 Lis Marques de Carvalho e Vieira1
Lis Marques de Carvalho e Vieira1 Victor Aguiar de Souza Penha2
Victor Aguiar de Souza Penha2 Kevin J. McGraw2*Amanda Vitória Dornelas da Silva1
Kevin J. McGraw2*Amanda Vitória Dornelas da Silva1 Erika Martins Braga1
Erika Martins Braga1- 1Departamento de Parasitologia, Universidade Federal de Minas Gerais, Belo Horizonte, Minas Gerais, Brazil
- 2Department of Integrative Biology, Michigan State University, East Lansing, MI, United States
Introduction: Pathogens and parasites play a crucial role in shaping ecological and evolutionary processes, influencing the behavior, physiology, and survival of their hosts across diverse ecosystems. Despite their taxonomic and functional diversity, non-passerine birds remain underrepresented in pathogen/parasite ecological research, providing an opportunity to explore how their unique life histories influence host-parasite dynamics. Investigating the susceptibility of non-passerines to infections, particularly in relation to physiological stress indicators such as heterophil-to-lymphocyte (H/L) ratios and microhematocrit levels, offers valuable insights into the complex interplay between health state, environmental conditions, and disease dynamics.
Methods: We evaluated the occurrence of haemosporidian parasite (avian malaria) infections (Plasmodium spp. and Haemoproteus spp.) in individuals from six non-passerine bird species – Red-legged Seriema (Cariama cristata), Crested Caracara (Caracara plancus), Yellow-headed Caracara (Daptrius chimachima), Dusky-legged Guan (Penelope obscura), Gray-necked Wood-Rail (Aramides cajaneus), and Tropical Screech Owl (Megascops choliba) – that were admitted to the Wildlife Screening Center in Belo Horizonte, Minas Gerais, Brazil. We assessed whether blood-parasite infection occurrence was associated with hematological parameters (e.g. microhematocrit, H/L ratio), major injuries, age, body condition, season, co-occurrence of Trichomonas spp. infections, and presence of ectoparasites.
Results: Of the 75 individuals analyzed, 37% were infected with haemosporidian parasites (Plasmodium spp. and Haemoproteus spp. combined). Age was a significant predictor of haemosporidian infection, with adults exhibiting higher overall haemosporidian parasite occurrence (both Plasmodium spp. and Haemoproteus spp. together), likely due to age-related chronic infection accumulation or higher mortality among infected juveniles. Also, individuals infected with Haemoproteus spp. only showed elevated H/L ratios, suggesting a physiological response to infection, and were more frequently infected during the rainy season, likely reflecting optimal vector conditions. No significant associations were observed between blood-parasite infection occurrence and other factors such as physical condition, major injuries, co-occurring Trichomonas spp., or the presence of ectoparasites.
Discussion: These findings highlight the importance of considering physiological, environmental, and life-history factors when investigating malaria infections in non-passerine birds. By advancing our understanding of host-parasite interactions in these underrepresented species, this study contributes valuable knowledge to inform conservation, rehabilitation, and wildlife-management strategies for these less-studied birds.
Introduction
Pathogens and parasites are fundamental drivers of host population dynamics, shaping behavior, physiology, and ecology across diverse ecosystems and host species (Woolhouse et al., 2002). Among these hosts, non-passerine birds stand out as a fascinating yet underexplored group in pathogen and parasite ecological research, with the potential to offer valuable insights into host-pathogen interactions across varied contexts (Santiago-Alarcon et al., 2014). Though comprising less than half of all bird species, non-passerine birds are generally large (with exceptions like hummingbirds) and include flightless runners (ratites) and swimmers (penguins), marshland waders (e.g. rails), and nocturnal hunters (owls), hence exposing them to a wide array of parasite and pathogen threats (Valkiunas, 2005; Gill, 2007).
Notably, non-passerines play a pivotal role in the dynamics of vector-borne diseases and the potential spillover of pathogens to other groups, including mammals (Valkiunas, 2005; Tsiodras et al., 2008). For example, when non-passerine species diversity is high, West Nile virus (WNV) transmission tends to decrease; this is because less competent hosts draw mosquitoes away from species that are better at amplifying the virus, which in turn reduces the risk of infection for other animals and humans (Ezenwa et al., 2006). Additionally, the foraging and nesting behaviors of non-passerines frequently bring them into contact with human-altered landscapes and livestock, thus increasing pathways for pathogen transmission. For example, waterfowl and shorebirds harbor avian influenza (Fenton et al., 2018) and coronaviruses, increasing the risk of spillover to farmed animals and humans (Rahman et al., 2021).
Among the many microbial threats to wild animals, haemosporidians represent a diverse group of vector-borne protozoan parasites that permit detailed examination of host susceptibilities, environmental selection pressures, and transmission dynamics. Haemosporidians are heteroxenous protozoa of broad ecological and veterinary importance, distributed worldwide except in Antarctica, and infecting various vertebrate hosts across different biomes (Valkiunas, 2005; Pacheco et al., 2011). In Brazil, they exhibit an average prevalence of ~28% in avian hosts (Ferreira-Junior et al., 2018; Oliveira et al., 2020; Fecchio et al., 2021c, Fecchio et al., 2021b, Fecchio et al., 2022; da Silva Rodrigues et al., 2021; de Angeli Dutra et al., 2023). These malaria parasites shape ecological dynamics by altering host physiology, behavior, and immune function, which in turn influence host interactions with vectors, conspecifics, and other species within their microenvironment and broader ecosystems (Carlson et al., 2015; Poulin, 2021). Although transmission dynamics have been well-studied in Passeriformes, research has primarily focused on their role in the spillover of virulent haemosporidian strains to naïve hosts via migration or introduction (Dinhopl et al., 2015; Niebuhr et al., 2016; Muriel et al., 2021), the crossing of wild-domestic boundaries (Ferreira-Junior et al., 2018), and interactions among unlikely host populations in rehabilitation centers and zoos (Sijbranda et al., 2017; Verwey et al., 2018; Ortiz-Catedral et al., 2019; Yoshimoto et al., 2021; González-Olvera et al., 2022). In contrast, much less is known about haemosporidian parasite transmission dynamics involving non-passerine birds. These interactions highlight the need to understand what makes non-passerine birds susceptible to disease, given their critical role in shaping disease dynamics and potential spillover to other animals. Given the diversity of non-passerines, it is likely that the risk and progression of haemosporidian infections in wild non-passerines are shaped by a multifaceted combination of biological, environmental, and anthropogenic factors.
Within avian communities, the impact of pathogens/parasites on hosts can be strongly influenced by age, with juveniles being particularly susceptible due to their relatively naïve immune systems or differences in behavior that may alter pathogen exposure (Wilson et al., 2002). For instance, low pathogenic influenza has been shown to cause greater severity in juvenile ostriches (Struthio camelus), primarily due to their limited capacity to immunologically cope with the virus (Capua et al., 2000). Similarly, juvenile Galapagos penguins (Spheniscus mendiculus) exhibit lower microbiome alpha diversity compared to adults, a difference likely driven by age-related behavioral patterns and potentially associated with increased susceptibility to pathogenic infections (Rohrer et al., 2023). Beyond age, an individual’s overall physical condition also plays an important role in infection risk. Birds in otherwise poor health may allocate fewer resources to immune defenses, increasing their susceptibility to infections and impairing their ability to manage pathogen burdens (da Silva Rodrigues et al., 2021). In American crows (Corvus brachyrhynchos), for example, poor body condition was related to a greater predisposition to several diseases (e.g., poxviral dermatitis, fungal pneumonia, enteritis) and consequent mortality (Townsend et al., 2010). Haemosporidian parasites have also been linked to decreased body condition in house martins (Delichon urbica; Marzal et al., 2013). However, it remains unclear whether individuals in poorer condition are more susceptible to infection, or if infection itself leads to declines in host condition.
Seasonal environmental fluctuations can further shape infection dynamics in wild birds, as changes in temperature, precipitation, and vector abundance can influence both host susceptibility and parasite transmission rates. Seasonality plays a crucial role in the transmission dynamics of avian malaria parasites, including Plasmodium, Haemoproteus, and Leucocytozoon (Valkiunas, 2005). In many regions, infection prevalence and parasite diversity fluctuate between the rainy and dry seasons, largely due to vector availability (Schultz et al., 2011; Hernández-Lara et al., 2017; Haghtalab et al., 2020; Fecchio et al., 2021a). The rainy season typically fosters higher transmission rates, as mosquito and biting midge populations—key vectors of Plasmodium and Haemoproteus, respectively—proliferate in response to increased humidity and water availability (Colombo and Joly, 2010; Tchoumbou et al., 2020). This seasonal peak in vector abundance often leads to a rise in infection prevalence and a higher likelihood of new infections, though the specific patterns depend on the haemosporidian parasite genus. For instance, Leucocytozoon spp. infections have been shown to increase with precipitation, likely due to the favorable conditions for black fly vectors, whereas Plasmodium spp. prevalence decreased overall in a global study, suggesting more complex seasonal drivers influencing its transmission in birds (Fecchio et al., 2021a).
