 Patricia Basile
Patricia Basile Federico Wallace
Federico Wallace Cristina Olivaro
Cristina Olivaro Nicolás De Palma
Nicolás De Palma Omar Borsani
Omar Borsani Arthur Fett-Neto
Arthur Fett-Neto- 1Espacio de Biología Vegetal del Noreste, CENUR Noreste, Universidad de la República, Tacuarembó, Uruguay
- 2Espacio de Ciencia y Tecnología Química, CENUR Noreste, Universidad de la República, Tacuarembó, Uruguay
- 3Plant Physiology Laboratory, Center for Biotechnology, Federal University of Rio Grande Do Sul (UFRGS), Porto Alegre, Brazil
- 4Departamento de Biología Vegetal, Facultad de Agronomía, Universidad de la República, Montevideo, Uruguay
Introduction: Cold weather poses a significant challenge to the growth of crops and subtropical tree species like Eucalyptus. Exposure of plants to stressful temperatures generates changes in their physiology resulting from modifications in gene expression and extensive metabolic reorganization. A direct comparison of several biochemical changes under cold exposure of leaf tissues of E. dunnii and E. grandis clones was carried out.
Methods: Leaf discs of E. grandis and E. dunnii were initially maintained for 24 h at 25°C and then 4 days at 6°C to induce cold stress. Sampling was conducted at 0 h (control condition), 2 and 4 days. Several biochemical parameters were measured, and an untargeted metabolomics approach based on ultra-high performance liquid chromatography (UHPLC) coupled to linear ion trap mass spectrometry fingerprinting was carried out.
Results: Results indicated distinct cold tolerance strategies in Eucalyptus grandis and Eucalyptus dunnii. Eucalyptus dunnii initiated protective mechanism activation after a 2-day exposure period with the accumulation of sugars and phenolic compounds, whereas E. grandis did so after 4 days, accumulating proline and anthocyanins. PLS-DA based on UHPLC-MS fingerprints revealed a clear species-specific effect across the metabolome. This effect was greater than the differences between cold temperatures. Additionally, this methodology allowed the putative identification of 16 phenolic marker compounds with high discriminant potential to differentiate the cold response in these two species.
Introduction
Eucalyptus L’H’er. (Myrtaceae), an Australian and Indonesian native genus, is one of the economically most essential hardwood crops worldwide. Approximately 90% of the Eucalyptus species planted around the world are dominated by ‘the big nine’ species (Eucalyptus camaldulensis Dehnh., Eucalyptus grandis W. Mill ex Maiden, Eucalyptus tereticornis Sm., Eucalyptus globulus Labill., Eucalyptus nitens (H. Deane y Maiden) Maiden, Eucalyptus urophylla S.T. Blake, Eucalyptus saligna Sm., Eucalyptus dunnii Maiden, and Eucalyptus pellita F. Muell.) and their hybrids. These species are of particular interest because of their widespread cultivation (Stanturf et al., 2013). In Uruguay, E. dunnii and E. grandis account for almost 80% of the Eucalyptus plantations (Ministerio de Ganadería, Agricultura y Pesca 2024).
Cold is a limiting factor for crop production and subtropical tree species such as Eucalyptus. Low temperatures can lead to physiological and biochemical damage, such as growth inhibition, dehydration, cell membrane damage, accumulation of reactive oxygen species (ROS), and electrolyte leakage (Thomashow, 1999; Yadav, 2010; Guo et al., 2018).
Exposure of plants to stressful temperatures generates changes in their physiology that result of modifications in gene expression and extensive metabolic reorganization. Metabolism adjustment allows plants to avoid damage such as chlorophyll loss and membrane impairment resulting in solute leakage. The protective adjustments include accumulation of compatible osmolytes, such as proline and soluble sugars, photosystem and membrane composition alterations, as well as an increase in antioxidant capacity provided by enzymatic and non-enzymatic antioxidants (Janská et al., 2010; Knaupp et al., 2011; Juszczak et al., 2016; Verslues and Sharma, 2010; Fürtauer et al., 2019). Phenolic compounds such as anthocyanins, which accumulate in vacuoles, can act as non-enzymatic antioxidants, scavenging ROS (Close et al., 2004; Pennycooke et al., 2005; Król et al., 2015; Gould et al., 2018; Oberschelp et al., 2020).
Some species such as Eucalyptus dunnii, Eucalyptus benthamii Maiden & Cambage, Eucalyptus gunnii Miq. and Eucalyptus nitens are generally considered cold tolerant. Other species such as Eucalyptus saligna Sm., Eucalyptus grandis W. Mill ex Maiden and Eucalyptus urophylla S.T. Blake are often regarded as cold sensitive (Jacobs, 1981; Jovanovic, Arnold, and Booth, 2000; Davidson, Battaglia, and Close, 2004; Floriani et al., 2013; Arnold et al., 2015). Selection and breeding for cold tolerance has been a major goal in eucalypt forestry for decades (Arnold et al., 2004), resulting in improved genetic materials both within species and/or by hybridization.
Previous studies have examined cold responses in E. dunnii and E. grandis. Domingues et al. (2019) showed that improved control of energy availability and sink relationships were associated with growth maintenance in young trees of E. globulus versus their E. grandis counterparts when exposed to 10°C for 24 h. Mokochinski et al. (2018) compared E. dunnii, E. grandis, and E. pellita saplings exposed to different temperatures (10°C, 20°C, and 30°C) for 1 week. These authors carried out growth and untargeted metabolomics analyses, showing the relevance of the latter to understand changes upon cold exposure. Oberschelp et al. (2020) examined acclimated and non-acclimated saplings of E. dunnii, E. benthamii, and E. grandis exposed to simulated frosts over a period of 9 h, followed by 15 days in nursery conditions to determine damage indexes. In these authors’ studies, biochemical quantifications, metabolomic analyses, and proteomic comparisons were conducted, indicating the importance of osmoprotectants and antioxidants in acclimation.
Nonetheless, the basis for cold tolerance differences between commercial clones of E. dunnii and E. grandis remains an open topic. A recent study did not find differences in cold tolerance between these species (Oberschelp et al., 2020). In addition, there is little information on cold experiments carried out under winter temperatures that are frequent in Uruguayan humid subtropical climate (Cfa, Koppen climate classification), i.e., between 0°C and 10°C (Kottek et al., 2006).
To help filling this information gap, herein a direct comparison of several biochemical changes under cold exposure of leaf tissues of E. dunnii and E. grandis clones was carried out. A leaf disk-based experimental system was proposed affording faster execution and demanding simpler resources for comparative evaluations. The main question addressed was whether these species differ in mechanisms of defense against low temperatures typically present in winter of mild temperate climates.
Materials and methods
Plant materials and cold treatments
Seedlings of commercial genotypes of Eucalyptus grandis Hill ex Maiden and Eucalyptus dunnii Maiden kindly donated by Lumin S.A and UPM Biofore, respectively, were used in this study.
The experiment was conducted in a growth chamber (Thermo Fisher model precision 818) under irradiance of 515 μmol s−1 (cold fluorescent lamps) and a photoperiod of 12 h/12 h (day/night) at the Tacuarembó campus of the Universidad de la República (UdelaR), Uruguay. For the experiment, leaf discs of plantlets were prepared as described in De Palma et al. (2023).
Briefly, healthy, fully expanded leaves were removed at the petiole insertion point, immersed in 1.5% (v/v) sodium hypochlorite for 15 min, and washed three times with distilled water. Subsequently, 1 cm diameter discs were cut using a steel cork borer and placed in 9 mm Petri dishes (30 discs per dish) with filter paper discs soaked in 0.1 × MS culture medium salts (20 mL per dish) (Murashige and Skoog, 1962).
Leaf discs were initially maintained for 24 h at 25°C and then 4 days at 6°C to induce cold stress. Sampling was conducted at 0 h (control condition, immediately before treatment application), 2 and 4 days. After harvest, disks were immediately frozen at −80°C and then liophilized. Henceforth, the term dry leaf disks refers to the ground lyophilized tissues.
Pigment analysis
Chlorophyll A (ChlA), chlorophyll B (ChlB), carotenoids, and anthocyanins were analyzed as described in De Palma et al. (2023). Dry leaf disks (50 mg) were placed in tubes with 1 mL of cold acetone/0.1 M tris buffer (80/20, v/v) (pH 7.8). These extracts were ground and ultrasonicated in ice water for 15 min and then placed in the dark at 4°C for 24 h. The next day, they were ultrasonicated again for 15 min. The extracts were centrifuged for 15 min at 10,000 × g and 4°C, and the supernatant was recovered, diluted with 100 µL of the extraction solution, and maintained in darkness. Absorbance readings were performed in a SpectraMax M2 UV–Vis spectrophotometer (Molecular Devices, San Jose, United States) at 470, 537, 647, and 663 nm. Concentrations of ChlA, ChlB, Total chlorophyll (TChl), carotenoid, and anthocyanin were estimated according to the equations proposed by Sims and Gamon (2002). ChlA/ChlB ratios were also determined.
