of a violet or blue halo (depending on the bioindicator) around the wells, indicating the existence of AHL in the medium. The absence of the halo was considered due to AHL degradation.
2.6 Minimum inhibitory concentration of B. pumilus-ECPs against Photobacterium damselae subsp. piscicida Lg 41/01
MIC of B. pumilus-ECPs against Phdp-Lg 41/01 was determined to assess their potential to inhibit Phdp-Lg 41/01 bacterial growth. Phdp-Lg 41/01 was cultured on TSAs plates at 23°C for 48 h. Bacterial cells were then collected and suspended in 10 mL TSBs tubes to achieve an OD595nm ~ 0.5.
For MIC determination, 20 µL of bacterial suspension was inoculated into flat-bottom polystyrene 96-well microplates (#D51588, Sarstedt, Nümbrecht, Germany) under the following conditions: a positive control (Phdp Control +) with wells filled up to 200 µL with TSBs containing only bacterial suspension (no ECPs). ECP-treated wells (Phdp + B. pumilus ECPs) with 20 µL of Phdp-Lg 41/01 bacterial suspension, 90 µL of double-concentrated TSBs, and 90 µL of ten-fold serial dilutions of B. pumilus ECPs (initial protein concentration adjusted to 30 µg mL-¹).
B. pumilus-ECPs were added at the beginning of incubation (0 h), and bacterial growth was assessed after 48 h by measuring absorbance at OD595nm using a microplate reader (Multiskan FC, Thermo Fisher Scientific). The same procedure was used to determine the inhibitory concentrations of ICs for each B. pumilus ECP condition (Phdp + ICs). Each condition was compared with the corresponding control wells containing only culture medium. Three independent experiments were conducted, and each experiment included five technical replicates.
2.7 Production of ECPs from Phdp-Lg 41/01 grown in the presence of B. pumilus-ECPs
Phdp-Lg 41/01 was cultured as described in section 2.1. Following incubation, one or two colonies were inoculated into 10 mL tubes of TSBs. Then, the B. pumilus ECP concentrations that did not inhibit Phdp-Lg 41/01 growth (as determined in the MIC assay) were selected to supplement the bacterial cultures. These conditions were established as Phdp-ECPs + B. pumilus-ECPs (Phdp-Lg 41/01 cultured in TSB supplemented with B. pumilus ECPs at 0 h); Phdp-ECPs + B. pumilus-ICs (Phdp-Lg 41/01 cultured in TSB supplemented with internal controls under the same conditions); Phdp-ECPs or Positive control (Phdp-Lg 41/01 cultured in TSB without B. pumilus ECPs or ICs) (Figure 1).

Figure 1. Protocol of extraction of Photobacterium damselae subsp. piscicida (Phdp-Lg 41/01) supernatant alone (Phdp-ECPs) and after adding ECPs from B. pumilus UMA 169 and UMA 216 isolates grown on different media. TSA: tryptic soy agar; MICRO: media containing 5% Microchloropsis gaditana; CHL: 5% Chlorella vulgaris; ATH: 5% Arthrospira platensis. Controls were obtained from the same culture media without bacterial inoculation.
This procedure was performed to extract the apoptosis-inducing protein AIP56, a secreted AB-type toxin, as previously described by Freitas et al. (2022). The protein concentration of each supernatant was determined using the Qubit Protein Assay Kit and Qubit 2.0 Fluorometer (Thermo Fisher Scientific, USA). To confirm the absence of bacterial growth, aliquots of each ECP sample were plated on TSA and incubated at 23°C for 24–48 h. All ECPs were stored at -80°C until further use.
2.8 Effects of B. pumilus-ECPs on the relative in vitro expression of virulence genes of Phdp-Lg 41/01 by Real-Time qPCR
Phdp-Lg 41/01 was cultured on TSAs plates at 23°C for 48 h, followed by inoculation of one or two Phdp-Lg 41/01 colonies into 10 mL tubes of TSBs. To evaluate the impact of B. pumilus-ECPs on relative Phdp-Lg 41/01 virulence gene expression, selected dilutions of B. pumilus-ECPs that did not inhibit bacterial growth were added to TSBs tubes at the beginning of incubation (0 h) and incubated at 23°C for 18 h under shaking (120 rpm). Simultaneously, 10 mL tubes of Phdp-Lg 41/01 cultures were added with the internal controls (B. pumilus-ICs) and subjected to identical incubation conditions. A Phdp-Lg 41/01 culture without B. pumilus-ECPs or ICs were maintained as a positive control. After 18 h of incubation, cells were harvested by centrifugation at 5000 × g for 10 min at 4°C. Three independent experiments were conducted, each with five technical replicates (n = 5 wells).
Total RNA was extracted from bacterial cells using the RNA Purification Kit (#K0731, Thermo Scientific™) following the manufacturer’s instructions. RNA quality was assessed by mixing 2 μL of extracted RNA with 2× RNA Loading Dye (#R0641, Thermo Scientific™), followed by heat shock at 95°C for 5 min and analysis on 1% (w/v) agarose gels. The concentration of extracted RNA was quantified using the Qubit 2.0 Fluorometer with the High Sensitivity RNA Quantification Kit (Thermo Scientific, Madrid, Spain). RNA samples were stored at -80°C until further use.
Complementary DNA (cDNA) was synthesized from 100 ng of total RNA per sample using the Maxima First Strand cDNA Synthesis Kit for RT-qPCR with dsDNase (#K1671, Thermo Scientific™). The synthesized cDNA was stored at -20°C for subsequent analysis. Relative expression levels of Phdp-Lg 41/01 virulence genes were determined by real-time quantitative PCR (qRT-PCR) using 16S rRNA as the reference gene (Núñez-Díaz et al., 2018). To obtain accurate results, PCR efficiency (E) was calculated using the following equation to ensure an optimized and reproducible assay. The data generated for reactions with efficiencies between 90 and 110% were used for further analysis (Supplementary Material 1).
qRT-PCR was performed using a CFX96 Touch Real-Time PCR Detection System (Bio-Rad Laboratories, Hercules, CA, USA). The reaction mixture contained 2 μL of cDNA, 62.5 U of Taq Accustart II Trough Mix (BioMerieux, Marcy-l’Étoile, France), and 20 pmol each of R_primer and F_primer (Table 1) in a final volume of 20 μL. The PCR program included an initial denaturation cycle at 95°C for 60 s, followed by 40 cycles at 95°C for 30 s, 55°C for 40 s, and 72°C for 60 s. Amplification was followed by a standard melting curve from 65°C to 95°C, in increments of 0.5°C for 5 s at each step. Changes in gene expression were quantified using the comparative Ct method (2-ΔΔCt). Expression levels were normalized to the 16S rRNA reference gene and compared to Phdp-Lg 41/01 cells grown in TSBs.
