 Dorys T. Chirinos1*
Dorys T. Chirinos1* Rossana Castro2
Rossana Castro2 Fernando David Sánchez-Mora1
Fernando David Sánchez-Mora1 Jessenia Castro-Olaya1
Jessenia Castro-Olaya1 Christina Mero Peñarrieta1Patricia Morán3
Christina Mero Peñarrieta1Patricia Morán3 Luz C. García Cruzatty1
Luz C. García Cruzatty1- 1Facultad de Ingenierías Agroambientales, Universidad Técnica de Manabí, Portoviejo, Provincia de Manabí, Ecuador
- 2Facultad de Ciencias Agrarias, Universidad Agraria del Ecuador, Guayaquil, Provincia de Guayas, Ecuador
- 3Facultad de Posgrado, Universidad Técnica de Manabí, Provincia de Manabí, Ecuador
Cacao (Theobroma cacao L.) is a tropical crop in high global demand. Ecuador is a significant producer and exporter of cacao where, there are several fine aroma cacao cultivars (type Nacional) and CCN51 (Trinitarian origin) in which the damages caused by pests in those genetic materials have so far been little evaluated. The objective of this research was to estimate the populations of sap pests and the damage caused to the pods in some genetic materials of cacao. This research was conducted during April 2022 - March 2023 in a 2160 m2 lot, planted in random blocks with four replicates and six families resulting from crosses: F1: L26-H64xL11-H19, F2: L26-H64xL21, F3: CCN-51xCCN-51, F4: L26-H64xCCN-51, F5: L21-H38xEET-103 y F6: L21-H38xL21-H38. Populations of Toxoptera aurantii, Planococcus sp., and Selenothrips rubrocintus were evaluated during the dry and rainy seasons. Damage to the pods was measured from S. rubrocintus and from Hemiptera species. Family F1 (National type) was the least infested by sap pests and showed moderate damage to the pods, while family F6 (National type) was the most infested. The dry season favored aphids, mealybugs and thrips, and damaged pods by S. rubrocintus. In the rainy season, the damaged pods from hemipterans was greater. To our knowledge, Leptoglossus zonatus and Guayaquilla gracillicornis are reported for the first time causing damage to cacao. Since one family of National type (F1) was less attacked by pests coupled with high yields, this fine aromatic cacao could represent a promising genetic material for the production of Ecuadorian cacao.
1 Introduction
Cacao (Theobroma cacao L.) (Malvales: Malvaceaae) is a tropical crop of high global demand whose grain production is destined for the chocolate industry and other products (Armengot et al., 2020). Originating from the humid neotropical forests of South America, its domestication has been controversial, attributed both to the native area and to Central America (Díaz-Valderrama et al, 2020). However, it has recently been demonstrated that the domestication of cacao dates back 5000 years in the native Amazon based on archaeogenomics and biochemistry analyses of ceramic residues taken from pre-Columbian cultures of South America and Central America (Lanaud et al., 2024).
In Ecuador, cacao is planted in 21 of the 24 provinces, being one of the main producing and exporting countries of cacao beans, with approximately 626,962 ha planted (Ministerio de Agricultura y Ganadería (MAG), 2025). It is cultivated 77% on the Pacific coast, 13% in the highlands (Andean region), and 10% in the Amazon region, where Manabí is the coastal province with the largest area at 117,080 ha (Ministerio de Agricultura y Ganadería (MAG), 2025). Various phytosanitary problems can affect cacao production, including diseases such as Monilia for which the causal agent is Moniliophthora roreri ((Cif.) H.C. Evans, Stalpers, Samson & Benny), witches broom caused by Moniliophthora perniciosa (Stahel) Aime & Phillips-Mora) (Marasmiaceae) and machete disease caused by Ceratocystis cacaofunesta Engelbr. (Ceratocystidaceae), the latter associated with the scolytine, Xyleborus ferrugineus (Fabricius) (Coleoptera: Curculionidae: Scolytinae) (Tarqui et al., 2017; Paladines-Rezabala et al., 2022).
There are insect pests attacking leaves, flowers, and pods such as the black citrus aphid, Toxoptera aurantii Boyer de Fonscolombe (Hemiptera: Aphididae), the redbanded thrips, Selenothrips rubrocintus (Giard) (Thysanoptera: Thripidae), pseudococcids (Hemiptera: Pseudococcidae), as well as the cocoa bug, Monalonion dissimulatum Distant (Hemiptera: Miridae) whose population densities could decrease cacao yield (Cañarte-Bermúdez and Navarrete-Cedeño, 2021; Morán et al., 2025). Furthermore, 14 taxa of scolytines associated with cacao have recently been reported both in monoculture and in agroforestry systems (Castro-Olaya et al, 2024).
Toxoptera aurantii develops its colonies on the underside of young leaves, flowers, and newly formed pods (Delgado et al., 2023). By extracting sap, leaves and flowers can deform, while small pods fall off or grow with difficulty. Additionally, the honeydew it excretes promotes the growth of sooty mold (Delgado et al., 2023). For its part, S. rubrocintus damages both young leaves and pods, and when feeding on leaves, it leaves black spots as a result of excrement falling on the surface (Denmark and Wolfenbarger, 2010). When feeding on pods, the sap rises to the surface, where it oxidizes upon contact with air, causing brown, corky-looking wounds (Cañarte-Bermúdez and Navarrete-Cedeño, 2021). These latter damages are referred to as aesthetic damages but make it difficult to evaluate the maturation of the pods (Walter et al., 2018). Pseudococcids extract sap from different plant organs, and some species inject toxins that cause deformities, with the most significant damage being virus transmission ( (Puig et al., 2021). Monalonion dissimulatum causes circular lesions on the pod, which also serve as an entry point for fungi and other pathogens (Cañarte-Bermúdez and Navarrete-Cedeño, 2021).
Given the importance of diseases and the need for increased productivity to meet national and international demand, genetic improvement programs are focused on resistance to relevant diseases (Tarqui et al., 2017), as well as achieving high yields (Sotomayor et al., 2017). So far, in Ecuador, there has been no genetic improvement for resistance to insect pests. Most studies conducted on pests are related to the existing entomofauna without reliable data on the levels that pests reach (Wright, 1984; Páliz et al, 1982; Cañarte-Bermúdez and Navarrete-Cedeño, 2021). There is a lack of studies comparing population densities and damage caused by pests in existing genotypes. In fact, research on the cacao genotype - herbivore interaction is scarce, outdated, and carried out on other continents (Campbell, 1990; Dibog et al., 2008) or the studies have been conducted in other countries in America (Cubillos, 2013; Alomía et al, 2021; Porcel et al., 2024).
Probably this is related to the fact that in the Neotropics pests have had more control, since being the center of origin of the crop, the phytophagous are consumed by their natural enemies which act as biological control agents, which is a consequence of the high biodiversity existing on this plant species in the region (Tscharntke et al., 2023). But in the Neotropics, pests have begun to increase their populations and damage (Fachin et al., 2024; Alomía et al, 2021; Delgado and Couturier, 2017) due to various factors, among which changes in climate, genetic improvement focused on high productivity, and monoculture planting stand out (Cilas and Bastide, 2020; Tscharntke et al., 2023). One of the strategies considered in integrated pest management programs consists of the plant s resistance to pest attacks. Throughout co-evolution, plants have developed defenses to protect themselves against herbivore attacks (Maron et al, 2019). Given the variety of cacao genotypes cultivated in Ecuador, it is important to estimate the extent of population densities and damage caused by pests.
In Ecuador, there are ancestral cultivars (native and non-native) and modern commercial cacao, among these, several types of national cacao that result in the production of fine chocolate of excellent quality and flavor (Jaimez et al., 2022; Thomas et al, 2024). So far, no research has been conducted that quantifies the population densities and the damage caused by insect pests in the cacao cultivars planted in Ecuador. The present research aimed to estimate the population densities of three sap-sucking pests and the damage caused to the pods in families of intraspecific hybrid cacao of National and Trinitarian origin. This study is unprecedented in the country, an important producer and exporter of cocoa in the world, which gives relevance to the present research.
2 Materials and methods
2.1 Location and experimental plot
During the period April 2022 - March 2023, this research was conducted on a 2160 m2 plot of three-year-old cacao located on the experimental campus of “La Teodomira”, Lodana, Manabí, Province, coordinates 01°09 S latitude and 80°21 W longitude, at an altitude of 60 m.a.s.l. The topography is flat, the soils are clay loam, with temperature ranges of 26-28.3°C, average precipitation of 1000 mm, and relative humidity fluctuating from 78 to 87%.
Lodana is included in the Santa Ana canton, which is characterized as an agricultural region, where temporary and perennial crops are grown, predominantly cacao, sugarcane (Saccharum officinarum L.) (Poaceae), broad beans (Vicia faba L.) (Fabaceae), beans (Phaseolus vulgaris L.) (Fabaceae), corn (Zea mays L.) (Poaceae), and bananas (Musa paradisiaca L.) (Musaceae) (Santa Ana, 2015). The life zone is classified as tropical dry forest (Holdridge, 1967) and has two well-defined climatic seasons, a rainy season from December to May and a dry season from June to November (Lopez et al., 2021). Precipitation data have been obtained from the meteorological station of the National Institute of Meteorology and Hydrology (INAMHI), located in “La Teodomira” (Supplementary Figure 1).
The plot contains six families of cacao intraspecific hybrids (Supplementary Figure 2), which were obtained through controlled crosses using genotypes obtained from the Tenguel-CCAT germplasm bank (various L-H), the EET-103 from the Tropical Experimental Station of Pichilingue (Ecuador), and the CCN51 genotype, selected for its high yield and disease tolerance (Sánchez-Mora et al., 2015; Jaimez et al., 2022; Thomas et al, 2024). In those coded with numbers preceded by the letters L and H, the letter L represents the line and the H, the row or the position of the plant within the collection (Sánchez-Mora et al., 2015). The families of cacao intra-specific hybrids and their respective origins are shown in Table 1.
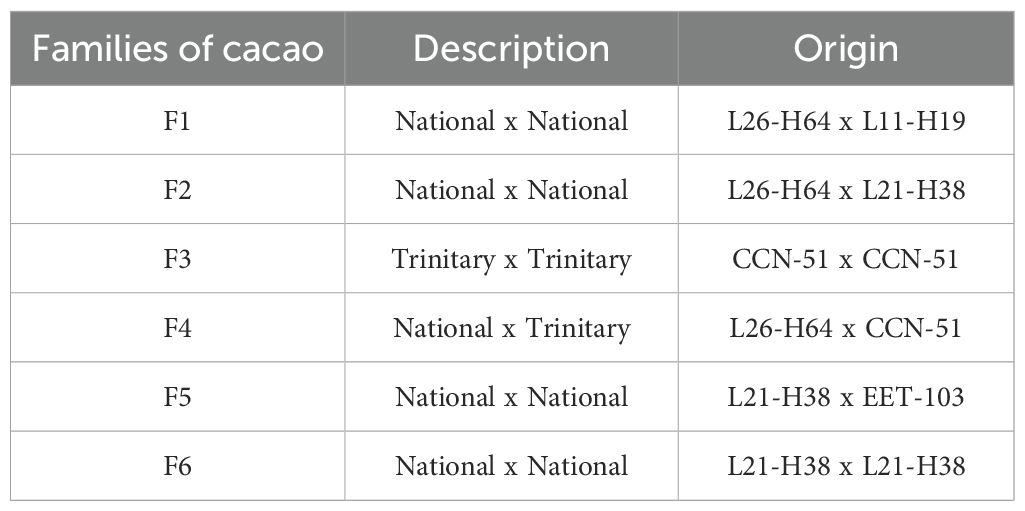
Table 1. Coding and origin of the different cacao materials evaluated.
