 Marius Wilhelm Baeken
Marius Wilhelm Baeken Fazilet Bekbulat
Fazilet Bekbulat Hagen Körschgen
Hagen Körschgen Albrecht Martin Clement
Albrecht Martin Clement Christian Behl
Christian Behl- Institute of Pathobiochemistry, The Autophagy Lab, University Medical Center of the Johannes Gutenberg University Mainz, Mainz, Germany
Gamma-aminobutyric acid receptor-associated protein (GABARAP) is a multifunctional member of the autophagy-related (ATG8) protein family, playing key roles in two distinct cellular pathways: macroautophagy and plasma membrane protein trafficking. In the context of autophagy, GABARAP modulates cargo recognition and supports the maturation and fusion of autophagosomes with lysosomes, a critical step in intracellular clearance and proteostasis. Separately, GABARAP also regulates vesicular receptor protein transport from the Golgi apparatus to the plasma membrane, contributing to proper surface localization and receptor recycling. Both tasks are especially vital for neurons, where protein turnover and receptor localization are tightly linked to synaptic plasticity and neuroprotection. We recently identified a direct interaction between GABARAP and the sigma-1 receptor (σ1R), an ER-resident receptor involved in diverse cellular stress responses, mitochondrial function, and protein homeostasis. Our findings suggest that σ1R acts as an upstream regulatory hub, influencing GABARAP’s functional commitment to either membrane trafficking or autophagy. Specifically, we hypothesize that ligand-dependent σ1R activation promotes GABARAP’s involvement in macroautophagy at the expense of its role in membrane transport. This regulatory switch may underline part of the neuroprotective effects observed with σ1R agonists in neurodegenerative disease models, where enhanced autophagy is often beneficial. Overall, we discuss the emerging molecular crosstalk between σ1R and GABARAP, its potential impact on neuronal homeostasis, and how σ1R’s pharmacological modulation might be leveraged to bias GABARAP function toward autophagy in diseases such as amyotrophic lateral sclerosis, Huntington’s, Parkinson’s, and Alzheimer’s disease.
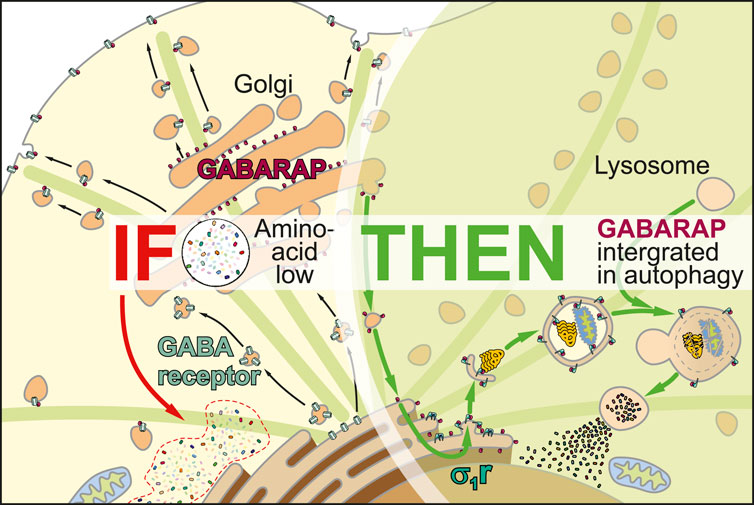
Introduction
Post-mitotic cells, especially neurons, have to co-orchestrate numerous delicate homeostatic systems, such as protein or ion homeostasis, to ensure organismal function. Hence, a cross-talk between these systems is essential.
We recently described the sigma-1 receptor (σ1R), a protein lacking any close homologs in the human proteome (Moebius et al., 1997), as a regulator that integrates gamma-aminobutyric acid receptor-associated protein (GABARAP) into macroautophagy (Baeken et al., 2025; Christ et al., 2019). In brief, σ1R-deficient HeLa cells were unable to incorporate GABARAP into autophagosomes, as evidenced by a complete loss of GABARAP-p62 co-localization and a breakdown of the GABARAP-associated autophagic flux (Baeken et al., 2025). However, we noticed that GABARAP’s lipidated form still persisted, which suggests an interaction with membranous structures. Interestingly, a previous study has shown that the G116A (amino acids in single letter code) GABARAP mutant, which cannot be C-terminally cleaved, was unable to transport GABAA receptors to the plasma membrane (Chen et al., 2007). This cleavage is a prerequisite for lipidation, which suggests that lipidation also facilitates the transport of associated vesicles.
Although still circumstantial, this evidence may broaden our understanding of σ1R’s physiological impact. As its bona fide cellular purpose remains elusive, we propose a regulatory role of σ1R in controlling GABARAP logistics, which affects neurotransmission as well as autophagy. In a broader context, this regulatory function might help explain the neuroprotective properties attributed to σ1R (Christ et al., 2020).
GABARAP’s role in mammalian macroautophagy
Macroautophagy (hereafter autophagy) is a highly conserved cellular degradation pathway, characterized by the formation of double-membraned vesicles – autophagosomes – that engulf cytosolic material and fuse with lysosomes, where contents are degraded by hydrolases and recycled for reuse (He and Klionsky, 2009). Dynamically regulated, autophagy enables cells to deal with changing demands, be it upholding energy homeostasis during nutrient deprivation or selectively removing misfolded proteins and damaged organelles. Thus, autophagy is an essential homeostatic and adaptive process promoting survival and renovation, especially for post-mitotic cells such as neurons (Mizushima and Komatsu, 2011). Autophagic efficiency declines with age and its dysfunction is associated with disease, including neurodegeneration (Aman et al., 2021; Palmer et al., 2025).
Autophagosome formation, maturation, and fusion with lysosomes are coordinated by core autophagy-related (ATG) proteins, including an initiation complex, a membrane expansion complex, and two ubiquitin-like conjugation systems, one being the LC3/GABARAP system (Ariosa and Klionsky, 2016). Microtubule-associated protein 1A/1B-light chain 3 (LC3) and GABARAP, both members of the ATG8-family, form two distinct subfamilies: LC3A, LC3B, and LC3C; and GABARAP, GABARAPL1, and GABARAPL2 (Schaaf et al., 2016).
ATG8 proteins are ubiquitously expressed, though GABARAPs are enriched in the central nervous system and endocrine glands (Lee and Lee, 2016). Although highly homologous, distinct residues mediate selectivity of GABARAP subfamily members towards interaction partners (Sugawara et al., 2004; Rogov et al., 2014); emerging evidence supports unique and complementary roles for LC3s and GABARAPs in autophagy. During autophagosome formation, LC3B is mainly involved in the elongation of the phagophore, general cargo recognition, and the transport of autophagosomes to lysosomes, while GABARAP has primarily been associated with late events such as autophagosome closure and autophagosome-lysosome fusion (Lee and Lee, 2016; Schaaf et al., 2016). Supporting its function in autophagosome maturation, GABARAP shows a higher affinity to Unc-51 like autophagy activating kinase 1 (ULK1) (Alemu et al., 2012), and mediates autophagosomal targeting of phosphatidylinositol 4-kinase type 2 alpha (PI4KIIα), thereby regulating autophagosome-lysosome fusion (Albanesi et al., 2015).
To function in autophagy, cytosolic ATG8s must undergo lipidation, i.e., a covalent conjugation to phosphatidylethanolamine (PE), enabling incorporation into autophagosomal membranes (Lee and Lee, 2016; Nguyen et al., 2016; Palmer et al., 2025). Initially synthesized as precursors, ATG8s are cleaved by cysteine endopeptidases of the ATG4 family, whereby a C-terminal glycine residue becomes accessible for lipidation (Nguyen et al., 2020). The exposed glycine is then adenylated, sequentially transferred through E1-like and E2-like ATG proteins, and finally conjugated to PE’s amino head group via an E3-like ATG conjugation complex (Maruyama and Noda, 2017; Martens and Fracchiolla, 2020).
