 Ryan B. Schmid
Ryan B. Schmid Kelton D. Welch
Kelton D. Welch Jonathan G. Lundgren
Jonathan G. Lundgren- Ecdysis Foundation, Estelline, SD, United States
Introduction: Regenerative pasture management seeks to maintain ranchers' economic viability by maximizing grassland biodiversity and ecosystem functionality to maintain production and reduce input costs through improved ecosystem services. Dung arthropod communities contribute to two important ecosystem services for ranchers: dung degradation and pest control.
Methods: We examined the effect of regenerative pasture management on key beneficial dung-dwelling arthropods (coprophages, predators, and parasitoids) and measured dung fouling of pastures and pest abundance in dung pats (fly pests and parasites). Bioinventories of the adult dung-dwelling arthropod communities were collected from regeneratively (n = 18) and conventionally managed pastures (n = 21) extending from northeastern South Dakota to central North Dakota.
Results: In total, 51,283 arthropod specimens were collected from 596 dung pats. Pats were also sampled for insect pests and cattle parasites: 20% of the pats contained adult fly pests, and 95% were infested with parasites. Regenerative pasture management did not increase the overall dung arthropod abundance or diversity, nor were there consistent effects on key beneficial arthropod groups throughout the grazing season. However, pasture fouling was significantly reduced in regeneratively managed pastures. Regenerative pastures also had low levels of insect pests, comparable to their conventional counterparts.
Discussion: Notably, the majority (76%) of conventional ranchers used parasiticides/insecticides as part of their pest control program, while only 11% of regenerative ranchers utilized these inputs, opting instead for management to achieve similar results. This work demonstrates that regenerative grassland management aimed at grassland functionality can foster ecosystem services that contribute to ranchers' economic viability.
Introduction
Grasslands are a dominant ecosystem of the Northern Plains that provide environmental, social, and economic benefits to the region (Wimberly et al., 2017). Managing this ecosystem for food production and fiber products has provided livelihoods for ranchers since European settlement began in the late 1800s. The conventional management practices typically enacted during this period included continuous and light rotational grazing, owing to the minimal labor requirements and relatively little infrastructure investments (Briske et al., 2011). However, because of natural resource base degradation, these long-practiced grazing systems now jeopardize the economic viability of ranches (Teague and Barnes, 2017; Wang et al., 2018). The main causes of grassland degradation associated with conventional management are overgrazing of localized areas or highly palatable plant species, while other areas and species are left relatively ungrazed, resulting in a buildup of undesirable plants (Briggs et al., 2005; Archer et al., 2017). This has a cascade of consequences for grassland systems, including soil degradation, erosion, an increased bare ground presence, a loss of forage biomass, weed proliferation, and a loss of plant diversity (Milchunas and Lauenroth, 1993; Webber et al., 2010; Teague et al., 2011; Hillenbrand et al., 2019; Apfelbaum et al., 2022). Continuously grazed grasslands' productivity and resilience are reduced as well as the biodiversity that facilitates energy/nutrient flow through the system (Teague and Kreuter, 2020).
Regenerative grassland management, intended to restore natural resources and economic viability of ranches via improved biodiversity and resulting ecosystem services, is an alternative to conventional management (Pecenka and Lundgren, 2019; Fenster et al., 2021; Apfelbaum et al., 2022; Johnson et al., 2022; Mosier et al., 2022; White et al., 2023). Regenerative grassland management has been implemented through various grazing systems, for example, adaptive multi-paddock grazing, that prescribe ecologically based practices meant to stimulate grassland production by restoring ecosystem processes (Teague and Barnes, 2017). Specific practices that define regenerative systems include high animal stock densities, short periods of grazing within paddocks, adequate recovery periods for vegetation following a grazing event, and reducing/eliminating pesticides applied to the herd (Fenster et al., 2021). Together, these grazing practices result in greater amounts of plant vegetation following grazing events, which leads to more rapid regrowth. When followed by adequate recovery periods that account for seasonal variation in herbaceous plant growth, the plant community diversity and forage biomass for livestock also increase (Teague et al., 2011; Apfelbaum et al., 2022).
Dung arthropod communities support plant communities in grassland systems. Specifically, coprophagous species are an important step in the process, whereby plant-based nutrients are recycled to support plant communities (Ridsdill-Smith and Edwards, 2011). Besides helping to facilitate dung degradation, the dung-dwelling arthropod community also provides pest control services for ranchers via predator and parasitoid species (Nichols et al., 2008). The parasiticides commonly used in conventional livestock management systems, for example, avermectin, can disrupt the dung arthropod community's structure through their direct toxicity and insect-repellent properties (Floate, 2007; Webb et al., 2010). Disrupting the community structure can result in a loss of ecosystem services for ranchers, for example, dung degradation and pest suppression (Ridsdill-Smith and Edwards, 2011; Pecenka and Lundgren, 2018). Regeneratively managing grasslands fosters dung arthropod community diversity by minimizing pesticide exposure in dung pats while improving the soil structure and increasing groundcover (Teague et al., 2011; Pecenka and Lundgren, 2019; Apfelbaum et al., 2022). However, whether the dung arthropod community fostered by regenerative grassland management leads to improved ecosystem services for ranchers is unclear. We examined the effects of regenerative pasture management on key beneficial dung-dwelling arthropods (coprophages, predators, and parasitoids) and measured dung pasture fouling rates and pest abundance in dung pats.
Materials and methods
Study sites
Dung-dwelling arthropods were sampled from 39 pastures (managed by 35 operators) that spanned an approximately 80-km-wide area extending from northeastern South Dakota/western-central Minnesota to central North Dakota (Figure 1). Pastures varied in size across our sampled sites from 5 to 445 ha. It should be noted that the pasture borders were defined by each rancher, and their definition of pasture borders affected their reported pasture size, as many of them owned pastures adjacent to the one we sampled that were separated by only a simple fence or a landscape feature. Consequently, we utilized metrics that did not rely on pasture size to describe management, as these techniques could be applied independently of the pasture's size. Stock density, herd rotation frequency, paddock recovery time, and using synthetic chemicals were uniquely applied to each pasture/herd, resulting in a continuum of management that produced a score, which was then used to categorize sites as conventional or regenerative (Table 1; sensu Pecenka and Lundgren, 2019; Fenster et al., 2021; Schmid et al., 2024a). Site stock density (animal units [AUs] per ha of paddock) was categorized as <5 AUs per ha (scored as 0), 5–10 AUs per ha (scored as 1), and >10 AUs per ha (scored as 2). Herd rotation scores were divided based on the animals moving between paddocks >30 d (scored 0), 10–30 d (scored as 1), and <10 d (scored as 2). The rest period was categorized as continuously grazed during the growing season (scored as 0), a rest period of 1–30 d (scored as 1), and a rest period of >30 d (scored as 2). Parasiticide and/or insecticide application was divided into multiple ubiquitous applications to the herd throughout the year, with one occurring proximally before ( ≤ 60 d) or during the grazing season (scored as 0), a single ubiquitous application to the herd not during or before (>60 d) the grazing season (scored as 1), and no parasiticide or insecticide use (scored as 2). Sites with cumulative scores of 5 and above were categorized as regenerative (Table 1). Management systems specific to each pasture had been established for at least 4 years prior to sampling.