The experimental lot is designed in randomized blocks with four repetitions and the six families of cacao intraspecific hybrids mentioned above; in each block, there are 10 individuals (tree) per family (90 m2), planted at a distance of 3 m x 3 m. The experimental lot included 60 trees per block covering an area of 540 m2 on a lot measuring 2160 m2 (30 m x 72 m; width x length) planted at a density of 1,111 plants per ha.
During the three years that the experimental plot has been planted, no chemical pesticide sprays have been applied. Weed control is carried out four times a year using a brush cutter. Fertilization is also carried out four times a year using an agrochemical containing N (12%), NO3 (5%), NH4 (7%), P2O5 (11%), K2O (18%), MgO (2.7%), S (8%), B (0.015%), Fe (0.2%), Mn (0.02%) and Zn (0.02%) at a dose of 300 g per plant per year. Training and sanitary pruning are carried out three months before the start of the rainy season. In addition, suckers (basal shoots from the graft rootstock) are removed monthly. The plot has drip irrigation that supplies 1 L per plant twice a week.
2.2 Samplings
2.2.1 Leaves and flowers
Every fifteen days, four plants per family were selected by block, and three young leaves (top of plants) collected from each, totaling 288 leaves in each sampling. Young leaves (top of the plant) were selected because they develop populations of aphids, thrips, and mealybugs, which are pests present in cacao crops in the area (Cañarte-Bermúdez and Navarrete-Cedeño, 2021).
On each tree, five flowers were also taken. Leaves and flowers were placed separately in airtight plastic bags for transport to the laboratory. There, they were observed under a Motic® SMZ 168 (Hong Kong, China) stereoscope with a magnification of 7.5-50X to quantify the number of aphids and thrips on leaves and flowers, as well as mealybugs on leaves.
Individuals from all taxa were counted directly on the organs, except for S. rubrocintus in the flowers, which were gently shaken against a sheet of white bond paper so that the present thrips would fall onto it. They were then collected with a fine natural bristle brush (No. 0) and placed in a Petri dish containing ethyl alcohol (70%) for counting. Insect pest populations were estimated per leaf, and for flowers, they were estimated for the five flowers sample per plant and three leaves per plant respectively.
2.2.2 Pods
2.2.2.1 Damage by thrips
To estimate the pods damaged by S. rubrocintus of the ten trees per block, four were selected and for each, all pods were counted. Subsequently, the pods showing thrips damage (corky epicarp) were counted on the same tree (non-destructive sampling). Thus, the percentage of damage by S. rubrocintus was estimated with the following formula:
Subsequently, the percentage of the area of the pod affected by thrips was determined using a visual scale, assigning it from 0 to 100% depending on the extent of the damage in the epicarp.
2.2.2.2 Damage by Hemiptera
To assess the damage caused by Hemiptera on the pods, in the lot, the total number of pods was counted on four plants per intraspecific hybrid family per block, as well as the pods damaged with punctures, proceeding to estimate the percentage of damaged pods similarly to the damage caused by thrips. Next, a damage scale was applied to the affected pods, which was based on the number of punctures observed in each one (Table 2), using the methodology described by Vargas et al (2005).
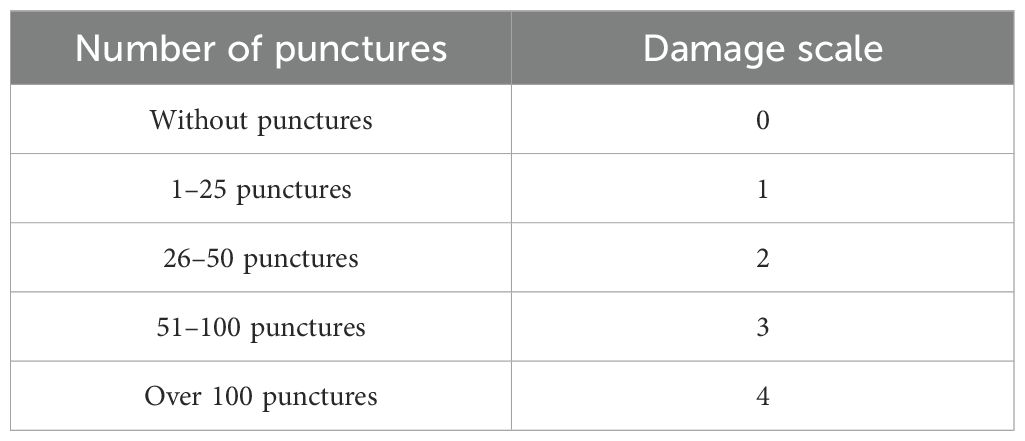
Table 2. Damage scale for affected pods by Hemiptera.
Since the hemipteran species observed damaging the pods are not the ones commonly reported as associated with cacao, the damage caused by them was documented photographically, and the species were identified using keys (Dietrich and Deitz, 1991; Brailovsky, 2014; Flórez-V et al, 2015; Collantes et al., 2016).
2.3 Data analysis
The variables were analyzed using an ANOVA in a randomized block design with four repetitions and six families of cacao intraspecific hybrids. The Shapiro-Wilk normality test was performed on the variables (p<0.05). Since the variables (numbers, percentages) were not normal, transformation was applied using the square root function (√X+1). Means were compared using Fisher s LSD test (p<0.05). A Spearman correlation analysis was performed between the different studied variables versus precipitation (p<0.05). The Chi-Square test was applied to the damage scales of hemipterans on the pods to estimate if there is an association between the scales and the families of cacao intraspecific hybrids (p<0.05). Furthermore, to associate the population densities of the pests and the damage with the families of cacao intraspecific hybrids, a principal component analysis was performed. To obtain the principal components, a standardization of the data was executed (mean 0 and standard deviation 1) to prevent the variables with higher variance from dominating the others. These analyses were carried out using software R Development Core ((R Core Team, 2022).
3 Results
3.1 Leaves and flowers
3.1.1 The black citrus aphid, Toxoptera aurantii
On the leaves, the population densities were high with ranges from 0 to 120 aphids, compared to those developed on the flowers where the populations did not exceed 13 aphids (Figure 1). The mean population of T. aurantii on cacao leaves was significantly different between cacao families (F = 3.93, df = 23, p-value = 0.01) (Supplementary Table 1). According to the Fisher LSD test, the families F4, F5, and F6 presented significantly higher T. aurantii population, while families F1, F2 and F3 presented significantly lower aphids population on leaves (Figure 1). The population densities of T. aurantii in flowers also showed significant differences according to the cacao family (F = 13.98, df = 23, p-value = 0.0001) (Supplementary Table 2). Fisher s LSD test revealed that families F4 and F5 had a significantly higher T. aurantii population in flowers compared to those detected in family F1 (Figure 1).
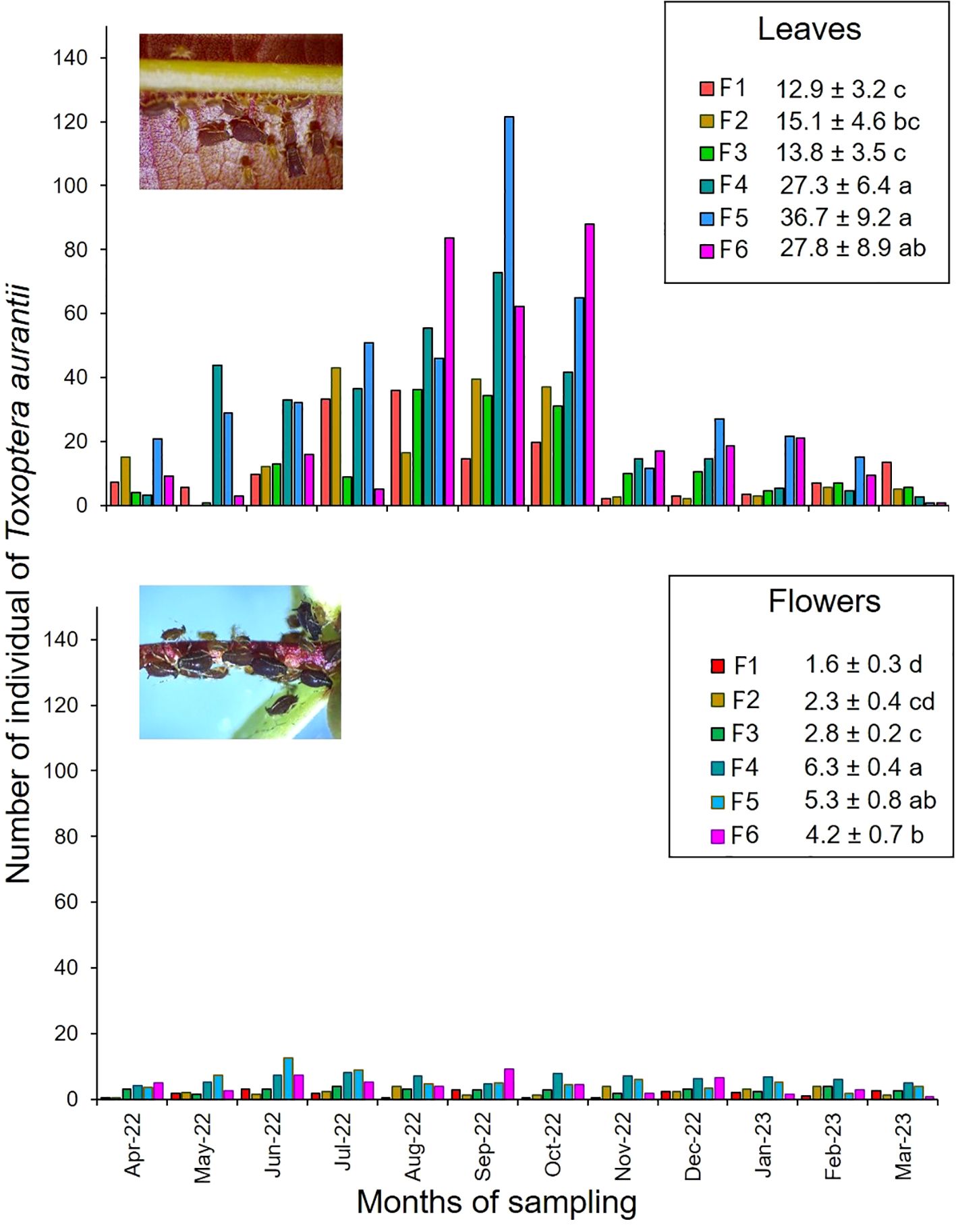
Figure 1. Number of individuals of Toxoptera aurantii observed on leaves and flowers on families of cacao intraspecific hybrids. Period April 2022 - March 2023. Mean ± standard error. Mean followed by different letters in each column is significantly different using the Fisher LSD test (p<0.05).
The highest population levels of T. aurantii were observed in the dry season, increasing from May and reaching their peak in September (Figure 1). These densities decreased from November, coinciding with the start of the rainy season (Supplementary Figure 1). On the leaves, a highly significant negative association (r = -0.58; p<0.01) between precipitation and the abundance of T. aurantii was detected, but not on the flowers (Figure 2).
