 Daniele Santana de Sousa Oliveira1*†
Daniele Santana de Sousa Oliveira1*† Elis Dionisio da Silva2*†
Elis Dionisio da Silva2*† Fábio Farias Galvão Júnior1†
Fábio Farias Galvão Júnior1† Danielle Maria Nascimento Moura3
Danielle Maria Nascimento Moura3 Policarpo Ademar Sales-Junior3
Policarpo Ademar Sales-Junior3 Maria Carolina Accioly Brelaz-de-Castro3,4
Maria Carolina Accioly Brelaz-de-Castro3,4 Valéria Rêgo Alves Pereira3*‡
Valéria Rêgo Alves Pereira3*‡- 1Postgraduation Program in Therapeutic Innovation, Federal University of Pernambuco, Recife, Pernambuco, Brazil
- 2Health and Biotechnology Institute, Federal University of Amazonas, Coari, Amazonas, Brazil
- 3Immunopathology and Molecular Biology Laboratory, Aggeu Magalhães Institute (FIOCRUZ-PE), Department of Immunology, Recife, Pernambuco, Brazil
- 4Parasitology Laboratory, Federal University of Pernambuco, Vitória de Santo Antão, Pernambuco, Brazil
Chagas disease is considered a public health issue, especially in Latin America. To this date, the course of the infection caused by the parasite Trypanosoma cruzi is yet to be completely understood and the conventional treatment do not promote a cure in the chronic phase, meaning there is an urgent need to discover new drugs. The expression of reporter genes by transgenic parasites has become an important tool in the screening of new compounds, whether in the study of the parasite, in the development of in vitro and in vivo assays, or in the application of High-Throughput Screening utilizing compounds collections. This review sought to gather information about transgenic T. cruzi applications in screening studies of compounds with action specifically against Chagas disease, the reporter genes in use, besides the highlighted characteristics of each one by the literature, including the performed assays, evolutionary forms and techniques applied, aiming to facilitate the identification of the reporter gene system or research model whose characteristics best adapt to the needs of new studies, contributing to the decisions about a framework adaptable to the reality of laboratories, in the screening of potentially trypanocidal compounds.
1 Introduction
Chagas disease (CD) caused by the parasite Trypanosoma cruzi (Chagas, 1909) (1) (Kinetoplastida, Trypanosomatidae), is a neglected disease affecting over 7 million people globally and frequently results in chronic heart conditions (2). The disease in the host progresses through two main phases: an initial acute phase characterized by high parasitemia and often few or no symptoms, followed by a chronic phase where parasitemia becomes intermittent. This chronic phase can remain asymptomatic for years or eventually lead to severe neurological and cardiodigestive complications (3, 4). Currently available treatments rely on nitroheterocyclic drugs which unfortunately present significant toxicity (5).
Preventive control of CD involves reducing human-vector contact (6), implementing blood bank screening programs (7), ensuring strict food and beverage hygiene coupled with public awareness campaigns (8), and ongoing efforts to develop vaccines to prevent/slow disease progression (6).
Post-infection control relies on pharmacological treatment, limited to two drugs: nifurtimox (commercialized as Lampit™ by Bayer HealthCare AG, Leverkusen, Germany) and benznidazole (commercialized as Benznidazol LAFEPE® in Brazil by LAFEPE, and as Abarax® in Argentina by Maprimed/ELEA) (9).
These drugs are primarily effective during the acute phase of the disease, but their multiple adverse reactions are frequently reported, leading to treatment discontinuation in approximately 20% of patients, which reinforces the urgent need for new therapeutic options (9).
Ideally, a new drug should demonstrate efficacy for all phases of CD, improved safety and tolerability compared to benznidazole, no contraindications during pregnancy or age groups, and shorter treatment regimen (10).
Achieving this goal requires overcoming significant gaps in knowledge of parasite biology, its interactions with the host immune system (11), and current lack of tools to assess treatment efficacy (12).
Methodological challenges to develop anti-T. cruzi drugs are the absence of standardized protocols for in vitro and in vivo screening and the difficulty in extrapolating animal model findings to human disease (7); also insufficient comprehension of CD pathogenesis, chronicity, and tissue tropism (13).
Romanha et al. (14) established some criteria to new compound’s identification in preclinical studies, providing decision-making steps for progression to later testing stages, facilitating data comparison across research groups (9).
Transgenic parasites expressing reporter genes were emphasized in that document (14), and since then, the importance of genetically modified parasites in drug screening has grown (15), facilitating studies on cellular signaling and gene expression (16) while enabling rapid data quantification and reducing manual labor (17).
In murine models, screening of anti-T. cruzi compounds has become faster and more efficient through the use of recombinant fluorescent or luminescent parasites (18), complemented by imaging systems (19).
This review aims to identify transgenic T. cruzi used in drug screening for CD, in vitro and in vivo, employing techniques such as imaging and High-Throughput Screening (HTS). Strains were analyzed in relation to their life stage, reporter genes, advantages and limitations of each approach, in order to provide information that may guide toward efficient, rapid, and quantitative tools for the screening of potentially trypanocidal compounds.
1.1 The reporter system
A reporter system codes for a detectable and quantifiable product in a living cell (20). For clarity, we have categorized reporter systems currently applied to T. cruzi into two groups: Enzymatic and Fluorescent (Figure 1): the enzymatic group, which includes chloramphenicol acetyltransferase (CAT), β-galactosidase (β-gal) reporters (21) and bioluminescent systems (20). The fluorescent group comprises green fluorescent protein (GFP); enhanced green fluorescent protein (EGFP); red fluorescent proteins such as Discosoma red fluorescent protein (DsRed) (20), tandem tomato (tdTomato) (22) and E2-crimson (23) (Table 1).