Measurement of other physiological indicators, such as heterophil-to-lymphocyte ratios (H/L) and microhematocrit levels, can provide deeper insights into an individual’s stress, immune function, and overall health, all of which influence their infection susceptibilities and disease dynamics (Ricklefs and Sheldon, 2007; Satyaningtijas et al., 2020). Elevated H/L ratios are commonly associated with chronic stress and reduced immune function, making individuals more susceptible to malaria. For example, owls (from six different species) that were infected with Leucocytozoon spp. had higher H/L ratio compared to uninfected individuals, suggesting that individuals under higher stress are more susceptible to Leucocytozoon infection, or infection with Leucocytozoon depleted B-cells, rapidly decreasing lymphocyte levels compared to heterophils (Martín-Maldonado et al., 2023). Also, higher H/L ratios were associated with higher haemosporidian parasitemia in a diverse group of five Brazilian bird species (Rodrigues et al., 2021). In addition, low microhematocrit levels, indicative of anemia or poor oxygen transport capacity, further compromise a bird’s overall condition and ability to combat infections (Mitchell and Johns, 2008). Anemia is a clinical condition that has been consistently found to be associated with the acute phase of malaria infection (Kocan, 1968; Valkiunas, 2005).
Here we investigated several environmental, physiological, and life-history predictors of haemosporidian parasite infections in several species of non-passerine birds from Brazil. Non-passerine birds are less frequently studied in the context of avian malaria and other related haemosporidian parasites compared to passerines, but research indicates that they can serve as both competent and incidental hosts for Plasmodium spp. parasites, with varying susceptibility across taxa (Bell et al., 2020; Ilgūnas et al., 2022; Lotta-Arévalo et al., 2023). Some groups, such as pigeons and doves (Columbiformes), appear to be relatively resistant to infections, while others, like raptors and shorebirds, exhibit a broader range of susceptibilities (Santiago-Alarcon et al., 2014; Soares et al., 2016b; Capasso et al., 2023). Differences in host competence may be influenced by immune defenses, life-history traits, and habitat use, as many non-passerines occupy ecological niches that may limit exposure to mosquito vectors (e.g., marine environments, high-altitude zones, nocturnal habitats; Valkiunas, 2005; Santiago-Alarcon et al., 2014). Additionally, studies suggest that certain avian malaria lineages exhibit host specificity, with some Plasmodium spp. rarely detected in non-passerines (Lotta-Arévalo et al., 2023; Capasso et al., 2023). Despite these insights, significant knowledge gaps remain regarding the transmission dynamics, pathological effects, and potential evolutionary consequences of avian malaria in non-passerine hosts, highlighting the need for further research, especially in tropical and highly anthropogenically modified environments.
The objective of the present study was to evaluate the occurrence and severity of a common blood parasite – haemosporidians (Haemosporida, genera Plasmodium and Haemoproteus) – in six different non-passerine bird species from Brazil: red-legged seriema (Cariama cristata), crested caracara (Caracara plancus), yellow-headed caracara (Daptrius chimachima), dusky-legged guan (Penelope obscura), gray-necked wood-rail (Aramides cajaneus), and tropical screech owl (Megascops choliba). We studied individuals that were taken into the Wildlife Screening Center from the Brazilian Institute of Environment and Renewable Natural Resources (IBAMA) in Belo Horizonte, Minas Gerais, Brazil. We tested whether parasite occurrence and severity were related to hematological parameters (H/L ratio and microhematocrit levels), presence/absence of major injuries (burns, bone displacements, and fractures), age (juvenile vs. adults), body condition upon arrival at the rehabilitation center (poor vs. good), season (rainy vs. dry), as well as the occurrence of trichomoniasis (Order Trichomonadida, genus Trichomonas spp.), and ectoparasites (ticks and mites). We predicted that individuals with poor hematological parameters (high H/L ratio, low microhematocrit levels), major injuries, poor arrival condition, of juvenile age, sampled during the rainy season, and also infected with trichomoniasis and ectoparasites will have the highest haemosporidian parasite occurrences and severities. Overall, developing this deeper understanding of malaria parasites in these non-passerines can inform conservation efforts, improve rehabilitation practices, and enhance knowledge of host-parasite dynamics in diverse, tropical avian species.
Methods
Data collection
We sampled individuals received at the Wildlife Screening Center (Centro de Triagem de Animais Silvestres – CETAS-BH) from the Brazilian Institute of Environment and Renewable Natural Resources (IBAMA) in Belo Horizonte, Minas Gerais, Brazil from August 2023 - January 2025. We studied six species from five non-passerine orders: Cariamiformes (Cariama cristata), Falconiformes (Caracara plancus and Daptrius chimachima), Galliformes (Penelope obscura), Gruiformes (Aramides cajaneus), and Strigiformes (Megascops choliba). These birds arrived at the center after being either discovered with significant injuries (as above; Figure 1) or found in homes within urban or suburban environments. In the latter scenario, residents often reach out to the city’s environmental services, a program designed to rescue wildlife in residential areas. All individuals originated from Minas Gerais state, primarily from the Belo Horizonte metropolitan area and nearby cities. Upon arrival at the center, veterinarians conducted thorough evaluations of each individual, assessing body condition based on keel prominence (poor condition = pronounced keel; good condition = normal keel). Bird intake into the facility also included: (a) checking for visual signs of trichomoniasis, such as oral plaques, throat lesions, or difficulty swallowing (Dudek et al., 2018; Alrefaei et al., 2019), (b) inspecting all plumage and bare parts for the presence of ectoparasites; and the presence of any major injuries, including bone displacements, fractures, skin or eye wounds, leg injuries, and burns. Additionally, the veterinarians categorized the age of each bird (as either juvenile or adult, based on plumage characteristics and skull ossification; Von-Matter et al., 2010) and collected blood samples from the brachial vein. Unfortunately, sex of each bird was not always evident or determined, but this is unlikely to significantly impact our findings, as most of the species examined exhibit minimal or no sexual dimorphism. The samples were used to prepare blood smear slides and fill microhematocrit capillaries, and the remainder was stored in microtubes with 70% ethanol for further molecular analysis. The smears were then screened for haemosporidian parasites and H/L ratios (see more below).

Figure 1. Selected images of birds taken into the screening center, with examples of different injuries. (A) Crested caracara (Caracara plancus) with a major injury on the right wing (yellow arrow); and (B) tropical screech-owl (Megascops choliba) with a major injury on the left eye (green arrow).
Hematological parameters
Microhematocrit evaluation involved measuring the proportion of red blood cells (RBCs) in a blood sample, which is a useful indicator of an individual’s overall health and oxygen-carrying capacity (Clark et al., 2009). To perform this procedure, a small volume of blood was drawn into a capillary tube, which was then sealed at one end with clay. The tube was placed in a centrifuge and spun at 10000 G for 3 minutes, to separate the blood into its components. Following centrifugation, blood separates into distinct layers according to density: erythrocytes form a dense layer at the bottom, plasma forms the upper layer, and a thin intermediate layer appears between them, composed mainly of leukocytes and thrombocytes. The length of the RBC column was then measured using a microhematocrit card reader and was expressed as a percentage of the total blood column length (Mondal and Zubair, 2025), which can indicate conditions such as anemia, dehydration, or polycythemia. To analyze H/L ratios, a blood smear was prepared by spreading a drop of blood thinly across a microscope slide. The smear was then air-dried, fixed with 100% methanol for 3 minutes, and stained using a 10% Giemsa solution for 70 minutes (Valkiunas, 2005). The stained slides were examined under an Olympus CX31 light microscope at 1000× magnification with oil immersion to identify heterophils (characterized by their lobed nuclei and granular cytoplasm) and lymphocytes (smaller cells with a large, round nucleus and scant cytoplasm). 22,000 total cells were counted per slide (i.e. per animal) to calculate the relative proportions of each type. The H/L ratio was then determined by dividing the number of heterophils by the number of lymphocytes. An elevated H/L ratio may indicate chronic stress, infection, or inflammation, whereas a lower ratio can reflect baseline immune conditions (Satyaningtijas et al., 2020).
Microscopy and blood parasite identification and quantification
The stained slides were also used to identify parasite forms under an Olympus CX31 light microscope at 1000× magnification with oil immersion. Haemosporidian parasite identification (Plasmodium spp. and Haemoproteus spp.) followed established and well-described protocols (Valkiunas, 2005). Infection severity was then quantified by analyzing 200 microscopic fields with monolayered blood cells, counting parasitized cells among 22,000 total cells.
