 Lorena Brito Pimentel Rodrigues dos Santos1*†
Lorena Brito Pimentel Rodrigues dos Santos1*† Naiane Oliveira-Santos1†
Naiane Oliveira-Santos1† Diogo Pereira Silva de Novais2†
Diogo Pereira Silva de Novais2† Valter Cruz-Magalhães3†
Valter Cruz-Magalhães3† Leandro Lopes Loguercio1*†
Leandro Lopes Loguercio1*†- 1Department of Biological Sciences, State University of Santa Cruz (UESC), Ilhéus, BA, Brazil
- 2Department of Education, Federal Institute of Education, Science and Technology of Bahia (IFBA), Porto Seguro, BA, Brazil
- 3Department of Phytophatology (DFP), Federal University of Lavras (UFLA), Lavras, MG, Brazil
Salinity, drought, and heavy metal contamination are abiotic stresses that pose significant challenges to agricultural productivity, compromising growth and development of plants. Strategies based on plants association with beneficial microorganisms, such as species of the Trichoderma genus, have been widely investigated due to their mitigating and protective potential against these adversities. This study aimed to organize and consolidate evidence on the contribution of Trichoderma spp. to plant resistance/tolerance against those three types of abiotic stresses. To achieve this, a series of meta-analysis were conducted on four groups of response variables in plants: growth and development, physiology/photosynthesis, enzymatic activities, and metabolites. Plants under abiotic stress conditions were compared with and without Trichoderma spp. inoculation. The presence of Trichoderma resulted in increased plant biomass, both in aerial and root parts, as well as in enhancement of photosynthesis-related parameters under all stress conditions. Specifically, positive effects were observed in chlorophyll fluorescence (ChlF), stomatal conductance (Gs), chlorophyll a (Chl a), and relative water content (RWC) under salinity stress; in Gs and RWC under drought; and in Chl a and Chl b under heavy metals exposure. Regarding antioxidant and enzymatic activities, we observed (i) increases in catalase (CAT) and total phenols, along with a reduction in malondialdehyde (MDA) and O2- under drought conditions; (ii) increases in ascorbate peroxidase (APx), superoxide dismutase (SOD) and total phenols, with a reduction in O2- and H2O2 under salinity stress; and (iii) increases in APx and SOD in the presence of heavy metals. Metabolite analysis revealed an increase in proline production under salinity stress and a reduction under drought. Taken together, the results highlight that associations with Trichoderma spp. tend to enhance plant adaptation to abiotic stresses by promoting plant growth and development, increasing chlorophyll production, controlling oxidative stress more efficiently by reducing the levels of reactive oxygen species (ROS), and contributing to osmotic balance under salinity conditions. Overall, this study reinforces the potential of Trichoderma spp. as a tool for managing abiotic stress in agriculture, in the context of environmental and climatic changes.
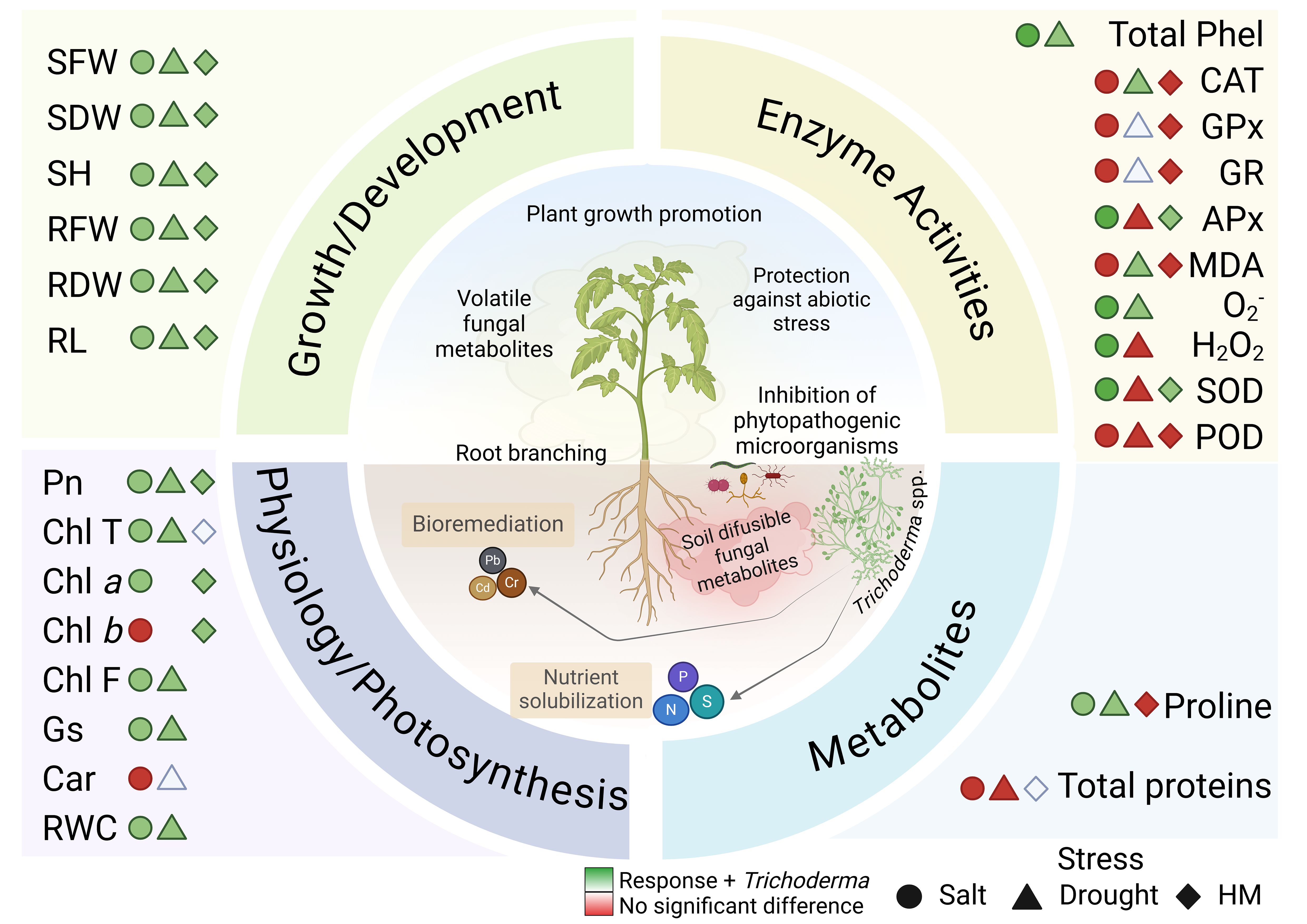
Graphical Abstract. Summary of beneficial Trichoderma-host interaction, based on the meta-analyses’ results. The symbols (circle, triangle, diamond) represent the types of stress studied (HM, heavy metals). The green color indicates positive responses from the interaction with Trichoderma spp., while the red color indicates that there was no significant difference between the control and treatments. Colorless symbols mean there have not been enough studies to measure the effect.
Introduction
Plants as we know nowadays have undergone coevolutionary processes with microorganisms, acquiring and preserving fundamental characteristics for their adaptation and establishment in the terrestrial environment (Rodriguez et al., 2019). This host-microbes interactions have occurred in a variety of forms throughout evolution, thus generating ‘units of cooperation’ deemed as holobionts (Catania et al., 2017); presumably, first eukaryotes have already maintained intimate relationships with prokaryotes and that, during the evolutionary process, their ‘immune systems’ became competent to distinguish beneficial from pathogenic microorganisms, giving them adaptive advantages (Rodriguez et al., 2019). With the advancement of ‘omics studies, it has been possible to understand ecological relationships of microorganisms, and mechanisms that regulate interactions between pathogenic, mutualistic and commensal microbes with plant hosts (Wiesmann et al., 2023). In this sense, regardless of the ecological relationship established, there are aspects of host defense, such as perception, signal transduction and activation of defensive responses, which involve primary and secondary metabolites, which operate in a similar way to what is observed in the innate immune system of animals (Haney et al., 2014; Mermigka et al., 2020) that help plants to distinguish between beneficial/neutral from harmful microbes.
Because of being sessile, plant species deal with adverse environmental changes (natural or anthropogenic) by specific means. Abiotic stresses in plants are caused by overlimit changes in physical or chemical factors that modulate ecosystems, such as radiation rates, water availability, composition of atmospheric gases, pH, sudden variations in temperature, composition of organic and mineral compounds, etc (Hasanuzzaman et al., 2020). These changes can affect the growth and development of plants, resulting in a decrease in quality and/or yield of crops, which poses a risk to global food security (Pereira, 2016; Raza et al., 2019). Beneficial host-associated microorganisms can help to mitigate these negative impacts of abiotic stresses through induced modifications in plants’ morphology, physiology, biochemistry, epigenetics, and overall gene expression patterns (Ogbe et al., 2020; González-Morales et al., 2021).
The Trichoderma genus includes several fungal species that interact with plants, helping them to cope with harmful effects of the environment. Studies have demonstrated that the presence of Trichoderma isolates improves the fitness of their hosts, induces tolerance to abiotic stresses, promotes plant growth and helps in the biocontrol of pests and diseases (dos Santos et al., 2022; Pacheco-Trejo et al., 2022). These phenotypes result from a diverse genetic arsenal of this fungal genus, capable of inducing genes related to plant defense, such as production of multiple secondary metabolites (Pacheco-Trejo et al., 2022), a variety of which with antifungal, antibiotic and hydrolytic activities (Durán and Marcato, 2011). Furthermore, many Trichoderma species are mycoparasites and/or compete for nutrients, thus directly or indirectly inhibiting growth of phytopathogens in the environment (Druzhinina et al., 2011; Mukherjee et al., 2022). These fungi are also known to induce systemic resistance in plants and promote tolerance to abiotic stresses such as drought, salinity and pollution by toxic metals (Zaidi et al., 2014; Hidangmayum and Dwivedi, 2018), thus help increasing root growth, nutrient absorption and protection against oxidative stress. Finally, Trichoderma spp. are capable of making various nutrients available in the soil; due to being saprophytes, these fungi produce a variety of lytic enzymes that break down insoluble nutrients, making them accessible to plant roots (Mehetre and Mukherjee, 2015; Vinale and Sivasithamparam, 2020).
In a previous study aiming to evaluate several mechanisms involved in different plant-Trichoderma interactions (dos Santos et al., 2022), it was observed that, depending on the plant species, the type of abiotic stress, and the Trichoderma isolate, this fungal genus can employ multiple and complex biological processes simultaneously to aid in plant survival. For a better understanding of the morphological, physiological and biochemical mechanisms of this stress-driven interaction, it is interesting to systematically examine such primary studies to determine the relative importance of each of those mechanisms. The magnitude of Trichoderma spp. effects on abiotic stress tolerance in plants varies between studies, as it depends on factors such as stress level, host types, strain and inoculum concentration of Trichoderma, environmental conditions and their complex interactions (dos Santos et al., 2022). Understanding the contribution of these factors to the plant-Trichoderma interaction is crucial to elucidate the mechanisms of stress tolerance mediated by this fungal genus. To identify trends and patterns of influence of Trichoderma spp. upon plants subjected to certain abiotic stresses, when compared to control conditions (stress in absence of the fungal treatment), it is essential to integrate the results of multiple studies, enabling the identification of underlying factors involved in these interactions (Gurevitch et al, 2018). With this objective, we conducted a meta-analysis to gauge the magnitude of the Trichoderma-driven effects in abiotic stress relief in plants, focusing on salt excess (high salinity), water deficit (drought) and high levels of heavy metals stresses. Our main question was what is the empirical primary evidence in the literature showing that plant species acquire resistance to abiotic stresses in the presence of Trichoderma species/isolates?
