 Mélanie Grosjean
Mélanie Grosjean Sophie Guénard1
Sophie Guénard1 Caroline Giraud
Caroline Giraud Paulo Juarez
Paulo Juarez- 1Département Recherche et Développement, Smaltis SAS, Besançon, France
- 2Laboratoire de Bactériologie, UMR CNRS 6249 Chrono-Environnement, Université Bourgogne Franche-Comté, Besançon, France
- 3Normandie Université, UNICAEN, U2RM, Caen, France
- 4Centre National de Référence de la Résistance aux Antibiotiques, Centre Hospitalier Régional Universitaire de Besançon, Besançon, France
Pseudomonas aeruginosa is a human opportunistic pathogen responsible for nosocomial infections, which is largely used as a model organism to study antibiotic resistance and pathogenesis. As other species of the genus, its wide metabolic versatility appears to be attractive to study biotechnological applications. However, its natural resistance to antibiotics and its capacity to produce a wide range of virulence factors argue against its biotechnological potential. By reducing the genome of the reference strain PAO1, we explored the development of four hypovirulent and hypersusceptible recombinant DNA hosts (rDNA hosts). Despite deleting up to 0.8% of the core genome, any of the developed strains presented alterations of fitness when cultured under standard laboratory conditions. Other features such as antibiotic susceptibility, cytotoxicity, in vivo pathogenesis, and expression of heterologous peptides were also explored to highlight the potential applications of these models. This work stands as the first stage of the development of a safe-platform strain of Pseudomonas aeruginosa that will be further optimized for biotechnological applications.
Introduction
Since the pioneering works of Bruce Holloway in the 1950s, Pseudomonas aeruginosa has become a model organism for exploring the genetics and physiological functions of genus Pseudomonas, and more generally Gram-negative non-fermenters. However, this microorganism stands out from most of related species by its capacity to adapt to and thrive in multiple environments, including the hospital (Moradali et al., 2017). This last adaptation makes of this bacterium an opportunistic pathogen able to cause life-threatening infections in immunocompromised patients such as those with respiratory infections, bacteremia and chronic diseases such as cystic fibrosis (Lister et al., 2009; de Bentzmann and Plésiat, 2011; McCarthy, 2015). Despite the development of specific molecular biology tools, the exploitation of the metabolic versatility of P. aeruginosa to produce recombinant proteins under safe conditions has been hampered by the potential pathogenicity of the bacterium (Moradali et al., 2017). The remarkable adaptability of P. aeruginosa is determined by a large genome of 6.4- to 7.4-Mbp in size (Hilker et al., 2015), the annotation of which is still in progress1 (Stover et al., 2000). It harbors genes encoding for a wide range of secreted virulence factors including lipases, proteases, exoenzymes and exotoxins (Dacheux et al., 2001; Diggle et al., 2006; Chemani et al., 2009; Hauser, 2009; Filloux, 2011; Burrows, 2012). Moreover, the cell envelope of this bacterium also contains several virulence factors such as adhesins, lectins, pili, flagella, and lipopolysaccharide (LPS) in the outer membrane (Feldman et al., 1998; Kohler et al., 2000; King et al., 2009; Wilder et al., 2011; Burrows, 2012; Kazmierczak et al., 2015). Additionally, to its pathogenesis, P. aeruginosa possesses an intrinsic resistance to several anti-Gram-negative antibiotics such as β-lactams and aminoglycosides (Lister et al., 2009; de Bentzmann and Plésiat, 2011; Li et al., 2015; McCarthy, 2015). This natural resistance is principally due to the low permeability of its membrane, the production of enzymes capable to degrade or modify antibiotics and the action of two efflux systems able to export multiple molecules outside the cell (Yonezawa et al., 1995; Poole, 2004; Juan et al., 2005, 2006; Lister et al., 2009; Vettoretti et al., 2009; Berrazeg et al., 2015; Ropy et al., 2015; Jeannot et al., 2017). All these genomic determinants were targeted in this work with the objective to build up four different rDNA hosts by reducing the genome of the reference strain PAO1 without causing any negative impact on bacterial growth very similar as the work done on P. putida (Lieder et al., 2015). This project is placed as a first stage to develop a safe-platform strain of Pseudomonas aeruginosa which will be further optimized for biotechnological applications such as expression of heterologous proteins or secondary metabolites.
Materials and Methods
Bacterial Strains, Plasmids, and Growth Conditions
Main features of bacterial strains and plasmids used in this study are listed in Table 1. Bacterial cultures of E. coli were performed in Luria-Bertani broth or on Luria-Bertani agar while those of P. aeruginosa were performed in Mueller-Hinton broth (MHB) with adjusted concentrations of Ca2+ (from 20 to 25 μg mL–1) and Mg2+ (from 10 to 12.5 μg mL–1), on Mueller-Hinton agar (MHA) or on Typticase Soy Broth (TSB). All media were purchased from Beckton-Dickinson. When needed, 50 μg mL–1 kanamycin or 50 μg mL–1 streptomycin was added to E. coli growth media. Recombinant plasmids were introduced into P. aeruginosa by triparental mating and mobilization with conjugative plasmid pRK2013 (KmR) provided by helper strain E. coli HB101 (Ditta et al., 1980) or electroporation (Dennis and Sokol, 1995).

Table 1. Strains and plasmids used in this study.
Genomic Deletions
Gene inactivation experiments were all carried out using the suicide plasmid pKNG101 (Kaniga et al., 1991) and homologous recombination events, Figure 1 schematizes all the steps involved in the gene deletion protocol. Additionally, the map of plasmid pKNG101 is presented in Supplementary Figure 1. Briefly, recombinant plasmids were constructed by assembly cloning using the NEBuilder Hi-Fi DNA Assembly Cloning Kit (New England Biolabs). Assembly products were directly used to transform competent E. coli CC118λpir strains. Once checked by DNA sequencing, recombinant plasmids with appropriate inserts were transferred to P. aeruginosa by conjugation. Transconjugants were selected on Pseudomonas Isolation Agar (PIA; Becton Dickinson) containing 500 or 2,000 μg mL–1 streptomycin. Excision of the undesired pKNG101 sequence was obtained by plating transformants on M9 plates (8.54 mM NaCl, 25.18 mM NaH2PO4, 18.68 mM NH4Cl, 22 mM KH2PO4, 2 mM MgSO4, 0.8% agar, pH 7.4) containing 5% (wt/vol) sucrose. Negative selection on streptomycin-containing MHA allowed the identification of transconjugants that had lost the plasmid. Finally, the allelic exchanges were verified by PCR and sequencing. All genomic modifications made to strain SM54 were verified by Whole Genome Sequencing using Illumina NextSeq sequencing (reads of 150 bp and 100× coverage). Reads were mapped against the PAO1 genome (Refseq NZ_CP053028.1) using CLC Genomic Workbench. Supplementary Table 1 lists the primers and temperatures employed to amplify the DNA fragments that were cloned in pKNG101, and that flanked targeted genes in the PAO1 genome. Detailed information about genomic deletions is found on Supplementary Table Gene Deletion.
