 Joseph Mulema1*
Joseph Mulema1* Privat Ndayihanzamaso1,2Placide Birushe3Epitace Bizimungu4Eustache Cimpaye5Elie Gikoro3Gaspard Hakizimana3Egide Hatungimana4Arlène Ingabire2Antoine Irakiza2Marie Goreth Itangishaka4Michel Manirakiza6Lucien Masabarakiza4
Privat Ndayihanzamaso1,2Placide Birushe3Epitace Bizimungu4Eustache Cimpaye5Elie Gikoro3Gaspard Hakizimana3Egide Hatungimana4Arlène Ingabire2Antoine Irakiza2Marie Goreth Itangishaka4Michel Manirakiza6Lucien Masabarakiza4 Bonaventure Minani7Dismas Minani6Alexis Mpawenimana2Jacqueline Nahaniyo4Pascal Ndayiragije2Fidèle Ndayishemeza6Dieudonné Ndayishimiye8Déo Ndikumana9Schadrack Nduwimana4
Bonaventure Minani7Dismas Minani6Alexis Mpawenimana2Jacqueline Nahaniyo4Pascal Ndayiragije2Fidèle Ndayishemeza6Dieudonné Ndayishimiye8Déo Ndikumana9Schadrack Nduwimana4 Nicolas Niko2Evariste Nkubaye2Dismas Nshimirimana2Neilla Ntawuyankira2Eliakim Sakayoya10Celestin Niyongere1,2Daniel Karanja1Morris Akiri1Edward Onkendi1Ivan Rwomushana1Samson Musonerimana5,9Janny G. M. Vos11
Nicolas Niko2Evariste Nkubaye2Dismas Nshimirimana2Neilla Ntawuyankira2Eliakim Sakayoya10Celestin Niyongere1,2Daniel Karanja1Morris Akiri1Edward Onkendi1Ivan Rwomushana1Samson Musonerimana5,9Janny G. M. Vos11 Willis Ochilo1
Willis Ochilo1- 1CAB International, Nairobi, Kenya
- 2Institut des Sciences Agronomiques du Burundi (ISABU), Bujumbura, Burundi
- 3Office National de Contrôle et de Certification des Semences (ONCCS), Bujumbura, Burundi
- 4Direction de la Protection des Végétaux du Burundi (DPV), Gitega, Burundi
- 5L’Autorité Burundaise de Régulation des Produits Vétérinaires, des Pesticides et des Aliments (ABREVPA), Bujumbura, Burundi
- 6Direction de Vulgarisation, Formation, Recherche et Développement (DVFRD), Bujumbura, Burundi
- 7Faculty of Agronomic and Veterinary Sciences, University of Ngozi, Ngozi, Burundi
- 8Department of Plant Science and Production at the Light University of Bujumbura, Bujumbura, Burundi
- 9Department of Crop Science and Production, University of Burundi, Bujumbura, Burundi
- 10Direction Générale de la Mobilisation pour l’Auto-développement et de la Vulgarisation Agricole (DGMAEVA), Bujumbura, Burundi
- 11CAB International, Leusden, Netherlands
The lack of information regarding potential invasions is one of the crucial factors contributing to the rise in pest incursions in sub-Saharan Africa, which was addressed through horizon scanning. Utilising the CABI Horizon Scanning Tool, 8,747 non-native pest species not previously recorded in Burundi were identified. The initial output was filtered to include only species with complete names, resulting in 8,541 species. This was further refined by selecting pests that affected major value chains in Burundi, leading to 1,803 species. Eight species were added from the analysis of the current list of regulated pests for Burundi, along with four soft rot species considered of phytosanitary concern, bringing the total to 1,815. An additional 104 vectors that transmit the assessed species and seven pathogenic organisms transmitted by those species were added, resulting in 1,926 species. The 1,926 pest species, including 416 classified as invasive, comprised 550 arthropods, 153 bacteria, 79 chromista, 518 fungi, 19 molluscs, 141 nematodes, nine protists, 16 viroids, and 437 viruses. The highest recorded overall risk score was 175, attributed to 41 species, while the lowest was 2, associated with 55 species. The majority (n = 1,030; 53%) of the assessed species could solely be introduced as contaminants, whereas the least (n = 347; 18%) were stowaways, and others (n = 435; 23%) could be introduced through multiple pathways. Using agreed-upon criteria for suggesting management actions, 58% (n = 1,123) of the pest species were deemed unsuitable for any action, 18% (n = 350) were recommended for detection surveillance, and 25% (n = 476) were considered for regulation. Additional actions included contingency planning, raising awareness about high-risk pests (risk communication), managing some species through industry involvement, and conducting research to inform phytosanitary measures. These horizon scanning results will be utilised to develop a Register of Priority Pests for Burundi for risk monitoring and to update the list of regulated pests to facilitate trade in Burundi.
Introduction
In recent years, the agricultural sector in sub-Saharan Africa (SSA) has faced significant challenges due to a variety of introduced non-native (alien or exotic) pests that have become invasive (Rwomushana et al., 2019; De Groote et al., 2020). This has led to considerable losses in crop yields and quality, as well as increased management costs (Paini et al., 2016; Kumar Rai and Singh, 2020; Ahmed et al., 2022). In an effort to address this problem, farmers indiscriminately use pest-management products, thereby indirectly threatening biological diversity, food safety and people’s well-being (Matowo et al., 2020; Atinkut Asmare et al., 2022). Non-native pests that become invasive also directly affect biological diversity and trade. Trade possibilities diminish when pests are regulated by trading partners (Campbell, 1929). Some of the invasive pests reported in SSA include the fall armyworm, Spodoptera frugiperda (De Groote et al., 2020); the golden apple snail, Pomacea canaliculata (Buddie et al., 2021); maize chlorotic mottle virus (Mahuku et al., 2015); the papaya mealybug, Paracoccus marginatus (Macharia et al., 2017); potato cyst nematodes, Globodera rostochiensis and G. pallida (Mburu et al., 2020); and the tomato leaf miner, Phthorimaea absoluta (synonym: Tuta absoluta) (Guimapi et al., 2016).
Global trade and international movement of plants, plant products and other regulated articles primarily facilitate the deliberate or accidental introduction of pests (Youm et al., 2011; Epanchin-Niell et al., 2021; Montgomery et al., 2023). Deliberate introductions involve species that are transported to a new area for a specific purpose, such as aesthetics, breeding, conservation, planting, pest management, and research (Hulme et al., 2008). These species include ornamental and horticultural plants, pets for breeding, biocontrol agents, and plants for conservation (Beckmann and Shine, 2009; Shine, 2010; Dechassa, 2020; Korda et al., 2023). Many invasive pests are unintentionally introduced as a secondary result of intentional transfer of another product, good, or service (Wabuyele et al., 2014). Most seed-borne or seed-transmitted pathogenic organisms, particularly bacteria, phytoplasma, viruses and viroids, are unintentionally introduced as contaminants in plant parts, plants intended for planting, and soil (Denancé and Grimault, 2022; Pagán, 2022; Franić et al., 2024; Wang et al., 2024). Other invasive pests can be transferred through natural mechanisms, such as those capable of flight, like arthropods, and by wind and global air currents, including spores (produced by certain fungi and oomycetes) and pollen (Aylor, 1990; Mims and Mims, 2004; Roper et al., 2010; Guimapi et al., 2016). Arthropods may also vector some pathogenic organisms, while pollen can further introduce viruses (Heck, 2018; Vilcinskas, 2019; Raffini et al., 2020; Wielkopolan et al., 2021; Wendimu and Gurmu, 2024).
The preponderance of introductions in SSA is mainly attributed to the porosity of borders, poor border biosecurity systems, country’s lack of capacity to stop or limit invasion, and the lack of reactive or timely response to pest invasions to prevent establishment (Early et al., 2016; Nagoshi et al., 2018; Graziosi et al., 2020). While it is unrealistic to expect countries in SSA to impede new incursions, collating information about the likely introductions is key for preparedness, prevention, early detection, and rapid response (Kenis et al., 2022; Mulema et al., 2022, 2024). Prevention remains the least costly, most cost-effective, efficient, and practical option for managing invasive species. Once an invasive species is established, containment and eradication become extremely difficult, given the nature of cropping systems and poor enforcement of regulations in SSA (Kueffer and Loope, 2009; Hulme et al., 2023; Roy et al., 2023). As a result, most invasive species naturalise and become part of ecosystems, as has been observed with S. frugiperda and P. absoluta, leaving management to limit effects on crop production as the only option (Matova et al., 2020; Desneux et al., 2022; Kumar et al., 2022).
Gathering information about high-risk invasive species is vital for developing strategies to limit incursions (Gallardo and Aldridge, 2013; Matthews et al., 2017; Saravanakumar et al., 2023). In this study, we employed horizon scanning to identify and prioritise invasive species not reported in Burundi but observed in other countries (globally) from which Burundi may source commodities. The horizon scanning method has been employed in numerous studies to prioritise invasive alien pests and plant species (Weber and Gut, 2004; Sutherland et al., 2008; Gassó et al., 2009; Roy et al., 2014, 2019; Gallardo et al., 2016; Bayón and Vilà, 2019; Kenis et al., 2022; Mulema et al., 2022). Horizon scanning involves a systematic search for potential biological invasions and an assessment of their possible impacts on the economy, society, and environment while considering potential opportunities to mitigate these impacts (Sutherland et al., 2008, 2010, 2020; Roy et al., 2014). The information generated from horizon scanning can be made available to risk managers and policymakers to support the planning and management of invasive species (Sutherland et al., 2010, 2020; Caffrey et al., 2014; Matthews et al., 2017).
Horizon scanning to prioritise invasive species has previously been conducted through an extensive literature search by a panel of experts who compile information on species reported as invasive in various geographical locations (Bayón and Vilà, 2019; Seymour et al., 2020; Dawson et al., 2023; Lieurance et al., 2023). The agreed list of species is subsequently subjected to risk assessment to determine which may require action either in the medium or long term.
To support decision-making regarding invasive species and identify potential risks in countries, provinces, and states, CABI has developed a Horizon Scanning Tool. This tool is accessible through the CABI Compendium, which is part of the CABI Digital Library. The Horizon Scanning Tool utilises CABI data to generate a list of species that are reported as not present (known not to occur) in a selected “area at risk” but are reported from “source areas”, which are geographic regions with similar climates, neighbouring territories, or selected trading partners. It is important to note that the lack of a report indicating presence does not confirm absence, as this may result from gaps in reporting. The Horizon Scanning Tool has previously been used to prioritise pests in Ghana (Kenis et al., 2022), Kenya (Mulema et al., 2022), the United States (Kendig et al., 2022; Lieurance et al., 2023), and Zambia (Mulema et al., 2024).
Materials and methods
Assessment team
A team of 29 subject matter experts (SMEs) conducted the assessment. The team was convened from regulatory agencies (L’Autorité Burundaise de Régulation des Produits Vétérinaires, des Pesticides et des Aliments (ABREVPA), Direction Générale de la Mobilisation pour l’Auto-développement et de la Vulgarisation Agricole (DGAEVMA), Direction de la Protection des Végétaux du Burundi (DPV), and Office National de Contrôle et de Certification des Semences (ONCCS); national (Institut des Sciences Agronomiques du Burundi (ISABU) and international agricultural research institutions (CABI); academia (Université du Burundi, Université Lumière de Bujumbura, and Université de Ngozi); and extension institutions (Direction de Vulgarisation, Formation, Recherche et Développement (DVFRD). The SMEs had experience in bacteriology, entomology, mycology, nematology, and virology.