One hallmark of autophagosome-mediated autophagy is cargo-specific sequestration, in which adaptor proteins bind distinct ubiquitinated cargo and directly interact with inner autophagosomal membrane-bound ATG8 proteins (Khaminets et al., 2016), enabling selective elimination of various cellular components. The latter interaction is assured by distinct binding motifs termed LC3-interacting regions (LIRs), which are recognized by LIR docking sites in ATG8s (Johansen and Lamark, 2020).
The core of the canonical LIR is defined by the hydrophobic consensus sequence [W/F/Y]-X1-X2-[L/V/I] (X for variable residue) (Birgisdottir et al., 2013). Its affinity is enhanced by phosphorylated or acidic residues flanking the motif, albeit preferentially on the N-terminus (Johansen and Lamark, 2020). The ubiquitin-binding protein p62, considered a bona fide cargo adaptor, directly binds to all ATG8s (Pankiv et al., 2007; Johansen and Lamark, 2020). Specificity of cargo adaptors toward particular ATG8s arises from unique hydrophobic contacts, salt bridges, and hydrogen bonds. Notably, selectivity towards GABARAP is mediated by a restriction of this core motif: [W/F]-[V/I]-X-V (Rogov et al., 2017; Rogov et al., 2023).
GABARAP’s role in receptor transport and deployment
Although widely known for its integration into autophagic processes, GABARAP was originally identified as a cellular agent to properly distribute GABAA receptors to the plasma membrane by interacting with the γ2 subunit (GABRG2) of the heteropentamer (Wang et al., 1999). Following studies confirmed that GABARAP interacts with γ1, but not with the γ3 or δ subunits in yeast hybrid systems (Kneussel et al., 2000). The primary interaction site of GABARAP with GABAA points to an intracellular loop between transmembrane regions 3 and 4 within the γ1/γ2 subunits (Nymann-Andersen et al., 2002; Chen et al., 2000). This interaction is facilitated by a classical LIR motif (414YECL417) (Ye et al., 2021). This core motif is also present in γ3 but lacks the favored acidic N-terminal residues (Rogov et al., 2017). Interestingly, Y412 in γ2 is phosphorylated by FYN and SRC kinases (Jurd et al., 2010), presumably regulating its affinity (Kittler et al., 2008). While Y414 can also be phosphorylated, the consequence for GABARAP binding remains unclear (Jurd et al., 2010). However, our in silico model suggests that this modification might affect binding negatively, whereas unmodified Y414 or pY412 would bind GABARAP (Figure 1A). Here, pY412 could enable intramolecular interaction with H443, condensing the C-terminal structure of γ2 and potentially stabilizing GABARAP binding.
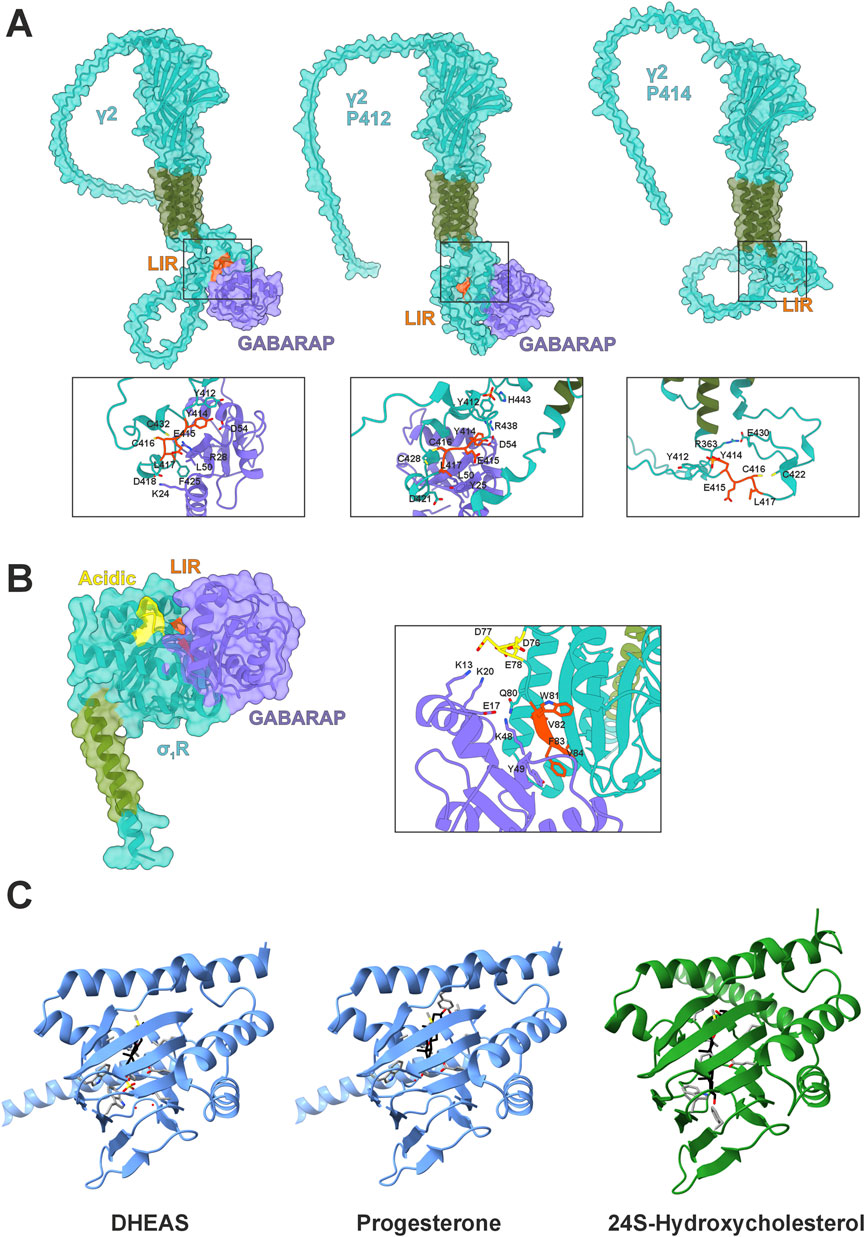
Figure 1. GABARAP interactions and σ1R structural context. (A) AlphaFold3 models of GABARAP (purple) in complex with the GABAA receptor subunit γ2 (teal with green transmembrane domain) with highlighted LIR (orange) in their native states including phosphorylated tyrosines Y412/Y414. (B) AlphaFold3 model of GABARAP (purple) in complex with σ1R (teal with green transmembrane domain), highlighting the LIR domain (orange) and acidic residues (yellow). (C) Crystal structure of Xenopus laevis σ1R (blue) with DHEAS (pdb:8WUE) or progesterone (pdb:8W4C) bound and Boltz-2 predicted interaction of Homo sapiens σ1R (green) with 24S-hydroxycholesterol.
Furthermore, GABARAP is involved in maintaining membrane GABAA localization by several mechanisms. (i) GABARAP can negatively regulate GABAA receptor reinternalization. This process relies on binding of the adaptor protein complex 2 (AP2) to the γ2 subunit (Comenencia-Ortiz et al., 2014). Coincidentally, GABARAP and AP2 compete for the same binding sites (Ye et al., 2021; Kittler et al., 2008). (ii) GABARAP partakes in GABAA receptor recycling by interacting with endosomal receptors, thus rerouting them toward the plasma membrane (Lüscher and Keller, 2001). (iii) GABARAP interacts with gephyrin and ankyrin 3, which facilitate postsynaptic receptor clusters and GABAA receptor stability (Nelson et al., 2020; Chen and Olsen, 2007). Moreover, lipidated GABARAP could potentially serve as an additional anchor for GABAA receptors via its PE-tail.