Proline quantification
Proline concentration was determined according to Lee et al. (2018), with minor modifications. Dry leaf disks (50 mg) were placed in tubes with 800 µL of 3% (w/v) sulfosalicylic acid and vortexed. The extracts were centrifuged for 15 min at 16,000 × g. One hundred µL of supernatant was mixed with 200 µL of 1.25% (w/v) ninhydrin in 80% (v/v) glacial acetic acid, vortexed, and incubated for 1 h in an oven at 100°C. The reaction was stopped by transferring the mixture to ice for 10 min and vortexing. Absorbance was read in a spectrophotometer at 520 nm. Proline concentration was calculated from a standard curve and expressed on a DW basis.
Lipid peroxidation analysis
Lipid peroxidation was analyzed according to Velikova et al. (2000) with minor modifications. One hundred mg of dry leaf disks were ground with 1,334 µL 0.1% (v/v) of trichloroacetic acid (TCA) and centrifuged at 16,000 × g at 4°C for 20 min. Then, 200 µL of supernatant were collected and mixed with 400 µL of 0.5% (w/v) thiobarbituric acid (TBA) in 20% (v/v) TCA solution. Next, the tubes were incubated in a hot water bath at 100°C for 20 min, after which the reaction was stopped in an ice bath. Finally, absorbance readings were performed at 532 and 600 nm. The lipid peroxidation was estimated as malondialdehyde (MDA) concentration using an extinction coefficient of 155 mM−1 cm−1 and expressed on a DW basis.
Hydrogen peroxide (H2O2) analysis
The hydrogen peroxide concentration was determined from the extract used for lipid peroxidation analysis as described in Velikova et al. (2000). Twenty µL of TCA extract were mixed with 100 µL of 10 mM potassium phosphate buffer and 400 µL 1 M potassium iodide (KI). Next, the reaction was incubated at 4°C for 1 h in the dark. The absorbance was read at 390 nm in a spectrophotometer. The hydrogen peroxide concentration was calculated and expressed on a DW basis.
Quantification of total soluble proteins (TSP)
Quantification of TSP followed the method of Bradford (1976). First, 50 mg of dry leaf disks were ground using a tissue disruptor with 1.5 mL of extraction buffer containing 1% (w/v) polyvinylpyrrolidone, 1 mM ethylenediaminetetraacetic acid, 1 mM phenylmethylsulfonyl fluoride, and 50 mM HEPES buffer (pH 7.4). Next, the ground material was centrifuged at 16,000 × g at 4°C for 15 min, and the resulting supernatants (protein extracts) were collected and transferred to new tubes. To quantify TSP, 30 μL of protein extract was mixed with 1.5 mL of Bradford reagent (PierceTM Bradford Protein Assay Kit, Thermo Scientific, Waltham, United States). Finally, absorbance readings were taken at 595 nm, and the TSP content was determined using a BSA standard curve.
Superoxide dismutase (SOD) activity
SOD activity was determined as outlined by Beyer and Irwin (1987), with slight modifications. Thirteen µL of protein extract were combined with 1 mL of 50 mM phosphate buffer (pH 7.8) containing 57 µM nitro blue tetrazolium (NBT), 9.9 mM L-methionine, 0.025% (w/v) Triton® X-100, and 2 mM riboflavin. The mixture was then placed for 15 min in a transilluminator with fluorescent light (20 W). Absorbance readings were obtained at 560 nm, and the SOD activity was calculated and expressed as SOD units. µg−1 protein.
Relative electrolyte leakage (EL%)
Relative electrolyte leakage was estimated according to Thalhammer et al. (2014), with some modifications. Previously washed leaf disks were placed in plastic tubes with ultrapure water and incubated in agitation for 24 h at room temperature. Then, the initial electric conductivity (Ci) was measured with a portable conductivity meter (HORIBA, model LaquaTwin, Kyoto, Japan). Afterwards, the tubes were incubated in a hot water bath for 30 min at 100°C. Subsequently, final electric conductivity (Cf) was measured, and EL% was calculated following the equation
Total soluble sugar content (TSS)
TSS quantification followed the protocol of Chow and Landhäusser (2004) with minor modifications. Thirteen mg of leaf disks were ground in liquid nitrogen and extracted two times in 750 µL ethanol 80% (w/v). Then, the extracts were incubated in a dry bath for 20 min at 75°C and centrifuged for 10 min at 16,000 × g. Next, the supernatants were collected and combined, constituting the total soluble sugar extracts. Afterwards, 75 µL of total soluble sugar extracts were diluted with 125 µL ethanol 80% (p/v) and mixed with 200 µL 5% (w/v) phenolic acid and 1 mL sulfuric acid. The reaction was incubated with agitation for 20 min at room temperature, and then the absorbances were read at 490 nm. The TSS concentration was expressed on a DW basis.
Total phenolics content
Total phenolics were determined as reported by Makkar (2000). Two hundred mg of dry leaf disks were ground with 10 mL 70% acetone (v/v) and ultrasonicated in ice water for 20 min. Next, extracts were centrifuged at 3,000 × g for 10 min at 4°C. Supernatants were collected and mixed with 450 µL distilled water, 250 µL 1 N Folin-Ciocalteu reagent and 1.25 mL 20% (w/v) sodium carbonate. The reaction mixture was vortexed and incubated for 40 min in darkness. Then, absorbance readings were done at 725 nm. Total phenolics concentration was expressed as mg gallic acid g−1DW.
Statistical analysis
Experiments were carried out in a totally randomized design using four biological replicates of each treatment, and statistical analysis was performed using R studio software version 2024.4.2. Normality tests and one-way Analysis of Variance (ANOVA) followed by Tukey’s, Mann–Whitney or Kruskal–Wallis tests were applied when appropriate. Multivariate analysis was performed with Principal Components Analysis (PCA) with “FactoMineR” package in R (R Core Team, 2023).
LC-MS/MS fingerprinting with a focus on phenolic compounds
Leaf disks were extracted according to Noleto-Dias et al. (2023) with some modifications. Succinctly, 30 mg of dry leaf disks were extracted with 750 µL water: methanol (50:50, v/v) and then centrifuged at 13,000 × g for 5 min. Supernatants were collected, filtered, and placed in glass vials for analysis.
Liquid chromatography coupled with tandem mass spectrometry (LC-MS/MS) was performed using a linear ion trap mass spectrometer (LTQ XL, Thermo Scientific, San José, United States) equipped with an electrospray ionization (ESI) interface. The liquid chromatography was conducted on a UHPLC system (Ultimate 3000 RSLC, Dionex, Waltham, United States). All data were collected and processed using XcaliburTM software (v3.063) (Thermo Scientific, 2013). The parameters of the source ESI were as follows: spray voltage 3.5 kV, capillary voltage −3 V, tube lens offset −228 V, sheath gas (nitrogen) flow rate 20 (arbitrary units, a.u.), auxiliary gas (nitrogen) flow rate 20 a.u., and capillary temperature 300°C. Mass spectrometry analysis was conducted in the negative ionization mode and scanned from m/z 150 to 900.
The separation column was a Hypersil Gold RP C18 (100 mm × 2.1 mm ID, 1.9 µm particle size) from Thermo Scientific (Waltham, United States). A 30 µL sample aliquot was injected into the UHPLC-MS system. The mobile phase consisted of a gradient of water and acetonitrile, both with 0.1% (v/v) formic acid. Initial conditions were 5% acetonitrile, and the gradient was run to 95% acetonitrile within 25 min and returned to 5% acetonitrile in the next 12 min. The flow rate was set to 0.25 mL/min, and the column was maintained at 40°C. In LC-MS/MS, the mass spectrometry was operated under Data Dependent™ scan mode. This acquisition mode included a single Full MS scan followed by MS/MS scans on the most abundant ions. The collision energy used was 35%.
Leaf disk samples were randomly analyzed using the pre-defined UHPLC-MS system. Eight records were obtained for each sample, including biological replicates. In addition, five quality control (QC) samples, prepared with 30 µL of each analyzed sample extract, were used to evaluate the chemometric results. QC samples and extracting solvent blanks were periodically injected into the sample sequence.
The data matrices for the chemometric analysis were obtained using MZMine 3.9.0 Software (Schmid et al., 2023). Partial Least Square Discriminant Analysis (PLS-DA) was performed using the SOLO 9.3 chemometrics Software (Eigenvector Research, Manson, WA, United States). Moreover, the average of the variable importance in projection (VIP) values obtained after building PLS-DA models were used to select variables with the most significant influence on the observed sample distribution using 1.0 as the cut-off value. Based on these variables, the MS2 spectrum of each one was used for metabolite identification. This was achieved by searching the NIST 20 Tandem Library (lr_msms_nist and nist_msms) using the MS/MS Identity Search function of the NIST MS Search Program (v.2.4) and by comparing the results with reported data in the literature for Eucalyptus samples. Finally, using those metabolites previously identified, a Heatmap was generated using “pheatmap” package in R (R Core Team, 2023). Hierarchical clustering was performed using Pearson correlation.