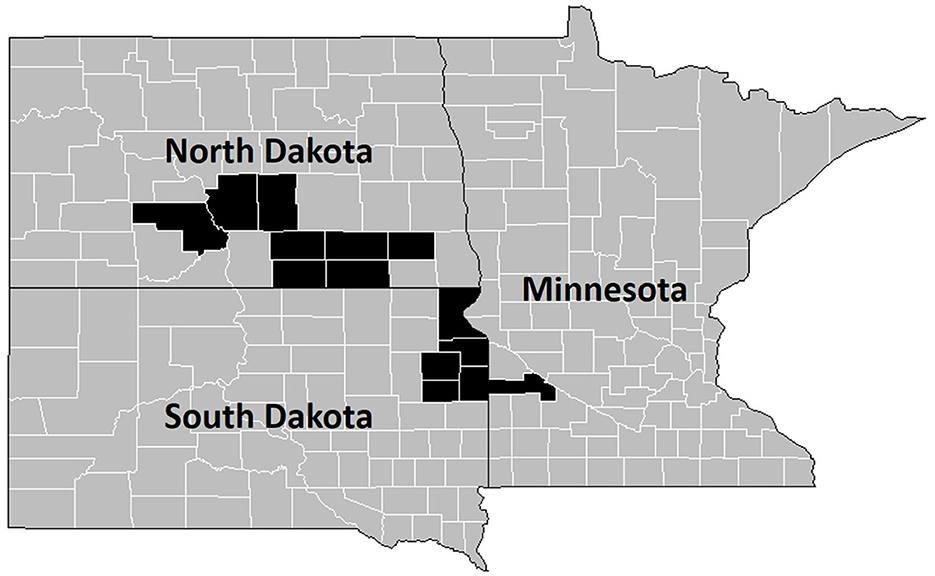
Figure 1. Counties containing sampled pastures (n = 39) for this study are highlighted in black on state county maps. Highlighted counties include Yellow Medicine County, MN; Burleigh County, ND; Dickey County, ND; Kidder County, ND; La Moure County, ND; Logan County, ND; McIntosh County, ND; Morton County, ND; Ransom County, ND; Codington County, SD; Deuel County, SD; Grant County, SD; Hamlin County, SD; and Roberts County, SD.
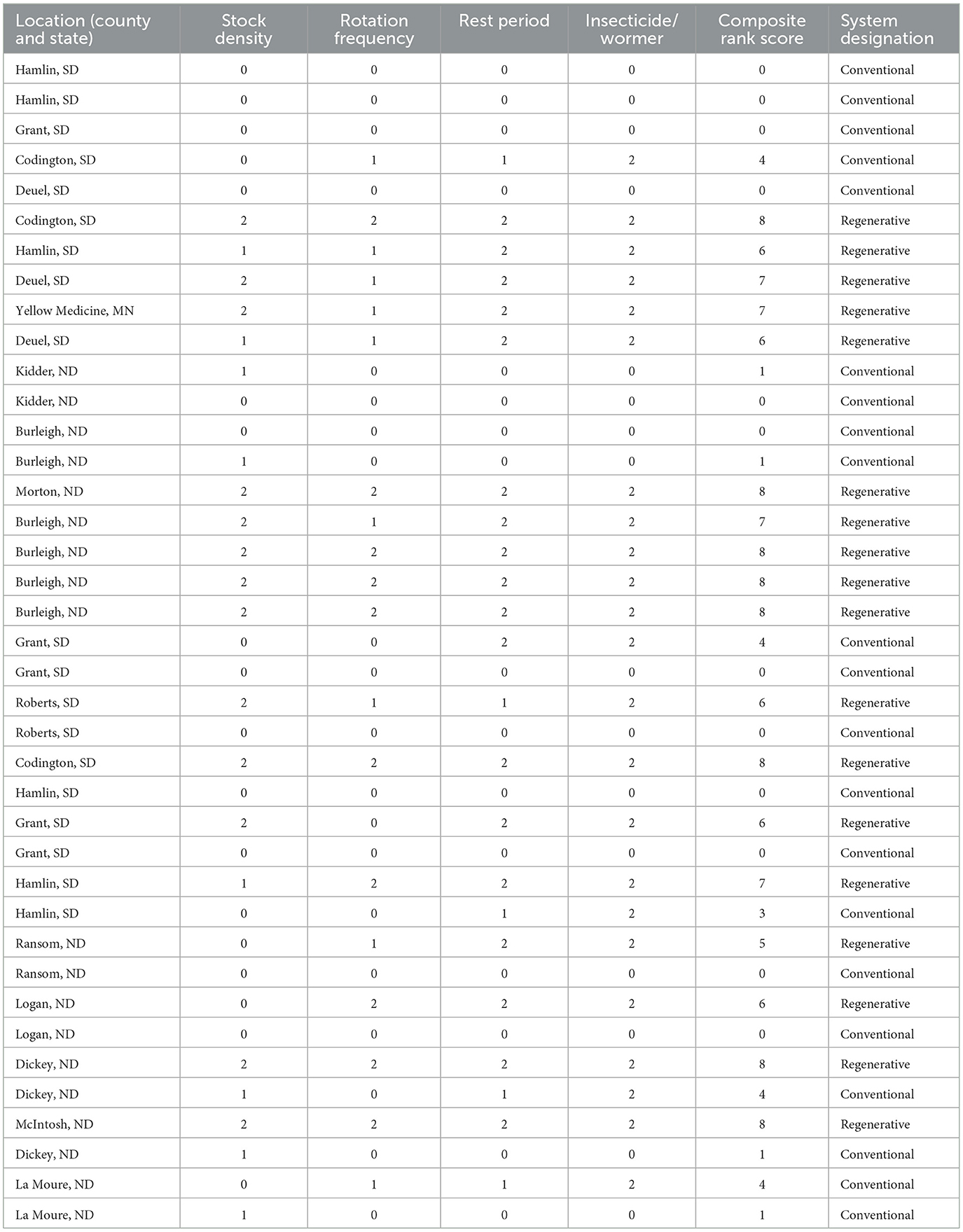
Table 1. Composite rank score and associated ranching system designation of pasture sites sampled for this study (n = 39).
Arthropod sampling procedure
Dung-dwelling arthropods were sampled three times at each site (early June, mid-July, and late August/early September) during both the 2019 (n = 19 sites) and 2020 (n = 20 sites) grazing seasons.
Arthropod samples were gathered by inserting a core cutter (10 cm diameter, 10 cm depth) at the center of a 2–4-d-old dung pat, when dung arthropod abundance is highest in the region (Pecenka and Lundgren, 2018). Dung cores were placed into individual plastic bags and kept on ice until they could be returned to the laboratory for extracting the arthropods. Once in the laboratory, the cores were placed in Berlese funnels for 10 d until completely dry and all arthropods had evacuated into vials filled with 70% isopropyl alcohol. A floatation method, modified from Ladell (1936), was used to further separate the arthropods from the soil and dung particles that fell into the vials during the Berlese process. Vial contents (arthropods and detritus) were separated from the alcohol using a 53-μm sieve before rinsing the contents into a glass beaker with an aqueous Epsom salt (MgSO4) solution of 1.13 specific gravity. The Epsom salt solution was then pumped into the bottom of the beaker to slowly raise the volume to the brim. The beaker was then placed on top of a 53-μm sieve, and the Epsom salt solution was slowly pumped into the bottom of the beaker for 30 s. This procedure permitted the arthropod specimens to float to the top of the solution while much of the soil sediment remained at the bottom. Arthropods were collected in the 53-μm sieve when the Epsom salt solution overflowed the beaker. The Epsom salt solution was pumped into the beaker for another 30 s following a 60-s rest period to ensure all the arthropods were collected from the solution. The arthropods were rinsed into vials using 70% isopropyl alcohol for storage until specimens could be identified.