Selection of non-native plant pests
The assessment was carried out across two workshops. The first workshop, held from 17 to 26 April 2023, focused on training the team in horizon scanning, utilising the CABI Horizon Scanning Tool, and conducting rapid risk assessments in accordance with the guidelines outlined in Supplementary File S1. The premium version of the Horizon Scanning Tool was used to create a preliminary list of plant pests that had not yet been reported as present in Burundi. The parameters set in the Horizon Scanning Tool involved determining the area at risk (Burundi) and selecting regions from which non-native pest species were likely to be introduced (source areas). The source areas included all geographical regions in Africa, Asia, Europe, North America, Oceania, and South America. All parameters except for the type of organism were left open. The considered pest organisms consisted of arthropods, bacteria (including phytoplasma), chromista, fungi, molluscs, protists, plant parasitic nematodes, and viruses (including viroids). Following the scan, the list was downloaded in Excel (.xlsx) format for further analysis. The current list of regulated pests for Burundi was also examined to determine if there were pests not included in the horizon scan output that could be considered for rapid risk assessment. The assessment conducted in the first workshop was validated in the second, held from 25 to 29 November 2024.
Description of the scoring system
The risk-scoring guidelines employed were based on the framework outlined by Roy et al. (2019) but have been modified by Kenis et al. (2022) and Mulema et al. (2022). Roy et al. (2019) evaluated the likelihood of introduction (entry), establishment, spread, and potential negative impacts on biodiversity and ecosystem services; however, this study examined the possibility of introduction (entry), establishment (including aspects of spread), and potential socio-economic and environmental (biodiversity) impacts. A five-point scoring system outlined in Supplementary File S1 was utilised for the four parameters (entry, establishment, socio-economic impact, and biodiversity impact).
Likelihood of entry
A score of one indicated that the organism was absent from Africa and unlikely to be found in an imported commodity; two indicated absence from Africa (the pest is not known to occur in Africa) but likely to be infrequently imported on a commodity; three was ascribed to three scenarios: either present in Africa (excluding neighbouring countries to Burundi) and spreading slowly; or absent from Africa but demonstrated to spread very quickly across several continents, or often associated with a commodity that is commonly imported, or frequently intercepted in Burundi; four, indicated presence in Africa (excluding neighbouring countries) with rapid spread, or presence in a neighbouring country with slow spread; and five, indicated presence in a neighbouring country (the Democratic Republic of the Congo (DR Congo), Rwanda, and Tanzania) with rapid spreads. A pest could have been present in Africa but absent from Burundi, suggesting that it was reported elsewhere on the African continent but not yet known to occur in Burundi. The global pest distribution data were obtained from the CABI Compendium, the European and Mediterranean Plant Protection Organisation (EPPO) Global Database, and the Global Biodiversity Information Facility (GBIF).
Pathway for introduction
Hulme et al. (2008) detailed the three mechanism and their respective pathways through which an alien species may enter new geographical or political areas. These mechanisms include the importation of a commodity (via three pathways: contaminant, escape, and release), the arrival of a transport vector (through one pathway: stowaway), and natural spread from a neighbouring region (via two pathways: corridor and unaided). The contaminant pathway involves organisms unintentionally introduced with a specific commodity, including obligate pathogens, parasites, and commensals of traded plants and animals. The escape pathway comprises organisms that are intentionally introduced but unintentionally escape, including feral crops and livestock, pets, garden plants, and live baits. The release pathway primarily involves organisms introduced deliberately for release, including biocontrol agents, game animals, and landscape plants. Only the contaminant pathway was considered because comprehensive risk assessments are typically conducted for organisms deliberately introduced into countries or regions.
The stowaway pathway, also considered in this assessment, involves organisms that are unintentionally introduced because they are attached to or within a transport vector, such as molluscs. The corridor pathway involves organisms that are unintentionally introduced as a result of human infrastructure development, linking previously unconnected regions. This pathway was not considered because it does not apply to the majority of pests. Lastly, the unaided pathway comprises organisms that are introduced through natural dispersal across political borders. Examples include flying insects and pollen, spores, and soil infested with pathogenic microorganisms that wind currents may carry. Therefore, the contaminant (CO), stowaway (ST), and unaided (UN) were considered for this study because they are more relevant to the introduction and spread of pests.
Likelihood of establishment
A score of one indicated that Burundi was climatically unsuitable or that host plants were absent; two, that only a few areas in Burundi were climatically suitable, or host plants were rare; three, that large regions of Burundi were climatically suitable, whilst host plants were rare; or that only a few areas in Burundi were climatically suitable, but host plants were at least moderately abundant; four, that large regions of Burundi were climatically suitable, but host plants were moderately abundant; and five, that large areas in Burundi were climatically suitable, whilst host plants were very abundant. To ensure consistency in assessing the capacity for the establishment, the SMEs weighted the likely hosts to be grown in Burundi (Supplementary File S2).
Assessment of potential socio-economic impact
A score of one indicated that the alien species did not attack plants that were cultivated or utilised; a score of two signified that the alien species damaged plants that were only occasionally cultivated or utilised; a score of three denoted that the alien species damaged plants that were regularly cultivated or utilised, but without threatening the cultivation, utilisation, or trade of this crop; a score of four meant that the alien species had the potential to threaten, at least locally, the cultivation of a plant that was regularly cultivated or utilised, or to periodically attack a crop that was key to the economy of Burundi without posing a threat to the latter; and a score of five indicated that the alien species had the potential to threaten, at least locally, a crop that was essential to the economy of Burundi.
Assessment of the potential impact on biodiversity
A score of one indicated that the alien species would not affect any native species; two meant the alien species would impact individuals of a native species without affecting its population level; three signified that the alien species had the potential to reduce the population levels of a native species; four implied that the alien species could eradicate a native species locally or affect populations of a protected or keystone species; and five denoted that the alien species possessed the potential to completely eliminate a native species or to locally eradicate a keystone species. Native species in this context include animals, plants, and other organisms, including pathogens.
Scoring of the prioritised species
All assessments were conducted in plenary by all SMEs. The scores for each species attribute were thoroughly discussed, and any discrepancies were resolved until a consensus was reached before the final score was confirmed. As detailed in Blackburn et al. (2014), a low, medium, and high confidence rating was provided for each of the four assessed attributes, the likely pathway of arrival, and the overall risk score. The vectors and vectored pathogenic organisms of prioritised pests were also evaluated.
Determining risk
Consequently, the overall risk score was calculated using the following formula, resulting in scores ranging from 250 to two.
Actions for management
An overall score of 54 was deemed the threshold for recommending a management action. This score was chosen because it is only achievable for species with a score of three for all assessable attributes or those exceeding three in at least three parameters. A score of three or more suggests a higher likelihood of entry, establishment, and significant impact (socio-economic or biodiversity). Scores below three are regarded as low risk as they indicate a reduced likelihood of entry, establishment, and impact; a score of three is classified as moderate, while scores above three (four and five) signify an increased likelihood of entry, establishment, and impact (contrasting with the lower scores of one and two). A no-action approach was recommended for species that recorded an overall risk score below 54. In contrast, a management action was proposed for all pests that recorded an overall risk score above 54. The suggested actions included detection surveillance for pests reported in a neighbouring country to establish pest status in Burundi, regulation supported by Pest Risk Analysis (PRA), contingency planning for preparedness, raising awareness, management by industry (relevant value chain system), and research.
Detection surveillance was also considered if the assessment team confirmed a previous interception or if there was a pest frequently affecting a host commodity imported from a country where the pest is endemic. Pest Risk Analysis is essential in providing guidelines for managing pathways to limit the introduction of quarantine pests. However, action was deemed necessary for certain species that recorded an overall risk score below 54. These are species currently regarded by trading partners as high-risk A1 or A2 quarantine pests, such as subspecies of Xylella fastidiosa. While a no-action recommendation would be made for some species that recorded an overall risk score above 54, the final decision on regulation or deregulation was left to the National Plant Protection Organisation (NPPO). This was for species whose primary pathway of introduction was deemed implausible. This category included pests transmitted by vectors rather than through seed (Seed-contaminated or seed-transmitted), yet the vectors were unlikely to be introduced alongside a viable pathogenic organism. Others in this category included soil-borne pests that are unlikely to accompany the commodity. A prime example would be soil-borne fungi that are not seed-transmitted. Furthermore, a no-action recommendation was also suggested when insufficient information was available for the assessment of a particular species or vectors of assessed species, as well as for pests vectored by assessed species already reported in Burundi.
Results
Evaluation of the horizon scanning output
A three-stage approach was utilised to develop the final list of non-native plant pest species that could impact crop and forest productivity if introduced to Burundi. Firstly, a global horizon scan identified 8,747 species that have not yet been reported as existing in Burundi. These species included 5,647 arthropods, 311 bacteria (including 54 phytoplasma), 171 chromista (oomycetes), 1,719 fungi, 27 molluscs, 206 nematodes, nine protists, 12 viroids, and 645 viruses. The initial output from the Horizon Scanning Tool was subsequently filtered to include only species with complete names, resulting in 8,541 species, which comprised 5,517 arthropods, 296 bacteria (including 54 phytoplasmas), 165 chromista, 1,678 fungi, 27 molluscs, 192 nematodes, nine protists, 28 viroids, and 629 viruses (Supplementary File S3). Of the 8,541 species, 641 had been reported as invasive. These 641 species consisted of 397 arthropods, 41 bacteria (including 14 phytoplasma), 15 chromista (oomycetes), 94 fungi, 11 molluscs, 38 nematodes, two viroids, and 43 viruses. This data is detailed in Supplementary File S2 (Sheet: All Horizon Scanning Data). The list of 8,541 species was further refined by selecting those that impacted major value chains cultivated in Burundi, resulting in 1,801 species.
Analysis of the current list of regulated pests
The current list of regulated pests for Burundi was developed in 2006, and since then, the status of some pests has changed. The list comprises 123 species, 93 of which were part of the horizon scanning output, except for 30 (27 of which had already been reported in Burundi). Of the 123 species, 110 are specific, while the remaining 13 are represented as general species, including Anastrepha spp., Ceratitis spp., Chilo spp., Cuscuta spp., Cylas spp., Dacus spp., Ditylenchus spp., Erwinia spp., Heterodora spp., Lyriomyza spp., Meloidogyne spp., Sclerospora spp., and Striga spp. Two of the remaining 110 species, Corynebacterium pisi and Elsinoe piperis, could not be determined, while for one species, Bostrynchopsis villosula, the available information was insufficient for assessment, leaving only 107 species. The 107 species included 23 not yet reported in Africa and 84 that have already been reported in Africa.
The 84 species documented in Africa included 24 species already recorded in Burundi. The remaining 60 species were reported in a neighbouring country. A total of 78 species were part of the horizon scanning output. The 78 species included 19 not reported in Africa, 59 reported in Africa, of which 27 were reported in a neighbouring country. Additionally, a number of species with the genera Anastrepha, Ceratitis, Chilo, Cylas, Dacus, Ditylenchus, Erwinia, Heterodora, Lyriomyza, Meloidogyne, and Sclerospora were also part of the output. However, eight species, including Allantophomopsiella pseudotsugae, Coffee blister spot agent, Helicobasidium brebissonii, Nilapavarta lugens, Popilia japonica, Robbsia andropogonis, Ustilaginoidella oedipigera, and Wheat striate mosaic virus were not and have been considered for rapid risk assessment. All data have been consolidated in Supplementary File S3.