Results
Time course changes in biochemical responses to cold stress in Eucalyptus grandis and Eucalyptus dunnii
In E. grandis, significant differences were observed only in total soluble protein (TSP) and proline content across the experimental days (Figures 1B,C). TSP increased over cold exposure time, nearly doubling on day 4 (Figure 1B). From day one to the fourth, proline concentration increased significantly, approximately three-fold (Figure 1C). Cold treatment did not induce changes in photosynthetic pigments such as TChl, ChlA, ChlB, ChlA/ChlB, carotenoids, TSS, phenolics, SOD activity, anthocyanins, MDA, H2O2, or EL% (Table 1; Figures 1A,D–F, 2).
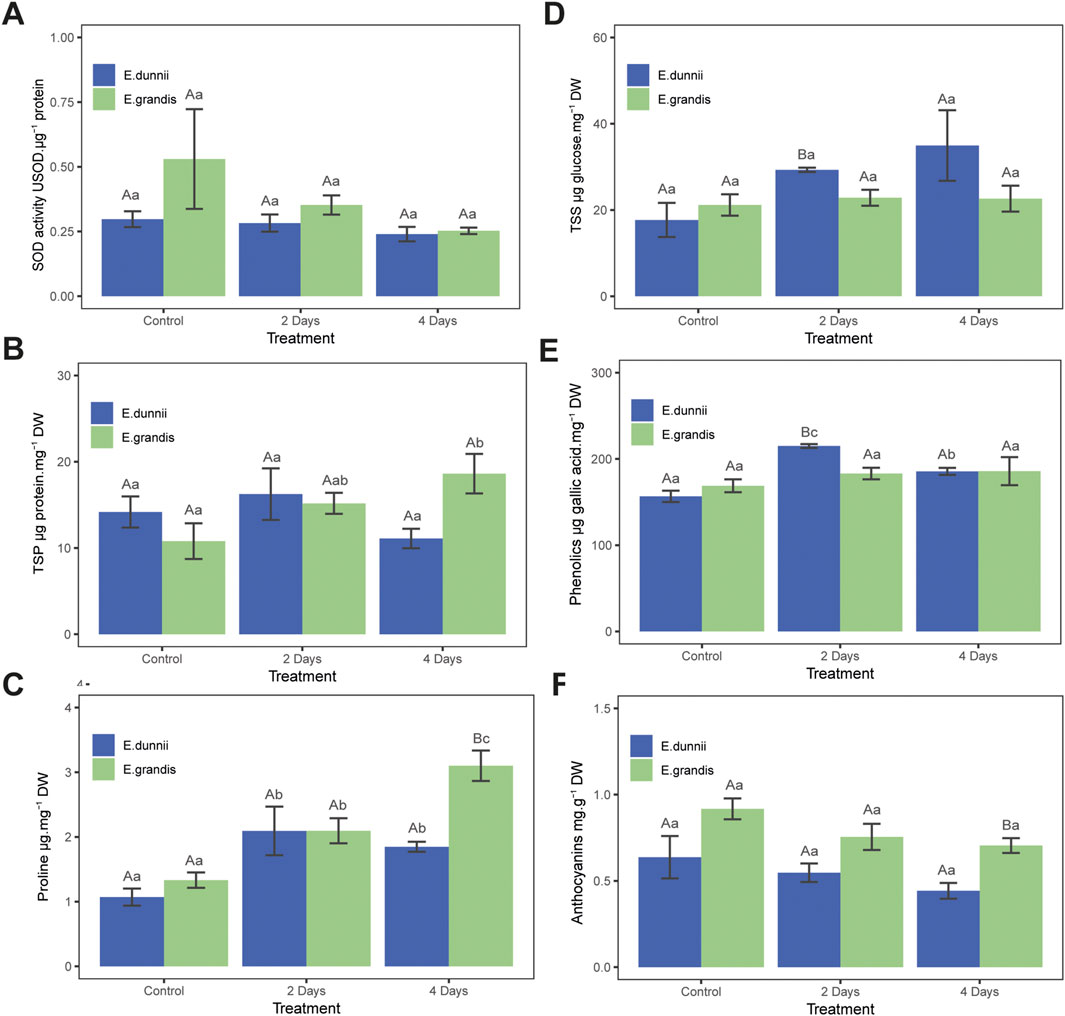
Figure 1. Stress protection parameters in Eucalyptus grandis and Eucalyptus dunnii leaf disks on the 2nd and 4th day of cold stress. Values correspond to mean ± SE (n = 4). Statistically different samples (p < 0.05) are indicated in capital or lowercase letters after inter-species or inter-treatment comparison, respectively. Lines on top of bars are standard errors. (A) Superoxide dismutase (SOD) activity. (B) Total soluble protein (TSP) concentration. (C) Proline concentration. (D) Total soluble sugars (TSS) concentration. (E) Total phenolic compounds concentration. (F) Anthocyanin concentration.
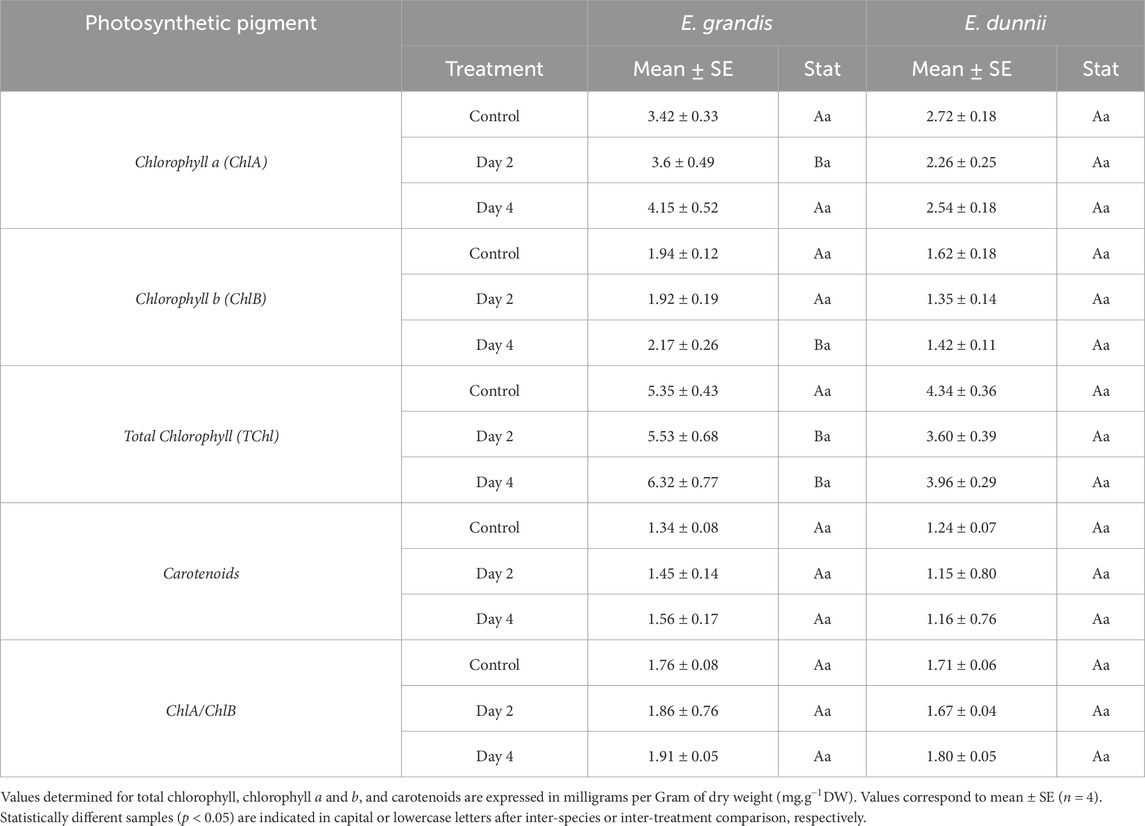
Table 1. Photosynthetic pigments in Eucalyptus grandis and Eucalyptus dunnii leaf disks on the 2nd and 4th day of cold stress.
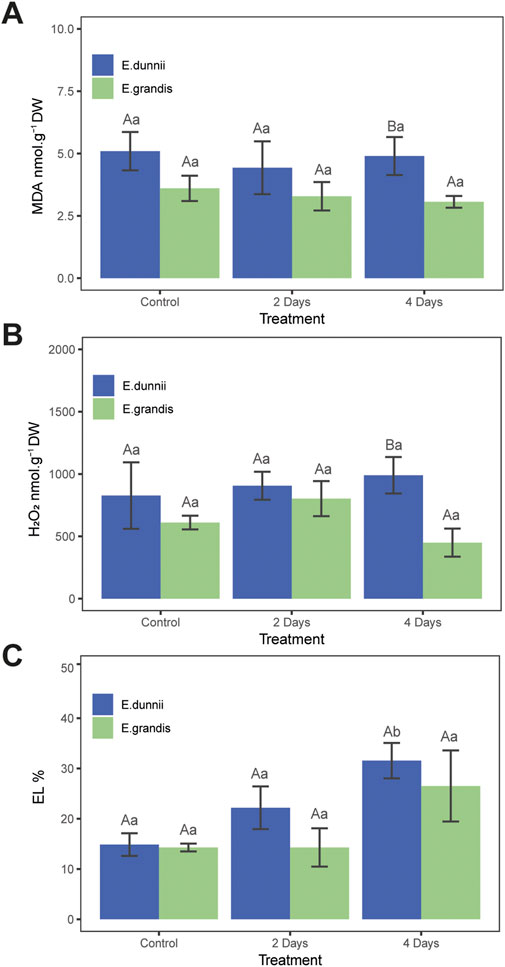
Figure 2. Stress indicator parameters in Eucalyptus grandis and Eucalyptus dunnii leaf disks on the 2nd and 4th day of cold stress. Values correspond to mean ± SE (n = 4). Statistically different samples (p < 0.05) are indicated in capital or lowercase letters after inter-species or inter-treatment comparison, respectively. Lines on top of bars are standard errors. (A) MDA levels. (B) H2O2 concentration. (C) Relative electrolyte leakage (EL%).