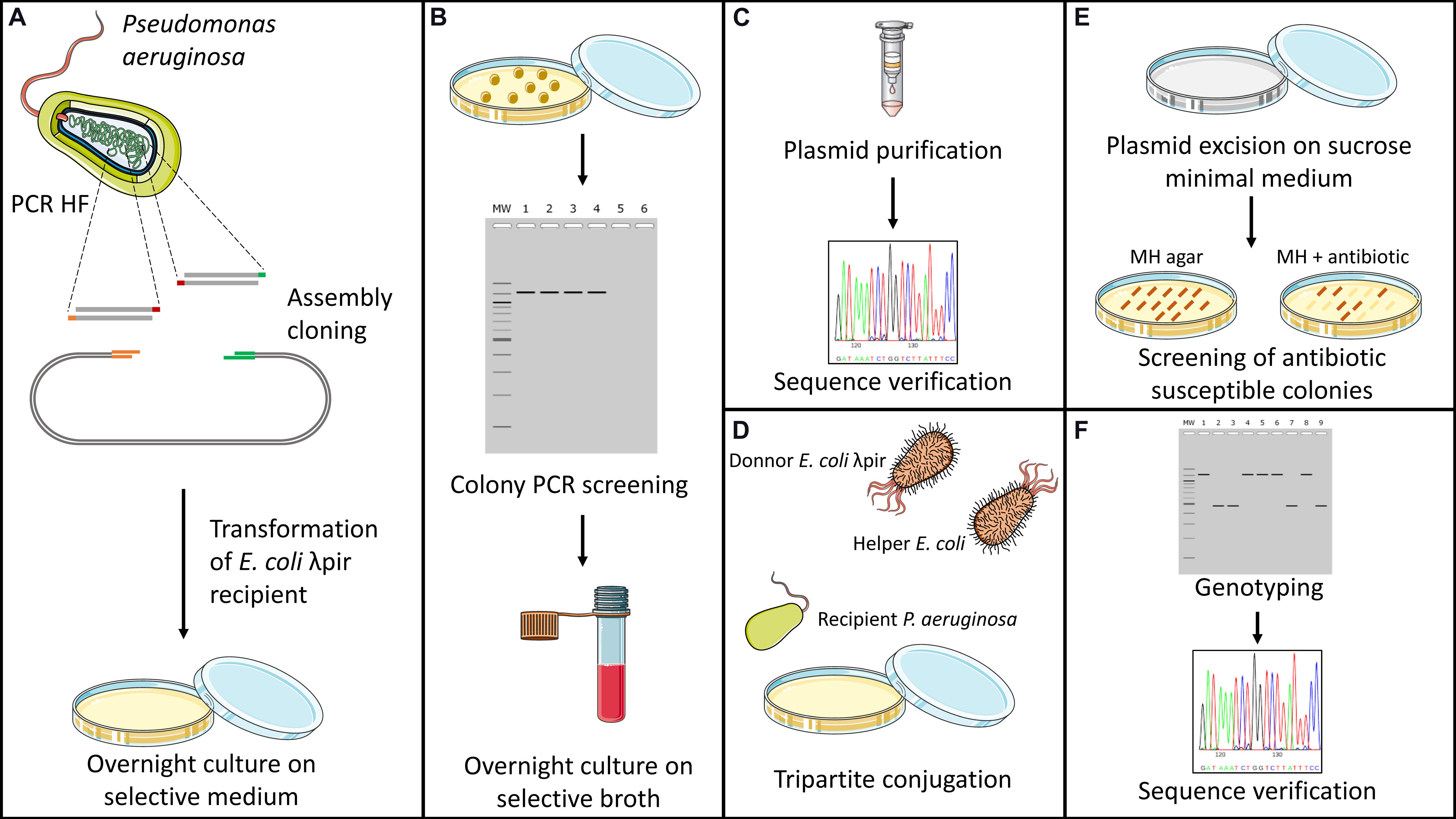
Figure 1. Schematic representation of genome deletion protocol. Genome deletions were carried out by homologous recombination using plasmid pKNG101 as a suicide vector (Kaniga et al., 1991). (A) Firstly, upstream and downstream regions of the targeted gene are amplified using a high-fidelity polymerase. Purified fragments are then inserted into linearized plasmid by assembly cloning and used to transformed CC118λpir competent cells of E. coli. (B) Antibiotic resistant colonies obtained after transformation are then screened by colony PCR to confirm the insertion of both fragments into the vector. (C) Recombinant plasmids are extracted and verified for absence of undesired SNPs through Sanger sequencing. (D) Transfer of recombinant plasmid to Pseudomonas aeruginosa is made by tripartite conjugation using the donor CC118 λpir strain of E. coli and HB101 helper strain containing plasmid pRK2013. (E) Transconjugants are then placed on M9 Minimal Medium containing sucrose as sole source of carbon. After several days of culture at room temperature, colonies are screened for plasmid excision through antibiotic susceptibility testing on nutritive media. (F) Antibiotic-susceptible colonies are screened through colony PCR for deletion of the targeted gene. Chromosomic deletions are verified by Sanger sequencing.
Antibiotic Susceptibility Testing
The minimal inhibitory concentrations (MICs) of selected antibiotics were determined by the standard serial twofold dilution method in MHA with inocula of 104 CFU per spot, according to the CLSI guidelines (Clinical and Laboratory Standards Institute, 2015). The presence of colonies was visually assessed after 18 h of incubation at 35°C.