DNA extraction and cytochrome b gene amplification
To ensure the detection of Haemoproteus spp. and Plasmodium spp. infections in cases where parasitemia was low and difficult to identify through microscopy, we also performed PCR screening for haemosporidian parasites. For DNA extraction, a 20 μL volume of blood was placed in 1.5 mL microtubes and incubated at 60°C until the alcohol was completely evaporated. Then, 100 μL of 5% Chelex 100 (Sigma-Aldrich, Saint Louis, Missouri, USA) and 2.75 μL of proteinase K (20 mg/mL) (Phoneutria, Minas Gerais, Brazil) were added to the samples. The tubes were briefly vortexed and incubated at 60°C for 2 hours. After digestion, an enzymatic denaturation was performed at 95°C for 10 minutes. The tubes were then cooled to room temperature, vortexed, and centrifuged at 11,000 rpm for 10 minutes. The supernatant containing DNA was collected and quantified using a NanoDrop 2000 spectrophotometer (Thermo Scientific, USA) to ensure DNA concentrations were within the optimal range (40–80 ng/µL) (Pacheco et al., 2018) for further analysis and stored at –20°C. Positive control samples (Plasmodium falciparum) and negative control samples (MilliQ water) were also subjected to the extraction process. A 1 µL aliquot of the extracted DNA was subjected to a nested PCR protocol targeting a 478 bp region of the mitochondrial Cyt b gene of Plasmodium spp., and Haemoproteus spp. species (Hellgren et al., 2004), using the recently extracted positive and negative controls, as well as specific controls to ensure the reaction’s functionality (P. falciparum, which was previously amplified in other reactions, and MilliQ water). PCR products were visualized on 6% polyacrylamide gels stained with silver nitrate to confirm amplification. An individual was classified as infected if PCR and/or microscopy yielded a positive result.
Statistical analysis
Haemosporidian parasite occurrence
All analyses were performed with R software (R Core Team, 2024). We visually assessed the two numerical variables, microhematocrit levels and H/L ratios, for homoscedasticity using histograms. H/L ratios showed a right-skewed distribution, which we corrected using a square-root transformation. We scaled microhematocrit levels and H/L ratios to have a mean of zero and standard deviation of one using the scale function (R Core Team, 2024) to make all numeric variables on similar scales. To account for potential temporal variations in infection likelihood, we divided the dataset into dry (April to September) and wet (October to March) seasons.
We used three measurements of malaria occurrence in our analyses: presence/absence of (1) only Plasmodium spp., (2) only Haemoproteus spp., and (3) either genus (i.e. combined parasite occurrence). Thus, we constructed three separate generalized linear mixed model (GLMM) using the glmer function from the lme4 package (Bates et al., 2015) to examine predictors of these three parasite-occurrence variables. For the Plasmodium spp. and Haemoproteus spp. specific models, total sample size was slightly smaller (n = 71 in each, compared to n = 75 for the combined parasite occurrence model) because we could not obtain clear PCR sequencing results for some samples, only confirming a positive infection status without genus-level identification. Each model included the following predictors: microhematocrit level, H/L ratio, occurrence of major injuries, age class, arrival body condition, tick occurrence, trichomoniasis occurrence, and season (dry vs. wet). We also included species identity as a random factor to account for both phylogenetic relatedness and interspecific variation. We tested for the absence of multicollinearity among predictors using the variance inflation factor (VIF), calculated with the VIF function from the regclass package (Petrie, 2020). A threshold of two for was used to identify collinear predictors, which made us exclude tick occurrence from all models. To determine the importance of explanatory variables, we employed an information-theoretic approach (Burnham and Anderson, 2002). Using the dredge function from the MuMIn package (Barton, 2019), we generated all possible models with the selected explanatory variables. For cases where the best model had a weight of less than 0.8, we calculated model-averaged estimates using the model.avg function from the same package. If the best model had a weight greater than 0.8, we focused exclusively on analyzing that model. Finally, we selected the most important explanatory variables based on their estimate, conditional standard error, and 95% confidence interval. We evaluated model performance using root mean square error (RMSE), with lower RMSE values indicating better model fit. Therefore, models with RMSE values closer to zero were considered to provide a good fit to the data (Norberg et al., 2019; Tobler et al., 2019).
We also evaluated predictors of hemoparasite severity – defined as the proportion of infected versus uninfected red blood cells (combining Plasmodium spp. and Haemoproteus spp.) – in the 28 malaria-infected birds. Due to the left-skewed distribution of severity values, we log-transformed both malaria severity and H/L ratio for analysis. Following this, we employed a similar modeling approach to that used for the occurrence model, focusing on model selection and identifying statistically significant predictors. We did not identify any predictors with a high Variance Inflation Factor (VIF), so all predictors were retained in the model, including ectoparasite occurrence.
Results
Across all sampled species, the average proportion of individuals infected with malarial parasites was 37%, with Aramides cajaneus exhibiting the highest prevalence (71%). Mean malaria severity was 0.002 ± 0.002, with Caracara plancus showing the highest value (0.006) (Table 1; Figure 2). Trichomoniasis prevalence was generally low (9%), with Daptrius chimachima having the highest occurrence (40%). Additionally, ectoparasitic were found in only 7% of individuals, specifically in four Caracara plancus and one Cariama cristata. Injuries were observed in 37% of individuals, with Cariama cristata showing the highest proportion (61.5%). Lastly, mean microhematocrit and H/L ratio were 0.45 ± 0.10 and 4.0 ± 5.2, respectively, with Daptrius chimachima having the highest H/L ratio (8.4 ± 13.4) and Penelope obscura the highest microhematocrit (0.50 ± 0.07).
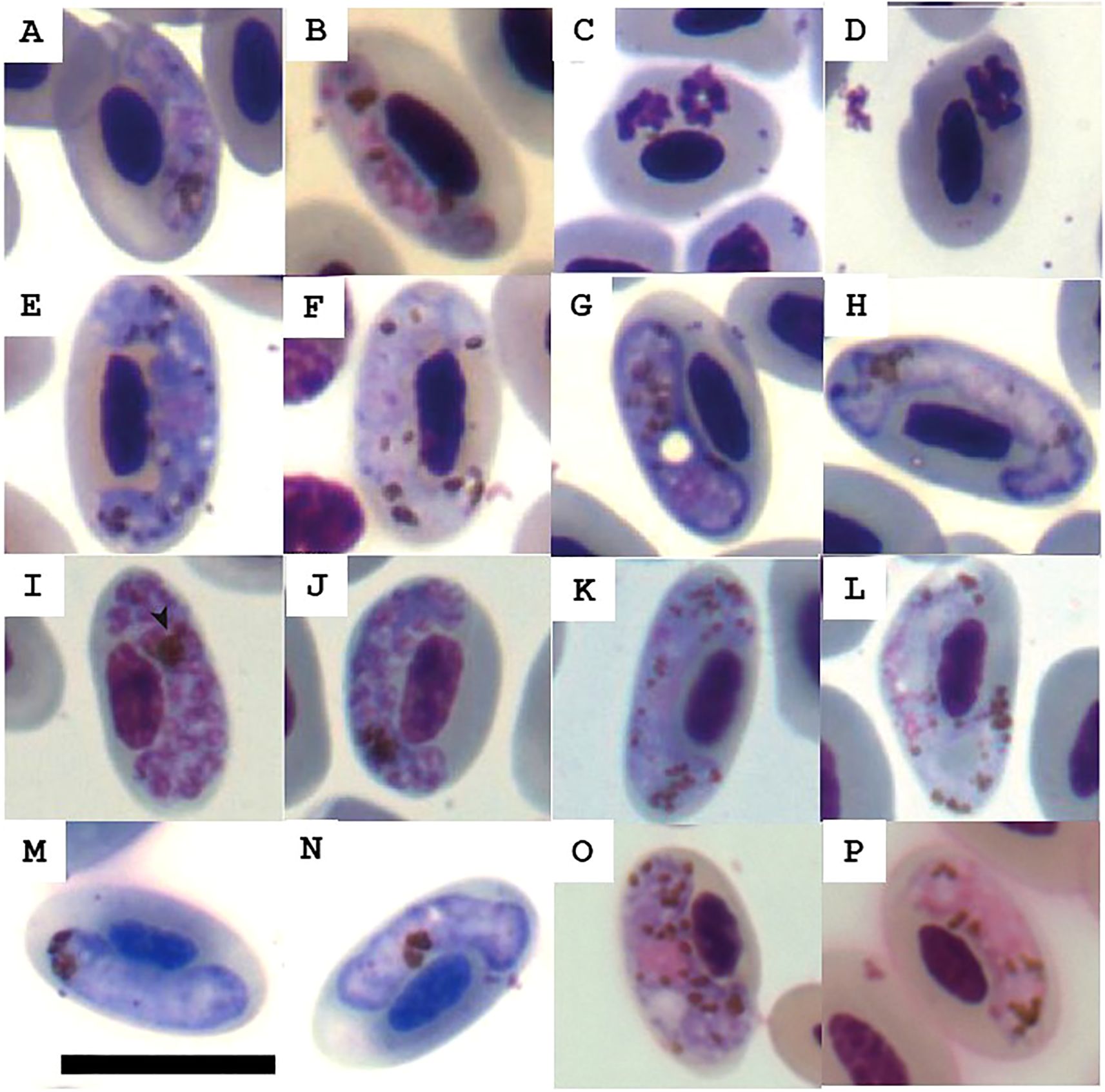
Figure 2. Haemosporidian morphotypes found in peripheral blood of avian hosts. (A-D) Plasmodium sp. from Aramides cajaneus; (E, F) Haemoproteus sp. from raptors (Caracara plancus and Daptrius chimachima); (G, H) Haemoproteus sp. from Penelope obscura; (I–L) Plasmodium sp. from Cariama cristata and D. chimachima; (M, N) Haemoproteus sp. from Megascops choliba; (O, P) Haemoproteus sp. from C. cristata. Scale bar = 10 mm.