In E. dunnii, significant differences were observed only for EL%, proline, and phenolic compounds across the experimental days. After 4 days of stress exposure, EL values doubled, while lipid peroxidation (MDA) and H2O2 content remained relatively stable (Figure 2). Concerning protective mechanisms, the highest accumulation of the osmolyte proline was observed on day two, in which it approximately doubled its initial value, remaining stable on day 4 (Figure 1C). Total phenolic amount increased significantly over time, peaking on day two but slightly decreasing on day four (Figure 1E). Photosynthetic pigments (TChl, ChlA, ChlB, ChlA/ChlB, and carotenoids), anthocyanins, and SOD activity did not exhibit significant changes during the experimental period in this species (Table 1; Figures 1A,F).
Comparison of biochemical changes under cold stress between species: Eucalyptus grandis versus Eucalyptus dunnii
Regarding stress indicators, there were no significant differences between species in EL% during the experiment (Figure 2C). Moreover, lipid peroxidation (MDA concentration) and H2O2 only exhibited significant differences on day 4, when E. dunnii showed higher values for both parameters (Figures 2A,B).
As for TChl, E. grandis showed higher values on day 2. Higher concentrations of ChlA and ChlB were also recorded in E. grandis on days 2 and 4, respectively. The ChlA/ChB ratio did not show significant differences between species (Table 1).
SOD activity, carotenoids, and TSP did not present differences between species throughout the experiments (Table 1; Figures 1A,B). After 2 days of stress, E. dunnii showed higher levels of TSS and phenolics (Figures 1D,E). The remaining parameters, proline and anthocyanin concentrations, displayed significant differences between species only on day 4 when E. grandis showed higher values than E. dunnii (Figures 1C,F).
Principal component analysis points to species and treatment trends
Principal component analysis (PCA) was performed with biochemical parameter data to study the relationships between species and treatments with metabolism and stress tolerance. 2 PC (54.7% variation) were used to generate a biplot (Figure 3). PC1 explained the highest proportion of variance (37.3%) and represented differences between species. Variables like proline, TSP, anthocyanins, carotenoids, SOD activity and chlorophylls (TChl, ChlA, ChlB, ChlA/ChlB) were positively associated with PC1 and E. grandis. Moreover, variables like TSS, phenolics, MDA, EL% and H2O2 were negatively correlated with PC1 and associated with E. dunnii.
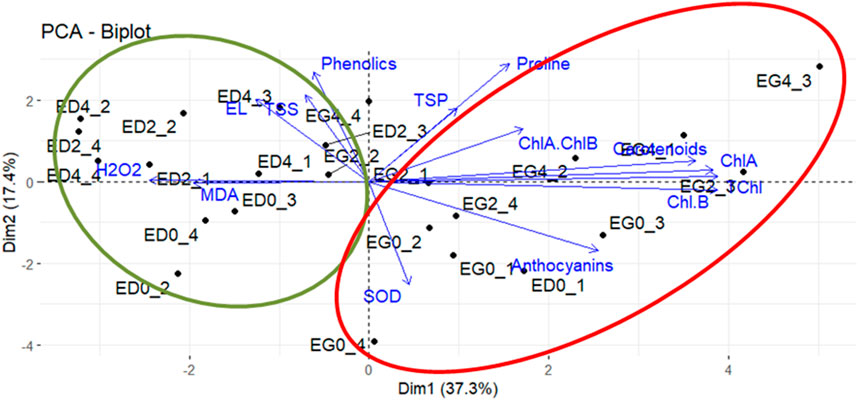
Figure 3. Principal components analysis. Biplot diagram for components 1 and 2 show sample observations and biochemical parameters. Vector lengths (original variables) indicates the strength of the relationship, and the angle between two vectors shows the degree of correlation. Ellipses were drawn to highlight sample observations (green ellipse: Eucalyptus dunnii; red ellipse: Eucalyptus grandis). Eucalyptus grandis control (EG0_1-EG0_4), Eucalyptus grandis on the second day of stress (EG2_1-EG2_4), Eucalyptus grandis on the fourth day of stress (EG4_1-EG4_4), Eucalyptus dunnii control (ED0_1-ED0_4), Eucalyptus dunnii on the second day of stress (ED2_1-ED2_4), Eucalyptus dunnii on the fourth day of stress (ED4_1-ED4_4).
PC2 explained 17.4% of the variance and pinpointed cold treatments. EL%, TSS phenolics, TSP, proline, carotenoids and ChlA/ChlB were positively correlated with PC2 and pointed to day 2 and day 4. PC2 was negatively correlated with variables such as anthocyanins and SOD activity, associated with non-stressed condition (i.e., day zero) (Figure 3).
Metabolomic analysis of phenolics compounds
A metabolomic analysis focused on obtaining a phenolic-focused fingerprint was performed to further disentangle the relationship between Eucalyptus species and cold treatments. The PLS-DA (8 LVs) score plot based on UHPLC-MS fingerprints revealed a clear separation between E. grandis and E. dunnii samples, as well as a distinction among stress days (days 0 and 2 vs. day 4) (Figure 4). These findings are further supported by the figure of merit in Supplementary Table S1, which shows 100% sensitivity and specificity, along with a classification accuracy of 100% within each sample group.
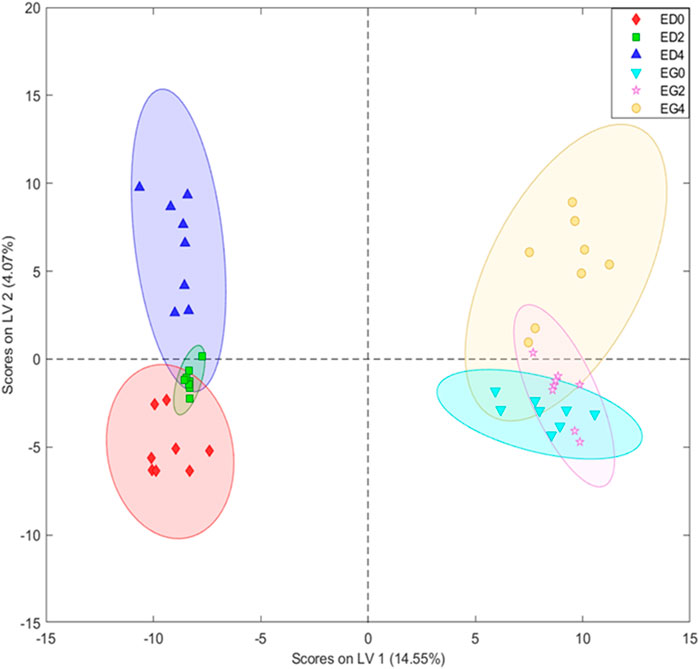
Figure 4. Partial least squares-discriminant analysis (PLS-DA) with eight latent variables (LVs) scores of the UHPLC-MS datasets derived from leaf disks extracts of Eucalyptus dunnii and Eucalyptus grandis maintained for 24 h at 25°C to simulate normal growth conditions and 4 days at 6°C to induce cold stress. Sampling was conducted at 0 h (control treatment), 2 and 4 days. Eucalyptus grandis control (EG0), Eucalyptus grandis on the 2nd day of stress (EG2), Eucalyptus grandis on the 4th day of stress (EG4), Eucalyptus dunnii control (ED0), Eucalyptus dunnii on the 2nd day of stress (ED2), Eucalyptus dunnii on the 4th day of stress (ED4).
Additionally, the UHPLC-ESI-IT-MS2 experiments in data-dependent mode allowed the putative identification of 16 significant phenolic compounds with high discriminant potential to differentiate the cold response in these two species. In total, seven flavonols, four ellagitannins, two gallotanins, one flavone, one phenolic acid and one cyclitol phenolic were identified (Supplementary Figure S1). These metabolite differences constitute potential markers of cold response differences.