Evaluation of vectors and vectored species
A total of 1,801 species from the horizon scanning output were assessed to identify those capable of transmitting or being transmitted by other species. The species that could act as vectors or be vectored but were not included in the horizon scanning output were also evaluated to determine their associated risks. The species capable of vectoring the assessed species numbered 105, while those that could be vectored numbered eight. The 105 species included 99 arthropods, one fungus (Olpidium bornovanus), four plants, all Cuscuta species (C. campestris, C. ceanothi, C. europaea, and C. reflexa), and one nematode (Paratrichodorus anemones). The arthropods belonged to the orders Acarida (one), Coleoptera (five), Hemiptera (88), Hymenoptera (four), and Thysanoptera (one). Most of the Hemiptera species were from the families Cicadellidae (54) and Aphididae (19). The eight species that could be vectored included two fungi (Microbotryum succisae and Neocosmospora euwallaceae), one protist (Gregarina sitophili), and five viruses (millet red leaf persistent luteovirus, raspberry leaf spot virus, watermelon mosaic potyvirus-II, wheat striate mosaic virus, and wheat yellow mosaic virus).
Inclusion of other soft rot Pectobacteriaceae
The Soft Rot Pectobacteriaceae, formerly known as the Soft Rot Enterobacteriaceae, includes two genera; Dickeya and Pectobacterium, both of which are part of the bacterial family Pectobacteriaceae. These two species cause highly destructive diseases in many value chains, particularly in the Solanaceae and Poaceae families. The horizon scanning analysis identified eight species of Dickeya and nine species of Pectobacterium. Six of the Dickeya species (D. chrysanthemi, D. dadantii, D. dianthicola, D. fangzhongdai, D. paradisiaca, and D. solani) affect potatoes, while one of each affects maize (D. zeae) and rice (D. oryzae). Although D. zeae was initially considered a single species, studies by Hugouvieux-Cotte-Pattat and Van Gijsegem (2021) demonstrated diversity among D. zeae strains, leading to two distinct clades, one of which was later elevated to D. oryzae. Seven of the nine identified species of Pectobacterium are known to affect potatoes. They include P. aroidearum, P. atrosepticum, P. betavasculorum, P. brasiliense, P. carotovorum, P. parmentieri and P. polaris. The previous species P. carotovorum subsp. carotovorum has been elevated to a number of species, including P. carotovorum, P. parvum, and P. peruviense (Waleron et al., 2018; Portier et al., 2019; Pasanen et al., 2020). Pectobacterium punjabense, recently isolated from potatoes, is a new species closely related to P. parmentieri (Sarfraz et al., 2018). Therefore, the inclusion of the species D. oryzae, P. parvum, P. peruviense and P. punjabense was guided by their close relationship to the species (D. zeae, P. carotovorum, and P. parmentieri) identified through horizon scanning.
Final list of pests suggested for rapid risk assessment
The total number of species suggested for rapid risk assessment was 1,924. This total included 1,803 species generated through horizon scanning. Eight species from the current list of regulated pests for Burundi, which were not part of the horizon scanning output, were added, bringing the total to 1,811. Additionally, four species from the genera Dickeya (D. oryzae) and Pectobacterium (P. parvum, P. peruviense, and P. punjabense) were included, as they were not part of the original horizon scanning output but were deemed essential due to their phytosanitary consequences, raising the number to 1,815. Furthermore, 104 vectors that transmit the assessed species and five pathogenic organisms transmitted by the assessed species were added, completing the total number of prioritised species for assessment. The 1,924 species included 416, which accounted for approximately 22%. More than half (n=992, 52%) of these species had already been reported in Africa, with about one-third of the 967 (n=356, 36%) documented in a neighbouring country. The 1,924 species comprised 550 arthropods, 153 bacteria, 79 chromista, 518 fungi, 19 molluscs, 141 nematodes, nine protists, 16 viroids, and 435 viruses. Of the 416 invasive species, there are 250 arthropods, 25 bacteria (including six phytoplasmas), seven chromista, 61 fungi, nine molluscs, 34 nematodes, one viroid, and 29 viruses. All data have been consolidated in Supplementary File S2 (Sheet: Assessment of prioritised pests).
Assessment of arthropods
The 550 arthropods prioritised for rapid risk assessment included 250 species (45%) recorded as invasive. Of these, 237 (43%) had been reported in Africa, and of that proportion, 96 (41%, n=237) were found in a neighbouring country (Table 1). Among the 250 species listed as invasive, 101 were reported outside Africa, while 150 were reported within Africa, including 70 in a neighbouring country. The highest overall risk score was 175, attributed to 23 species, whereas the lowest was two, noted for 28 species. Six of the species, namely Dieuches humilis, Nysius euphorbiae, Phyllocoptes arcani, Unkanodes albifascia, Unkanodes sapporonus, and Uroleucon escalantii, could not be assessed due to lack of enough sufficient information. Most arthropods were likely to be introduced solely as contaminants (84%, n=461) (Table 2). The remaining 18% could be introduced through various pathways, with 5% (n=30) entering as contaminants or stowaways and 10% (n=53) as contaminants, stowaways, or unaided (Table 2). The remaining six species not assessed accounted for 1%. The 450 assessed species also included 313 (57%) known vectors. All data have been consolidated in Supplementary File S4.
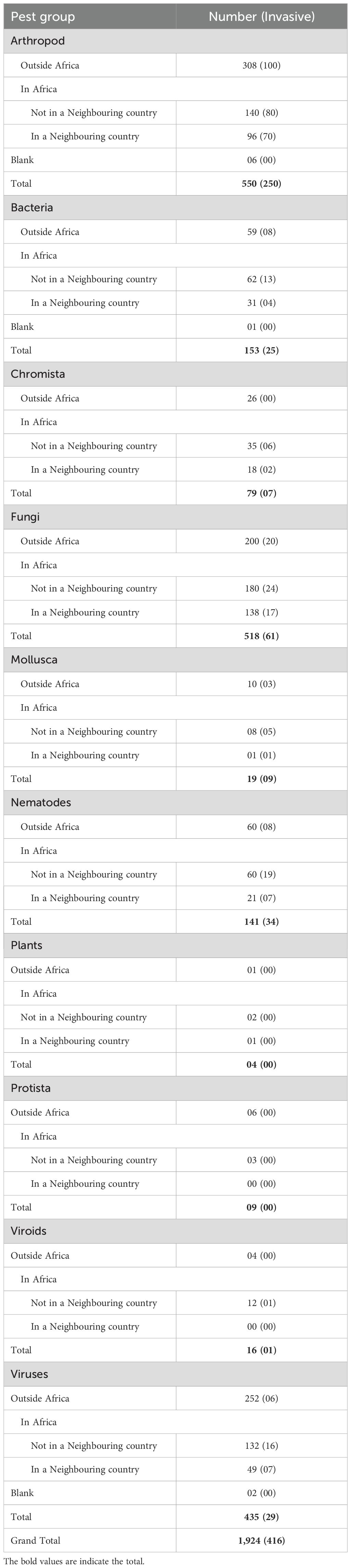
Table 1. Pest groups identified by the Horizon Scanning Tool.
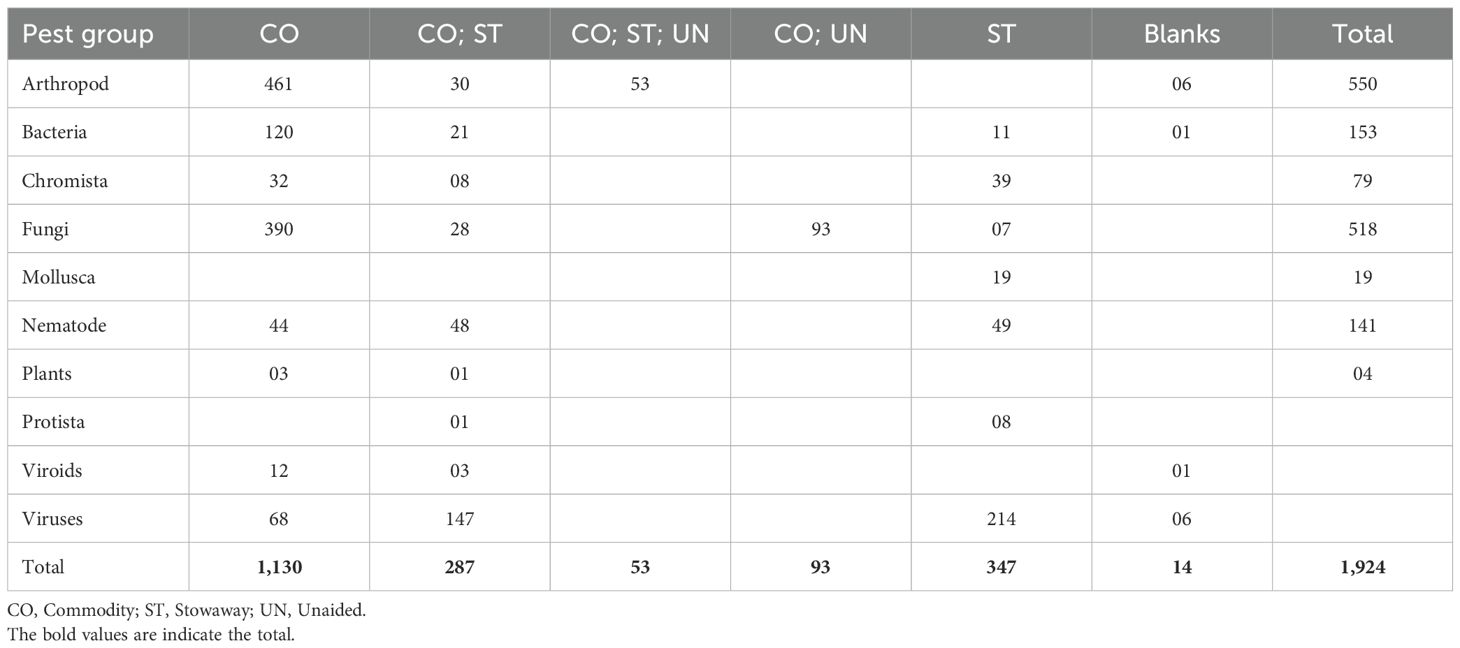
Table 2. Likely pathways of arrival for assessed pests.
Assessment of bacteria
The 153 bacteria species prioritised for assessment included 25 (16%) already documented as invasive. Of these, 61% (n=93) were reported in Africa, with 33% (n=31) reported in a neighbouring country (Table 1). The highest overall risk score was 140, recorded for 18 species, while the lowest was two, recorded for four species: Candidatus Phytoplasma rubi, Pseudomonas syringae pv. atropurpurea, Xanthomonas axonopodis pv. betlicola, and Xanthomonas translucens pv. arrhenatheri. One species (sesame phyllody phytoplasma) was not assessed due to a lack of sufficient information. The majority of assessed bacteria (78%, n=120) were likely introduced solely as contaminants, while the fewest (7%, n=11) could be introduced only as stowaways. The remaining 14% (n=21) could enter through either pathway as contaminants or stowaways (Table 2). The pathway of introduction could not be determined for one species (Sesame phyllody phytoplasma), as indicated above. Thirty-two (21%) of the assessed species were vectored, of which all were phytoplasmas, except for Candidatus Liberibacter americanus, Candidatus Liberibacter asiaticus, Candidatus Liberibacter solanacearum, Dickeya zeae, Pantoea ananatis, Pantoea stewartii subsp. stewartii, Rathayibacter rathayi, Rathayibacter tritici, Spiroplasma citri, Spiroplasma kunkelii, Xylella fastidiosa subsp. fastidiosa (Xff), Xylella fastidiosa subsp. multiplex (Xfm), and Xylella fastidiosa subsp. pauca (Xfp). All data have been consolidated in Supplementary File S5.