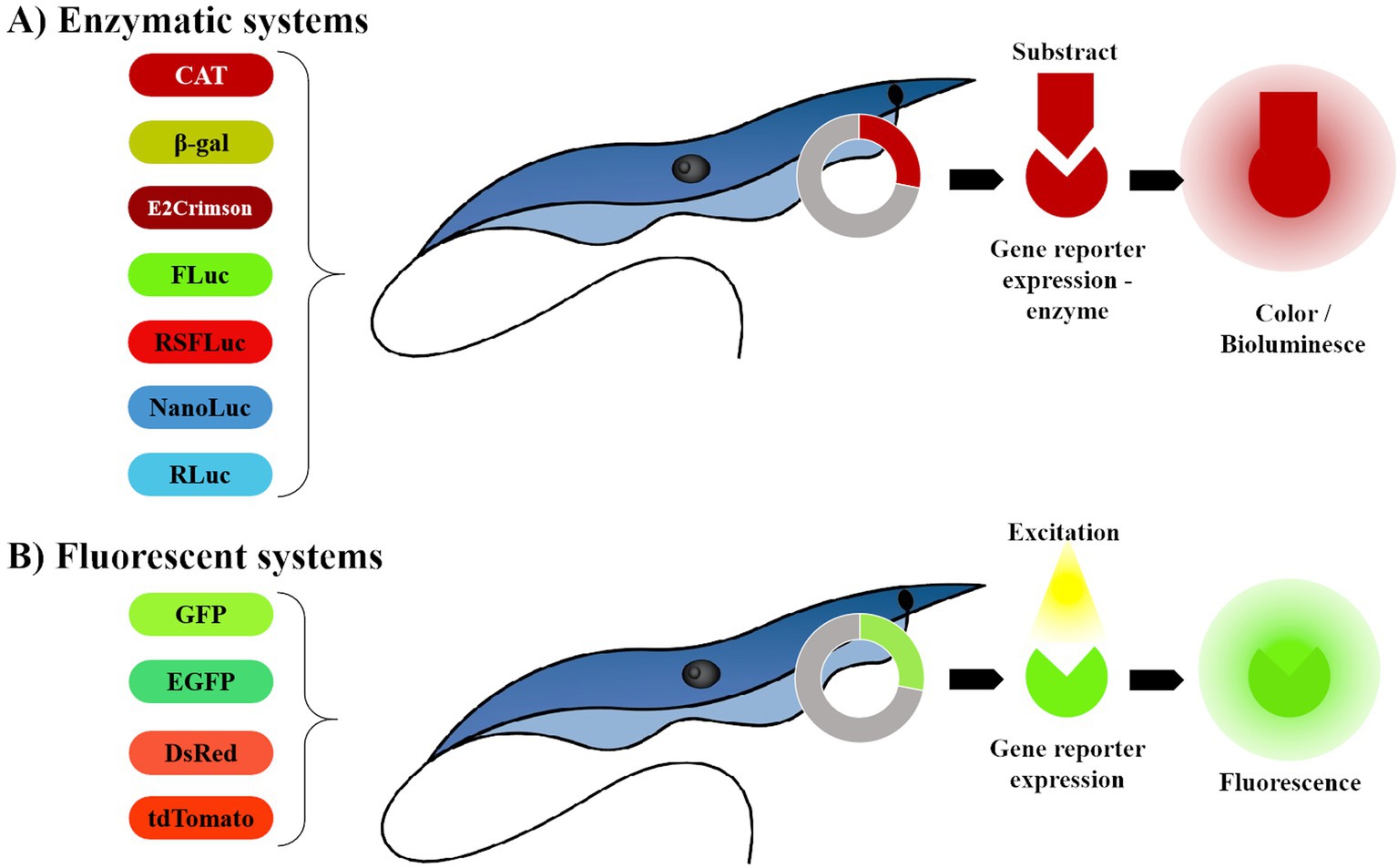
Figure 1. Representation of reporter systems categories, currently in use for anti- T. cruzi drug screening. (A) The enzymatic system depends on the enzyme reaction in the presence of the substract to express bioluminescence. (B) The fluorescent system requires no substrates, enzymes or cofactors to express fluorescence. The image was created by the authors using the following programs: AutoCAD 2024 - educational version, for drawing; WPS Office free version for assembling the figure (https://www.wps.com/) and GIMP 2.10.38 (gimp.org) for exporting with higher resolution (300dpi).
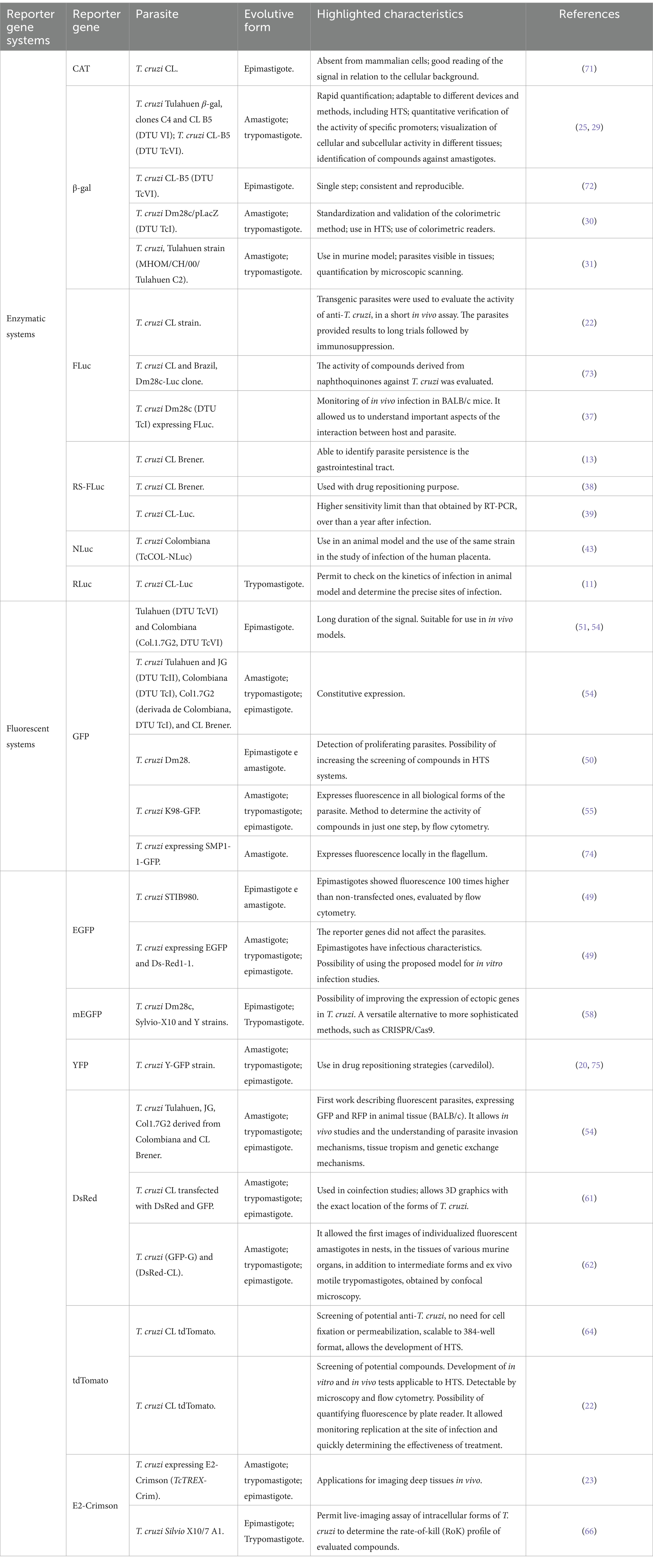
Table 1. Reporter genes systems highlighted characteristics, parasite strains and forms mentioned by the literature.