GABARAP-mediated protein trafficking extends beyond GABAA receptors; it is also involved in depositing angiotensin II receptor type 1 (AGTR1) at the plasma membrane (Cook et al., 2008). Mutagenesis analyses identified F309, L316, and L317 as critical residues, characterizing the motif as a canonical LIR (313FLQL316). This sequence, however, is not flanked by acidic but basic residues, defying the paradigm (Rogov et al., 2017). Notably, AGTR1 is highly expressed in dopaminergic neurons of the substantia nigra pars compacta that are highly susceptible to Parkinson’s disease (Kamath et al., 2022). Furthermore, GABARAP is responsible for the installation of transient receptor potential vanilloid 1 (TRPV1) at the plasma membrane (Laínez et al., 2010), which is important for microglial function in Alzheimer's disease (AD) (Lu et al., 2021). Here, the cytosolic 791FALV794 sequence might serve as a LIR. GABARAP also couples receptor homeostasis and autophagy by regulating epidermal growth factor receptor degradation via its LIR motif 1086FLPV1089 (Dobner et al., 2020; Üffing et al., 2024).
Dormant GABARAP localizes mainly to the Golgi, where it binds golgin subfamily A member 2 (GOLGA2) (Joachim et al., 2015), and to the ER, suggesting a role at the intersection of autophagy and receptor transport (Nakamura et al., 2008). Upon amino acid starvation, uncleaved GABARAP is displaced from GOLGA2 and rerouted from the Golgi to the ER via the centrosome (Joachim et al., 2015). How GABARAP is ultimately integrated into autophagy, and how these pathways are coordinated, remains unclear, though recent findings suggest a role for σ1R (Baeken et al., 2025).
Structure-function relation of σ1R
As a modulatory receptor pre-dominantly localized at the mitochondrial associated membrane (MAM) (Aishwarya et al., 2021), the σ1R is involved in multiple cell physiological processes such as calcium signaling, NMDA receptor activity, G-protein-mediated signaling pathways, and autophagy (Couly et al., 2023). It thus contributes to neuronal excitability, plasticity, proteostasis, and resilience. Accordingly, σ1R agonists have been proposed as therapeutic candidates for neurodegenerative diseases (Couly et al., 2024; Lachance et al., 2023). First identified over four decades ago as a unique opioid receptor class due to its atypical ligand binding properties (Martin et al., 1976; Tam, 1983), the σ1R represents an evolutionary orphan, sharing no close homology with any other human protein.
Phylogenetically, σ1R shares ∼69% structural similarity with yeast sterol isomerases that are involved in ergosterol biosynthesis (Hanner et al., 1996). It is highly conserved among vertebrates, with 80% sequence identity to zebrafish (Danio rerio) (Schmidt et al., 2016). X-ray crystallography of purified σ1R revealed a homotrimeric organization, where each protomer contains a cupin-like β-barrel ligand-binding domain (LBD) flanked by four α-helices and a deeply buried ligand-binding site (Schmidt et al., 2016). Membrane anchoring likely occurs via a single, poorly conserved N-terminal transmembrane helix linked to E102 of the LBD by V36 and F37. The protein’s topology remains debated - type II (C-terminus in ER-lumen) versus type III (C-terminus cytosolic) – likely due to artifacts from protein tagging (Aydar et al., 2002; Hayashi and Su, 2007). Most structural studies support a type II orientation, placing the C-terminal LBD in the ER-lumen, consistent with its interaction with the luminal chaperone heat shock 70 kDa protein 5 (HSPA5) (Hayashi and Su, 2007; Mavylutov et al., 2018; Sharma et al., 2021). However, this poses questions about the function of the N-terminal di-basic ER retention motif (1MQWAVGRR8) (Seth et al., 1998; Aishwarya et al., 2021). Notably, since most experiments to date have employed tagged proteins, the in situ topology of endogenous σ1R remains unresolved.
While no specific signal transduction pathway has been defined for σ1R, evidence suggests that agonists promote its monomeric, active form, facilitating interactions with client proteins, while antagonists stabilize an inactive oligomeric state. Agonist binding leads to dissociation from HSPA5, allowing σ1R to stabilize inositol trisphosphate receptor (IP3R) at the MAM, consequently enhance Ca2+ efflux, or modulate NMDA receptor activity (Hayashi and Su, 2007; Balasuriya et al., 2013; Choi et al., 2017). Prolonged activation further drives σ1R translocation to nuclear and plasma membranes, expanding its functional repertoire (Tsai et al., 2015).
Agonist treatment features a biphasic dose response, suggesting that high concentrations may suppress σ1R activity (Maurice, 2021), this response varies by cell and agonist type and dosage. Overall, σ1R activation promotes the unfolded protein response (Alam et al., 2017) and autophagy (Christ et al., 2019; Yang et al., 2019; Wang et al., 2021), reducing ER stress and supporting proteostasis. Several mutations in σ1R disrupt this function and are linked to motoneuron diseases; E102Q, for instance, is implicated in juvenile-onset ALS (Al-Saif et al., 2011; Vollrath et al., 2014; Dreser et al., 2017) and F83L in an ALS-like condition (Ma et al., 2020). The latter mutation resides in a sequence we have recently identified as a LIR motif, 81WVFV84 with specificity for GABARAP (Figure 1B) (Baeken et al., 2025). AlphaFold modeling supports experimental data by predicting π-stacking between F83 of σ1R and Y49 of GABARAP, potentially explaining binding specificity despite F83’s variable motif position (Figure 1B). Notably, the agonist-mediated induction of σ1R’s monomeric state enables this interaction; we did not observe any interaction in an oligomeric state. This specific interaction between σ1R and GABARAP may provide a causal link for the well-established effects of σ1R on autophagy (Christ et al., 2019; Yang et al., 2019; Wang et al., 2023).
σ1R as pharmaceutical target
The discovery that agonistic compounds such as alazocine [(+)-SKF-10047] and (+)-pentazocine (Martin et al., 1976; Su, 1982) selectively bind σ1R sparked its cellular and molecular characterization while highlighting it as a potent druggable target. It’s versatile roles as a regulator of Ca2+-homeostasis at the MAM, as a modulator of transmembrane receptors, and as an enhancer of the unfolded protein response and autophagy underline the potential neuroprotective properties of σ1R activation. Over time, extensive preclinical evidence has demonstrated that σ1R agonists confer neuroprotection across a broad spectrum of acute and chronic neurological disorders, including nerve injury, ALS, AD, Parkinson’s, and Huntington’s disease (Drewes et al., 2025).
Notably, neuroprotective σ1R agonists are structurally diverse. Classic benzomorphan-based ligands such as (+)-pentazocine (Li et al., 2021) and (+)-SKF-10047 (Phan et al., 2005), as well as chemically distinct molecules such as blarcamesine (Anavex 2-73), PRE-084 (Lahmy et al., 2013), and pridopidine (Ryskamp et al., 2019; Estévez-Silva et al., 2022), have all demonstrated beneficial behavioral and neuroprotective effects in various pathogenic mouse models. Several of these compounds have progressed to clinical trials. Pridopidine is currently in phase III for Huntington’s disease (NCT04556656) and phase II for ALS (NCT04615923); however, the HEALEY-ALS platform trial reported no significant clinical improvement (Shefner et al., 2025). In contrast, a phase IIB/III trial of blarcamesine in early AD has demonstrated significant improvements across multiple clinical outcomes (Macfarlane et al., 2025).