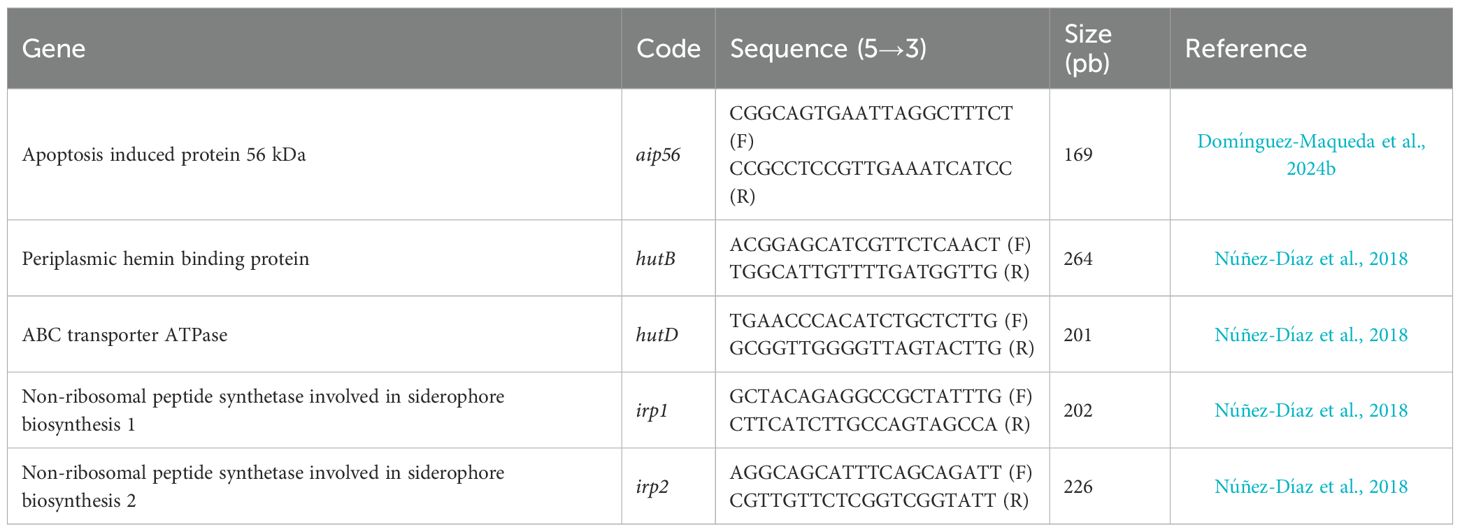
Table 1. List of primers used.
2.9 Statistical analysis
Statistical analyses were conducted using IBM SPSS Statistics 22.0. The normality and homogeneity of variance of the data were assessed using the Shapiro-Wilk and Levene’s tests, respectively. Differences between groups were evaluated by using one-way analysis of variance (ANOVA), followed by Tukey and Games-Howell post hoc tests when the assumptions of normality and homogeneity of variance were met. For non-normally distributed data, the Kruskal-Wallis test was employed, followed by a multiple comparison test. Statistical significance was established at p ≤ 0.05.
3 Results
3.1 Biofilm production
The influence of B. pumilus UMA 169 and UMA 216 ECPs obtained under different culture conditions and internal controls on the biofilm formation by the pathogenic bacterial species tested is shown in Figure 2. The biofilm formation by T. maritimum was statistically increased in the presence of ECPs from MICRO 169 and Chlorella-supplemented media (both CHL 216 and its control). With respect to V. anguillarum, all ECPs, except for MICRO 216, exhibited statistically significant reductions in biofilm formation, while all ECPs decreased A. hydrophila biofilm.

Figure 2. Inhibition of biofilm formation by T. maritimum, V. anguillarum, and A. hydrophila in the presence of ECPs from UMA 169 and UMA 216 isolates grown on algal-supplemented media. TSA: tryptic soy agar; MICRO: media containing 5% Microchloropsis gaditana; CHL: 5% Chlorella vulgaris; ATH: 5% Arthrospira platensis. Controls were obtained from the same media without bacterial inoculation. Black bars represent the biofilm formation of each bacterium (control group). The results are representative of three independent experiments and are expressed as mean ± SD (n = 5). Hash marks (#) and asterisks (*) indicate reduction and proliferation, respectively, of biofilm formation between treatments and bacterial control (p < 0.05). Different letters indicate significant differences between treatments and their internal controls (p < 0.05).
3.2 DNase activity
DNase activity was detected in MICRO 216, with 226 µg protein mL-1, being the lowest protein concentration with detectable activity (Table 2). Similarly, DNase activity was also present in CHL 216 (408 µg protein mL-1) and MICRO 169 (720 µg protein mL-1) ECPs, whilst no DNase activity was observed for either the internal controls or ATH 216 or TSA 216 ECPs (Table 2).
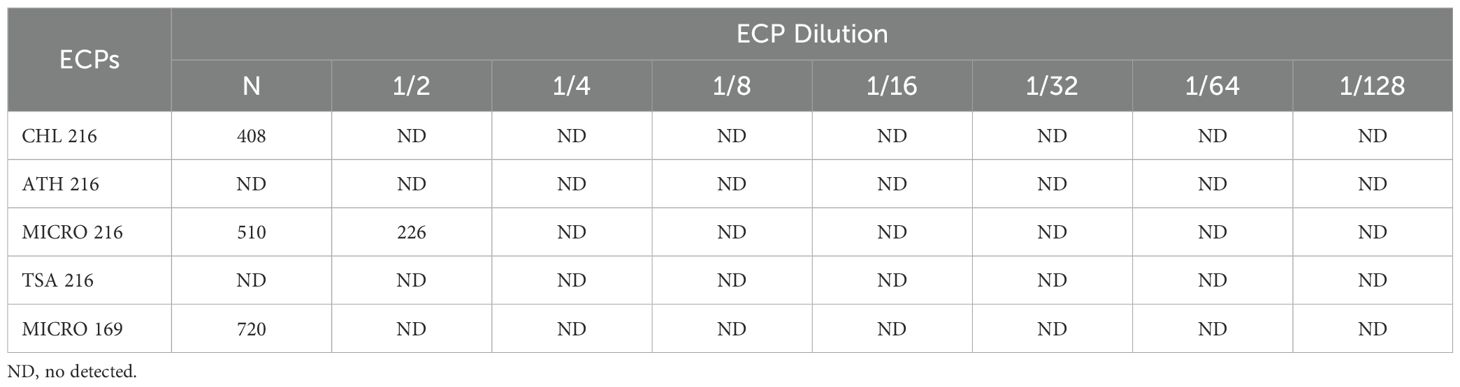
Table 2. DNase activity produced by the different ECPs (µg protein mL-1).
3.3 Quorum quenching activity
No HSL hydrolysis was detected in the ECPs assayed.
3.4 Phdp-Lg 41/01 growth in the presence of B. pumilus ECPs
The dose-response curve of Phdp-Lg 41/01 growth in the presence of ECPs showed varied inhibitory effects (Figure 3). The most potent inhibition was observed with TSA 216 ECPs, which completely suppressed bacterial growth up to a 1/16 dilution (1.88 µg mL−1 protein concentration) (Supplementary Material 2). In contrast, CHL 216, ATH 216, and MICRO 216 ECPs inhibited growth up to a 1/8 dilution (3.75 µg mL−1 protein concentration) (Supplementary Material 2). Interestingly, MICRO 169 ECPs did not show any inhibitory effect and, in fact, appeared to promote bacterial growth. For subsequent assays, we selected the dilution immediately following the minimum inhibitory concentration (MIC). This corresponded to a 1/16 dilution for ATH 216, CHL 216, and MICRO 216, and a 1/32 dilution for TSA 216. For the non-inhibitory MICRO 169 ECPs, the undiluted sample (N) was used.