2 Enzymatic systems
2.1 Chloramphenicol acetyltransferase
Chloramphenicol acetyltransferase (CAT), the first reporter used to assess mammalian transcriptional activity of mammalian is stable and absent in mammalian cells (24). Early assays were lengthy, costly, and required radioactive tracers (20), later replaced by non-radioactive fluorescent or immunosorbent methods (24). Thus, this system proves to be inadequate for automated analyses and high-throughput screening, which are essential for the discovery of new drugs for Chagas disease.
2.2 β-galactosidase
The Escherichia coli lacZ gene encodes β-galactosidase (β-gal), the first reporter gene used in T. cruzi for in vitro screening (21). Buckner et al. (25) developed the Tulahuen β-gal strain (clone C4 and CL clone B5, DTU VI), detectable in host cells, enabling compound screening against intracellular parasite by colorimetric assay (25), thus allowing the measurement of parasite proliferation under compound exposure.
The Tulahuen strain is compatible with microplate readers assays (12) and adaptable to High-Content (26) and HTS systems (27), as demonstrated by GlaxoSmithKline HTS campaign, which identified 500 non-cytotoxic compounds (28). Broad Institute MLPCN T. cruzi Inhibition Project (29) used Tulahuen β-gal strain with the GalScreen luminescent reporter system on 303,224 compounds, identifying 4,394 hits (12).
Colorimetric reporters may interfere with enzymatic detection, but fluorometric or luminescent readings can resolve it (30). Gulin et al. (30) minimized interference by removing the supernatant before adding Chlorophenol red-β-D-galactopyranoside (CPRG) substrate and validated an in vitro assay using the transfected T. cruzi Dm28c/pLacZ strain (DTU TcI). The findings enphasized both the reliability and reproducibility of the assay and its suitability for HTS systems.
Also, β-galactosidase enables the study of parasitism throughout all CD stages in animal models: using 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside (X-gal) as a substrate, parasites become visible after fixation and blue staining, allowing quantification by microscopic scanning post-euthanasia (31). Although well-stablished and reliable, the lacZ system is limited by the requirement for cell lysis in order to assess enzymatic activity, preventing real-time analysis.
2.3 Luciferase systems
Luciferases catalyze light production by converting a supplemented substrate (16). Their efficient oxidation, bioluminescence and quantifiable activity make them valuable as reporters (32), particularly in T. cruzi transfection, where they reduce interference from colored compounds in HTS assays (12). In vivo, luciferase is ideal since animals lack endogenous activity, enabling reliable, multiple measurements. However, for identifying or quantifying reporter-expressing cells, fluorescent proteins are preferable remain more suitable (33).
The studies here presented enabled more accurate host–parasite interaction analysis in deep tissues and drug screening, and demonstrated the reporters effectiveness in assessing parasite replication. However, those systems require specific substrate addition, with possible limited bioavailability. It also requires specialized bioluminescent detection equipment.
2.3.1 Firefly luciferase
Firefly luciferase (FLuc) emerges as valuable reporter for anti-T. cruzi drug development (18) as it detects trace ATP levels and emits light efficiently through mammalian tissues (34). Although FLuc requires luciferin addition (12), it allows in vivo imaging without cell lysis, supporting whole-animal and HTS assays (21). However, its sensitivity decreases in deep tissues due to its <600 nm emission, which is absorbed by hemoglobin—a limitation that can be addressed with reporter emitting above 600 nm (35). Also, FLuc can be inhibited by small molecules structurally related to D-luciferin, such as benzothiazoles, benzimidazoles, benoxazoles, and biaryl oxadiazoles, resulting in false positives during inhibition assays (36).
Despite these limitations, significant advance has been achieved. Canavaci et al. (22) used a luciferase-expressing CL strain T. cruzi to infect Balb/c mice and developed a 12-day assay comparable to traditional 80-days protocols with immunosuppression (22), representing an advancement that allows for rapid and non-invasive assessment of the drug’s efficacy through bioluminescence.
In another work, Henriques et al. (37) tracked the progression of in vivo infection using T. cruzi (Dm28c-luc) in BALB/c mice, identifying new infection sites for the first time: the luminescent signal was observed at the inoculation site, reaching the peritoneal cavity 1 day post-infection and spreading to abdominal organs and adjacencies, shedding light on both the pathology and the interaction between parasite and host. Studies like these advanced the understanding of T. cruzi’s pathogenesis with importance for new drugs development.
2.3.2 Red-shifted firefly luciferase
Red-shifted firefly luciferase (RS-FLuc) is a variant of luciferase with enhanced sensitivity and stability, offering improved visualization in deep mammalian tissues (38); it has been used to identify T. cruzi tropisms in mice, as reported by Lewis et al. (13), who integrated the thermostable red-shifted luciferase gene into the parental CL Brener strain, allowing the monitoring of animals over a year, with a detection limit of 100 parasites, and revealed gastrointestinal tissue as primary site of parasite persistence.
The luciferase permitted to assess posaconazole’s efficacy against acute and chronic CD, Francisco et al. (38) inoculated mice with CL-Brener strain and evaluated them using an in vivo/ex vivo imaging system after an strategic use of cyclophosphamide-induced immunosuppression to uncover residual in vivo infection, in which posaconazole demonstrated inferior performance compared to benznidazole in both infection phases. In other study, Calvet et al. CL Brenner strain expressing red-shifted luciferase (T. cruzi CL-luc) which enabled the detection of live parasites in mouse tissues surpassing RT-PCR sensitivity, for over a year post-infection (39), also enabling drug efficacy tracking through different phases of the CD.
2.3.3 NanoLuc
NanoLuc (NLuc) derived from the luciferase of Oplophorus gracilirostris (A. Milne-Edwards, 1881) (40) (Decapoda, Oplophoridae) and furimazine (41), is used in in vivo Bioluminescence Imaging due to its sensitivity and intense luminescence (42), enabling multiplexing with longer-wavelength reporters (41). Colombian strain of T. cruzi expressing NLuc (TcCOL-NLuc) has been applied to study placental crossing in 3D cell culture models (43), providing insights into tissue tropism and highlighting NLuc’s utility for ADMET evaluation in drug screening, under physiologically relevant conditions.
2.3.4 Renilla luciferase
Renilla reniformis (Pallas, 1766) (44) (Scleralcyonacea, Renillidae) (RLuc) luciferase catalyzes the oxidation of coelenterazine producing “bioluminescence, coelenteramide and CO2” (32), and serves as a reporter for bioluminescent imaging in animal models; however, its sensitivity depends on the depth of the tissue investigated (45). The applications of RLuc are discussed in section 4 (Multiple Gene Reporter Systems), to which we direct the reader for further details.