Assessment of chromista
The 79 assessed Chromista (oomycetes) included seven species noted as invasive. Most (67%, n=53) of the species had been reported in Africa, with 18 of these (34%, n=53) documented in a neighbouring country (Table 1). The highest overall risk score was 175, recorded for Peronosclerospora maydis and Phytophthora nicotianae, while the lowest was six, recorded for Peronospora radii, Phytophthora aleatoria, Phytophthora pinifolia, and Pythium sulcatum. Half (49%, n=39) of the species were likely to be introduced solely as stowaways, while 41% (n=32) were likely to be introduced as contaminants. The remaining 10% (n=8) could potentially be introduced through multiple pathways, specifically as contaminants or stowaways (Table 2). All data have been consolidated in Supplementary File S6.
Assessment of fungi
The 518 fungi prioritised for assessment included 61 (12%) documented as invasive. A proportion of 61% (n=318) had been reported in Africa, of which 43% (n=138) were recorded in a neighbouring country (Table 1). The majority of assessed fungi (75%, n=390) were likely introduced exclusively as contaminants, while the fewest (1%, n=7) could be introduced solely as stowaways. The remaining 24% (n=121) could be introduced through multiple pathways (Table 2). This group includes 28 (5%) that could be introduced as both contaminants and stowaways and 93 (18%) that could be both contaminants and unaided. The highest overall risk score was 175, recorded for Lasiodiplodia theobromae, Phakopsora pachyrhizi, Puccinia polysora, Uromyces appendiculatus, and Ustilago maydis, while the lowest was two, recorded for seven species including Erysiphe biocellata, Exobasidium vaccinii, Myrothecium verrucaria, Passalora sequoiae, Phomopsis vaccinii, Sirococcus clavigignenti-juglandacearum, and Tilletia lolii. Nine of the assessed fungal species, which include Albonectria rigidiuscula, Amylostereum areolatum, Fusarium oxysporum f.sp. cubense Tropical Race 4 (Foc TR4), Haematonectria haematococca, Leptographium procerum, Microbotryum succisae, Microbotryum violaceum, Neocosmospora euwallaceae, and Raffaelea lauricola, were likely to be disseminated by vectors. All data have been consolidated in Supplementary File S7.
Assessment of molluscs
The 19 molluscs prioritised for assessment included nine species documented as invasive, representing 47% (Table 1). Nine (47%) of the assessed molluscs had been reported in Africa, but only one had been documented in a neighbouring country (Tanzania) (Table 1). Species recorded in Africa included Achatina fulica, Arion hortensis Ferussac, Bradybaena similaris, Cornu aspersum, Deroceras laeve, Deroceras reticulatum, Limax maximus, Pomacea canaliculata, and Rumina decollata with P. canaliculata also reported in Kenya. The highest overall risk score was 120, recorded for Achatina fulica, while the lowest was six, recorded for Meghimatium pictum. All the assessed molluscs could be introduced exclusively as stowaways (Table 2). All data have been consolidated in Supplementary File S8.
Assessment of nematodes
The 141 prioritised plant-parasitic nematodes included 34 (24%) species identified as invasive. Of these, 57% (n=81) were reported in Africa, with 27% (n=21) documented in a neighbouring country (Table 1). The highest overall risk score was 140, recorded for 13 species, while the lowest was 2, noted for two species: Longidorus macrosoma and Trophotylenchulus piperis. A proportion of 31% (n=44) was likely to be introduced exclusively as contaminants, while 35% (n=49) was likely introduced as stowaways. The remaining 34% (n=48) could be introduced either as contaminants or stowaways. Four nematodes, Bursaphelenchus fungivorus, Bursaphelenchus mucronatus, Bursaphelenchus xylophilus, and Rhadinaphelenchus cocophilus, could be vectored by arthropods, while 17 were known vectors for bacteria and viruses. All data have been consolidated in Supplementary File S9.
Assessment of parasitic plants
The four evaluated Cuscuta species (C. campestris, C. ceanothi, C. europaea, and C. reflexa) were not part of the horizon scanning output but were included for assessment due to their role as vectors for the species being evaluated. With the exception of C. ceanothi, the remaining three Cuscuta species were reported in Africa, and only C. campestris was found in a neighbouring country, Tanzania. All were likely to be introduced as contaminants, except for C. campestris, which could be introduced as either a contaminant or stowaway. The highest overall risk score recorded was 100 recorded for C. campestris, while the lowest was 35, attributed to Cuscuta ceanothi. All data have been consolidated in Supplementary File S10.
Assessment of protists
The nine protists prioritised for assessment included three species, Plasmodiophora brassicae, Polymyxa betae, and Polymyxa graminis, all reported in Africa, with none observed in a neighbouring country (Table 1). The highest overall risk score was 105, recorded for Polymyxa graminis, while the lowest was 12, recorded for two species, Physarum cinereum and Phytomonas ananas. Eight of the assessed protists could be introduced exclusively as stowaways, whereas only Ligniera vasculorum could be introduced as either a contaminant or stowaway (Table 2). Two protists, Polymyxa betae and Polymyxa graminis, are known vectors of viruses. Polymyxa betae is known to vector beet necrotic yellow vein virus, while Polymyxa graminis vectors soilborne wheat mosaic virus, wheat spindle streak mosaic virus, wheat streak mosaic virus, and wheat yellow mosaic virus. Additionally, three protists, Gregarina sitophili, Phytomonas leptovasorum and Phytomonas staheli can spread by arthropod vectors. All data have been consolidated in Supplementary File S11.
Assessment of viroids
The 16 viroids prioritised for assessment included only one viroid, the potato spindle tuber viroid, identified as invasive. Twelve viroids, accounting for 75% of the total, were reported in Africa, with none recorded in a neighbouring country. The majority (75%, n=12) of the viroids were likely introduced as contaminants, while the remaining 19% (n=3) were introduced either as contaminants or stowaways. For one viroid, the pathway could not be determined based on the available information. The highest overall risk score was 140, recorded for potato spindle tuber viroid and tomato apical stunt viroid, while the lowest was three, recorded for one species (coconut cadang-cadang viroid). All data have been consolidated in Supplementary File S12.
Assessment of viruses
The 435 virus species prioritised for assessment included 29 identified as invasive. Forty-two percent of the total assessed viruses (n=181) were reported in Africa, along with 49 reported in a neighbouring country (Table 1). Most of the viruses were likely introduced exclusively as stowaways (49%, n=214), while the least were contaminants (16%, n=68). The remaining 34% (n=147) could have been introduced as either contaminants, stowaways, or both. Due to insufficient information, the potential pathways for six viruses- including cowpea mosaic virus, millet red leaf persistent luteovirus, okra yellow vein mosaic virus, pepper chlorotic spot virus, pigeon pea yellow mosaic virus, and wheat yellow mosaic virus- could not be determined. The highest overall risk score recorded was 175, noted for 11 viruses, while the lowest was two, noted for 14 viruses. A proportion of 84% (n=366) of the viruses are transmitted by known vectors, while 21 are expected to be vectorised; however, the specific vectors are yet to be identified. All data have been consolidated in Supplementary File S13.
Assessment of vector species
The total number of known vector species assessed was 354. These included 328 arthropods, three fungi, 12 nematodes, four plants, and three protists. Of the 350 species, 26 were not assessed because they had already been reported as present in Burundi, while an additional six, including Dieuches humilis, Nysius euphorbiae, Phyllocoptes arcani, Unkanodes albifascia, Unkanodes sapporonus, and Uroleucon escalantii, were not assessed due to insufficient information to conduct the assessment leaving 317. The 317 species comprised 296 arthropods (33%), three fungi, 12 nematodes, four plants, and two protists. The fungi included Olpidium bornovanus, Olpidium brassicae, and Olpidium radicale; the plants included Cuscuta. campestris, Cuscuta ceanothi, Cuscuta europaea, and Cuscuta reflexa; and the protists included Polymyxa betae and Polymyxa graminis. The nematodes belonged to the genera Longidorus, Paratrichodorus, Trichodorus, and Xiphinema. Most of the arthropods were Hemiptera, specifically from the family Cicadellidae, followed by the Aphididae. The highest overall risk score recorded was 175 for eight species, which included Apis mellifera, Bemisia afer, Bemisia tabaci, Chilo patellus, Cicadulina mbila, Frankliniella schultzei, Peregrinus maidis, and Thrips tabaci. The lowest overall risk score recorded was two for 28 species. All data have been consolidated in Supplementary File S14 (Sheet: Vector species assessment).
Assessment of vectored organisms
The total number of vectored species was 215, comprising 28 bacteria (including phytoplasma), nine fungi, five nematodes, one protist, and 172 viruses. Two species, Ralstonia solanacearum and tobacco rattle virus, were not assessed as they were already present in Burundi. Additionally, three species, millet red leaf persistent luteovirus, sesame phyllody phytoplasma, and wheat yellow mosaic virus- were excluded from the assessment due to insufficient information. One species, Angiostrongylus cantonensis, was also not assessed because it affects humans rather than crops. The remaining 210 species included 109 (52%) reported as present in Africa, of which 24 were noted in a neighbouring country. The majority of the vectors for the assessed species were arthropods, accounting for 90% (n=195). However, other vectors included nematodes from the genera Anguina (A. agrostis and A. tritici), Longidorus (L. attenuatus and L. elongatus), Paratrichodorus (P. anemones, P. minor, P. pachydermus, P. porosus, and P. teres), Trichodorus (T. primitivus, T. similis, and T. viruliferus), and Xiphinema (X. americanum, X. diversicaudatum, and X. rivesi); fungi from the genus Euwallacea (E. fornicatus and E. perbrevis); molluscs from the genus Pomacea (P. canaliculata); and protists in the genus Polymyxa (P. betae and P. graminis). The highest overall risk score was 175, reported for six species (alfalfa mosaic virus, bean yellow mosaic virus, maize mosaic virus, potato virus A, potato virus M, and potato virus Y), while the lowest was two, recorded for eight species, all viruses, including black raspberry necrosis virus, blueberry leaf mottle virus, blueberry shoestring virus, chickpea chlorotic dwarf Syria virus, parsnip mosaic virus, raspberry leaf mottle virus, raspberry leaf spot virus, and ryegrass mosaic virus. All data have been consolidated in Supplementary File S14 (Sheet: Vectored species assessment).
Actions for management
No-action
Of the 1,926 species assessed, 1,123, accounting for 58% of the total, were not considered for any action, although the risk associated with certain pests could be monitored. This category included pests that received an overall risk score below 54. The 1,123 pests consisted of 341 arthropods, 66 bacteria, 40 Chromista, 290 fungi, 12 molluscs, 78 nematodes, seven protists, four viroids, and 292 viruses (Table 3). This group also included 60 species that recorded an overall risk score above 54. All data have been consolidated in Supplementary File S15 (Sheet: No action).