Methods
A local database of articles (Supplementary Table S1) was established from a previous systematic review (dos Santos et al., 2022) to verify empirical evidence in the peer-reviewed literature on the ability of Trichoderma species to confer improved resistance/tolerance to abiotic stresses in plants. The publications were extracted from the PubMed, Scopus, Web of Science databases and from Google Scholar through the Publish or Perish software (Harzing, 2007); the keywords “Trichoderma” and “abiotic stress” were used.
The exclusion criteria to select the articles of this study presented one or more of the following characteristics: (i) review articles, simple or expanded abstracts, book chapters, dissertations and theses; (ii) studies that included only in vitro assays, without a plant species; (iii) articles that did not report variation parameters (se, sd, CV, etc.) or number of replications; (iv) articles on biocontrol or biotic stress factors (e.g., phytopathogens); (v) studies with absence of control groups and/or inoculated Trichoderma species grown under stress conditions; (vi) articles not aligned with the study objective and/or with flaws or inconsistencies between methodology and results/conclusions. For the selected articles, the Trichoderma species, the plant host and the observations reported were assessed and summarized (Supplementary Table S2). Based on this assessment, the effect measures were grouped into three categories, based on stress type, i.e., saline, water deficit and heavy metal stresses; a summary of these and other related scientometric information, including articles retrieved and selected, number and characteristics of experiments addressed, effect measures and entries for the meta-analyses are presented in Table 1, Supplementary Table S3.
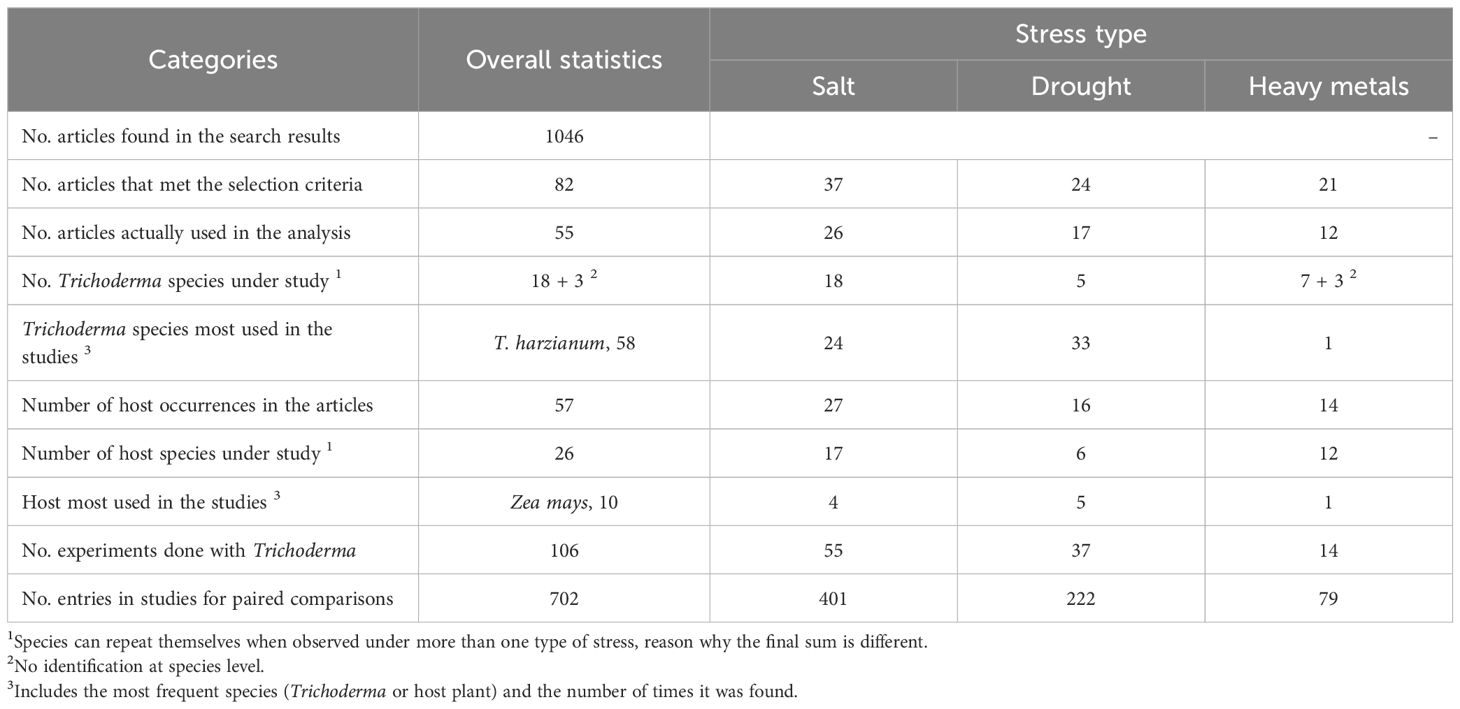
Table 1. General scientometric data found on the effects of the Trichoderma on plant fitness under conditions of saline, drought and heavy metal stresses.
The data for each effect measure was obtained, including the mean values, variation measures (‘sd’, ‘95%CI’, ‘se’ etc., which were all converted to ‘sd’ for the meta-analyses) and the number of replicates for plants under abiotic stress conditions without and with Trichoderma sp. isolates (control and treatment, respectively). These data were extracted directly from the tables, text, or graphs, using the GetData Graph Digitizer software (v. 2.26; http://getdata-graph-digitizer.com). All the collected values were associated with the categorical information in the database (Supplementary Table S3). To better assess the effect measures found, the data were categorized into four major groups of variables: (i) biomass (fresh and dry weight of shoot/roots, and length of shoot and root); (ii) physiology and photosynthesis [chlorophyll fluorescence, transpiration, net photosynthesis, stomatal conductance, relative water content, photosynthetic pigments (chlorophyll a, chlorophyll b, total chlorophyll, and total carotenoid)]; (iii) enzyme activities [superoxide dismutase (SOD), catalase (CAT), peroxidase (POD), glutathione peroxidase (GPx), glutathione reductase (GR), ascorbate peroxidase (Apx), malondialdehyde (MDA), superoxide radical (O2–), hydrogen peroxide (H2O2), and total phenol); and (iv) other metabolites (proline, protein) (Supplementary Table S3; dos Santos et al., 2022).
Data for meta-analysis was obtained from each study, according to the following criteria: (i) if data were reported on several species, different types of contaminants or in different locations in the same publication, all information was recorded and used; some individual publications, therefore, contributed to a larger number of studies to the analyses. (ii) When a publication contained several treatments that were not related to the core theme of this study, only the data of interest were selected; (iii) when data were reported with several concentrations of the stressing agent or spores of Trichoderma strains, the lowest and highest concentrations were ignored in order to have single and average concentrations to compare with control. These values collected from the articles were arranged in MS Excel® spreadsheets, and meta-analyses were performed using the OpenMEE software (http://www.cebm.brown.edu/openmee/), calculating the mean difference of the various effects’ sizes in each pairwise comparison.
For the scope of this work, a ‘study’ (also referred as an ‘entry’ for a meta-analysis) corresponds to primary data from individual pairwise comparisons between treatment and control (i.e., an experiment) obtained from the selected articles; hence, a single article may show pairwise data for one or more experiments, thus providing one or various entries to the analysis. To visualize the results of the meta-analyses, forest plots were used (Supplementary Figure S1); studies (entries in the graph) with the difference between treatment and control means showing values > 0 indicate an effect in favor of the treatment, and values < 0 indicate the opposite effect. These difference means for the treatment effect were weighed (%) in the meta-analysis according to the level of experimental variation in the corresponding study; this was depicted by the confidence intervals (CI), which were indicated by the horizontal lines of each study (entry). The higher the variation (larger CI), the lower the weight of that study in the meta-analysis. At the bottom of each forest plot (Supplementary Figure S1), the diamond represents the meta-analytic mean (MAm), which indicates the weighted average of the differences between treatments and control of the included studies (i.e., the total effect magnitude); the diamond’s width indicates the CI for the meta-analysis. The figures were generated and summarized by OpenMEE, and R Studio (ggplot2 3.5.1 package -Wickham, 2016- https://ggplot2.tidyverse.org) and edited in InkScape (https://inkscape.org/pt-br/).
Results
Scientometric analysis
After applying all the selection criteria, the final database for this meta-analysis comprised 55 articles (Supplementary Table S1), which were fully analyzed for the extraction of quali- and quantitative data (Supplementary Tables S2, S3). Despite the search had not applied a time restriction to the past, the pertinent selected studies were distributed between the years 2010 and 2020 (last year of our search), with the largest number of publications (13 articles) recorded in 2017 (Supplementary Table S2), evidencing global published research on abiotic stresses and plant-beneficial microorganisms in that decade. During the processing of the primary data collected, a greater number of articles for the meta-analyses were mostly related to saline (26), drought (17) and heavy metals (12) stresses on the plants (Table 1). Moreover, the metals-related stresses could be divided into Arsenic (6), Cadmium and Lead (4 each), Copper and Zinc (2 each) and Chromium and Nickel (1 each) (Supplementary Table S2; dos Santos et al., 2022). It is worth noting that the amount of heavy metals is greater than the corresponding amount of related articles, because single articles have addressed more than one metal (Supplementary Table S2). The search also identified other individual and combined stresses (dos Santos et al., 2022), which were not further assessed due to a very small, non-representative number of entries (data not shown).
Aiming at addressing the frequency and distribution of distinct Trichoderma species investigated in the main theme, our analysis identified 106 strains (see no. experiments in Table 1), distributed in 18 species, in addition to three entries showing no identification at the species level (Table 1; Supplementary Table S2). The most common species found in this study was Trichoderma harzianum, with 58 occurrences, followed by T. asperellum (11), T. atroviride (6), T. afroharzianum (5), T. longibrachiatum (4), T. virens (3), and T. hamatum, T. koningiopsis and T. viride (2 each). The species T. aggressivum, T. brevicompactum, T. britannicum, T. citrinoviride, T. crassum, T. koningii, T. reesei, T. saturnisporum, and T. viridescens were identified with one occurrence each (Supplementary Table S2). The names of these fungal species were updated based on the most recent designations, considering the described strains found in the articles and in taxonomic updating studies (e.g., Cai and Druzhinina, 2021). Some of the T. harzianum, T. asperellum, and T. atroviride strains were from bioproducts originally formulated for other purposes and were tested in these studies to analyze their effects on plant response to abiotic stresses.