3 Fluorescent systems
Green fluorescent protein (GFP) chromophore emits light without needing cofactors or substrates (46), however all fluorescent proteins photobleach under prolonged excitation, making photostability essential repetitive imaging experiments (47). Mutations in the GFP gene produce reporters with varied colors and intensities based on aminoacid sequences (48). This led to the development of T. cruzi strains enhanced green fluorescent Protein (EGFP) (49) and red fluorescent proteins, such as Discosoma striata (Corallimorpharia, Discosomidae) (DsRed) (21), tandem tomato fluorescent protein (tdTomato) (22) and E2-crimson (23) which have been applied in drug screening for CD (50). Fluorescent proteins enable real-time imaging without cell lysis but require specialized equipment and may be affected by tissue autofluorescence.
3.1 Green fluorescent protein
Encoded by a single gene, requiring no substrates, enzymes or cofactors (34), GFP is a low cost, non-toxic reporter for in vitro and in vivo imaging (23), detectable via microplate readers, fluorescence microscope, Fluorescence-Activated Cell Sorter (FACS) or fluorimetry (21). Da Rocha et al. (51) stably expressed GFP in Tulahuen (DTU TcVI) and Col.1.7G2 (DTU TcVI) epimastigotes, with persistent signal for over 5 weeks without drug selection. Despite its stability, GFP’s excitation can damage cells (52) and detection is limited to ~1 mm depth from the surface, suitable for small/transparent models (53).
Nevertheless, GFP-expressing strains like Tulahuen and JG (DTU TcII), Colombiana (DTU TcI), Col1.7G2 (DTU TcI) and CL Brener hybrids have shown infectivity in vitro and in vivo, as seen in confocal and fluorescence microscopy (54).
The pBEX/GFP strain, derived from Dm28 (DTU TcI) by Kessler et al. (50), exclusively expresses GFP in replicative forms of T. cruzi, allowing their growth tracking. Kessler et al. (50) also validated a screening method based on GFP signal intensity for HTS. Miranda et al. (55) developed the K98-GFP strain (DTU TcI) fluorescent in all T. cruzi stages facilitating single-step screening. GFP mutations have expanded emission spectra, improving signal quality and broadening application possibilities (48).
3.2 Enhanced green fluorescent protein (EGFP and mEGFP)
EGFP displays stronger fluorescence than GFP (48). Florêncio-Martínez et al. (56) used T. cruzi expressing EGFP and Ds-Red1-1 to investigate the infection process showing that reporter expression did not affect parasites infectivity. All forms infected NIH-3 T3 fibroblasts with similar kinetics, validating the model for in vitro infection studies (56).
Fesser et al. (49) monitored EGFP-expressing T. cruzi STIB980 amastigotes every 4 h for 6 days in mouse embrionic fibroblasts, using high-content imaging and. Pharmacodynamic analysis and flow cytometry revealed EGFP-expressing epimastigotes exhibited 100-fold higher autofluorescence than non-transfected cells (49). This temporal assessment of parasite growth is key in evaluating compounds effect on parasite replication, particularly as amastigotes represent the main stage for therapeutic targeting (9, 14) and epimastigotes once considered non-infective are now recognized for their potential in infection studies (57).
Niemirowicz et al. (58) further advanced the genetic engineering of T. cruzi by comparing conventional multi-mRNAs vectors to those based on 2A self-cleaving peptides for mEGFP expression in Dm28c, Sylvio-X10 and Y strains, offering a more efficient alternative to CRISPR/Cas9 for endogenous gene labeling and expanding the toolkit for reporter-based drug screening platform.
3.3 Discosoma sp. (DsRED)
Red fluorescent proteins (RFPs) expanded the imaging spectrum beyond GFP, with the first RFP exhibiting excitation/emission peaks at 555/585 nm (59). DsRed, isolated from (Discosoma sp.) (20), was the first fluorescent protein from a non-photosynthetic organism and is widely used due to its high photostability and compatibility with confocal microscopy and flow cytometry (52). Pires et al. (54) engineered T. cruzi strains expressing RFP or GFP (pROCKRFPNeo and pROCKGFPNeo vectors) in epimastigotes of Tulahuen, Col1.7G2 and CL Brener. Fluorescence remained stable for over 6 months and dual-color imaging revealed coinfection of host cells by different strains, enabling studies on invasion dynamics, tissue tropism and genetic exchange.
Usign T. cruzi CL strain coexpressing DsRed and GFP, researchers visualized parasite differentiation within Leishmania amazonensis Lainson & Shaw, 1972 (L. amazonensis) (60) revealing that metacyclic trypomastigotes developed into amastigote-like forms, failing to reach the cytosol (61). These findings highlight species-specific requirements for intracellular differentiation, offering insights into host–parasite relations that may critically interfere in drug targeting.
Ferreira et al. (62) observed T. cruzi in vivo in BALB/c or C57BL/6 mice infected with G strain (DTU TcI) parasites transfected with GFP (GFP-G) or CL strain (DTU TcVI) trypomastigotes transfected with DsRed (DsRed-CL). The method provided the first images of fluorescent amastigotes in tissue nests as well as intermediate forms and motile ex vivo trypomastigotes, visualized by confocal microscopy (62).
These studies demonstrate the potential for direct monitoring of tissue infection using DsRed reporter, a critical step in evaluating the efficacy of new drug candidates, and potential understanding of the infection, refining the design of preclinical assays.
3.4 Tandem dimeric tomato red fluorescent protein
Among the brightest and photostable fluorescent proteins, tdTomato (a DsRed variant) exhibits excitation/emission peaks at 554/581 nm (63). Bustamante et al. (64) used the T. cruzi CL tdTomato strain for anti-T. cruzi screening, highlighting its compatibility with HTS abd the advantage of not requiring cell fixation or permeabilization (64).
Canavaci et al. (22) developed a T. cruzi CL tdTomato strain constitutively expressing tdTomato suitable for in vitro and in vivo HTS. The strong fluorescent signal across all stages enabled replication monitoring and rapid treatment efficacy assessment by microscopy, flow cytometry and plate reader (22). Trypanosoma cruzi tdTomato strains allow non-invasive in vivo monitoring of parasitemia tracking and parasite distribution via fluorescent imaging (65).
3.5 E2-Crimson
E2-Crimson, a DsRed fluorescent protein is non-toxic photostable rapidly maturing, and suitable for live-animal imaging, flow cytometry and stimulated emission depletion (STED) microscopy (23). Aiming applications for imaging deep tissues in vivo infected by T. cruzi, Goyard et al. (23) developed T. cruzi Y expressing E2-Crimson fluorescence (TcTREX-Crimson), offering another tool for drug screening, particularly in the in vivo context.
In other hand, by combining an in vivo imaging assay that allows for real-time, detailed evaluation of parasite clearance without invasive techniques and the use of cardiomyocytes with nuclear EGFP, Svensen et al. (66) enabled visualization of the parasite (Tc-X10/7-E2Crimson) and host interaction as well as the effect of the compound on target cells, leading to a more precisa, efficient drug screening.
