 John Birger Ulvund
John Birger Ulvund Aslak Smalås
Aslak Smalås Rita Strand1
Rita Strand1- 1Department of Life Sciences, Scandinavian Nature Surveillance a DNV company (Det Norske Veritas), Trondheim, Norway
- 2Arctic and Marine Biology, UiT - The Arctic University of Norway, Tromsø, Norway
Migrations between different habitats to maximise fitness are well documented in many organisms, although such migrations imply a potentially great risk when moving between habitats. For anadromous salmonids, these migrations occur between the freshwater stage in their natal rivers to coastal waters during spring/summer, where they can potentially experience high mortality rates. This study investigated the riverine migration dynamics of Atlantic salmon (Salmo salar) and sea trout (Salmo trutta) smolts with a multiple camera array over three consecutive years (2021–2023). A total of 19,238 individuals were recorded, and the dynamics of schooling behaviour and diel activity patterns were examined with respect to both intra- and interspecific traits associated with migratory behaviour. The observed pattern describes the migration dynamics for sympatric natural stocks of Atlantic salmon and sea trout over a 3-year period with little-to-no interferences with natural behaviour. Results indicate a clear diel pattern in schooling behaviour, governed by the influx of natural light, with clear species-specific differences in migratory behaviour. The observed diel pattern was less dominant later in the migration period, revealing daylight length as a factor altering migratory behaviour during the smolt run. The observed changes in migration dynamics could possibly be adapted as an anti-predator behaviour during river migration, where night/dark offers protection from visual predators, whereas safety in numbers aids predator detection and avoidance during daytime.
Introduction
The life history and migration patterns of anadromous Atlantic salmon (Salmo salar) and brown trout (Salmo trutta), hereafter sea trout, have fascinated and attracted scientific interest for decades. Both species exhibit high plasticity in life history traits, where perhaps the most intriguing and diverse part of their life history is anadromy. Prior to the initiation of the seaward migration, smolt experience a change in both morphological and physiological attributes (smoltification) to adapt to both the saline levels of the ocean and the ability to cover long distances in search for food (Hoar, 1988). The seaward migration from their natal rivers typically occurs from spring to early summer; the size and age of these smolts are between 12 and 25 cm and between 1 and 6 years, respectively (Thorstad et al., 2010; 2012).
Seaward migration of smolt is a critical life history event where the timing of sea entry to optimise favourable marine environmental conditions is crucial for maximising the survival and fitness of these salmonids (McCormick et al., 1998; Harvey et al., 2020). Migratory cues triggering downstream migration are well documented in the literature, and factors such as photoperiod, water discharge, temperature, and the interaction effects between these are often seen as main drivers for migration (McCormick et al., 1998; Thorstad et al., 2010; Bjerck et al., 2021; Hawley et al., 2024). The downstream migration in the freshwater phase is shown to be an active migration, i.e., the fish moves faster than the water currents (Newton et al., 2021). Atlantic salmon and sea trout smolts have been proven to respond to the same environmental triggers for migration (Harvey et al., 2020). In Norwegian rivers, Atlantic salmon smolt typically start their migration period somewhat earlier than sea trout, but the differences have shown to be negligible for some rivers (Jensen et al., 2012; Haraldstad et al., 2017; Harvey et al., 2020).
Visual predators like birds [gulls (Laridae sp.) and mergansers (Mergus sp.)], mammals [otters (Lutra lutra) and mink (Neovison vison)], and other fish [brown trout and pike (Esox lucius)] are the most common predators of salmonid smolt migrating toward the sea, and predator-induced mortality rates during the riverine migration phase are reported to be between 0.3 % and 7 % km−1 (Thorstad et al., 2012). Mortality rates from these predators are dependent on environmental factors such as water discharge, light, turbidity, and temperature (Thorstad et al., 2012; Gauld et al., 2013: Haraldstad et al., 2017). However, anti-predator behaviour such as nocturnal migration and schooling are considered normal (Thorstad et al., 2012). The smolt run is shown to be predominantly nocturnal in the beginning, with a transition to more daytime migration later in the season due to prolonged daylight and rising temperatures, especially in northern latitudes (Ibbotson et al., 2006; Haraldstad et al., 2017). However, migration in relation to daylight and its impact on the dynamics of smolt migration and inter-species differences are not fully understood. Schooling behaviour is a well-documented anti-predator strategy in many fish species (Marras and Domenici, 2013), but the interspecific dynamics is scarcely documented for descending Atlantic salmon and sea trout smolts in sympatric populations. A better understanding of not only the environmental factors affecting the degree of schooling during riverine smolt migration but also the dynamics of migratory strategies is needed to enhance the understanding of migratory behaviour in salmonids.
Data on natural schooling behaviour of salmonid smolts are a concept that is notoriously difficult to document throughout the migration period without invasive studies. PIT-tagged fish are prone to not be detected if they pass the PIT antenna during the same/short period (Holcombe et al., 2019; Zetner et al., 2021), acoustic tagged fish studies typically do not have the numbers of tagged fish to estimate such effects due to the cost of tags, and tagged experiments typically operate in the space of 50–200 tagged individuals (Hussey et al., 2015). In addition, both PIT and acoustic tagging of fish involve capture, handling and post-operation practises that may harm the fish and their natural behaviour (Thorstad et al., 2013; Vollset et al., 2020). The current study used a video surveillance system detecting close to 20,000 descending smolts from a river system in the western part of Norway from 2021 to 2023. We investigate if and how environmental factors (water discharge, temperature, and light intensity) impact anti-predator behaviour like nocturnal migration and schooling, and if there are any behavioural differences between descending Atlantic salmon and sea trout smolts. Understanding both the behavioural mechanisms of migration and anti-predator strategies, and factors affecting this critical life stage, is of vital importance for better management and preservation of these ecologically and economically valuable species.
Material and methods
River Ervika, a part of Dalsbøvassdraget, is located on the outermost part of the Norwegian coast in the middle of Western Norway (62°9.732 N; 5°6.710 E) (Figure 1). The total accumulated drainage basin for the river is 32.8 km2, and the river has a total length of 11.6 km. The water course consists of multiple river stretches and four lakes (Morkavatnet, Sætrevatnet, Dalsbøvatnet, and Ervikvatnet). The only two anadromous species of this river are Atlantic salmon and sea trout. Sea-run fish can access a total anadromous stretch of 9.3 km up to the final migration barrier located just downstream of Morkavatnet, where fluvial habitats account for 5.2 km (Figure 1). Possible visual predators on smolts inhabiting the system include gulls, otters, mink, and brown trout.