Seven distinct flavonols were detected in peaks L5, L7, L9, L12, L13, L14, and L15. Notably, these included myricetin glucuronide (L5), myricetin hexoside (L7), quercetin galloyl hexoside (L9), quercetin glucuronide (L12), kaempferol glucuronide (L13), kaempferol (L14), and quercetin deoxyhexoside (L15) (Table 2). Four ellagitannins were identified in peaks L1, L4, L6, and L8. These were characterized as HHDP-galloyl-hexoside (L1), ellagic acid hexoside (L4), ellagic acid pentoside (L6), and HHDP digalloyl hexoside (L8). Two gallotannins were detected in peaks L3 and L11, corresponding to trigalloylhexoside and tetragalloylhexoside, respectively. Finally, a single cyclitol phenolic, phenolic acid and flavone were identified in peaks L2, L10, and L16, respectively. Tentative identifications suggest coumaroylquinic acid as the cyclitol phenolic, ellagic acid as the phenolic acid, and apigenin glucuronide as the flavone.
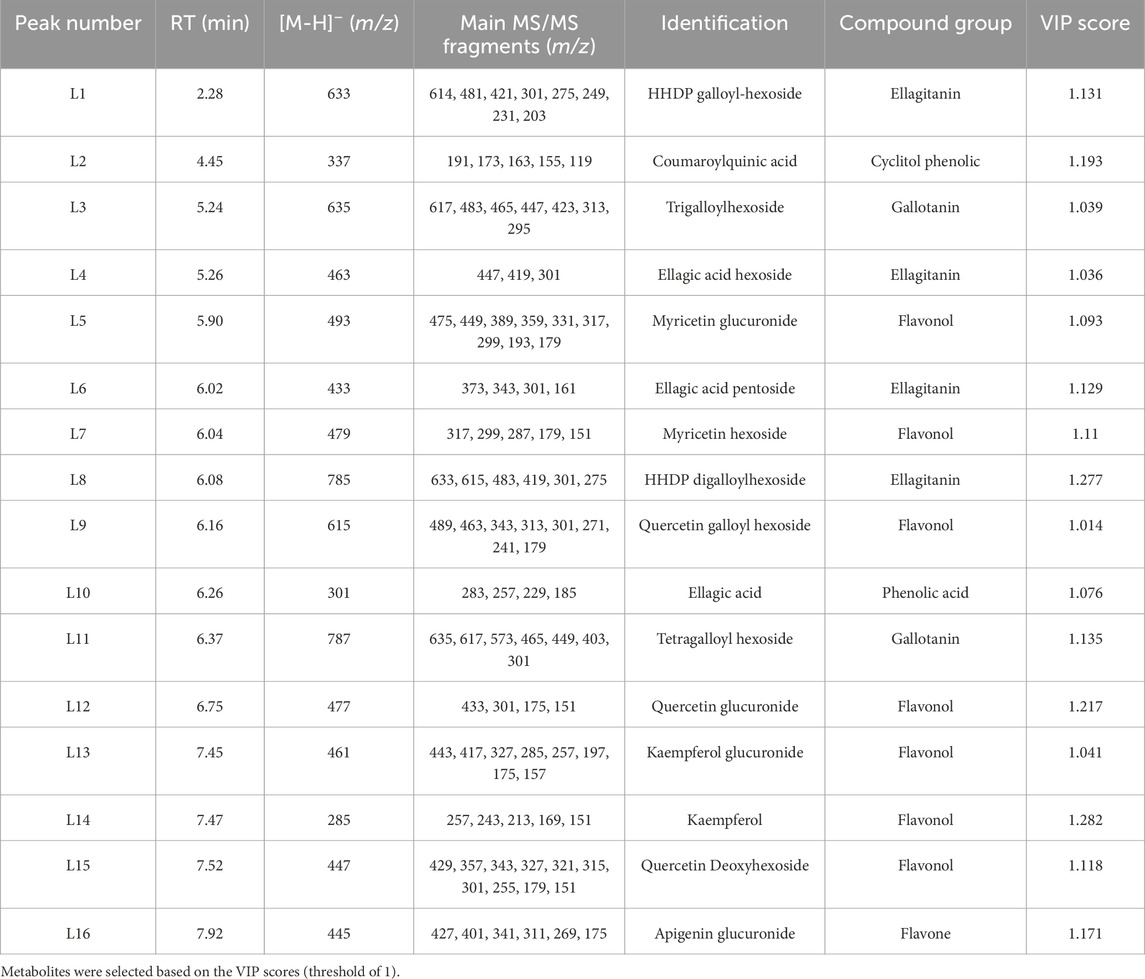
Table 2. Peak assignments in Eucalyptus dunnii and Eucalyptus grandis leaf extracts by LC-ESI-IT-MS2 in negative ionization mode.
To further explore the metabolic differences, heatmap and hierarchical clustering analyses were done for the 16 tentatively identified metabolites. These analyses revealed distinct patterns associated with species and cold treatment (Figure 5). Two clusters emerged from the hierarchical clustering. The first cluster grouped all E. grandis treatments (EG0, EG2, and EG4), while the second cluster contained only the E. dunnii 2-day cold treatment (ED2) (Figure 5, bottom dendrogram).
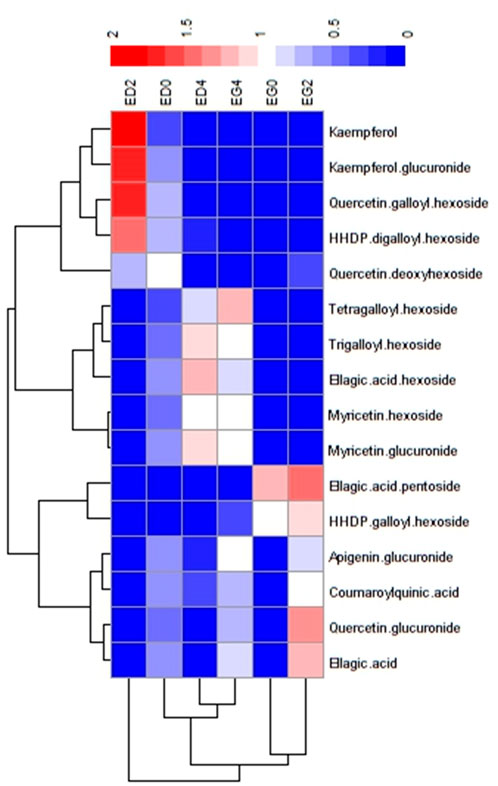
Figure 5. Heatmap representation of the putative identified metabolites significantly discriminant between species and treatments. Data was clustered by samples and metabolites.
Metabolites were categorized into three groups based on abundance patterns. Group I comprised metabolites that were significantly more abundant in the E. dunnii control (ED0) and 2-day cold treatment (ED2) like kaempferol, kaempferol glucuronide, quercetin galloyl hexoside, quercetin deoxyhexoside and HHDP digalloylhexoside (Figure 5, lateral dendrogram). Group II included metabolites more abundant in the 4-day cold treatment of both species (ED4 and EG4) such as myricetin glucuronide, myricetin hexoside, ellagic acid hexoside, trigalloyl hexoside, and tetragalloylhexoside. Finally, Group III consisted of metabolites that were more abundant in E. grandis control (EG0) and 2-day cold treatment (EG2), like ellagic acid, quercetin glucuronide, coumaroylquinic acid, apigenin glucuronide, HHDP galloyl hexoside and ellagic acid pentoside.
Discussion
EL%, MDA, H2O2, and SOD
Over the experimental period, EL% increased only in E. dunnii after 4 days of cold stress, while E. grandis remained unaffected. Similarly, timewise the lipid peroxidation end-product MDA and H2O2 amounts did not exhibit significant changes in either species. These observations contrast with those reported by Liu et al. (2014) for E. dunnii, in which H2O2 levels increased by 50% after 3 h at 4°C. This discrepancy may be a function of the distinct time scales of sampling used in the two studies (hours vs. days).
Overall data from this study indicated significant interspecific differences in a limited number of parameters. Whereas no significant temporal variations in MDA and H2O2 concentrations were observed between species, E. dunnii exhibited elevated lipid peroxidation (MDA) and H2O2 amounts on day 4. However, E. dunnii appeared to maintain relatively higher basal levels of these parameters in the preceding days.
Given the absence of increased lipid peroxidation or H2O2 content throughout the assay time, the elevated EL% observed in E. dunnii could primarily be the result of K+ efflux rather than membrane damage (Palta et al., 1977; Demidchik et al., 2014; De Palma et al., 2023). Nonetheless, the loss of K+ may affect cell turgor and disturb growth. Alternatively, the observed increase in EL% might be attributed to alterations in membrane fluidity induced by low temperatures. In this context, a reduction in membrane fluidity is known to enhance membrane permeability, facilitating the efflux of water and solutes (Xin and Browse, 2000).
The activity of SOD remained unchanged in E. dunnii and E. grandis. These data differ from the findings of Sales et al. (2013) and Oberschelp et al. (2020) which observed an increase in SOD activity under chilling and freezing temperatures. As SOD serves as a primary defense against superoxide radicals generated during photoinhibition (Mittler, 2002) by converting them to H2O2, the stable levels of the latter could be at least partly linked to the SOD activity profile.
Photosynthetic pigments
Under low-temperature conditions, all major components of photosynthesis, including thylakoid electron transport, the carbon reduction cycle and control of stomatal conductance, can be disrupted (Allen and Ort, 2001; Butnor et al., 2019). Plants adjust the composition of their antenna complexes and pigments to avoid photooxidative damage caused by the overproduction of ROS (Ort, 2001). In the present study, there were no significant alterations in chlorophyll and carotenoid concentrations in either species throughout the experimental period. This observation suggests defense mechanisms against cold stress were effective in both species.