4 Multiple gene reporter systems
Reporter systems with different substrates, spectra and emission kinetics can be used simultaneously in the same animal without cross-reaction, enabling distinct emissions and timeline readings (34), a allowing monitoring of parasite behavior and drug efficacy without interference between one another.
EGFP and DsRed1-1 fluorescent proteins were used as T. cruzi reporters in Florêncio-Martinez et al. (56) to study infectious process in live cells. This model provided insights into molecular mechanisms of intracellular microorganism infection (56), permitting real-time observation of the infection progress through treatment, in vitro and in vivo, as shown by Canavaci et al. (22).
Costa et al. (11) investigated T. cruzi role in CD progression by generating a reporter strain, CL-Luc, incorporating a red-shifted luciferase/GFP fusion protein (Luc-mNeonGreen). Luminescence and fluorescence enabled monitoring of infection kinetics, infection sites, and parasite–host interactions at cellular level (11). The strain was further modified with CRISPR/Cas9 to generate null parasites with fluorescence (11) enabling analysis of gene function and drug efficacy at molecular levels.
Taylor et al. (67) used T. cruzi CL-Luc:Neon strains expressing chimeric bioluminescent and fluorescent protein to visualize individual parasites in mouse tissue and investigate replication in host cells. Ward et al. (68) developed a transgenic T. cruzi expressing bioluminescent and fluorescent fusion proteins, proposing a model for heart disease development during chronic phase. Precise information on parasite persistence site was needed. Using murine tissue, ex vivo imaging and confocal microscopy, they visualized host cells infected with two strains: T. cruzi CL-Luc:Neon, a CL Brener clone (DTU TcIV) expressing red-shifted luciferase linked to mNeonGreen; and the JR Clone (DTU TcI) expressing red-shifted luciferase (68).
Dual systems have gained space in drug repositioning: Rivero et al. (69) generated the Tulahuen Luc-mNeonGreen strain (DTU TcVI), expressing a double reporter gene, and infected mice to characterize carvedilol’s efficacy as a promising hit.
Olmo et al. (70) developed a panel of transfected T. cruzi strains, expressing bioluminescent/fluorescent fusion proteins—a tool expected to enhance data from experimental infection models, in vitro and in vivo, and enable studies of mixed infections in CD drug development (70).
5 Conclusion
In conclusion, based on the reviewed literature, genetically modified parasites expressing fluorescent and luminescent proteins represent a promising approach for real-time monitoring of infection and treatment response. The simultaneous use of multiple reporter systems, despite potential challenges related to cost and optimization, appears to be a promising strategy to combine the advantages of different techniques while reducing their limitations. These reporter systems facilitate compounds screening with trypanocidal potential and enhance the analysis of parasite–host interactions, significantly contributing to the development of new therapies against T. cruzi.
Transgenic parasites, as crucial tool for drug discovery, offers benefits such as reduced costs, time and labor. This review correlates the evolutionary forms of transgenic T. cruzi, reporter genes, and their applications as valuable tools for screening drug candidates against CD, providing a framework for future research models.
Advancements in the application of transgenic parasites mark a significant step forward in pharmacological research. However, continuous methodological optimization is necessary to maximize their potential in the development of new treatments for CD.
Author contributions
DS: Conceptualization, Investigation, Writing – original draft. ES: Conceptualization, Investigation, Writing – original draft. FG: Conceptualization, Investigation, Writing – original draft. DM: Formal analysis, Writing – review & editing. PS: Formal analysis, Writing – review & editing. MB: Formal analysis, Writing – review & editing. VP: Conceptualization, Formal analysis, Project administration, Supervision, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by Fundação para o Desenvolvimento Científico e Tecnológico em Saúde - Fiotec.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Chagas, C. Nova tripanozomiaze humana: estudos sobre a morfolojia e o ciclo evolutivo do Schizotrypanum cruzi n. gen., n. sp., ajente etiolojico de nova entidade morbida do homem. Mem Inst Oswaldo Cruz. (1909) 1:159–218. Available online at: https://www.biodiversitylibrary.org/item/49879#page/7/mode/1up (Accessed April 23, 2025).
2. Word Health Organization. Chagas’ disease (American Trypanosomiasis) fact sheet. (2023). Available online at: https://www.who.int/news-room/fact-sheets/detail/chagas-disease-(american-trypanosomiasis) (Accessed April 22, 2025).
3. Sales Junior, PA, Molina, I, Fonseca Murta, SM, Sánchez-Montalvá, A, Salvador, F, Corrêa-Oliveira, R, et al. Experimental and clinical treatment of Chagas disease: a review. Am J Trop Med Hyg. (2017) 97:1289–303. doi: 10.4269/ajtmh.16-0761
4. Nunes, MCP, Bern, C, Clark, EH, Teixeira, AL, and Molina, I. Clinical features of Chagas disease progression and severity. Lancet Reg Health-Am. (2024) 37:100832. doi: 10.1016/j.lana.2024.100832
5. de Gonzaga, BMS, Ferreira, RR, Coelho, LL, ACC, C, Garzoni, LR, and Araujo-Jorge, TC. Clinical trials for Chagas disease: etiological and pathophysiological treatment. Front Microbiol. (2023) 14:17. doi: 10.3389/fmicb.2023.1295017
6. Dumonteil, E, and Herrera, C. The case for the development of a Chagas disease vaccine: why? How? When? Trop Med Infect Dis. (2021) 6:16. doi: 10.3390/tropicalmed6010016
7. Chatelain, E, and Konar, N. Translational challenges of animal models in Chagas disease drug development: a review. Drug Des Devel Ther. (2015) 9:4807–23. doi: 10.2147/DDDT.S90208
8. Hernández-Flores, A, Elías-Díaz, D, Cubillo-Cervantes, B, Ibarra-Cerdeña, CN, Morán, D, Arnal, A, et al. Fighting strategies against Chagas’ disease: a review. Pathogens. (2025) 14:183. doi: 10.3390/pathogens14020183
9. de Soeiro, MNC. Perspectives for a new drug candidate for Chagas disease therapy. Mem Inst Oswaldo Cruz. (2022) 117:e220004. doi: 10.1590/0074-02760220004
10. DNDi. DNDi factsheet Chagas disease [Internet]. (2023) Available online at: https://dndi.org/wp-content/uploads/2024/09/DNDi-Factsheet-ChagasDisease-2023.pdf (Accessed April 22, 2025).