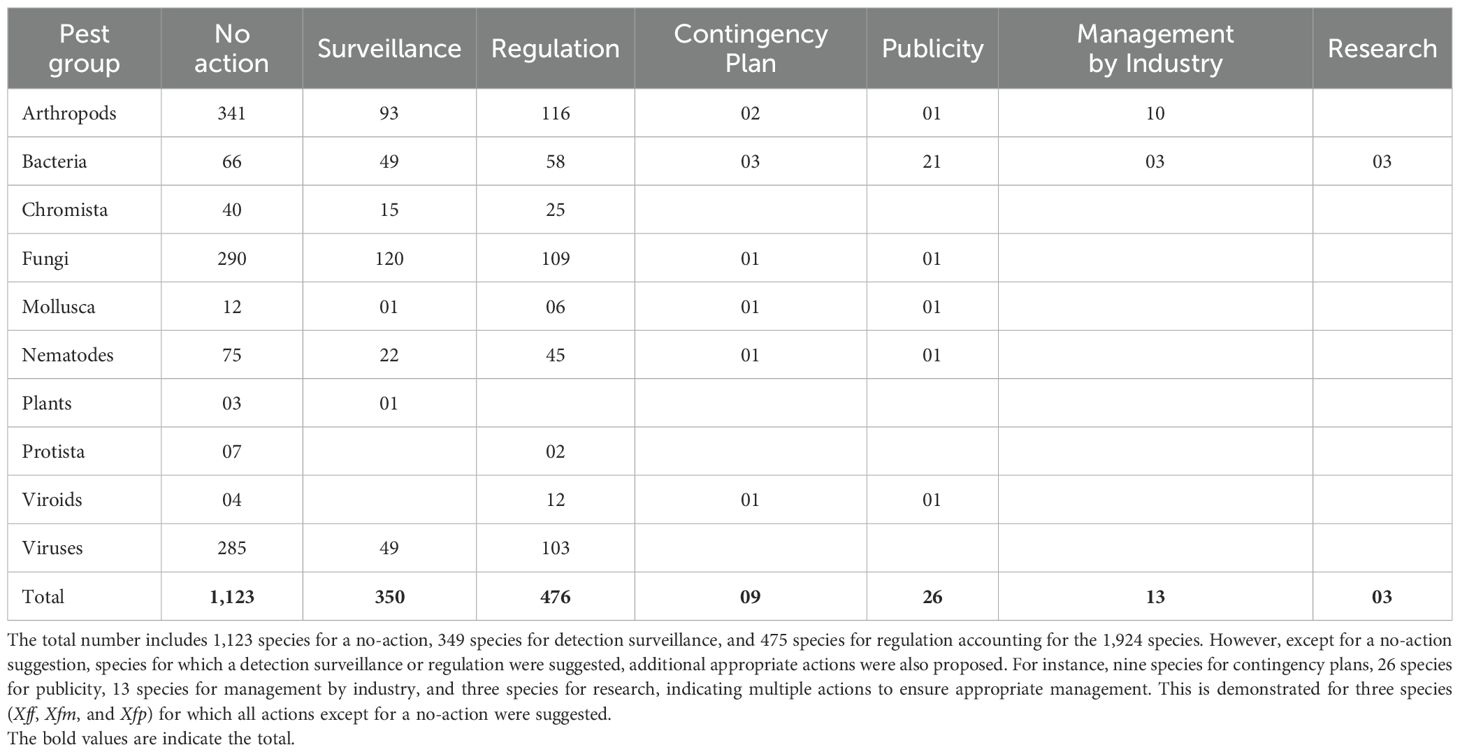
Table 3. Actions suggested for assessed pests.
Target surveillance
Detection surveillance was proposed for 350 species, representing 18% of the total assessed species. The 350 pests included 93 arthropods, 49 bacteria, 15 Chromista, 120 fungi, one mollusc, 22 nematodes, one plant, and 49 viruses (Table 3). While most of these species were reported in neighbouring countries, some pests not reported in a neighbouring country were included to confirm freedom from these pests in the country. Examples of such pests include Candidatus Liberibacter asiaticus, D. chrysanthemi, D. dadantii, D. dianthicola, D. zeae, Globodera pallida, Foc TR4, P. betavasculorum, P. brasiliense, P. carotovorum, P. cypripedii, and P. parmentieri, sweet potato latent virus, Xff, Xfm, and Xfp. All data have been consolidated in Supplementary File S15 (Sheet: Surveillance).
Regulation
A proportion of 25% (n=476) of the assessed species is recommended for regulation. The 476 species include 116 arthropods, 58 bacteria, 25 Chromista, 109 fungi, six molluscs, 45 nematodes, two protists, 12 viroids, and 103 viruses (Table 3). Most of these species were likely introduced primarily as contaminants (n=339) and stowaways (n=57), or through both pathways (n=80, as contaminants and stowaways). All data have been consolidated in Supplementary File S15 (Sheet: Regulation). Pest risk analysis (PRA) documents to inform importing guidelines will be developed for these pests.
Suggested additional actions
Additional actions may include developing contingency plans, raising awareness about the species, managing the industry, and conducting research to provide further phytosanitary evidence to support either regulation or deregulation. A contingency plan has been proposed for nine pest species, including two arthropods, three bacteria, one fungus, one mollusc, one viroid, and one nematode (Table 3). Awareness-raising has been proposed for 26 species, including one arthropod, 21 bacteria, one fungus, one mollusc, one nematode, and one viroid (Table 3). Thirteen pests, consisting of 10 arthropods and three bacteria, were recommended for management by the industry (Table 3). Phytosanitary research is crucial for improving phytosanitary decisions; therefore, research has been proposed for three bacterial species, all part of the Xylella fastidiosa subspecies (Table 3). All data have been consolidated in Supplementary File S15 (Sheets: Contingency plan, Publicity, Management by Industry, and Research).
Discussion
Burundi has already encountered various plant biosecurity challenges similar to those faced by many other nations in Sub-Saharan Africa. This is evidenced by the introduction of numerous non-native species, which have adversely affected agriculture and livelihoods (Paudel Timilsena et al., 2022). Effective management of non-native pests classified as quarantine relies on an updated list of regulated pests that could inform phytosanitary measures to limit their introduction. Law No. 1/23 of November 23, 2017, on Plant Protection in Burundi addresses the protection of plants, plant products, and articles; the prevention and official control of harmful organisms at both the introduction and spread levels within the national territory; the dissemination and promotion of plant protection techniques aimed at enhancing plant production; and support for the export and import of plants and plant products. Article 4 prohibits the introduction, retention or transport of plant pests into the national territory, whatever the stage of their development.
The current list of regulated pests for Burundi was developed in 2006, and since then, the status of most pests in the country has changed (Legg and Fauquet, 2004; Niyongere et al., 2013; Nakato et al., 2023). Additionally, new threats are emerging that require urgent action (Amer and Mahmoud, 2020; Viljoen et al., 2020; Aguayo et al., 2021; Castro et al., 2021). To address these two gaps, pests not reported as present in Burundi were identified using the CABI Horizon Scanning Tool. Furthermore, a rapid risk assessment was conducted based on four attributes: the likelihood of entry (introduction) and establishment (including components of spread), and the magnitude of socio-economic and environmental (biodiversity) impacts (Kenis et al., 2022; Mulema et al., 2022, 2024).
The assessment of the introduction pathways demonstrated that the contaminant pathway, which is relevant for seed-borne and seed-transmitted pests, was considered more plausible for species within and beyond Africa (Denancé and Grimault, 2022). Equally, the stowaway pathway, which is more appropriate for vector- and soil-borne pests (Montagnani et al., 2022), was considered more likely for species reported in a neighbouring country. The information supporting the assessment was gathered from CABI Compendium enhanced datasheets, reviewed and published resources (journals and reviews), EPPO datasheets, grey literature, and expert opinions. Although these did not impact the overall risk score, the probable pathways of arrival and associated confidence levels informed discussions regarding the potential for entry and establishment. A minimum overall risk score was suggested to guide the selection of pest species for which management action was necessary.
A no-action was suggested for pests that recorded an overall score below the set minimum. However, this was not true for some pests, especially those considered high-risk due to their ability to establish and spread, and with significant socio-economic impacts. A no-action was also suggested for nine species, including six arthropods (Dieuches humilis, Nysius euphorbiae, Phyllocoptes arcani, Unkanodes albifascia, Unkanodes sapporonus, and Uroleucon escalantii), one bacterium (Sesame phyllody phytoplasma), and two viruses (Millet red leaf persistent luteovirus and Wheat yellow mosaic virus). These pest species were not assessed due to a lack of sufficient information to support a meaningful evaluation. All these species were not part of the horizon scanning, except for sesame phyllody phytoplasma. They were included because they serve as vectors (arthropods) or are vectored (bacteria and viruses) by other assessed pests. Although not evaluated, the risk associated with these species was considered very low.
For instance, species in the genus Unkanodes (U. albifascia, U. sapporonus, and U. escalantii) have not been reported in Africa. Dieuches humilis is a vector of Phytomonas leptovasorum, a protist reported on coffee species only in the Dominican Republic, Guyana, and Suriname (CABI, 2019a). Millet red leaf persistent luteovirus was vectored by Sitobion miscanthi, which is only reported in Asia, Oceania, and the Americas (CABI, 2022). Nysius euphorbiae is a vector of Phytomonas leptovasorum and Phytomonas staheli, but neither of these protists has been reported in Africa (CABI, 2019a, b). Phyllocoptes arcani vectors rose rosette virus, a virus reported only in Canada, India, and the United States (Vázquez-Iglesias, 2024). Additionally, roses are not a priority value chain for Burundi. Sesame phyllody phytoplasma is vectored by Neoaliturus haematoceps, which has only been reported in Egypt, Morocco, and Tunisia in Africa (CABI, 2021a). Wheat yellow mosaic virus is vectored by the soil-borne protist Polymyxa graminis. Although it has been reported in Africa (Burkina Faso, Côte d’Ivoire, Mali, Niger, and Senegal) (CABI, 2021b), an overall risk score of 105 was realised, Burundi does not source wheat seed in the mentioned countries, and it’s practically implausible to introduce Polymyxa graminis-contaminated soil either with seed or transportation vectors, especially vehicles.
Surveillance was considered for pests reported as present in neighbouring countries, as well as some in the East African region. This was suggested to ensure that phytosanitary measures are not imposed on pests that may already be present in Burundi, thereby creating a technical barrier to trade. This is in line with Article VII, Sections g and j of the International Plant Protection Convention (IPPC). This action was also deemed necessary for emerging pests prioritised by the region due to their high phytosanitary concern. Hence, confirming freedom from such pests is deemed essential. This is important not only for preventing entry but also for preparedness. A complete list of pests for which surveillance should be undertaken has been suggested. It is after their status has been established as negative that the NPPO can decide if regulation is necessary.
Regulation has been considered for certain pests, and it is also suggested that PRAs be conducted to guide importing requirements. This is crucial to ensure that the likely pathways of introduction are restricted (Simberloff et al., 2013). The majority of pests proposed for regulation were likely to be introduced via seed (seed-borne or seed-transmitted), particularly for bacteria, phytoplasma, viroids, and viruses, or through soil (soil-borne), especially in the case of tubers (Denancé and Grimault, 2022; Pagán, 2022; Franić et al., 2024). Nevertheless, some pests were earmarked for regulation, yet the final decision regarding this action was left to the NPPO. This is because these pests are not reported in the region and are not transmitted through seed but by vector, which diminishes their likelihood of introduction. Examples of such pests include Pantoea stewartii subsp. stewartii, rice dwarf virus, rice gall dwarf virus, rice grassy stunt virus, rice hoja blanca virus, rice ragged stunt virus, rice stripe necrosis virus, rice tungro bacilliform virus, and rice tungro spherical virus, among others. None of these rice viruses have yet been reported in Africa (Hull, 1996; Bolaños et al., 2017). Interestingly, vectors, which including Nephotettix cincticeps, Nephotettix malayanus, Nephotettix parvus, Nephotettix virescens, Nilaparvata bakeri, Nilaparvata lugens, Nilaparvata muiri, Recilia dorsalis, Unkanodes albifascia, and Unkanodes sapporonus, that vector the indicated rice viruses have not been reported as present in Africa or in the region especially for Nephotettix nigropictus (Cameroon), Polymyxa graminis (Burkina Faso, Côte d’Ivoire, Mali, Niger, and Senegal), and Sogatella furcifera (Egypt). A complete list of pests for regulation has been suggested.