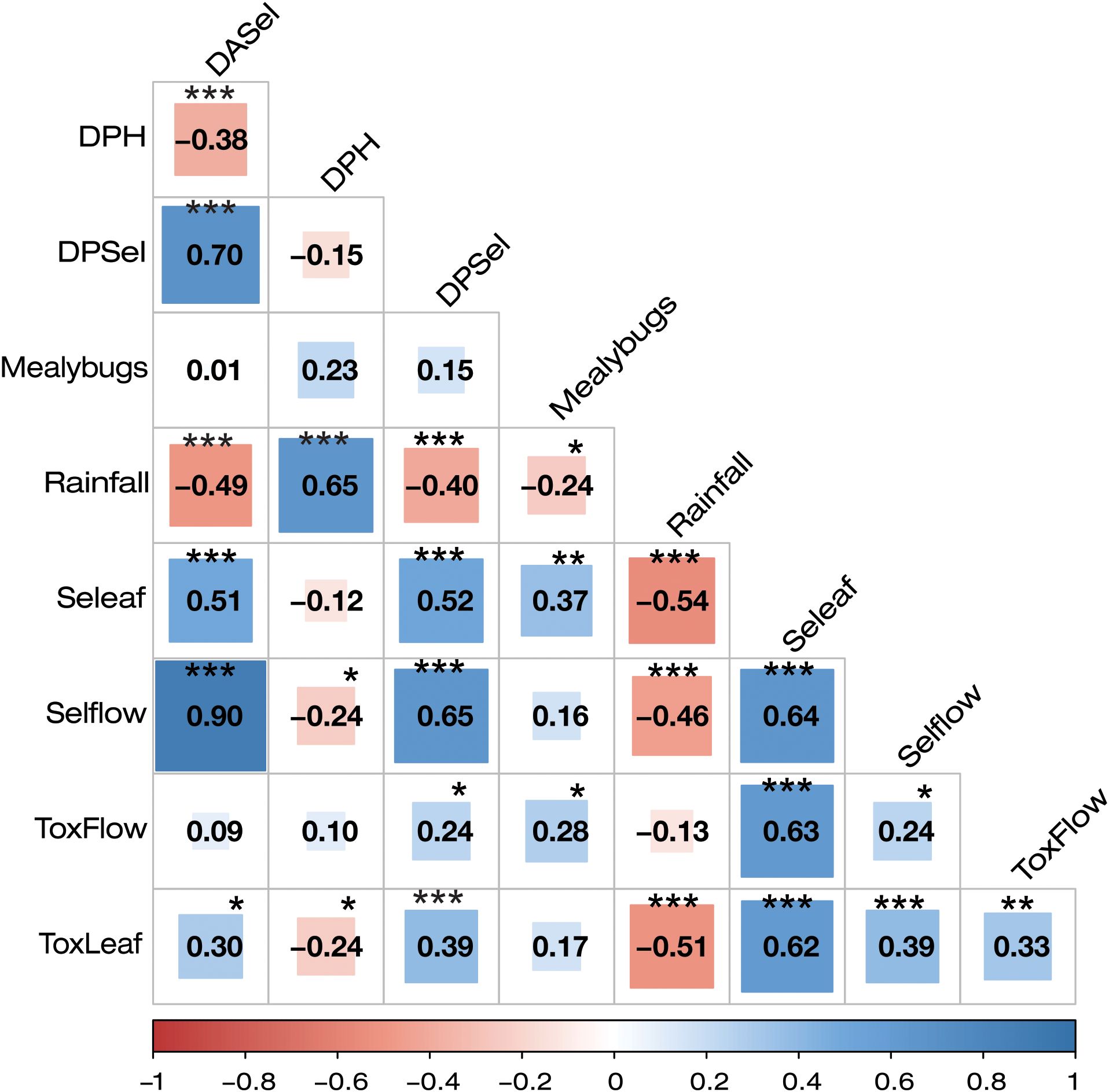
Figure 2. Spearman correlation analysis between population densities, damages and rainfall. ToxLeaf, Toxoptera on leaves; ToxFlow, Toxoptera on flowers; Seleaf, Selenothrips on leaves; Selflow, Selenothrips on flowers; DPH, pods damaged by Hemiptera; DPSel, pods damaged by Selenothrips; DASel, Area of the pod damaged by Selenothrips. p<0.05 (*), p<0.01 (**), p<0.001 (***).
3.1.2 Mealybugs, Planococcus sp.
Mealybugs of the genus Planococcus were only observed on leaves with maximum levels of 14 individuals (Figure 3). The analysis of variance showed differences in the population densities of pseudococcids depending on the cacao family tested (F = 28.21, df = 23, p-value = 0.0001) (Supplementary Table 3). Families F5 and F6 exhibited the highest densities of mealybugs while in F1, F2, and F3, the densities were significantly lower according the Fisher LSD test (Figure 3). Although with a low value in the correlation coefficient (r: -0.24), a significant negative association (p<0.05) was detected between precipitation and the abundance of these sucking insects (Figure 2).
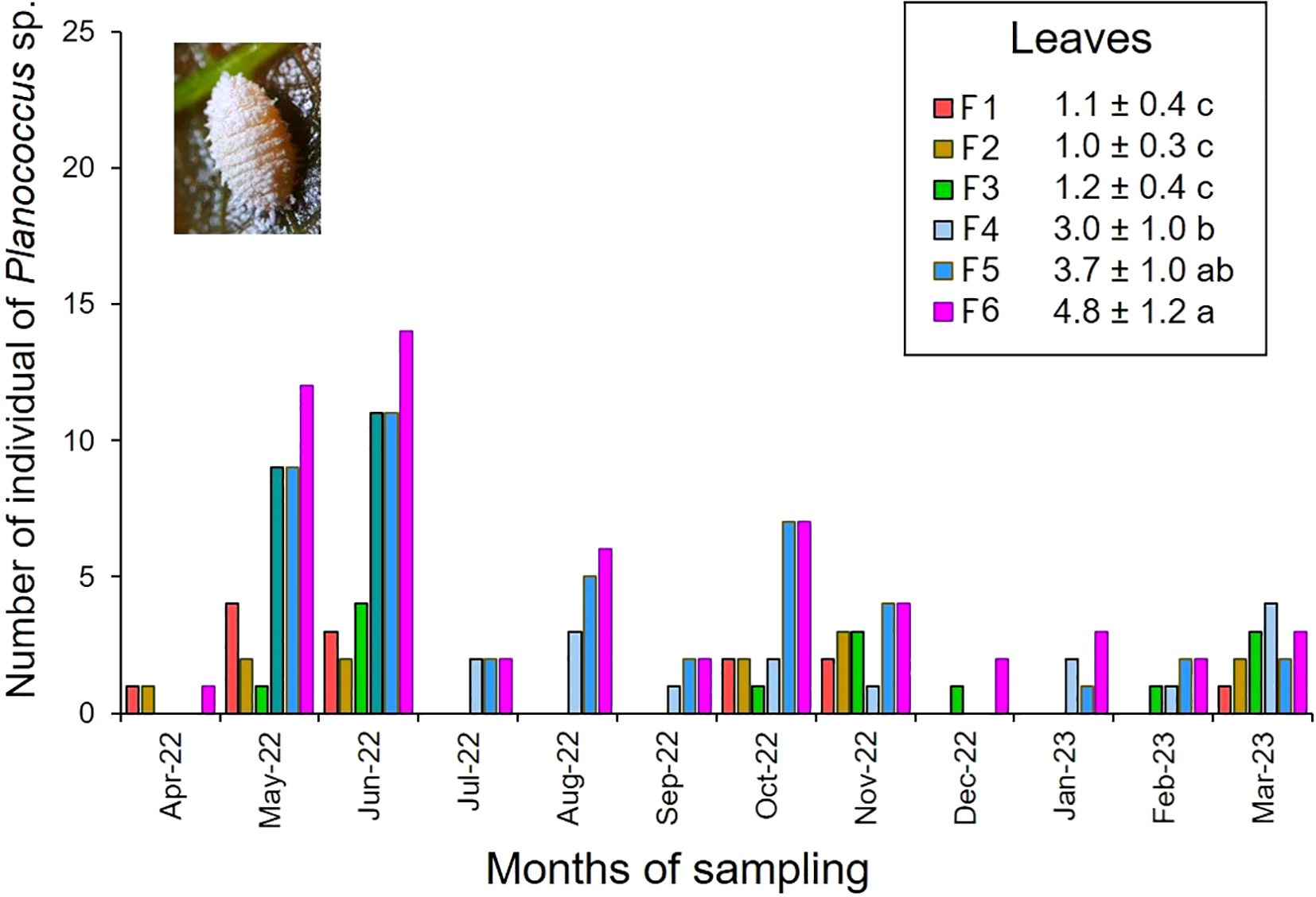
Figure 3. Number of individuals of Planococcus sp. observed on leaves of intra-specific hybrid cacao families. Period April 2022 - March 2023. Mean ± standard error. Mean followed by different letters in each column is significantly different using the Fisher LSD test (p<0.05).
3.1.3 The redbanded thrips, Selenothrips rubrocintus
On the leaves, the highest population densities of S. rubrocintus were observed in the period from May to August, while on the flowers, they were detected from May to October (Figure 4). In July 2022, maximum peaks were observed both on leaves and on flowers, ranging from 7 to 35 thrips per leaf and from 1 to 15 individuals per flower (Figure 4).
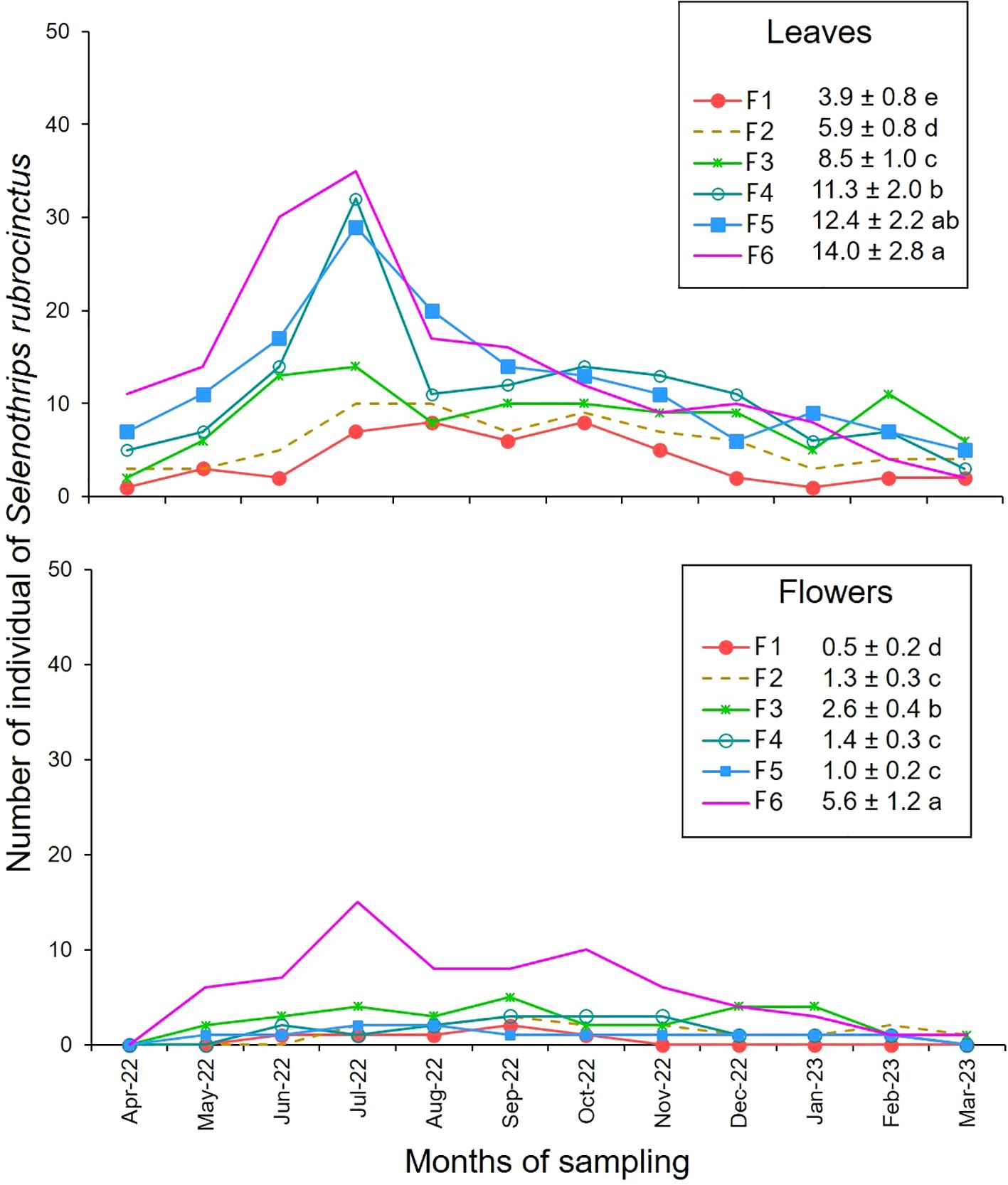
Figure 4. Number of individuals of Selenothrips rubrocintus observed on leaves and flowers of intra-specific hybrid cacao families. Period April 2022 - March 2023. Mean ± standard error. Mean followed by different letters in each column is significantly different using the Fisher LSD test (p<0.05).