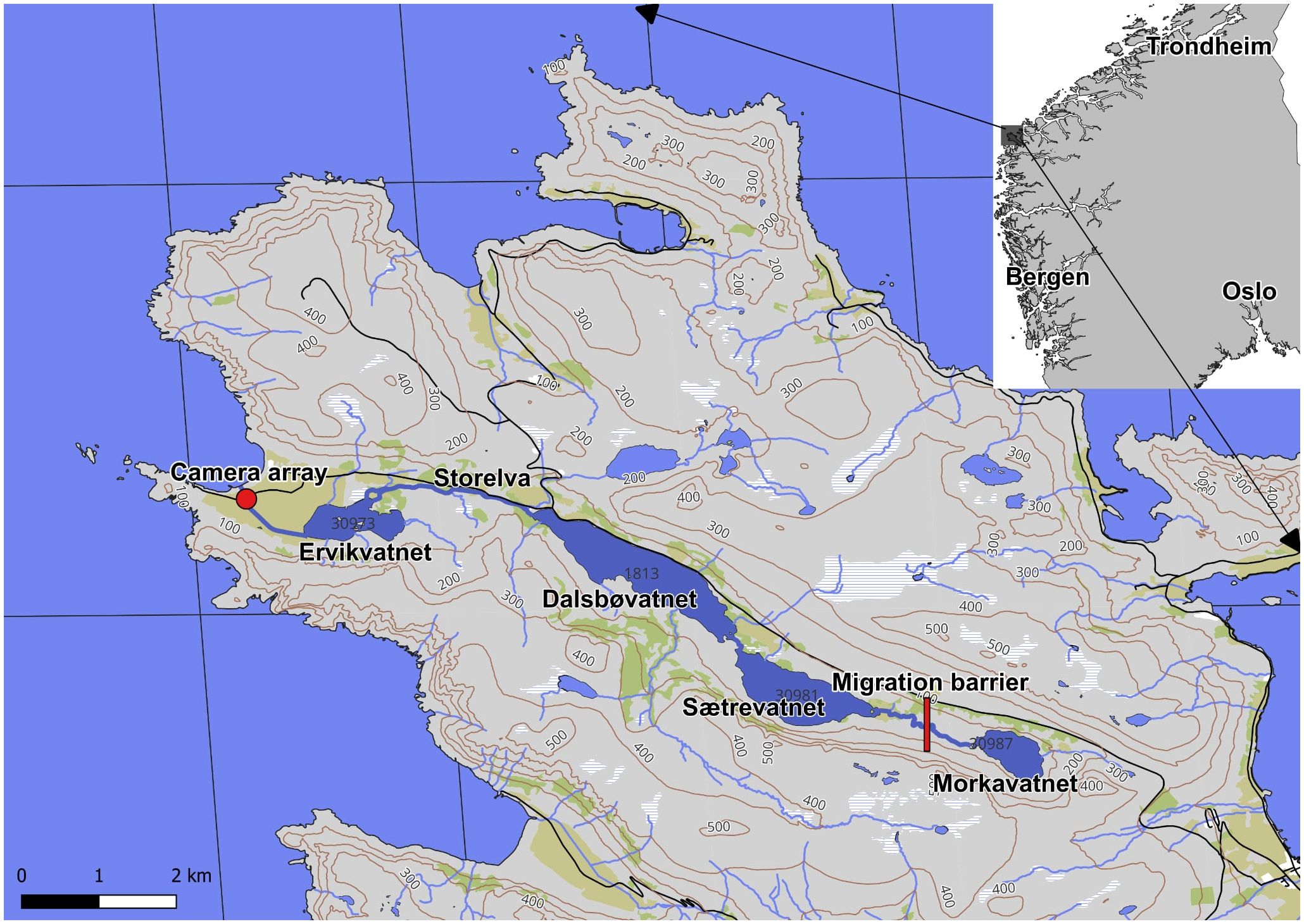
Figure 1. Map over Dalsbøvassdraget; river Ervika is located from Ervikvatnet to the sea. Main spawning and nursing area are located in Storelva. The camera array are located only 200 m above the sea, and the anadromous migration barrier is located in the river stretch between Morkavatn and Sætrevatn.
Video technology has been established as an acknowledged methodology for gathering information in fish monitoring programs during the past decades (e.g., Davidsen et al., 2005; Svenning et al., 2017), and the method relies on visual information and classification of species and life stages. The multi-camera array was placed at the same location in all consecutive years (Figure 1), and the total smolt-run period (mid-April until mid-June) was surveyed each year (2021–2023). A total of four cameras were placed in the narrowest part of the river. Camera platforms were moored to the bottom of the riverbed with an effective weight in water of 8–10 kg and with an adjacent white LED light mounted on a bracket next to the cameras. Each camera (Minicam V2 from Skynordic) had a horizontal field of view under water of 60°. The underwater lights have a beam angle of 135° and a colour temperature of 5,700 K and 1,500 lumens of light. The maximum depth at the camera array varied between 40 and 60 cm depending on water discharge, and all cameras were positioned less than 1 m apart from each other to ensure overlapping line of sight over the three subsequent cameras (Figure 2). Each camera delivered a phase alternating line (PAL) continuous stream with a resolution of 720 × 576 pixels. All cameras were set up with a hardwire connection to a stationary land base equipped with a receiver and transmitter, storing the video at 3 frames per second (FPS) for the whole duration of the experimental period in external hard drives.

Figure 2. Camera array with measured distance between camera platforms. All cameras had overlapping line of sight for maximum detection probability of passing fish. Created in https://BioRender.com.
All material was analysed through manual continuous playback, varying between 6× and 12× the actual speed. All anadromous fish passing were logged, and uncertainties regarding fish species identification of the fish passing were cross-checked by extracting still images from the video and cross-validated by multiple persons subsequently. For each detected anadromous fish, data were recorded including the date and time of passage (HH:MM:SS), species identification, camera unit responsible for detection, and the observed swimming direction. Schooling behaviour was defined as three or more fish passing the camera array within a timeframe of 12 s for comparison to earlier published data (Fernandes et al., 2015). Nevertheless, in the majority of observations, the smolts were observed within visual range of each other, and the maximum time between registered individuals within a school never exceeded 4 s.
To account for daylight, influx solar irrigation (hereafter solar radiation) was extracted for the time period in question and the Ervika location through the package “httr” (Wickham, 2023) and “jsonlite” (Ooms, 2014) in R [R version 4.4.0; R Core Team (2024)] and applied to the raw data on an hourly basis. Solar radiation in this context is the global solar irrigation (direct + diffuse) measured in kW per square meter. Minimum and maximum values throughout the time series for the Ervika location were 0 and 801.1 kW m−2, respectively, where solar radiation levels follow, on average, a bell-shaped curve during the day with the peak at noon (mean = 449.5 kW m−2) when the sun reaches its highest point (see Appendix Figure A2). Solar radiation is therefore used as a proxy for daylight. For the whole time series, average solar radiation was 173.8 kW m−2. Water discharges were readily available from an NVE station (Norwegian Water Resources and Energy Directorate) in Dalsbøvassdraget, and water temperatures were collected by HOBO loggers attached to the camera platforms, both logging on an hourly basis.