At the molecular level, σ1R agonists exhibit distinct modes of action, likely due to subtle differences at the agonist-σ1R interface, which may explain divergent cellular responses. Recent findings show that benzomorphan-based compounds – but not phencyclidine-derived compounds such as PRE-084 – induce monomerization of endogenous, untagged σ1R in vitro and in mouse tissues (Couly et al., 2024), at least under the experimental conditions and compound concentrations used. Moreover, the nature of the cellular stressor influences this response: oxidative stress (H2O2), but not ER stress (thapsigargin), promotes σ1R monomerization, even though both stressors activate the receptor (Couly et al., 2024). These results suggest that structurally diverse agonists and endogenous ligands regulate the switch between the “open-like” and closed σ1R conformation. In line, we recently observed that H2O2 and (+)-SKF-10047, but not PRE-084, promote σ1R binding to GABARAP (Baeken et al., 2025). An important consideration for the development of novel σ1R agonist is whether induction of the σ1R–GABARAP interaction is required for their neuroprotective effects, particularly in the context of autophagy modulation. For example, although PRE-084 does not induce σ1R–GABARAP interaction in HeLa cells (Baeken et al., 2025), it enhances autophagic flux in both HeLa cells and C. elegans, possibly via ULK1 modulation (Christ et al., 2019; Wang et al., 2021). These findings support the idea that σ1R agonists can elicit ligand-specific effects, including differential engagement of the σ1R–GABARAP axis.
σ1R in GABARAP logistics
A peculiarity concerning σ1R and GABARAP is their phylogenetic co-emergence in the Parahoxozoa, the earliest metazoan clade with neuronal circuits, suggesting a fundamental role for their interplay in neuronal homeostasis. Intriguingly, choline has been identified as an endogenous σ1R agonist (Brailoiu et al., 2019). In neurons, intracellular choline levels rise under oxidative or metabolic stress (e.g., phosphatidylcholine breakdown, hypoxia, or low acetyl-CoA), conditions that typically demand autophagy activation to degrade damaged components or replenish carbon sources. Thus, σ1R activation might redirect GABARAP toward autophagic pathways. It remains unclear whether σ1R activation sequesters Golgi-resident GABARAP or acts only on newly synthesized protein. The latter could explain σ1R’s neuroprotective effects, as it would preserve the Golgi’s GABARAP pool needed for plasma membrane homeostasis. Testing GABARAP–GOLGA2 interactions after σ1R activation may resolve this question.
Another group of molecules that modulate this system are steroid hormones: dehydroepiandrosterone (DHEA) acts as a σ1R agonist, while progesterone functions as an antagonist (Moriguchi et al., 2011; Johannessen et al., 2011). This aligns with σ1R’s evolutionary relationship with ergosterol-synthesizing sterol isomerases in yeast (Hanner et al., 1996). Notably, DHEA also serves as a negative allosteric regulator of GABAA receptors (Genud et al., 2009), suggesting that simultaneous σ1R activation and GABARAP rerouting could alter synaptic tone. Conversely, progesterone acts as a positive GABAA receptor modulator (Kapur and Joshi, 2021), effectively mirroring DHEA’s opposing effects on this dual regulatory axis.
Interestingly, crystal structures of Xenopus laevis σ1R in complex with DHEA sulfate (DHEAS) or progesterone demonstrate deeper penetration of the antagonist toward the C-terminal, membrane-associated helix (Fu et al., 2024) (Figure 1C). Here, progesterone probably interacts with Y203, while DHEAS engages W161. Former interaction might tighten the β-barrel, thereby blocking GABARAP interaction. Notably, DHEA levels rise during fasting (Grasfeder et al., 2009), suggesting σ1R activation in extrahepatic tissues could promote autophagy and thereby couple cellular energy responses to systemic metabolic status. In this light, σ1R may help precondition peripheral tissues for nutrient scarcity. Consequently, σ1R agonists might mimic aspects of the fasting response by shifting GABARAP from anabolic trafficking roles toward catabolic autophagy (Figures 2A,B). Mutational analysis of Y203 and W161 is needed to confirm their roles in agonist vs. antagonist binding. The relevance of this regulatory axis could further be tested in vivo by monitoring autophagy and σ1R monomerization under altered hormonal conditions.
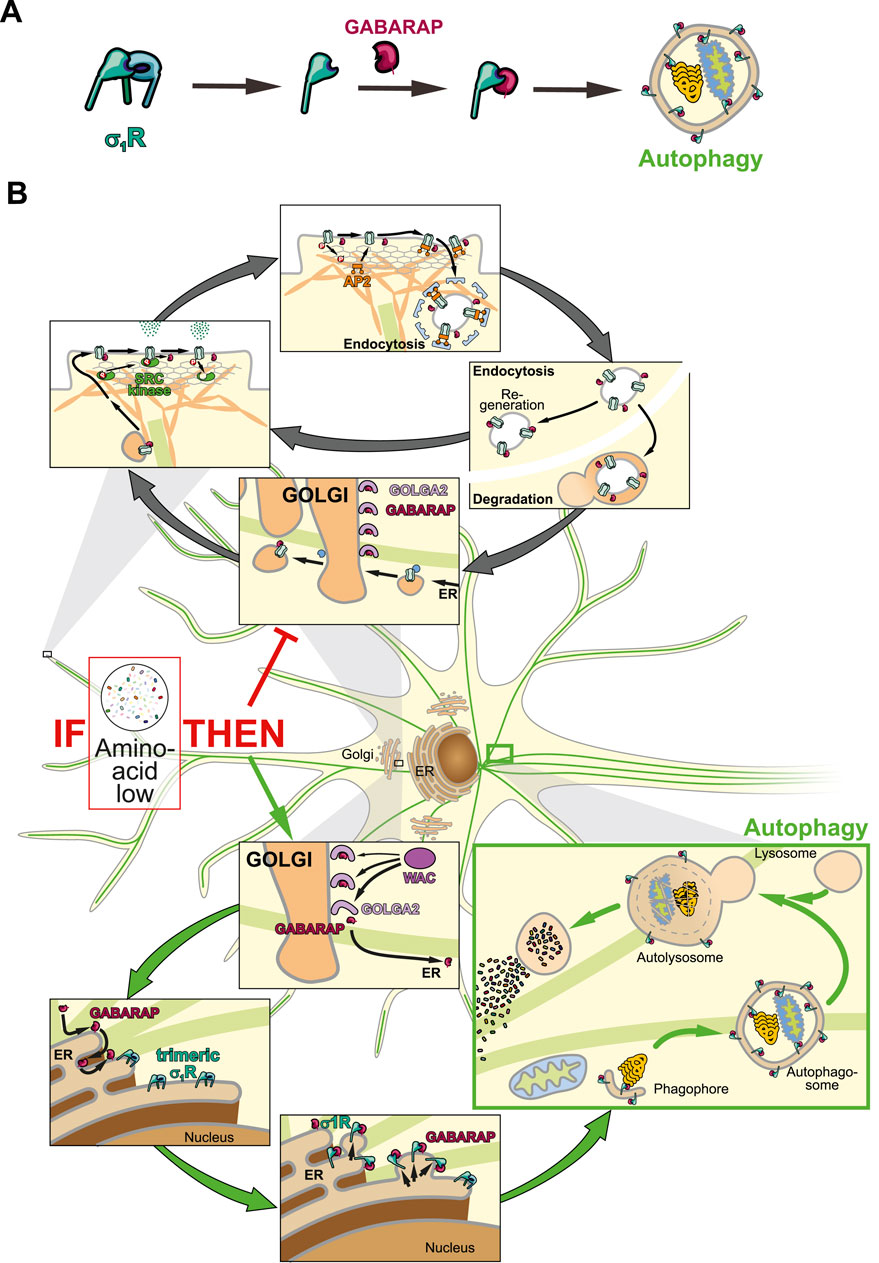
Figure 2. Schematic illustration of neuronal GABARAP logistics. (A) Step by step illustration of σ1R activation leading to autophagosome-associated GABARAP. (B) GABARAP is stored at the Golgi and either mediates receptor transport or integrates into autophagy via σ1R.