Based on the cacao family, the population averages were significantly different in both leaves (F = 32.48, df = 23, p-value = 0.0001) (Supplementary Table 4) and flowers (F = 33.16, df = 23, p-value = 0.0001) (Supplementary Table 5). The Fisher LSD test detected higher populations of S. rubrocintus in both organs in Family F6 compared to the populations observed in family F1 (Figure 4). A highly significant negative correlation between S. rubrocintus and precipitation was detected for both leaves (r = -0.54; p<0.01) and flowers (r = -0.46; p<0.01) (Figure 2).
3.2 Damage to pods and the number of pods produced by cacao family
3.2.1 Damage to the pods by Selenothrips rubrocintus
Percentage of pods damaged by S. rubrocintus was statistically different depending on the cacao family evaluated (F = 4.86, df = 23, p-value = 0.004) (Supplementary Table 6). The highest percentages of damage were detected in families F3 and F6, while family F1 exhibited significantly lower damage (Figure 5A). On the other hand, according to the analysis of variance, the external pod area damaged by S. rubrocintus differed depending on the family studied (F = 28.89, df = 23, p-value = 0.004) (Supplementary Table 7). According to the Fisher LSD test, families F3 and F6 showed significantly higher damage, while damage was lower in pods from cacao family F5 (Figure 5B).
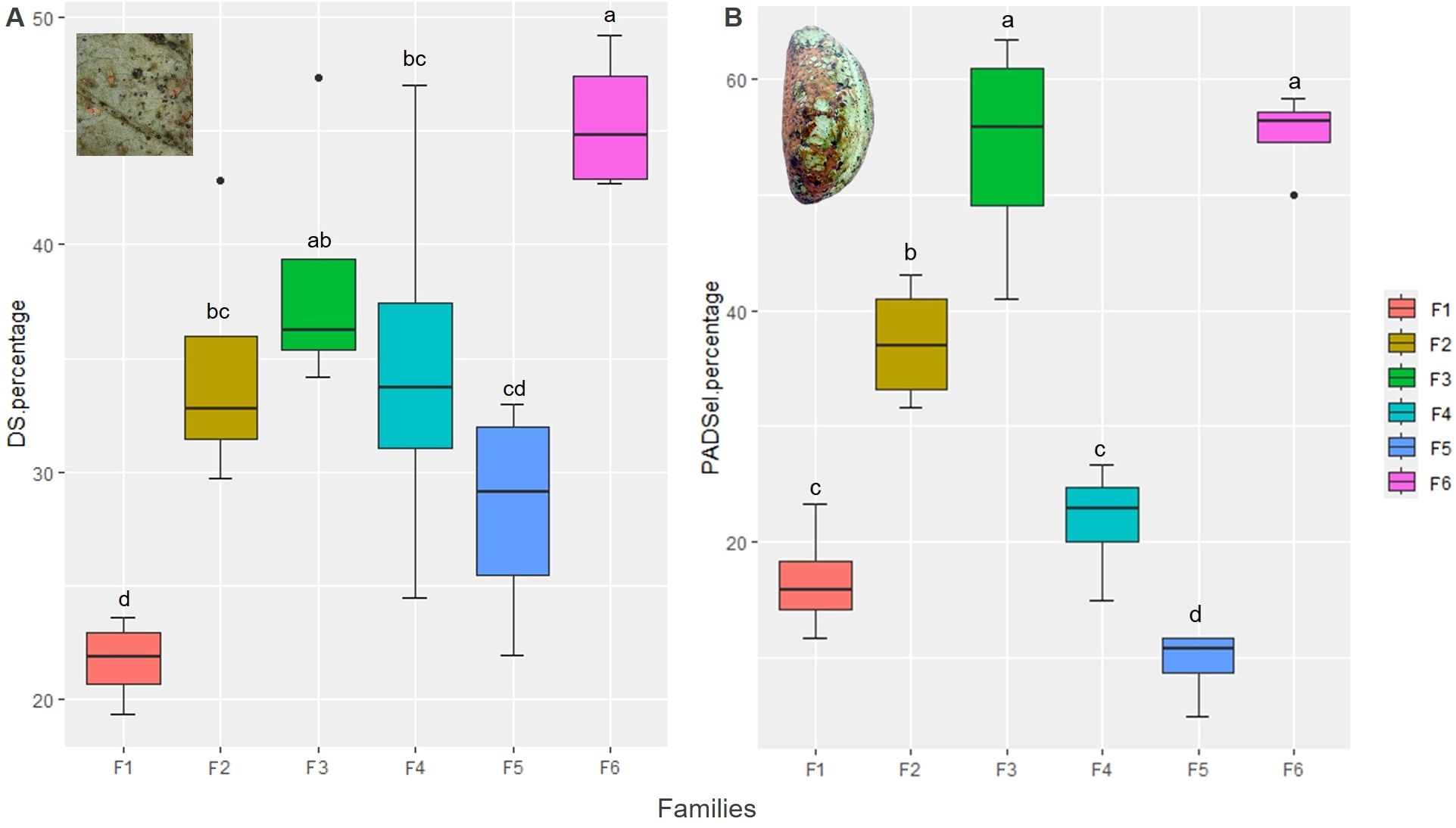
Figure 5. Percentage of damaged pods by Selenothrips rubrocintus (A) and area of damaged pod (B) in the families of cacao intraspecific hybrids. Mean followed by different letters in each column is significantly different using the Fisher LSD test (p<0.05).
Interestingly, populations of S. rubrocintus on flowers show a positive association with both the percentage of damaged pods (r: 0.65; p<0.01) and the percentage of damaged area of the pod by this species, the latter showing a very high correlation coefficient (r: 0.90; p<0.01) (Figure 2). Additionally, no association was found between the damage to cacao pods and the populations of S. rubrocintus developed on leaves even though on the latter organ the populations were higher (Figure 2).
3.2.2 Damage in the pods by Hemiptera
The damage to the pods by Hemiptera was caused by Leptoglossus zonatus (Dallas) (Hemiptera: Coreidae) and by Guayaquila gracillicornis (Stål) (Hemiptera: Membracidae) (Figure 6; Supplementary Figures 3-5). These damages were easily recognizable due to the concentric halo of brown coloration that forms around the puncture, which can vary in shade from light brown (center) to dark brown (periphery) (L. zonatus) or be completely brown without tonal changes (G. gracillicornis). Additionally, G. gracillicornis was found associated with bees of the genus Trigona (Figure 6; Supplementary Figure 5).

Figure 6. Adult (A), nymphs (B) and damage by Leptoglossus zonatus (C). Adult and its eggs (D) adults and nymphs together with bees of the genus Trigona (E) and damage by Guayaquila gracillicornis (F).
Significant differences for Hemiptera damage were found for the evaluated cacao families (F = 18.16, df = 23, p-value = 0.0001) (Supplementary Table 8). The percentage of pods damaged by Hemiptera was higher in F6, significantly differing from F1 and F2 (p<0.05) (Figure 7). Upon analyzing the scales of damage caused by Hemiptera (Figure 8), it is observed that families F1 and F2 have the highest percentages of undamaged pods (grade 0), with grade 1 being predominant over the rest of the damage scales, with variations according to family of cacao intraspecific hybrids. The Chi-square test = 126.75 was significant (p-value< 2.2e-16), indicating that the damage scales differ according to the family of cacao. Damage by Hemiptera was positively correlated with precipitation (Figure 2).
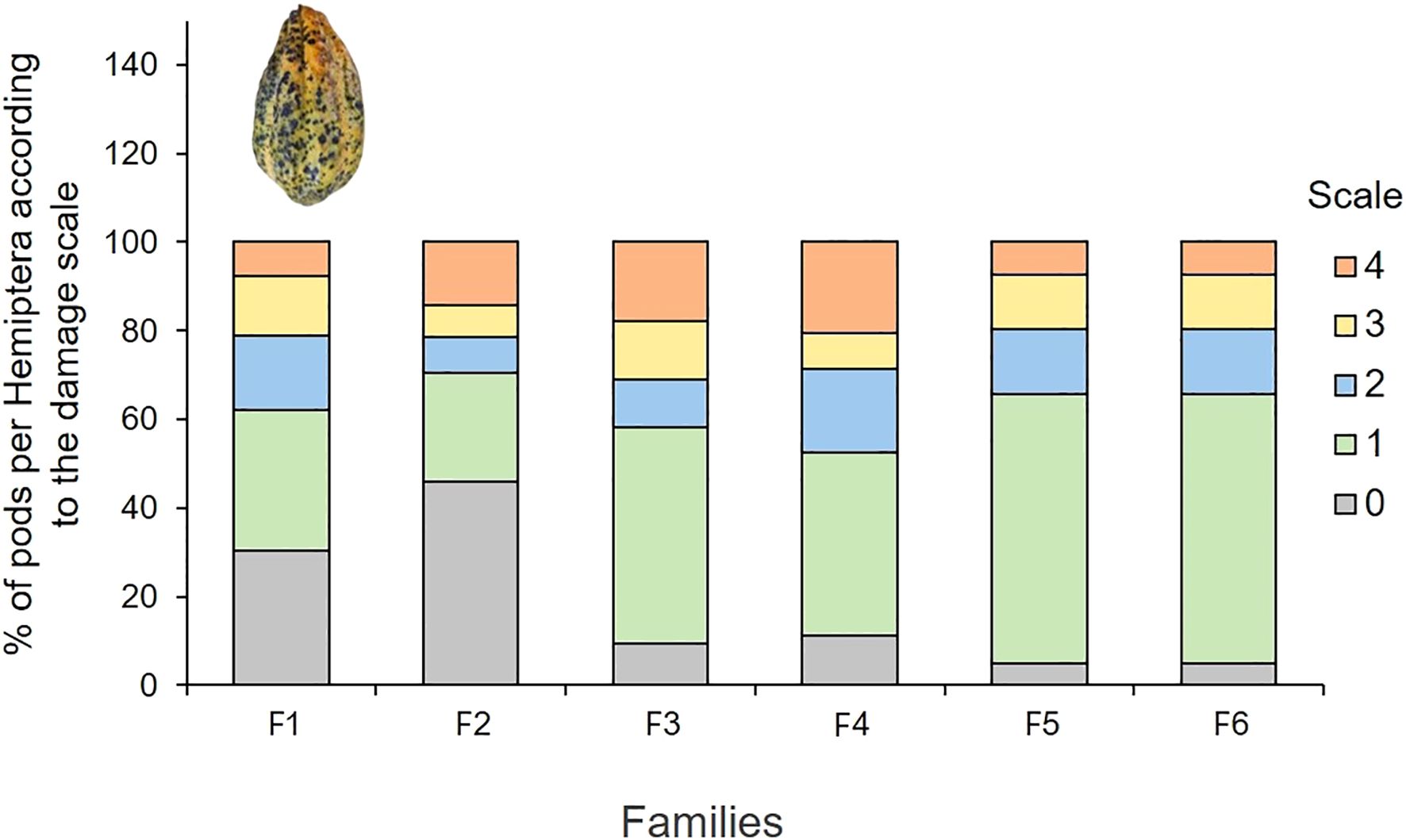
Figure 7. Percentage of Hemiptera damage in the families of in the families of cacao intraspecific hybrids. Mean followed by different letters in each column is significantly different using the Fisher LSD test (p<0.05).