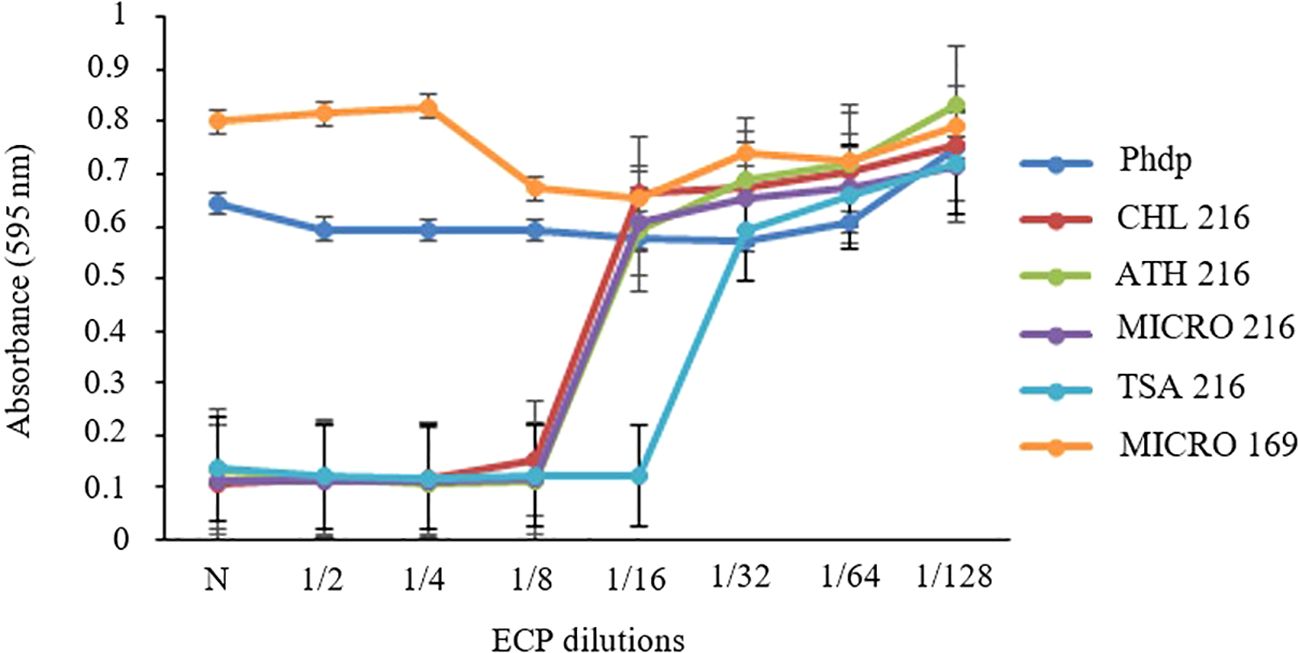
Figure 3. Dose-response curve of Phdp-Lg 41/01 growth after incubation for 48 h in the presence of ECPs from the selected isolates (B. pumilus UMA 169 and UMA 216) grown on TSA: tryptic soy agar; MICRO: media containing 5% Microchloropsis gaditana; CHL: 5% Chlorella vulgaris; ATH: 5% Arthrospira platensis. Controls were obtained from the same media without bacterial inoculation. Growth estimated based on OD595nm. N: undiluted ECPs.
3.5 Effects of B. pumilus-ECPs on the virulence genes of Phdp-Lg 41/01
All ECP conditions added to the Phdp-Lg 41/01 cultures significantly affected the aip56 gene transcription (Figure 4), except for the MICRO internal control (Control MICRO). The rest of the internal controls (Control CHL, Control ATH, and Control TSA) significantly upregulated the aip56 gene transcription compared to the control. Conversely, all ECP conditions significantly reduced the aip56 gene expression.
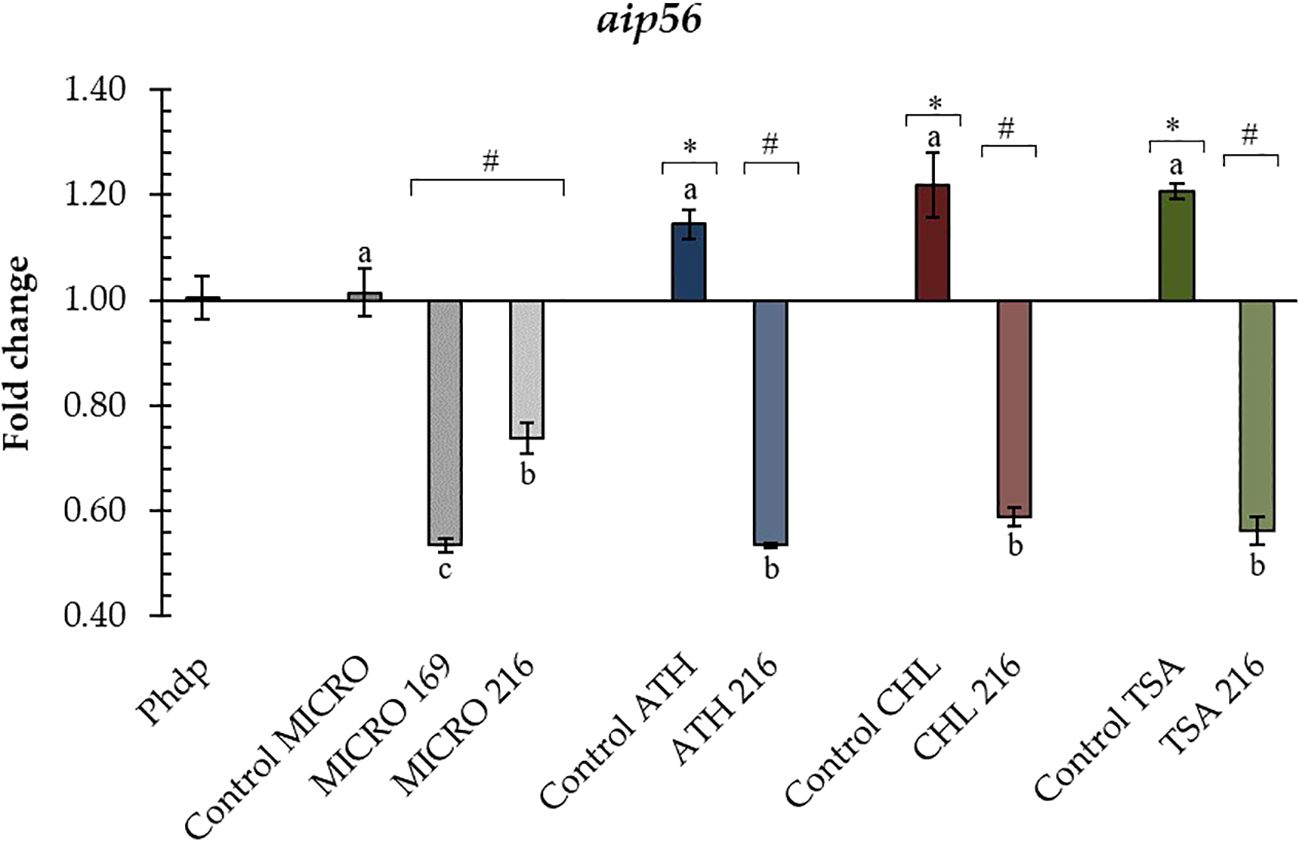
Figure 4. Relative transcription of Phdp-Lg 41/01 aip56 gene incubated with B. pumilus UMA 169 and UMA 216 ECPs. In all cases, transcription values are shown for internal controls and ECP treatments of the selected isolates. TSA: tryptic soy agar; MICRO: media containing 5% Microchloropsis gaditana; CHL: 5% Chlorella vulgaris; ATH: 5% Arthrospira platensis. Controls were obtained from the same media without bacterial inoculation. Letters (a, b) indicate significant differences (one-way ANOVA; p < 0.05) between treatments and their internal controls. Asterisks (*) and hash marks (#) indicate up- or down-regulation, respectively (one-way ANOVA; p < 0.05) compared to the control (Phdp). Values represent the mean ± standard error of the mean (SEM) of four technical replicates and two biological replicates.
Figure 5 summarizes Phdp-Lg 41/01 transcription of genes involved in iron acquisition. Significant differences were observed for hutB and hutD genes between Phdp-Lg 41/01 incubated in the presence of ECPs and their respective internal controls (Figures 5A, B), except when incubated with MICRO 216 for both genes and TSA 216 for hutB. Notably, a significant downregulation of the expression of these two genes was observed with CHL 216, ATH 216, compared to their respective controls, but not in the case of MICRO 169. Additionally, while the Control TSA significantly increased the expression of hutB but not hutD, TSA 216 downregulated the expression of the hutD gene (Figures 5A, B).