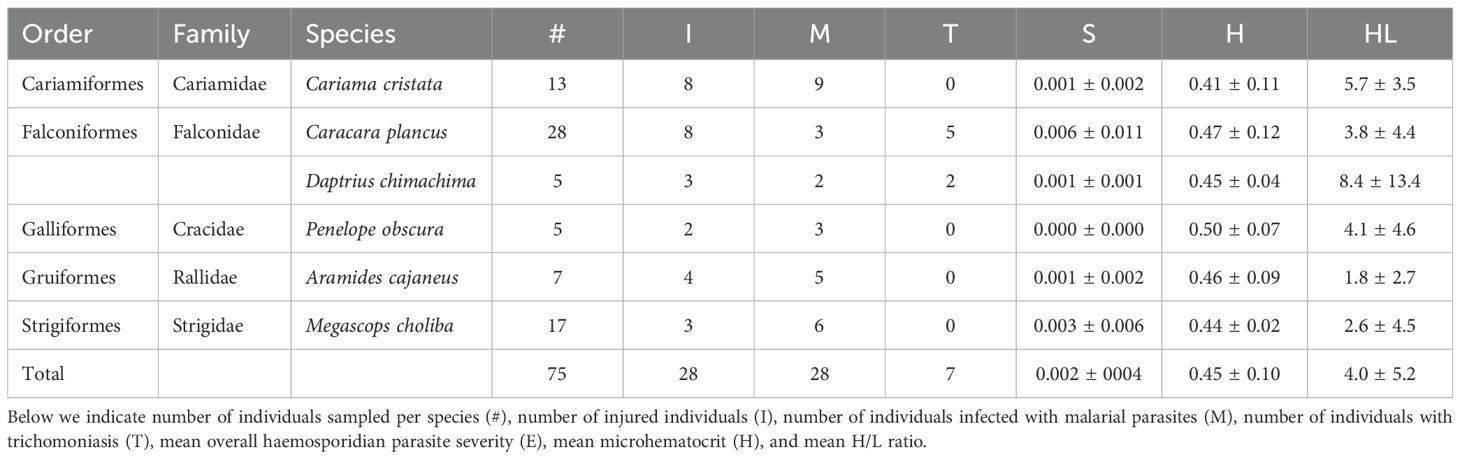
Table 1. Data summary for the study species, alongside their taxonomic classification.
The best model predicting overall haemosporidian parasite occurrence (Plasmodium spp. and Haemoproteus spp. combined; RMSE = 0.96) included only age (Supplementary Table 1), with juveniles being less likely than adults to be infected (Table 2; Figure 3). For Haemoproteus spp. (RMSE = 0.67), the best model included age, H/L ratio, and season, though only H/L ratio and season were significant predictors (Table 2). Individuals captured during the rainy season and those with higher H/L ratios were more likely to be infected with Haemoproteus spp. (Table 2; Figure 3). We did not identify any significant predictors for Plasmodium spp. occurrence (RMSE = 0.75) or for severity of malaria infection (RMSE = 1.65; Supplementary Table 1; Table 2).
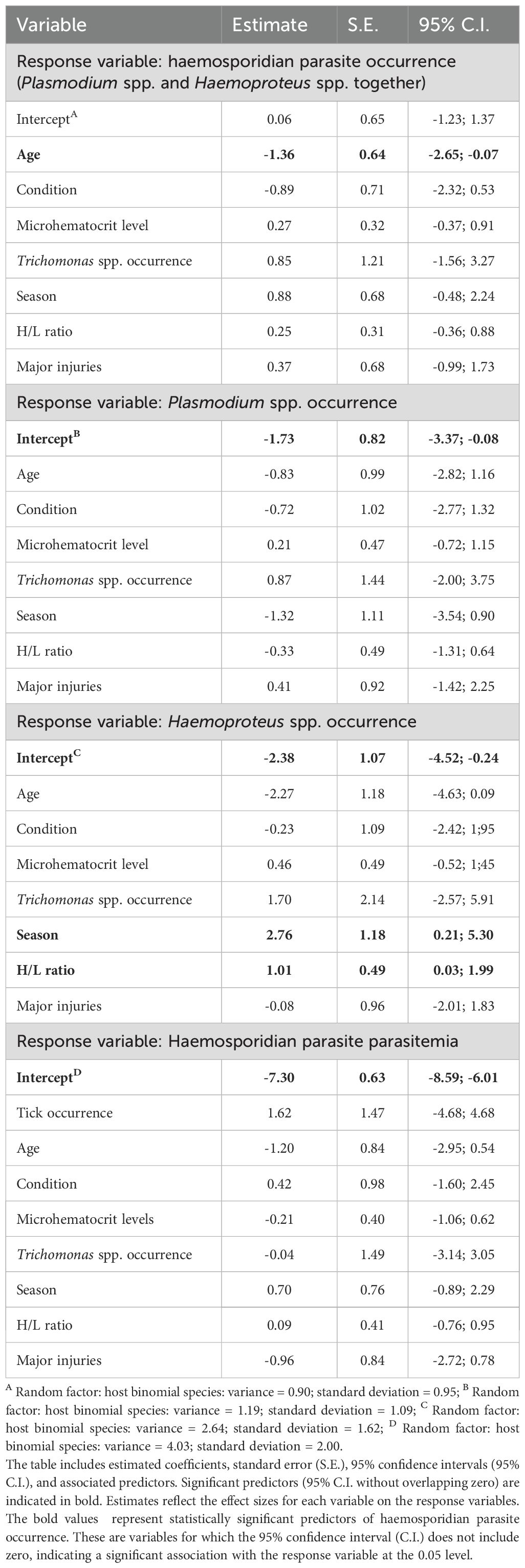
Table 2. Summary of the generalized linear mixed models (GLMM) results for the occurrence of haemosporidian parasites (Plasmodium spp. and Haemoproteus spp. occurrence together), Plasmodium spp., and Haemoproteus spp., as well as haemosporidian severity (Plasmodium spp. and Haemoproteus spp. combined) in non-passerine birds.
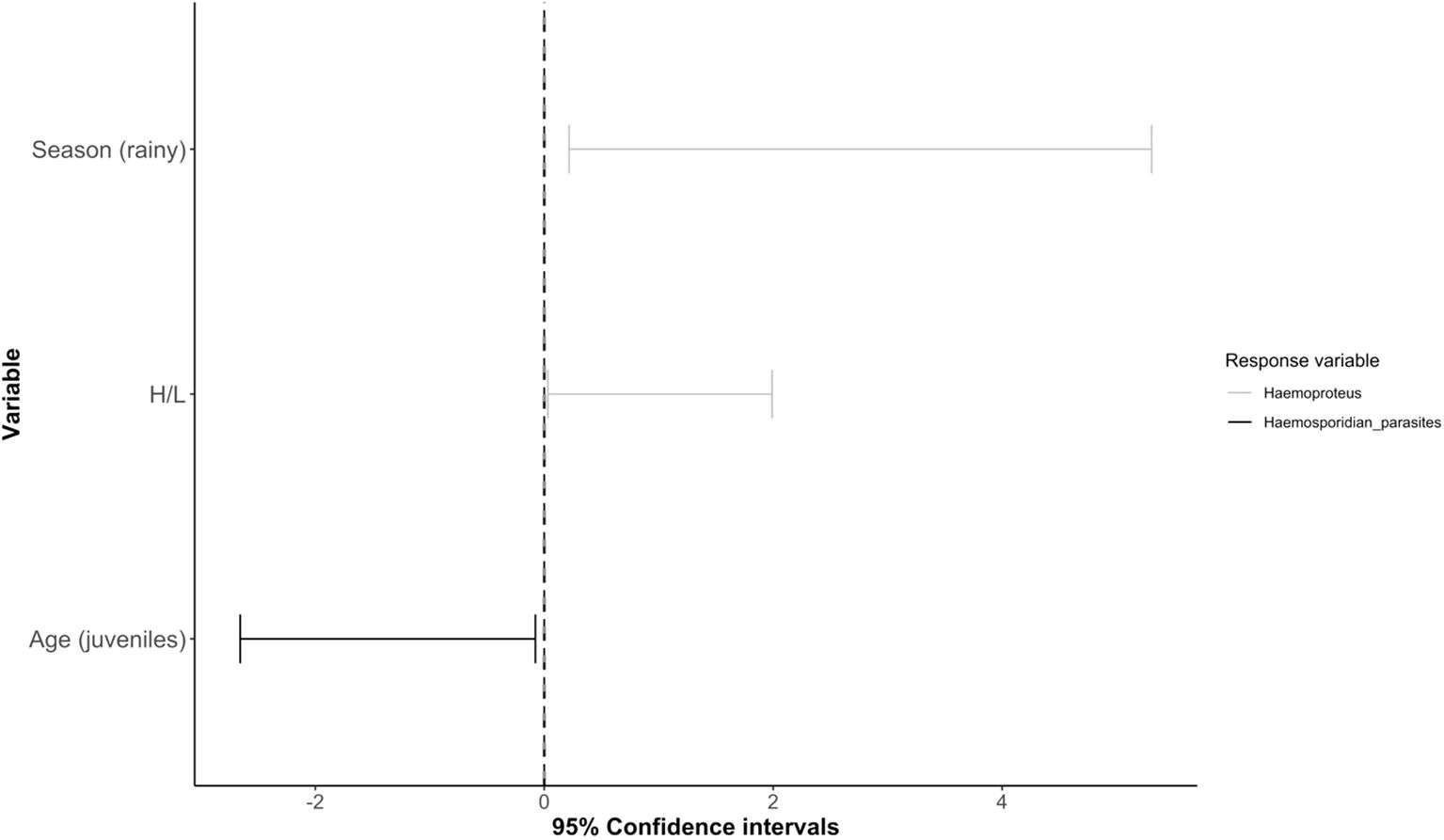
Figure 3. Forest plot showing the generalized linear mixed models (GLMM) 95% confidence intervals for models predicting the occurrence of haemosporidian parasites (Plasmodium spp. and Haemoproteus spp. together) and Haemoproteus spp. in non-passerine birds. Significant predictors are shown on the y-axis; the responses are represented by separate lines, with bars indicating confidence intervals (Haemoproteus spp. occurrence in gray and haemosporidian parasites in black). For complete list of predictors, please refer to Table 2.