A common plant strategy to mitigate photodamage involves reducing photosystem size (Mokochinski et al., 2018; Schimpl et al., 2018; Oberschelp et al., 2020; Oberschelp et al., 2022). However, our study suggested that the two species examined did not appear to employ this mechanism, as E. grandis displayed higher chlorophyll amount (TChl, ChlA, ChlB) on days 2 and 4, while these parameters remained stable in E. dunnii. On the other hand, the absence of significant chlorophyll loss is an indicator of stress tolerance by these species.
TSP, proline, and sugars
Regarding potential protective mechanisms, a notable increase in total soluble protein (TSP) content was observed in E. grandis on day 4, while it remained relatively stable in E. dunnii. This disparity suggests distinct protein synthesis strategies between the two species. The elevated TSP in E. grandis may be associated with the synthesis of regulatory and functional proteins, such as Late Embryogenesis-abundant (LEA) proteins, Heat Shock Proteins (HSPs), Cold-Inducible Proteins (KINs), and other cold stress-related proteins (Seki et al., 2002).
Proline, a compatible osmolyte known to accumulate in plant cells, offers protection against dehydration and lipid peroxidation under various abiotic stresses (Verslues and Sharma, 2010). In this study, both species exhibited increased proline amounts over time, consistent with previous reports (Kaplan et al., 2007; Liu et al., 2014; Primo-Capella et al., 2022). However, the peak accumulation occurred earlier in E. dunnii (day 2) compared to E. grandis (day 4).
Unlike E. dunnii, E. grandis showed a pronounced increase in proline content on day 4, suggesting its pivotal role in this species’ stress response. Similarly, E. grandis displayed higher anthocyanin content on day 4. Despite a report that did not detect significant interspecific differences in anthocyanin content between E. grandis and E. dunnii (Oberschelp et al., 2020), profiles of these flavonoids are known to vary with clone type within species.
On the second day, E. dunnii exhibited significantly higher levels of TSS and phenolics than E. grandis. Our findings regarding TSS are consistent with those of Oberschelp et al. (2020) but diverge from the results of Floriani et al. (2013), who did not observe significant differences between the two species. Conversely, our results for phenolics contradict those of Oberschelp et al. (2020), as they reported no significant differences on the concentration of these metabolites between E. grandis and E. dunnii.
Akin to proline, sugars also act as compatible osmolytes, playing an essential role in protecting cells from chilling injury (Couée et al., 2006; Yuanyuan et al., 2009; Tarkowski and Wim, 2015). In the present work, TSS concentration remained relatively stable among days in both species. In contrast, other authors have observed increased sugar amount after 5 days of cold treatment (Mokochinski et al., 2018; Oberschelp et al., 2022). These results could be explained by shorter cold exposure in the current experiments, variations in stress intensity or different relative importance of defense mechanisms in the clones evaluated.
Total phenolics and anthocyanins
Plants possess diverse light-absorbing metabolites, phenolic compounds, including anthocyanins, which protect photosynthetic tissues from photooxidative stress caused by excessive light energy. These metabolites function as photochemical energy dissipators, converting excess absorbed light into heat. This mechanism provides an alternative to the xanthophyll cycle, offering an additional layer of photoprotection (Close and Beadle, 2003; Tarkowski and Wim, 2015; Naing et al., 2017; Gould et al., 2018; Kumar et al., 2023).
In the present work, anthocyanin concentration did not change between days of cold treatment in any of the species. Anthocyanins confer a beneficial role when plants are unable to dissipate excess thermal energy and require additional mechanisms to quench excessive light, particularly under prolonged exposure to intense full-spectrum white light or natural sunlight at low temperatures (Gould et al., 2018). Our study did not detect significant changes in chlorophyll or carotenoid amounts in either E. grandis or E. dunnii across the experimental days. These findings suggest that the existing pigment quantity may be sufficient to prevent photodamage. Alternatively, the relatively low irradiance intensity and sample harvest frequency may not have been sufficient to afford measurable changes in anthocyanin levels.
The concentration of phenolics increased in E. dunnii, while levels remained stable in E. grandis across the experimental days. The elevated phenolic levels in E. dunnii suggest that the accumulation of these compounds may represent a significant mechanism for mitigating photoinhibition in this species. These results differ from those reported by Oberschelp et al. (2020) and Oberschelp et al. (2022), who found an increase in phenolics in both species under cold stress conditions. Again, this may reflect distinct experimental conditions and/or genetic differences in defense strategies among eucalypt commercial clones.
Comparison of cold-response profiles and phenolic metabolomics
Our results indicate distinct cold tolerance strategies in the clones of E. grandis and E. dunnii. Notably, the timing of protective mechanism activation differed between the two species. Eucalyptus dunnii initiated these responses after a 2-day exposure period, whereas E. grandis did so only after 4 days. Furthermore, the specific protective parameters deployed by each species also differed. Data point out to E. dunnii primarily relying on the accumulation of sugars and phenolic compounds, with E. grandis favoring accumulation of proline and anthocyanins.
Indeed, Principal Component Analysis (PCA) of the biochemical parameter dataset further corroborates these divergent cold tolerance strategies between the two species. In line with this finding, the Partial Least Squares-Discriminant Analysis (PLS-DA) score plot derived from metabolomic analysis revealed a clear species-specific effect across the metabolome. Furthermore, this effect was greater than the differences between cold temperatures.
The phenolic compounds primarily responsible for the observed discrimination were flavonols, ellagitannins, gallotannins, flavones, phenolic acids, and cyclitol phenolics, where flavonols constituted the most abundant class of identified metabolites (Supplementary Figure S1). All these metabolites have been previously reported in Eucalyptus leaves (Mokochinski et al., 2018; Noleto-Dias et al., 2023) and wood (Santos et al., 2013). Evidence that phenolic compounds are temperature-responsive metabolites was previously shown by Mokochinski et al. (2018), comparing E. pellita, E. grandis, and E. dunnii. Some polyphenols identified by the authors’ metabolomic analysis increased after low temperature exposure, particularly in the case of E. dunnii. In the present study E. dunnii 2-day cold treatment showed an accumulation of kaempferol and quercetin glycosides. These flavonols are known to absorb UVB light and play a significant role in photodamage avoidance (Ryan et al., 2002). Flavonols can contribute to mitigate damage caused by cold-induced photoinhibition (Mishra et al., 2023). Indeed, accumulation of flavonols and phenolic acids is a well-established adaptive response to chilling in plants (Rao and Zheng, 2025).
Taken together, overall data highlight that the accumulation of TSS and phenolic compounds is a distinctive cold tolerance strategy of E. dunnii and that phenolic fingerprint under cold stress differs in E. dunnii and E. grandis. Moreover, the latter species seems to rely more intensely on proline and anthocyanin accumulation.
Considerations on distinct methodology comparisons
The leaf disk-based method herein described is considerably faster and demands less infrastructure for its implementation. Similar approaches to investigate the impact of tree stress on secondary metabolism have been used in previous studies, mostly showing agreement with whole plant data (e.g., De Costa et al., 2013; Palma et al., 2023). A leaf disk method also proved valuable to estimate frost tolerance variability in Eucalyptus regnans F. Muell. and Eucalyptus nitens (Deane & Maiden) Maiden as a tool for breeding purposes (Raymond et al., 1992). However, like any other experimental method, it is not free of some limitations (Piotrowska and Kacperska, 1990). Among these, one could list the lack of correlative influences among plant organs, the absence of whole plant metabolic coordination, and the non-adequacy to long time span assays. Nonetheless, useful as well as novel information can emerge from the approach used in the present study, particularly for direct comparisons, as described above. Indeed, several consistencies with available literature regarding cold responses in eucalypts were recorded. Admittedly, caution should be always exercised before extrapolating data from simplified to more complex experimental approaches (e.g., organs vs. whole plants, laboratory versus field tests).
Conclusion
Distinct cold tolerance biochemical strategies were found in E. grandis and E. dunnii leaf tissues exposed to mild temperate climate winter cold conditions. These differences encompass both the specific protective agents employed by each species and the timing of protective mechanism deployment. Whereas E. dunnii appeared to rely predominantly on the accumulation of sugars and phenolic compounds, the accumulation of proline and anthocyanins was primarily present in E. grandis. Moreover, 16 phenolic compounds with high discriminant potential were identified, allowing for the differentiation of the cold response in these two species and providing potential future tools for cold tolerance screening. Further studies will delve in detailed exploration of the metabolites and genes taking part in cold tolerance responses in commercial clones of these high interest forest species.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
PB: Conceptualization, Formal Analysis, Investigation, Project administration, Writing – original draft, Writing – review and editing. FW: Formal Analysis, Investigation, Writing – review and editing. CO: Formal Analysis, Resources, Writing – review and editing. ND: Investigation, Writing – review and editing. OB: Conceptualization, Methodology, Project administration, Writing – review and editing. AF: Conceptualization, Methodology, Project administration, Resources, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by the Comisión Sectorial de Investigación Científica (CSIC)-Programa Iniciación a la Investigación 2021 (22320200200271UD), Universidad de la República (Uruguay), the Programa de Desarrollo de las Ciencias Básicas (PEDECIBA), and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq-Brazil, grant 310775/2021-3).