11. Costa, FC, Francisco, AF, Jayawardhana, S, Calderano, SG, Lewis, MD, Olmo, F, et al. Expanding the toolbox for Trypanosoma cruzi: a parasite line incorporating a bioluminescence-fluorescence dual reporter and streamlined CRISPR/Cas9 functionality for rapid in vivo localisation and phenotyping. PLoS Negl Trop Dis. (2018) 12:e0006388. doi: 10.1371/journal.pntd.0006388
12. Alonso-Padilla, J, Abril, M, de Noya, BA, Almeida, IC, Angheben, A, Jorge, TA, et al. Target product profile for a test for the early assessment of treatment efficacy in Chagas disease patients: an expert consensus. PLoS Negl Trop Dis. (2020) 14:e0008035. doi: 10.1371/journal.pntd.0008035
13. Lewis, MD, Francisco, AF, Taylor, MC, and Kelly, JM. A new experimental model for assessing drug efficacy against Trypanosoma cruzi infection based on highly sensitive in vivo imaging. SLAS Discov. (2015) 20:36–43. doi: 10.1177/1087057114552623
14. Romanha, AJ, de Castro, SL, de Soeiro, M, Lannes-Vieira, J, Ribeiro, I, Talvani, A, et al. In vitro and in vivo experimental models for drug screening and development for Chagas disease. Mem Inst Oswaldo Cruz. (2010) 105:233–8. doi: 10.1590/S0074-02762010000200022
15. Sharma, R, Silveira-Mattos, PS, Ferreira, VC, Rangel, FA, Oliveira, LB, Celes, FS, et al. Generation and characterization of a dual-reporter transgenic Leishmania braziliensis line expressing eGFP and luciferase. Front Cell Infect Microbiol. (2020) 9:68. doi: 10.3389/fcimb.2019.00468
16. Neefjes, M, Housmans, B, van den Akker, G, van Rhijn, L, Welting, T, and van der Kraan, P. Reporter gene comparison demonstrates interference of complex body fluids with secreted luciferase activity. Sci Rep. (2021) 11:1359. doi: 10.1038/s41598-020-80451-6
17. Zulfiqar, B, Shelper, TB, and Avery, VM. Leishmaniasis drug discovery: recent progress and challenges in assay development. Drug Discov Today. (2017) 22:1516–31. doi: 10.1016/j.drudis.2017.06.004
18. Andriani, G, Chessler, ADC, Courtemanche, G, Burleigh, BA, and Rodriguez, A. Activity in vivo of anti-Trypanosoma cruzi compounds selected from a high throughput screening. PLoS Negl Trop Dis. (2011) 5:e1298. doi: 10.1371/journal.pntd.0001298
19. Scarim, CB, Olmo, F, Ferreira, EI, Chin, CM, Kelly, JM, and Fortes, FA. Image-based in vitro screening reveals the Trypanostatic activity of Hydroxymethylnitrofurazone against Trypanosoma cruzi. Int J Mol Sci. (2021) 22:6930. doi: 10.3390/ijms22136930
20. Boy, RL, Hong, A, Aoki, JI, Floeter-Winter, LM, and Laranjeira-Silva, MF. Reporter gene systems: a powerful tool for Leishmania studies. Curr Res Microb Sci. (2022) 3:100165. doi: 10.1016/j.crmicr.2022.100165
21. Dube, A, Gupta, R, and Singh, N. Reporter genes facilitating discovery of drugs targeting protozoan parasites. Trends Parasitol. (2009) 25:432–9. doi: 10.1016/j.pt.2009.06.006
22. Canavaci, AMC, Bustamante, JM, Padilla, AM, Brandan, CMP, Simpson, LJ, Xu, D, et al. In vitro and in vivo high-throughput assays for the testing of anti-Trypanosoma cruzi compounds. PLoS Negl Trop Dis. (2010) 4:e740. doi: 10.1371/journal.pntd.0000740
23. Goyard, S, Dutra, PL, Deolindo, P, Autheman, D, D’Archivio, S, and Minoprio, P. In vivo imaging of trypanosomes for a better assessment of host-parasite relationships and drug efficacy. Parasitol Int. (2014) 63:260–8. doi: 10.1016/j.parint.2013.07.011
24. Alam, J, and Cook, JL. Reporter genes for monitoring gene expression in mammalian cells. New comprehensive biochemistry. (2003) 38:291–308. doi: 10.1016/S0167-7306(03)38018-4
25. Buckner, FS, Verlinde, CL, La Flamme, AC, and Van Voorhis, WC. Efficient technique for screening drugs for activity against Trypanosoma cruzi using parasites expressing beta-galactosidase. Antimicrob Agents Chemother. (1996) 40:2592–7. doi: 10.1128/AAC.40.11.2592
26. Dantas, RF, Torres-Santos, EC, and Silva, FP Jr. Past and future of trypanosomatids high-throughput phenotypic screening. Mem Inst Oswaldo Cruz. (2022) 117:e210402. doi: 10.1590/0074-02760210402
27. Bettiol, E, Samanovic, M, Murkin, AS, Raper, J, Buckner, F, and Rodriguez, A. Identification of three classes of heteroaromatic compounds with activity against intracellular Trypanosoma cruzi by chemical library screening. PLoS Negl Trop Dis. (2009) 3:e384. doi: 10.1371/journal.pntd.0000384
28. Peña, I, Pilar Manzano, M, Cantizani, J, Kessler, A, Alonso-Padilla, J, Bardera, AI, et al. New compound sets identified from high throughput phenotypic screening against three Kinetoplastid parasites: an open resource. Sci Rep. (2015) 5:8771. doi: 10.1038/srep08771
29. National Center for Biotechnology Information. PubChem bioassay record for AID 493197, source: Broad Institute [Internet]. Bethesda (MD): National Library of Medicine (US), National Center for Biotechnology Information; 2004. Available from: https://pubchem.ncbi.nlm.nih.gov/bioassay/493197 (Accessed April 23, 2025).
30. Gulin, JEN, Rocco, DM, Alonso, V, Cribb, P, Altcheh, J, and García-Bournissen, F. Optimization and biological validation of an in vitro assay using the transfected Dm28c/pLacZ Trypanosoma cruzi strain. Biol Methods Protoc. (2021) 6:bpab004. doi: 10.1093/biomethods/bpab004
31. Buckner, FS, Wilson, AJ, and Van Voorhis, WC. Detection of live Trypanosoma cruzi in tissues of infected mice by using histochemical stain for beta-galactosidase. Infect Immun. (1999) 67:403–9. doi: 10.1128/IAI.67.1.403-409.1999
32. Zhang, H, Bai, H, Jiang, T, Ma, Z, Cheng, Y, Zhou, Y, et al. Quenching the firefly bioluminescence by various ions. Photochem Photobiol Sci. (2016) 15:244–9. doi: 10.1039/c5pp00432b
33. Sedzro, DM. Structure, Function, Application and Modification Strategy of β-Galactosidase. J Multidis Res Rev. (2018) 1:10–16.