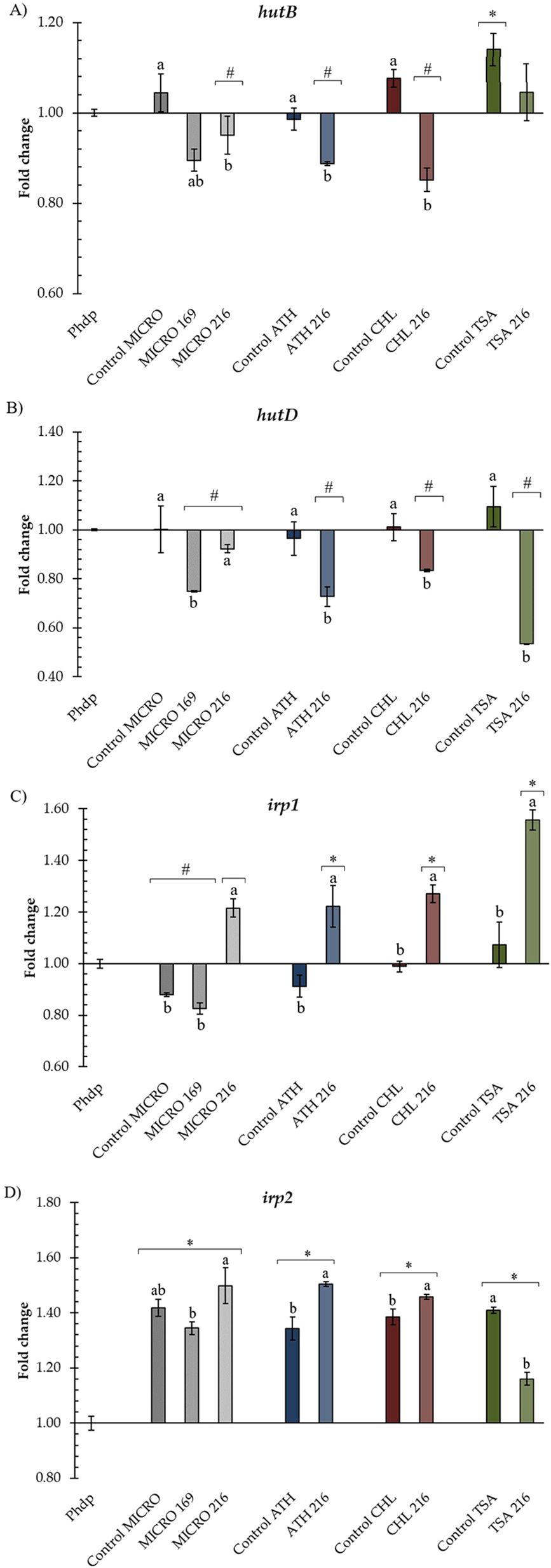
Figure 5. Relative transcription of genes (A) hutB, (B) hutD, (C) irp1, and (D) irp2 involved in iron acquisition in Phdp-Lg 41/01 incubated with B. pumilus UMA 169 and UMA 216 ECPs. TSA: tryptic soy agar; MICRO: media containing 5% Microchloropsis gaditana; CHL: 5% Chlorella vulgaris; ATH: 5% Arthrospira platensis. Controls were obtained from the same media without bacterial inoculation. In all cases, transcription values are shown for internal controls and ECP treatments of the selected isolates. Letters (a, b) indicate significant differences (one-way ANOVA; p < 0.05) between treatments and their internal controls. Asterisks (*) and hash marks (#) indicate up- or down-regulation, respectively (one-way ANOVA; p < 0.05) compared to the control (Phdp). Values represent the mean ± standard error of the mean (SEM) of four technical replicates and two biological replicates.
In general, ECP conditions (excluding their internal controls) upregulated the expression of irp1 and irp2 genes (Figures 5C, D). In the case of the irp1 gene, except for MICRO 169, upregulation with ECPs was observed compared to their respective internal controls (Figure 5C). In the case of irp2, CHL 216 and ATH 216 statistically increased the transcription of the irp2 gene compared to their respective internal controls (Figure 5D). Conversely, MICRO 169 and TSA 216 decreased the transcription of irp2 relative to their internal controls.
4 Discussion
In this study, we evaluated the ECPs derived from B. pumilus UMA 169 and UMA 216, cultured in microalgal- and cyanobacterial-enriched media, for their antibiofilm, QQ, and virulence activities against major fish pathogens. Our findings contribute to the potential of Bacillus-derived ECPs as biocontrol agents in aquaculture.
Bacterial biofilms are a major challenge in aquaculture, as they provide protection against environmental stressors and antimicrobial treatments (Naji et al., 2024). Our results demonstrate that most of the tested ECPs, except for MICRO 216, significantly inhibited biofilm formation by A. hydrophila and V. anguillarum. This suggests that bioactive compounds in ECPs interfere with biofilm development, potentially through enzymatic degradation of extracellular polymeric substances (EPS) or QS disruption (García-Márquez et al., 2025). Conversely, ECPs obtained from Microchloropsis and Chlorella-enriched media enhanced T. maritimum biofilm formation, indicating a complex interaction between ECP composition and pathogen physiology. Previous studies have reported strain-dependent biofilm modulation as well as specific EPS components variations among bacterial species (Pérez-Gómez et al., 2025). Escribano et al. (2023) also observed that ECPs from T. maritimum can enhance its own biofilm formation, reinforcing the notion that some extracellular components promote bacterial aggregation rather than inhibition.
Additionally, the presence of specific carbohydrates (e.g., mannose, galactose, glucosamine, fructose) in the culture media has been linked to increased biofilm formation in T. discolor (Schoina et al., 2024). It is possible that B. pumilus ECPs contain metabolic by-products that act as nutritional sources or signalling molecules for T. maritimum, thereby enhancing its biofilm formation. To confirm this hypothesis, further compositional analysis of UMA 169 and UMA 216 ECPs is needed.
On the other hand, the inhibition of A. hydrophila and V. anguillarum biofilms by ECPs suggests the presence of biofilm-disrupting metabolites in B. pumilus isolates (UMA 169 and UMA 216). Previous studies have demonstrated that B. pumilus-derived compounds, such as proteases and lipases, can interfere with biofilm integrity (García-Márquez et al., 2025). In addition, several studies have reported the production of secondary metabolites by Bacillus strains, including B. pumilus, with anti-biofilm activity against some bacterial pathogens (Nithya et al., 2011; Sabino et al., 2023).
Our study confirms the DNase activity in the ECPs from B. pumilus cultivated in M. gaditana and C. vulgaris-enriched media, which could contribute to biofilm degradation. Outer membrane vesicles from B. pumilus have been reported to be highly dependent on the culture medium, and they may contain lipopolysaccharides, phospholipids, and periplasmic proteins (Kurdy et al., 2024). Given that EPS, proteins, and DNA are key structural components of biofilms, it is reasonable to hypothesize that the enzymatic activities detected in ECPs from Bacillus strains contribute to biofilm inhibition, as previously suggested by other authors (Nithya et al., 2011; Monzón-Atienza et al., 2024; Pérez-Gómez et al., 2025).
To avoid the effects of growth variations among the tested strains under the different assay conditions, protein concentration was used for normalization across the various assays. However, this approach does not account for the complexity of the extracellular product composition, which exhibits variations in the content of numerous components depending on the physiological and cultural conditions of the bacteria. A detailed quantitative and qualitative analysis of the selected ECP composition must therefore be performed to specifically identify components involved in the ECP activities of the studied strains.
Despite the observed biofilm reduction in certain pathogens, no direct correlation was found between DNase activity and biofilm inhibition across all strains tested. Our assays focused primarily on biofilm formation ability, rather than on its structural integrity. DNase activity has been shown to alter biofilm biomass, architecture, and morphology, rather than inhibiting initial biofilm development (Al-Madboly et al., 2024). This observation is consistent with findings by Domínguez-Maqueda et al. (2024a), who reported that biofilm formation in T. maritimum was less affected compared to V. anguillarum and A. hydrophila. The strong surface adhesion properties of T. maritimum may contribute to its resilience, as hydrophobic interactions and rapid aggregation have been observed in in vitro biofilm assays (Levipan et al., 2019).