Virulence Assays
Motility tests. Swarming and swimming motilities of PAO1 and SM mutants were assessed as previously described (Kohler et al., 2000). Evaluation of twitching motility was performed by vertically inoculating the center of a Luria-Bertani agar plate (1% agar) down to the plastic with a colony collected on a toothpick (Déziel et al., 2001). After 24 h of growth at 37°C, the bacteria adherent to polystyrene were stained with a 1% (wt/vol) solution of crystal violet. Motility assays were all repeated twice. Rhamnolipid production. Production of rhamnolipids was detected by using a cetyltrimethylammonium (CTAB) agar medium (Kohler et al., 2000). Diffusion of rhamnolipids from bacterial spots after 24 h incubation at 37°C generated blue halos in the medium. Hemolytic activity. The capacity of strains to lyse red blood cells was assessed by streaking bacteria on Columbia Blood Agar plates. The test was considered positive when a clear halo was apparent around the colonies after an incubation of 18 h at 37°C.
Determination of Bacterial Growth
Growth curves in different culture media. Bacteria were grown in MHB or TSB, respectively at 30oC and 37oC with shaking (225 rpm). Bacterial density was recorded at DO600nm in covered PS 96-well microplates (Nest), using a Spark 10M microplate reader (Tecan). Biomass determination. Strains were grown aerobically in 50 mL of TSB during 24 h, then pelleted at 11,000 × g for 10 min, and dried at 85°C until the pellet weight remained constant over 72 h. Presented results are means of two independent experiments and are expressed in grams of dehydrated bacteria per L of culture. Bacterial counting. Strains were aerobically cultured in MHB or TSB for 24 h at 30°C or 37°C. Colony forming units were then enumerated on agar plates in quadruplicates, by serial dilutions. Presented results are means of three independent experiments ± standard deviations.
Transmission Electron Microscopy
Bacterial overnight cultures were adjusted at DO600nm = 1 in MHB. Bacteria were recovered by centrifugation prior to fixation 2.5% glutaraldehyde for 1 h at room temperature. Negative staining was done with a 1% aqueous solution of ammonium molybdate. Subsequently, the length of bacteria was measured in 100 randomly selected cells in each sample using a Hitachi H7800 at 80 kV equipped with LaB6 electron source.
Recombinant Protein Production by Arabinose
The gene encoding the mCherry fluorescent protein was cloned on the arabinose-inducible expression vector pJN105 (Newman and Fuqua, 1999). The recombinant plasmid was next introduced into strains PAO1 and SM54 by electroporation. Log phase bacteria were resuspended at a cell density of OD600nm = 0.1 in MHB supplemented with 10 μg mL–1 of gentamicin for PAO1 or 2 μg mL–1 for SM54, and incubated with shaking (225 rpm) for 8 h at 37°C. The fluorescence of induced (0.5% wt/vol arabinose) and non-induced cultures was recorded with a Tecan Spark 10M microplate reader (excitation A590nm/emission A610nm). In parallel, bacterial growth was spectrophotometrically monitored at OD600nm.
Cytotoxic Activity on J774 A.1 Macrophages
Cell viability of J774 A.1 murine macrophages after bacterial infection was determined by using the Cytotoxicity Detection Kit (Sigma Aldrich), following suppliers’ recommendations. Briefly, 4 × 104 cells were cultivated in RPMI 1640 medium (Gibco) supplemented with 1% of fetal bovine serum (FBS, Gibco) at 37°C for 24 h in a humid atmosphere (95%) enriched with 5% CO2. After incubation, macrophages were infected during 3 h with a Multiplicity of Infection of 5 (MOI = 5). The cytotoxicity of each bacterial strain was reported as a percentage of the total cell lysis caused by a Triton X-100 treatment. Tests were performed three times with two replicates.
Adhesion on A549 Human Pulmonary Cells
Adhesion of bacteria was evaluated on human pulmonary cell lines A549. Briefly, 2.5 × 104 cells were cultured in DMEM F-12 medium (Gibco) supplemented with 2 mM GlutaMAX (Gibco) and 10% FBS, at 37oC for 48 h in a humid atmosphere (95%) containing 5% CO2. Once at confluence, cells were infected during 2 h at MOI = 10. Supernatants containing non-adherent bacteria were collected, and cells were washed three times with 100 μL PBS (Gibco). Cell monolayers with adherent bacteria were then lysed using Triton X-100 0.1%. Bacteria of the collected aliquots were counted on MH plates by serial dilution. Bacterial adherence was reported as the percentage of CFU present in cell lysates, compared to the total of CFU counted in aliquots (lysate plus supernatants).
Galleria mellonella Infection Assay
In-house reared Galleria mellonella larvae were infected subcutaneously using a syringe pump (KD scientific) with strains PAO1 and SM54, respectively. Bacteria were collected from overnight cultures by centrifugation, then washed and resuspended in physiological water in order to inoculate 30 CFU into each larva. Twenty insects were used per strain, and the experiments were repeated four times. Larvae killing was then monitored between 19 and 24 h post-infection. Data and statistical analysis were performed using the Kaplan-Meier R package and the logrank test, respectively.
In vivo Evaluation of Pathogenesis in Murine Model
A total of 20 mice CD1 (female 28–32 g) were used to evaluate pathogenesis of strains PAO1 and SM54 of P. aeruginosa. Inocula were prepared by adjusting a bacterial suspension to OD595nm = 0.45 from an overnight solid culture. An additional 1:10 serial dilution was performed using cold physiological water to obtain a solution equivalent to 5 × 107 CFU/mL. Each individual received 500 μL intraperitoneal injections equivalent to 2.5 × 107 CFU. After inoculation, animals were monitored for mortality every 12 h for 4 days. The animal study was reviewed and approved by the Comité D’Ethique Régional (C2EA) and the Ministère de l’Enseignement Supérieur et de la Recherche (ref APAFIS#27692-2020101510342409 v2). Statistical analysis was performed using the logrank test.
Results
Targeted Gene Reduction Strategy
The strategy used in this work for gene deletion was the classical genome editing tool by homologous recombination, using a suicide plasmid with R6K origin and the counter-selection gene sacB responsible for saccharose susceptibility in various Gram-negative bacteria (Kaniga et al., 1991). The main molecular mechanisms driving antibiotic resistance and virulence factor production in P. aeruginosa have been largely studied and characterized (Lister et al., 2009; McCarthy, 2015). However, as P. aeruginosa possesses one of the most complex regulation network (Schuster and Greenberg, 2006; Huang et al., 2019; Kostylev et al., 2019) and some regulation pathways await to be described, targeting regulatory genes was not favored because of the probability to have non-desired effects. Instead, the main effector genes conducing to these traits of pathogenicity were removed in different blocks. First, two blocks of antibiotic resistance-associated genes were targeted; the four clinically significant RND efflux pumps were deleted in strain SMEff, namely MexAB, MexCD, MexEF and MexXY while other resistance genes concerning enzymatic (ampC, aph) and membrane-modification (arnBCDATEFugd) mechanisms were additionally removed in strain SMRes. A third block of gene deletions was performed to obtain strain SMVir in which virulence-associated genes were removed such as those involved in mobility and attachment (fliEFG, pilQ), secreted toxins (plcH, exoT, exoS, exoY, toxA), the Pseudomonas quinolone system (pqsA) and production of rhamnolipids (rhlA) and elastase (lasA, lasB). Finally, in addition to the above mentioned genes, a fourth block was targeted in strain SM54 and consisted in cytotoxicity related genes (pcrVHpopBD) and adhesion related genes (lecA, lecB). In SM54, a total of 37 genes were deleted (≈51 kb) which represents 0.8% of the core genome of the species. A scheme identifying all the effector genes removed in SM54 is shown in Figure 2.