Regarding the plants used in the experiments, a clear trend was observed in the studies, which have mostly worked with model species known for having short life cycles, small size, easy propagation, considerable economic relevance and wide use in research in the area of plant biology/agronomy. A total of 26 species were identified, the most commonly employed being Zea mays, with 10 occurrences in the articles (Table 1). Other plant species more frequently found were Oryza sativa (7), Arabidopsis thaliana (5), Triticum aestivum and Solanum lycopersicum (4 each) (Supplementary Table S2). Taking all 57 occurrences of studies involving plant species, the represented taxonomic families and occurrences were grasses (Poaceae, 23), legumes (Fabaceae, 10), vegetables (Brassicaceae, 6), Solanaceae (6), Cucurbitaceae (4), Asteraceae (3), Salicaceae (2) and Amaryllidaceae, Convoloulaceae e Resedaceae (1 each) (Supplementary Table S2).
In order to assess the collective effects of Trichoderma spp. in mediating plant responses to the three major abiotic stresses found in the scientific literature, 27 parameters (effect measures) were addressed under those different stresses (Supplementary Tables S3). Data on plant biomass, including dry and fresh weights of shoots and roots, shoot height and root length were the most frequently reported effect measures in the articles for the observation of results from the plant x abiotic stress x Trichoderma interactions (Supplementary Table S3). In the second group of response variables related to physiology and photosynthesis, the most frequently analyzed data, with higher number of entries, are associated with total chlorophyll (43) and relative water content (23), followed by net photosynthesis and stomata conductance (17 entries each). In the third group referring to enzymatic activities, studies addressing antioxidant enzymes stood out, mainly superoxide dismutase (SOD; 20 entries), catalase (CAT) and malondialdehyde (MDA; 18 entries each), and hydrogen peroxide (H2O2; 15 entries). Finally, in the metabolites group, the production of proline (21) was the one mostly assessed (Supplementary Table S3).
Effects on growth of plants subjected to abiotic stresses
Response variables related to growth and development (plant biomass parameters) correspond to important phenotypes of agronomic interest for several crops, since they are directly related to the end-products and yield (Habyarimana et al., 2020). Thus, through meta-analyses, we studied the effects of Trichoderma spp. presence on plant tolerance to important types of abiotic stress. Taken together, the interaction with Trichoderma spp. significantly increased plant biomass and growth (roots and shoots weight and height), under all three stress conditions studied (saline, drought and heavy metal stresses) (Figure 1). A large variability was observed for all biomass variables studied, both in relation to the magnitude of the differences in favor of Trichoderma, as indicated by the meta-analytic means (MAm), and to the experimental variation between the studies, observed through the varied sizes of the confidence intervals (CIs) (Figure 1). However, it is noted that for root and shoot dry weights (RDW and SDW, respectively), for the three stress types, a general trend of smaller magnitudes for MAm and for experimental variability in the studies (entries) was observed in favor of the Trichoderma spp. treatment (Figure 1). In terms of variation among studies included in the meta-analyses, it is worth mentioning that water-deficit stress was the independent variable showing greater variability in plant responses (larger CIs) for the fresh weights and root and shoot lengths variables (RFW, RL, SFW, SH). On the other hand, for RDW and SDW, greater magnitudes of MAm (with greater variability among entries) were observed for heavy metal compared to the other stresses (Figure 1).
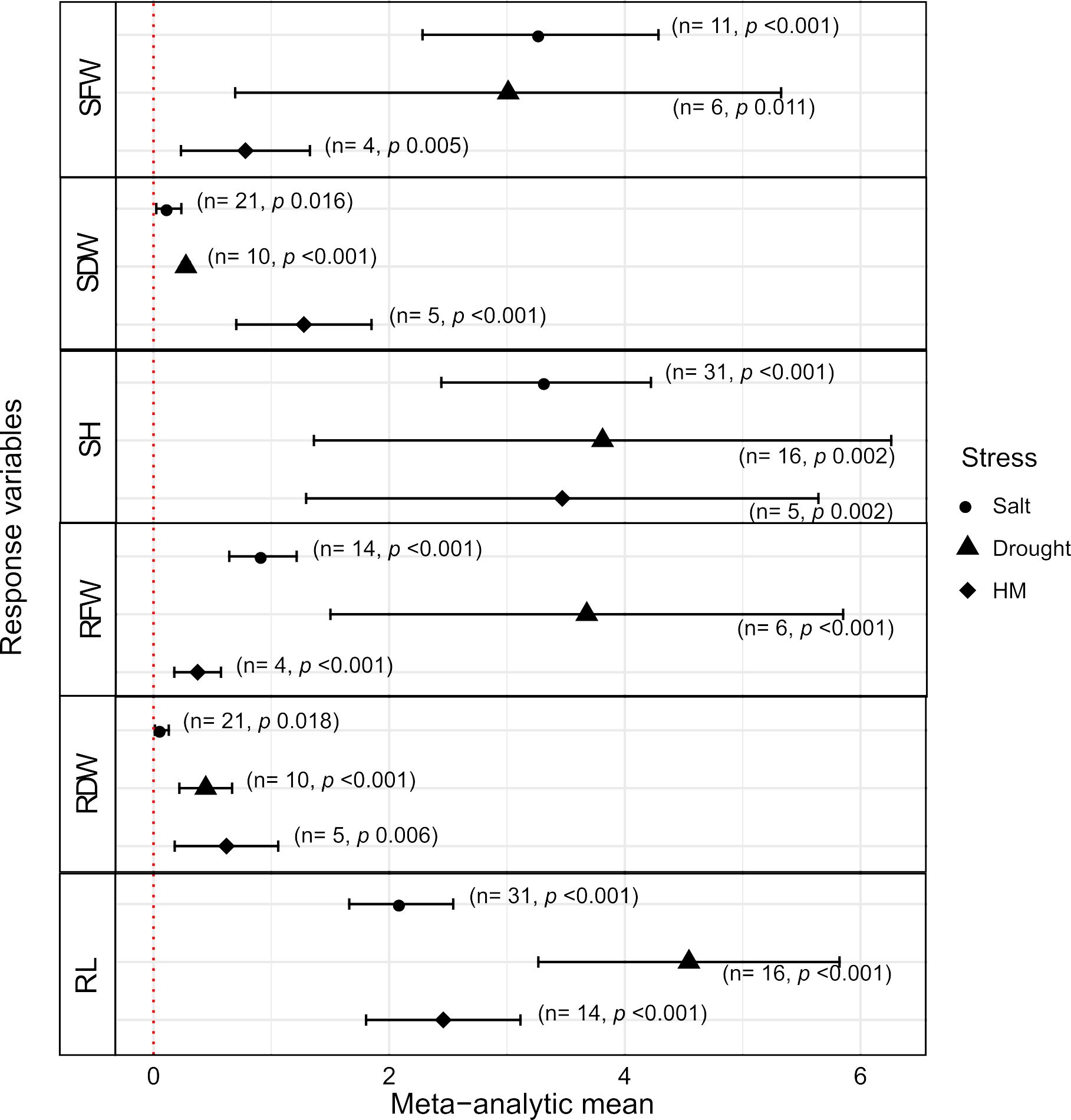
Figure 1. Effects of Trichoderma spp. on biomass and length of shoots and roots of plants under salt, drought and heavy metal (HM) stresses conditions. The effect measures correspond to the growth/development response variables: SFW – shoots fresh weight, SDW – shoots dry weight, SH – shoots height; RFW – roots fresh weight, RDW – roots dry weight, RL – roots length. The black symbols (circle, triangle, diamond) correspond to the meta-analytic means (differences between the average effects with and without Trichoderma), including the ±95% Confidence Interval (CI) horizontal bars. When CIs do not overlap the ‘zero’ on the horizontal scale of meta-analytic means, the effects are statistically significant (p < 0.05). Positive meta-analytic means indicate that the growth responses of plants + Trichoderma were significantly greater than those of plants without the fungus. In parentheses, ‘n’ is the number of studies included in the meta-analysis and ‘p’ is the p-value, that indicates the statistical significance of the meta-analytic mean.
Some details of the analysis regarding the magnitude of effects and their differences are important to highlight. First, let us consider the response variables for shoots; for SFW, the effect size in favor of Plant + Trichoderma spp interaction under saline (MAm = 3.284) and heavy metal (MAm = 0.78) stresses were different from each other, with no overlap of the respective CIs, the first being of greater magnitude (Figure 1; Supplementary Table S4). For SDW, the effects in favor of Trichoderma presence under heavy metal stress (MAm = 1.276) were significantly more prominent than under saline (MAm = 0.13) and water deficit (MAm = 0.275) stresses, which were of same magnitude, based on their CIs (Figure 1; Supplementary Table S4). Finally, for SH, there was no difference in magnitude among the positive effects of Trichoderma for the three types of stress, based on the overlapping CI values (Figure 1; Supplementary Table S4). Second, concerning the variables related to roots, the effects of Trichoderma were different from those for shoots, based on the respective CIs and MAm: (i) for RFW, the three stresses produced significantly different results, with a greater magnitude of MAm (3.677) for drought stress; (ii) for RDW, there was no difference in the Trichoderma effect among the three stresses, with all MAm < 0.62; and (iii) for RL, the beneficial effect of Trichoderma for water deficit stress (MAm = 4.544) was significantly greater than the others, which did not differ statistically from each other (Figure 1; Supplementary Table S4).
Effects on physiology/photosynthesis in plants subjected to abiotic stress
A second set of parameters evaluated in this systematic survey refers specifically to physiological response variables linked to photosynthesis. Typically, this is a first layer of in-depth analysis of the effects of some treatment on plants, since photosynthesis is closely linked to growth, development and yield parameters (e.g., Figure 1) (Razi and Muneer, 2021). To verify the effects of plant-Trichoderma interaction on usual photosynthetic parameters as descriptors of physiological responses to environmental stimuli, the primary data on these parameters in the selected database were evaluated (Figure 2). For the total chlorophyll (ChlT) and net photosynthesis (Pn) variables, we found data for the three stress types, although only 1 study (entry) was found for the former under heavy metal stress (Supplementary Table S3). For the other physiological/photosynthetic variables, we combined the data from two out of the three stresses available for the meta-analyses (Figure 2; Supplementary Tables S3, S4). The results showed that, for the three types of stress tested, the Trichoderma effects on Pn and ChlT were significantly greater than the absence of fungus application (Supplementary Table S4). For Pn, statistical difference was only observed between heavy metal and saline stress, the latter with the greatest difference in favor of Trichoderma treatments; on the other hand, for ChlT, the three studied stresses were significantly different from each other, with the greatest difference in favor of Trichoderma for heavy metal stress (Supplementary Table S4).