Statistical analysis
To assess differences in timing of downstream migration between Atlantic salmon and sea trout across multiple years, daily counts were transformed into proportional values within species–year–groups to ensure the focus on relative timing patterns with migration dynamics independent of total abundance. Dates were assigned a day-of-year value (Julian date). To account for temporal distribution of migration events, each date was converted to day-of-year format and weighted by its proportional contribution to the total migration. A weighted dataset was constructed by replicating day-of-year values according to their proportional weights, simulating the temporal distribution of migration events. An analysis of variance (ANOVA) was performed to test for effects on species, year, and interaction effects between species and year on migration dates.
To evaluate how environmental variables on an hourly scale influenced the probability of smolt migrating in schools, we fitted a generalised linear mixed-effects model (GLMM) using the glmmTMB package in R (Brooks et al., 2017). Each fish migrating past the camera array was classified as either schooling or solitary, meaning that the response variable “schooling” was binary. To account for potential non-linear effects of solar radiation on schooling behaviour, we modelled solar radiation influx (kW m−2) using a natural cubic spline with four degrees of freedom (df = 4) from the “splines” package in R (Bates and Venables, 2014). Natural splines were chosen because they provide smooth, flexible fits while constraining the function to be linear beyond the boundary knots, reducing the risk of overfitting at the extremes of the data range (Hastie and Tibshirani, 1990). Increasing the degrees of freedom beyond four did not substantially improve model fit [as assessed by the Bayesian information criterion (BIC)] and risked overfitting, while fewer degrees of freedom failed to capture the relationship between solar radiation and schooling probability.
Model selection was performed using a backward stepwise procedure based on the BIC, which penalises model complexity more strongly than the Akaike information criterion (AIC). The full model included fish species (Atlantic salmon vs. sea trout), solar radiation influx (modelled as a spline), river discharge (m³ s−1), and interaction terms between species and both environmental variables. At each step, the variable or interaction contributing least to model performance (as measured by BIC) was removed, and the process continued until no further improvement in BIC was achieved. The final model retained species and solar radiation as main effects, while water discharge and the interaction terms were excluded, indicating that these terms did not substantially improve model fit relative to their complexity. Year was included as a random intercept to account for repeated observations within years and unmeasured annual differences. Therefore, the final model included the predictors species and solar radiation influx with year as a random intercept in the GLMM. The model used a binomial error distribution with a logit link. Model assumptions were evaluated using the DHARMa package (Hartig, 2025). Normality of residuals was assessed via a Kolmogorov–Smirnov test on simulated residuals (p = 0.394), variance homogeneity was checked using a dispersion test (p = 0.82), an outlier test showed no residual outliers with extreme values (p = 0.69), and a visual inspection of residuals versus predicted values showed no heteroscedasticity in the residuals (Appendix Figure A1). These diagnostics indicated no significant deviations from model assumptions. The model equation for the final model was:
where is the probability of being in a school for observation i, is the intercept, is the effect of trout compared to salmon, is the natural spline-transformed effect of solar radiation, and is the random intercept for year.
Results
During the entire study period, the visibility was consistently greater than 1.5–2 m; therefore, at least two cameras had an overlapping field of view to ensure a continuous visual coverage of the full width and depth of the water column. A total of 19,238 individual smolts were registered over the period 2021-2023 (Table 1), with the median time between observations of schools and single fish varied between 11.9 and 12.8 min (see Appendix Table A1). The migration period had a relatively short time span in all years, with significant variations between species and year, where the first smolts were observed passing the camera array on 18, 19, and 15 April and with the last detection of smolt migration on 15, 11, and 14 June in 2021, 2022 and 2023, respectively. An accumulation of 90 % of all smolts passed the video arrays within 28 May (2021), 24 May (2022), and 4 June (2023) for Atlantic salmon and 20 May (2021), 24 May (2022), and 3 June (2023) for sea trout (Figure 3).
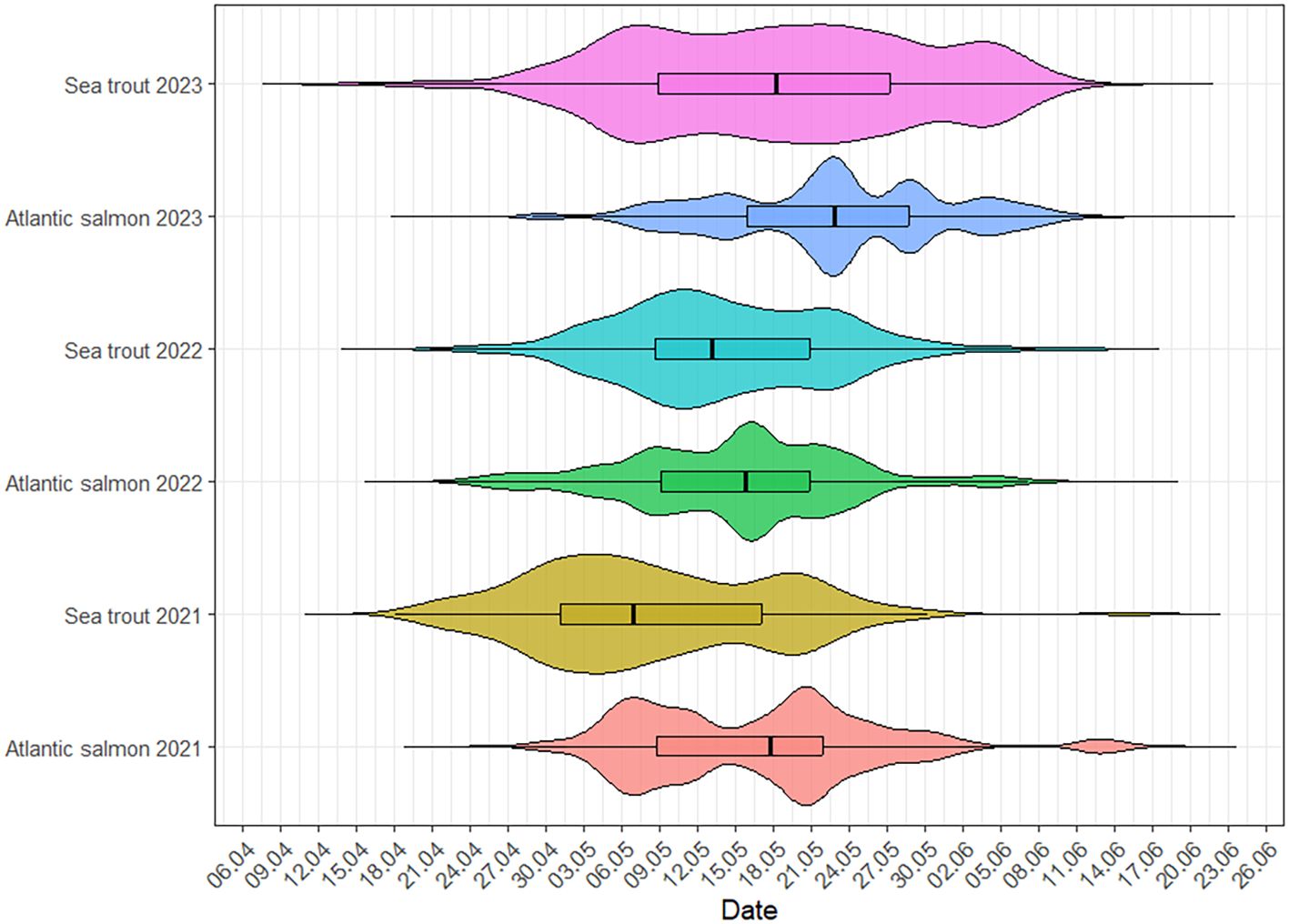
Figure 3. Distribution of arrival dates to the camera sector for Atlantic salmon and sea trout smolts by year. Each horizontal violin represents the distribution of individual arrival date times for a given species and year. The width of each violin reflects the density of arrivals (wider sections indicate more arrivals at that time). The embedded boxplot within each violin shows the interquartile range (IQR; the middle 50% of the data), with the thick horizontal line indicating the median arrival date time. Whiskers extend to the most extreme data points within 1.5 × IQR from the box.