However, under the conditions tested in this study, none of the tested ECPs showed direct QQ activity. This suggests that the ECPs obtained did not contain molecules capable of interfering with QS signalling pathways, at least at the concentrations tested. Nevertheless, most studies have reported QQ activity predominantly in the bacterial culture supernatant, suggesting that the responsible enzymes are generally secreted (Sikdar and Elias, 2020). However, QQ activity may be strain-dependent, as notable differences in QQ capacity have been observed among Bacillus strains (Wang et al., 2004; Liu et al., 2008). In our experimental conditions, no QQ activity was detected in the ECPs, consistent with the findings of Monzón-Atienza et al. (2024) but contrasting with the observation of Santos et al. (2021b), who reported ECP-mediated QQ activity of Bacillus spp. This result does not necessarily indicate a lack of QQ potential in the B. pumilus strains, as the active molecules may remain associated with the cells (Reina et al., 2021) or be retained during the filtration process used for ECP preparation, as reported by other authors (Rehman and Leiknes, 2018).
P. damselae subsp. piscicida is the causative agent of photobacteriosis (or pasteurellosis), a prevalent disease affecting various marine fish species, including gilthead seabream (S. aurata), European seabass (Dicentrarchus labrax), and Senegalese sole (S. senegalensis) (Romalde, 2002; Santos et al., 2022; Valsamidis et al., 2023). Pathogenic P. damselae subsp. piscicida strains possess multiple virulence factors, including the ability to invade non-phagocytic cells, evade immune responses, and produce the AIP56 toxin (Abushattal et al., 2020; Freitas et al., 2025).
In the present study, all tested ECPs significantly downregulated the transcription of the aip56 gene. This suggests that the ECPs from B. pumilus UMA169 and UMA 216 contain compounds that can directly interfere with the pathogen’s ability to produce the toxin, a finding consistent with previous studies using ECPs from other probiotic bacteria. For instance, Domínguez-Maqueda et al. (2024b) reported that ECPs from the probiotic strain Shewanella putrefaciens Pdp11 grown on tryptone soy agar and media supplemented with a partial replacement of aquafeed by 25% of a blend of microalgae and cyanobacteria also downregulated Phdp-Lg 41/01 aip56 gene transcription.
Similar anti-virulence effects have been described in other Bacillus species. In B. subtilis, compounds such as surfactin have been shown to inhibit genes associated with the bacterial cell cycle (Kim et al., 2007), while fengycins—cyclic lipopeptides produced by marine B. subtilis strains—have been linked to reduced expression of ROS-scavenging enzymes in Magnaporthe grisea (Chakravarty and Massé, 2019). Furthermore, recent studies have demonstrated that cell-free supernatants (CFS) from Bacillus species can modulate virulence in diverse pathogens: CFS from B. subtilis GS3 suppressed the expression of virulence genes, including listeriolysin O, in Listeria monocytogenes (Behbahani et al., 2024), while CFS from B. thuringiensis exhibited anti-biofilm activity against Staphylococcus aureus (Ray et al., 2023).
Although the specific mechanism underlying the aip56 downregulation remains to be elucidated, the current findings indicate that B. pumilus-derived postbiotics produced using microalgal and cyanobacterial culture media represent a promising approach to modulate bacterial metabolism and attenuate virulence. Moreover, their application for the biocontrol of P. damselae subsp. piscicida introduces a novel mechanistic perspective, offering insights into how these compounds interfere with pathogen physiology. This strategy could ultimately enhance understanding of the infection process and support the development of more effective interventions, including future in vivo validation trials.
Interestingly, our results also revealed a complex regulatory effect of the ECPs on the pathogen’s iron acquisition system. We investigated the relative expression of genes involved in haem utilization (hutB and hutD) and the production of the siderophore piscibactin (irp1 and irp2) in Phdp-Lg 41/01. Our results indicated significant differences in gene expression between the pathogen incubated with assayed ECPs and their respective internal controls, suggesting a potential regulatory role of these ECPs in the expression of virulence-related genes. The downregulation of hutB and hutD genes in the presence of ECP conditions such as CHL 216, ATH 216, and MICRO 169 is noteworthy, as both genes are essential components of the haem utilization system, which allows bacteria to acquire iron from host hemin and haemoglobin (Magariños et al., 1994; Osorio et al., 2010, 2015). On the other hand, the upregulation of the irp1 and irp2 genes in most ECP conditions suggests a potential activation of piscibactin production pathways.
The differential effects of ECP conditions on gene expression highlight the complexity of bacterial regulatory mechanisms. For instance, ECP conditions CHL 216 and ATH 216 exhibited downregulation of genes associated with haem utilization (hutB and hutD) compared to both Phdp-Lg 41/01 and their respective control media (Control CHL and Control ATH). Conversely, despite the downregulation of haem utilization genes, the same ECP conditions (CHL 216 and ATH 216) induced the transcription of genes involved in the production of the siderophore piscibactin (irp1 and irp2) relative to Phdp-Lg 41/01 and their respective control media. The upregulation of irp1 and irp2 genes suggests that these ECPs may stimulate the ability of P. damselae subsp. piscicida to acquire iron through piscibactin production (Lemos and Balado, 2020). It is conceivable that the composition of ECPs derived from different culture conditions modulate specific signalling pathways involved in iron acquisition in Phdp-Lg 41/01, leading to differential gene expression patterns. Further studies should be carried out to elucidate the specific mechanisms controlling these regulatory effects and to evaluate the impact of ECPs on P. damselae subsp. piscicida virulence in vivo.
In conclusion, this study provides strong evidence that ECPs from B. pumilus UMA 216 and UMA 169 cultivated in microalgal-supplemented media are promising tools for sustainable pathogen management in aquaculture. These ECPs effectively inhibit biofilm formation in certain pathogens such as V. anguillarum, A. hydrophila, and P. damselae subsp. piscicida, and can modulate the expression of key virulence-related genes, such as the AIP56 toxin gene, despite lacking direct quorum-quenching activity. These findings highlight the potential of developing Bacillus-derived postbiotics as a valuable alternative to conventional antimicrobials.