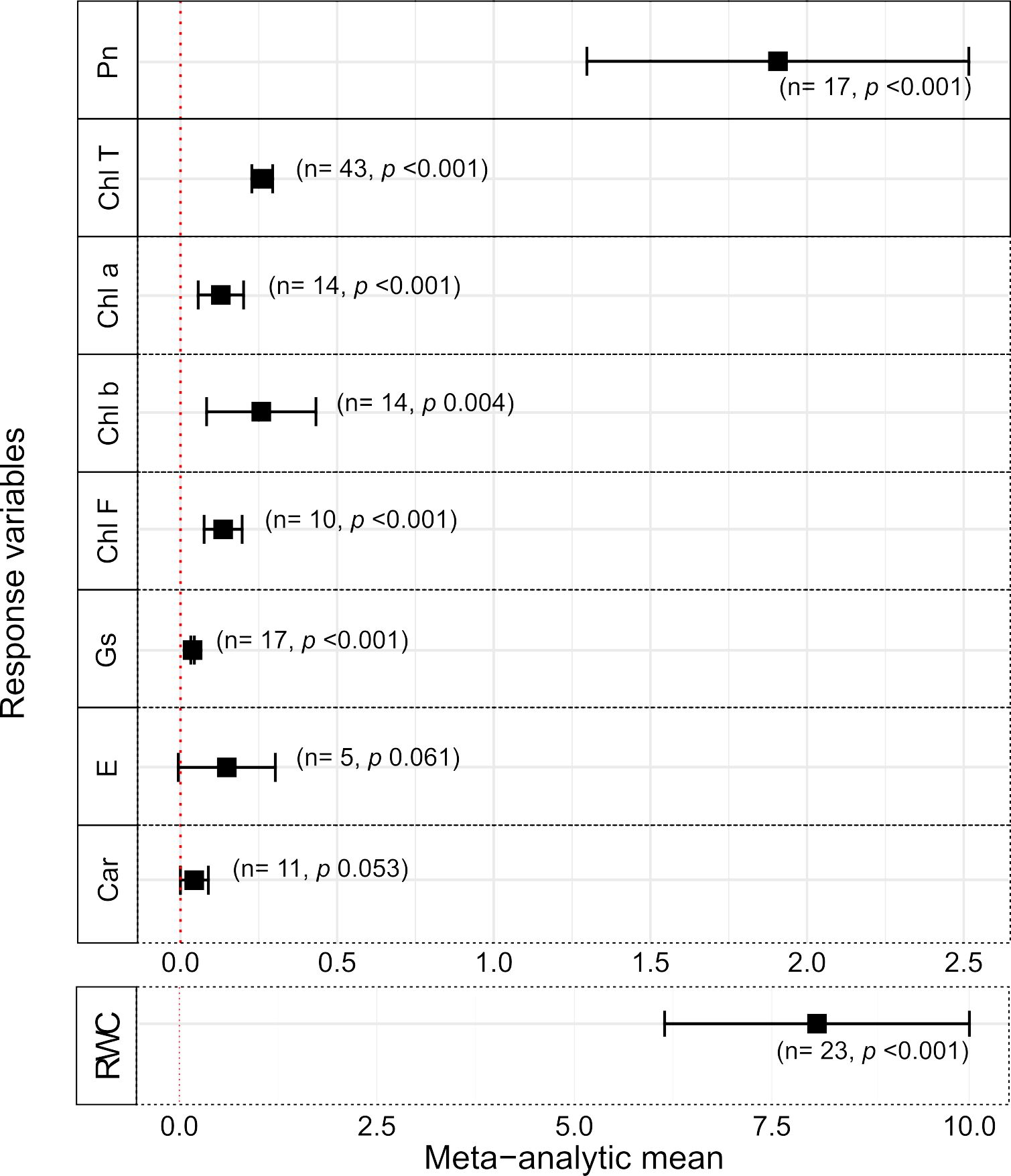
Figure 2. Effects of Trichoderma spp. on physiological/photosynthetic variables of plants under abiotic stresses. Black squares indicate the meta-analystic mean (MAm) for studies’ data of all stress types taken together, with corresponding ±95% CIs shown by horizontal bars (see legend of Figure 1 for other details). Pn, Net photosynthesis; Chl T, total chlorophyll; Chl a, chlorophyll a; Chl b, chlorophyll b; Chl F, chlorophyll fluorescence; Gs, stomatal conductance; E, transpiration; Car, carotenoids; and RWC, relative water content.
A variety of statistical results were observed for the other physiology/photosynthesis parameters studied, considering the response variables, type of stress and statistical significance of the differences (Supplementary Table S3). It is important to highlight that most of the physiological variables and primary data found in our survey showed results for only two of the three stress types, i.e., saline, drought and heavy metals (Supplementary Table S3). In these cases, the respective comparison groups (with vs without Trichoderma) were analyzed, and only those with a minimum number (n) of two entries were considered (Supplementary Table S3). Although mostly of small magnitudes (Supplementary Table S4), positive and significant effects (p < 0.05) of Trichoderma spp treatments in the meta-analytical means (MAm) were observed for the variables under stress as follows: (i) chlorophyll fluorescence (ChlF; MAm = 0.192), stomatal conductance (Gs; 0.011), chlorophyll a (Chl a; 0.299) and relative water content (RWC; 5.496) for salt stress; (ii) Gs (0.02) and RWC (14.794), for water deficit; and (iii) Chl a (0.145) and Chl b (0.61) for heavy metal stress. Only a single study (entry; n = 1) was found for ChlT under heavy metal stress, transpiration (E) under salt stress, and carotenoid levels (Car) under drought (Supplementary Table S3), therefore, they were not included in the meta-analyses. On the other hand, when the stresses were taken together, significant meta-analytic means in favor of Trichoderma effect were found for ChlF, Chl a, Chl b, Gs, and RWC, the latter with the largest magnitude of difference in the MAm (8.072) between the comparison groups (Figures 2; Supplementary Tables S1, S4). Only two variables, E (drought) and Car (salt stress) did not show a significant difference between the comparison groups (Figure 2). Finally, in the set of evaluated articles, we did not find data for the Chl a and b variables under drought, and for ChlF, E, Gs, Car and RDW under heavy metal stress.
Effects on antioxidant and enzymatic activities of plants subjected to abiotic stresses
At the biochemical/molecular level, the analysis of effect variables linked to enzymatic reactions and metabolisms in response to environmental stimuli offers insights into associated gene expression patterns, which may be potential targets for biotechnological manipulation (Hasanuzzaman et al., 2020). Thus, based on the primary data available in the selected article database, we aimed at assessing the effects of Trichoderma spp in the modulation of enzymatic and antioxidant activities, usually associated with the regulation of plant response to a variety of stresses. Overall, depending on the stress type, the favorable effects of Trichoderma were less pronounced, with some antioxidants and other enzymatic activities not differing significantly between treatments with and without the fungus (Figure 3). The antioxidant activities, i.e., superoxide dismutase (SOD), ascorbate peroxidase (APx), glutathione reductase (GR), peroxidases (POD) and catalase (CAT), showed distinct responses. In the presence of Trichoderma, APx (MAm = 5.847, p < 0.001) and SOD (MAm = 35.903, p = 0.023) showed significantly altered responses in plants under salt and heavy metal stresses, but no significant effect under drought conditions, whereas no significant effect was noticed for POD (Figures 3, Supplementary Tables S1, S4). The responses of guaiacol peroxidase (GPx), GR and CAT were not significantly different between the controls and fungal treatments, with the exception of water deficit stress for the CAT response (MAm = 1.515, p < 0.001; Supplementary Table S4).
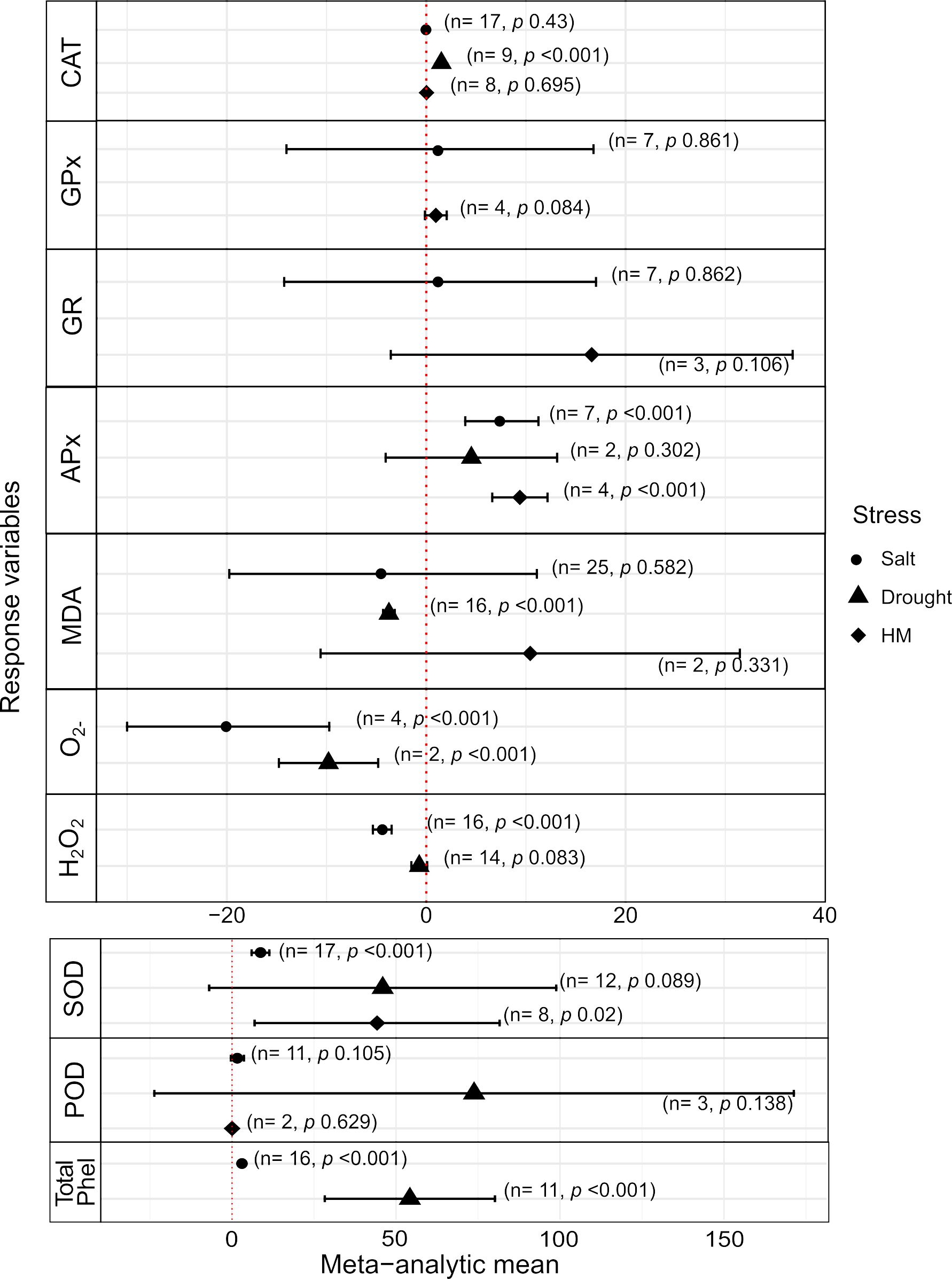
Figure 3. Effects of Trichoderma spp. on plant enzymatic activities under three types of abiotic stresses. See legend of Figure 1 for black symbols, horizontal bars, and information in parenthesis. CAT, catalase; GPx, guaiacol peroxidase; GR, glutathione reductase; APx, ascorbate peroxidase; MDA, malondialdehyde; O2–, superoxide radical; H2O2, hydrogen peroxide; SOD, superoxide dismutase; POD, peroxidase; Total Phel, total phenolics content.