The proposed actions included above focus on prevention and preparedness, eradicating pest species if detected early and containing them if their boundaries are established through delimiting surveillance. The eradication or containment of a pest for which its boundaries have been established is possible through the utilisation of low-risk pesticides that protect non-target biodiversity and/or biological control (Venette et al., 2021). Developing contingency plans is essential to ensure that a structured preparedness strategy is in place (Roy et al., 2024). A list of pests that require a contingency plan has been suggested. The public plays a significant role in the introduction of new pests due to porous borders and inadequate biosecurity measures, and it is also crucial for individuals to report new pests. Therefore, prioritising awareness of emerging and high-risk pests is essential, which is why a list of pests requiring publicity has been proposed. For certain high-risk pests, the NPPO, in collaboration with national agricultural research systems and academia, may allocate resources to phytosanitary research. This is vital for refining and guiding phytosanitary decisions. Research may focus on developing evidence notes to inform policy, understanding the population dynamics or behaviour of specific pest species, adapting and implementing diagnostic protocols, discovering vectors, and trialling environmentally friendly management actions (MacLeod and Lloyd, 2020; Turner et al., 2021).
The suggested actions are crucial for preventing the introduction and management of incursions. Prevention is the most cost-effective strategy for management. Although eradication is more expensive than prevention, its economic advantages significantly outweigh the associated costs. Once a non-native pest species becomes established and widespread, eradication becomes impossible. There is no practical justification for attempting to create pest-free areas or regions of low pest prevalence. The focus shifts to managing pest populations at a level that has minimal impact on productivity. While prevention and preparedness are the responsibilities of the NPPO, managing established pests is a burden to the farmer, who needs professional management advice on pest management solutions from public and private extension advisers and other key players in the relevant value chains. CABI has collaborated with the Ministry of Environment, Agriculture and Livestock and other actors in Burundi to establish Plant Health Clinics aimed at addressing the scourge of established pests (Ochilo et al., 2022; Toepfer et al., 2023). Plant Health Clinics play an important role in general surveillance and can be incorporated into an early detection and rapid response system (Rambauli and Antwi, 2021).
Conclusion
Horizon scanning has been utilised in several countries to prioritise high-risk non-native pests for which management actions, particularly the prevention of their introduction, are recommended. This method has now also been applied in Burundi. The results from the assessment have enabled the establishment of a register of priority pests, which will be key in facilitating risk monitoring. This will enhance real-time updates of risk based on new available information about the status of specific pests in countries in SSA that have similar biosecurity constraints, as well as from trading partners. This assessment has also enabled the updating of the list of regulated pests, last developed in 2006, ensuring that high-risk species are prevented from being introduced. The assessment results will also be utilised to support risk communication, particularly in raising awareness about high-risk and emerging pests, and in developing contingency and response plans for preparedness. To effectively tackle all these tasks, the NPPO needs to initiate strategic collaborative efforts with other key stakeholders in the Plant Health System, including academia, extension delivery organisations (both public and private), research institutions (public, private, and international), and regional NPPOs.
A regional rather than country-based approach in pest risk assessment and prioritisation will result in effective resource utilisation among countries and enhance protection against foreign and emerging risks. An observation across countries where the study has been conducted, such as Kenya, Ghana, Zambia (published), and Rwanda and Uganda (not yet published), demonstrates that there is an overlap in pests prioritised as high risk; however, many pests are also reported in neighbouring countries. With porous borders and weak border biosecurity, it is improbable that such pests are not present in those countries, which demonstrates a gap in pest reporting. The observation of many pests in neighbouring countries and the overlap of many high-risk prioritised pests underscores the regional pest-risk assessment and prioritisation approach. CABI has worked with partners to conduct rapid risk assessments at the regional levels of East Africa (EAC, East African Community), Central Africa (ECCA, Economic Community of Central African States) and West Africa (ECOWAS, Economic Community of West African States). However, the results from Burundi will enhance regional efforts for managing incursions in the East African Community.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
JM: Conceptualization, Data curation, Formal analysis, Funding acquisition, Methodology, Validation, Visualization, Writing – original draft, Writing – review & editing. PN: Formal analysis, Validation, Visualization, Writing – review & editing. PB: Formal analysis, Validation, Visualization, Writing – review & editing. EB: Formal analysis, Validation, Visualization, Writing – review & editing. EC: Formal analysis, Validation, Visualization, Writing – review & editing. EG: Formal analysis, Validation, Visualization, Writing – review & editing. GH: Formal analysis, Validation, Visualization, Writing – review & editing. EH: Formal analysis, Validation, Visualization, Writing – review & editing. ArI: Formal analysis, Validation, Visualization, Writing – review & editing. AnI: Formal analysis, Validation, Visualization, Writing – review & editing. MI: Formal analysis, Validation, Visualization, Writing – review & editing. MM: Formal analysis, Validation, Visualization, Writing – review & editing. LM: Formal analysis, Validation, Visualization, Writing – review & editing. BM: Formal analysis, Validation, Visualization, Writing – review & editing. DM: Formal analysis, Validation, Visualization, Writing – review & editing. AM: Formal analysis, Validation, Visualization, Writing – review & editing. JN: Formal analysis, Visualization, Writing – review & editing. PN: Formal analysis, Validation, Visualization, Writing – review & editing. FN: Formal analysis, Validation, Visualization, Writing – review & editing. DN: Formal analysis, Validation, Visualization, Writing – review & editing. DeN: Formal analysis, Validation, Visualization, Writing – review & editing. SN: Formal analysis, Validation, Visualization, Writing – review & editing. NN: Formal analysis, Validation, Visualization, Writing – review & editing. EN: Formal analysis, Validation, Visualization, Writing – review & editing. DiN: Funding acquisition, Visualization, Writing – review & editing. NN: Formal analysis, Validation, Visualization, Writing – review & editing. ES: Formal analysis, Validation, Visualization, Writing – review & editing. CN: Formal analysis, Validation, Visualization, Writing – review & editing. DK: Funding acquisition, Resources, Writing – review & editing. MA: Funding acquisition, Resources, Writing – review & editing. EO: Formal analysis, Validation, Visualization, Writing – review & editing. IR: Formal analysis, Validation, Visualization, Writing – review & editing. SM: Formal analysis, Validation, Visualization, Writing – review & editing. JV: Formal analysis, Validation, Visualization, Writing – review & editing. WO: Formal analysis, Resources, Validation, Visualization, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This research was implemented under the CABI-led PlantwisePlus Burundi programme, with financial support from the Embassy of the Kingdom of the Netherlands in Burundi.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fagro.2025.1604493/full#supplementary-material
References
Aguayo J., Cerf-Wendling I., Folscher A. B., Fourrier-Jeandel C., Ioos R., Mathews M. C., et al. (2021). First report of Fusarium oxysporum f. sp. cubense tropical race 4 (TR4) causing banana wilt in the Island of Mayotte. Plant Dis. 105, 219. doi: 10.1094/PDIS-06-20-1196-PDN
Ahmed D. A., Hudgins E. J., Cuthbert R. N., Kourantidou M., Diagne C., Haubrock P. J., et al. (2022). Managing biological invasions: the cost of inaction. Biol. Invasions 24, 1927–1946. doi: 10.1007/s10530-022-02755-0
Amer M. A. and Mahmoud S. Y. (2020). First report of Tomato brown rugose fruit virus on tomato in Egypt. New Rep. 41, 2044–0588.2020. doi: 10.5197/j.2044-0588.2020.041.024
Atinkut Asmare B., Freyer B., and Bingen J. (2022). Women in agriculture: pathways of pesticide exposure, potential health risks and vulnerability in sub-Saharan Africa. Environ. Sci. Eur. 34, 89. doi: 10.1186/s12302-022-00638-8
Aylor D. E. (1990). The role of intermittent wind in the dispersal of fungal pathogens. Annu. Rev. Phytopathol. 28, 73–92. doi: 10.1146/annurev.py.28.090190.000445
Bayón Á. and Vilà M. (2019). Horizon scanning to identify invasion risk of ornamental plants marketed in Spain. NeoBiota 52, 47–86. doi: 10.3897/neobiota.52.38113
Beckmann C. and Shine R. (2009). Impact of invasive cane toads on Australian birds. Conserv. Biol. 23, 1544–1549. doi: 10.1111/j.1523-1739.2009.01261.x, PMID: 19508674
Blackburn T. M., Essl F., Evans T., Hulme P. E., Jeschke J. M., Kühn I., et al. (2014). A unified classification of alien species based on the magnitude of their environmental impacts. PLOS Biology. 12, e1001850. doi: 10.1371/journal.pbio.1001850, PMID: 24802715
Bolaños C., Leiva A. M., Saavedra J., Bruzzone C., Cruz M., and Cuellar W. J. (2017). Occurrence and molecular detection of Rice hoja blanca virus (Genus Tenuivirus) in Peru. Plant Dis. 101, 1070–1070. doi: 10.1094/PDIS-12-16-1797-PDN
Buddie A. G., Rwomushana I., Offord L. C., Kibet S., Makale F., Djeddour D., et al. (2021). First report of the invasive snail Pomacea canaliculata in Kenya. CABI Agric. Biosci. 2, 11. doi: 10.1186/s43170-021-00032-z
CABI (2019a). Phytomonas leptovasorum. CABI Compend (Wallingford, United Kingdom: CABI Compendium), 40308. doi: 10.1079/cabicompendium.40308
CABI (2019b). Phytomonas staheli. CABI Compend (Wallingford, United Kingdom: CABI Compendium), 40309. doi: 10.1079/cabicompendium.40309