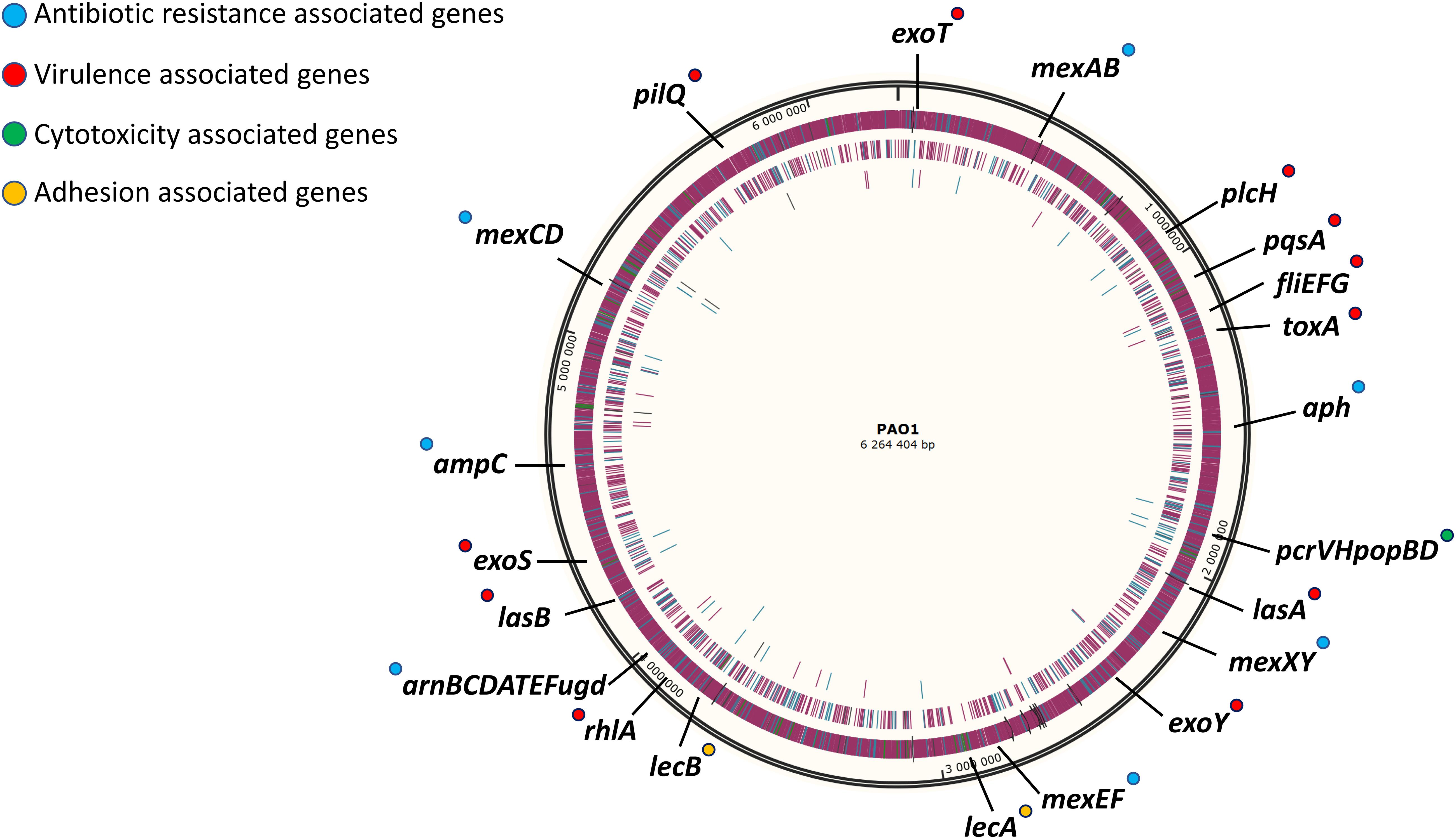
Figure 2. Schematic representation of targeted genomic regions for gene inactivation in strain SM54. Approximative position of the 37 genes deleted from the wild-type strain PAO1 in the physical map of its chromosome (Accession No. NC_002516). Genes related to antibiotic resistance are marked in blue, those related to virulence factors are marked in red, those related to cytotoxicity are marked in green and finally those genes related to adhesion are marked in yellow.
Reduction of Basal Antibiotic Resistance and Virulence Factor Production
As several effector genes were targeted for deletion in the different strains, determination of minimal inhibitory concentrations (MIC) and specific in vitro virulence assays were performed to verify the impairment of antibiotic resistance and production of virulence factors, respectively. As shown in Table 2, the efflux deficient strain SMEff was, from 2- to 32-fold, more susceptible to all antibiotics tested, these included antibiotics commonly used as resistance markers in molecular biology (ampicillin, chloramphenicol, tetracycline, streptomycin, and kanamycin) and clinically used antipseudomonals (gentamicin, tobramycin, ceftazidime, meropenem, aztreonam and colistin). It was interesting to notice that even though several other genes were deleted from SMRes and SM54 strains, only their susceptibility to ampicillin and kanamycin was highly increased (more than 1000-fold and 64-fold, respectively) in comparison to SMEff. Concerning strain SMVir, its susceptibility to antibiotics remained unchanged as expected. The production of several virulence factors was also assessed by performing different in vitro tests. As it was intended, different types of motilities such as swarming, swimming, and twitching dependent on flagellum and pili were abolished in strains SMVir and SM54. The same was observed for the production of hemolytic factors and rhamnolipids (Supplementary Figure 2). Not surprisingly, none of these features were altered in strains SMEff and SMRes (data not shown).