The timing of downstream migration differed significantly, both between species and years. A Levene’s test revealed significant heterogeneity of variance across groups (p < 0.005), violating the assumptions of the ANOVA test. To account for this, a Welch’s ANOVA was used and confirmed significant differences in migration across species–year combination (F = 27.49, p < 0.001). A Kruskal–Wallis test supported this result, indicating significant differences in the distribution of migration timing (χ2 = 113.11, p < 0.001). The results demonstrated significant differences both between species, year, and interaction and between year and species. A post-Tukey HSD was performed to investigate all years, and the only year without a significant effect on migration timing between species was 2022 (Tukey HSD; p = 0.97) (Figure 3, Appendix Tables A2-A5).
Across all years, a total of 1,467 schools of fish were recorded: there were 1,014 (69.1 %) schools of Atlantic salmon, 102 (7.0 %) schools of sea trout, and 351 (23.9 %) mixed-species schools. Results reveal a diel pattern in schooling behaviour for both species, where a total of 76.9 %–88.8 % of all Atlantic salmon observations and 41.7 %–57.8 % of all sea trout observations were classified to exhibit schooling behaviour in 2021–2023 (Figure 4, Table 1).
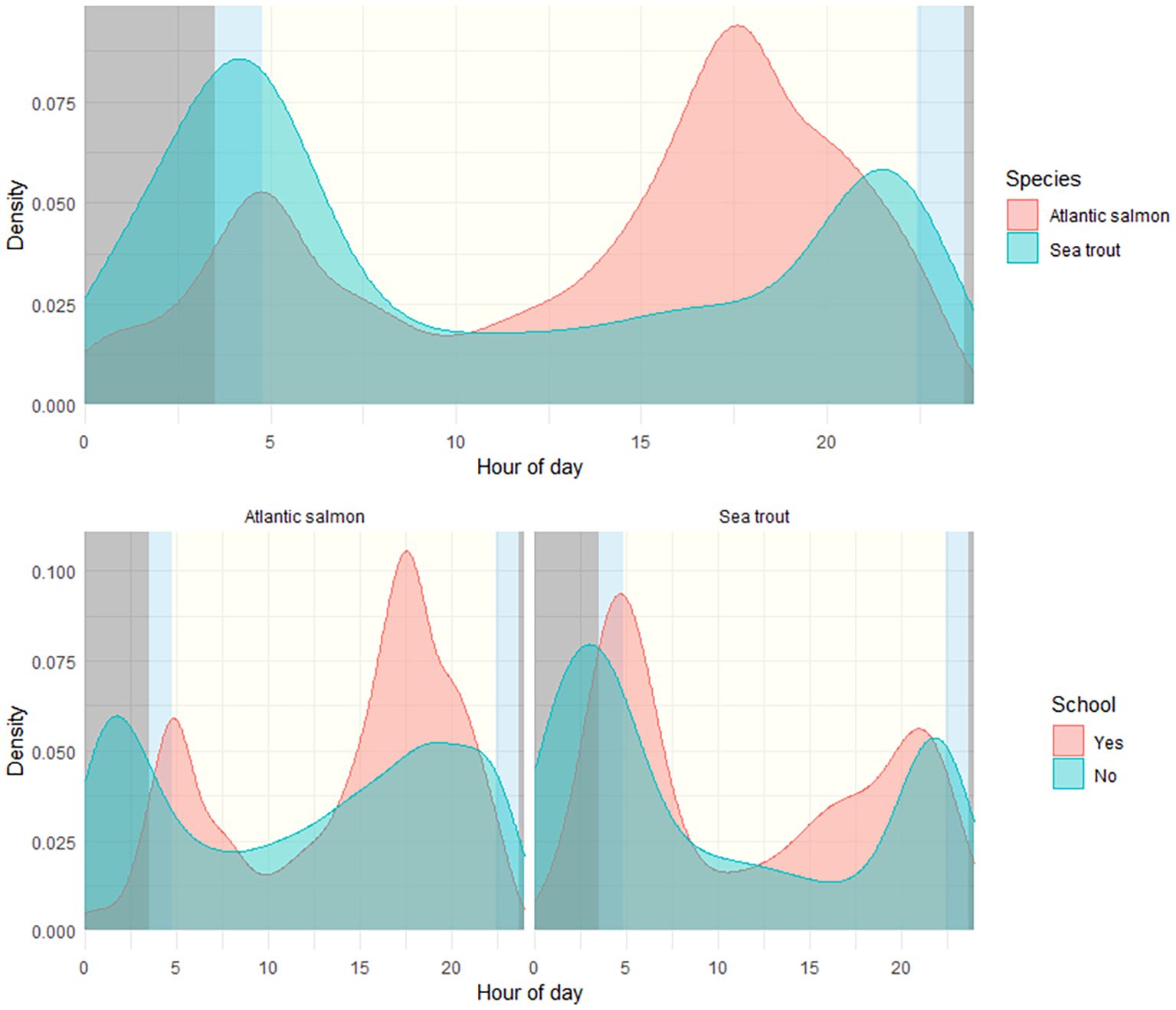
Figure 4. Observed diel migration patterns for all fish in 2021–2023, all raw-data combined. Density represents portions of the total number of observed fish in each separate category. The upper panel shows total portions of all passages for Atlantic salmon and sea trout; all fish included for all years. The lower panel shows schooling vs. single fish passages for Atlantic salmon (left) and sea trout (right). Daylight times were derived using the “suncalc” (Thieurmel and Elmarhraoui, 2022) package in R for May 15, representing approximately 50% cumulative migration across all years.