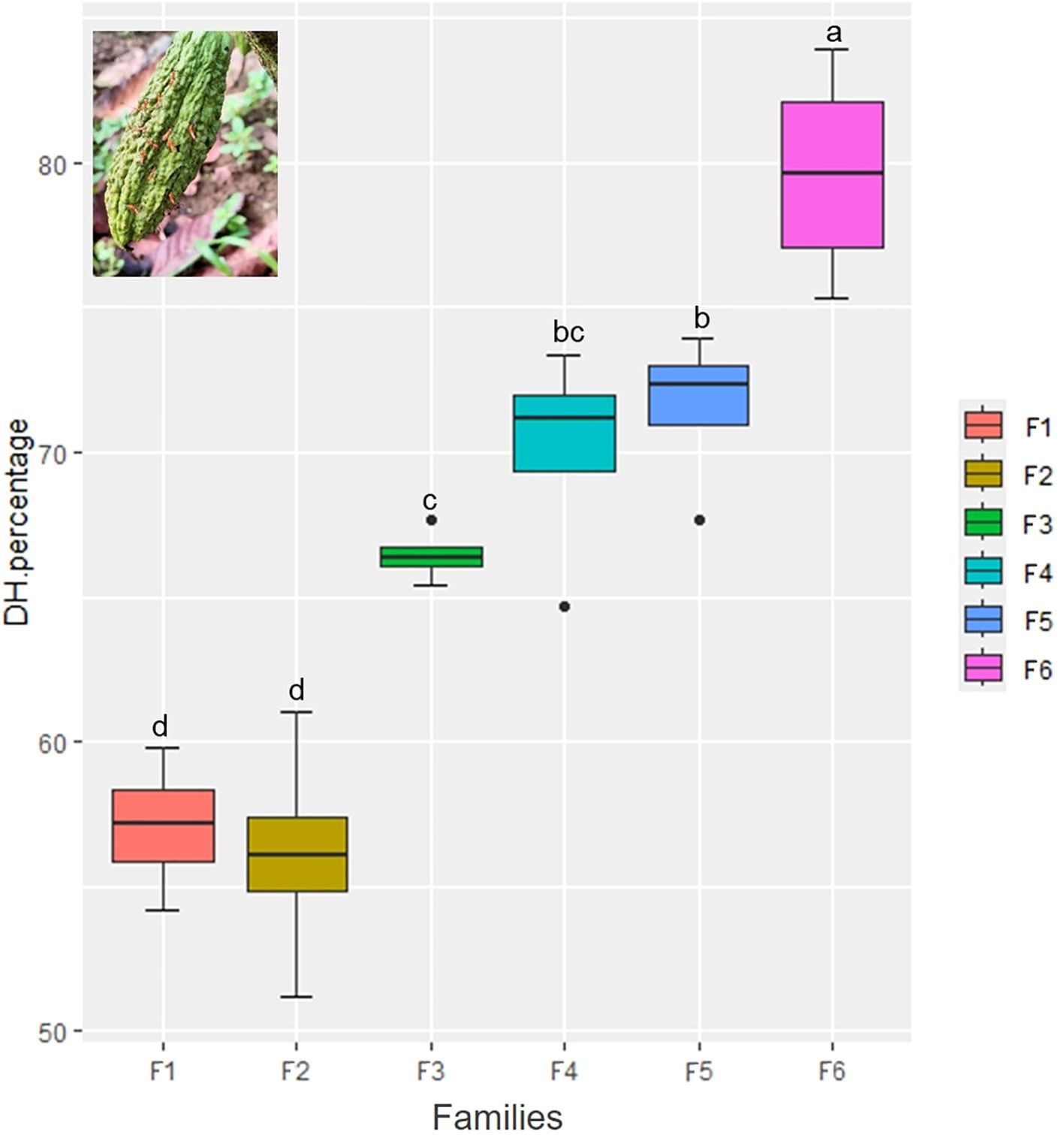
Figure 8. Percentage of Hemiptera-damaged pods in the different damage scales according to the number of punctures by the families of cacao intraspecific hybrids.
3.2.3 Number of pods
The number of pods varied significantly depending on the cacao family tested as indicated by the analysis of variance (F = 76.4, df = 23, p-value = 0.0001) (Supplementary Table 9). Families F1, F4, and F5 produced the highest number of pods, followed by families F3 and F6, and the number of pods was significantly lower in Family F2 (Table 3; p<0.05).
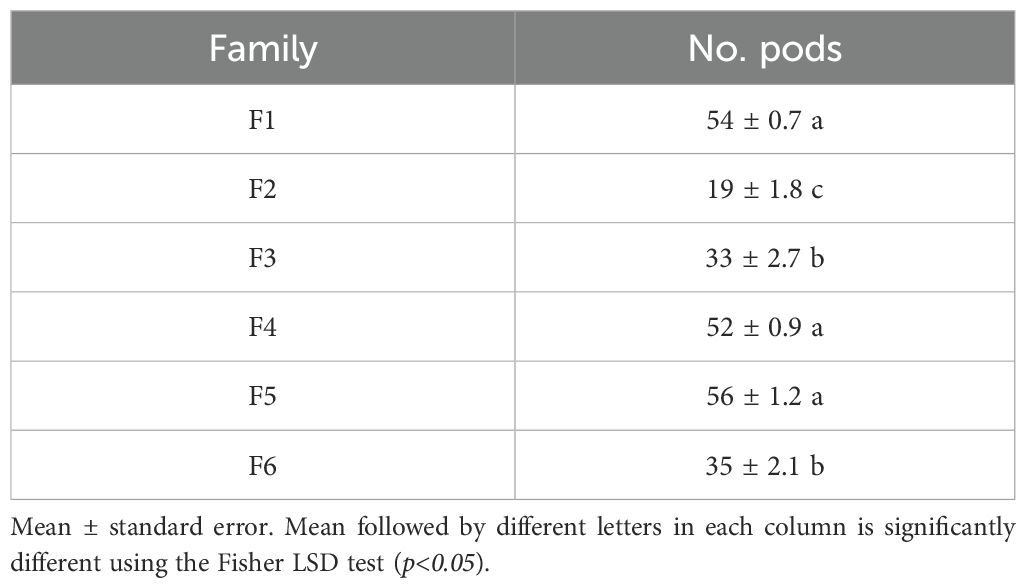
Table 3. Number of pods produced in one year per family of cacao intraspecific hybrid.
3.3 Comparison between families of intra-specific hybrids
The principal component analysis (Figure 9) shows that the populations of S. rubrocintus in flowers, the damage caused in the pod by this species, and the damage caused by Hemiptera are more associated with family F6.
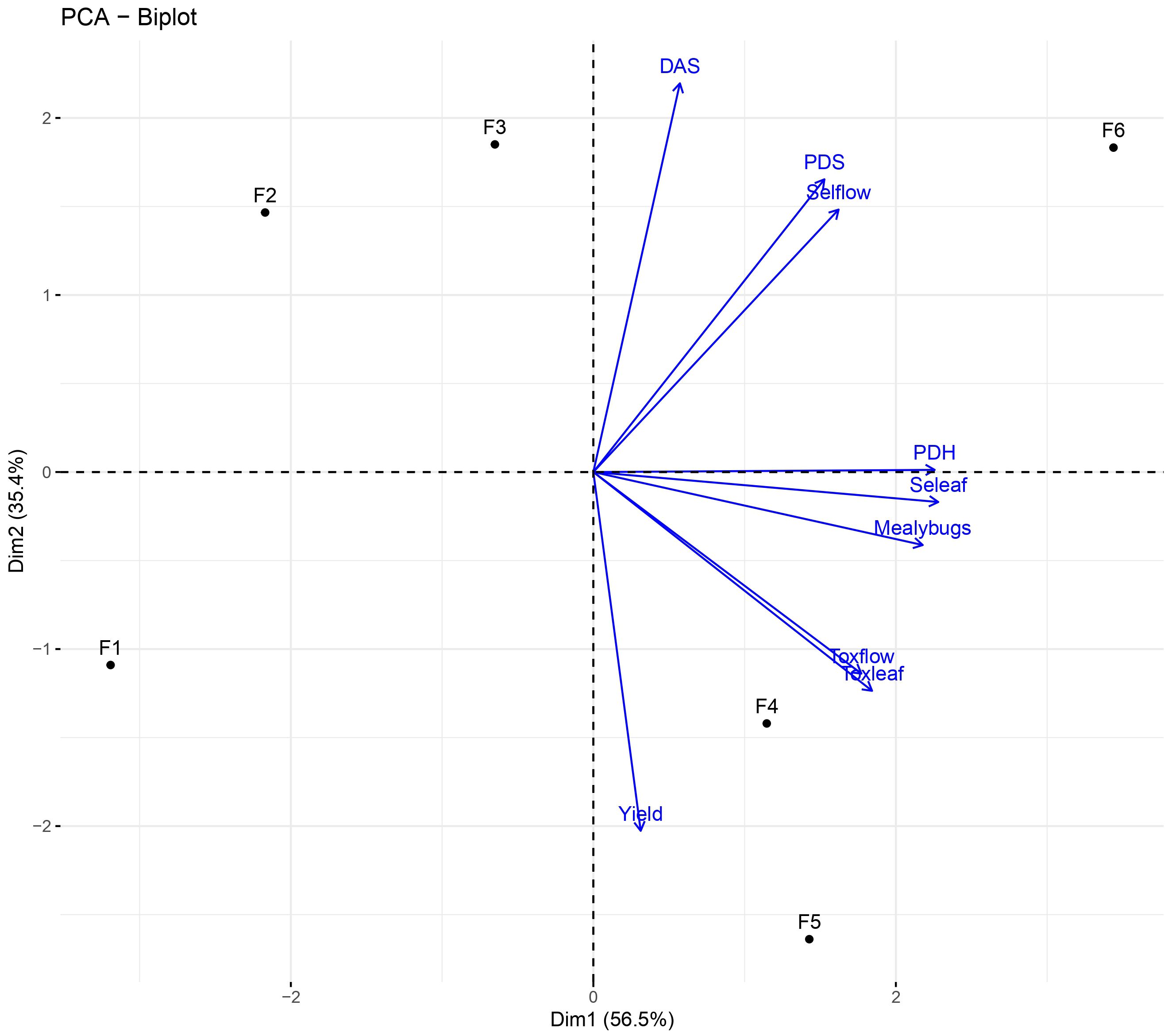
Figure 9. Principal component analysis for sucking pest population densities, damage and number of pods per in families of cacao intraspecific hybrids. ToxLeaf, Toxoptera on leaves; ToxFlow, Toxoptera on flowers; Seleaf, Selenothrips on leaves; Selflow, Selenothrips on flowers; PDH, Hemiptera damaged pods; PDS, Selenothrips damaged pods; DAS, Pod area damaged by Selenothrips.
Meanwhile, aphids on leaves and flowers, as well as scale insects, are primarily present in families F4 and F5. Population densities and damage caused by the evaluated insects are not associated with families F1, F2, and F3. Finally, the number of pods (yield) per year is more related to families F1, F4, and F5. In summary, although families F2 and F3 are less related to pests and damages, their yields are lower compared to the other families. Families F4 and F5 have high yields but also presence of pests, while family F6 combines pest presence with average yields. Additionally, family F1 is not associated with high population densities, and the yields are high.
4 Discussion
The population densities of sucking insects (T. aurantii, Planococcus sp., and S. rubrocintus) observed on leaves and flowers were influenced by the cacao family. With some variations, these insects were more abundant in families F6 and F5, while family F1 had fewer individuals detected. For other genetic materials of cacao, differential populations of several homopteran pests have also been observed. Campbell (1990) evaluated the incidence of three species of Pseudococcidae and T. aurantii in some progenies resulting from crosses between foreign cacao and found differential aphid populations, classifying them as susceptible or resistant depending on the bugs population densities found in the cacao progeny.