In cell culture, GABARAP is rerouted toward autophagy during starvation despite the absence of DHEAS, suggesting either σ1R bypass of or activation by alternative ligands. In neurons, Cytochrome P450 46A1 (CYP46A1), which is also upregulated during starvation (Shinohara et al., 2022), generates oxysterols potentially fulfilling this role. The biomolecular foundation model Boltz-2 predicts that 24(S)-hydroxycholesterol, a CYP46A1 product, binds human σ1R (Figure 1C), possibly explaining how CYP46A1 enhances autophagy (Nóbrega et al., 2019) and how H2O2 induces σ1R activity via sterol oxidation. Although further tests with oxysterol treatments and monitoring of σ1R and autophagy are required, current evidence highlights σ1R as a key regulator of GABARAP-mediated cell functions.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
MWB: Conceptualization, Visualization, Writing – original draft, Writing – review and editing. FB: Writing – original draft, Writing – review and editing. HK: Writing – original draft, Writing – review and editing. AMC: Writing – original draft, Writing – review and editing. CB: Funding acquisition, Writing – original draft, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the Hanna Bragard-Apfel Foundation, the Peter Beate Heller Foundation of the German Stifterverband, the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) - 530063157, and the COST Action SIGMA-1 EUROPE - CA23156.
Acknowledgments
The authors thank Michael Plenikowski for the graphical illustrations.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. The authors declare that AlphaFold3 (Abramson et al., 2024) and Boltz-2 (Passaro et al., 2025) were used to model molecular structures of proteins.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abramson, J., Adler, J., Dunger, J., Evans, R., Green, T., Pritzel, A., et al. (2024). Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 630 (8016), 493–500. doi:10.1038/s41586-024-07487-w
Aishwarya, R., Abdullah, C. S., Morshed, M., Remex, N. S., and Bhuiyan, M. S. (2021). Sigmar1's molecular, cellular, and biological functions in regulating cellular pathophysiology. Front. Physiol. 12, 705575. doi:10.3389/fphys.2021.705575
Al-Saif, A., Al-Mohanna, F., and Bohlega, S. (2011). A mutation in sigma-1 receptor causes juvenile amyotrophic lateral sclerosis. Ann. Neurol. 70 (6), 913–919. doi:10.1002/ana.22534
Alam, S., Abdullah, C. S., Aishwarya, R., Orr, A. W., Traylor, J., Miriyala, S., et al. (2017). Sigmar1 regulates endoplasmic reticulum stress-induced C/EBP-homologous protein expression in cardiomyocytes. Biosci. Rep. 37 (4), BSR20170898. doi:10.1042/BSR20170898
Albanesi, J., Wang, H., Sun, H. Q., Levine, B., and Yin, H. (2015). GABARAP-mediated targeting of PI4K2A/PI4KIIα to autophagosomes regulates PtdIns4P-dependent autophagosome-lysosome fusion. Autophagy 11 (11), 2127–2129. doi:10.1080/15548627.2015.1093718
Alemu, E. A., Lamark, T., Torgersen, K. M., Birgisdottir, A. B., Larsen, K. B., Jain, A., et al. (2012). ATG8 family proteins act as scaffolds for assembly of the ULK complex: sequence requirements for LC3-interacting region (LIR) motifs. J. Biol. Chem. 287 (47), 39275–39290. doi:10.1074/jbc.M112.378109
Aman, Y., Schmauck-Medina, T., Hansen, M., Morimoto, R. I., Simon, A. K., Bjedov, I., et al. (2021). Autophagy in healthy aging and disease. Nat. Aging. 1 (8), 634–650. doi:10.1038/s43587-021-00098-4
Ariosa, A. R., and Klionsky, D. J. (2016). Autophagy core machinery: overcoming spatial barriers in neurons. J. Mol. Med. (Berl.) 94 (11), 1217–1227. doi:10.1007/s00109-016-1461-9
Aydar, E., Palmer, C. P., Klyachko, V. A., and Jackson, M. B. (2002). The sigma receptor as a ligand-regulated auxiliary potassium channel subunit. Neuron 34 (3), 399–410. doi:10.1016/s0896-6273(02)00677-3
Baeken, M. W., Christ, M., Schmitt, D., Trein, W., Nagel, H., Clement, A. M., et al. (2025). Conserved LIR-specific interaction of Sigma-1 receptor and GABARAP. iScience 28, 113287. doi:10.1016/j.isci.2025.113287
Balasuriya, D., Stewart, A. P., and Edwardson, J. M. (2013). The σ-1 receptor interacts directly with GluN1 but not GluN2A in the GluN1/GluN2A NMDA receptor. J. Neurosci. 33 (46), 18219–18224. doi:10.1523/JNEUROSCI.3360-13.2013
Birgisdottir, Å. B., Lamark, T., and Johansen, T. (2013). The LIR motif - crucial for selective autophagy. J. Cell Sci. 126 (Pt 15), 3237–3247. doi:10.1242/jcs.126128
Brailoiu, E., Chakraborty, S., Brailoiu, G. C., Zhao, P., Barr, J. L., Ilies, M. A., et al. (2019). Choline is an intracellular messenger linking extracellular stimuli to IP3-Evoked Ca2+ signals through Sigma-1 receptors. Cell Rep. 26 (2), 330–337.e4. doi:10.1016/j.celrep.2018.12.051
Chen, Z. W., and Olsen, R. W. (2007). GABAA receptor associated proteins: a key factor regulating GABAA receptor function. J. Neurochem. 100 (2), 279–294. doi:10.1111/j.1471-4159.2006.04206.x
Chen, L., Wang, H., Vicini, S., and Olsen, R. W. (2000). The gamma-aminobutyric acid type A (GABAA) receptor-associated protein (GABARAP) promotes GABAA receptor clustering and modulates the channel kinetics. Proc. Natl. Acad. Sci. U. S. A. 97 (21), 11557–11562. doi:10.1073/pnas.190133497
Chen, Z. W., Chang, C. S., Leil, T. A., and Olsen, R. W. (2007). C-terminal modification is required for GABARAP-mediated GABA(A) receptor trafficking. J. Neurosci. 27 (25), 6655–6663. doi:10.1523/JNEUROSCI.0919-07.2007
Choi, S. R., Moon, J. Y., Roh, D. H., Yoon, S. Y., Kwon, S. G., Choi, H. S., et al. (2017). Spinal D-Serine increases PKC-dependent GluN1 phosphorylation contributing to the Sigma-1 receptor-induced development of mechanical allodynia in a mouse model of neuropathic pain. J. Pain 18 (4), 415–427. doi:10.1016/j.jpain.2016.12.002
Christ, M. G., Huesmann, H., Nagel, H., Kern, A., and Behl, C. (2019). Sigma-1 receptor activation induces autophagy and increases proteostasis capacity in vitro and in vivo. Cells 8 (3), 211. doi:10.3390/cells8030211