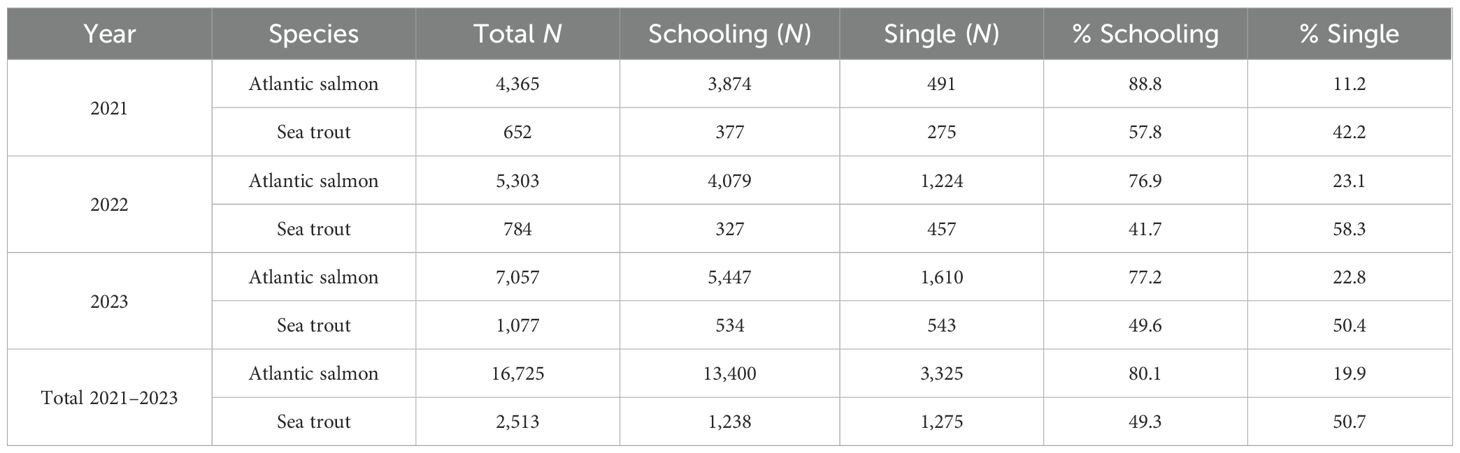
Table 1. Total numbers of registered smolts categorised as “schooling” and “single” fish passages in River Ervika 2021–2023.
The observed schooling behaviour correlates with daylight and reveals a bimodal trend where higher densities of fish passages are pronounced around dusk for Atlantic salmon, whereas sea trout have higher densities of fish passages during night/dawn (Figure 4).
The final GLMM included the predictors species and solar radiation modelled as a natural cubic spline with four degrees of freedom (BIC = 19,159.7), with year as a random intercept in the GLMM. Model selection excluded the interaction between discharge and species (BIC = 19,223.3), the interaction between solar radiation and species (BIC = 19,175.4), and discharge as a main effect (BIC = 19,172.6). Results from the GLMM analysis reveal that the probability of migrating together with other fish, “schooling”, differed significantly between salmon and sea trout smolts. Disregarding environmental factors, but keeping year as a random intercept, salmon smolts had 81% probability and sea trout had only 51% probability of migrating in a school. This corresponds to 77 % lower odds of schooling in trout compared to salmon (odds ratio = 0.23, t = −32.35, p < 0.001). Including solar radiation as a natural spline (df = 4) significantly improved model fit compared to a linear effect (likelihood ratio test: χ2 = 642.3, df = 3, p < 0.001), demonstrating a non-linear relationship between solar radiation and probability of schooling for both species (Figure 5). While the relationship between solar radiation and schooling was not linear, increased solar radiation promoted schooling, but the effect was strongest at intermediate levels of solar radiation. For the highest levels of solar radiation, the effect on schooling reached a plateau (Figure 5). The random intercept for year had a variance of 0.15, indicating modest to moderate annual variation (see Appendix Table A6 for model output results).
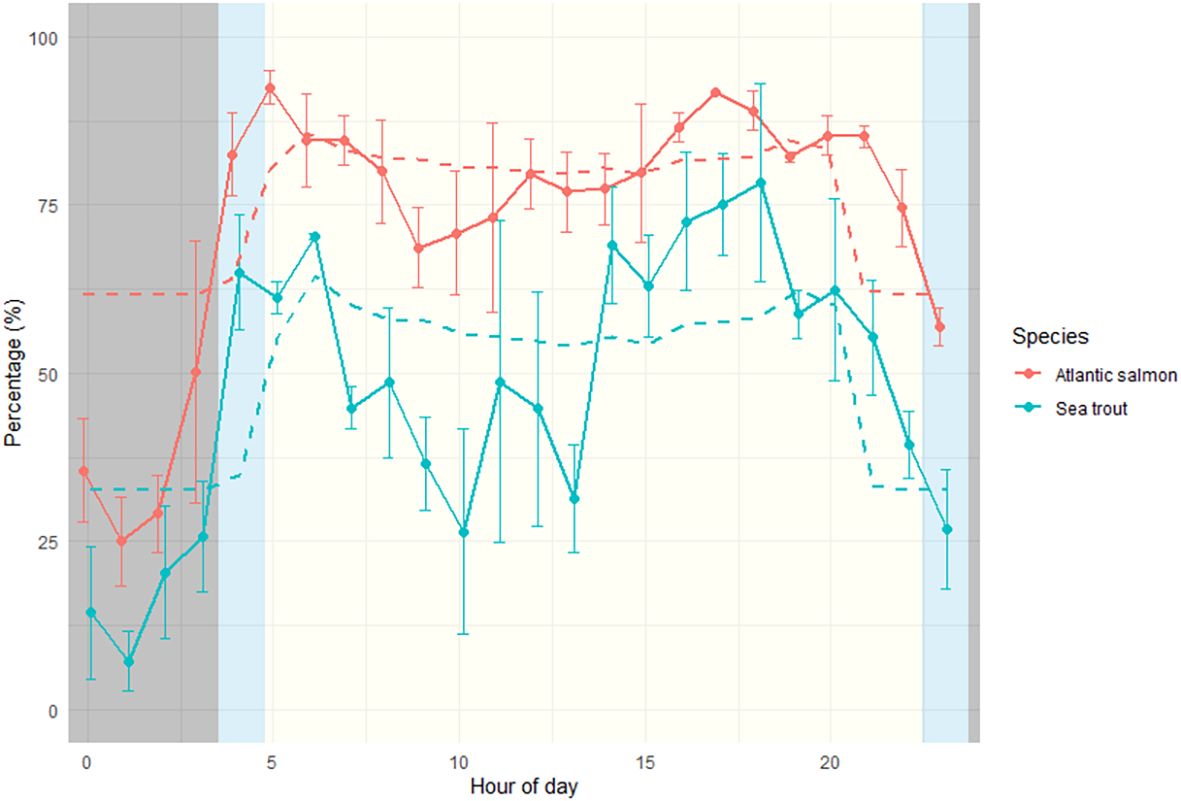
Figure 5. Observed (solid lines) and predicted probability (dashed lines) of schooling by hour of day for Atlantic salmon (red) and sea trout (blue) smolts. Observed percentages represent average values across years 2021–2023 for each species, with whiskers showing standard error (SE) between years. Predictions are based on the generalised linear mixed-effects model results. Beige, blue, and grey background shading indicate daytime, dawn/dusk, and night, respectively. Daylight times were derived using the “suncalc” package in R for May 15, representing approximately 50 % cumulative migration across all years.