Arthropod community identification and classification
Multiple resources, including peer-reviewed literature, taxonomic keys, and consultation with taxonomic experts, were used to identify the arthropod specimens to the lowest taxonomic level possible. When these resources were either unavailable or presented an unrealistic time frame, some arthropod groups were only identified to the following respective levels: mites (Arachnida: Acari) and Protura were identified to the class level, thrips (Insecta: Thysanoptera) were identified to the ordinal level, springtails (Hexapoda: Collembola) were identified to the family level, and immature specimens were identified to ordinal or family level. These arthropod groups were excluded from data analysis because their inclusion would cause inaccurate diversity metrics. All specimens not in the aforementioned groups were identified to the genus and species level or assigned a unique morphospecies number. Specimens belonging to arthropod orders of ecological importance within cattle dung (Araneae, Coleoptera, Diptera, and Hymenoptera) were assigned to functional groups according to peer-reviewed literature and current hypotheses of these organisms' ecology. Functional groups assigned to these orders included coprophagous (broken down further into dweller, roller, and tunneller for dung beetles), Diptera, parasitoid, and predator. Voucher specimens are deposited in the Mark F. Longfellow Ecological Reference Collection housed at Blue Dasher Farm (Estelline, South Dakota, USA), and digital photos of vouchers are archived on BugBox (Welch and Lundgren, 2024).
Pasture fouling
Pasture fouling was assessed by counting the number of dung pats present in 1 m of a 50-m transect line (n = 2 transects/pasture). Transect locations were chosen randomly at the beginning of the grazing season and were maintained for all three sample observations. One fouling sample was omitted from the analysis owing to the paddock being grazed the day before sampling at exceptionally high stock density. The area where the dung pats were counted along transect lines was also measured. Both the number and the area of the dung pats were adjusted for herd size using the AUs present in the pasture.
Cattle fecal parasites
The dung pats sampled for arthropods were simultaneously sampled for fecal parasites. A 10-g sample was extracted from the remainder of the same pats the arthropod samples were collected. Fecal samples were kept on ice, but not frozen, in the field and were then stored at 2°C until further processing. Fecal samples were prepared for enumerating parasite eggs and oocysts within 3 d of sampling from pastures using the Wisconsin double centrifugation flotation procedure with saturated sucrose as a flotation medium (Cox and Todd, 1962). Helminth eggs and Coccidia oocyst counts were expressed per gram of sample.
Data analysis
The effects of pasture management on arthropod community characteristics (abundance, species richness, diversity [Shannon H'], dung beetle, fly, parasitoid, predator, and pest fly), pasture fouling (dung pat counts and dung pat area), and parasite abundance (helminth and Coccidia) were compared using a repeated measures analysis of variance (rm-ANOVA). The date sampled within each grazing season was the within-subjects factor, and the grazing treatment and year were the between-subjects factors. All data conformed to the assumptions of rm-ANOVA. The statistical significance was set at α = .05. All data were analyzed using RStudio statistic software (version 4.3.2).
Results
Arthropod community
The results of the dung-dwelling arthropod community survey are described in detail by Schmid et al. (2024a), along with the data of the collected specimens and their abundance in the Supplementary material. In brief, 51,283 arthropod specimens were identified to 787 different morphospecies. Mites, Collembola, Protura, Thysanoptera, and immature specimens were found in the samples but not included in the results due to a lack of taxonomic keys and time to identify them to the morphospecies. The remaining arthropod community was composed of seven classes (Arachnida, Chilopoda, Diplopoda, Diplura, Insecta, Malacostraca, and Symphyla) and 20 orders (Araneae, Coleoptera, Dermaptera, Dicellurata, Diptera, Geophilomorpha, Hemiptera, Hymenoptera, Isopoda, Julida, Lepidoptera, Lithobiomorpha, Neuroptera, Opiliones, Polydesmida, Pseudoscorpiones, Psocodea, Rhabdura, Symphyla, and Trichoptera).
Arthropod community abundance, species richness, and diversity fluctuated significantly between years (Table 2). However, grazing management had no significant effect on the overall arthropod community abundance, species richness, or diversity (Table 2), with the arthropod community metrics tracking closely together between grazing treatments within grazing seasons.
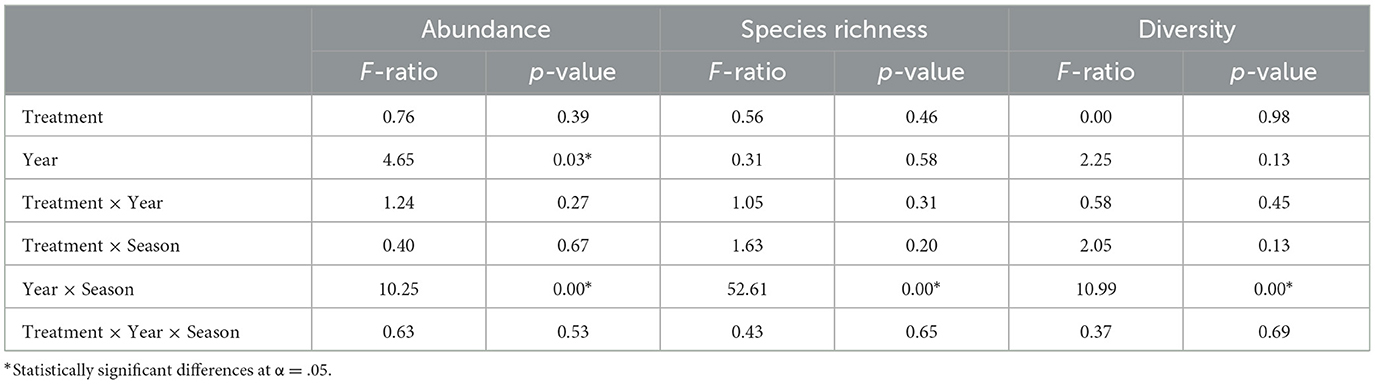
Table 2. Repeated measures analysis of variance comparing dung arthropod community abundance, species richness, and diversity (Shannon H') in regeneratively and conventionally grazed pastures.
While grazing management did not affect the overall arthropod community abundance, it did have a significant effect on dung beetle and predator abundance (Table 3). Conventionally grazed pastures had a higher mean abundance of both dung beetles (17.71 ± 1.19 per pat) and predators (20.35 ± 1.26 per pat) compared to the regenerative pastures (dung beetles: 14.42 ± 1.05 per pat, predators: 16.59 ± 1.13 per pat). Pest abundance did not differ significantly between grazing treatments during any portion of either grazing season (Table 3). However, it bears pointing out that adult fly pest (Haematobia irritans, Musca domestica, and Musca autumnalis) abundance in the regeneratively grazed pastures decreased as the season progressed in both the 2019 and 2020 grazing seasons, while the fly pest abundance in the conventionally grazed pastures increased as the season progressed during both grazing seasons (Figure 2).