34. Widder, EA, and Falls, B. Review of bioluminescence for engineers and scientists in biophotonics. IEEE J Sel Top Quantum Electron. (2014) 20:232–41. doi: 10.1109/JSTQE.2013.2284434
35. McLatchie, AP, Burrell-Saward, H, Myburgh, E, Lewis, MD, Ward, TH, Mottram, JC, et al. Highly sensitive in vivo imaging of Trypanosoma brucei expressing “red-shifted” luciferase. PLoS Negl Trop Dis. (2013) 7:e2571. doi: 10.1371/journal.pntd.0002571
36. Syed, AJ, and Anderson, JC. Applications of bioluminescence in biotechnology and beyond. Chem Soc Rev. (2021) 50:5668–705. doi: 10.1039/D0CS01492C
37. Henriques, C, Henriques-Pons, A, Meuser-Batista, M, Ribeiro, AS, and de Souza, W. In vivo imaging of mice infected with bioluminescent Trypanosoma cruzi unveils novel sites of infection. Parasit Vectors. (2014) 7:89. doi: 10.1186/1756-3305-7-89
38. Francisco, AF, Lewis, MD, Jayawardhana, S, Taylor, MC, Chatelain, E, and Kelly, JM. Limited ability of Posaconazole to cure both acute and chronic Trypanosoma cruzi infections revealed by highly sensitive in vivo imaging. Antimicrob Agents Chemother. (2015) 59:4653–61. doi: 10.1128/AAC.00520-15
39. Calvet, CM, Choi, JY, Thomas, D, Suzuki, B, Hirata, K, Lostracco-Johnson, S, et al. 4-aminopyridyl-based lead compounds targeting CYP51 prevent spontaneous parasite relapse in a chronic model and improve cardiac pathology in an acute model of Trypanosoma cruzi infection. PLoS Negl Trop Dis. (2017) 11:e0006132. doi: 10.1371/journal.pntd.0006132
40. Calvet, CM, Choi, JY, Thomas, D, Suzuki, B, Hirata, K, Lostracco-Johnson, S, et al. 4-aminopyridyl-based lead compounds targeting CYP51 prevent spontaneous parasite relapse in a chronic model and improve cardiac pathology in an acute model of Trypanosoma cruzi infection. PLoS Negl Trop Dis. (2017). 11:e0006132. Available online at: https://journals.plos.org/plosntds/article?id=10.1371/journal.pntd.0006132 (Accessed March 6, 2025).
41. WoRMS. World Register of Marine Species-Oplophorus gracilirostris A. Milne-Edwards. (1881). Available online at: https://www.marinespecies.org/aphia.php?p=taxdetails&%20id=212900 (Accessed April 25, 2025).
42. Mezzanotte, L, van’t Root, M, Karatas, H, Goun, EA, and Löwik, CWGM. In vivo molecular bioluminescence imaging: new tools and applications. Trends Biotechnol. (2017) 35:640–52. doi: 10.1016/j.tibtech.2017.03.012
43. Silberstein, E, Kim, KS, Acosta, D, and Debrabant, A. Human placental trophoblasts are resistant to Trypanosoma cruzi infection in a 3D-culture model of the maternal-fetal Interface. Front Microbiol. (2021) 12:626370. doi: 10.3389/fmicb.2021.626370
44. Hall, MP, Unch, J, Binkowski, BF, Valley, MP, Butler, BL, Wood, MG, et al. Engineered luciferase reporter from a Deep Sea shrimp utilizing a novel Imidazopyrazinone substrate. ACS Chem Biol. (2012) 7:1848–57. Available online at: https://pubs.acs.org/doi/full/10.1021/cb3002478 (Accessed January 17, 2025).
45. Loening, AM, Dragulescu-Andrasi, A, and Gambhir, SS. A red-shifted Renilla luciferase for transient reporter-gene expression. Nat Methods. (2010) 7:5–6. doi: 10.1038/nmeth0110-05
46. WoRMS. World Register of Marine Species-Renilla reniformis (1766). Available online at: https://www.marinespecies.org/aphia.php?p=taxdetails&id=290965 (Accessed April 14, 2025).
47. Shaner, NC, Steinbach, PA, and Tsien, RY. A guide to choosing fluorescent proteins. Nat Methods. (2005) 2:905–9. doi: 10.1038/nmeth819
48. Sommer, JR, Alderson, J, Laible, G, and Petters, RM. Reporter system for the detection of in vivo gene conversion: changing colors from blue to green using GFP variants. Mol Biotechnol. (2006) 33:115–22. doi: 10.1385/MB:33:2:115
49. Fesser, AF, Braissant, O, Olmo, F, Kelly, JM, Mäser, P, and Kaiser, M. Non-invasive monitoring of drug action: a new live in vitro assay design for Chagas’ disease drug discovery. PLoS Negl Trop Dis. (2020) 14:e0008487. doi: 10.1371/journal.pntd.0008487
50. Kessler, RL, Gradia, DF, de Pontello Rampazzo, R, Lourenço, ÉE, Fidêncio, NJ, Manhaes, L, et al. Stage-regulated GFP expression in Trypanosoma cruzi: applications from host-parasite interactions to drug screening. PLoS One. (2013) 8:e67441. doi: 10.1371/journal.pone.0067441
51. DaRocha, WD, Silva, RA, Bartholomeu, DC, Pires, SF, Freitas, JM, Macedo, AM, et al. Expression of exogenous genes in Trypanosoma cruzi: improving vectors and electroporation protocols. Parasitol Res. (2004) 92:113–20. doi: 10.1007/s00436-003-1004-5
52. Jaiswal, AK, Rao, KB, Kushwaha, P, Rawat, K, Modukuri, RK, Khare, P, et al. Development of Leishmania donovani stably expressing DsRed for flow cytometry-based drug screening using chalcone thiazolyl-hydrazone as a new antileishmanial target. Int J Antimicrob Agents. (2016) 48:695–702. doi: 10.1016/j.ijantimicag.2016.09.018
53. Farhadi, A, Sigmund, F, Westmeyer, GG, and Shapiro, MG. Genetically encodable materials for non-invasive biological imaging. Nat Mater. (2021) 20:585–92. doi: 10.1038/s41563-020-00883-3
54. Pires, SF, DaRocha, WD, Freitas, JM, Oliveira, LA, Kitten, GT, Machado, CR, et al. Cell culture and animal infection with distinct Trypanosoma cruzi strains expressing red and green fluorescent proteins. Int J Parasitol. (2008) 38:289–97. doi: 10.1016/j.ijpara.2007.08.013
55. Miranda, CG, Solana, ME, de Curto, M, Lammel, EM, Schijman, AG, and Alba Soto, CD. A flow cytometer-based method to simultaneously assess activity and selectivity of compounds against the intracellular forms of Trypanosoma cruzi. Acta Trop. (2015) 152:8–16. doi: 10.1016/j.actatropica.2015.08.004