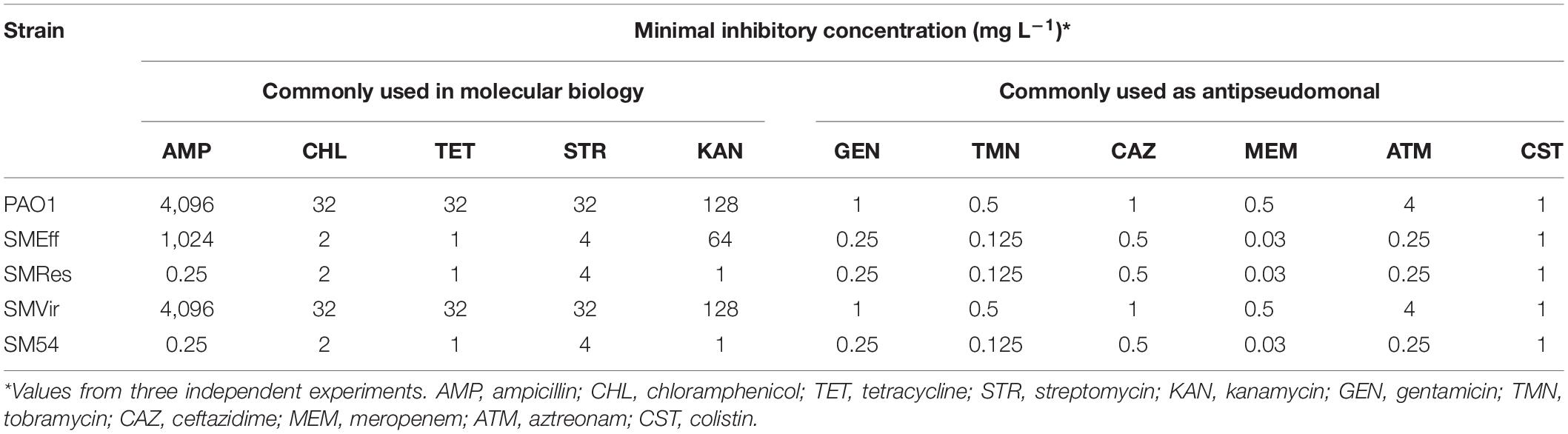
Table 2. Antibiotic susceptibility of selected strains.
Effect of Genome Reduction on Biomass and Bacterial Morphology
As genome reduction may provoke deleterious effects for biotechnological applications such as growth, biomass production and cellular morphology (Kurokawa et al., 2016), some of these parameters were evaluated. Bacterial counting and dry weight determination were thus performed for all the PAO1-derived strains. As P. aeruginosa is a non-fastidious bacterium to growth, two nutritive broths were evaluated: Mueller-Hinton (MHB) and Trypticase Soy (TSB) at two temperatures, 30°C and 37°C. Interestingly, as shown in Figure 3, after 24 h of incubation strains SMVir and SM54 produced significantly more bacterial counting (from 1.3- to 6.1-fold) and more dry weight (from 1.5- to 1.8-fold) compared to the parental strain PAO1. An important remark is that the best performance for these strains was observed when cultured in TSB at 37°C. Concerning strains SMRes and SMEff, their behavior was very similar to that of the wild-type strain PAO1. Finally, cell morphology was studied by transmission electron microscopy to evaluate if genomic modifications could modify this trait. Even though, 0.8% of the genome was withdrawn in SM54, no significant changes were observed compared to PAO1 (p-value >0.05) (Supplementary Figure 3).

Figure 3. Bacterial counting and dry weight at 24 h of growth. Bacterial cells were counted after 24 h of culture at 30°C (light gray) or at 37°C (dark gray) in (A) Mueller-Hinton Broth and (B) Trypticase Soy Broth. All cultures were agitated at 225 rpm to allow proper oxygenation. Results represent mean values of three independent experiments. Error bars represent standard deviation and asterisks (*) represent a significant difference in the number of CFU counted between the two strains compared according to a Student’s t test (p < 0.01). Below each graph, dry weights produced by these cultures are indicated.
Genome Reduction Does Not Impair Heterologous Protein Production in SM54
As strain SM54 showed interesting characteristics in all the aspects studied above, a step forward was taken with this strain to evaluate other features. The natural capacity of the modified strain to produce heterologous proteins was evaluated as this feature might be also affected by genome reduction (Lieder et al., 2015; Zhu et al., 2017). For this, the encoding sequence of the red fluorescent protein mCherry was inserted into the arabinose-inducing plasmid pJN105 and fluorescence was followed during 8 h. As shown in Figure 4 the ratio fluorescence/DO600 emitted by SM54 harboring this recombinant plasmid was very similar to that produced by the parental strain PAO1, thus providing evidence that the smaller genome of SM54 does not significantly impair protein synthesis.