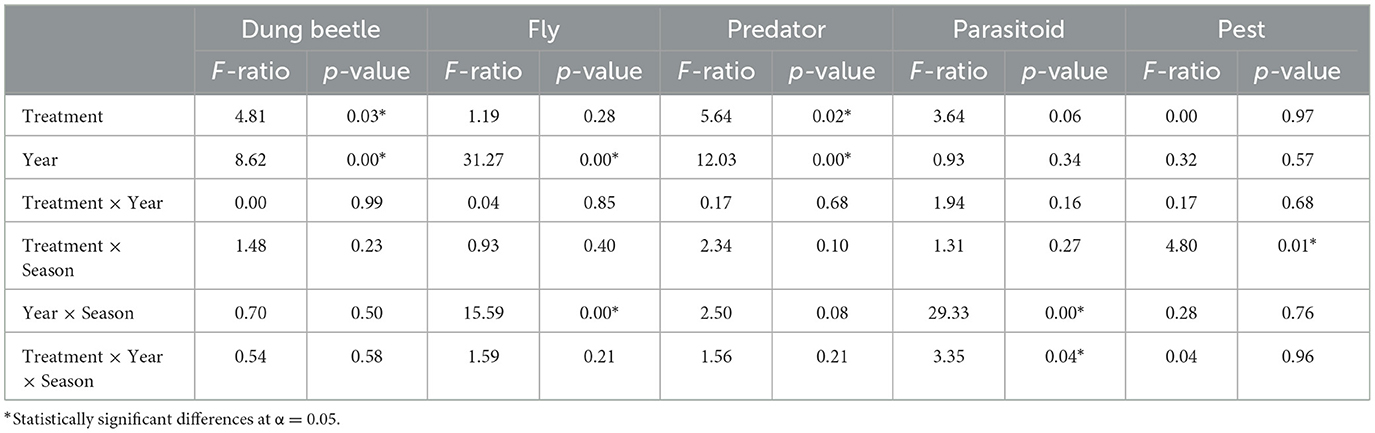
Table 3. Repeated measures analysis of variance comparing dung-dwelling arthropod functional groups, that is, dung beetles, flies, predators, parasitoids, and adult fly pests in regeneratively and conventionally grazed pastures.
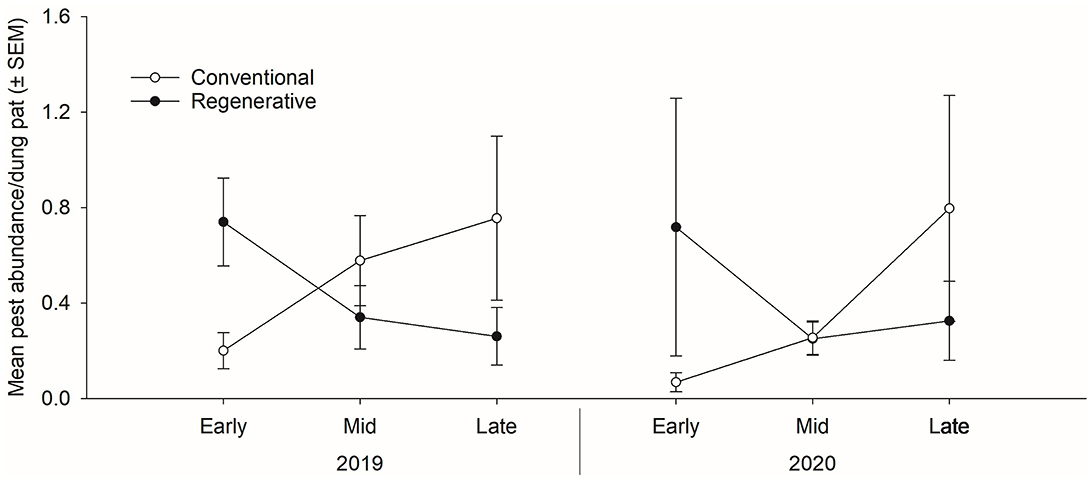
Figure 2. Mean ± standard error of the mean (SEM) adult fly pest abundance per dung pat in regeneratively and conventionally grazed pastures (n = 39) measured in the early (early June), middle (mid-July), and late (late August/early September) portions of the 2019 and 2020 grazing seasons. Statistical analysis was performed using repeated measures analysis of variance, with statistical significance set at α = 0.05.
Pasture fouling
Pasture fouling (both dung pat counts and the area of the pastures covered by dung pats) was consistently higher in conventionally grazed pastures throughout both grazing seasons, significantly so when the grazing season transitioned into the middle and late portions of each year (Table 4 and Figure 3). Furthermore, both metrics of pasture fouling bifurcated between grazing treatments, with regeneratively grazed pasture fouling decreasing as the grazing season progressed, while conventionally grazed pasture fouling increased.
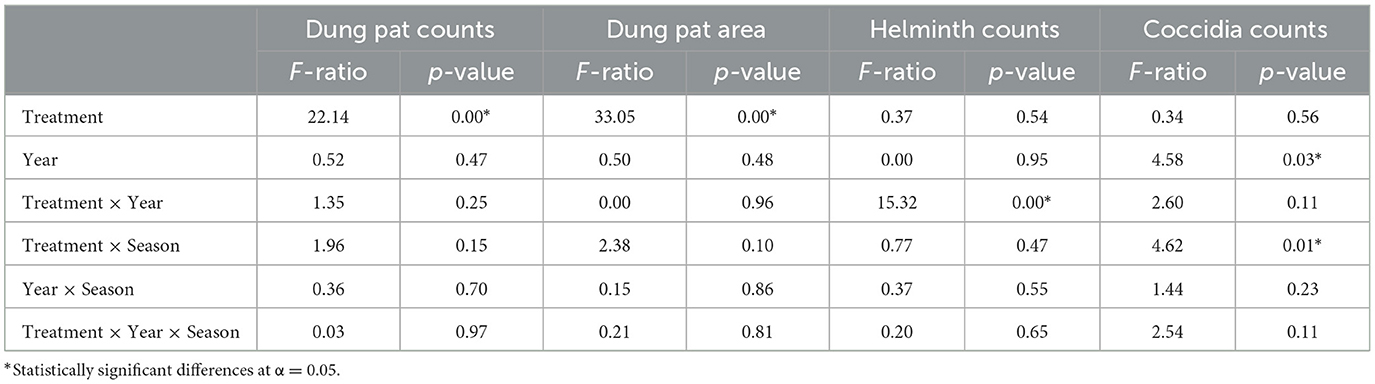
Table 4. Repeated measures analysis of variance comparing pasture fouling (dung pat counts and area) and dung parasite loads (helminth and Coccidia counts) in regeneratively and conventionally grazed pastures.