Acknowledgments
The authors gratefully acknowledge the contribution of Cibele Tesser Da Costa of the Plant Physiology Laboratory, Center for Biotechnology, Federal University of Rio Grande do Sul (UFRGS), Porto Alegre, Brazil, for assistance with laboratory analyses. We would also acknowledge the support of the Agencia Nacional de Investigación e Innovación (ANII-POS_NAC_2021_1_170869).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmolb.2025.1584132/full#supplementary-material
References
Allen, D. J., and Ort, D. R. (2001). Impacts of chilling temperatures on photosynthesis in warm-climate plants. Trends Plant Sci. 6 (1), 36–42. doi:10.1016/S1360-1385(00)01808-2
Arnold, R., Li, B., Luo, J., Bai, F., and Baker, T. (2015). Selection of cold-tolerant Eucalyptus species and provenances for inland frost-susceptible, humid subtropical regions of southern China. Aust. For. 78 (3), 180–193. doi:10.1080/00049158.2015.1063471
Arnold, R. J., Clarke, B., and Luo, J. (2004). “Trials of cold-tolerant eucalypt species in cooler regions of south Central China,” in Australian Center for International Agricultural Research Technical Reports no. 57, 106. doi:10.22004/ag.econ.113922
Beyer, W. F., and Irwin, F. (1987). Assaying for superoxide dismutase activity: some large consequences of minor changes in conditions. Anal. Biochem. 161 (2), 559–566. doi:10.1016/0003-2697(87)90489-1
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72 (7), 248–254. doi:10.1016/0003-2697(76)90527-3
Butnor, J. R., Johnsen, K. H., Anderson, P. H., Hall, K. B., Halman, J. M., Hawley, G. J., et al. (2019). Growth, photosynthesis, and cold tolerance of Eucalyptus benthamii planted in the piedmont of North Carolina. For. Sci. 65 (1), 59–67. doi:10.1093/forsci/fxy030
Chow, P. S., and Landhäusser, S. M. (2004). A method for routine measurements of total sugar and starch content in woody plant tissues. Tree Physiol. 24 (10), 1129–1136. doi:10.1093/treephys/24.10.1129
Close, D. C., and Beadle, C. L. (2003). Alternate energy dissipation? Phenolic metabolites and the xanthophyll cycle. J. Plant Physiol. 160 (4), 431–434. doi:10.1078/0176-1617-00915
Close, D. C., Beadle, C. L., and Battaglia, M. (2004). Foliar anthocyanin accumulation may be a useful indicator of hardiness in eucalypt seedlings. For. Ecol. Manag. 198 (1–3), 169–181. doi:10.1016/j.foreco.2004.03.039
Couée, I., Sulmon, C., Gouesbet, G., and Amrani, A. E (2006). Involvement of soluble sugars in reactive oxygen species balance and responses to oxidative stress in plants. J. Exp. Bot. 57 (3), 449–459. doi:10.1093/jxb/erj027
Davidson, N. J., Battaglia, M., and Close, D. C. (2004). Photosynthetic responses to overnight frost in Eucalyptus nitens and E. Globulus. Trees - Struct. Funct. 18 (3), 245–252. doi:10.1007/s00468-003-0298-3
De Costa, F., Yendo, A. C. A., Fleck, J. D., Grace, G., and Fett-Neto, A. G. (2013). Accumulation of a bioactive triterpene saponin fraction of quillaja brasiliensis leaves is associated with abiotic and biotic stresses. Plant Physiology Biochem. 66, 56–62. doi:10.1016/j.plaphy.2013.02.003
De Palma, N., Yendo, A. C. A., Vilasboa, J., Chacon, D. S., and Fett-Neto, A. G. (2023). Biochemical responses in leaf tissues of alkaloid producing psychotria brachyceras under multiple stresses. J. Plant Res. 136 (3), 397–412. doi:10.1007/s10265-023-01441-z
Demidchik, V., Straltsova, D., Medvedev, S. S., Pozhvanov, G. A., Sokolik, A., and Yurin, V. (2014). Stress-induced electrolyte leakage: the role of K+-Permeable channels and involvement in programmed cell death and metabolic adjustment. J. Exp. Bot. 65 (5), 1259–1270. doi:10.1093/jxb/eru004
Domingues, A. P., Daloso, D. de M., Machado, M., Rosado-Souza, L., Perez de Souza, L., Fernie, A. R., et al. (2019). A cold change: how short low temperature exposure affects primary metabolism in leaves and stems of two Eucalyptus species. Theor. Exp. Plant Physiol. 31 (4), 429–444. doi:10.1007/s40626-019-00156-8
Floriani, M. M. P., André Steffens, C., Miler Chaves, D., Talamini do Amarante, C. V., Pikart, T. G., and dos Santos Ribeiro, M. (2013). Relação entre concentrações foliares de carboidratos solúveis totais e tolerância ao frio em diferentes espécies de Eucalyptus spp. Cienc. Florest. 23 (1), 165–174. doi:10.5902/198050988450
Fürtauer, L., Weiszmann, J., Weckwerth, W., and Nägele, T. (2019). Dynamics of plant metabolism during cold acclimation. Int. J. Mol. Sci. 20 (21), 5411. doi:10.3390/ijms20215411
Gould, K. S., Jay-Allemand, C., Logan, B. A., Baissac, Y., and Bidel, L. P. R. (2018). When are foliar anthocyanins useful to plants? Re-evaluation of the photoprotection hypothesis using arabidopsis thaliana mutants that differ in anthocyanin accumulation. Environ. Exp. Bot. 154, 11–22. doi:10.1016/j.envexpbot.2018.02.006
Guo, X., Liu, D., and Chong, K. (2018). Cold signaling in plants: insights into mechanisms and regulation. J. Integr. Plant Biol. 60 (9), 745–756. doi:10.1111/jipb.12706
Janská, A., Maršík, P., Zelenková, S., and Ovesná, J. (2010). Cold stress and acclimation - what is important for metabolic adjustment? Plant Biol. 12 (3), 395–405. doi:10.1111/j.1438-8677.2009.00299.x
Jovanovic, T., Arnold, R., and Booth, T. (2000). Determining the climatic suitability of Eucalyptus dunnii for plantations in Australia, China and central and south America. New For. 19 (3), 215–226. doi:10.1023/A:1006662718206
Juszczak, I., Cvetkovic, J., Zuther, E., Hincha, D. K., Baier, M., Peter, N., et al. (2016). Natural variation of cold deacclimation correlates with variation of cold-acclimation of the plastid antioxidant system in Arabidopsis thaliana accessions. Front. Plant Sci. 7 (March), 305–317. doi:10.3389/fpls.2016.00305
Kaplan, F., Kopka, J., Sung, D. Y., Zhao, W., Popp, M., Porat, R., et al. (2007). Transcript and metabolite profiling during cold acclimation of arabidopsis reveals an intricate relationship of cold-regulated gene expression with modifications in metabolite content. Plant J. 50 (6), 967–981. doi:10.1111/j.1365-313X.2007.03100.x
Knaupp, M., Mishra, K. B., Nedbal, L., and Heyer, A. G. (2011). Evidence for a role of raffinose in stabilizing photosystem II during freeze-thaw cycles. Planta 234 (3), 477–486. doi:10.1007/s00425-011-1413-0
Kottek, M., Grieser, J., Beck, C., Rudolf, B., and Rubel, F. (2006). World map of the Köppen-Geiger climate classification updated. Meteorol. Z. 15 (3), 259–263. doi:10.1127/0941-2948/2006/0130
Król, A., Amarowicz, R., and Weidner, S. (2015). The effects of cold stress on the phenolic compounds and antioxidant capacity of grapevine (Vitis vinifera L.) leaves. J. Plant Physiol. 189, 97–104. doi:10.1016/j.jplph.2015.10.002
Kumar, K., Debnath, P., Singh, S., and Kumar, N. (2023). An overview of plant phenolics and their involvement in abiotic stress tolerance. Stresses 3 (3), 570–585. doi:10.3390/stresses3030040
Lee, Mi R., Kim, C. S., Park, T., Choi, Y. S., and Lee, K. H. (2018). Optimization of the ninhydrin reaction and development of a multiwell plate-based high-throughput proline detection assay. Anal. Biochem. 556, 57–62. doi:10.1016/j.ab.2018.06.022
Liu, Y., Jiang, Y., Lan, J., Zou, Y., and Gao, J. (2014). Comparative transcriptomic analysis of the response to cold acclimation in Eucalyptus dunnii. PLoS ONE 9 (11), 113091–e113111. doi:10.1371/journal.pone.0113091
Makkar, H. P. S. (2000). Quantification of tannins in tree and shrub foliage. Quantification Tannins Tree Shrub Foliage. doi:10.1007/978-94-017-0273-7