Moreover, this study highlights the use of microalgae as a sustainable substitute for conventional media in probiotic cultivation—a promising strategy to enhance postbiotic production. Microalgal biomass, rich in essential nutrients and bioactive compounds, serves as a natural substrate that stimulates bacterial growth and modulates probiotic metabolism, thereby promoting the secretion of novel or higher concentrations of bioactive molecules with postbiotic potential. This approach harnesses renewable resources to generate postbiotics with unique functional properties. Future research should focus on identifying the specific compounds responsible for these effects and validating their efficacy in vivo.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
JG-M: Formal analysis, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing. MD-M: Formal analysis, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing. IL: Data curation, Funding acquisition, Investigation, Validation, Writing – review & editing. ST: Data curation, Investigation, Writing – review & editing. SA: Investigation, Validation, Writing – review & editing. MM: Conceptualization, Funding acquisition, Investigation, Project administration, Resources, Supervision, Validation, Writing – review & editing. MB: Conceptualization, Funding acquisition, Investigation, Project administration, Resources, Supervision, Validation, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was funded by Junta de Andalucía and co-financed by FEDER funds (Grant P20-00085). This work was also funded by the Agencia Estatal de Investigación (grants PID2020-113637RB-C22/AEI/10.13039/501100011033 and PID2019-106704RB-100/AEI/10.13039/501100011033).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1705909/full#supplementary-material
References
Abushattal S., Vences A., and Osorio C. R. (2020). A virulence gene typing scheme for Photobacterium damselae subsp. piscicida, the causative agent of fish photobacteriosis, reveals a high prevalence of plasmid-encoded virulence factors and of type III secretion system genes. Aquaculture 521, 735057. doi: 10.1016/J.AQUACULTURE.2020.735057
Aguilar-Toalá J. E., Garcia-Varela R., Garcia H. S., Mata-Haro V., González-Córdova A. F., Vallejo-Cordoba B., et al. (2018). Postbiotics: An evolving term within the functional foods field. Trends Food Sci. Technol. 75, 105–114. doi: 10.1016/J.TIFS.2018.03.009
Al-Madboly L. A., Aboulmagd A., El-Salam M. A., Kushkevych I., and El-Morsi R. M. (2024). Microbial enzymes as powerful natural anti-biofilm candidates. Microb. Cell Factories 23, 343. doi: 10.1186/s12934-024-02610-y
Austin B. (2019). Methods for the diagnosis of bacterial fish diseases. Mar. Life Sci. Technol. 1, 41–49. doi: 10.1007/S42995-019-00002-5/TABLES/3
Barros C. P., Guimarães J. T., Esmerino E. A., Duarte M. C. K., Silva M. C., Silva R., et al. (2020). Paraprobiotics and postbiotics: concepts and potential applications in dairy products. Curr. Opin. Food Sci. 32, 1–8. doi: 10.1016/J.COFS.2019.12.003
Behbahani B. A., Noshad M., Vasiee A., and Brück W. M. (2024). Probiotic Bacillus strains inhibit growth, biofilm formation, and virulence gene expression of Listeria monocytogenes. Lwt 191, 115596. doi: 10.1016/j.lwt.2023.115596
Borges Lopes L. D., Soares R. C., de Freitas R. M., Amarante D. O., de Carvalho F. C. T., Cavalcante K. M. D. S. P., et al. (2025). Co-culture of microalgae and bacteria for the production of bioactive compounds. Ann. Microbiol. 75, 16. doi: 10.1186/s13213-025-01804-y
Cerezo I. M., Pérez-Gómez O., Rohra-Benítez S., Domínguez-Maqueda M., García-Márquez J., and Arijo S. (2025). Postbiotics of marine origin and their therapeutic application. Mar. Drugs 23, 335. doi: 10.3390/md23090335
Chakravarty S. and Massé E. (2019). RNA-dependent regulation of virulence in pathogenic bacteria. Front. Cell Infect. Microbiol. 9. doi: 10.3389/fcimb.2019.00337
Chilton M. D., Currier T. C., Farrand S. K., Bendich A. J., Gordon M. P., and Nester E. W. (1974). Agrobacterium tumefaciens DNA and PS8 bacteriophage DNA not detected in crown gall tumors. Proc. Natl. Acad. Sci. 71, 3672–3676. doi: 10.1073/PNAS.71.9.3672
Croxatto A., Lauritz J., Chen C., and Milton D. L. (2007). Vibrio Anguillarum colonization of rainbow trout integument requires a DNA locus involved in exopolysaccharide transport and biosynthesis. Environ. Microbiol. 9, 370–382. doi: 10.1111/J.1462-2920.2006.01147.X
Díaz-Rosales P., Chabrillón M., Moriñigo M. A., and Balebona M. C. (2003). Survival against exogenous hydrogen peroxide of Photobacterium damselae subsp. piscicida under different culture conditions. J. Fish Dis. 26, 305–308. doi: 10.1046/J.1365-2761.2003.00455.X
Domínguez-Maqueda M., Espinosa-Ruíz C., Esteban M.Á., Alarcón F. J., Tapia-Paniagua S. T., Balebona M. C., et al. (2024b). An ex vivo approach in European seabass leucocytes supports the in vitro regulation by postbiotics of aip56 gene expression of Photobacterium damselae subsp. piscicida. Probiotics Antimicrob. Proteins, 1–15. doi: 10.1007/S12602-024-10255-X/FIGURES/8
Domínguez-Maqueda M., García-Márquez J., Tapia-Paniagua S. T., González-Fernández C., Cuesta A., Espinosa-Ruíz C., et al. (2024a). Evaluation of the differential postbiotic potential of Shewanella putrefaciens Pdp11 cultured in several growing conditions. Mar. Biotechnol. 26, 1–18. doi: 10.1007/S10126-023-10271-Y/FIGURES/4
Escribano M. P., Balado M., Toranzo A. E., Lemos M. L., and Magariños B. (2023). The secretome of the fish pathogen Tenacibaculum maritimum includes soluble virulence-related proteins and outer membrane vesicles. Front. Cell Infect. Microbiol. 13. doi: 10.3389/fcimb.2023.1197290
FAO (Food and Agriculture Organization of the United Nations) (2022). The State of World Fisheries and Aquaculture 2022. Towards Blue Transformation (Rome, Italy: FAO). doi: 10.4060/cc0461en
Freitas I. L., Macedo M. F., Oliveira L., Oliveira P., do Vale A., and Dos Santos N. M. (2025). AIP56, an AB toxin secreted by Photobacterium damselae subsp. piscicida, has tropism for myeloid cells. Front. Immunol. 15. doi: 10.3389/fimmu.2024.1527088
Freitas I. L., Teixeira A., Loureiro I., Lisboa J., Saraiva A., Dos Santos N. M. S., et al. (2022). Susceptibility of sea bream (Sparus aurata) to AIP56, an AB-type toxin secreted by Photobacterium damselae subsp. piscicida. Toxins 14, 119. doi: 10.3390/TOXINS14020119
Ganesh P. S., Vishnupriya S., Vadivelu J., Mariappan V., Vellasamy K. M., and Shankar E. M. (2020). Intracellular survival and innate immune evasion of Burkholderia cepacia: Improved understanding of quorum sensing-controlled virulence factors, biofilm, and inhibitors. Microbiol. Immunol. 64, 87–98. doi: 10.1111/1348-0421.12762
García-Márquez J., Domínguez-Maqueda M., Torres M., Cerezo I. M., Ramos E., Alarcón F. J., et al. (2023). Potential effects of microalgae-supplemented diets on the growth, blood parameters, and the activity of the intestinal microbiota in Sparus aurata and Mugil cephalus. Fishes 8, 409. doi: 10.3390/FISHES8080409/S1
García-Márquez J., Galafat A., Molina-Roque L., Domínguez-Maqueda M., De las Heras V., Simó-Mirabet P., et al. (2025). Microalgal and cyanobacterial biomasses modified the activity of extracellular products from Bacillus pumilus: an in vitro and in vivo assessment. Probiotics Antimicrob. Proteins 17(4), 2179–2196. doi: 10.1007/s12602-024-10350-z