Christ, M. G., Clement, A. M., and Behl, C. (2020). The Sigma-1 receptor at the crossroad of proteostasis, neurodegeneration, and autophagy. Trends Neurosci. 43 (2), 79–81. doi:10.1016/j.tins.2019.12.002
Comenencia-Ortiz, E., Moss, S. J., and Davies, P. A. (2014). Phosphorylation of GABAA receptors influences receptor trafficking and neurosteroid actions. Psychopharmacol. (Berl.) 231 (17), 3453–3465. doi:10.1007/s00213-014-3617-z
Cook, J. L., Re, R. N., deHaro, D. L., Abadie, J. M., Peters, M., and Alam, J. (2008). The trafficking protein GABARAP binds to and enhances plasma membrane expression and function of the angiotensin II type 1 receptor. Circ. Res. 102 (12), 1539–1547. doi:10.1161/CIRCRESAHA.108.176594
Couly, S., Yasui, Y., and Su, T. P. (2023). SIGMAR1 confers innate resilience against neurodegeneration. Int. J. Mol. Sci. 24 (9), 7767. doi:10.3390/ijms24097767
Couly, S., Yasui, Y., Foncham, S., Grammatikakis, I., Lal, A., Shi, L., et al. (2024). Benzomorphan and non-benzomorphan agonists differentially alter sigma-1 receptor quaternary structure, as does types of cellular stress. Cell Mol. Life. Sci. 81 (1), 14. doi:10.1007/s00018-023-05023-z
Dobner, J., Simons, I. M., Rufinatscha, K., Hänsch, S., Schwarten, M., Weiergräber, O. H., et al. (2020). Deficiency of GABARAP but not its paralogs causes enhanced EGF-induced EGFR degradation. Cells 9 (5), 1296. doi:10.3390/cells9051296
Dreser, A., Vollrath, J. T., Sechi, A., Johann, S., Roos, A., Yamoah, A., et al. (2017). The ALS-linked E102Q mutation in sigma receptor-1 leads to ER stress-mediated defects in protein homeostasis and dysregulation of RNA-binding proteins. Cell Death Differ. 24 (10), 1655–1671. doi:10.1038/cdd.2017.88
Drewes, N., Fang, X., Gupta, N., and Nie, D. (2025). Pharmacological and pathological implications of Sigma-1 receptor in neurodegenerative diseases. Biomedicines 13 (6), 1409. doi:10.3390/biomedicines13061409
Estévez-Silva, H. M., Cuesto, G., Romero, N., Brito-Armas, J. M., Acevedo-Arozena, A., Acebes, Á., et al. (2022). Pridopidine promotes synaptogenesis and reduces spatial memory deficits in the alzheimer's disease APP/PS1 mouse model. Neurotherapeutics 19 (5), 1566–1587. doi:10.1007/s13311-022-01280-1
Fu, C., Xiao, Y., Zhou, X., and Sun, Z. (2024). Insight into binding of endogenous neurosteroid ligands to the sigma-1 receptor. Nat. Commun. 15 (1), 5619. doi:10.1038/s41467-024-49894-7
Genud, R., Merenlender, A., Gispan-Herman, I., Maayan, R., Weizman, A., and Yadid, G. (2009). DHEA lessens depressive-like behavior via GABA-ergic modulation of the mesolimbic system. Neuropsychopharmacology 34 (3), 577–584. doi:10.1038/npp.2008.46
Grasfeder, L. L., Gaillard, S., Hammes, S. R., Ilkayeva, O., Newgard, C. B., Hochberg, R. B., et al. (2009). Fasting-induced hepatic production of DHEA is regulated by PGC-1alpha, ERRalpha, and HNF4alpha. Mol. Endocrinol. 23 (8), 1171–1182. doi:10.1210/me.2009-0024
Hanner, M., Moebius, F. F., Flandorfer, A., Knaus, H. G., Striessnig, J., Kempner, E., et al. (1996). Purification, molecular cloning, and expression of the mammalian sigma1-binding site. Proc. Natl. Acad. Sci. U. S. A. 93 (15), 8072–8077. doi:10.1073/pnas.93.15.8072
Hayashi, T., and Su, T. P. (2007). Sigma-1 receptor chaperones at the ER-mitochondrion interface regulate Ca(2+) signaling and cell survival. Cell 131 (3), 596–610. doi:10.1016/j.cell.2007.08.036
He, C., and Klionsky, D. J. (2009). Regulation mechanisms and signaling pathways of autophagy. Annu. Rev. Genet. 43, 67–93. doi:10.1146/annurev-genet-102808-114910
Joachim, J., Jefferies, H. B., Razi, M., Frith, D., Snijders, A. P., Chakravarty, P., et al. (2015). Activation of ULK kinase and autophagy by GABARAP trafficking from the centrosome is regulated by WAC and GM130. Mol. Cell 60 (6), 899–913. doi:10.1016/j.molcel.2015.11.018
Johannessen, M., Fontanilla, D., Mavlyutov, T., Ruoho, A. E., and Jackson, M. B. (2011). Antagonist action of progesterone at σ-receptors in the modulation of voltage-gated sodium channels. Am. J. Physiol. Cell Physiol. 300 (2), C328–C337. doi:10.1152/ajpcell.00383.2010
Johansen, T., and Lamark, T. (2020). Selective autophagy: ATG8 family proteins, LIR motifs and cargo receptors. J. Mol. Biol. 432 (1), 80–103. doi:10.1016/j.jmb.2019.07.016
Jurd, R., Tretter, V., Walker, J., Brandon, N. J., and Moss, S. J. (2010). Fyn kinase contributes to tyrosine phosphorylation of the GABA(A) receptor gamma2 subunit. Mol. Cell Neurosci. 44 (2), 129–134. doi:10.1016/j.mcn.2010.03.002
Kamath, T., Abdulraouf, A., Burris, S. J., Langlieb, J., Gazestani, V., Nadaf, N. M., et al. (2022). Single-cell genomic profiling of human dopamine neurons identifies a population that selectively degenerates in parkinson's disease. Nat. Neurosci. 25 (5), 588–595. doi:10.1038/s41593-022-01061-1
Kapur, J., and Joshi, S. (2021). Progesterone modulates neuronal excitability bidirectionally. Neurosci. Lett. 744, 135619. doi:10.1016/j.neulet.2020.135619
Khaminets, A., Behl, C., and Dikic, I. (2016). Ubiquitin-dependent and independent signals in selective autophagy. Trends Cell Biol. 26 (1), 6–16. doi:10.1016/j.tcb.2015.08.010
Kittler, J. T., Chen, G., Kukhtina, V., Vahedi-Faridi, A., Gu, Z., Tretter, V., et al. (2008). Regulation of synaptic inhibition by phospho-dependent binding of the AP2 complex to a YECL motif in the GABAA receptor gamma2 subunit. Proc. Natl. Acad. Sci. U. S. A. 105 (9), 3616–3621. doi:10.1073/pnas.0707920105
Kneussel, M., Haverkamp, S., Fuhrmann, J. C., Wang, H., Wässle, H., Olsen, R. W., et al. (2000). The gamma-aminobutyric acid type A receptor (GABAAR)-associated protein GABARAP interacts with gephyrin but is not involved in receptor anchoring at the synapse. Proc. Natl. Acad. Sci. U. S. A. 97 (15), 8594–8599. doi:10.1073/pnas.97.15.8594
Lachance, V., Bélanger, S. M., Hay, C., Le Corvec, V., Banouvong, V., Lapalme, M., et al. (2023). Overview of sigma-1R subcellular specific biological functions and role in neuroprotection. Int. J. Mol. Sci. 24 (3), 1971. doi:10.3390/ijms24031971