CABI (2021a). Neoaliturus haematoceps. CABI Compend (Wallingford, United Kingdom: CABI Compendium), 36364. doi: 10.1079/cabicompendium.36364
CABI (2021b). Polymyxa graminis. CABI Compend (Wallingford, United Kingdom: CABI Compendium), 42776. doi: 10.1079/cabicompendium.42776
CABI (2022). Sitobion miscanthi (Indian grain aphid). CABI Compend (Wallingford, United Kingdom: CABI Compendium), 51740. doi: 10.1079/cabicompendium.51740
Caffrey J. M., Baars J. R., Barbour J. H., Boets P., Boon P., Davenport K., et al. (2014). Tackling invasive alien species in Europe: The top 20 issues. Manage. Biol. Invasions 5, 1–20. doi: 10.3391/mbi.2014.5.1.01
Campbell W. G. (1929). Quarantine measures as trade barriers. Ann. Am. Acad. Pol. Soc Sci. 141, 30–35. doi: 10.1177/000271622914100104
Castro C., DiSalvo B., and Roper M. C. (2021). Xylella fastidiosa: A reemerging plant pathogen that threatens crops globally. PloS Pathog. 17, e1009813. doi: 10.1371/journal.ppat.1009813, PMID: 34499674
Dawson W., Peyton J. M., Pescott O. L., Adriaens T., Cottier-Cook E. J., Frohlich D. S., et al. (2023). Horizon scanning for potential invasive non-native species across the United Kingdom Overseas Territories. Conserv. Lett. 16, e12928. doi: 10.1111/conl.12928
Dechassa N. (2020). Origin, distribution, impact and management of Water hyacinth (Eichhornia crassipes (Martius) Solm): A review. J. Geogr. Environ. Earth Sci. Int. 10, 13–18. doi: 10.7176/JEES/10-10-02
De Groote H., Kimenju S. C., Munyua B., Palmas S., Kassie M., and Bruce A. (2020). Spread and impact of fall armyworm (Spodoptera frugiperda J.E. Smith) in maize production areas of Kenya. Agric. Ecosyst. Environ. 292, 106804. doi: 10.1016/j.agee.2019.106804, PMID: 32308246
Denancé N. and Grimault V. (2022). Seed pathway for pest dissemination: The ISTA Reference Pest List, a bibliographic resource in non-vegetable crops. EPPO Bull. 52, 434–445. doi: 10.1111/epp.12834
Desneux N., Han P., Mansour R., Arnó J., Brévault T., Campos M. R., et al. (2022). Integrated pest management of Tuta absoluta: practical implementations across different world regions. J. Pest Sci. 95, 17–39. doi: 10.1007/s10340-021-01442-8
Early R., Bradley B. A., Dukes J. S., Lawler J. J., Olden J. D., Blumenthal D. M., et al. (2016). Global threats from invasive alien species in the twenty-first century and national response capacities. Nat. Commun. 7, 12485. doi: 10.1038/ncomms12485, PMID: 27549569
Epanchin-Niell R., McAusland C., Liebhold A., Mwebaze P., and Springborn M. R. (2021). Biological invasions and international trade: Managing a moving target. Rev. Environ. Econ. Policy 15, 180–190. doi: 10.1086/713025
Franić I., Cleary M., Aday Kaya A. G., Bragança H., Brodal G., Cech T. L., et al. (2024). The biosecurity risks of international forest tree seed movements. Curr. For. Rep. 10, 89–102. doi: 10.1007/s40725-023-00211-3
Gallardo B. and Aldridge D. C. (2013). Priority setting for invasive species management: risk assessment of Ponto-Caspian invasive species into Great Britain. Ecol. Appl. 23, 352–364. doi: 10.1890/12-1018.1, PMID: 23634587
Gallardo B., Zieritz A., Adriaens T., Bellard C., Boets P., Britton J. R., et al. (2016). Trans-national horizon scanning for invasive non-native species: a case study in western Europe. Biol. Invasions 18, 17–30. doi: 10.1007/s10530-015-0986-0
Gassó N., Sol D., Pino J., Dana E. D., Lloret F., Sanz-Elorza M., et al. (2009). Exploring species attributes and site characteristics to assess plant invasions in Spain. Divers. Distrib. 15, 50–58. doi: 10.1111/j.1472-4642.2008.00501.x
Graziosi I., Tembo M., Kuate J., and Muchugi A. (2020). Pests and diseases of trees in Africa: A growing continental emergency. Plants People Planet 2, 14–28. doi: 10.1002/ppp3.31
Guimapi R. Y. A., Mohamed S. A., Okeyo G. O., Ndjomatchoua F. T., Ekesi S., and Tonnang H. E. Z. (2016). Modeling the risk of invasion and spread of Tuta absoluta in Africa. Ecol. Complex. 28, 77–93. doi: 10.1016/j.ecocom.2016.08.001
Heck M. (2018). Insect transmission of plant pathogens: a systems biology perspective. mSystems 3, 1–5. doi: 10.1128/mSystems.00168-17, PMID: 29629417
Hugouvieux-Cotte-Pattat N. and Van Gijsegem F. (2021). Diversity within the Dickeya zeae complex, identification of Dickeya zeae and Dickeya oryzae members, proposal of the novel species Dickeya parazeae sp. nov. Int. J. Syst. Evol. Microbiol. 71, 1–10. doi: 10.1099/ijsem.0.005059, PMID: 34726587
Hull R. (1996). Molecular biology of rice tungro viruses. Annu. Rev. Phytopathol. 34, 275–297. doi: 10.1146/annurev.phyto.34.1.275, PMID: 15012544
Hulme P. E., Bacher S., Kenis M., Klotz S., Kühn I., Minchin D., et al. (2008). Grasping at the routes of biological invasions: a framework for integrating pathways into policy. J. Appl. Ecol. 45, 403–414. doi: 10.1111/j.1365-2664.2007.01442.x
Hulme P. E., Beggs J. R., Binny R. N., Bray J. P., Cogger N., Dhami M. K., et al. (2023). Emerging advances in biosecurity to underpin human, animal, plant, and ecosystem health. iScience 26, 107462. doi: 10.1016/j.isci.2023.107462, PMID: 37636074
Kendig A. E., Canavan S., Anderson P. J., Flory S. L., Gettys L. A., Gordon D. R., et al. (2022). Scanning the horizon for invasive plant threats using a data-driven approach. NeoBiota 74, 129–154. doi: 10.3897/neobiota.74.83312
Kenis M., Agboyi L. K., Adu-Acheampong R., Ansong M., Arthur S., Attipoe P. T., et al. (2022). Horizon scanning for prioritising invasive alien species with potential to threaten agriculture and biodiversity in Ghana. NeoBiota 71, 129–148. doi: 10.3897/neobiota.71.72577
Korda M., Ripka G., Hradil K., Glavendekic M., Matosevic D., Hrasovec B., et al. (2023). Alien eating alien - rapid spread of Aceria fraxiniflora, a non-native gall mite of the invasive green ash (Fraxinus pennsylvanica) in Central-Eastern Europe. Exp. Appl. Acarol. 91, 405–412. doi: 10.1007/s10493-023-00849-5, PMID: 37819594
Kueffer C. and Loope L. (2009). Prevention, early detection and containment of invasive, nonnative plants in the Hawaiian Islands: current efforts and needs. Available online at: http://pubs.er.usgs.gov/publication/70175010 (Accessed May 3, 2023).
Kumar R. M., Gadratagi B.-G., Paramesh V., Kumar P., Madivalar Y., Narayanappa N., et al. (2022). Sustainable management of invasive fall armyworm, Spodoptera frugiperda. Agronomy 12, 1–17. doi: 10.3390/agronomy12092150
Kumar Rai P. and Singh J. S. (2020). Invasive alien plant species: Their impact on environment, ecosystem services and human health. Ecol. Indic. 111, 106020. doi: 10.1016/j.ecolind.2019.106020, PMID: 32372880
Legg J. P. and Fauquet C. M. (2004). Cassava mosaic geminiviruses in Africa. Plant Mol. Biol. 56, 585–599. doi: 10.1007/s11103-004-1651-7, PMID: 15630622
Lieurance D., Canavan S., Behringer D. C., Kendig A. E., Minteer C. R., Reisinger L. S., et al. (2023). Identifying invasive species threats, pathways, and impacts to improve biosecurity. Ecosphere 14, e4711. doi: 10.1002/ecs2.4711
Macharia I., Kimani E., Koome F., Kosiom T., Heya H., Otipa M., et al. (2017). First report and distribution of the papaya mealybug, Paracoccus marginatus, in Kenya. J. Agric. Urban Entomol. 33, 142–150. doi: 10.3954/jaue17-02.1
MacLeod A. and Lloyd S. (2020). The emergence of prioritisation systems to inform plant health biosecurity policy decisions. Emerg. Top. Life Sci. 4, 463–471. doi: 10.1042/ETLS20200341, PMID: 33210712
Mahuku G., Lockhart B. E., Wanjala B., Jones M. W., Kimunye J. N., Stewart L. R., et al. (2015). Maize lethal necrosis (MLN), an emerging threat to maize-based food security in sub-Saharan Africa. Phytopathology 105, 956–965. doi: 10.1094/PHYTO-12-14-0367-FI, PMID: 25822185
Matova P. M., Kamutando C. N., Magorokosho C., Kutywayo D., Gutsa F., and Labuschagne M. (2020). Fall-armyworm invasion, control practices and resistance breeding in Sub-Saharan Africa. Crop Sci. 60, 2951–2970. doi: 10.1002/csc2.20317, PMID: 33328691
Matowo N. S., Tanner M., Munhenga G., Mapua S. A., Finda M., Utzinger J., et al. (2020). Patterns of pesticide usage in agriculture in rural Tanzania call for integrating agricultural and public health practices in managing insecticide-resistance in malaria vectors. Malar. J. 19, 257. doi: 10.1186/s12936-020-03331-4, PMID: 32677961
Matthews J., Beringen R., Creemers R., Hollander H., Kessel N., H.H. V., et al. (2017). A new approach to horizon-scanning: Identifying potentially invasive alien species and their introduction pathways. Manage. Biol. Invasions 8, 1–16. doi: 10.3391/mbi.2017.8.1.04
Mburu H., Cortada L., Haukeland S., Ronno W., Nyongesa M., Kinyua Z., et al. (2020). Potato cyst nematodes: A new threat to potato production in East Africa. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.00670, PMID: 32523602
Mims S. A. and Mims F. M. (2004). Fungal spores are transported long distances in smoke from biomass fires. Atmos. Environ. 38, 651–655. doi: 10.1016/j.atmosenv.2003.10.043
Montagnani C., Gentili R., Brundu G., Caronni S., and Citterio S. (2022). Accidental introduction and spread of top invasive alien plants in the European Union through human-mediated agricultural pathways: What should we expect? Agronomy 12, 1–28. doi: 10.3390/agronomy12020423
Montgomery K., Walden-Schreiner C., Saffer A., Jones C., Seliger B. J., Worm T., et al. (2023). Forecasting global spread of invasive pests and pathogens through international trade. Ecosphere 14, e4740. doi: 10.1002/ecs2.4740
Mulema J., Day R., Nunda W., Akutse K. S., Bruce A. Y., Gachamba S., et al. (2022). Prioritization of invasive alien species with the potential to threaten agriculture and biodiversity in Kenya through horizon scanning. Biol. Invasions. 24, 2933–2949. doi: 10.1007/s10530-022-02824-4
Mulema J., Phiri S., Bbebe N., Chandipo R., Chijikwa M., Chimutingiza H., et al. (2024). Rapid risk assessment of plant pathogenic bacteria and protists likely to threaten agriculture, biodiversity and forestry in Zambia. NeoBiota 91, 145–178. doi: 10.3897/neobiota.91.113801
Nagoshi R. N., Goergen G., Tounou K. A., Agboka K., Koffi D., and Meagher R. L. (2018). Analysis of strain distribution, migratory potential, and invasion history of fall armyworm populations in northern Sub-Saharan Africa. Sci. Rep. 8, 3710–3710. doi: 10.1038/s41598-018-21954-1, PMID: 29487323
Nakato G. V., Okonya J. S., Kantungeko D., Ocimati W., Mahuku G., Legg J. P., et al. (2023). Influence of altitude as a proxy for temperature on key Musa pests and diseases in watershed areas of Burundi and Rwanda. Heliyon 9, 1–12. doi: 10.1016/j.heliyon.2023.e13854, PMID: 36895396