Analyses on solar radiation reveals a fixed diel pattern for all years, where hours with no solar influx have dominantly more single fish passages as also predicted by the GLMM. This pattern breaks up during dawn when fish passages for both species predominantly are together with others (schooling). Sea trout migrated more often during hours with less solar influx and dawn compared to Atlantic salmon, which favoured dusk, and generally had less migration activity during total darkness compared to sea trout. Nevertheless, diel patterns with less midday movement and prominently less “schooling” and more single fish detections during nighttime were observed for both species (Figure 5).
Discussion
The results from River Ervika offer a novel insight into the diel migration patterns in sympatric salmonids and reveal a correlation between schooling behaviour and natural light regimes, and its effect on the migratory timing during seaward migration for Atlantic salmon and sea trout smolts. The observed pattern describes the migration dynamics for sympatric natural stocks of Atlantic salmon and sea trout over a 3-year period with little-to-no interferences with natural behaviour.
Results indicate significant differences in the timing of the “smolt run” between species within years, where sea trout migrated earlier than Atlantic salmon in 2021 and 2023. There were no significant differences, but the same trend in the data was observed in 2022. Significant variation between years was also detected for both species. The timing of the “smolt run” and the effects of environmental triggers have been thoroughly studied during the last decades, and environmental factors such as photoperiod, temperature, water discharge, and turbidity have shown to be amongst the most pronounced effects triggering migration (Jonsson and Ruud-Hansen, 1985; McCormick et al., 1998; Urke et al., 2013; Harvey et al., 2020; Bjerck et al., 2021). The timing of migration between species considering day of year have often shown that Atlantic salmon are somewhat earlier than sea trout, but with considerable overlap between species (e.g., Aldvén et al., 2015; Haraldstad et al., 2017). Nonetheless, there are also examples where sea trout smolt start their migration period before Atlantic salmon (Sortland et al., 2025); thus, the results from River Ervika are formally recognisable compared to earlier work and further establish and strengthen the hypothesis of coinciding migration patterns for sympatric Atlantic salmon and sea trout.
Concerning migration behaviour, analyses disclose a diel pattern in schooling behaviour, with clear species-specific differences in migratory behaviour concerning time of day. Mixed-species schools were frequently observed to contain higher proportions of Atlantic salmon compared to sea trout. Observations within these schools suggest that sea trout individuals typically occupied positions toward the rear of the school, often passing the camera array as the last individuals. In addition, Atlantic salmon exhibited a more direct and coordinated movement, suggesting a decisive migratory strategy. In contrast, sea trout displayed less structured behaviour, occasionally lingering or moving erratically within the school before passing the camera array, but within the observed school. These observations could indicate species-specific differences in schooling dynamics and migratory decisiveness, which may reflect underlying ecological or behavioural adaptation traits. The shift from territorial behaviour to formation of schools in salmonid fish such as Atlantic salmon and sea trout is associated with the transition from parr to the smolt migratory life stage, influenced by habitat conditions, water temperature, and food availability (Kasumyan and Pavlov, 2023). Okasaki et al. (2020) also found that Chinook salmon (Oncorhynchus tshawytscha) exhibited increased efficiency in navigation fishways at manmade dams when migrating in schools rather than as a single fish, suggesting that the conspecific social cues could facilitate better route identification for safe passages. Nevertheless, in the same study, there was no evidence that the sockeye salmon (Oncorhynchus nerka) had higher passing success; thus, interspecific differences in salmonids must be expected (Okasaki et al., 2020).
Results from the current study show a correlation between schooling behaviour and influx of natural light, where “single fish” are more prone to be detected during darker hours. However, there were discrepancies between observed and predicted values (GLMM) during the darkest hours of the night, where the observed probabilities for schooling were lower than predicted from the model. Even though the model showed a good fit to the data, a model will always be a simplification of ecological reality. Several likely possibilities could explain these differences: first, data on global solar radiation were extracted for the coordinates matching River Ervika, but these are modelled data and not accurate measurements on site, meaning that there could be a difference between actual solar radiation and modelled values. In addition, other environmental and ecological factors that promote or discourage schooling behaviour in salmonid smolts but not measured or modelled here could play a part in addition to solar radiation. The model prediction showed that schooling probability increased with increasing solar radiation, but the effect plateaued for the highest values of solar radiation for both species. The observed values support these findings, which indicate a threshold in solar radiation that does not promote additional probability of schooling. In a biological sense, it could be interpreted such that solar radiation or light influx acts as a factorial variable of “light or dark” instead of a continuous variable, with a linear effect on schooling probability only during the transition between “light and dark” periods.
Species-specific differences reveal a bimodal trend for passing the camera location during dawn/dusk and darker periods rather than in full natural sunlight, where sea trout are more prone to pass at dawn and Atlantic salmon are more prone to pass in the afternoon/at dusk. This migration regime breaks up in late May when daylight length increases, and the darkest hours of the night could only be considered as dusk or dawn. These results are in accordance with earlier research where the diel patterns become less prominent later in the migration period with longer natural daylight regimes (Thorstad et al., 2012; Haraldstad et al., 2017). The observed changes in migration dynamics could possibly be adaptations in anti-predator behaviour during river migration, where night/dark offers protection from visual predators, whereas safety in numbers aids predator detection and avoidance during daytime. Previous research has demonstrated that Atlantic salmon smolts are more likely to migrate in schools during daylight hours than at night (Riley et al., 2014; Simmons et al., 2021), concluding that it is an adaptive response to predation risk, where darkness offers protection for solitary individuals from visually oriented predators, while the group migration during the day enhances vigilance and safety through collective behaviour. The current data collected and analysed further support this hypothesis and contribute to a deeper understanding of one of the most vulnerable life history stages in anadromous salmonids. Moreover, results from River Ervika reveal species-specific differences in diel migration patterns, with salmon predominantly migrating in the afternoon and evening, while sea trout show a stronger preference for migration during the morning hours. Such interspecific variation in sympatric Atlantic salmon and sea trout has been poorly documented in previous literature. The observed species composition has remained stable throughout the 3-year monitoring period in River Ervika, where 87 % of all registered smolts in 2023 were Atlantic salmon, compared to 81 % in 2020, and 87 % in both 2021 and 2022 (Ulvund et al., 2025). The consistently high smolt counts observed in the current study suggest that the river may be approaching its maximum production potential, given its size, and that recruitment is strong with good survival from egg to smolt. These are indicators of a system with a production potential in equilibrium and, thereby, the consistency of the observed pattern can be seen as a natural behavioural trait for both species.