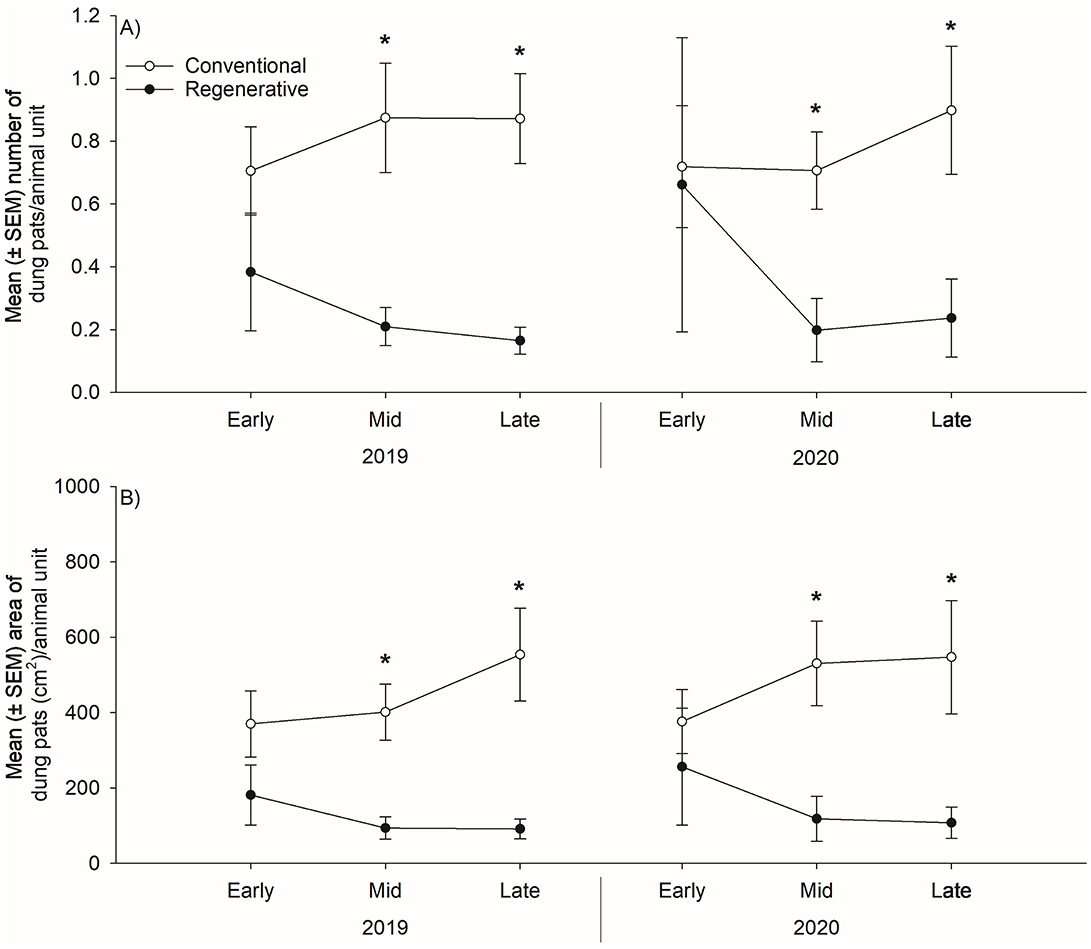
Figure 3. Mean ± standard error of the mean (SEM) number (A) and area (B) of dung pats per animal unit in regeneratively and conventionally grazed pastures (n = 39) measured in the early (early June), middle (mid-July), and late (late August/early September) portions of the 2019 and 2020 grazing seasons. Statistical analysis was performed using repeated measures analysis of variance. * denotes statistical significance at α = 0.05.
Cattle fecal parasites
Throughout the region, helminth and Coccidia parasitism was common in both grazing treatments (Figure 4), with the prevalence of helminth eggs in the dung pats ranging from 80 to 97.5% and the Coccidia oocysts prevalence ranging from 50 to 90%. While both groups of cattle parasites were commonly found in the dung pats, the mean parasite count per gram was relatively low for each grazing treatment. Grazing treatments also showed inconsistent effects on parasite abundance in the dung pats, with the dung pats from regenerative pastures containing significantly higher helminth egg counts in 2019, while conventional pastures had significantly higher egg counts during the 2020 grazing season (Table 4 and Figure 4A). Conventional grazing also resulted in significantly higher Coccidia oocyst counts during the early and middle portions of the 2020 grazing season (Table 4 and Figure 4B).
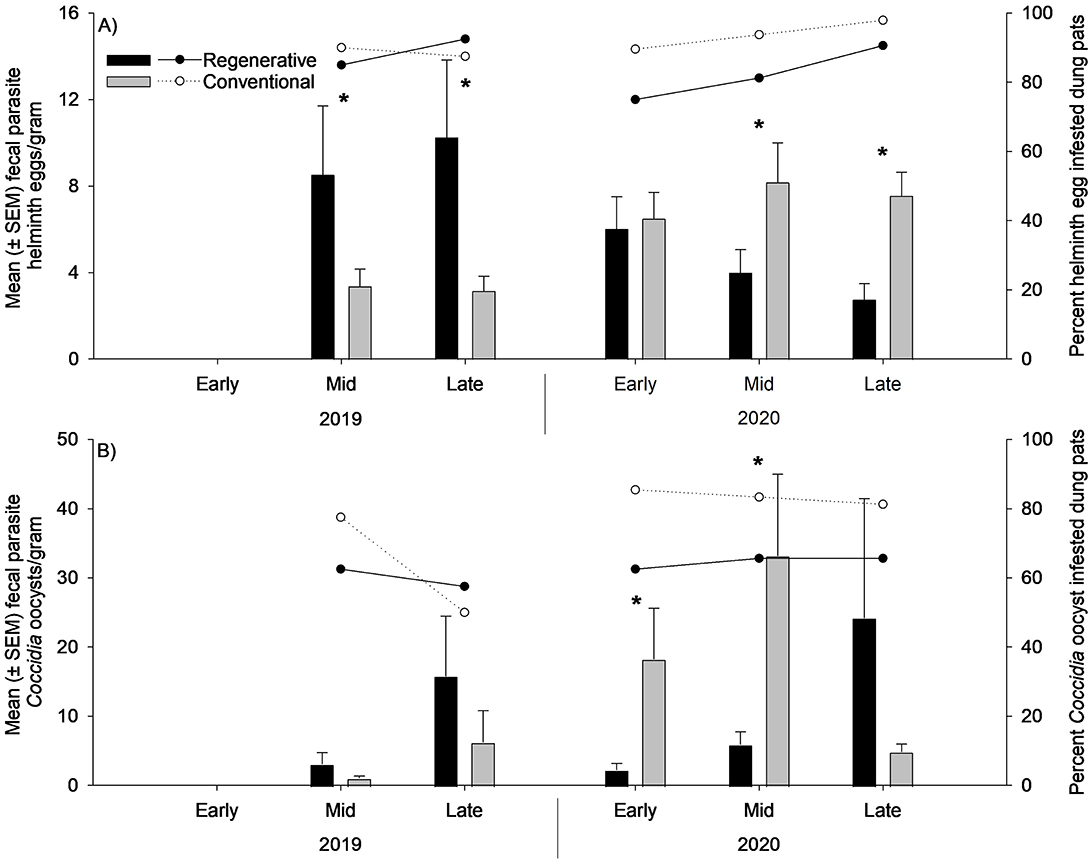
Figure 4. Mean ± standard error of the mean of fecal parasite abundance per gram of helminth eggs (A) and Coccidia oocysts (B) found in dung pats from regeneratively and conventionally grazed pastures (n = 39) sampled during the early (early June), middle (mid-July), and late (late August/early September) portions of the 2019 and 2020 grazing seasons. The percentage of dung pats infested with helminth or Coccidia is represented by the line graphs at the top of each bar graph. Statistical analysis was performed on the mean helminth egg and Coccidia oocyst counts per pat using repeated measures analysis of variance. * denotes statistical significance at α = 0.05.