Lahmy, V., Meunier, J., Malmström, S., Naert, G., Givalois, L., Kim, S. H., et al. (2013). Blockade of tau hyperphosphorylation and Aβ1-42 generation by the aminotetrahydrofuran derivative ANAVEX2-73, a mixed muscarinic and σ1 receptor agonist, in a nontransgenic mouse model of Alzheimer's disease. Neuropsychopharmacology 38 (9), 1706–1723. doi:10.1038/npp.2013.70
Laínez, S., Valente, P., Ontoria-Oviedo, I., Estévez-Herrera, J., Camprubí-Robles, M., Ferrer-Montiel, A., et al. (2010). GABAA receptor associated protein (GABARAP) modulates TRPV1 expression and channel function and desensitization. FASEB J. 24 (6), 1958–1970. doi:10.1096/fj.09-151472
Lee, Y. K., and Lee, J. A. (2016). Role of the mammalian ATG8/LC3 family in autophagy: differential and compensatory roles in the spatiotemporal regulation of autophagy. BMB Rep. 49 (8), 424–430. doi:10.5483/bmbrep.2016.49.8.081
Li, L., He, S., Liu, Y., Yorio, T., and Ellis, D. Z. (2021). Sigma-1R protects retinal ganglion cells in optic nerve crush model for glaucoma. Invest. Ophthalmol. Vis. Sci. 62 (10), 17. doi:10.1167/iovs.62.10.17
Lu, J., Zhou, W., Dou, F., Wang, C., and Yu, Z. (2021). TRPV1 sustains microglial metabolic reprogramming in Alzheimer's disease. EMBO Rep. 22 (6), e52013. doi:10.15252/embr.202052013
Lüscher, B., and Keller, C. A. (2001). Ubiquitination, proteasomes and GABA(A) receptors. Nat. Cell Biol. 3 (10), E232–E233. doi:10.1038/ncb1001-e232
Lystad, A. H., Ichimura, Y., Takagi, K., Yang, Y., Pankiv, S., Kanegae, Y., et al. (2014). Structural determinants in GABARAP required for the selective binding and recruitment of ALFY to LC3B-positive structures. EMBO Rep. 15 (5), 557–565. doi:10.1002/embr.201338003
Ma, M. T., Chen, D. H., Raskind, W. H., and Bird, T. D. (2020). Mutations in the SIGMAR1 gene cause a distal hereditary motor neuropathy phenotype mimicking ALS: report of two novel variants. Neuromuscul. Disord. 30 (7), 572–575. doi:10.1016/j.nmd.2020.05.005
Macfarlane, S., Grimmer, T., Teo, K., O'Brien, T. J., Woodward, M., Grunfeld, J., et al. (2025). Blarcamesine for the treatment of early Alzheimer's disease: results from the ANAVEX2-73-AD-004 phase IIB/III trial. J. Prev. Alzheimers Dis. 12 (1), 100016. doi:10.1016/j.tjpad.2024.100016
Martens, S., and Fracchiolla, D. (2020). Activation and targeting of ATG8 protein lipidation. Cell Discov. 6, 23. doi:10.1038/s41421-020-0155-1
Martin, W. R., Eades, C. G., Thompson, J. A., Huppler, R. E., and Gilbert, P. E. (1976). The effects of morphine- and nalorphine-like drugs in the nondependent and morphine-dependent chronic spinal dog. J. Pharmacol. Exp. Ther. 197 (3), 517–532. doi:10.1016/s0022-3565(25)30536-7
Maruyama, T., and Noda, N. N. (2017). Autophagy-regulating protease Atg4: structure, function, regulation and inhibition. J. Antibiot. (Tokyo) 71 (1), 72–78. doi:10.1038/ja.2017.104
Maurice, T. (2021). Bi-phasic dose response in the preclinical and clinical developments of sigma-1 receptor ligands for the treatment of neurodegenerative disorders. Expert Opin. Drug Discov. 16 (4), 373–389. doi:10.1080/17460441.2021.1838483
Mavylutov, T., Chen, X., Guo, L., and Yang, J. (2018). APEX2-tagging of sigma 1-receptor indicates subcellular protein topology with cytosolic N-terminus and ER luminal C-terminus. Protein Cell 9 (8), 733–737. doi:10.1007/s13238-017-0468-5
Mizushima, N., and Komatsu, M. (2011). Autophagy: renovation of cells and tissues. Cell 147 (4), 728–741. doi:10.1016/j.cell.2011.10.026
Moebius, F. F., Striessnig, J., and Glossmann, H. (1997). The mysteries of sigma receptors: new family members reveal a role in cholesterol synthesis. Trends Pharmacol. Sci. 18 (3), 67–70. doi:10.1016/s0165-6147(96)01037-1
Moriguchi, S., Yamamoto, Y., Ikuno, T., and Fukunaga, K. (2011). Sigma-1 receptor stimulation by dehydroepiandrosterone ameliorates cognitive impairment through activation of CaM kinase II, protein kinase C and extracellular signal-regulated kinase in olfactory bulbectomized mice. J. Neurochem. 117 (5), 879–891. doi:10.1111/j.1471-4159.2011.07256.x
Nakamura, T., Hayashi, T., Nasu-Nishimura, Y., Sakaue, F., Morishita, Y., Okabe, T., et al. (2008). PX-RICS mediates ER-to-Golgi transport of the N-cadherin/beta-catenin complex. Genes Dev. 22 (9), 1244–1256. doi:10.1101/gad.1632308
Nelson, A. D., Caballero-Florán, R. N., Rodríguez Díaz, J. C., Hull, J. M., Yuan, Y., Li, J., et al. (2020). Ankyrin-G regulates forebrain connectivity and network synchronization via interaction with GABARAP. Mol. Psychiatry 25 (11), 2800–2817. doi:10.1038/s41380-018-0308-x
Nguyen, T. N., Padman, B. S., Usher, J., Oorschot, V., Ramm, G., and Lazarou, M. (2016). Atg8 family LC3/GABARAP proteins are crucial for autophagosome-lysosome fusion but not autophagosome formation during PINK1/Parkin mitophagy and starvation. J. Cell Biol. 215 (6), 857–874. doi:10.1083/jcb.201607039
Nguyen, N., Olivas, T. J., Mires, A., Jin, J., Yu, S., Luan, L., et al. (2020). The insufficiency of ATG4A in macroautophagy. J. Biol. Chem. 295 (39), 13584–13600. doi:10.1074/jbc.RA120.013897
Nóbrega, C., Mendonça, L., Marcelo, A., Lamazière, A., Tomé, S., Despres, G., et al. (2019). Restoring brain cholesterol turnover improves autophagy and has therapeutic potential in mouse models of spinocerebellar ataxia. Acta neuropathol. 138 (5), 837–858. doi:10.1007/s00401-019-02019-7
Nymann-Andersen, J., Wang, H., Chen, L., Kittler, J. T., Moss, S. J., and Olsen, R. W. (2002). Subunit specificity and interaction domain between GABA(A) receptor-associated protein (GABARAP) and GABA(A) receptors. J. Neurochem. 80 (5), 815–823. doi:10.1046/j.0022-3042.2002.00762.x
Palmer, J. E., Wilson, N., Son, S. M., Obrocki, P., Wrobel, L., Rob, M., et al. (2025). Autophagy, aging, and age-related neurodegeneration. Neuron 113 (1), 29–48. doi:10.1016/j.neuron.2024.09.015
Pankiv, S., Clausen, T. H., Lamark, T., Brech, A., Bruun, J. A., Outzen, H., et al. (2007). p62/SQSTM1 binds directly to Atg8/LC3 to facilitate degradation of ubiquitinated protein aggregates by autophagy. J. Biol. Chem. 282 (33), 24131–24145. doi:10.1074/jbc.M702824200