To assess whether uneven species representation influenced our results, we re-ran all models using only the four most represented species (Caracara plancus, Megascops choliba, Cariama cristata, and Aramides cajanea), each with more than five individuals in the dataset. The results were highly consistent with those from the full dataset: in the general haemosporidian model, age remained marginally significant, and in the Haemoproteus-specific model, season was again marginally associated with infection risk. These findings suggest that the main patterns are not driven solely by uneven sampling across species. Additionally, we examined the distribution of individuals across species, age classes, and seasons (Supplementary Table S4). Notably, species with high infection prevalence—such as Aramides cajanea and Cariama cristata—were not disproportionately represented in the rainy season or in a particular age group. For example, Aramides cajanea was represented only by adults and almost evenly between dry (n = 4) and rainy (n = 3) seasons. These results indicate that the observed effects of age and season are not artifacts of biased sampling but instead reflect broader trends in our data.
Discussion
We found that the occurrence of haemosporidian parasite infections (Plasmodium spp. and Haemoproteus spp. combined) in non-passerine birds was influenced by the age of the avian hosts, with juvenile birds less likely to be infected than adults. We also observed that individuals infected specifically with Haemoproteus spp. had a higher H/L ratio and were more often brought into the facility during the rainy season. Taken together, these findings highlight the complex interplay between host age, physiological condition, and environmental factors in shaping the dynamics of malaria parasite infections, offering new insights into the eco-immunological and life-history drivers of hemoparasite infection in tropical non-passerine birds.
First, we found that juvenile birds exhibited a lower overall occurrence of avian malaria (Plasmodium spp. and Haemoproteus spp. combined) compared to adults, contrary to our initial prediction. We propose two non-mutually exclusive explanations for this pattern: (a) chronic infections may accumulate as birds age, which have been demonstrated in several bird species, such as in white-banded tanagers (Neothraupis fasciata; Fecchio et al., 2015), house martins (Marzal et al., 2016), dark-eyed juncos (Junco hyemalis; Slowinski et al., 2022), European bee-eaters (Merops apiaster; Emmenegger et al., 2020), and in a community-level avian study (Ellis et al., 2014), or (b) Infections may disproportionately impact the survival of younger birds (e.g. due to their developing immune systems; Ricklefs, 1992), leading to their reduced representation in the population. For instance, in yellowthroats (Geothlypis trichas), older males had a higher likelihood of surviving and managing chronic haemosporidian infections compared to their younger counterparts (Freeman-Gallant and Taff, 2017). Additionally, mortality linked to haemosporidian parasites has been documented in younger cranes (from 10 different species) at a zoo in China, further highlighting the vulnerability of younger individuals to these infections (Jia et al., 2018). These findings underscore the need for further research to disentangle the mechanisms driving age-related patterns of haemosporidian parasite occurrence in passerine and non-passerine bird populations.
Second, as we predicted, individuals infected with Haemoproteus spp. exhibited significantly higher H/L ratios compared to uninfected individuals. We hypothesize that Haemoproteus spp. infections may deplete B-cells, which are responsible for humoral immunity, leading to reduced lymphocyte levels in infected individuals (Delhaye et al., 2018). Similar patterns have been observed across various passerine and non-passerine species, including reed warblers (Acrocephalus scirpaceus) and sedge warblers (Acrocephalus schoenobaenus) infected with multiple blood parasites (Wojczulanis-Jakubas et al., 2012), various owl species (Martín-Maldonado et al., 2023), and several Neotropical birds such as ruddy ground-dove (Columbina talpacoti), great kiskadee (Pitangus sulphuratus), lesser elaenia (Elaenia chiriquensis), flavescent warblers (Myiothlypis flaveola), and brown-crested flycatchers (Myiarchus tyrannulus; da Silva Rodrigues et al., 2021), as well as house sparrows (Passer domesticus; Santiago-Alarcon et al., 2018) and house finches (Haemorhous mexicanus; Aguiar De Souza Penha et al., 2024). However, some studies have reported no such associations in red crossbills (Loxia curvirostra; Cornelius et al., 2014), American redstarts (Setophaga ruticilla), gray catbirds (Dumetella carolinensis), cedar waxwings (Bombycilla cedrorum), red-eyed vireos (Vireo olivaceus; Granthon and Williams, 2017), and rosy starlings (Pastor roseus; Dimitrov et al., 2019), indicating that this relationship can vary across species and contexts. In addition, also consistent with our predictions, we found that Haemoproteus spp. infections occurred more frequently in birds brought in during the rainy season. This result suggests that the higher prevalence of Haemoproteus spp. during the rainy season may overlap with optimal conditions for vector development and proliferation, given the more humid and wetter conditions (Meneghim et al., 2021). The tropical-bird literature offers mixed support for this pattern, with some studies reporting a higher malaria prevalence at the end of the dry season (Ferreira et al., 2017; Pulgarin et al., 2018; de Angeli Dutra et al., 2023), others finding no seasonal trends (Fecchio et al., 2015), and yet others reporting higher occurrence during the wet season (Schultz et al., 2011; Rivero de Aguilar et al., 2018). These findings illustrate the importance of considering environmental factors like climate and water availability when studying host-parasite interactions and emphasize the complexity of Haemoproteus spp. epidemiology.
We did not find any evidence that body condition, trichomoniasis, tick presence, or major injuries were statistically significant predictors of haemosporidian parasite occurrence, either overall (Plasmodium spp. and Haemoproteus spp. combined) or when analyzed separately. These results suggest that poor physical condition in affected individuals may be attributed to other causes, such as accidental collisions or encounters with predators (Stenkat et al., 2013), rather than parasitic infection. The lack of association with Trichomonas spp. or ectoparasites also underscores the variability in susceptibility to co-infections, which may be shaped by species-specific differences in ecology, physiology, and immune function. Associations in other bird species between haemosporidian parasites and pathogens such as Mycoplasma gallisepticum (Reinoso-Perez et al., 2020), Trypanosoma spp (Soares et al., 2016a), Isospora sp (Aguiar De Souza Penha et al., 2024), microfilariae (Clark et al., 2016), and Trichomonas gallinae (Thomas et al., 2022) demonstrate the complexity of host-pathogen interactions. This variability makes it challenging to establish consistent patterns of co-infection and emphasizes the need for targeted experimental studies to better understand the mechanisms driving these interactions and the broader ecological and evolutionary dynamics of co-infections across clades and geographic regions. It is also noteworthy that the lack of significant patterns in Trichomonas spp. and tick infections may be attributable to the low prevalence of these parasites in our sample (9% and 7%, respectively). Interestingly, microhematocrit levels were also not an important predictor of any of our haemosporidian parasite measurements (occurrence and severity), despite the fact that, based on other studies, we expected it would be reduced in infected individuals (Ellis et al., 2014; but see Martín-Maldonado et al., 2023).
Lastly, we found no significant predictors of hemoparasite severity in the 28 malaria-infected birds in this study. This contrasts with findings from studies of other non-passerine and/or tropical species, which have identified various factors influencing malaria severity, such as body condition, H/L ratio, total white-blood cell count, and reproductive effort (Knowles et al., 2009; da Silva Rodrigues et al., 2021). These results suggest that the drivers of hemoparasite severity may be context-dependent and demonstrate the need for further research with larger sample sizes and broader ecological contexts to better understand the patterns and determinants of avian malaria severity.