The generation of malondialdehyde (MDA), a product of lipid peroxidation, under stress is related to the production of reactive oxygen species (ROS) in plant tissues, including hydrogen peroxide (H2O2) and the superoxide radical (O2–) (Hasanuzzaman et al., 2020). The comparative MDA content for heavy metal stress came from a very low number of studies found (n = 2), and with no statistical significance for the difference (Figure 3); in addition, no data were found for H2O2 and O2– under heavy metals. Under the other two stress conditions, plants inoculated with Trichoderma produced significantly less MDA only for drought, and less ROS (H2O2 and O2–) for both salt excess and water deficit conditions, with exception of H2O2 under drought (Figure 3). Overall, the reduction levels of O2– were more pronounced than for H2O2 in plants+Trichoderma for both stresses, despite a relatively low number of inputs for salt (n = 4) and drought (n = 2) (Figure 3; Supplementary Table S4). Finally, total phenol production generally indicates an adaptive response of plants to stress, as they have the ability to neutralize free radicals (Lakhdari et al., 2023). In the present study, a significant increase in the production of these compounds (simple phenols, flavonoids, and tannins) was observed in the plant-Trichoderma interaction, considering the saline and drought stresses (MAm = 3.144 and 54.212, respectively; p = <0.001); such a greater magnitude increases in phenols observed in water deficit stress stands out (Figure 3; Supplementary Table S4).
Considering previous evidence of tissue/organ-specific activities in several plant systems for the studied enzymes related to stress responses (e.g., Yu et al., 2017; Anzano et al., 2022; Liang et al., 2023), we assessed the proportion of organs/plant parts investigated in the studies selected for the meta-analysis. The analyses of these investigated activities (see Figure 3) indicated a marked predominance of primary evaluations performed in leaves (79.5%), followed in a much smaller proportion by roots (14.4%) and seedlings (6.1%) (Supplementary Table S5). In this context, specific meta-analyses were performed by plant organs in relation to enzymatic activities, with and without the presence of Trichoderma spp. (Supplementary Table S5). The results confirmed that the responses of enzymatic activities to stresses varied according to the enzyme and the plant organ analyzed, for both Trichoderma treatments. For instance, it was observed that SOD and APx activity was significantly increased in the presence of Trichoderma spp. (p < 0.05; Supplementary Table S5), both in leaves and roots, while there was no significant effect in seedlings only for SOD. On the other hand, GPx showed a significant effect in favor of Trichoderma only for seedlings (Supplementary Table S5). The enzymes CAT and POD showed a significant increase in activity in leaves in the presence of Trichoderma spp. (p < 0.001), but for roots, the increase in these enzymes under stress occurred in the absence of Trichoderma spp. (p = 0.006 and p < 0.001, respectively). Furthermore, compounds indicative of oxidative stress, such as MDA, O2- and H2O2, showed significant reduction in leaves and roots for treatment with Trichoderma species (Supplementary Table S5).
Effects on metabolites production of plants subjected to abiotic stress
The production of two types of cellular compounds, proline and protein contents, were those observed in the selected primary studies; these variables were significantly different in plants with and without Trichoderma under abiotic stresses (Figure 4; Supplementary Table S4). In plants, proline is an amino acid that plays important roles of regulating osmotic balance, protecting cell membranes against damage, and stabilizing structural proteins (Hosseinifard et al., 2022). The general interest of research appeared to be relevant for osmotic stress responses in plants, since the numbers of entries involving proline for the meta-analyses were large in this systematic research, i.e., n = 36 for salt stress, n = 14 for water deficit, and n = 3 for heavy metals (Supplementary Table S3), in a total of n = 53. The results on proline content in plants were interesting; for those subjected to saline stress, this variable responded positively to the presence of Trichoderma, significantly increasing the production of this small osmolyte (MAm = 4.027). However, under drought stress, the result was the opposite: there was a significant decrease in proline production in the presence of the fungus (MAm = –6.96). Finally, under heavy metal stress, there was no significant difference in proline content between control and treatment (p = 0.788), although the number of studies for this case was very low (Figure 4).
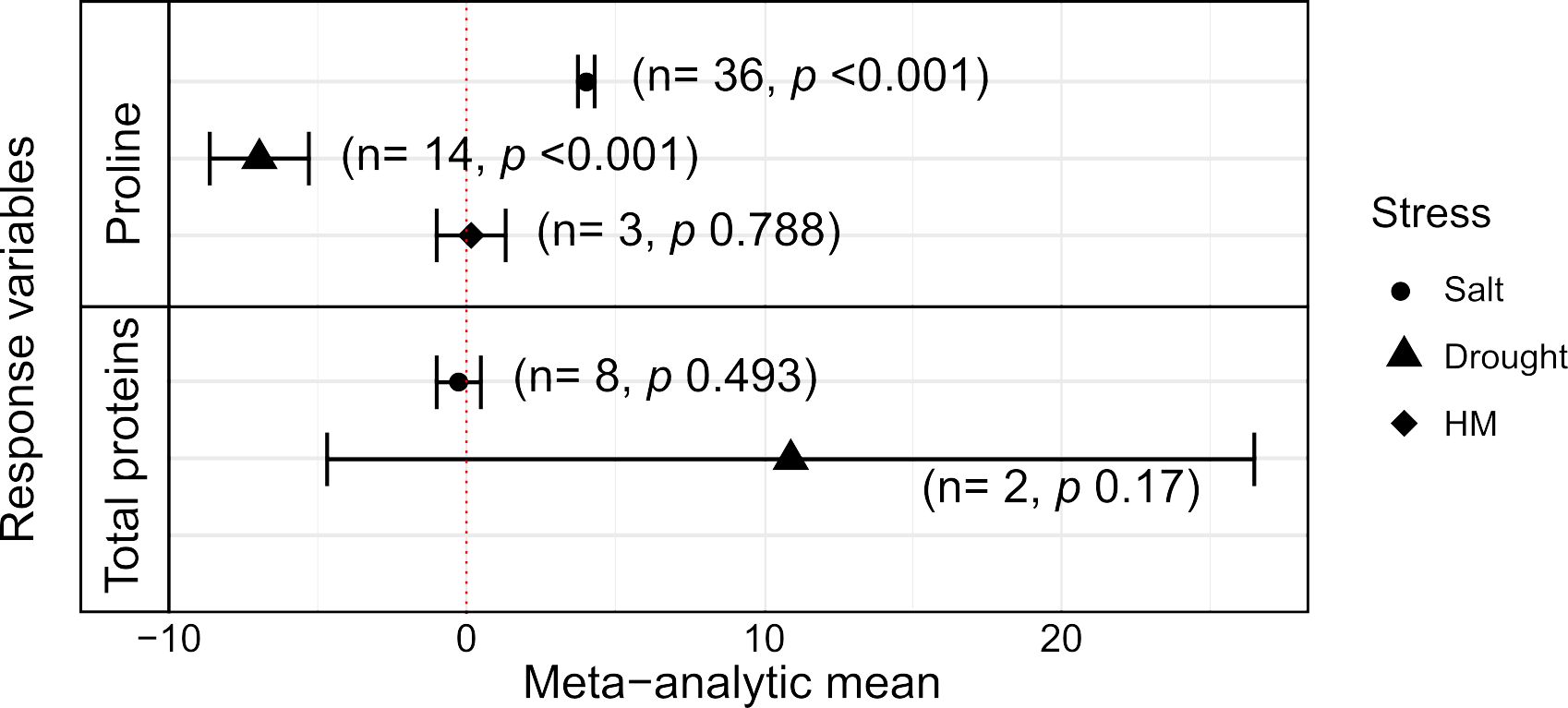
Figure 4. Effects of Trichoderma spp. on proline and total protein content in plants under three types of abiotic stresses. Black symbols and horizontal bars indicate the meta-analytic mean for each type of stress and the corresponding ±95% CIs, respectively; In parenthesis, ‘n’ is the number of studies (entries) included, and ‘p’ is the p-value for statistical significance of the meta-analyses.
The total protein content is a response variable that can be considered as providing information about the general health of the plants; the meta-analysis results, however, indicated that, under salt and drought stresses, there were no significant differences between treatments with and without Trichoderma spp. applied to the plants (Figure 4). We have not addressed the condition of heavy metal stress for this variable, as only a single study was found for this condition (Supplementary Table S3).
The analysis of differential content of these metabolites among the plant tissues was also addressed in relation to presence/absence of Trichoderma in plants under stress (Supplementary Table S5). Similarly to the enzymatic activities described above, proline was mostly studied in leaves (79.7% of studies), followed by roots and seedlings (14.1% and 6.2%, respectively). This cellular osmolyte showed a significant increase in all organs analyzed for the presence of Trichoderma spp. (p < 0.001), with a notably high magnitude of effect on seedlings, although caution must be taken with these results due to the very low number of studies in this sub-group (Supplementary Table S5). On the other hand, the total proteins variable did not show significant differences in any of the organs evaluated, although there were not that many studies reporting this type of information (Supplementary Table S5).
Discussion
Climate change poses major challenges to plant species, since scenarios of severe droughts, extremely high or low temperatures, or excessive short-term rainfalls are predicted to occur more frequently in coming years (Zandalinas et al., 2021). These abiotic stresses can significantly impact crop productivity and quality, which can generate food insecurity for human populations (Wang and Frei, 2011; Kumar, 2020). In this context, it is essential to devise approaches that aim to improve plant fitness and, consequently, their productivity. Microorganisms with multiple beneficial relationships with plants has been extensively researched, such as fungi of the Trichoderma genus. Several studies have reported these species are multifunctional, displaying various activities such as biocontrol, biofertilization and phytostimulation; they have shown abilities to produce different beneficial bioactive compounds and elicitors that interact with host plants, thus increasing plant growth, development and resistance/tolerance to various abiotic stresses (Zhang et al., 2018; Sood et al., 2020; Abdenaceur et al., 2022; dos Santos et al, 2022). In this study, a meta-analysis was carried out on empirical evidence in the literature related to benefits that plant species acquire in the presence of Trichoderma spp. under abiotic stresses. Strengths and gaps in this topic, as well as consequences and potential applications of our findings were discussed.
Abiotic stresses can cause up to 60% or more reduction in crop yields (Kopecká et al., 2023); the stresses addressed in this meta-analysis – salinity, drought, and heavy metals – represent major factors that limit agricultural production quali- and quantitatively on a global scale (Li et al., 2023). These environmental stress factors generate osmotic stress, due to an imbalanced concentration of solutes inside and outside the cells, and oxidative stress, due to the generation of ROS, which can disrupt the integrity of the cell wall/membranes (Nadarajah, 2020; Atta et al., 2021; Askari-Khorasgani et al., 2021). In brief, salinity stress is deleterious to plants, due to reduction in the osmotic potential of water in the soil, turning it less available to roots. Moreover, excess of salts in the soil affects its physical structure, reducing aeration and water permeability, besides increasing concentration of Na+ and Cl– ions in the plant, which can be toxic to metabolism (Badar et al., 2015; Dastogeer et al., 2020), resulting in deleterious physiological effects. Drought also causes osmotic stress in plants by reducing absorption of water and nutrients from the soil, which hinders plant respiration, photosynthesis and stomatal movement, among other essential functions (Yang et al., 2021; Seleiman et al., 2021). Finally, heavy metals excess (pollution) turn the soil into a toxic environment for plants. When such metals penetrate inner layers of plants, they cause inactivation and denaturation of intracellular enzymes, changes in protein structures and disruption of membrane integrity, although these effects vary according to the metal and plant species (Feng et al., 2023). This, hence, affects metabolic/physiological processes, and so, normal plant growth.