Passaro, S., Corso, G., Wohlwend, J., Reveiz, M., Thaler, S., Somnath, V. R., et al. (2025). Boltz-2: towards accurate and efficient binding affinity prediction. BioRxiv 06.14, 2025.06.14.659707. doi:10.1101/2025.06.14.659707
Phan, V. L., Miyamoto, Y., Nabeshima, T., and Maurice, T. (2005). Age-related expression of sigma1 receptors and antidepressant efficacy of a selective agonist in the senescence-accelerated (SAM) mouse. J. Neurosci. Res. 79 (4), 561–572. doi:10.1002/jnr.20390
Rogov, V., Dötsch, V., Johansen, T., and Kirkin, V. (2014). Interactions between autophagy receptors and ubiquitin-like proteins form the molecular basis for selective autophagy. Mol. Cell 53 (2), 167–178. doi:10.1016/j.molcel.2013.12.014
Rogov, V. V., Stolz, A., Ravichandran, A. C., Rios-Szwed, D. O., Suzuki, H., Kniss, A., et al. (2017). Structural and functional analysis of the GABARAP interaction motif (GIM). EMBO Rep. 18 (8), 1382–1396. doi:10.15252/embr.201643587
Rogov, V. V., Nezis, I. P., Tsapras, P., Zhang, H., Dagdas, Y., Noda, N. N., et al. (2023). Atg8 family proteins, LIR/AIM motifs and other interaction modes. Autophagy Rep. 2 (1), 2188523. doi:10.1080/27694127.2023.2188523
Ryskamp, D., Wu, L., Wu, J., Kim, D., Rammes, G., Geva, M., et al. (2019). Pridopidine stabilizes mushroom spines in mouse models of alzheimer's disease by acting on the sigma-1 receptor. Neurobiol. Dis. 124, 489–504. doi:10.1016/j.nbd.2018.12.022
Schaaf, M. B., Keulers, T. G., Vooijs, M. A., and Rouschop, K. M. (2016). LC3/GABARAP family proteins: autophagy-(un)related functions. FASEB J. 30 (12), 3961–3978. doi:10.1096/fj.201600698R
Schmidt, H. R., Zheng, S., Gurpinar, E., Koehl, A., Manglik, A., and Kruse, A. C. (2016). Crystal structure of the human σ1 receptor. Nature 532 (7600), 527–530. doi:10.1038/nature17391
Seth, P., Fei, Y. J., Li, H. W., Huang, W., Leibach, F. H., and Ganapathy, V. (1998). Cloning and functional characterization of a sigma receptor from rat brain. J. Neurochem. 70 (3), 922–931. doi:10.1046/j.1471-4159.1998.70030922.x
Sharma, N., Patel, C., Shenkman, M., Kessel, A., Ben-Tal, N., and Lederkremer, G. Z. (2021). The Sigma-1 receptor is an ER-localized type II membrane protein. J. Biol. Chem. 297 (5), 101299. doi:10.1016/j.jbc.2021.101299
Shefner, J. M., Oskarsson, B., Macklin, E. A., Chibnik, L. B., Quintana, M., Saville, B. R., et al. (2025). Pridopidine in amyotrophic lateral sclerosis: the HEALEY ALS platform trial. JAMA 333 (13), 1128–1137. doi:10.1001/jama.2024.26429
Shinohara, Y., Ando, H., Maekawa, M., Arai, M., Horibata, Y., Satou, M., et al. (2022). Transcription of cytochrome P450 46A1 in NIH3T3 cells is negatively regulated by FBS. Biochimica biophysica acta. Mol. cell Biol. lipids 1867 (6), 159136. doi:10.1016/j.bbalip.2022.159136
Su, T. P. (1982). Evidence for sigma opioid receptor: binding of [3H]SKF-10047 to etorphine-inaccessible sites in guinea-pig brain. J. Pharmacol. Exp. Ther. 223 (2), 284–290. doi:10.1016/s0022-3565(25)33330-6
Sugawara, K., Suzuki, N. N., Fujioka, Y., Mizushima, N., Ohsumi, Y., and Inagaki, F. (2004). The crystal structure of microtubule-associated protein light chain 3, a mammalian homologue of Saccharomyces cerevisiae Atg8. Genes cells. 9 (7), 611–618. doi:10.1111/j.1356-9597.2004.00750.x
Tam, S. W. (1983). Naloxone-inaccessible sigma receptor in rat central nervous system. Proc. Natl. Acad. Sci. U. S. A. 80 (21), 6703–6707. doi:10.1073/pnas.80.21.6703
Tsai, S. Y., Chuang, J. Y., Tsai, M. S., Wang, X. F., Xi, Z. X., Hung, J. J., et al. (2015). Sigma-1 receptor mediates cocaine-induced transcriptional regulation by recruiting chromatin-remodeling factors at the nuclear envelope. Proc. Natl. Acad. Sci. U. S. A. 112 (47), E6562–E6570. doi:10.1073/pnas.1518894112
Üffing, A., Weiergräber, O. H., Schwarten, M., Hoffmann, S., and Willbold, D. (2024). GABARAP interacts with EGFR - supporting the unique role of this hAtg8 protein during receptor trafficking. FEBS Lett. 598 (21), 2656–2669. doi:10.1002/1873-3468.14997
Vollrath, J. T., Sechi, A., Dreser, A., Katona, I., Wiemuth, D., Vervoorts, J., et al. (2014). Loss of function of the ALS protein SigR1 leads to ER pathology associated with defective autophagy and lipid raft disturbances. Cell death. Dis. 5 (6), e1290. doi:10.1038/cddis.2014.243
Wang, H., Bedford, F. K., Brandon, N. J., Moss, S. J., and Olsen, R. W. (1999). GABA(A)-receptor-associated protein links GABA(A) receptors and the cytoskeleton. Nature 397 (6714), 69–72. doi:10.1038/16264
Wang, M., Wan, C., He, T., Han, C., Zhu, K., Waddington, J. L., et al. (2021). Sigma-1 receptor regulates mitophagy in dopaminergic neurons and contributes to dopaminergic protection. Neuropharmacology 196, 108360. doi:10.1016/j.neuropharm.2020.108360
Wang, S. M., Wu, H. E., Yasui, Y., Geva, M., Hayden, M., Maurice, T., et al. (2023). Nucleoporin POM121 signals TFEB-mediated autophagy via activation of SIGMAR1/sigma-1 receptor chaperone by pridopidine. Autophagy 19 (1), 126–151. doi:10.1080/15548627.2022.2063003
Yang, H., Shen, H., Li, J., and Guo, L. W. (2019). SIGMAR1/Sigma-1 receptor ablation impairs autophagosome clearance. Autophagy 15 (9), 1539–1557. doi:10.1080/15548627.2019.1586248
Keywords: sigma-1 receptor, GABARAP, autophagy, GABAa receptor, LIR
Citation: Baeken MW, Bekbulat F, Körschgen H, Clement AM and Behl C (2025) The sigma-1 receptor as a neurohomeostatic decision hub for GABARAP-mediated receptor trafficking and macroautophagy. Front. Mol. Biosci. 12:1673249. doi: 10.3389/fmolb.2025.1673249
Received: 25 July 2025; Accepted: 15 October 2025;
Published: 30 October 2025.
Edited by:
Maurizio Renna, University of Naples Federico II, ItalyReviewed by:
Zafiroula Georgoussi, National Centre of Scientific Research Demokritos, GreeceTao Wang, North Sichuan Medical College, China
Copyright © 2025 Baeken, Bekbulat, Körschgen, Clement and Behl. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marius Wilhelm Baeken, bWJhZWtlbkB1bmktbWFpbnouZGU=