Discussion
Regenerative grazing management is intended to enhance the ecological function of grassland ecosystems by optimizing grazing disturbances and fostering biodiversity. While studies have shown these practices increase the diversity of several communities in the grassland habitat, for example, soil microbes, plants, and birds (Davis et al., 2020; Wang et al., 2021; Johnson et al., 2022; White et al., 2023), our study found no significant difference between grazing treatments in the dung-dwelling arthropod community (Table 2). In fact, the conventional pastures actually had a higher abundance of dung beetles and predators. However, this effect was largely due to the middle portion of the 2020 grazing season when both dung beetles and predators were significantly higher in the conventional pastures, while the communities remained relatively the same between treatments for the remaining grazing seasons. Despite this, the ecosystem services provided by the dung arthropod community were either better in the regenerative pastures (i.e., reduced pasture fouling) or comparable between the two grazing treatments (i.e., pest fly and parasite abundance). The ecosystem service results would seem to contradict the dung arthropod community results; however, there are potential reasons for this discrepancy, and it comes down to key differences between adult and juvenile dung arthropods.
To understand why pats from the regenerative pastures did not have a higher arthropod abundance or diversity than the conventional pastures, understanding how two traits of adult dung arthropods can affect their abundance in an area is important. First, adults are highly mobile, especially relative to the juvenile stages (Simmons and Ridsdill-Smith, 2011), and second, most parasiticide products are not directly lethal to adult dung arthropods a month after application (Steel and Wardhaugh, 2002; Floate et al., 2005; Beynon, 2012). When these two points are considered in the context of our experiment, it provides an explanation for why there was little to no difference in the arthropod communities between grazing treatments. Even if the parasiticides applied to the herds immediately before or during the grazing season affected the dung arthropod community, the highly mobile adult forms, which were the only form sampled in this study, would have been able to migrate from the surrounding area into our sampled pastures to take advantage of the valuable dung resources. Furthermore, parasiticides (except for slow-release bolus formulations) would have had little to no direct lethal effect on the adult dung arthropods by the second sampling date of each year. Consequently, these results indicate that regenerative grazing will not overcome these two facets of adult dung arthropods to increase the adult community's abundance and diversity.
While the overall arthropod abundance and diversity were comparable between grazing treatments, their functionality within the ecosystem was hindered in the conventional pastures. Ecosystem services in regeneratively grazed pastures were either improved (i.e., reduced pasture fouling) or comparable to (i.e., pest fly and parasite abundance) the conventional grazed pastures (Figures 2–4). If the dung arthropod community was similar between grazing treatments, why was the community better able to provide ecosystem services in the regenerative pastures? Again, the potential reason comes down to differences between adult and juvenile dung arthropods and our excluding juveniles from our sampling procedure. The juvenile form of the most abundant dung-dwelling arthropod guilds, for example, dung-feeding beetles, dung-feeding flies, and mixed-diet flies, consume and degrade far greater quantities of dung pats than the adult form. This is especially true in northern temperate regions like the study area, which contain mostly dweller dung beetles in the population and lack an abundance of the roller and tunneler adult forms that quickly degrade the pats (Hanski, 1991). It is important to note that parasiticides were applied much more to conventional herds immediately before or during the grazing season (Conventional: 76% usage; Regenerative: 11% usage), and 89% of the parasiticide products used were from the avermectin class. Avermectins are especially harmful to juvenile dung beetles, having both direct and indirect effects (Steel and Wardhaugh, 2002; Floate et al., 2005; Jacobs and Scholtz, 2015). Avermectins cause the direct mortality of juveniles at lower concentrations and for longer periods after livestock are treated compared to adult beetles (Steel and Wardhaugh, 2002; Floate et al., 2005). The indirect effects of avermectins on juvenile dung beetles include prolonged larval development and reduced emergence (Ridsdill-Smith, 1988; Lumaret et al., 1993; Kruger and Scholtz, 1997; Cruz Rosales et al., 2012). Adult beetles' fecundity is also inhibited by avermectin exposure (Weaving et al., 2019). In fact, Pérez-Cogollo et al. (2015) showed that exposing Onthophagus landolti adults to an avermectin product in dung led to reduced fecundity and significant declines of adult emergence from brood masses, coupled with slower juvenile development and significantly lower dung removal. This study highlights that the direct and indirect effects of avermectin, which was prevalently used in our study (especially in the conventional sites), have downstream effects on ecosystem services, like dung removal. Pérez-Cogollo et al.'s (2015) results lend credence to our results that showed significantly less pasture fouling in the regenerative pastures that used substantially less parasiticide during or immediately before the grazing season, despite having similar adult dung arthropod community metrics to the conventional pastures. If juveniles were not excluded from our sample processing, it might have added clarity to our understanding of why the regenerative pastures experienced significantly less pasture fouling as each season progressed. In addition to the dung-dwelling arthropod community, regenerative pastures also foster many activities and organisms that aid in decomposing dung pats, for example, more active soil arthropod and microbial communities, insectivorous birds, and trampling by livestock (Wratten and Forbes, 1996; Johnson et al., 2022; McGraw et al., 2024; Schmid et al., 2024b). While we did not measure the juvenile dung arthropod community or the soil microbial community, it is plausible that they are reasons we observed significantly less pasture fouling in the regenerative pastures despite having a similar abundance of dung arthropods per pat.
The second ecosystem service measured by this experiment was pest control. For both parasite abundance and adult fly pests, the regenerative and conventional grazing treatments were similar. However, parasite abundance was influenced by year (Table 4), with Coccidia oocyst abundance being significantly higher in 2020 than in 2019, while helminth egg abundance was significantly higher in the regenerative pastures during 2019 but significantly lower in the regenerative pastures in 2020. The precipitation difference between years is a potential cause of these inconsistent results, with 2019 experiencing well-above-average precipitation while 2020 had below-normal precipitation levels during the grazing season (National Oceanic and Atmospheric Administration, National Oceanic and Atmospheric Administration). Regardless of the differences between years, it is important to note that both parasite and pest abundance were relatively low in both grazing treatments. The question becomes, how did each of these grazing treatments achieve low pest abundance? The relatively high level of parasiticide usage in the conventional pastures, compared to the regenerative pastures, indicates that conventional ranchers were relying on chemical inputs to maintain low pest levels. Alternatively, regenerative ranchers were utilizing management techniques to achieve similar results. The reason the management techniques were able to achieve these results in the regenerative pastures is likely two-fold. First, regenerative management minimizes overgrazing forage, keeping livestock from grazing close to the soil surface, and moves livestock from fouled paddocks, allowing for adequate rest periods before livestock returns (Teague and Kreuter, 2020). This has key benefits for gastrointestinal parasite control because these parasites are typically found on the lower 6–8 in. of plants, and their survival is dramatically reduced when outside their host for extended periods (Waller, 2006; Smith et al., 2009; Hildreth and McKenzie, 2020). Second, a quick breakdown of dung reduces pest fly abundance in pastures (Ridsdill-Smith and Edwards, 2011). The reduced amount of pasture fouling in the regenerative pastures would indicate that regenerative pastures are cycling dung into the soil at a rate that maintains low levels of fly pests. It is also important to note that the management techniques used in the regenerative pastures led to declining fly pests over the grazing season (mean: −0.44 adult fly pests/pat), while fly pests increased in the conventional pastures (mean: +0.64 adult fly pests/pat). This indicates that regenerative management was resilient to fly buildup over the grazing season, while conventional management's reliance on insecticides was potentially breaking down as the grazing season progressed.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
RS: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Writing – original draft, Writing – review & editing. KW: Data curation, Investigation, Methodology, Writing – review & editing. JL: Conceptualization, Funding acquisition, Investigation, Project administration, Resources, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by NCR-SARE, grant number LNC18-410.