The need to understand mechanisms of resistance to abiotic stress to promote sustainable agriculture in a changing climate seems to reflect the growing number of publications about crops focusing on this theme. Since 2010, there has been a noticeable increase in such studies, linked with the recognition of Trichoderma genus as bearing useful fungi beyond a more frequent use as biocontrol agents of plant diseases (dos Santos et al., 2022). Since then, studies have been unraveling mechanisms and abilities of Trichoderma spp to modulate multiple stress factors in plants, and in their seeds and seedlings, in addition to the ability of inducing plant systemic resistance (Saba et al., 2012; Dutta et al., 2022). Trichoderma-based products have grown in the agricultural market during this period, such that in 2014 there were more than 250 of such bioproducts registered worldwide (Woo et al., 2014), many with an increased economic interest due to their multiple functions and applications (Dutta et al., 2022).
According to the selected material for this meta-analysis, the most frequent plant hosts under study share some characteristics: considerable economic relevance, wide use as model species in biological research use, rapid growth, cosmopolitan distribution, shorter life cycles (annual plants), smaller sizes, and easy propagation. Trichoderma spp. tend to be part of dominant mycoflora in soils from cold temperate to tropical climates (Dutta et al., 2022); the presence of these fungal species in the rhizosphere protects and assists plants in two ways: (i) a pre-colonization interaction, where a “dialogue” between hosts and fungi occurs through diffusible soil metabolites and volatile organic compounds, in addition to the availability of nutrients, due to the saprophytic action of Trichoderma (Contreras-Cornejo et al., 2024); and (ii) effective colonization, where fungal hyphae interact with plant root/shoot tissues, allowing modulation of the host response to an array of biotic and abiotic stressors (Salwan et al., 2022; Contreras-Cornejo et al., 2024). Interestingly, there are evidences that beneficial interactions between plants and Trichoderma can be inherited by the host, being activated in subsequent generations in a facilitated manner, thus acting as an immune-boosting “memory” (Morán-Diez et al., 2021; Contreras-Cornejo et al., 2024). Hence, all the effects observed on our meta-analyses have a well described (though incomplete) knowledge framework that help explaining most of our findings.
Stressing conditions result in changes at the morphophysiological, photosynthetic, biochemical and molecular levels that affect plant development and growth. With this conceptual framework in mind, we have divided the stress-response plant variables assessed into four groups of parameters, due to a similarity in their biological functions within each group (Figures 1 to 4; dos Santos et al., 2022). The first dataset analyzed in this study focused on the length and biomass of plants subjected to abiotic stress conditions (Figure 1). These phenotypes tend to be primary characteristics to be observed in plants due to being major parameters related to plant yield and to the ease of measurements/visual assessments. Plant treatments with and without Trichoderma isolates were compared, with the meta-analytic results demonstrating an ability of these fungi to mitigate negative effects of stress on plants by an overall increase in roots/shoots biomasses and length (Figure 1). These results can be explained by the fact that, in general, Trichodermas improve the growth and fitness of host plants, directly and indirectly, by using resources available in the soil (Supplementary Table S2; dos Santos et al., 2022). It has been shown that Trichoderma spp. can solubilize macro and micronutrients and make them available to plants. For example, Babu et al. (2014) have demonstrated that T. virens PDR-28 promoted both the growth of maize and bioremediation of heavy metals due to the production of siderophores, which in addition to making iron available to the host, also chelated the heavy metals, turning them into less toxic forms to the plant. Trichoderma spp. also produce phytohormones and metabolites that influence the host’s hormonal balance, including auxins (indole-3-acetic acid), gibberellins, and the ACC-deaminase enzyme (Reghmit, 2023). Zhang et al. (2015) have shown that transgenic Arabidopsis expressing ACC-deaminase from T. asperellum grew faster under normal and saline conditions, and improved salt tolerance, likely because this enzyme reduces ethylene, a regulatory phytohormone produced by plants under stress conditions, which tends to inhibit root elongation and cause plant death in high concentrations. Moreover, Trichoderma produces bioactive metabolites, including organic molecules (ketones, esters, aldehydes, alcohols, fatty and organic acids), hydrocarbons, non-ribosomal peptides (NRPs), peptaibols and other compounds that can positively modulate physiological aspects in the host plant, thereby maintaining its integrity and well-being (Contreras-Cornejo et al., 2024).
The second group of evaluated parameters was related to physiological and photosynthetic response variables of plants under the different stress conditions taken together, with the results pointing to an overall positive effect of Trichoderma treatments (Figure 2). Plants subjected to abiotic stresses suffer damage to their photosystems and physiological processes, such as chlorophyll biosynthesis, electron transport, and gas exchange (mainly caused by oxidative stress), thus decreasing their photosynthetic capacity (Muhammad et al., 2021). Research by Harman et al. (2021) shows that Trichoderma strains are capable of increasing the photosynthetic efficiency of host plants under environmental stresses, by promoting synthesis and accumulation of photosynthetic pigments, and by upregulating expression of genes that regulate the biosynthesis of chlorophyll and proteins of the light-harvesting complexes. In addition, plants colonized by Trichoderma spp. improve the efficiency of water use by roots, as they significantly increase the production of growth hormones, as mentioned above; these are responsible for root elongation, which leads to greater absorption of water and nutrients from the soil, resulting in increased levels of photosynthetic pigments (Gupta et al., 2022). These plants also increase carbon sink activity through root exudates, positively manipulating the rate of photosynthesis in leaves (Oljira et al., 2020; Harman et al., 2021).
The third group of plant response variables assessed is associated with the enzymatic and non-enzymatic antioxidant activities that the Trichoderma-host interaction under abiotic stress can produce. The results suggest that, taken together, these parameters tended to be less sensitive in general to detect Trichoderma effects, since the statistical significance detected has varied according to the stress type and the variable assessed (Figure 3). Nevertheless, when significant differences were found, the favorable effect of Trichoderma was evident, although at distinct magnitudes, depending on the variable. The results also allowed us to evaluate conditions related to oxidative stress caused by those abiotic stressors in plants. In general, oxidative stress is defined by the overproduction of ROS, such as superoxide (O2–), hydroxyl (OH–) and hydrogen peroxide (H2O2) (Mittler, 2002). These molecules react chemically with practically all plant metabolites, including proteins, lipids and nucleic acids, causing damage to them. The result of ROS excess in plants is an increased metabolic toxicity, damage to membranes, inhibition of photosynthesis, changes in phytohormonal levels, among other deleterious effects (Selvakumar et al., 2012). On the other hand, in homeostasis, low levels of ROS act as cellular signals, promoting acclimation and an adjusted tolerance to stress (Foyer, 2018). Our results demonstrated that plant species subjected to abiotic stresses tend to produce more ROS compared to those inoculated with Trichoderma spp., thereby pointing to an ameliorating action of these fungi. The same occurs with the production of MDA under water deficit conditions, an indicator of membrane lipid peroxidation caused by ROS. The ROS processing system in plants keeps the balance between conditions of ROS-causing damages and its detoxification; this is maintained by the fine-modulation mechanism of antioxidant enzymes and non-enzymatic compounds (Mittler, 2002; Foyer, 2018).
Studies have suggested that Trichoderma spp. use and depend upon the ROS processing system for several cellular processes and responses to environmental changes (Cruz-Magalhães et al., 2019). To maintain this redox balance in the cells, species with sequenced genomes, such as T. reesei, T. atroviride and T. virens, have shown to display a robust antioxidant system, with three SOD genes (one Mn SOD and two Cu/Zn SODs), seven catalases and three catalase peroxidases (Schmoll et al., 2016). These enzymes are homologous to the redox system of plants, which convert ROS into less toxic molecules and participate in the water-water cycle and the ascorbate (AsA) and/or glutathione (GSH) regeneration cycle (Mittler, 2002; Foyer, 2018). These data and information corroborate a major hypothesis proposed in this study, which is that Trichoderma species essentially deploy similar mechanisms of plants to survive adversities. During the process of plant-Trichoderma communication, such mechanisms allow the establishment of specialized interactions, likely with a modulatory interference on the host. Also, in response to oxidative stress, Trichodermas accumulate phenolic compounds, which are associated with the biochemical defense of plants (Lakhdari et al., 2023). Increased phenol synthesis has a direct effect on antioxidant activity, acting as ROS scavengers and contributing to the formation of cell wall, thereby strengthening physical barriers of plants (Lakhdari et al., 2023).
Finally, we have assessed a couple of metabolites involved in this interaction, which could be identified in the selected primary studies (Supplementary Table S1). A recurrent mechanism of action found for Trichoderma spp. in this study was the production and accumulation of proline, an amino acid that acts as an osmolyte for cellular osmotic adjustment (Harman et al., 2021). In addition, proline also functions as a chaperone, stabilizing the structure of cytoplasmic proteins, and its accumulation buffers the cytosolic pH, maintains the redox state of the cell and ensures adequate NADP+/NADPH ratios compatible with metabolism (Hayat et al., 2012). This mechanism is particularly effective under salt stress conditions, as observed in the studies evaluated in this meta-analysis and in other reviews (Hayat et al., 2012; Rawat et al., 2013; Kumar et al., 2017; Pehlivan et al., 2017; Yasmeen and Siddiqui, 2017). Apparently, though, the mechanism of proline production and accumulation has not been observed in treatments with Trichoderma under water deficit (Figure 4). Under drought, host plants without fungal isolates seem to respond more efficiently with this mechanism. Shukla et al. (2012) justify these findings by suggesting that Trichoderma species may use other mechanisms to improve host fitness under stress, which may otherwise induce a negative regulation of proline production. This is corroborated by experimental results showing a better response in physiological and photosynthetic parameters in the rice-Trichoderma interaction under water deficit, with plants demonstrating greater resistance to wilting in response to drought (Shukla et al., 2012).
Several studies have been showing that genes encoding antioxidant enzymes, as well as synthesis of stress-response metabolites, all display tissue/organ specific patterns of expression (Yu et al., 2017; Anzano et al., 2022; Liang et al., 2023). In the scope of our meta-analyses, we were limited to the studies with data on application of Trichoderma spp. on plants under stress (see supplemental material), and consequently, to the organ types used in those studies. The majority of our collected data were related to research on leaves, which is likely a consequence of their relevance as a physiology-regulatory organ in terms of photosynthesis, gas exchange, water absorption and evapotranspiration. Moreover, the general ease of primary data collection and analyses in leaves may also contribute to the observed preference of studies with this organ. Nevertheless, despite such unequal distribution of collected data among plant organs (Supplementary Table S5), the meta-analytical results confirmed tissue/organ-specific responses to stresses in relation to presence of Trichoderma spp., both for enzymatic activities and specific metabolites related to homeostasis maintenance in plants (Nadarajah, 2020; Singh et al., 2022). The results, though, suggest that further studies on stress-responsive enzymes and metabolites in other under- or non-represented tissues are warranted to deepen our understanding of Trichoderma spp ameliorating effects in plants under abiotic stresses.