Schooling of migrating salmonids has been documented in earlier research (e.g., Riley et al., 2014; Simmons et al., 2021) but not as extensively as in the current study, in terms of number of individuals and consistency. In addition, the current study collected observational data with a method with little-to-no interferences with the study objects’ natural behaviour, compared to tagging studies and studies using fish traps, where tagging effects and the typical laminar flow over fish traps could affect the natural behaviour of the fish. Data collection and different methodologies for observation and remote logging of animal behaviour have weaknesses, strengths, and limitations. In the case of video surveillance as a method for detecting the presence of fish and observing their behaviour, the most profound limiting factor is visibility in the water column. Nonetheless, the consistency and the precision of the data collected over multiple years in River Ervika suggest that video surveillance as a method works well in small rivers. Size differences among individuals can influence migratory behaviour, particularly through size-dependent predation risk (Ibbotson et al., 2011). In the present study, although measurements indicated negligible size variation between Atlantic salmon and sea trout smolts when comparing individuals migrating in mixed schools to those observed in single-fish frames, these estimates are subject to inherent uncertainties in size determination. Nevertheless, the limited observed variation suggests that potential confounding effects of body size on behaviour and seaward migration were likely minimal. Consequently, the behavioural patterns reported are more plausibly driven by ecological or environmental factors rather than size-related vulnerability. The bimodal migration patterns observed in River Ervika are a poorly documented and understood phenomenon, and further understanding of the migration patterns and schooling behaviour could assist in improved management plans through facilitating better migration possibilities in rivers with anthropogenic challenges such as power plants with better practices for bypass flow for fish migration concerning natural light regimes.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
JU: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. AS: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. RS: Conceptualization, Data curation, Funding acquisition, Methodology, Validation, Writing – original draft, Writing – review & editing, Project administration. AL: Conceptualization, Data curation, Funding acquisition, Methodology, Supervision, Validation, Writing – original draft, Writing – review & editing, Project administration.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. The authors declare that this study received funding from SalmonTracking 2030, Vestland Fylkeskommune (County) and FHF - Norwegian Seafood Research Fund. The smolt monitoring project in Ervikelva was financed by the research project Salmon Tracking 2030 (www.salmontracking.no).
Acknowledgments
We would like to express our gratitude to Øyvind K. Hanssen for valuable feedback on the manuscript. We also thank the two reviewers for their constructive comments and valuable suggestions, which greatly improved the quality and clarity of this manuscript. We appreciate local aid in the maintenance of the camera array from Svein P. Ervik and Gry H. Strandvik.
Conflict of interest
Authors JU, AS, RS and AL were all employed by the company Scandinavian Nature Surveillance a DNV company (Det Norske Veritas).
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1716743/full#supplementary-material
References
Aldvén D., Degerman E., and Höjesjö J. (2015). Environmental cues and downstream migration of anadromous brown trout (Salmo trutta) and Atlantic salmon (Salmo salar) smolts. Boreal Environ. Res. 20, 35–44. doi: 10.60910/zc9u-w5ck
Bates D. and Venables W. N. (2025). splines: Regression Spline Functions and Classes (Version 4.6.0) (R Foundation for Statistical Computing). Available online at: https://stat.ethz.ch/R-manual/R-devel/library/splines/html/splines-package.html (Accessed October 30, 2025).
Bjerck H. B., Urke H. A., Haugen T. O., Alfredsen J. A., Ulvund J. B., and Kristensen T. (2021). Synchrony and multimodality in the timing of Atlantic salmon smolt migration in two Norwegian fjords. Sci. Rep. 11. doi: 10.1038/s41598-021-85941-9
Brooks M. E., Kristensen K., van Benthem K. J., Magnusson A., Berg C. W., Nielsen A., et al. (2017). glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 9, 378–400. doi: 10.32614/RJ-2017-066
Davidsen J., Svenning M.-A., Orell P., Yoccoz N., Dempson J. B., Niemelä E., et al. (2005). Spatial and temporal migration of wild Atlantic salmon smolts determined from a video camera array in the sub-Arctic River Tana. Fisheries Res. 74, 210–222. doi: 10.1016/j.fishres.2005.02.005
Fernandes W. P. A., Ibbotson A. T., Griffiths S. W., Maxwell D. L., Davison P. I., and Riley W. D. (2015). Does relatedness influence migratory timing and behaviour in Atlantic salmon smolts? Anim. Behav. 106, 191–199. doi: 10.1016/j.anbehav.2015.06.006
Gauld N. R., Campbell R. N. B., and Lucas M. C. (2013). Reduced flow impacts salmonid smolt emigration in a river with low-head weirs. Sci. Total Environ., 458–460. doi: 10.1016/j.scitotenv.2013.04.063
Haraldstad T., Kroglund F., Kristensen T., Jonsson B., and Haugen T. O. (2017). Diel migration pattern of Atlantic salmon (Salmo salar) and sea trout (Salmo trutta) smolts: an assessment of environmental cues. Ecol. Freshw. Fish 26, 541–551. doi: 10.1111/eff.12298
Hartig F. (2025). DHARMa: Residual Diagnostics for Hierarchical (Multi-Level / Mixed) Regression Models (Version 0.4.7). Available online at: https://CRAN.R-project.org/package=DHARMa (Accessed October 30, 2025).
Harvey A. C., Glover K. A., Wennevik V., and Skaala Ø. (2020). Atlantic salmon and sea trout display synchronised smolt migration relative to linked environmental cues. Sci. Rep. 10. doi: 10.1038/s41598-020-60588-0
Hastie T. J. and Tibshirani R. J. (1990). Generalized additive models (New York, NY: Chapman and Hall).
Hawley K. L., Urke H. A., Kristensen T., and Haugen T. O. (2024). Balancing risks and rewards of alternate strategies in the seaward extent, duration and timing of fjord use in contemporary anadromy of brown trout (Salmo trutta). BMC Ecol. Evol. 24. doi: 10.1186/s12862-023-02179-x
Hoar W. S. (1988). “The physiology of smolting salmonids,” in Fish Physiology, Vol. XIB. Eds. Hoar W. S. and Randall D. J. (Academic press, New York), 275–343.
Holcombe E. F., Borsky A. J., Biron J. M., Bentley P. J., and Sanford B. P. (2019). Detection of PIT-tagged juvenile salmonids migrating in the Columbia River Estuar (Portland, Oregon: Bonneville Power Administration).