Ghanei-Motlagh R., Mohammadian T., Gharibi D., Menanteau-Ledouble S., Mahmoudi E., Khosravi M., et al. (2020). Quorum quenching properties and probiotic potentials of intestinal associated bacteria in Asian sea bass Lates calcarifer. Mar. Drugs 18, 23. doi: 10.3390/MD18010023
Gonzales M., Jacquet P., Gaucher F., Chabriere E., Plener L., and Daude D. (2024). AHL-based quorum sensing regulates the biosynthesis of a variety of bioactive molecules in bacteria. J. Nat. Prod. 87, 1268–1284. doi: 10.1021/acs.jnatprod.3c00672
Guo M., Meng L., Li F., Liu Y., Zhang Z., Zhang Q., et al. (2025). Effects of dietary Bacillus pumilus on the growth, intestinal health, lipid metabolism, and mTOR signaling pathway of juvenile coho salmon (Oncorhynchus kisutch). Water Biol. Secur. 4, 100313. doi: 10.1016/j.watbs.2024.100313
Harrison A. M. and Soby S. D. (2020). Reclassification of Chromobacterium violaceum ATCC 31532 and its quorum biosensor mutant CV026 to Chromobacterium subtsugae. AMB Express 10, 1–7. doi: 10.1186/S13568-020-01140-1/FIGURES/3
Hill C., Guarner F., Reid G., Gibson G. R., Merenstein D. J., Pot B., et al. (2014). The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 11, 506–514. doi: 10.1038/nrgastro.2014.66
Juszczuk-Kubiak E. (2024). Molecular aspects of the functioning of pathogenic bacteria biofilm based on quorum sensing (QS) signal-response system and innovative non-antibiotic strategies for their elimination. Int. J. Mol. Sci. 25, 2655. doi: 10.3390/ijms25052655
Kim S. Y., Kim J. Y., Kim S. H., Bae H. J., Yi H., Yoon S. H., et al. (2007). Surfactin from Bacillus subtilis displays anti-proliferative effect via apoptosis induction, cell cycle arrest and survival signaling suppression. FEBS Lett. 581, 865–871. doi: 10.1016/j.febslet.2007.01.059
Kurdy W., Zelenikhin P. V., Yakovleva G. Y., Sinyagina M. N., Kolpakov A. I., and Ilinskaya O. N. (2024). The proteome of extracellular membrane vesicles from Bacillus pumilus 3-19. Mol. Biol. 58, 157–164. doi: 10.1134/S0026893324010059
Lemos M. L. and Balado M. (2020). Iron uptake mechanisms as key virulence factors in bacterial fish pathogens. J. Appl. Microbiol. 129, 104–115. doi: 10.1111/JAM.14595
Levipan H. A., Tapia-Cammas D., Molina V., Irgang R., Toranzo A. E., Magariños B., et al. (2019). Biofilm development and cell viability: an undervalued mechanism in the persistence of the fish pathogen Tenacibaculum maritimum. Aquaculture 511, 734267. doi: 10.1016/j.aquaculture.2019.734267
Li J., Jia T., and Yang L. (2025). Targeting anti-virulence factor strategies of bacterial pathogens. Biosaf. Health 7, 1–4. doi: 10.1016/j.bsheal.2025.01.006
Liu P. V. (1957). Survey of hemolysin production among species of Pseudomonads. J. Bacteriol. 74, 718. doi: 10.1128/JB.74.6.718-727.1957
Liu D., Momb J., Thomas P. W., Moulin A., Petsko G. A., Fast W., et al. (2008). Mechanism of the Quorum-Quenching lactonase (AiiA) from Bacillus thuringiensis. 1. Product-bound structures. Biochemistry 47, 7706. doi: 10.1021/BI800368Y
Lu S., Na K., Li Y., Zhang L., Fang Y., and Guo X. (2022). Bacillus-derived probiotics: metabolites and mechanisms involved in bacteria–host interactions. Crit. Rev. Food Sci. Nutr. 64, 1701–1714. doi: 10.1080/10408398.2022.2118659
Ma M. and Hu Q. (2023). Microalgae as feed sources and feed additives for sustainable aquaculture: Prospects and challenges. Rev. Aquac. 16, 818–835. doi: 10.1111/RAQ.12869
Mabrok M., Algammal A. M., Sivaramasamy E., Hetta H. F., Atwah B., Alghamdi S., et al. (2023). Tenacibaculosis caused by Tenacibaculum maritimum: Updated knowledge of this marine bacterial fish pathogen. Front. Cell Infect. Microbiol. 12. doi: 10.3389/FCIMB.2022.1068000/BIBTEX
Magariños B., Romalde J. L., Lemos M. L., Barja J. L., and Toranzo A. E. (1994). Iron uptake by Pasteurella piscicida and its role in pathogenicity for fish. Appl. Environ. Microbiol. 60, 2990–2998. doi: 10.1128/AEM.60.8.2990-2998.1994
McClean K. H., Winson M. K., Fish L., Taylor A., Chhabra S. R., Camara M., et al. (1997). Quorum sensing and Chromobacterium violaceum: Exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology 143, 3703–3711. doi: 10.1099/00221287-143-12-3703/CITE/REFWORKS
Monzón-Atienza L., Bravo J., Torrecillas S., Gómez-Mercader A., Montero D., Ramos-Vivas J., et al. (2024). An in-depth study on the inhibition of quorum sensing by Bacillus velezensis D-18: its significant impact on Vibrio biofilm formation in aquaculture. Microorganisms 12, 890. doi: 10.3390/microorganisms12050890
Moradi M., Molaei R., and Guimarães J. T. (2021). A review on preparation and chemical analysis of postbiotics from lactic acid bacteria. Enzyme Microb. Technol. 143, 109722. doi: 10.1016/J.ENZMICTEC.2020.109722
Morohoshi T., Kato M., Fukamachi K., Kato N., and Ikeda T. (2008). N-Acylhomoserine lactone regulates violacein production in Chromobacterium violaceum type strain ATCC 12472. FEMS Microbiol. Lett. 279, 124–130. doi: 10.1111/J.1574-6968.2007.01016.X
Naji N. P., Chandrakala N., Eswari J., Shanmugapriya S., Geetha R. V., Gopal R. K., et al. (2024). “Biofilm in Aquaculture Microorganisms,” in Aquatic Ecosystems and Microbial Biofilms (Boca Raton, Florida, United States of America: CRC Press), 264–281). doi: 10.1201/9781003487203
Nithya C., Devi M. G., and Karutha Pandian S. (2011). A novel compound from the marine bacterium Bacillus pumilus S6–15 inhibits biofilm formation in gram-positive and gram-negative species. Biofouling 27, 519–528. doi: 10.1080/08927014.2011.586127
Núñez-Díaz J. A., Fumanal M., Do Vale A., Fernández-Díaz C., Moriñigo M.Á., and Balebona M. C. (2018). Transcription of IVIAT and virulence genes in Photobacterium damselae subsp. piscicida infecting Solea Senegalensis. Microorganisms 6, 67. doi: 10.3390/MICROORGANISMS6030067
Osorio C. R., Juiz-Río S., and Lemos M. L. (2010). The ABC-transporter hutCD genes of Photobacterium damselae subsp. piscicida are essential for haem utilization as iron source and are expressed during infection in fish. J. Fish Dis. 33, 649–655. doi: 10.1111/J.1365-2761.2010.01169.X
Osorio C. R., Rivas A. J., Balado M., Fuentes-Monteverde J. C., Rodríguez J., Jiménez C., et al. (2015). A transmissible plasmid-borne pathogenicity island confers piscibactin biosynthesis in the fish pathogen Photobacterium damselae subsp. piscicida. Appl. Environ. Microbiol. 81, 5867. doi: 10.1128/AEM.01580-15
Pazos F., Santos Y., Macías A. R., Núñez S., and Toranzo A. E. (1996). Evaluation of media for the successful culture of Flexibacter maritimus. J. Fish Dis. 19, 193–197. doi: 10.1111/J.1365-2761.1996.TB00701.X
Pereira W. A., Mendonça C. M. N., Urquiza A. V., Marteinsson V.Þ., LeBlanc J. G., Cotter P. D., et al. (2022). Use of probiotic bacteria and bacteriocins as an alternative to antibiotics in aquaculture. Microorganisms 10, 1705. doi: 10.3390/MICROORGANISMS10091705
Pérez-Gómez O., Domínguez-Maqueda M., García-Márquez J., Moriñigo M.Á., and Tapia-Paniagua S. T. (2025). Metabolite-driven modulation of biofilm formation in shewanella: insights from shewanella sp. Pdp11 extracellular products. Microb. Ecol. 88, 55. doi: 10.1007/s00248-025-02552-x