We must also address the fact that our study was conducted on a sample of rescued and rehabilitated birds, many of which were injured or otherwise compromised, and not of free-ranging, wholly uninjured individuals. As a result, our findings should be interpreted cautiously, as health/stress conditions related to injury or a home invasion could have influenced parasitological and physiological measures. Sampling free-ranging, uninjured birds in the wild would provide a valuable comparison to evaluate whether the patterns observed in these rescued individuals hold true in natural populations. Additionally, lack of data on the sex of the birds was a limitation, as we were unable to account for potential sex differences in infection or physiology; we recommend that future studies of this nature incorporate sex of the animals.
In summary, our study provides novel insight into the eco-immunological correlates of haemosporidian infections in non-passerine birds, a group that remains largely underrepresented in avian malaria research. Although the sample size was limited, our results consistently point to age and seasonality—particularly in Haemoproteus spp. infections—as key drivers of infection patterns, with additional support for the role of host stress physiology (via H/L ratio). These patterns were robust even when accounting for interspecific differences and restricting analyses to the most represented species. While not all variables yielded significant associations, our findings underscore the multifactorial nature of infection risk and highlight the importance of integrating life-history, physiological, and environmental factors to understand host–parasite dynamics in tropical bird assemblages. Further research with larger and free-ranging populations will be essential to refine these associations and explore their broader conservation and epidemiological implications.
Data availability statement
The original contributions presented in the study are publicly available. The data presented in the study are deposited in the GitHub repository, accessible at https://github.com/victoraspenha/FrontPara_Lis-et-al-2025-.
Ethics statement
The animal study was approved by Federal University Minas Gerais, Brazil. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
LM: Investigation, Writing – review & editing. VA: Writing – original draft, Writing – review & editing. KM: Writing – review & editing. AV: Writing – review & editing. EB: Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Acknowledgments
The authors express their gratitude to volunteers, interns, and environmental analysts at CETAS-BH, especially Nikolas Costa Pinto Felizardo, João Victor Martins Almeida, Sheila Cristina Ferreira da Silva, and Thiago Lima Stehling, for their assistance in capturing and collecting the animals used in this study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpara.2025.1568852/full#supplementary-material
References
Aguiar De Souza Penha V., Manica L. T., Barrand Z. A., Hepp C. M., and McGraw K. J. (2024). Correlates of co-infection with coccidiosis and avian malaria in house finches (Haemorhous mexicanus). J. Wildl. Dis. 61, 1–13. doi: 10.7589/JWD-D-23-00175
Alrefaei A. F., Low R., Hall N., Jardim R., Dávila A., Gerhold R., et al. (2019). Multilocus analysis resolves the european finch epidemic strain of trichomonas gallinae and suggests introgression from divergent trichomonads. Genome Biol. Evol. 11, 2391–2402. doi: 10.1093/gbe/evz164
Barton K. (2019). MuMIn: Multi-model inference (R package version 1.43.15) [Computer software]. (Vienna, Austria: R Foundation for Statistical Computing). Available online at: https://CRAN.R-project.org/package=MuMIn (Accessed January 18, 2025).
Bates D., Maechler M., Bolker B., and Walker S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw 67, 1–48. doi: 10.18637/jss.v067.i01
Bell J. A., González-Acuña D., and Tkach Vasyl V. (2020). Haemosporidian parasites of Chilean ducks: the importance of biogeography and nonpasserine hosts. J. Parasitol. 106, 211. doi: 10.1645/19-130
Burnham K. P. and Anderson D. R. (2002). Model selection and inference: A practical information-theoretic approach. 2nd edn (New York: Springer).
Capasso S., Schumm Y. R., Quillfeldt P., Bonsergent C., Malandrin L., Lorenti E., et al. (2023). Surveillance of avian malaria and related haemoparasites in common terns (Sterna hirundo) on the Atlantic coast of South America. Parasitology 150, 498–504. doi: 10.1017/S0031182023000185
Capua I., Mutinelli F., Bozza M. A., Terregino C., and Cattoli G. (2000). Highly pathogenic avian influenza (H7N1) in ostriches (Struthio camelus). Avian Pathol. 29, 643–646. doi: 10.1080/03079450020016913
Carlson J. S., Walther E., Trout Fryxell R., Staley S., Tell L. A., Sehgal R. N. M., et al. (2015). Identifying avian malaria vectors: Sampling methods influence outcomes. Parasit Vectors 8, 365. doi: 10.1186/s13071-015-0969-0
Clark P., Boardman W., and Raidal S. (2009). Atlas of clinical avian hematology (Hoboken, New Jersey: Wiley-Blackwell).
Clark N. J., Wells K., Dimitrov D., and Clegg S. M. (2016). Co-infections and environmental conditions drive the distributions of blood parasites in wild birds. J. Anim. Ecol. 85, 1461–1470. doi: 10.1111/1365-2656.12578
Colombo A. F. and Joly C. A. (2010). Brazilian Atlantic Forest lato sensu: the most ancient Brazilian forest, and a biodiversity hotspot, is highly threatened by climate change. Braz. J. Biol. 70, 697–708. doi: 10.1590/S1519-69842010000400002
Cornelius E. A., Davis A. K., and Altizer S. A. (2014). How important are hemoparasites to migratory songbirds? Evaluating physiological measures and infection status in three neotropical migrants during stopover. Physiol. Biochem. Zoology 87, 719–728. doi: 10.1086/677541
da Silva Rodrigues R., de Souza Penha V. A., Miwa R. Y., Branco J. O., and Junior O. M. (2021). Stress and body condition predict haemosporidian parasitaemia in birds from cerrado, southeastern Brazil. Ardea 109, 175–183. doi: 10.5253/arde.v109i3.a7
de Angeli Dutra D., Khan A. U., Ferreira F. C., Beirão M. V., Pichorim M., Moreira P. A., et al. (2023). Host phylogeny and seasonality shapes avian haemosporidian prevalence in a Brazilian biodiverse and dry forest: the Caatinga. Parasitology 150, 1277–1285. doi: 10.1017/S0031182023000549
Delhaye J., Jenkins T., Glaizot O., and Christe P. (2018). Avian malaria and bird humoral immune response. Malar J. 17, 1–7. doi: 10.1186/s12936-018-2219-3
Dimitrov D., Marinov M. P., Bobeva A., Ilieva M., Bedev K., Atanasov T., et al. (2019). Haemosporidian parasites and leukocyte profiles of pre-migratory rosy starlings (Pastor roseus) brought into captivity. Anim. Migration 6, 41–48. doi: 10.1515/ami-2019-0005
Dinhopl N., Nedorost N., Mostegl M. M., Weissenbacher-Lang C., and Weissenböck H. (2015). In situ hybridization and sequence analysis reveal an association of Plasmodium spp. with mortalities in wild passerine birds in Austria. Parasitol. Res. 114, 1455–1462. doi: 10.1007/s00436-015-4328-z
Dudek B. M., Kochert M. N., Barnes J. G., Bloom P. H., Papp J. M., Gerhold R. W., et al. (2018). Prevalence and risk factors of Trichomonas gallinae and trichomoniasis in golden eagle (Aquila chrysaetos) nestlings in western North America. J. Wildl Dis. 54, 755–764. doi: 10.7589/2017-11-271
Ellis V. A., Kunkel M. R., and Ricklefs R. E. (2014). The ecology of host immune responses to chronic avian haemosporidian infection. Oecologia 176, 729–737. doi: 10.1007/s00442-014-3048-x
Emmenegger T., Alves J. A., Rocha A. D., Costa J. S., Schmid R., Schulze M., et al. (2020). Population- and age-specific patterns of haemosporidian assemblages and infection levels in European bee-eaters (Merops apiaster). Int. J. Parasitol. 50, 1125–1131. doi: 10.1016/j.ijpara.2020.07.005
Ezenwa V. O., Godsey M. S., King R. J., and Guptill S. C. (2006). Avian diversity and West Nile virus: testing associations between biodiversity and infectious disease risk. Proc. R. Soc. B: Biol. Sci. 273, 109–117. doi: 10.1098/rspb.2005.3284
Fecchio A., Clark N. J., Bell J. A., Skeen H. R., Lutz H. L., De La Torre G. M., et al. (2021a). Global drivers of avian haemosporidian infections vary across zoogeographical regions. Global Ecol. Biogeography 30, 2393–2406. doi: 10.1111/geb.13390