Hussey N. E., Kessel S. T., Aarestrup K., Cooke S. J., Cowley P. D., Fisk A. T., et al. (2015). Aquatic animal telemetry: A panoramic window into the underwater world. Science 348. doi: 10.1126/science.1255642
Ibbotson A. T., Beaumont W. R. C., and Pinder A. (2011). A size-dependent migration strategy in Atlantic salmon smolts: Small smolts favour nocturnal migration. Environ. Biol. Fishes 92, 544–551. doi: 10.1007/s10641-011-9824-0
Ibbotson A. T., Beaumont W. R. C., Pinder A., Welton S., and Ladle M. (2006). Diel migration patterns of Atlantic salmon smolts with particular reference to the absence of crepuscular migration. Ecol. Freshw. Fish 15, 544–551. doi: 10.1111/j.1600-0633.2006.00194.x
Jensen A. J., Finstad B., Fiske P., Hvidsten N. A., Rikardsen A. H., and Saksgård L. (2012). Timing of smolt migration in sympatric populations of Atlantic salmon (Salmo salar), brown trout (Salmo trutta), and Arctic char (Salvelinus alpinus). Can. J. Fisheries Aquat. Sci. 69, 711–723. doi: 10.1139/f2012-005
Jonsson B. and Ruud-Hansen J. (1985). Water temperature as the primary influence on timing of seaward migrations of atlantic salmon (Salmo salar) smolts. Can. J. Fisheries Aquat. Sci. 42, 593–595. doi: 10.1139/f85-076
Kasumyan A. O. and Pavlov D. S. (2023). Development of schooling behavior in fish ontogeny. J. Ichthyology 63. doi: 10.1134/S0032945223070111
Marras S. and Domenici P. (2013). Schooling fish under attack are not all equal: some lead, others follow. PloS One 8. doi: 10.1371/journal.pone.0065784
McCormick S. D., Hansen L. P., Quinn T. P., and Saunders R. L. (1998). Movement, migration, and smolting of Atlantic salmon (Salmo salar). Can. J. Fisheries Aquat. Sci. 55, 77–92. doi: 10.1139/d98-011
Newton M., Barry J., Lothian A., Main R., Honkanen H., Mckelvey S., et al. (2021). Counterintuitive active directional swimming behaviour by Atlantic salmon during seaward migration in the coastal zone. ICES J. Mar. Sci. 78, 1730–1743. doi: 10.1093/icesjms/fsab024
Okasaki C., Keefer M. L., Westley P. A. H., and Berdahl A. M. (2020). Collective navigation can facilitate passage through human-made barriers by homeward migrating Pacific salmon. Proc. R. Soc. B 287. doi: 10.1098/rspb.2020.2137
Ooms J. (2014). The jsonlite Package: A Practical and Consistent Mapping Between JSON Data and R Objects. Available online at: https://cran.r-project.org/web/packages/jsonlite/index.html (Accessed October 30, 2025).
R Core Team (2024). R: A Language and Environment for Statistical Computing (Vienna, Austria: R Foundation for Statistical Computing). Available online at: https://www.R-project.org/ (Accessed October 30, 2025).
Riley W. D., Ibbotson A. T., Maxwell D. L., Davison P. I., Beaumont W. R. C., and Ives M. J. (2014). Development of schooling behaviour during downstream migration of Atlantic salmon Salmo salar in a chalk stream. J. Fish Biol. 85, 1042–1059. doi: 10.1111/jfb.12457
Simmons O. M., Gregory S. D., Gillingham P. K., Riley W. D., Scott L. J., and Britton J. R. (2021). Biological and environmental influences on the migration phenology of Atlantic salmon Salmo salar smolts in a chalk stream in southern England. Freshw. Biol. 66, 1581–1594. doi: 10.1111/fwb.13776
Sortland L. K., Aarestrup K., and Birnie-Gauvin K. (2025). Comparing the migration behavior and survival of Atlantic salmon (Salmo salar) and brown trout (Salmo trutta) smolts. J. Fish Biol. 106, 1360–1376. doi: 10.1111/jfb.15749
Svenning M.-A., Lamberg A., Dempson B., Strand R., Hanssen Ø.K., and Fauchald P. (2017). Incidence and timing of wild and escaped farmed Atlantic salmon (Salmo salar) in Norwegian rivers inferred from video surveillance monitoring. Ecolology Freshw. Fish 26, 360–370. doi: 10.1111/eff.12280
Thieurmel B. and Elmarhraoui A. (2022). suncalc: Compute Sun Position, Sunlight Phases, Moon Position and Lunar Phase (R package version 0.5.1). Available online at: https://cran.r-project.org/web/packages/suncalc/index.html (Accessed October 30, 2025).
Thorstad E. B., Whoriskey F., Rikardsen A. H., and Aarestrup K. (2010). “Aquatic nomads: The life and migrations of the Atlantic salmon,” in Atlantic Salmon Ecology. Ed. Aas Ø., et al (West Sussex, UK: Wiley-Blackwell), 1–32. doi: 10.1002/9781444327755.ch1
Thorstad E. B., Whoriskey F., Uglem I., Moore A., Rikardsen A. H., and Finstad B. (2012). A critical life stage of the Atlantic salmon Salmo salar: behaviour and survival during the smolt and initial post-smolt migration. J. Fish Biol. 81, 500–542. doi: 10.1111/j.1095-8649.2012.03370.x
Thorstad E. B., Rikardsen A. H., Alp A., and Økland F. (2013). The use of electronic tags in fish research–an overview of fish telemetry methods. TRJFAS 13, 881–896. doi: 10.4194/1303-2712-v13_5_13
Ulvund J. B., Smalås A., and Lamberg A. (2025). Videoovervåking av utvandring, innsig og vandringsforløp av anadrom laksefisk i Ervikelva i 2023, SNA-rapport 07/2025–25 s. (Trondheim: Scandinavian Nature Surveillance).
Urke H. A., Kristensen T., Ulvund J. B., and Alfredsen J. A. (2013). Riverine and fjord migration of wild and hatchery-reared Atlantic salmon smolts. Fisheries Manage. Ecol. 20, 544–552. doi: 10.1111/fme.12042
Vollset K. W., Lennox R. J., Thorstad E. B., Auer S., Bär K., Larsen M. H., et al. (2020). Systematic review and meta-analysis of PIT tagging effects on mortality and growth of juvenile salmonids. Rev. Fish Biol. Fisheries 30, 553–568. doi: 10.1007/s11160-020-09611-1
Wickham H. (2023). httr: Tools for Working with URLs and HTTP (R package version 1.4.7). Available online at: https://cran.r-project.org/web/packages/httr/index.html (Accessed October 30, 2025).
Keywords: smolt migration, schooling behaviour, seasonal patterns, fish behaviour, migration dynamics, camera surveillance
Citation: Ulvund JB, Smalås A, Strand R and Lamberg A (2025) Diel patterns and migration behaviour of sympatric Atlantic salmon (Salmo salar) and brown trout (Salmo trutta) smolts during seaward migration. Front. Mar. Sci. 12:1716743. doi: 10.3389/fmars.2025.1716743
Received: 30 September 2025; Accepted: 17 November 2025; Revised: 07 November 2025;
Published: 10 December 2025.
Edited by:
Tomas Chalde, CONICET Centro Austral de Investigaciones Científicas (CADIC), ArgentinaReviewed by:
Céline Artero, Muséum National d’Histoire Naturelle, FranceRyo Futamura, Leibniz-Institute of Freshwater Ecology and Inland Fisheries (IGB), Germany
Copyright © 2025 Ulvund, Smalås, Strand and Lamberg. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: John Birger Ulvund, am9obi5iaXJnZXIudWx2dW5kQGRudi5jb20=