Adomako and Ackonor (2003) evaluated ten cacao selections from the upper Amazon, detecting differences in the susceptibilities of the evaluated selections to attacks by various homopteran insects. These latter researchers mentioned that one of the genotypes was less attractive to homopteran pests and consequently indicated that it could be used to reduce the damage degree from these pests. Based on our results, family F6 proved suitable for the population development of sap pests, while family F1 did not allow the increase of the population densities of these suckers, making its use promising.
Regarding S. rubrocintus, a field and laboratory study evaluated the behavior of two cacao clones,RT-18 and ICS-1 (both clones selected from the high Amazon) in the presence of S. rubrocintus on leaves (Fennah, 1965). The laboratory experiment demonstrated that the leaves of RT-18 were less attacked by S. rubrocintus, which was associated with characteristics of the leaves such as greater weight, more solid material, and higher water content compared to ICS-1. In fact, the RT-18 clone is considered a thrips-resistant clone (Müntzing, 1959; Fennah, 1965).
The importance of finding a genetic material of cacao less suitable for the development of populations of aphids, mealybugs, and thrips lies in mitigating the damage that these insects may cause to the plant. Delgado et al. (2023) indicated that T. aurantii is located on the underside of the leaves, in flowers, and also on newly formed pods, from which it extracts large amounts of sap. Thus, the high populations of T. aurantii can cause deformation of leaves and flowers, as well as premature dropping of the pods.
Mealybugs (Hemiptera: Pseudococcidae) insert their mouthparts, sucking sap from different organs of the plant, and some species inject toxins causing deformities that reduce the growth and vigor of the plant (Puig et al., 2021). Although the main economic impact associated with mealybugs is the transmission of viruses from the Badnavirus genus (Puig et al., 2021) no viruses associated with cacao have been reported in Ecuador.
Selenothrips rubrocintus feeds on leaves as well as on cacao pods and flowers; thrips are considered possible pollinators of cacao (Wolcott et al., 2023). Denmark and Wolfenbarger (2010) indicate that S. rubrocintus feeds on young leaves, leaving black spots that are the result of droppings falling onto the surface. Similarly, these researchers point out that high populations of this thrips can cause distortion and dropping of leaves.
Besides the genetic constitution of the plant, environmental conditions also seem to be determining factors in the abundances of cacao pests (Adomako and Ackonor, 2003). In this study, aphids, mealybugs, and thrips were more abundant in the dry season. Previous studies show that an increase in precipitation and temperature can decrease populations of aphids (Hasan et al., 2009; Behi et al, 2019). For this reason, high precipitation constitutes a regulatory factor of the population densities of aphids (Kaakeh and Dutcher, 1993). Crossley et al. (2022) indicate that when temperature and precipitation are high, the flight activity of aphids decreases, affecting the phenology of these sap-sucking insects.
In the study area, the rainy season is characterized by high temperatures (27-28°C) (Suplementary Figure 1). Thus, the lower population densities of T. aurantii detected in the rainy season could be attributed to the fact that high precipitation can wash away nymphs and adults, as well as delay flight, consequently decreasing the population, which is aided by the high temperatures that occur in the rainy season in the area.
Mealybugs also seem to be affected by rain. Moghaddam et al (2021) report that the arrival of rains and warm weather influence the decline of populations of mealybugs. The mealybug, Paracoccus marginatus Williams & Granara de Willink, an important pest of papaya, Carica papaya L., was more abundant in the dry season compared to the rainy season in a study conducted in Deli, Indonesia (Pramayudi et al., 2021). The research notes the mechanical control exerted by precipitation in washing away eggs and small nymphs; and consequently, the researchers estimate that to reduce the damage caused by this species, controls should be more intensive in the dry season (Pramayudi et al., 2021).
Selenothrips rubrocintus also showed a negative correlation with precipitation in studies evaluating cacao leaves in the Tabasco region, Mexico (Capetillo-Concepción et al., 2014), as well as in a study of population fluctuation conducted on leaves and flowers of cashew plants (Anacardium occidentale L.) in the Anton district, Panama (Atencio-Valdespino et al., 2023). Although in this research, the populations of Planococcus sp. were only observed on leaves, T. aurantii and S. rubrocintus were detected on leaves and flowers, with greater abundance on leaves. This agrees with other studies in which both insects were more abundant on leaves compared to flowers (Hasan et al., 2009; Atencio-Valdespino et al., 2023; Delgado et al., 2023).
The percentage of pods damaged by S. rubrocintus was also higher in family F6 and lower in F1. The affected area of the damaged pods was also highly significant in family F6, but the smallest affected area was detected in family F5.
The detection of genetic materials where the pod is less damaged by this species is relevant as a management practice for this pest. Cañarte-Bermúdez and Navarrete-Cedeño (2021) mentioned that the damage by S. rubrocintus is important in both leaves and pods, and on the latter, by feeding with their rasping-sucking mouthparts, the sap rises to the surface where it oxidizes upon contact with air, causing brown wounds with a cork-like appearance. Walter et al. (2018) mention that although these are cosmetic damages, it complicates the assessment of pod maturation. We estimate that the damage caused by this species could exceed cosmetic damages. Our observations suggest that, depending on the degree of impact and the age at which the damage occurs, the growth of the pod could be limited (unpublished data). It is important to emphasize that S. rubrocintus can move from flowers to newly formed pods, and under these conditions, if high populations develop, the damage could reduce pod growth, which could affect yield.
Following a similar pattern to population densities, pod damage caused by thrips was greater in the dry season. Additionally, populations of S. rubrocintus developed in flowers were positively correlated with the damaged pods and especially with the affected area of the pod. This suggests that thrips populations in flowers could be an indicator of damage to pods, and therefore, sampling of thrips on this organ is important to predict possible pod damages.
The damage caused to pods by the hemipteran species, L. zonatus and G. gracillicornis detected in this study is reported for the first time in cacao cultivation. The characteristics of adults and nymphs of L. zonatus coincide with those described in other studies (Brailovsky, 2014; Collantes et al., 2016; Joyce et al., 2017). The characteristics of G. gracillicornis are consistent with those reported by Flórez-V et al (2015). In this study, this species was found feeding on cacao pods in symbiosis with Trigona sp. The symbiosis of G. gracillicornis with ants (Atencio-Valdespino et al., 2023) or with wasps (Flórez-V et al, 2015) had been reported. Both reports constitute interesting findings for Ecuador as a center of origin for this crop and could indicate pests associated with other crops adapting to cacao.
In Ecuador, the membracid species Amastris dissimilis Broomfield, Bolbonota pictipennis Fairmaire, Membracis foliata subsp. c-album Fairemaire, and Horiola picta Coquebert had been previously reported on cacao leaves (Wright, 1984) without reports of damage to the pods. In Peru, a neighboring country to Ecuador, three membracid species associated with cacao pods have been reported: H. picta, Cyphonia clavata Fabricius, and Bolbonota globosa Fairmaire, but without information on population levels and damage caused by these species (Delgado et al., 2023). During our evaluations, we found damage consistent with the presence of G. gracillicornis in symbiosis with Trigona sp., a membracid species that is new to the country and causes damage to cacao pods. In our exhaustive searches of researches conducted in this and other cacao-producing countries, we found no reports of this insect damaging the pods of this crop. This demonstrates the relevance of this study.
In Ecuador, M. dissimulatum had been reported causing damage to the pod (Riera et al, 2013; Gamboa et al, 2020; Cañarte-Bermúdez and Navarrete-Cedeño, 2021). However, there is scarce information regarding the taxonomic identification of this species in cacao-producing areas in the country. There are references for the taxonomic identification of M. dissimulatum in Quevedo, the province of Los Ríos (Ecuadorian coast) and in Quito (capital of Ecuador, Andean region) (Riera et al, 2013; Gamboa et al, 2020). Quevedo and Lodana are located at an altitude of 74 and 60 m.a.s.l. respectively, while Quito is at 2800 m.a.s.l. For M. dissimulatum, there are no references regarding altitudinal distribution, but for other Monalonion species, altitudinal distributions ranging from 600 to 1800 m.a.s.l. are reported (Gamboa et al, 2020).
Since Quevedo and Lodana are at low altitudes (less than 100 m), and the insect is present in Quevedo, elevation is not the factor that determines the absence of M. dissimulatum in Lodana. Both regions are part of the Ecuadorian coast, but the life zone of Lodana is a tropical dry forest while Quevedo is characterized as a tropical wet forest (Holdridge, 1967). Thus, the annual precipitation of Quevedo (3000 mm) triples that of Lodana (1000 mm). Additionally, in Quevedo, relative humidity exceeds 90%. Previous studies report that M. dissimulatum has been found in South American countries in coastal and mountainous areas characterized by high humidity, elevated temperatures, and excessive shade favoring its abundance (Figueroa, 1952; Vargas et al, 2005). Consequently, Lodana does not seem to have the appropriate climatic conditions for the presence of the species.
Damage from hemipterans was more abundant in family F6 and lower in F1. Of the pods with punctures, in family F1 the highest scale was scale 1, which means there were between 1 and 25 punctures. This agrees with what was observed by Mahob et al. (2019) who evaluated the damage caused by the bug, Sahlbergella singularis Haglund (Hemiptera: Miridae) in the pods of various cacao genotypes, finding that the damage averaged 30 punctures. In general, losses of 4-5% are reported due to the damage caused to pods by other species of hemipterans (Babin et al., 2012; Mahob et al., 2019).
Contrasting with what was found for the population densities of sucking pests and damage from thrips, damage from Hemiptera was greater during the rainy season. This seems to be attributed to the habits of the insects causing this damage. Membracid species have been found to be more abundant in the rainy season with high temperatures (Lopes, 1995). For L. zonatus, the increase in temperature shortens the cycle and allows for a greater number of generations per year. Tepole-García et al. (2016) determined a shorter biological cycle duration of L. zonatus as the temperature increased, which is attributed to the fact that the increase activates the functioning of enzymes, accelerating chemical reactions and thus decreasing development time.
Leptoglossus zonatus is a polyphagous pest associated with several host plants in South America, which causes significant losses in some crops (Foresti et al., 2017). For its part, species within the family Membracidae, in cacao, have been reported as secondary pests infesting leaves, stems, flowers, and pods (Sánchez-López and Sánchez-Soto, 2019; Da Silva et al., 2020). In this study, the membracid caused damage to the pods similar to that caused by L. zonatus. Both species imply direct damage from the insertion of the mouthparts and also indirect damage. Feeding injuries from heteropterans and membracids in some crops provide a point of entry for fungal spores and also vectorize other types of pathogens (Mitchell, 2004; Atencio-Valdespino et al., 2023).
Damage from hemipterans was more abundant during the rainy season, where temperatures seem to favor the occurrence of the insects. Unlike aphids, scale insects, and thrips, these insects are larger and have greater mobility, which possibly contributes to their protection from the rain during their occurrence, a situation that is limited in the first group of insects, where small size, being apterous or poor flyers limits their movement and thus they tend to be swept away by water during rainy periods.
Families F4, F5, and F6 had a greater abundance of sucking herbivores and generally greater damage to the pods. On the other hand, the number of pods from the families of interspecific hybrids of cacao were also differential. Family F1 produced 1.6 times more pods than family F6. Since these are new hybrids resulting from crosses between national and Trinitarian cacaos, the evaluation of the population densities of the herbivores is essential to classify these hybrids depending on the infestations reached, thus determining them as susceptible, tolerant, or resistant to pests. Having been scarcely infested throughout the entire study year, F1 could be classified as resistant to the population densities of sucking pests and tolerant to damage in the pods (20 to 55%), while F6 would be classified as susceptible both to pest populations and to damage in the pods (45 to 75%).