Pokorzynski N. D. and Groisman E. A. (2023). How bacterial pathogens coordinate appetite with virulence. Microbiol. Mol. Biol. Rev. 87, e00198–e00122. doi: 10.1128/mmbr.00198-22
Porras M., Hernández D., and Boto A. (2025). Short synthesis of structurally diverse N-acylhomoserine lactone analogs and discovery of novel quorum quenchers against gram-negative pathogens. Int. J. Mol. Sci. 26, 1775. doi: 10.3390/ijms26041775
Priya P. S., Boopathi S., Murugan R., Haridevamuthu B., Arshad A., and Arockiaraj J. (2023). Quorum sensing signals: Aquaculture risk factor. Rev. Aquac. 15, 1294–1310. doi: 10.1111/RAQ.12774
Rahman M. A., Ashrafudoulla M., Akter S., Park S. H., and Ha S. D. (2024). Probiotics and biofilm interaction in aquaculture for sustainable food security: A review and bibliometric analysis. Crit. Rev. Food Sci. Nutr. 64, 12319–12335. doi: 10.1080/10408398.2023.2249114
Ramírez-Larrota J. S. and Eckhard U. (2022). An introduction to bacterial biofilms and their proteases, and their roles in host infection and immune evasion. Biomolecules 12, 306. doi: 10.3390/BIOM12020306
Rasmussen-Ivey C. R., Figueras M. J., McGarey D., and Liles M. R. (2016). Virulence factors of Aeromonas hydrophila: In the wake of reclassification. Front. Microbiol. 7. doi: 10.3389/FMICB.2016.01337/BIBTEX
Ray S., Jin J. O., Choi I., and Kim M. (2023). Cell-free supernatant of Bacillus thuringiensis displays anti-biofilm activity against Staphylococcus aureus. Appl. Biochem. Biotechnol. 195, 5379–5393. doi: 10.1007/s12010-022-03971-z
Rehman Z. U. and Leiknes T. (2018). Quorum-quenching bacteria isolated from red sea sediments reduce biofilm formation by pseudomonas aeruginosa. Front. Microbiol. 9. doi: 10.3389/fmicb.2018.01354
Reina J. C., Romero M., Salto R., Cámara M., Llamas I., and Aha P. (2021). A Quorum Quenching Acylase from Psychrobacter sp. M9-54–1 That Attenuates Pseudomonas aeruginosa and Vibrio coralliilyticus Virulence. Mar. Drugs 19, 16. doi: 10.3390/md19010016
Ricós-Muñoz N., Rivas Soler A., Castagnini J. M., Moral R., Barba F. J., and Pina-Pérez M. C. (2023). Improvement of the probiotic growth-stimulating capacity of microalgae extracts by pulsed electric fields treatment. Innov. Food Sci. Emerg. Technol. 83, 103256. doi: 10.1016/J.IFSET.2022.103256
Romalde J. L. (2002). Photobacterium damselae subsp. piscicida: An integrated view of a bacterial fish pathogen. Int. Microbiol. 5, 3–9. doi: 10.1007/S10123-002-0051-6/METRICS
Sabino Y. N. V., Araújo Domingues K. C., and Mathur H. (2023). Exopolysaccharides produced by Bacillus spp. inhibit biofilm formation by Staphylococcus aureus strains associated with bovine mastitis. Int J Biol Macromol. 253(Pt 2), 126689. doi: 10.1016/j.ijbiomac.2023.126689
Santos R. A., Monteiro M., Rangel F., Jerusik R., Saavedra M. J., Carvalho A. P., et al. (2021b). Bacillus spp. Inhibit Edwardsiella tarda Quorum-Sensing and Fish Infection. Mar. Drugs 19, 602. doi: 10.3390/md19110602
Santos R. A., Oliva-Teles A., Pousão-Ferreira P., Jerusik R., Saavedra M. J., Enes P., et al. (2021a). Isolation and characterization of fish-gut Bacillus spp. as source of natural antimicrobial compounds to fight aquaculture bacterial diseases. Mar. Biotech. 23, 276–293. doi: 10.1007/S10126-021-10022-X/FIGURES/1
Santos P., Peixoto D., Ferreira I., Passos R., Pires P., Simões M., et al. (2022). Short-term immune responses of gilthead seabream (Sparus aurata) juveniles against Photobacterium damselae subsp. piscicida. Int. J. Mol. Sci. 23, 1561. doi: 10.3390/IJMS23031561/S1
Schoina E., Marchand L., Zykwinska A., Sinquin C., Leroi F., Delbarre-Ladrat C., et al. (2024). A thorough insight into the biofilm properties of potential fish pathogen Tenacibaculum discolor strain FMCC B487. Aquac. J. 4, 76–91. doi: 10.3390/aquacj4020006
Shaw P. D., Ping G., Daly S. L., Cha C., Cronan J. E., Rinehart K. L., et al. (1997). Detecting and characterizing N-acyl-homoserine lactone signal molecules by thin-layer chromatography. Proc. Natl. Acad. Sci. 94, 6036–6041. doi: 10.1073/pnas.94.12.6036
Sikdar R. and Elias M. (2020). Quorum quenching enzymes and their effects on virulence, biofilm, and microbiomes: a review of recent advances. Expert. Rev. Anti Infect. Ther. 18, 1221–1233. doi: 10.1080/14787210.2020.1794815
Thakur K., Singh B., Kumar S., Sharma D., Sharma A. K., Jindal R., et al. (2025). Potential of probiotics and postbiotics in aquaculture: connecting current research gaps and future perspectives. Microbe 8, 100431. doi: 10.1016/j.microb.2025.100431
Torres M., Rubio-Portillo E., Antón J., Ramos-Esplá A. A., Quesada E., and Llamas I. (2016). Selection of the N-acylhomoserine lactone-degrading bacterium Alteromonas stellipolaris PQQ-42 and of its potential for biocontrol in aquaculture. Front. Microbiol. 7. doi: 10.3389/FMICB.2016.00646/BIBTEX
Valsamidis M. A., White D. M., Kokkoris G. D., and Bakopoulos V. (2023). Immune response of European sea bass (Dicentrarchus labrax L.) against combination of antigens from three different pathogens. Vet. Immunol. Immunopathol. 256, 110535. doi: 10.1016/J.VETIMM.2022.110535
Vivas J., Padilla D., Real F., Bravo J., Grasso V., and Acosta F. (2008). Influence of environmental conditions on biofilm formation by Hafnia alvei strains. Vet. Microbiol. 129, 150–155. doi: 10.1016/j.vetmic.2007.11.007
Wang L. H., Weng L. X., Dong Y. H., and Zhang L. H. (2004). Specificity and enzyme kinetics of the quorum-quenching N-Acyl homoserine lactone lactonase (AHL-lactonase). J. Biol. Chem. 279, 13645–13651. doi: 10.1074/jbc.M311194200
Whiteley M., Diggle S. P., and Greenberg E. P. (2017). Progress in and promise of bacterial quorum sensing research. Nature 551, 313–320. doi: 10.1038/nature24624
Yan R., Zeng X., Shen J., Wu Z., Guo Y., Du Q., et al. (2024). New clues for postbiotics to improve host health: a review from the perspective of function and mechanisms. J. Sci. Food Agric. 104, 6376–6387. doi: 10.1002/jsfa.13444.38450745
Yi Y., Zhang Z., Zhao F., Liu H., Yu L., Zha J., et al. (2018). Probiotic potential of Bacillus velezensis JW: Antimicrobial activity against fish pathogenic bacteria and immune enhancement effects on Carassius auratus. Fish Shellfish Immunol. 78, 322–330. doi: 10.1016/J.FSI.2018.04.055
Keywords: Bacillus pumilus, biofilm inhibition, extracellular products, postbiotics, quorum sensing
Citation: García-Márquez J, Domínguez-Maqueda M, Llamas I, Tapia-Paniagua ST, Arijo S, Moriñigo MÁ and Balebona MC (2025) Extracellular products derived from Bacillus pumilus cultured on microalgal and cyanobacterial supplemented media: potential for controlling four specific aquaculture pathogens. Front. Mar. Sci. 12:1705909. doi: 10.3389/fmars.2025.1705909
Received: 15 September 2025; Accepted: 13 November 2025; Revised: 05 November 2025;
Published: 26 November 2025.
Edited by:
Bin Wu, Zhejiang University, ChinaReviewed by:
Fenglin Wang, Ningbo University, ChinaKarla Alejandra Soto Marfileño, Universidad Autónoma de Nuevo León, Mexico
Copyright © 2025 García-Márquez, Domínguez-Maqueda, Llamas, Tapia-Paniagua, Arijo, Moriñigo and Balebona. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Miguel Ángel Moriñigo, bW9yaW5pZ29AdW1hLmVz
†These authors have contributed equally to this work