56. Florencio-Martínez, L, Márquez-Dueñas, C, Ballesteros-Rodea, G, Martínez-Calvillo, S, and Manning-Cela, R. Cellular analysis of host cell infection by different developmental stages of Trypanosoma cruzi. Exp Parasitol. (2010) 126:332–6. doi: 10.1016/j.exppara.2010.04.015
57. De Souza, W, and Barrias, ES. May the epimastigote form of Trypanosoma cruzi be infective? Acta Trop. (2020) 212:105688. doi: 10.1016/j.actatropica.2020.105688
58. Niemirowicz, GT, Carlevaro, G, Campetella, O, Bouvier, LA, and Mucci, J. A versatile 2A peptide-based strategy for ectopic expression and endogenous gene tagging in Trypanosoma cruzi. Heliyon. (2024) 10:e24595. doi: 10.1016/j.heliyon.2024.e24595
59. Mishin, AS, Subach, FV, Yampolsky, IV, King, W, Lukyanov, KA, and Verkhusha, VV. The first mutant of the Aequorea victoria green fluorescent protein that forms a red chromophore. Biochemistry. (2008) 47:4666–73. doi: 10.1021/bi702130s
60. Lainson, R, and Shaw, JJ. Leishmaniasis of the New World: taxonomic problems. Br Med Bull. (1972) 28:44–8. doi: 10.1093/oxfordjournals.bmb.a070892
61. Pessoa, CC, Ferreira, ÉR, Bayer-Santos, E, Rabinovitch, M, Mortara, RA, and Real, F. Trypanosoma cruzi differentiates and multiplies within chimeric Parasitophorous vacuoles in macrophages Coinfected with Leishmania amazonensis. Infect Immun. (2016) 84:1603–14. doi: 10.1128/IAI.01470-15
62. Ferreira, BL, Orikaza, CM, Cordero, EM, and Mortara, RA. Trypanosoma cruzi: single cell live imaging inside infected tissues. Cell Microbiol. (2016) 18:779–83. doi: 10.1111/cmi.12553
63. Morris, LM, Klanke, CA, Lang, SA, Lim, FY, and Crombleholme, TM. TdTomato and EGFP identification in histological sections: insight and alternatives. Biotech Histochem. (2010) 85:379–87. doi: 10.3109/10520290903504753
64. Bustamante, JM, and Tarleton, RL. Methodological advances in drug discovery for Chagas disease. Expert Opin Drug Discov. (2011) 6:653–61. doi: 10.1517/17460441.2011.573782
65. Fonseca-Berzal, C, Arán, VJ, Escario, JA, and Gómez-Barrio, A. Experimental models in Chagas disease: a review of the methodologies applied for screening compounds against Trypanosoma cruzi. Parasitol Res. (2018) 117:3367–80. doi: 10.1007/s00436-018-6084-3
66. Svensen, N, Wyllie, S, Gray, DW, and Rycker, MD. Live-imaging rate-of-kill compound profiling for Chagas disease drug discovery with a new automated high-content assay. PLoS Negl Trop Dis. (2021) 15:e0009870. doi: 10.1371/journal.pntd.0009870
67. Taylor, MC, Francisco, AF, Jayawardhana, S, Mann, GS, Ward, AI, Olmo, F, et al. Exploiting genetically modified dual-reporter strains to monitor experimental Trypanosoma cruzi infections and host-parasite interactions. Methods Mol Biol. (2019) 1955:147–63. doi: 10.1007/978-1-4939-9148-8_11
68. Ward, AI, Lewis, MD, Khan, AA, McCann, CJ, Francisco, AF, Jayawardhana, S, et al. In vivo analysis of Trypanosoma cruzi persistence foci at single-cell resolution. MBio. (2020) 11:e01242–20. doi: 10.1128/mBio.01242-20
69. Rivero, CV, Martínez, SJ, Novick, P, Cueto, JA, Salassa, BN, Vanrell, MC, et al. Repurposing carvedilol as a novel inhibitor of the Trypanosoma cruzi autophagy flux that affects parasite replication and survival. Front Cell Infect Microbiol. (2021) 11:657257. doi: 10.3389/fcimb.2021.657257
70. Olmo, F, Jayawardhana, S, Khan, AA, Langston, HC, Francisco, AF, Atherton, RL, et al. A panel of phenotypically and genotypically diverse bioluminescent: fluorescent Trypanosoma cruzi strains as a resource for Chagas disease research. PLoS Negl Trop Dis. (2024) 18:e0012106. doi: 10.1371/journal.pntd.0012106
71. Lu, HY, and Buck, GA. Expression of an exogenous gene in Trypanosoma cruzi epimastigotes. Mol Biochem Parasitol. (1991) 44:109–14. doi: 10.1016/0166-6851(91)90226-V
72. Vega, C, Rolón, M, Martínez-Fernández, AR, Escario, JA, and Gómez-Barrio, A. A new pharmacological screening assay with Trypanosoma cruzi epimastigotes expressing beta-galactosidase. Parasitol Res. (2005) 95:296–8. doi: 10.1007/s00436-005-1300-3
73. Lara, LS, Moreira, CS, Calvet, CM, Lechuga, GC, Souza, RS, Bourguignon, SC, et al. Efficacy of 2-hydroxy-3-phenylsulfanylmethyl-[1, 4]-naphthoquinone derivatives against different Trypanosoma cruzi discrete type units: identification of a promising hit compound. Eur J Med Chem. (2018) 144:572–81. doi: 10.1016/j.ejmech.2017.12.052
74. Won, MM, Krüger, T, Engstler, M, and Burleigh, BA. The intracellular amastigote of Trypanosoma cruzi maintains an actively beating flagellum. MBio. (2023) 14:e0355622. doi: 10.1128/mbio.03556-22
Keywords: Chagas disease, Trypanosoma cruzi, transgenic parasite, reporter gene, drug screening
Citation: Sousa Oliveira DS, da Silva ED, Galvão Júnior FF, Moura DMN, Sales-Junior PA, Brelaz-de-Castro MCA and Pereira VRA (2025) Reporter genes and transgenic Trypanosoma cruzi (Kinetoplastida, Trypanosomatidae): applications for screening new drugs against Chagas disease. Front. Med. 12:1591148. doi: 10.3389/fmed.2025.1591148
Edited by:
Nicola Pugliese, University of Bari Aldo Moro, ItalyReviewed by:
Kaio Cesar Chaboli Alevi, São Paulo State University, BrazilAngel Ramos-Ligonio, Universidad Veracruzana, Mexico
Copyright © 2025 Sousa Oliveira, da Silva, Galvão Júnior, Moura, Sales-Junior, Brelaz-de-Castro and Pereira. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Daniele Santana de Sousa Oliveira, ZGFuaWVsZXNhbkBnbWFpbC5jb20=; Elis Dionisio da Silva, ZWxpc2Rpb25pc2lvQHVmYW0uZWR1LmJy; Valéria Rêgo Alves Pereira, dmFsZXJpYXBoQGdtYWlsLmNvbQ==
†These authors have contributed equally to this work and share first authorship
‡Senior author