Niyongere C., Losenge T., Ateka E. M., Ntukamazina N., Ndayiragije P., Simbare A., et al. (2013). Understanding banana bunchy top disease epidemiology in Burundi for an enhanced and integrated management approach. Plant Pathol. 62, 562–570. doi: 10.1111/j.1365-3059.2012.02676.x
Ochilo W. N., Toepfer S., Ndayihanzamaso P., Mugambi I., Vos J., and Niyongere C. (2022). Assessing the plant health system of Burundi: What it is, who matters and why. Sustainability 14, 1–19. doi: 10.3390/su142114293
Pagán I. (2022). Transmission through seeds: The unknown life of plant viruses. PloS Pathog. 18, e1010707. doi: 10.1371/journal.ppat.1010707, PMID: 35951532
Paini D. R., Sheppard A. W., Cook D. C., De Barro P. J., Worner S. P., and Thomas M. B. (2016). Global threat to agriculture from invasive species. Proc. Natl. Acad. Sci. 113, 7575–7579. doi: 10.1073/pnas.1602205113, PMID: 27325781
Pasanen M., Waleron M., Schott T., Cleenwerck I., Misztak A., Waleron K., et al. (2020). Pectobacterium parvum sp. nov., having a Salmonella SPI-1-like Type III secretion system and low virulence. Int. J. Syst. Evol. Microbiol. 70, 2440–2448. doi: 10.1099/ijsem.0.004057, PMID: 32100697
Paudel Timilsena B., Niassy S., Kimathi E., Abdel-Rahman E. M., Seidl-Adams I., Wamalwa M., et al. (2022). Potential distribution of fall armyworm in Africa and beyond, considering climate change and irrigation patterns. Sci. Rep. 12, 539. doi: 10.1038/s41598-021-04369-3, PMID: 35017586
Portier P., Pédron J., Taghouti G., Fischer-Le Saux M., Caullireau E., Bertrand C., et al. (2019). Elevation of Pectobacterium carotovorum subsp. odoriferum to species level as Pectobacterium odoriferum sp. nov., proposal of Pectobacterium brasiliense sp. nov. and Pectobacterium actinidiae sp. nov., emended description of Pectobacterium carotovorum and description of Pectobacterium versatile sp. nov., isolated from streams and symptoms on diverse plants. Int. J. Syst. Evol. Microbiol. 69, 3207–3216. doi: 10.1099/ijsem.0.003611, PMID: 31343401
Raffini F., Bertorelle G., Biello R., D’Urso G., Russo D., and Bosso L. (2020). From nucleotides to satellite imagery: Approaches to identify and manage the invasive pathogen Xylella fastidiosa and its insect vectors in Europe. Sustainability 12, 1–38. doi: 10.3390/su12114508
Rambauli M. and Antwi M. (2021). Review of plant health clinics: a recent phenomenon for effective plant pests and diseases diagnosis. South Afr. J. Agric. Ext. SAJAE 49, 90–103. doi: 10.17159/2413-3221/2021/v49n3a11245
Roper M., Seminara A., Bandi M. M., Cobb A., Dillard H. R., and Pringle A. (2010). Dispersal of fungal spores on a cooperatively generated wind. Proc. Natl. Acad. Sci. U. S. A. 107, 17474–17479. doi: 10.1073/pnas.1003577107, PMID: 20880834
Roy H. E., Bacher S., Essl F., Adriaens T., Aldridge D. C., Bishop J. D. D., et al. (2019). Developing a list of invasive alien species likely to threaten biodiversity and ecosystems in the European Union. Glob. Change Biol. 25, 1032–1048. doi: 10.1111/gcb.14527, PMID: 30548757
Roy H. E., Pauchard A., Stoett P., Renard Truong T., Lipinskaya T., and Vicente J. R. (2023). “Chapter 1: Introducing biological invasions and the IPBES thematic assessment of invasive alien species and their control,” in Thematic Assessment Report on Invasive Alien Species and their Control of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. Eds. Roy H. E., Pauchard A., Stoett P., and Truong T.R. (IPBES Secretariat, Bonn, Germany). doi: 10.5281/zenodo.7430723
Roy H. E., Pauchard A., Stoett P., Renard Truong T., Lipinskaya T., and Vicente J. R. (2024). IPBES Invasive Alien Species Assessment: Chapter 1. Introducing biological invasions and the IPBES thematic assessment of invasive alien species and their control (Bonn, Germany). doi: 10.5281/zenodo.10677041
Roy H. E., Peyton J., Aldridge D. C., Bantock T., Blackburn T. M., Britton R., et al. (2014). Horizon scanning for invasive alien species with the potential to threaten biodiversity in Great Britain. Glob. Change Biol. 20, 3859–3871. doi: 10.1111/gcb.12603, PMID: 24839235
Rwomushana I., Pratt C., González-Moreno P., Beale T., Godwin J., Makale F., et al. (2019). Tomato leaf miner (Tuta absoluta): impacts and coping strategies for Africa (Nairobi, Kenya).
Saravanakumar D., Bartholomew E. S., Seepersad G., Gore-Francis J., Goldsmith J., Ramnanan N., et al. (2023). Prioritisation of quarantine pest list for the Caribbean using a multi-criteria decision approach. NeoBiota 88, 1–16. doi: 10.3897/neobiota.88.102673
Sarfraz S., Riaz K., Oulghazi S., Cigna J., Sahi S. T., Khan S. H., et al. (2018). Pectobacterium punjabense sp. nov., isolated from blackleg symptoms of potato plants in Pakistan. Int. J. Syst. Evol. Microbiol. 68, 3551–3556. doi: 10.1099/ijsem.0.003029, PMID: 30239330
Seymour C. L., Gillson L., Child M. F., Tolley K. A., Curie J. C., da Silva J. M., et al. (2020). Horizon scanning for South African biodiversity: A need for social engagement as well as science. Ambio 49, 1211–1221. doi: 10.1007/s13280-019-01252-4, PMID: 31564051
Shine R. (2010). The ecological impact of invasive cane toads (Bufo marinus) in Australia. Q. Rev. Biol. 85, 253–291. doi: 10.1086/655116, PMID: 20919631
Simberloff D., Martin J.-L., Genovesi P., Maris V., Wardle D. A., Aronson J., et al. (2013). Impacts of biological invasions: what’s what and the way forward. Trends Ecol. Evol. 28, 58–66. doi: 10.1016/j.tree.2012.07.013, PMID: 22889499
Sutherland W. J., Albon S. D., Allison H., Armstrong-Brown S., Bailey M. J., Brereton T., et al. (2010). Review: The identification of priority policy options for UK nature conservation. J. Appl. Ecol. 47, 955–965. doi: 10.1111/j.1365-2664.2010.01863.x
Sutherland W. J., Bailey M. J., Bainbridge I. P., Brereton T., Dick J. T. A., Drewitt J., et al. (2008). Future novel threats and opportunities facing UK biodiversity identified by horizon scanning. J. Appl. Ecol. 45, 821–833. doi: 10.1111/j.1365-2664.2008.01474.x
Sutherland W. J., Dias M. P., Dicks L. V., Doran H., Entwistle A. C., Fleishman E., et al. (2020). A horizon scan of emerging global biological conservation issues for 2020. Trends Ecol. Evol. 35, 81–90. doi: 10.1016/j.tree.2019.10.010, PMID: 31813647
Toepfer S., Niyongere C., Ndayihanzamaso P., Ndikumana D., Irakoze W., Cimpaye E., et al. (2023). Sustainable improvements in diagnostic capabilities of plant health practitioners through short In-service training. Sustainability 15, 1–12. doi: 10.3390/su151712956
Turner R. M., Brockerhoff E. G., Bertelsmeier C., Blake R. E., Caton B., James A., et al. (2021). Worldwide border interceptions provide a window into human-mediated global insect movement. Ecol. Appl. 31, e02412. doi: 10.1002/eap.2412, PMID: 34255404
Vázquez-Iglesias I. (2024). rose rosette virus. CABI Compend (CABI Compendium), 47834. doi: 10.1079/cabicompendium.47834
Venette R. C., Gordon D. R., Juzwik J., Koch F. H., Liebhold A. M., Peterson R. K. D., et al. (2021). “Early intervention strategies for invasive species management: connections between risk assessment, prevention efforts, eradication, and other rapid responses,” in Invasive species in forests and rangelands of the United States: A comprehensive science synthesis for the United States forest sector. Eds. Poland T. M., Patel-Weynand T., Finch D. M., Miniat C. F., Hayes D. C., and Lopez V. M. (Springer International Publishing, Cham), 111–131. doi: 10.1007/978-3-030-45367-1_6
Vilcinskas A. (2019). Pathogens associated with invasive or introduced insects threaten the health and diversity of native species. Pests Resist. • Behav. Ecol. 33, 43–48. doi: 10.1016/j.cois.2019.03.004, PMID: 31358194
Viljoen A., Mostert D., Chiconela T., Beukes I., Fraser C., Dwyer J., et al. (2020). Occurrence and spread of the banana fungus Fusarium oxysporum f. sp. cubense TR4 in Mozambique. South Afr. J. Sci. 116, 1–11. doi: 10.17159/sajs.2020/8608
Wabuyele E., Lusweti A., Bisikwa J., Kyenune G., Clark K., Lotter W., et al. (2014). A roadside survey of the invasive weed Parthenium hysterophorus (Asteraceae) in East Africa. J. East Afr. Nat. Hist. 103, 49–57. doi: 10.2982/028.103.0105
Waleron M., Misztak A., Waleron M., Franczuk M., Wielgomas B., and Waleron K. (2018). Transfer of Pectobacterium carotovorum subsp. carotovorum strains isolated from potatoes grown at high altitudes to Pectobacterium Peruviense sp. nov. Syst. Appl. Microbiol. 41, 85–93. doi: 10.1016/j.syapm.2017.11.005, PMID: 29325987
Wang R., Bai B., Li D., Wang J., Huang W., Wu Y., et al. (2024). Phytoplasma: A plant pathogen that cannot be ignored in agricultural production-Research progress and outlook. Mol. Plant Pathol. 25, e13437. doi: 10.1111/mpp.13437, PMID: 38393681
Weber E. and Gut D. (2004). Assessing the risk of potentially invasive plant species in central Europe. J. Nat. Conserv. 12, 171–179. doi: 10.1016/j.jnc.2004.04.002
Wendimu G. Y. and Gurmu A. K. (2024). Insect vectors of plant viruses: Host interactions, their effects, and future opportunities. Adv. Agric. 2024, 6006985. doi: 10.1155/aia/6006985
Wielkopolan B., Jakubowska M., and Obrępalska-Stęplowska A. (2021). Beetles as plant pathogen vectors. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.748093, PMID: 34721475
Keywords: invasive alien species, horizon scanning, pest identification, pest risk analysis, pest risk management
Citation: Mulema J, Ndayihanzamaso P, Birushe P, Bizimungu E, Cimpaye E, Gikoro E, Hakizimana G, Hatungimana E, Ingabire A, Irakiza A, Itangishaka MG, Manirakiza M, Masabarakiza L, Minani B, Minani D, Mpawenimana A, Nahaniyo J, Ndayiragije P, Ndayishemeza F, Ndayishimiye D, Ndikumana D, Nduwimana S, Niko N, Nkubaye E, Nshimirimana D, Ntawuyankira N, Sakayoya E, Niyongere C, Karanja D, Akiri M, Onkendi E, Rwomushana I, Musonerimana S, Vos JGM and Ochilo W (2025) Rapid risk assessment to prioritise potentially high-risk non-native plant pests to protect agriculture and forestry, safeguard biodiversity, and facilitate trade in Burundi. Front. Agron. 7:1604493. doi: 10.3389/fagro.2025.1604493
Received: 01 April 2025; Accepted: 08 August 2025;
Published: 08 September 2025.
Edited by:
Murray B. Isman, University of British Columbia, CanadaReviewed by:
Jozsef Kiss, Szent István University, HungarySatya Singh, National Innovation Foundation, India
Phumudzo Patrick Tshikhudo, University of South Africa, South Africa
Ahmed Imam, Desert Research Center, Egypt
Copyright © 2025 Mulema, Ndayihanzamaso, Birushe, Bizimungu, Cimpaye, Gikoro, Hakizimana, Hatungimana, Ingabire, Irakiza, Itangishaka, Manirakiza, Masabarakiza, Minani, Minani, Mpawenimana, Nahaniyo, Ndayiragije, Ndayishemeza, Ndayishimiye, Ndikumana, Nduwimana, Niko, Nkubaye, Nshimirimana, Ntawuyankira, Sakayoya, Niyongere, Karanja, Akiri, Onkendi, Rwomushana, Musonerimana, Vos and Ochilo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Joseph Mulema, ai5tdWxlbWFAY2FiaS5vcmc=