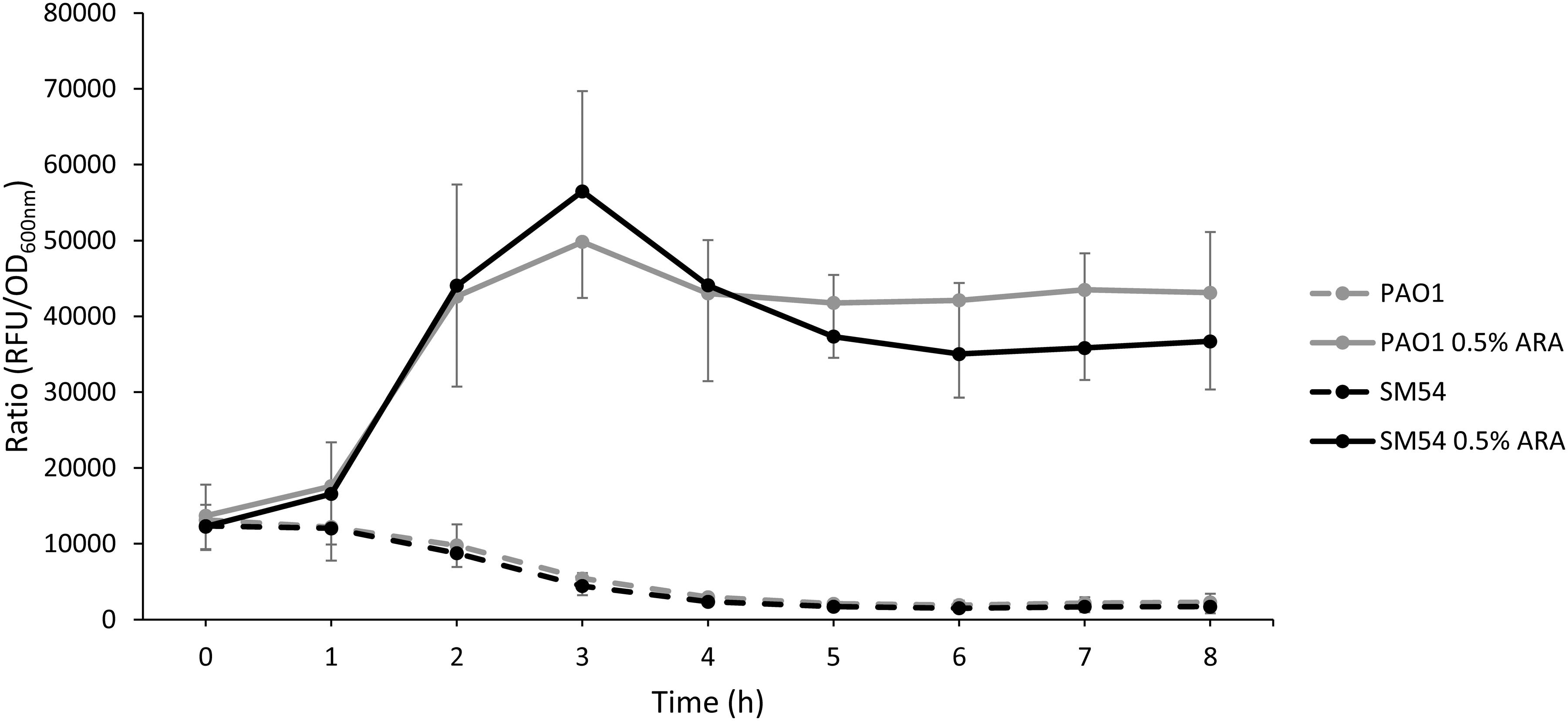
Figure 4. Accumulation of the red fluorescent protein mCherry. Strains PAO1 and SM54 were grown in Mueller-Hinton broth containing 10 or 2 μg mL– 1 of Gentamicin, respectively, used as selection marker of plasmid pJN105. The transcription of the mCherry coding gene was induced with 0.5% of arabinose and the accumulation of the fluorescent protein was monitored using a microplate reader (Ex 590 nm, Em 610 nm) (solid lines). Non-induced cultures were used as controls (dashed lines). Results represent mean values of three independent experiments and error bars represent standard deviation.
In vitro and in vivo Evaluation of SM54 Pathogenicity
As SM54 showed potential for further biotechnological applications, it seemed to be essential to evaluate additional traits of pathogenicity. Thus, in vitro assays of cytotoxicity and cell adhesion were performed using murine macrophages (J774 A.1 cells) and human pulmonary cells (A549 cells), respectively. Once again, strain SM54 showed a reduction of these two pathogenic traits (Figure 5). A significant decrease of cytotoxicity was observed compared to PAO1 (27 vs 80%, respectively. p-value <0.05). On the other hand, concerning adhesion to pulmonary cells, even though a reduction of this trait was noted, the difference between these two strains was not statistically significant (PAO1 86% vs SM54 65%; p-value >0.05). Finally, to test whether the pathogenicity of SM54 was significantly attenuated, two in vivo models were studied. Firstly, larvae of Galleria mellonella were infected with 30 CFU and secondly CD1 mice were challenged with intraperitoneal injections of 2.5 × 107 CFU of each strain; for both cases survival of individuals was followed over time (Figure 5). In the case of G. mellonella infection, it was observed that larvae infected with SM54 significantly survived 50% better than those infected with PAO1 after 24 h of infection (p-value <0.001). For CD1 mice infection, even though 100% of mortality was observed after 48 h of infection, the mortality of individuals infected with SM54 was significantly slower than those infected with PAO1 (0 vs 80% at 15 h, p-value <0.05).
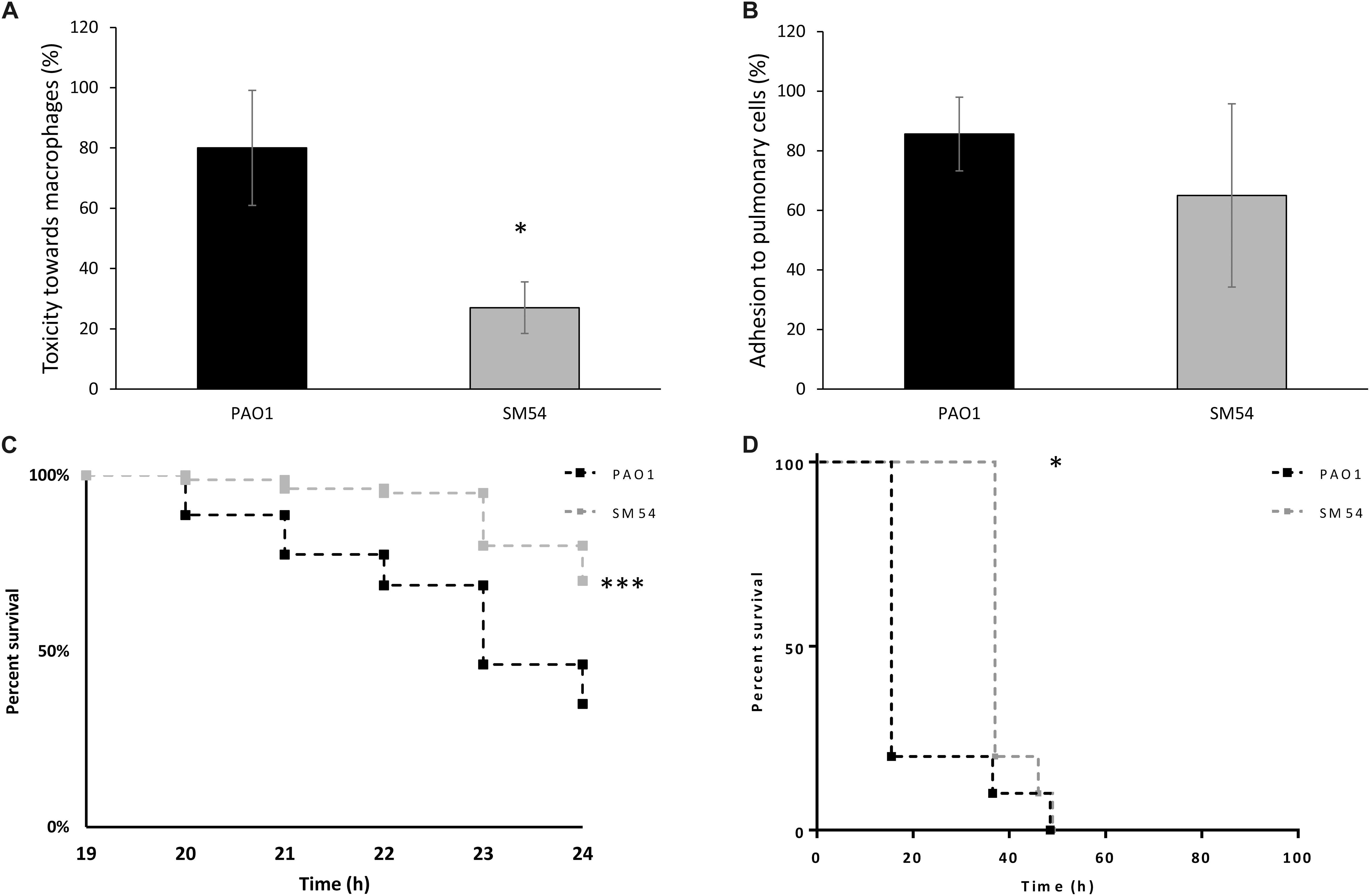
Figure 5. Evaluation of SM54 pathogenesis. (A) cytotoxicity tests performed on J774 A.1 murine macrophages using a MOI of 5. Results represent mean values of three independent experiments for which a significant difference was shown according to the Mann-Whitney test (p < 0.01) (*). (B) Adhesion tests performed on A549 human pulmonary cells using a MOI of 10. Results represent mean values of three independent experiments. Error bars represent standard deviation. (C) Kaplan-Meier survival curves of G. mellonella larvae infected with the WT strain PAO1 (black) or SM54 (gray). Results represent mean values from four independent experiments for which a significantly difference (***) was shown according to the logrank test (p < 0.001). (D) Kaplan-Meier survival curves of mice CD3 infected with the WT strain PAO1 (black) or SM54 (gray). Results represent the survival of 20 individuals for each condition. Survival between the two groups of organisms was shown to be significantly different after 15 h (*) of infection according to the logrank test (p < 0.05).
Discussion
Several biotechnological applications of the genus Pseudomonas have been largely studied, especially for P. putida (Poblete-Castro et al., 2012; Molina et al., 2013; Martinez-Garcia and de Lorenzo, 2019; Weimer et al., 2020). The species of this genus are highly adaptative to a wide-range of environments fact that could be explained by their large genome containing variable metabolic pathways which become active in response to the environmental conditions in which the bacterium is growing (Silby et al., 2011; Mielko et al., 2019). In the case of P. aeruginosa, the biotechnological applications have been poorly studied mainly because its character of opportunistic pathogen. In this study, we have sought to develop an attenuated strain derived from P. aeruginosa PAO1 as a first step to create a chassis for biotechnological applications. Genome reduction was the strategy used to target specific effector genes related to antibiotic resistance and pathogenicity. Thus, strain SM54 was engineered by sequentially deleting four blocks of genes related to these traits. Additionally, three other strains are also described, SMEff, SMRes and SMVir, which could be helpful to study isolated mechanisms of antibiotic resistance or virulence. Overall, the effects of these genomic deletions on antibiotic resistance and virulence showed the expected results. In detail, our results showed that when antibiotic related genes were targeted, as in SMEff, SMRes or SM54, the basal susceptibility to antibiotics was highly increased and, on the same logic, when virulence factor genes were deleted in strains SMVir or SM54 their capacities to produce these factors was abolished. It is important to notice that not all pathogenic traits were targeted in this first stage development. For example, another well-characterized determinant of antibiotic resistance of