On the other hand, the lower population densities and damage in F1 could be attributed to antixenosis. Throughout our sampling, we found no evidence of antibiotic effects or dead insects once the feeding of the insects began. Then some families were more suitable than others. It is important to highlight that the different evaluated cacao families were randomly located within the same experimental plot designed in blocks at random. Consequently, the insects had the same probability of choosing among the existing family plants. Antixenosis is a term proposed by Kogan and Oatman (Kogan and Ortman, 1978), to refer to the “non-preference” of insects for the plants. In this type of resistance, the plant avoids colonization or the initiation of feeding due to strategies that include morphological changes, such as variations in plant surface, color changes, taste, wax, or pubescent leaves or gummy exudates, among others (Kogan and Ortman, 1978).
However, antibiosis or adverse effects from a particular family on the biology of the insect that may have contributed to the lower populations in those genetic materials cannot be ruled out. Future studies should focus on evaluating population densities in different genetic materials in various production areas, as well as yield estimates that include grain weight, to calculate predictive models between pest population densities and the effect on cacao productivity. While one of the hybrids that contains CCN-51 (F4) as a parent proved to be highly productive, they also showed an abundance of associated phytophagous, including aphids, thrips, and scale insects, especially in the dry season, as well as damage to the pods caused by hemipterans in the rainy season.
Therefore, under a possible scenario of mass plantings and potential applications of pesticides, infestations in these genetic materials could increase due to factors such as climate (especially temperature) and the reduction of natural enemies resulting from the imbalances caused by frequent pesticide spraying. This research represents an important contribution as studies on the plant-herbivore-climate interaction had not been conducted in Ecuador until now. Additionally, L. zonatus and G. gracillicornis are reported for the first time as causing damage to cacao pods in Ecuador.
5 Conclusion
Regarding the abundance of sucking pests and pod damage, these results allow for the separation of families (intraspecific hybrids) into two groups. One group includes families F6 > F5 > F4, which showed higher populations and damage, contrasting with a second group composed of F3 > F2 > F1, families where generally the lowest populations and damage occurred. Among the families least affected by pests, family F1 produced a high number of pods during the evaluation year. Thus, family F1 (L26-H64 x L11-H19), which has national cacao as parents, proved to be as productive as the one with CCN-51 as one of the parents (F4 = L26-H64 x CCN-51), and F1 was also less damaged by pests. This makes this genetic material promising for pest resistance with good productivity, highlighting the importance of national genotypes in Ecuador. To our knowledge, Leptoglossus zonatus and Guayaquila gracillicornis are reported for the first time attacking cacao pods.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
DC: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Validation, Visualization, Writing – original draft, Writing – review & editing. RC: Conceptualization, Investigation, Methodology, Validation, Visualization, Writing – original draft, Writing – review & editing. FS: Data curation, Formal analysis, Methodology, Validation, Writing – original draft, Writing – review & editing. JC-O: Conceptualization, Investigation, Validation, Visualization, Writing – original draft, Writing – review & editing. CM: Conceptualization, Data curation, Validation, Writing – original draft, Writing – review & editing. PM: Data curation, Investigation, Methodology, Validation, Writing – original draft, Writing – review & editing. LG: Funding acquisition, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. To the Technical University of Manabí for the academic support given to this research. Grant No. PYTAUTO3607-2020-FIAG0023
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fagro.2025.1588450/full#supplementary-material
References
Adomako B. and Ackonor J. (2003). Genetic analysis of attractiveness of Upper Amazon cocoa selections to homopterous insect pests. Trop. Sci. 43, 39–43. doi: 10.1002/ts.86
Alomía J., Alomía C., and Vega B. (2021). Carmenta foraseminis Eichlin y Phytophthora palmivora en frutos de Theobroma cacao L. en Satipo, Perú. Manglar 18, 283–288. doi: 10.17268/manglar.2021.037
Armengot L., Ferrari L., Milz J., Hohmann P., and Schneider M. (2020). Cacao agroforestry systems do not increase pest and disease incidence compared with monocultures under good cultural management practices. Crop Prot. 130, 105047. doi: 10.1016/j.cropro.2019.105047
Atencio-Valdespino R., Aguilera-Cogley V., Barba-Alvarado A. A., Jáen M., and Herrera-Vásquez J. A. (2023). Presencia de Guayaquila gracilicornis (Stål) (Hemiptera: Membracidae) y su asociación con hormigas en el cultivo de marañón (Anacardium occidentale L.) en Panamá. IDESIA (Chile) 41, 61–68. doi: 10.4067/S0718-34292023000300061
Babin Y. R., Djieto-Lordon C., Cilas C., Dibog L., Mahob R., and Bilong Bilong C. F. (2012). True bug (Heteroptera) impact on cocoa fruit mortality and productivity. J. Economic Entomology 105, 1285–1292. doi: 10.1603/EC12022
Behi F., Souissi R., and Boukhris-Bouhachem S. (2019). Temporal changes in the aphid–natural enemy complex in Tunisian citrus over two decades. J. Entomological Sci. 54, 357–369. doi: 10.18474/JES18-97
Brailovsky H. (2014). Illustrated key for identification of the species included in the genus Leptoglossus (Hemiptera: Heteroptera: Coreidae: Coreinae: Anisoscelini), and descriptions of five new species and new synonyms. Zootaxa 3794, 143–178. doi: 10.11646/zootaxa.3794.1.7
Campbell C. A. M. (1990). The susceptibility of cocoa to mealybugs (Pseudococcidae) and other honeydew-producing Homoptera in Ghana. Bull. Entomological Res. 80, 137–151. doi: 10.1017/S0007485300013353
Cañarte-Bermúdez E. and Navarrete-Cedeño B. (2021). Reconocimiento de artrópodos - plaga y controladores biológicos como herramienta para el manejo ecológico de plagas en cacao. 1 edición (Portoviejo, Ecuador: Instituto Nacional de Investigaciones Agropecuarias). Available online at: http://repositorio.iniap.gob.ec/handle/41000/5750 (Accessed April 4, 2025).
Capetillo-Concepción E., Córdova Avalos V., Sánchez Soto S., Romero Nápoles J., Pérez de la Cruz M., and Mendoza Hernández J.H.R. (2014). Fluctuación poblacional de Selenothrips rubrocinctus (Giard) en cultivo de cacao en Huimanguillo Vol. 49 (Tabasco, México: Revista Peruana de Entomología), 137–142. Available online at: https://www.revperuentomol.com.pe/index.php/rev-peru-entomol/article/view/244 (Accessed April 4, 2025).
Castro-Olaya J., Chirinos D. T., and Kondo T. (2024). Scolytines (Coleoptera: curculionidae: scolytinae) associated with cacao monoculture and agroforestry systems. Sarhad J. Agric. 39, 93–102. doi: 10.17582/journal.sja/2023/39/s2.93.102
Cilas C. and Bastide P. (2020). Challenges to cocoa production in the face of climate change and the spread of pests and diseases. Agronomy 10, 1–8. doi: 10.3390/agronomy10091232
Collantes González R. C., Rodríguez P., Romero B., and Rodríguez E. (2016). Estados inmaduros de Leptoglossus zonatus (Hemiptera, Coreidae): agente relacionado con la caída de naranjas (Citrus sinensis) en Azuero, Panamá. Aporte Santiaguino 9, 93–100. doi: 10.32911/as.2016.v9.n1.216
Crossley M. S., Lagos‐Kutz D., Davis T. S., Eigenbrode S. D., Hartman G. L., Voegtlin D. J., et al. (2022). Precipitation change accentuates or reverses temperature eff ects on aphiddispersal. Ecol. Appl. 35, e2593. doi: 10.1002/eap.2593
Cubillos G. (2013). Manual del perforador de la mazorca del cacao: Carmenta foraseminis (Busck) Eichlin. Ed. Cubillos G. (edellín: Compañía Nacional de Chocolates S.A.S). Available online at: https://repository.agrosavia.co/handle/20.500.12324/13314 (Accessed April 4, 2025).
Da Silva A. F. C., de Oliveira Moreira S. M.D.C., da Silva Cunha R. D.S., Moreira D. A., Teodoro A., do Nascimento A. H.C., et al. (2020). Effect of isolates of Beauveria bassiana pathogenic to leafhopper cacao Horiola picta (Hemiptera: Membracidae). Amazonian J. Plant Res. 4, 456–461. doi: 10.26545/ajpr.2020.b00054x
Delgado C. and Couturier G. (2017). Primer registro de Xylosandrus compactus (Coleoptera: Curculionidae: Scolytinae) sobre cacao en Perú. Rev. Colombiana Entomologia 43, 121–124. doi: 10.25100/socolen.v43i1.6659
Delgado C., Couturier G., Balcazar L., and Chichipe A. (2023). Insect pests of Theobroma cacao (Malvaceae) in the Peruvian Amazon. Trop. Agric. 100, 110–114. Available online at: https://journals.sta.uwi.edu/ojs/index.php/ta/article/view/7854 (Accessed April 4, 2025).
Denmark H. A. and Wolfenbarger D. O. (2010). “Redbanded thrips, selenothrips rubrocinctus (Giard) (Insects: thysanoptera: thripidae),” in Division of plant industry, florida department of agriculture and consumer service, gainesville, florida, entomology circular, University of Florida, USA, 108. doi: 10.32473/edis-in256-1999
Díaz-Valderrama J. R., Leiva-Espinoza S. T., and Aime M. C. (2020). The history of cacao and its diseases in the Americas. Phytopathology 110, 1604–1619. doi: 10.1094/PHYTO-05-20-0178-RVW
Dibog L., Babin R., Mbang J. A.A., Decazy B., Nyassé S., Cilas C., et al. (2008). Effect of genotype of cocoa (Theobroma cacao) on attractiveness to the mirid Sahlbergella singularis (Hemiptera: Miridae) in the laboratory. Pest Manage. Sci. 64, 977–980. doi: 10.1002/ps.1589
Dietrich C. H. and Deitz L. L. (1991). Revision of the neotmpical treehopper tribe aconophorini (Homoptera: membracidae). Eds. Dietrich C. H. and Deitz L. L. (North Carolina: North Carolina State University). Available online at: https://d.lib.ncsu.edu/collections/catalog/ua101_001_301502_20230127_57612?c=&m=&cv=&xywh=-5035%2C-257%2C13654%2C5131 (Accessed April 4, 2025).
Fachin G., Pinedo K., Vásquez J., Flores E., Doria M., Alvarado J., et al. (2024). Factores ambientales y su relación con la incidencia de Carmenta foraseminis (Busck) Eichlin (Lepidoptera: Sesiidae) en frutos de Theobroma cacao “cacao” en San Martín, Perú. Boletín Científico del Centro Museos 28, 133–145. doi: 10.17151/bccm.2019.23.2.6
Fennah R. G. (1965). The influence of environmental stress on the cacao tree in predetermining the feeding sites of cacao thrips, Selenothrips rubrocinctus (Giard), on leaves and pods. Bull. Entomological Res. 56, 333–349. doi: 10.1017/S000748530005642X
Figueroa A. (1952). Monalonion sp. plaga importante en el cacao del valle del Cauca – Colombia. Acta Agronomica II, 183–193. Available online at: https://revistas.unal.edu.co/index.php/acta_agronomica/article/view/49142 (Accessed April 4, 2025) (Accessed April 4, 2025).