Conclusions
The scientific reports related to the various biological activities of Trichoderma spp. with beneficial effects on host plants can add value to existing bioproducts and to derivatives of species of this fungal genus. Discussing the practical applications and feasibility of using Trichoderma spp. in relation to the benefits they can promote in sustainable agriculture is of paramount importance. There are more than 250 agricultural bioproducts based on Trichoderma registered and commercialized worldwide (Woo et al., 2014). These products are marketed as biopesticides, biofungicides, bioprotectants, bioinoculants, biostimulants, biodecomposers, biofertilizers and plant growth promoters.
The meta-analytical approach offers the opportunity to gain knowledge of secondary nature on the informational spaces left among the primary studies, based on the empirical data accessible through literature databases. By organizing, quantifying and consolidating primary evidence of Trichoderma contribution to resistance and adaptation of plant species to drought, salt excess and heavy metals abiotic stresses, we provided a glance at the biotechnological potential of this fungal species beyond its common use as biocontrol agent. The series of meta-analyses we performed revealed that interactions with Trichoderma spp. tended to increase roots/shoots biomass and length, as well as total chlorophyll and net photosynthesis of plants under all three stress types. Moreover, control of oxidative stress in plants with a reduction in ROS production, and regulation of osmotic balance in drought or high salinity conditions through proline production and accumulation were observed for Trichoderma treatments. For the stress-responsive enzymatic activities and metabolites, organ-specific results were noticed under presence/absence of Trichoderma spp. This work brought to light and integrated the evidence in the literature on the potential use of Trichoderma species as the basis of bioproducts aiming to manage abiotic stress conditions in plants of agricultural and environmental interests. We believe the information presented here is valuable for the current context of undergoing climate and natural environment changes.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Author contributions
LS: Conceptualization, Formal analysis, Data curation, Investigation, Methodology, Software, Writing – original draft, Writing – review & editing. NO-S: Conceptualization, Methodology, Writing – review & editing. DN: Data curation, Formal analysis, Methodology, Software, Writing – review & editing. VM: Conceptualization, Writing – review & editing. LL: Conceptualization, Formal analysis, Funding acquisition, Methodology, Project administration, Supervision, Validation, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Acknowledgments
The authors are grateful to the Post-graduation Program of Genetics and Molecular Biology (PPG-GBM) from the State University of Santa Cruz (UESC) for infrastructural support. Supporting scholarships for living expenses were granted as doctoral fellowships to L.B.P.R.S by the Bahia State Research Support Foundation (FAPESB) and to N.O-S. by the Coordination for the Improvement of Higher Education Personnel - Brazil (CAPES) - Financing Code 001; V.C-M. received a post-doctoral fellowship from PNPD-CAPES.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphgy.2025.1569221/full#supplementary-material
References
Abdenaceur R., Farida B. T., Mourad D., Rima H., Zahia O., Fatma S. H. (2022). Effective biofertilizer Trichoderma spp. isolates with enzymatic activity and metabolites enhancing plant growth. I. nt. Microbiol. 25, 817–829. doi: 10.1007/s10123-022-00263-8
Anzano A., Bonanomi G., Mazzoleni S., Lanzotti V. (2022). Plant metabolomics in biotic and abiotic stress: a critical overview. Phytochem. Rev. 21, 503–524. doi: 10.1007/s11101-021-09786-w
Askari-Khorasgani O., Rehmani M. I. A., Wani S. H., Kumar A. (2021). “Osmotic stress: an outcome of drought and salinity,” in Handbook of plant and crop physiology (Boca Raton, Florida: CRC Press), 445–464.
Atta K., Pal A. K., Jana K. (2021). Effects of salinity, drought and heavy metal stress during seed germination stage in ricebean [Vigna umbellata (Thunb.) Ohwi and Ohashi. Plant Physiol. Rep. 26, 109–115. doi: 10.1007/s40502-020-00542-4
Babu A. G., Shim J., Bang K. S., Shea P. J., Oh B. T. (2014). Trichoderma virens PDR-28: a heavy metal-tolerant and plant growth-promoting fungus for remediation and bioenergy crop production on mine tailing soil. J.E.M. 132, 129–134. doi: 10.1016/j.jenvman.2013.10.009
Badar R., Batool B., Ansari A., Mustafa S., Ajmal A., Perveen S. (2015). Amelioration of salt affected soils for cowpea growth by application of organic amendments. J. Pharmacogn. Phytochem. 3, 87–90.
Cai F., Druzhinina I. S. (2021). In honor of John Bissett: authoritative guidelines on molecular identification of Trichoderma. Fungal. Divers. 107, 1–69. doi: 10.1007/s13225-020-00464-4
Catania F., Krohs U., Chittò M., Ferro D., Ferro K., Lepennetier G., et al. (2017). The hologenome concept: we need to incorporate function. Theor. Biosci. 136, 89–98. doi: 10.1007/s12064-016-0240-z
Contreras-Cornejo H. A., Schmoll M., Esquivel-Ayala B. A., González-Esquivel C. E., Rocha-Ramírez V., Larsen J. (2024). Mechanisms for plant growth promotion activated by Trichoderma in natural and managed terrestrial ecosystem. Microbiol. Res. 281, 127621. doi: 10.1016/j.micres.2024.127621
Cruz-Magalhães V., Nieto-Jacobo M. F., van Zijll de Jong E., Rostás M., Padilla-Arizmendi F., Kandula D., et al. (2019). The NADPH oxidases Nox1 and Nox2 differentially regulate volatile organic compounds, fungistatic activity, plant growth promotion and nutrient assimilation in Trichoderma atroviride. Front. Microbiol. 9, 3271. doi: 10.3389/fmicb.2018.03271
Dastogeer K. M., Zahan M. I., Tahjib-Ul-Arif M., Akter M. A., Okazaki S. (2020). Plant salinity tolerance conferred by arbuscular mycorrhizal fungi and associated mechanisms: a meta-analysis. Front. Plant Sci. 11, 588550. doi: 10.3389/fpls.2020.588550
dos Santos L. B. P. R., Oliveira-Santos N., Fernandes J. V., Jaimes-Martinez J. C., De Souza J. T., Cruz-Magalhães V., et al. (2022). “Tolerance to and alleviation of abiotic stresses in plants mediated by Trichoderma spp,” in Advances in Trichoderma biology for agricultural applications (Springer International Publishing, Cham), 321–359.
Druzhinina I. S., Seidl-Seiboth V., Herrera-Estrella A., Horwitz B. A., Kenerley C. M., Monte E., et al. (2011). Trichoderma: the genomics of opportunistic success. Nat. Rev. Microbiol. 9, 749–759. doi: 10.1038/nrmicro2637
Durán N., Marcato P. D. (2011). “Biotechnological routes to metallic nanoparticles production: mechanistic aspects, antimicrobial activity, toxicity and industrial applications,” in Nano-antimicrobials: progress and prospects (Springer Berlin Heidelberg, Berlin, Heidelberg), 337–374.
Dutta P., Deb L., Pandey A. K. (2022). Trichoderma-from lab bench to field application: Looking back over 50 years. Front. Agron. 4, 932839. doi: 10.3389/fagro.2022.932839
Feng D., Wang R., Sun X., Liu P., Tang J., Zhang C., et al. (2023). Heavy metal stress in plants: Ways to alleviate with exogenous substances. Sci. Total Environ. 897, 165397. doi: 10.1016/j.scitotenv.2023.165397
Foyer C. H. (2018). Reactive oxygen species, oxidative signaling and the regulation of photosynthesis. Environ. Exp. Bot. 154, 134–142. doi: 10.1016/j.envexpbot.2018.05.003
González-Morales S., Solís-Gaona S., Valdés-Caballero M. V., Juárez-Maldonado A., Loredo-Treviño A., Benavides-Mendoza A. (2021). Transcriptomics of biostimulation of plants under abiotic stress. Front. Genet. 12, 583888. doi: 10.3389/fgene.2021.583888
Gupta R., Singh M., Khan B. R. (2022). Photosynthetic electron transport rate and root dynamics of finger millet in response to Trichoderma harzianum. Plant Signal. Behav. 17, 2146373. doi: 10.1080/15592324.2022.2146373
Gurevitch J., Koricheva J., Nakagawa S., Stewart G. (2018). Meta-analysis and the science of research synthesis. Nat. 555, 175–182. doi: 10.1038/nature25753
Habyarimana E., Lopez-Cruz M., Baloch F. S. (2020). Genomic selection for optimum index with dry biomass yield, dry mass fraction of fresh material, and plant height in biomass sorghum. Genes 11, 61. doi: 10.3390/genes11010061
Haney C. H., Urbach J. M., Ausubel F. M. (2014). Differences and similarities: Innate immunity in plants and animals. Biochem. (Lond) 36, 40–45. doi: 10.1042/BIO03605040
Harman G. E., Doni F., Khadka R. B., Uphoff N. (2021). Endophytic strains of Trichoderma increase plants’ photosynthetic capability. J. Appl. Microbiol. 130, 529–546. doi: 10.1111/jam.v130.2
Harzing A. W. (2007). Publish or Perish. Available online at: from https://harzing.com/resources/publish-or-perish.
Hasanuzzaman M., Bhuyan M. B., Zulfiqar F., Raza A., Mohsin S. M., Mahmud J. A., et al. (2020). Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 9, 681. doi: 10.3390/antiox9080681
Hayat S., Hayat Q., AlYemeni M. N., Wani A. S., Pichtel J., Ahmad A. (2012). Role of proline under changing environments: A review. Plant Signal. Behav. 7, 1456–1466. doi: 10.4161/psb.21949
Hidangmayum A., Dwivedi P. (2018). Plant responses to Trichoderma spp. and their tolerance to abiotic stresses: a review. J. Pharmacogn. Phytochem. 7, 758–766.
Hosseinifard M., Stefaniak S., Ghorbani Javid M., Soltani E., Wojtyla Ł., Garnczarska M. (2022). Contribution of exogenous proline to abiotic stresses tolerance in plants: a review. Int. J. Mol. Sci. 23, 5186. doi: 10.3390/ijms23095186
Kopecká R., Kameniarová M., Černý M., Brzobohatý B., Novák J. (2023). Abiotic stress in crop production. Int. J. Mol. Sci. 24, 6603. doi: 10.3390/ijms24076603
Kumar S. (2020). Abiotic stresses and their effects on plant growth, yield and nutritional quality of agricultural produce. Int. J. Food Sci. Agric. 4, 367–378. doi: 10.26855/ijfsa.2020.12.002
Kumar K., Manigundan K., Amaresan N. (2017). Influence of salt tolerant Trichoderma spp. on growth of maize (Zea mays) under different salinity conditions. J. Basic Microbiol. 57, 141–150. doi: 10.1002/jobm.201600369
Lakhdari W., Benyahia I., Bouhenna M. M., Bendif H., Khelafi H., Bachir H., et al. (2023). Exploration and evaluation of secondary metabolites from Trichoderma harzianum: GC-MS analysis, phytochemical profiling, antifungal and antioxidant activity assessment. Molecules 28, 5025. doi: 10.3390/molecules28135025
Li Z., Chen W., Tan Q., Hou Y., Farooq T. H., Iqbal B., et al. (2023). “Interaction of heavy metal with drought/salinity stress in plants,” in Heavy metal toxicity and tolerance in plants: A biological, omics, and genetic engineering approach, (Hoboken, New Jersey: John Wiley & Sons) 407–423.