P. aeruginosa is gene oprD (PA0958) (Lister et al., 2009). It was here considered not remove oprD from SM54 as its deletion would result in a decrease of strain susceptibility to carbapenems. Indeed, this gene codes for the OprD substrate-specific porin which serves as the main portal of entry for basic amino acids and carbapenems (Fukuoka et al., 1993) and which when altered results in significant resistance to these antibiotics. Furthermore, P. aeruginosa also harbors in its genome 8 additional RND efflux systems (representing ∼ 44.8 kb) for which the contribution to antibiotic resistance and bacterial physiology has been poorly studied (Poole, 2014). For this reason, it was considered here that they should not be removed from SM54 to avoid undesirable effects on fitness or physiology. As previous studies have shown that genome reduction might have either positive or negative effects on bacterial development (Lieder et al., 2015; Kurokawa et al., 2016), we studied the effect of these modifications on biomass and bacterial morphology. Our results showed that when virulence related genes are deleted, the dry weight and the number of CFUs recovered after 24 h of culture are significantly increased when compared to strain PAO1. These results suggest that strains SMVir and SM54 may eventually produce larger amounts of biomass than a wild-type strain when cultured in rich media such as MH or TSB. It is important to notice that this yield could be modified following the nutrient level of the medium used (e.g., rich vs minimal media) as it was shown for genome reduced E. coli (Kurokawa et al., 2016; Tsuchiya et al., 2018) and further characterization of SM54 will explore this behavior. Concerning TEM observation of bacterial cells, even though the difference between PAO1 and SM54 was not statistically significant, SM54 cells are slightly bigger in length and width which is in accordance with other studies evaluating the impact of genome reduction on cell morphology (Hutchison et al., 2016; Kurokawa et al., 2016). Based on these observations, it would be expected that if additional deletions are made to SM54 genome, the morphology of the cells would have tendance to be bigger. Previous work done on P. putida showed that following genome reduction, the capacities of the strain to produce heterologous proteins were enhanced (Lieder et al., 2015). Following the same line of thinking, we evaluate the capacity of SM54 to produce the fluorescent protein mCherry after induction with arabinose. The difference between PAO1 and SM54 was not significant (p-value >0.05) suggesting that the genomic modifications done to SM54 did not have any effect on this trait. The deletion of non-essential genes was shown to improve the expression of heterologous genes in P. putida (Martinez-Garcia et al., 2014) it would be interesting to explore if such deletions would have the same positive effects on SM54. Bioinformatic analysis has shown that SM54 still harbors genes coding for two prophages (cluster 1 from PA0617 to PA0641 and cluster 2 from PA0720 to PA0728), two endonucleases (endA, nth), one restriction-modification system (hdsMSR) and about 7 kb coding for insertion sequences representing a total of ∼43 kb. Targeting these regions would be the next step to consider in the improvement of SM54 toward the application of recombinant protein production. Regarding pathogenesis experiments, our in vitro results showed that SM54 is no longer capable to induce cell death when tested against murine macrophages and is less adhesive to human pulmonary cells. We sought to further reduce the adhesion of SM54 by deleting genes related to biofilm formation (chaperone usher systems- strain SM59) (Vallet et al., 2001; Giraud and de Bentzmann, 2012). However, in SM59, bacterial growth was importantly impacted (data not shown) and deletion of the cup clusters provoked clumps in liquid cultures and roughly colonies clearly; showing that some deletions can be deleterious for bacterial fitness and further applications. Regarding these results, we kept SM54 to evaluate its pathogenesis in vivo using Galleria mellonella larvae and CD1 mice as models. The first has been mainly used to evaluate the response of innate immunity to bacterial infections (Tsai et al., 2016) and more specifically to study the role of the type III secretion system of P. aeruginosa pathogenesis (Miyata et al., 2003). The murine model has been now extensively used for study a wide variety of bacterial infections and allows the study of both innate and adaptative immune responses (Stotland et al., 2000). Although our results showed a decreased of mortality in both models, individuals infected with SM54 kept dying at a slower rate than those infected with PAO1. Recently, it has been showed that targeting five key pathogenicity genes from the P. aeruginosa chromosome was enough to produce an attenuated strain incapable to kill mice (Valentine et al., 2020). Among these genes, phzM encoding for a phenazine-specific methyltransferase required for pyocyanin production (Rada and Leto, 2013) and wapR encoding for a rhamnosyltransferase involved in the synthesis of the O antigen