Acknowledgments
We thank Tia Busenitz, Amy Heibult, Will Hillery, RaeAnn Krull, Braden Kullman, Kirstyn Loyva, Alex Marra, Alex Michels, Dan Pecenka, Pauline Reby, Nolan Ryne, Liz Schmid (Adee), Jay Skaar, Sierra Stendahl, and Julia Stueven for their help with field collection of specimens and preparations of specimens from dung samples for identification.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Apfelbaum, S. I., Thompson, R., Wang, F., Mosier, S., Teague, R., Byck, P., et al. (2022). Vegetation, water infiltration, and soil carbon response to adaptive multi-paddock and conventional grazing in southeastern USA ranches. J. Environ. Manage. 308:114576. doi: 10.1016/j.jenvman.2022.114576
Archer, S., Andersen, E., Predick, K., Schwinning, S., Steidl, R. J., Woods, S. R., et al. (2017). “Woody plant encroachment: causes and consequences,” in Rangeland Systems: Processes, Management and Challenges. Environmental Management (Springer Open), 25–84. doi: 10.1007/978-3-319-46709-2_2
Beynon, S. A. (2012). Potential environmental consequences of administration of anthelmintics to sheep. Vet. Parasitol. 189, 113–124. doi: 10.1016/j.vetpar.2012.03.040
Briggs, J. M., Knapp, A. K., Blair, J. M., Heisler, J. L., Hoch, G. A., Lett, M. S., et al. (2005). An ecosystem in transition: causes and consequences of the conversion of mesic grassland to shrubland. BioScience. 55:243. doi: 10.1641/0006-3568(2005)055[0243:AEITCA]2.0.CO;2
Briske, D. D., Sayre, N. F., Huntsinger, L., Fernandez-Gimenez, M., Budd, B., Derner, J. D., et al. (2011). Origin, persistence, and resolution of the rotational grazing debate: Integrating human dimensions into rangeland research. Rangeland Ecol. Manage. 64, 325–334. doi: 10.2111/REM-D-10-00084.1
Cox, D., and Todd, A. (1962). Survey of gastrointestinal parasitism in Wisconsin dairy cattle. J. Am. Vet. Med. Assoc. 141, 706–709.
Cruz Rosales, M., González-Hernández, H., and Fajersson, P. (2012). Effect of ivermectin on the survival and fecundity of Euoniticellus intermedius (Coleoptera: Scarabaeidae). Rev. Biol. Trop. 60, 333–345. doi: 10.15517/rbt.v60i1.2765
Davis, K. P., Augustine, D. J., Monroe, A. P., Derner, J. D., and Aldridge, C. L. (2020). Adaptive rangeland management benefits grassland birds utilizing opposing vegetation structure in the shortgrass steppe. Ecol. Appl. 30:e02020. doi: 10.1002/eap.2020
Fenster, T. L. D., LaCanne, C. E., Pecenka, J. R., Schmid, R. B., Bredeson, M. M., Busenitz, K. M., et al. (2021). Defining and validating regenerative farm systems using a composite of ranked agricultural practices. F1000Res. 10:115. doi: 10.12688/f1000research.28450.1
Floate, K. D. (2007). Endectocide residues affect insect attraction to dung from treated cattle: Implications for toxicity tests. Med. Vet. Entomol. 21, 312–322. doi: 10.1111/j.1365-2915.2007.00702.x
Floate, K. D., Wardhaugh, K. G., Boxall, A. B. A., and Sherratt, T. N. (2005). Fecal residues of veterinary parasiticides: nontarget effects in the pasture environment. Annu. Rev. Entomol. 50, 153–179. doi: 10.1146/annurev.ento.50.071803.130341
Hanski, I. (1991). North temperate dung beetles,” in Dung Beetle Ecology (Princeton, NJ: Princeton University Press), p. 75-96. doi: 10.1515/9781400862092.75
Hildreth, M. B., and McKenzie, J. B. (2020). Epidemiology and control of gastrointestinal nematodes of cattle in northern climates. Vet. Clin. North Am. Food Anim. Pract. 36, 59–71. doi: 10.1016/j.cvfa.2019.11.008
Hillenbrand, M., Thompson, R., Wang, F., Apfelbaum, S., and Teague, R. (2019). Impacts of holistic planned grazing with bison compared to continuous grazing with cattle in South Dakota shortgrass prairie. Agric. Ecosyst. Environ. 279, 156–168. doi: 10.1016/j.agee.2019.02.005
Jacobs, C. T., and Scholtz, C. H. (2015). A review on the effect of macrocyclic lactones on dung-dwelling insects: toxicity of macrocyclic lactones to dung beetles. Onderstepoort J Vet Res. 82, 8. doi: 10.4102/ojvr.v82i1.858
Johnson, D. C., Teague, R., Apfelbaum, S., Thompson, R., and Byck, P. (2022). Adaptive multi-paddock grazing management's influence on soil food web community structure for: increasing pasture forage production, soil organic carbon, and reducing soil respiration rates in southeastern USA ranches. PeerJ. 10:e13750. doi: 10.7717/peerj.13750
Kruger, K., and Scholtz, C. (1997). Lethal and sublethal effects of ivermectin on the dung-breeding beetles Euoniticellus intermedius (Reiche) and Onitis alexis Klug (Coleoptera, Scarabaeidae). Agric. Ecosyst. Environ. 61, 123–131. doi: 10.1016/S0167-8809(96)01108-5
Ladell, W. (1936). A new apparatus for separating insects and other arthropods from the soil. Ann. Appl. Biol. 23, 862–879. doi: 10.1111/j.1744-7348.1936.tb06132.x
Lumaret, J. P., Galante, E., Lumbreras, C., Mena, J., Bertrand, M., Bernal, J. L., et al. (1993). Field effects of ivermectin residues on dung beetles. J. Appl. Ecol. 30:428. doi: 10.2307/2404183
McGraw, M. J., Apfelbaum, S. I., Thompson, R., Wang, F., Szuter, M. A., Teague, R., et al. (2024). Breeding bird response to adaptive multi-paddock and continuous grazing practices in Southeastern United States. Ecosphere 15:e70107. doi: 10.1002/ecs2.70107
Milchunas, D. G., and Lauenroth, W. K. (1993). Quantitative effects of grazing on vegetation and soils over a global range of environments. Ecol. Monogr. 63, 327–366. doi: 10.2307/2937150
Mosier, S., Apfelbaum, S., Byck, P., Ippolito, J., and Cotrufo, M. F. (2022). Improvements in soil properties under adaptive multipaddock grazing relative to conventional grazing. Agron. J. 114, 2584–2597. doi: 10.1002/agj2.21135
National Oceanic and Atmospheric Administration. Climate data online. Available online at: https://www.ncei.noaa.gov/cdo-web/ (accessed April 1 2025).