Fecchio A., Dias R. I., Ferreira T. V., Reyes A. O., Dispoto J. H., Weckstein J. D., et al. (2022). Host foraging behavior and nest type influence prevalence of avian haemosporidian parasites in the Pantanal. Parasitol. Res. 121, 1407–1417. doi: 10.1007/s00436-022-07453-3
Fecchio A., Lima M. R., Bell J. A., Schunck F., Corrêa A. H., Beco R., et al. (2021b). Loss of forest cover and host functional diversity increases prevalence of avian malaria parasites in the Atlantic Forest. Int. J. Parasitol. 51, 719–728. doi: 10.1016/j.ijpara.2021.01.001
Fecchio A., Lima M. R., Silveira P., Ribas A. C. A., Caparroz R., and Marini M. Â. (2015). Age, but not sex and seasonality, influence Haemosporida prevalence in White-banded Tanagers (Neothraupis fasciata) from central Brazil. Can. J. Zool 93, 71–77. doi: 10.1139/cjz-2014-0119
Fecchio A., Ribeiro R. M., Ferreira F. C., de Angeli Dutra D., Tolesano-Pascoli G., Alquezar R. D., et al. (2021c). Higher infection probability of haemosporidian parasites in Blue-black Grassquits (Volatinia jacarina) inhabiting native vegetation across Brazil. Parasitol. Int. 80, 102204. doi: 10.1016/j.parint.2020.102204
Fenton H., McManamon R., and Howerth E. W. (2018). “Anseriformes, ciconiiformes, charadriiformes, and gruiformes,” in Pathology of wildlife and zoo animals (London, United Kingdom: Elsevier), 697–721. doi: 10.1016/B978-0-12-805306-5.00029-8
Ferreira F. C., Rodrigues R. A., Ellis V. A., Leite L. O., Borges M. A. Z., and Braga E. M. (2017). Habitat modification and seasonality influence avian haemosporidian parasite distributions in southeastern Brazil. PloS One 12, 1–18. doi: 10.1371/journal.pone.0178791
Ferreira-Junior F. C., de Angeli Dutra D., Silveira P., Pacheco R. C., Witter R., de Souza Ramos D. G., et al. (2018). A new pathogen spillover from domestic to wild animals: Plasmodium juxtanucleare infects free-living passerines in Brazil. Parasitology 145, 1949–1958. doi: 10.1017/S003118201800077X
Freeman-Gallant C. R. and Taff C. C. (2017). Age-specific patterns of infection with haemosporidians and trypanosomes in a warbler: implications for sexual selection. Oecologia 184, 813–823. doi: 10.1007/s00442-017-3919-z
González-Olvera M., Hernandez-Colina A., Himmel T., Eckley L., Lopez J., Chantrey J., et al. (2022). Molecular and epidemiological surveillance of Plasmodium spp. during a mortality event affecting Humboldt penguins (Spheniscus humboldti) at a zoo in the UK. Int. J. Parasitol. Parasites Wildl 19, 26–37. doi: 10.1016/j.ijppaw.2022.06.010
Granthon C. and Williams D. A. (2017). Avian malaria, body condition, and blood parameters in four species of songbirds. Wilson J. Ornithol 129, 492–508. doi: 10.1676/16-060.1
Haghtalab N., Moore N., Heerspink B. P., and Hyndman D. W. (2020). Evaluating spatial patterns in precipitation trends across the Amazon basin driven by land cover and global scale forcings. Theor. Appl. Climatol 140, 411–427. doi: 10.1007/s00704-019-03085-3
Hellgren O., Waldenström J., and Bensch S. (2004). a New Pcr Assay for Simultaneous Studies of Leucocytozoon, Plasmodium, and Haemoproteus From Avian Blood. J. Parasitol. 90, 797–802. doi: 10.1645/GE-184R1
Hernández-Lara C., González-García F., and Santiago-Alarcon D. (2017). Spatial and seasonal variation of avian malaria infections in five different land use types within a Neotropical montane forest matrix. Landsc Urban Plan 157, 151–160. doi: 10.1016/j.landurbplan.2016.05.025
Ilgūnas M., Himmel T., Harl J., Dagys M., Valkiūnas G., and Weissenböck H. (2022). Exo-erythrocytic development of avian haemosporidian parasites in european owls. Animals 12, 2212. doi: 10.3390/ani12172212
Jia T., Huang X., Valkiūnas G., Yang M., Zheng C., Pu T., et al. (2018). Malaria parasites and related haemosporidians cause mortality in cranes: a study on the parasites diversity, prevalence and distribution in Beijing Zoo. Malar J. 17, 234. doi: 10.1186/s12936-018-2385-3
Knowles S. C. L., Nakagawa S., and Sheldon B. C. (2009). Elevated reproductive effort increases blood parasitaemia and decreases immune function in birds: A meta-regression approach. Funct. Ecol. 23, 405–415. doi: 10.1111/j.1365-2435.2008.01507.x
Kocan R. M. (1968). Anemia and mechanism of erythrocyte destruction in ducks with acute leucocytozoon infections. J. Protozool 15, 455–462. doi: 10.1111/j.1550-7408.1968.tb02156.x
Lotta-Arévalo I. A., González A. D., Gamboa-Suárez B. A., Pacheco M. A., Escalante A. A., Moreno C., et al. (2023). Haemosporidians in non-passerine birds of Colombia: an overview of the last 20 years of research. Diversity (Basel) 15, 57. doi: 10.3390/d15010057
Martín-Maldonado B., Mencía-Gutiérrez A., Andreu-Vázquez C., Fernández R., Pastor-Tiburón N., Alvarado A., et al. (2023). A four-year survey of hemoparasites from nocturnal raptors (Strigiformes) confirms a relation between leucocytozoon and low hematocrit and body condition scores of parasitized birds. Vet. Sci. 10, 54. doi: 10.3390/vetsci10010054
Marzal A., Balbontín J., Reviriego M., García-Longoria L., Relinque C., Hermosell I. G., et al. (2016). A longitudinal study of age-related changes in Haemoproteus infection in a passerine bird. Oikos 125, 1092–1099. doi: 10.1111/oik.02778
Marzal A., Reviriego M., Hermosell I. G., Balbontín J., Bensch S., Relinque C., et al. (2013). Malaria infection and feather growth rate predict reproductive success in house martins. Oecologia 171, 853–861. doi: 10.1007/s00442-012-2444-3
Meneghim R. L. F. d. S., Madeira N. G., Ribolla P. E. M., Padovani C. R., and Schellini S. A. (2021). Flies as possible vectors of inflammatory trachoma transmission in a Brazilian municipality. Rev. Inst Med. Trop. Sao Paulo 63, e66. doi: 10.1590/s1678-9946202163066
Mitchell E. B. and Johns J. (2008). Avian hematology and related disorders. Veterinary Clinics North America: Exotic Anim. Pract. 11, 501–522. doi: 10.1016/j.cvex.2008.03.004
Mondal H. and Zubair M. (2025). “Hematocri,” in StatPearls (StatPearls Publishing, Treasure Island (FL). Available online at: https://www.ncbi.nlm.nih.gov/books/NBK542276/.
Muriel J., Marzal A., Magallanes S., García-Longoria L., Suarez-Rubio M., Bates P. J. J., et al. (2021). Prevalence and diversity of avian haemosporidians may vary with anthropogenic disturbance in tropical habitats in Myanmar. Diversity (Basel) 13, 111. doi: 10.3390/d13030111
Niebuhr C. N., Poulin R., and Tompkins D. M. (2016). Is avian malaria playing a role in native bird declines in New Zealand? Testing hypotheses along an elevational gradient. PloS One 11, e0165918. doi: 10.1371/journal.pone.0165918
Norberg A., Abrego N., Blanchet F. G., Adler F. R., Anderson B. J., Anttila J., et al. (2019). A comprehensive evaluation of predictive performance of 33 species distribution models at species and community levels. Ecol. Monogr. 89, e01370. doi: 10.1002/ecm.1370
Oliveira L., Dias R. J. P., Rossi M. F., D’Agosto M., and Santos H. A. (2020). Molecular diversity and coalescent species delimitation of avian haemosporidian parasites in an endemic bird species of South America. Parasitol. Res. 119, 4033–4047. doi: 10.1007/s00436-020-06908-9
Ortiz-Catedral L., Brunton D., Stidworthy M. F., Elsheikha H. M., Pennycott T., Schulze C., et al. (2019). Haemoproteus minutus is highly virulent for Australasian and South American parrots. Parasit Vectors 12, 40. doi: 10.1186/s13071-018-3255-0
Pacheco M. A., Cepeda A. S., Bernotienė R., Lotta I. A., Matta N. E, Valkiūnas G, et al. (2018). Primers targeting mitochondrial genes of avian haemosporidians: PCR detection and differential DNA amplification of parasites belonging to different genera. Int. J. Parasitol. 48 (8), 657–670. doi: 10.1016/j.ijpara.2018.02.003
Pacheco M. A., Escalante A. A., Garner M. M., Bradley G. A., and Aguilar R. F. (2011). Haemosporidian infection in captive masked bobwhite quail (Colinus virginianus ridgwayi), an endangered subspecies of the northern bobwhite quail. Vet. Parasitol. 182, 113–120. doi: 10.1016/j.vetpar.2011.06.006
Petrie A. (2020). “regclass: tools for an introductory class in regression and modeling,” in R package version 1.6. (Vienna, Austria: R Foundation for Statistical Computing) Available online at: https://cran.r-project.org/web/packages/regclass/. (Accessed January 18, 2025).