Liang J., Lu L., Zhang W., Chi M., Shen M., An C., et al. (2023). Comprehensive characterization and expression analysis of enzymatic antioxidant gene families in passion fruit (Passiflora edulis). Iscience 26 (11), 108329. doi: 10.1016/j.isci.2023.108329
Mehetre S. T., Mukherjee P. K. (2015). “Trichoderma improves nutrient use efficiency in crop plants,” in Nutrient use efficiency: from basics to advances. (New Delhi: Springer), 173–180.
Mermigka G., Amprazi M., Mentzelopoulou A., Amartolou A., Sarris P. F. (2020). Plant and animal innate immunity complexes: fighting different enemies with similar weapons. Trends Plant Sci. 25, 80–91. doi: 10.1016/j.tplants.2019.09.008
Mittler R. (2002). Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 7, 405. doi: 10.1016/S1360-1385(02)02312-9
Morán-Diez M. E., Martinez de Alba A. E., Rubio M. B., Hermosa R., Monte E. (2021). Trichoderma and the plant heritabl priming responses. J. Fungi 7, 318. doi: 10.3390/jof7040318
Muhammad I., Shalmani A., Ali M., Yang Q. H., Ahmad H., Li F. B. (2021). Mechanisms regulating the dynamics of photosynthesis under abiotic stresses. Front. Plant Sci. 11, 615942. doi: 10.3389/fpls.2020.615942
Mukherjee P. K., Mendoza-Mendoza A., Zeilinger S., Horwitz B. A. (2022). Mycoparasitism as a mechanism of Trichoderma-mediated suppression of plant diseases. Fungal Biol. Rev. 39, 15–33. doi: 10.1016/j.fbr.2021.11.004
Nadarajah K. K. (2020). ROS homeostasis in abiotic stress tolerance in plants. Int. J. Mol. Sci. 21, 5208. doi: 10.3390/ijms21155208
Ogbe A. A., Finnie J. F., Van Staden J. (2020). The role of endophytes in secondary metabolites accumulation in medicinal plants under abiotic stress. S. Afr. J. Bot. 134, 126–134. doi: 10.1016/j.sajb.2020.06.023
Oljira A. M., Hussain T., Waghmode T. R., Zhao H., Sun H., Liu X., et al. (2020). Trichoderma enhances net photosynthesis, water use efficiency, and growth of wheat (Triticum aestivum L.) under salt stress. Microorganisms 8, 1565. doi: 10.3390/microorganisms8101565
Pacheco-Trejo J., Aquino-Torres E., Reyes-Santamaría M. I., Islas-Pelcastre M., Pérez-Ríos S. R., Madariaga-Navarrete A., et al. (2022). Plant defensive responses triggered by Trichoderma spp. as tools to face stressful conditions. Horticulturae 8, 1181. doi: 10.3390/horticulturae8121181
Pehlivan N., Yesilyurt A. M., Durmus N., Karaoglu S. A. (2017). Trichoderma lixii ID11D seed biopriming mitigates dose dependent salt toxicity in maize. Acta Physiol. Plant 39, 1–12. doi: 10.1007/s11738-017-2375-z
Pereira A. (2016). Plant abiotic stress challenges from the changing environment. Front. Plant Sci. 7, 1123. doi: 10.3389/fpls.2016.01123
Rawat L., Singh Y., Shukla N., Kumar J. (2013). Salinity tolerant Trichoderma harzianum reinforces NaCl tolerance and reduces population dynamics of Fusarium oxysporum f. sp. ciceri in chickpea (Cicer arietinum L.) under salt stress conditions. . Arch. Phytopathol. Pflanzenschutz 46, 1442–1467. doi: 10.1080/03235408.2013.769316
Raza A., Razzaq A., Mehmood S. S., Zou X., Zhang X., Lv Y., et al. (2019). Impact of climate change on crops adaptation and strategies to tackle its outcome: A review. Plants 8, 34. doi: 10.3390/plants8020034
Razi K., Muneer S. (2021). Drought stress-induced physiological mechanisms, signaling pathways and molecular response of chloroplasts in common vegetabe crops. Crit. Rev. Biotechnol. 41, 669–691. doi: 10.1080/07388551.2021.1874280
Reghmit A. (2023). “Phytohormones and biomolecules produced by trichoderma strains as eco-friendly alternative for stimulation of plant growth,” in New insights into phytohormones (London, UK: IntechOpen).
Rodriguez P. A., Rothballer M., Chowdhury S. P., Nussbaumer T., Gutjahr C., Falter-Braun P. (2019). Systems biology of plant-microbiome interactions. Mol. Plant 12, 804–821. doi: 10.1016/j.molp.2019.05.006
Saba H., Vibhash D., Manisha M., Prashant K. S., Farhan H., Tauseef A. (2012). Trichoderma–a promising plant growth stimulator and biocontrol agent. Mycosphere 3, 524–531. doi: 10.5943/mycosphere/3/4/14
Salwan R., Sharma A., Kaur R., Sharma R., Sharma V. (2022). The riddles of Trichoderma induced plant immunity. BioControl 174, 105037. doi: 10.1016/j.biocontrol.2022.105037
Schmoll M., Dattenböck C., Carreras-Villaseñor N., Mendoza-Mendoza A., Tisch D., Alemán M. I., et al. (2016). The genomes of three uneven siblings: footprints of the lifestyles of three Trichoderma species. Microbiol. Mol. Biol. Rev. 80, 205–327. doi: 10.1128/MMBR.00040-15
Seleiman M. F., Al-Suhaibani N., Ali N., Akmal M., Alotaibi M., Refay Y., et al. (2021). Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 10, 259. doi: 10.3390/plants10020259
Selvakumar G., Panneerselvam P., Ganeshamurthy A. N. (2012). “Bacterial mediated alleviation of abiotic stress in crops,” in Bacteria in agrobiology: stress management. Ed. Maheshwari D. (Springer, Berlin, Heidelberg), 205–224.
Shukla N., Awasthi R. P., Rawat L., Kumar J. (2012). Biochemical and physiological responses of rice (Oryza sativa L.) as influenced by Trichoderma harzianum under drought stress. Plant Physiol. Biochem. 54, 78–88. doi: 10.1016/j.plaphy.2012.02.001
Singh A., Mehta S., Yadav S., Nagar G., Ghosh R., Roy A., et al. (2022). How to cope with the challenges of environmental stresses in the era of global climate change: An update on ROS stave off in plants. Int. J. Mol. Sci. 23, 1995. doi: 10.3390/ijms23041995
Sood M., Kapoor D., Kumar V., Sheteiwy M. S., Ramakrishnan M., Landi M., et al. (2020). Trichoderma: the “secrets. multitalented Biocontrol agent. Plants 9, 762. doi: 10.3390/plants9060762
Vinale F., Sivasithamparam K. (2020). Beneficial effects of Trichoderma secondary metabolites on crops. Phytother. Res. 34, 2835–2842. doi: 10.1002/ptr.v34.11
Wang Y., Frei M. (2011). Stressed food–The impact of abiotic environmental stresses on crop quality. Agric. Ecosyst. Environ. 141, 271–286. doi: 10.1016/j.agee.2011.03.017
Wickham H. (2016). ggplot2: elegant graphics for data analysis (Springer-Verlag New York). Available online at: https://ggplot2.tidyverse.org (Accessed April 11, 2024).
Wiesmann C. L., Wang N. R., Zhang Y., Liu Z., Haney C. H. (2023). Origins of symbiosis: shared mechanisms underlying microbial pathogenesis, commensalism and mutualism of plants and animals. FEMS Microbiol. Rev. 47, fuac048. doi: 10.1093/femsre/fuac048
Woo S. J., Ruoccco M., Vinale F., Nigro M., Marra R., Lombardi N., et al. (2014). Trichoderma-based products and their widespread use in agriculture. Open Mycol. J. 8, 71–126. doi: 10.2174/1874437001408010071
Yang X., Lu M., Wang Y., Wang Y., Liu Z., Chen S. (2021). Response mechanism of plants to drought stress. Horticulturae 7, 50. doi: 10.3390/horticulturae7030050
Yasmeen R., Siddiqui Z. (2017). Physiological responses of crop plants against Trichoderma harzianum in saline environment. Acta Bot. Croat. 76, 154–162. doi: 10.1515/botcro-2016-0054
Yu L., Ma J., Niu Z., Bai X., Lei W., Shao X., et al. (2017). Tissue-specific transcriptome analysis reveals multiple responses to salt stress in Populus euphratica seedlings. Genes 8, 372. doi: 10.3390/genes8120372
Zaidi N. W., Dar M. H., Singh S., Singh U. S. (2014). “Trichoderma species as abiotic stress relievers in plants,” in Biotechnology and biology of trichoderma (Amsterdam: Elsevier), 515–525.
Zandalinas S. I., Fritschi F. B., Mittler R. (2021). Global warming, climate change, and environmental pollution: recipe for a multifactorial stress combination disaster. Trends Plant Sci. 26, 588–599. doi: 10.1016/j.tplants.2021.02.011
Zhang F., Huo Y., Cobb A. B., Luo G., Zhou J., Yang G., et al. (2018). Trichoderma biofertilizer links to altered soil chemistry, altered microbial communities, and improved grassland biomass. Front. Microbiol. 9, 848. doi: 10.3389/fmicb.2018.00848
Keywords: fungal-host interaction, growth and development, physiology and photosynthesis, enzymatic activities, plant metabolites, systematic review
Citation: dos Santos LBPR, Oliveira-Santos N, Novais DPSd, Cruz-Magalhães V and Loguercio LL (2025) Beneficial plants-Trichoderma interactions on host tolerance to abiotic stresses: a meta-analysis. Front. Plant Physiol. 3:1569221. doi: 10.3389/fphgy.2025.1569221
Received: 31 January 2025; Accepted: 03 March 2025;
Published: 20 May 2025.
Edited by:
Fan Yang, Hainan University, ChinaReviewed by:
Simon Zebelo, University of Maryland Eastern Shore, United StatesTihana Marček, Josip Juraj Strossmayer University of Osijek, Croatia
Copyright © 2025 dos Santos, Oliveira-Santos, Novais, Cruz-Magalhães and Loguercio. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lorena Brito Pimentel Rodrigues dos Santos, cGltZW50ZWwxODEyQGdtYWlsLmNvbQ==; Leandro Lopes Loguercio, bGVhbmRyb0B1ZXNjLmJy
†ORCID: Lorena Brito Pimentel Rodrigues dos Santos, orcid.org/0000-0002-8457-1999
Naiane Oliveira-Santos, orcid.org/0000-0001-5138-9042
Diogo Pereira Silva de Novais, orcid.org/0000-0001-9301-509X
Valter Cruz Magalhães, orcid.org/0000-0001-5552-1442
Leandro Lopes Loguercio, orcid.org/0000-0002-2326-2797