Mishra, A., Mishra, K. B., Surá, K., Veselá, B., Klem, K., and Urban, O. (2023). Non-photochemical quenching in natural accessions of Arabidopsis thaliana during cold acclimation. Environ. Exp. Bot. 211 (March), 105372. doi:10.1016/j.envexpbot.2023.105372
Mittler, R. (2002). Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 7 (9), 405–410. doi:10.1016/S1360-1385(02)02312-9
Mokochinski, J. B., Mazzafera, P., Sawaya, A. C. H. F., Mumm, R., Hall, R. D., and Hall, R. D. (2018). Metabolic responses of Eucalyptus species to different temperature regimes. J. Integr. Plant Biol. 60 (5), 397–411. doi:10.1111/jipb.12626
Murashige, T., and Skoog, F. (1962). A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 15, 473–497. doi:10.1111/j.1399-3054.1962.tb08052.x
Naing, A. H., Park, K.Il, Ai, T. N., Chung, Mi Y., Han, J. S., Kang, Y. W., et al. (2017). Overexpression of snapdragon delila (del) gene in tobacco enhances anthocyanin accumulation and abiotic stress tolerance. BMC Plant Biol. 17 (1), 65–14. doi:10.1186/s12870-017-1015-5
Noleto-Dias, C., Picoli, E. A. de T., Porzel, A., Wessjohann, L. A., Tavares, J. F., and Farag, M. A. (2023). Metabolomics characterizes early metabolic changes and markers of tolerant Eucalyptus Ssp. clones against drought stress. Phytochemistry 212 (February), 113715. doi:10.1016/j.phytochem.2023.113715
Oberschelp, G. P. J., Beatriz Guarnaschelli, A., Teson, N., Harrand, L., Podestá, F. E., and Margarit, E. (2020). Cold acclimation and freezing tolerance in three Eucalyptus species: a metabolomic and proteomic approach. Plant Physiol. Biochem. 154 (May), 316–327. doi:10.1016/j.plaphy.2020.05.026
Oberschelp, G. P. J., Morales, L. L., Montecchiarini, M. L., Harrand, L., Podestá, F. E., and Margarit, E. (2022). Harder, better, faster, stronger: frost tolerance of Eucalyptus benthamii under cold acclimation. Plant Physiol. Biochem. 186 (April), 64–75. doi:10.1016/j.plaphy.2022.06.022
Palta, J. P., Jacob, L., and Stadelmann, E. J. (1977). Freezing injury in onion bulb cells. Plant Physiol. 60 (3), 398–401. doi:10.1104/pp.60.3.398
Pennycooke, J. C., Cox, S., and Stushnoff, C. (2005). Relationship of cold acclimation, total phenolic content and antioxidant capacity with chilling tolerance in petunia. Environ. Exp. Bot. 53 (2), 225–232. doi:10.1016/j.envexpbot.2004.04.002
Piotrowska, G., and Kacperska, A. (1990). Utility of leaf disks cultured in vitro for studies on frost resistance. Plant Cell, Tissue Organ Cult. 22 (1), 21–26. doi:10.1007/BF00043694
Primo-Capella, A., Forner-Giner, M. Á., Martínez-Cuenca, M. R., and Terol, J. (2022). Comparative transcriptomic analyses of citrus cold-resistant vs. sensitive rootstocks might suggest a relevant role of ABA signaling in triggering cold scion adaption. BMC Plant Biol. 22 (1), 1–26. doi:10.1186/s12870-022-03578-w
R Core Team (2023). R: a language and environment for statistical computing. (R Foundation for Statistical Computing).
Rao, M. J., and Zheng, B. (2025). The role of polyphenols in abiotic stress tolerance and their antioxidant properties to scavenge reactive oxygen species and free radicals. Antioxidants 14 (1), 74. doi:10.3390/antiox14010074
Raymond, C. A., Owen, J. V., Eldridge, K. G., and Harwood, C. E. (1992). Screening eucalypts for frost tolerance in breeding programs. Can. J. For. Res. 22 (1), 1271–1277. doi:10.1139/x92-169
Ryan, K. G., Swinny, E. E., Markham, K. R., and Winefield, C. (2002). Flavonoid gene expression and UV photoprotection in transgenic and mutant petunia leaves. Phytochemistry 59 (1), 23–32. doi:10.1016/S0031-9422(01)00404-6
Sales, C. R. G., Ribeiro, R. V., Silveira, J. A. G., Machado, E. C., Martins, M. O., and Lagôa, A. M. M. A. (2013). Superoxide dismutase and ascorbate peroxidase improve the recovery of photosynthesis in sugarcane plants subjected to water deficit and low substrate temperature. Plant Physiol. Biochem. 73, 326–336. doi:10.1016/j.plaphy.2013.10.012
Santos, S. A. O., Vilela, C., Freire, C. S. R., Neto, C. P., and Silvestre, A. J. D. (2013). Ultra-high performance liquid chromatography coupled to mass spectrometry applied to the identification of valuable phenolic compounds from Eucalyptus wood. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 938, 65–74. doi:10.1016/j.jchromb.2013.08.034
Schimpl, F. C., Ribeiro, R. V., Pereira, L., Rodrigues, H. S., and Mazzafera, P. (2018). Photochemical responses to abrupt and gradual chilling treatments in Eucalyptus species. Theor. Exp. Plant Physiol. 30 (1), 9–17. doi:10.1007/s40626-018-0097-2
Schmid, R., Heuckeroth, S., Korf, A., Smirnov, A., Myers, O., Dyrlund, T. S., et al. (2023). Integrative analysis of multimodal mass spectrometry data in MZmine 3. Nat. Biotechnol. 41 (4), 447–449. doi:10.1038/s41587-023-01690-2
Seki, M., Narusaka, M., Ishida, J., Nanjo, T., Fujita, M., Oono, Y., et al. (2002). Monitoring the expression profiles of 7000 arabidopsis genes under drought, cold and high-salinity stresses using a full-length CDNA microarray. Plant J. 31 (3), 279–292. doi:10.1046/j.1365-313X.2002.01359.x
Sims, D. A., and Gamon, J. A. (2002). Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 81 (2–3), 337–354. doi:10.1016/S0034-4257(02)00010-X
Stanturf, J. A., Vance, E. D., Fox, T. R., and Kirst, M. (2013). Eucalyptus beyond its native range: environmental issues in exotic bioenergy plantations. Int. J. For. Res. 2013, 1–5. doi:10.1155/2013/463030
Tarkowski, Ł. P., and Wim, V. den E. (2015). Cold tolerance triggered by soluble sugars: a multifaceted countermeasure. Front. Plant Sci. 6 (APR), 1–7. doi:10.3389/fpls.2015.00203
Thalhammer, A., Hincha, D. K., and Zuther, E. (2014). “Measuring freezing tolerance: electrolyte leakage and chlorophyll fluorescence assays,” in Plant cold acclimation, methods in molecular biology, ed. D. Hincha, and E. Zuther (New York, NY: Humana Press), Vol. 1166, 33–44. doi:10.1007/978-1-4939-0844-8_3
Thomashow, M. F. (1999). Plant cold acclimation: freezing tolerance genes and regulatory mechanisms. Annu. Rev. Plant Biol. 50, 571–599. doi:10.1146/annurev.arplant.50.1.571
Velikova, V., Yordanov, I., and Edreva, A. (2000). Oxidative stress and some antioxidant systems in acid rain-treated bean plants. Plant Sci. 151 (1), 59–66. doi:10.1016/S0168-9452(99)00197-1
Verslues, P. E., and Sharma, S. (2010). Proline metabolism and its implications for plant-environment interaction. Arabidopsis Book 8 (8), e0140. doi:10.1199/tab.0140
Xin, Z., and Browse, J. (2000). Cold comfort farm: the acclimation of plants to freezing temperatures. Plant Cell Environ. 23 (9), 893–902. doi:10.1046/j.1365-3040.2000.00611.x
Yadav, S. K. (2010). Cold stress tolerance mechanisms in plants. A review. Agron. Sustain. Dev. 30, 515–527. doi:10.1051/agro/2009050
Keywords: Eucalyptus grandis, Eucalyptus dunnii, cold stress, biochemical responses, metabolomic
Citation: Basile P, Wallace F, Olivaro C, De Palma N, Borsani O and Fett-Neto A (2025) Cold-induced biochemical changes in leaves of two commercial clones of Eucalyptus. Front. Mol. Biosci. 12:1584132. doi: 10.3389/fmolb.2025.1584132
Received: 26 February 2025; Accepted: 21 May 2025;
Published: 04 June 2025.
Edited by:
Martin Aran, IIBBA-CONICET Leloir Institute Foundation, ArgentinaReviewed by:
Inês Maria Valente, LAQV Network of Chemistry and Technology, PortugalLiliana Beatriz Pena, University of Buenos Aires, Argentina
Copyright © 2025 Basile, Wallace, Olivaro, De Palma, Borsani and Fett-Neto. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Arthur Fett-Neto, ZmV0dC5uZXRvQHVmcmdzLmJy; Patricia Basile, cGF0cmljaWEuYmFzaWxlQGN1dC5lZHUudXk=