Poulin R. (2021). The rise of ecological parasitology: twelve landmark advances that changed its history. Int. J. Parasitol. 51, 1073–1084. doi: 10.1016/j.ijpara.2021.07.001
Pulgarín-R P. C., Gómez J. P., Robinson S., Ricklefs R. E., and Cadena C. D. (2018). Host species, and not environment, predicts variation in blood parasite prevalence, distribution, and diversity along a humidity gradient in northern South America. Ecol. Evol. 8, 3800–3814. doi: 10.1002/ece3.3785
Rahman M. M., Talukder A., Chowdhury M. M. H., Talukder R., and Akter R. (2021). Coronaviruses in wild birds – A potential and suitable vector for global distribution. Vet. Med. Sci. 7, 264–272. doi: 10.1002/vms3.360
R Core Team (2024). R: A language and environment for statistical computing [Computer software]. (Vienna, Austria: R Foundation for Statistical Computing). Available online at: https://www.R-project.org/. (Accessed January 18, 2025).
Reinoso-Pérez M. T., Dhondt K. V., Sydenstricker A. V., Heylen D., and Dhondt A. A. (2020). Complex interactions between bacteria and haemosporidia in coinfected hosts: An experiment. Ecol. Evol. 10, 5801–5814. doi: 10.1002/ece3.6318
Ricklefs R. E. (1992). Embryonic development period and the prevalence of avian blood parasites. Proc. Natl. Acad. Sci. U.S.A. 89, 4722–4725. doi: 10.1073/pnas.89.10.4722
Ricklefs R. E. and Sheldon K. S. (2007). Malaria prevalence and white-blood-cell response to infection in a tropical and in a temperate thrush. Auk 124, 1254–1266. doi: 10.1093/auk/124.4.1254
Rivero de Aguilar J., Castillo F., Moreno A., Peñafiel N., Browne L., Walter S. T., et al. (2018). Patterns of avian haemosporidian infections vary with time, but not habitat, in a fragmented Neotropical landscape. PloS One 13, e0206493. doi: 10.1371/journal.pone.0206493
Rodrigues R. A., Felix G. M. F., Pichorim M., Moreira P. A., and Braga E. M. (2021). Host migration and environmental temperature influence avian haemosporidians prevalence: A molecular survey in a Brazilian Atlantic rainforest. PeerJ 9, e11555. doi: 10.7717/peerj.11555
Rohrer S. D., Jiménez-Uzcátegui G., Parker P. G., and Chubiz L. M. (2023). Composition and function of the Galapagos penguin gut microbiome vary with age, location, and a putative bacterial pathogen. Sci. Rep. 13, 5358. doi: 10.1038/s41598-023-31826-y
Santiago-Alarcon D., Carbó-Ramírez P., Macgregor-Fors I., Chávez-Zichinelli C. A., and Yeh P. J. (2018). The prevalence of avian haemosporidian parasites in an invasive bird is lower in urban than in non-urban environments. Ibis 162, 201–214. doi: 10.1111/ibi.12699
Santiago-Alarcon D., Rodríguez-Ferraro A., Parker P. G., and Ricklefs R. E. (2014). Different meal, same flavor: cospeciation and host switching of haemosporidian parasites in some non-passerine birds. Parasit Vectors 7, 286. doi: 10.1186/1756-3305-7-286
Satyaningtijas A. S., Suprayogi A., Darusman H. S., Nurdin A., and Hanadhita D. (2020). Relative white blood cell counts, heterophil-to-lymphocyte ratio, and discovery of blood parasites in wild dugong (Dugong dugon) at Lingayan Island, Toli-toli, Indonesia. Vet. World 13, 1849–1853. doi: 10.14202/vetworld.2020.1849-1853
Schultz A., Underhill L. G., Earlé R., and Underhill G. (2011). Seasonality, distribution and taxonomic status of avian haemosporidian parasites within the Greater Cape Town area, South Africa. Ostrich 82, 141–153. doi: 10.2989/00306525.2011.603478
Sijbranda D., Hunter S., Howe L., Lenting B., Argilla L., and Gartrell B. (2017). Cases of mortality in little penguins (Eudyptula minor) in New Zealand associated with avian malaria. N Z Vet. J. 65, 332–337. doi: 10.1080/00480169.2017.1359124
Slowinski S. P., Geissler A. J., Gerlach N., Heidinger B. J., and Ketterson E. D. (2022). The probability of being infected with haemosporidian parasites increases with host age but is not affected by experimental testosterone elevation in a wild songbird. J. Avian Biol. 2022, e02819. doi: 10.1111/jav.02819
Soares L., Ellis V. A., and Ricklefs R. E. (2016a). Co-infections of haemosporidian and trypanosome parasites in a North American songbird. Parasitology 143, 1930–1938. doi: 10.1017/S0031182016001384
Soares L., Escudero G., Penha V. A. S., and Ricklefs R. E. (2016b). Low prevalence of haemosporidian parasites in shorebirds. Ardea 104, 129–141. doi: 10.5253/arde.v104i2.a8
Stenkat J., Krautwald-Junghanns M.-E., and Schmidt V. (2013). Causes of morbidity and mortality in free-living birds in an urban environment in Germany. Ecohealth 10, 352–365. doi: 10.1007/s10393-013-0868-9
Tchoumbou M. A., Mayi M. P. A., Malange E. N. F., Foncha F. D., Kowo C., Fru-cho J., et al. (2020). Effect of deforestation on prevalence of avian haemosporidian parasites and mosquito abundance in a tropical rainforest of Cameroon. Int. J. Parasitol. 50, 63–73. doi: 10.1016/j.ijpara.2019.10.006
Thomas R. C., Dunn J. C., Dawson D. A., Hipperson H., Horsburgh G. J., Morris A. J., et al. (2022). Assessing rates of parasite coinfection and spatiotemporal strain variation via metabarcoding: Insights for the conservation of European turtle doves Streptopelia turtur. Mol. Ecol. 31, 2730–2751. doi: 10.1111/mec.16421
Tobler M. W., Kéry M., Hui F. K. C., Guillera-Arroita G., Knaus P., Sattler T., et al. (2019). Joint species distribution models with species correlations and imperfect detection. Ecology 100, e02754. doi: 10.1002/ecy.2754
Townsend A. K., Clark A. B., McGowan K. J., Miller A. D., and Buckles E. L. (2010). Condition, innate immunity and disease mortality of inbred crows. Proc. R. Soc. B: Biol. Sci. 277, 2875–2883. doi: 10.1098/rspb.2010.0480
Tsiodras S., Kelesidis T., Kelesidis I., Bauchinger U., and Falagas M. E. (2008). Human infections associated with wild birds. J. Infection 56, 83–98. doi: 10.1016/j.jinf.2007.11.001
Verwey J., Peters A., Monks D., and Raidal S. (2018). Spillover of avian haemosporidian parasites (Haemosporidia: Plasmodium) and death of captive psittacine species. Aust. Vet. J. 96, 93–97. doi: 10.1111/avj.12671
Von-Matter S., Straube F. C., Accordi I. A., Piacentini V. Q., and Cândido-Jr J. F. (2010). Ornitologia e Conservação: Ciência aplicada, técnicas de pesquisa e levantamento (Rio de Janeiro: Technical Books Editora).
Wilson K., Bjørnstad O. N., Dobson A. P., Merler S., Poglayen G., Randolph S. E., et al. (2002). “Heterogeneities in macroparasite infections: patterns and processes,” in The ecology of wildlife diseases, eds. Hudson P. J., Rizzoli A., Grenfell B. T., Heesterbeek H., and Dobson A. P., (Oxford, United Kingdom: Oxford University Press Oxford), 6–44. doi: 10.1093/oso/9780198506201.003.0002
Wojczulanis-Jakubas K., Jakubas D., Czujkowska A., Kulaszewicz I., and Kruszewicz A. G. (2012). Blood parasite infestation and the leukocyte profiles in adult and immature reed warblers (Acrocephalus scirpaceus) and sedge warblers (Acrocephalus schoenobaenus) during autumn migration. Ann. Zool Fennici 49, 341–349. doi: 10.5735/086.049.0507
Woolhouse M. E. J., Webster J. P., Domingo E., Charlesworth B., and Levin B. R. (2002). Biological and biomedical implications of the co-evolution of pathogens and their hosts. Nat. Genet. 32, 569–577. doi: 10.1038/ng1202-569
Keywords: blood parasites, plasmodium, haemoproteus, heterophil-to-lymphocyte ratio (H/L), parasitemia, microhematocrit, malaria
Citation: Vieira LMdCe, Aguiar de Souza Penha V, McGraw KJ, da Silva AVD and Braga EM (2025) Physiological, environmental, and life-history drivers of haemosporidian infections in non-passerine birds from a rehabilitation center. Front. Parasitol. 4:1568852. doi: 10.3389/fpara.2025.1568852
Received: 11 February 2025; Accepted: 17 July 2025;
Published: 04 August 2025.
Edited by:
Samson Mukaratirwa, Ross University School of Veterinary Medicine, Saint Kitts and NevisReviewed by:
Santiago Merino, Spanish National Research Council (CSIC), SpainLuis Gerardo Herrera M., Universidad Nacional Autónoma de México, Mexico
Copyright © 2025 Vieira, Aguiar de Souza Penha, McGraw, da Silva and Braga. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kevin J. McGraw, bWNncmF3azVAbXN1LmVkdQ==