Nichols, E., Spector, S., Louzada, J., Larsen, T., Amezquita, S., Favila, M. E., et al. (2008). Ecological functions and ecosystem services provided by Scarabaeinae dung beetles. Biol. Conserv. 141, 1461–1474. doi: 10.1016/j.biocon.2008.04.011
Pecenka, J. R., and Lundgren, J. G. (2018). The importance of dung beetles and arthropod communities on degradation of cattle dung pats in eastern South Dakota. PeerJ. 6:e5220. doi: 10.7717/peerj.5220
Pecenka, J. R., and Lundgren, J. G. (2019). Effects of herd management and the use of ivermectin on dung arthropod communities in grasslands. Basic Appl. Ecol. 40, 19–29. doi: 10.1016/j.baae.2019.07.006
Pérez-Cogollo, L. C., Rodríguez-Vivas, R. I., Delfín-González, H., et al. (2015). Lethal and sublethal effects of ivermectin on Onthophagus landolti (Coleoptera: Scarabaeidae). Environ. Entomol. 44, 1634–1640. doi: 10.1093/ee/nvv139
Ridsdill-Smith, T. J. (1988). Survival and reproduction of Musca vetustissima Walker (Diptera: Muscidae) and a Scarabaeine dung beetle in dungs of cattle treated with avermectin B1. Aust. J. Entomol. 27, 175–178. doi: 10.1111/j.1440-6055.1988.tb01517.x
Ridsdill-Smith, T. J., and Edwards, P. (2011). “Biological control: ecosystem functions provided by dung beetles,” in Ecology and Evolution of Dung Beetles. 1st Edn (Chichester, UK: Blackwell Publishing), 245–266. doi: 10.1002/9781444342000.ch12
Schmid, R. B., Welch, K. D., and Lundgren, J. G. (2024a). A survey of the dung-dwelling arthropod community in the pastures of the Northern Plains. Insects. 15:38. doi: 10.3390/insects15010038
Schmid, R. B., Welch, K. D., Teague, R., and Lundgren, J. G. (2024b). Adaptive multipaddock (AMP) pasture management increases arthropod community guild diversity without increasing pests. Rangeland Ecol. Manage. 94, 141–148. doi: 10.1016/j.rama.2024.03.001
Simmons, L., and Ridsdill-Smith, T. J. (2011). Ecology and Evolution of Dung Beetles. Oxford, UK: Blackwell Publishing. doi: 10.1002/9781444342000
Smith, L. A., Marion, G., Swain, D. L., White, P. C. L., and Hutchings, M. R. (2009). The effect of grazing management on livestock exposure to parasites via the faecal-oral route. Prevent. Vet. Med. 91, 95–106. doi: 10.1016/j.prevetmed.2009.05.026
Steel, J., and Wardhaugh, K. (2002). Ecological impact of macrocyclic lactones on dung fauna. in Macrocyclic Lactones in Antiparasitic Therapy (Wallingford, U.K.: CABI Publishing), 141–162. doi: 10.1079/9780851996172.0141
Teague, R., and Barnes, M. (2017). Grazing management that regenerates ecosystem function and grazingland livelihoods. Afr. J. Range Forage Sci. 34, 77–86. doi: 10.2989/10220119.2017.1334706
Teague, R., and Kreuter, U. (2020). Managing grazing to restore soil health, ecosystem function, and ecosystem services. Front. Sustain. Food Syst. 4:534187. doi: 10.3389/fsufs.2020.534187
Teague, W. R., Dowhower, S. L., Baker, S. A., Haile, N., DeLaune, P. B., Conover, D. M., et al. (2011). Grazing management impacts on vegetation, soil biota and soil chemical, physical and hydrological properties in tall grass prairie. Agric. Ecosyst. Environ. 141, 310–322. doi: 10.1016/j.agee.2011.03.009
Waller, P. J. (2006). Sustainable nematode parasite control strategies for ruminant livestock by grazing management and biological control. Anim. Feed Sci. Technol. 126, 277–289. doi: 10.1016/j.anifeedsci.2005.08.007
Wang, F., Apfelbaum, S. I., Thompson, R. L., Teague, R., and Byck, P. (2021). Effects of adaptive multiple paddock and continuous grazing on fine-scale spatial patterns of vegetation species and biomass in commercial ranches. Landscape Ecol. 36, 2725–2741. doi: 10.1007/s10980-021-01273-z
Wang, T., Richard Teague, W., Park, S. C., and Bevers, S. (2018). Evaluating long-term economic and ecological consequences of continuous and multi-paddock grazing - A modeling approach. Agric. Syst. 165, 197–207. doi: 10.1016/j.agsy.2018.06.012
Weaving, H., Sands, B., and Wall, R. (2019). Reproductive sublethal effects of macrocyclic lactones and synthetic pyrethroids on the dung beetle Onthophagus similis. Bull. Entomol. Res. 110, 195–200. doi: 10.1017/S0007485319000567
Webb, L., Beaumont, D. J., Nager, R. G., and McCracken, D. I. (2010). Field-scale dispersal of Aphodius dung beetles (Coleoptera: Scarabaeidae) in response to avermectin treatments on pastured cattle. Bull. Entomol. Res. 100, 175–183. doi: 10.1017/S0007485309006981
Webber, D. F., Mickelson, S. K., Ahmed, S. I., Russell, J. R., Powers, W. J., Schultz, R. C., et al. (2010). Livestock grazing and vegetative filter strip buffer effects on runoff sediment, nitrate, and phosphorus losses. J. Soil Water Conserv. 65, 34–41. doi: 10.2489/jswc.65.1.34
Welch, K. D., and Lundgren, J. G. (2024). Introducing bugbox: a platform for AI-assisted bioinventories of arthropods. Am. Entomol. 70, 31–33. doi: 10.1093/ae/tmae047
White, L. J., Yeater, K. M., and Lehman, R. M. (2023). Soil microorganisms respond distinctively to adaptive multi-paddock and conventional grazing in the southeastern United States. Soil Sci. Soc. Am. J. 87, 1096–1108. doi: 10.1002/saj2.20573
Wimberly, M. C., Janssen, L. L., Hennessy, D. A., Luri, M., Chowdhury, N. M., Feng, H., et al. (2017). Cropland expansion and grassland loss in the eastern Dakotas: new insights from a farm-level survey. Land Use Policy. 63, 160–173. doi: 10.1016/j.landusepol.2017.01.026
Keywords: grassland, ranching, ecosystem services, insects, adaptive multi-paddock grazing
Citation: Schmid RB, Welch KD and Lundgren JG (2025) Regenerative management increases the efficacy of dung arthropod communities. Front. Sustain. Resour. Manag. 4:1523963. doi: 10.3389/fsrma.2025.1523963
Received: 06 November 2024; Accepted: 02 April 2025;
Published: 24 April 2025.
Edited by:
Johann G. Zaller, University of Natural Resources and Life Sciences Vienna, AustriaReviewed by:
Shujian Gao, Ocean University of China, ChinaFred Provenza, Utah State University, United States
Copyright © 2025 Schmid, Welch and Lundgren. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ryan B. Schmid, cnlhbi5zY2htaWRAZWNkeXNpcy5iaW8=