Flórez-V C., Wolff M. I., and Cardona-Duque J. (2015). Contribution to the taxonomy of the family Membracidae Rafinesque (Hemiptera: Auchenorrhyncha) in Colombia. Zootaxa 3910, 001–261. doi: 10.11646/zootaxa.3910.1.1
Foresti J., Bastos C., Fernandes F. L., and Da Silva P. R. (2017). Economic Injury Levels and Economic Thresholds for Leptoglossus zonatus (Dallas) (Hemiptera: Coreidae) infesting seed maize. Pest Manage. Sci. 74, 149–158. doi: 10.1002/ps.4671
Gamboa J., Serna F., and Morales I. (2020). Estado actual del conocimiento taxonómico del género Monalonion Herrich-Schaeffe 1850(Hemiptera: Heteroptera: Miridae: Bryocorinae: Monaloniini) , Boletín Científico. Centro Museos. Museo Hist. Natural 24, 144–168. doi: 10.17151/bccm.2020.24.2.11
Hasan M. R., Ahmad M., Rahman M. H., and Haque M. A. (2009). Aphid incidence and its correlation with different environmental factors. J. Bangladesh Agric. Univ. 7, 15–18. Available online at: https://ageconsearch.umn.edu/record/208329/?v=pdf (Accessed April 4, 2025).
Holdridge L. (1967). Life zone ecology. San José, Costa Rica: Tropical Science Center. Available online at: https://reddcr.go.cr/sites/default/files/centro-de-documentacion/holdridge_1966_-_life_zone_ecology.pdf (Accessed April 4, 2025).
Jaimez R. E., Barragan L., Fernández-Niño M., Wessjohann L. A., Cedeño-Garcia G, Sotomayor Cantos I., et al. (2022). Theobroma cacao L. cultivar CCN 51: a comprehensive review on origin, genetics, sensory properties, production dynamics, and physiological aspects. PeerJ, e12676. doi: 10.7717/peerj.12676
Joyce A. L., Higbee B. S., Haviland D. R., and Brailovsky H. (2017). Genetic variability of two leaffooted bugs, Leptoglossus clypealis and Leptoglossus zonatus (Hemiptera: Coreidae) in the Central Valley of California. J. Economic Entomology 110, 2576–2589. doi: 10.1093/jee/tox222
Kaakeh W. and Dutcher J. D. (1993). Effect of rainfall on population abundance of aphids (Homoptera: aphididae) on pecan. J. Entomological Sci. 28, 283–286. Available online at: https://jes.kglmeridian.com/meridian/jes/published/rest/pdf-watermark/v1/journals/ents/28/3/article-p283.pdf/watermark-pdf/ (Accessed April 4, 2025).
Kogan M. and Ortman E. F. (1978). Antixenosis: a new term proposed to define Painter s “nonpreference” modality of resistance. Bull. Entomological Soc. America 24, 175–176. doi: 10.1093/%20besa/24.2.175
Lanaud C., Vignes H., Utge J., Valette G., Rhoné B., et al. (2024). A revisited history of cacao domestication in pre−Columbian times revealed by archaeogenomic approaches. Sci. Rep. 14, 2972. doi: 10.1038/s41598-024-53010-6
Lopes B. (1995). Treehoppers (Homoptera: Membracidae) in southeastern Brazil: use of host plants. Rev. Bras. Zoologia 12, 595–608. doi: 10.1590/S0101-81751995000300015
Lopez G., Gaiser T., Ewert F., and Srivastava A. (2021). Effects of recent climate change on maize yield in Southwest Ecuador. Atmosphere 12, 299. doi: 10.3390/atmos12030299
Mahob R. J., Ngah I. M., Feumba R. D., Mahot H. C., Bassogog C. B., Bilong C. F., et al. (2019). Field evaluation of the impact of Sahlbergella singularis Haglund infestations on the productivity of different Theobroma cacao L. genotypes in the Southern Cameroon. J. Plant Dis. Prot. 126, 203–210. doi: 10.1007/s41348-019-00221-z
Maron J. L., Agrawal A. A., and Schemske D. W. (2019). Plant–herbivore coevolution and plant speciation. Ecol. 100, e02704. doi: 10.1002/ecy.2704
Ministerio de Agricultura y Ganadería (MAG) (2025). Agroproductive data, Ministerio de Agricultura y Ganadería Available online at: http://sipa.agricultura.gob.ec/index.php/cifras-agroproductivas (Accessed February 15, 2024).
Mitchell P. L. (2004). Heteroptera as vectors of plant pathogens. Neotropical Entomology 33, 519–545. doi: 10.1590/S1519-566X2004000500001
Moghaddam M., Abdollahipour M., and Fathipour Y. (2021). Scale insects , in Omkar (ed.) Polyphagous pests of crops (Singapure: Springer), 273–309. doi: 10.1007/978-981-15-8075-8_6
Morán P., Agrawal A. A., and Schemske D. W. (2025). “Sustainable pest management of cacao in the Neotropics. Challenges and opportunities,” in Sustainable cacao cultivation in Latin America, 1st edn. Ed. Garcia L. C. (Routledge, New York), 168–190. doi: 10.4324/9781003381761-12
Müntzing A. (1959). Results of field trials with cacao at hacienda clementina, Ecuador. I. Disease resistance. Acta Agriculturae Scandinavica 9, 129–148. doi: 10.1080/00015125909434241
Paladines-Rezabala A., Moreira-Morrillo A. A., Mieles A., and Garcés-Fiallos F. R. (2022). Avances en la comprensión de la interacción entre Ceratocystis cacaofunesta y Xyleborus ferrugineus (Coleoptera: Curculionidae: Scolytinae) en árboles de cacao. Scientia Agropecuaria 13, 43–52. doi: 10.17268/sci.agropecu.2022.004
Páliz V., Mendoza J., Cansing V., and INIAP (1982). “Insectos asociados al cultivo del cacao en el Ecuador,” in Comunicaci (INIAP - Estación Pichilingue, Pichilingue, Ecuador).
Porcel M., Miranda T. C., Pisco‐Ortiz C., Camargo H., Moreno J., and Gutiérrez Y. (2024). A scarab beetle (Coleoptera: Scarabaeidae) causes tree mortality, delayed growth, and yield reduction in cacao with genotype-specific susceptibility to herbivory. Pest Manage. Sci. 80, 2179–2187. doi: 10.1002/ps.7957
Pramayudi N., Bakti D., and Tobing M. C. (2021). “The attack intensity of Paracoccus marginatus Williams & Granara de Willink (Hemiptera: Pseudococcidae) on papaya,” in IOP conf. Series: earth and environmental science, IOP Publishing, Banda Aceh, Indonesia, 951. doi: 10.1088/1755-1315/951/1/012033
Puig A., Wurzel S., Suarez S., Niogret J., and Marelli J. P.v (2021). Molecular identification of mealybug species (Hemiptera: pseudococcidae) affecting theobroma cacao for improved pest management. Proceedings 68, 10399. doi: 10.3390/iece-10399
R Core Team (2022). “R: A language and environment for statisticalcomputing,” in R foundation for statistical computing, (Vienna, Austria). Available online at: https://www.r-project.org.
Riera C. A., Paredes J. R., and Peralta E. L. (2013). “Situación actual y determinación de mecanismos de antixenosis sobre el chinche del cacao Monalonion dissimulatum Distant,” in Artículos de Tesis de Grado – FIMCP, Espol., 1–8. Available online at: https://www.dspace.espol.edu.ec/handle/123456789/52.
Sánchez-López S. and Sánchez-Soto S. (2019). “Identificación e incidencia de un membrácido (Hemiptera: Membracidae) y de hormigas asociadas (Hymenoptera: Formicidae), en cultivo de cacao en Tabasco,” in Revista nicaragüense de entomología, Leon, Nicaragua, vol. 64. , 1–17.
Sánchez-Mora F. D., Medina-Jara S. M., Díaz-Coronel G. T., Ramos-Remache R. A., Vera-Chang J. F., Vásquez-Morán V. F., et al. (2015). Potencial sanitario y productivo de 12 clones de cacao en Ecuador. Rev. Fitotecnia Mexicana 38, 265–274. Available online at: https://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S0187-73802015000300005 (Accessed April 4, 2025).
Santa Ana G. (2015). Actualización del plan de desarrollo y ordenamiento territorial de la parroquia Santa Ana. Diagnóstico. Available online at: https://santana.gob.ec/images/septiembre2018/plan_terriorial_santa_ana.pdf.
Sotomayor I. A., Tarqui Freire O. M., Loor Solorzano R. G., Amores Puyutaxi F. M., and Motamayor J. C. (2017). Variación fenotípica y selección de genotipos de cacao de alto rendimiento en Ecuador. EspamCiencia 8, 23–33.
Tarqui O. M., Sotomayor Cantos Casanova Mendoza I. A.T.J., Rodríguez Zamora G. A., Plaza Avellán L. F., and Zambrano Flores F. G. (2017). Selection of cocoa genotypes (Theobroma cacao L) resistant to witch broom disease (Moniliophthora perniciosa) in Los Ríos, Ecuador. Ciencia y Tecnología UTEQ 10, 17–26. doi: 10.18779/cyt.v10i1.197
Tepole-García R. E., Ramírez-Rojas S., Bartolo-Reyes J. C., and Castrejón-Gómez V. R. (2016). Ciclo de vida y análisis de riesgo climático de Leptoglossus zonatus Dallas (Hemiptera: Coreidae) para las zonas productoras de sorgo en el estado de Morelos. México Acta Zoológica Mexicana 32, 300–309. Available online at: https://www.scielo.org.mx/scielo.php?pid=S0065-17372016000300300&script=sci_arttext (Accessed April 4, 2025).
Thomas E., Zambrano F., and Loor R. (2024). Catálogo de cacaos de Ecuador (Quevedo, Ecuador: Bioversity International, MOCCA y el Instituto Nacional de Investigaciones Agropecuarias (INIAP).
Tscharntke T., Ocampo-Ariza C., Vansynghel J., Ivañez-Ballesteros B., Aycart P., Rodriguez L., et al. (2023). Socio-ecological benefits of fine-flavor cacao in its center of origin. Conserv. Lett. 16, 1–10. doi: 10.1111/conl.12936
Vargas A., Somarriba E., and Carballo M. (2005). Dinámica poblacional del chinche (Monalonion dissimulatum Dist.) y daño de mazorcas en plantaciones orgánicas de cacao del Alto Beni, Bolivia. Agroforestería en las Américas 43–44), 72–76. Available online at: http://www.sidalc.net/repdoc/A2335E/A2335E.PDF (Accessed April 4, 2025).
Walter N. T., Oluwaseun Adeleye Muthomi V. P.K., Ortiz Rojas R. J., Strzyzewski I., Funderburk J., and Martini X. (2018). Toxicity of different insecticides against two thrips (Thysanoptera: Thripidae) pests of concern in Central America. Florida Entomologist 101, 627–633. doi: 10.1653/024.101.0426
Wolcott K. A., Stanley E. L., Gutierrez O. A., Wuchty S., and Whitlock B. A. (2023). 3D pollination biology using micro-computed tomography and geometric morphometrics in Theobroma cacao. Appl. Plant Sci. 11, e11549. doi: 10.1002/aps3.11549
Keywords: antixenosis, population densities, intra-specific hybrids, resistance, susceptibility
Citation: Chirinos DT, Castro R, Sánchez-Mora FD, Castro-Olaya J, Mero Peñarrieta C, Morán P and García Cruzatty LC (2025) National versus Trinitarian cacao hybrids in Ecuador: which is less infested by sap pests and which is less damaged by pod pests. Front. Agron. 7:1588450. doi: 10.3389/fagro.2025.1588450
Received: 07 March 2025; Accepted: 14 July 2025;
Published: 01 September 2025.
Edited by:
Emmanouil Roditakis, Hellenic Mediterranean University, GreeceReviewed by:
Eustachio Tarasco, Faculty of Agricultural Science, University of Bari Aldo Moro, ItalyLuis F. Aristizabal, Consultant, Kailua-kona, Hawaii, United States
Copyright © 2025 Chirinos, Castro, Sánchez-Mora, Castro-Olaya, Mero Peñarrieta, Morán and García Cruzatty. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dorys T. Chirinos, ZG9yeXMuY2hpcmlub3NAdXRtLmVkdS5lYw==