of LPS (Poon et al., 2008) could be two interesting targets to delete from SM54 to finally produce an attenuated strain. Overall, further genetic modifications of strain SM54 is undeniably needed to reduce its pathogenesis and to improve its capacities for heterologous gene expression. It should be noted that these modifications could have an impact on bacterial fitness and choices would be needed to equilibrate efficacy and yield of the desired product. The development of bacterial chassis requires a certain degree of engineering for which the actual state of SM54 might not be suitable to use this terminology. Indeed, as Synthetic Biology develops as science, the community tries to establish what a bacterial chassis is. Recently, it has been proposed that the term “chassis” should imply that the organisms labeled as such have undergone through several degrees of optimization from a genomic and metabolomic points of view. This kind of organisms should have a genome encoding a number of basic functions for stable self-maintenance, growth and optimal operation under specific environmental conditions (de Lorenzo et al., 2021). Following this definition, it is clear that SM54 is not and should not be called a chassis but rather a rDNA host for which one can use or develop specific plasmids to study heterologous gene expression, protein function or other fundamental studies. It is important to notice that by developing an attenuated strain, it is possible to make a pathogen like P. aeruginosa user-friendly and suitable for the use of classical tools of molecular biology (e.g., antibiotic cassettes). From a medical point of view, some studies have raised the possibility to use attenuated bacteria to deliver DNA plasmid vaccines (Dietrich et al., 2003). This approach has been already been explored for P. aeruginosa in which an attenuated strain deleted of gene aroA, involved in the synthesis of aromatic amino acids, was proven to be protective in a pre-clinical setting after challenging mice using models of corneal and acute lung infection (Priebe et al., 2002, 2003; Zaidi et al., 2006; Kamei et al., 2011). One can imagine that developing an unarmed chassis of P. aeruginosa such as SM54 could be a novel approach to further engineer this strain and explore this kind of medical applications. Even though the early stage of this project, SM54 proves to have some qualities which can be of interest for further optimization. As mentioned above, additional genomic modifications are planned (retirement of prophages, endonucleases, restriction-modification systems, insertion sequences, and abolishment of phenazine production and O antigen synthesis) to make of SM54 a safe-bacterial platform.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Ethics Statement
The animal study was reviewed and approved by Comité D’Ethique Régional (C2EA) and the Ministère de l’Enseignement Supérieur et de la Recherche (ref APAFIS#27692-2020101510342409 v2).
Author Contributions
MG, SG, and PJ performed the experimental procedures. CG performed in vivo survival tests on Galleria mellonella. CM, PP, and PJ designed the experiments. PJ wrote the manuscript. All the authors read the manuscript and agreed with its submission.
Funding
MG was supported with grants from the French “Association Nationale de la Recherche et de la Technologie” through the 2017/0994 CIFRE agreement between Smaltis and UMR 6249 Chrono-Environnement from Université Bourgogne Franche-Comté.
Conflict of Interest
This work was done under industrial support from SMALTIS SAS; the distribution of biological material will be regulated by the establishment of Material Transfer Agreements after discussion with the company authorities.
Acknowledgments
The authors acknowledge Delphine Croisier (Vivexia S.A.R.L, Résidence Richelieu, 10 Blv Carnot, 21000 Dijon, France) for the evaluation of in vivo pathogenesis of strains in murine model and Laure Avoscan (Plate-forme DImaCell, AgroSup Dijon, INRAE, Univ. Bourgogne Franche-Comté, F-21000 Dijon, France) for TEM observations and analysis.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fbioe.2021.640450/full#supplementary-material
Footnotes
References
Berrazeg, M., Jeannot, K., Ntsogo Enguene, V. Y., Broutin, I., Loeffert, S., Fournier, D., et al. (2015). Mutations in beta-lactamase AmpC increase resistance of Pseudomonas aeruginosa isolates to antipseudomonal cephalosporins. Antimicrob. Agents Chemother. 59, 6248–6255. doi: 10.1128/AAC.00825-15
Burrows, L. L. (2012). Pseudomonas aeruginosa twitching motility: type IV pili in action. Annu. Rev. Microbiol. 66, 493–520. doi: 10.1146/annurev-micro-092611-150055
Chemani, C., Imberty, A., de Bentzmann, S., Pierre, M., Wimmerova, M., Guery, B. P., et al. (2009). Role of lecA and lecB lectins in Pseudomonas aeruginosa-induced lung injury and effect of carbohydrate ligands. Infect. Immun. 77, 2065–2075. doi: 10.1128/IAI.01204-08
Clinical and Laboratory Standards Institute (2015). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard, 10th Edn. Wayne, PA: Clinical and Laboratory Standards Institute.
Dacheux, D., Goure, J., Chabert, J., Usson, Y., and Attrée, I. (2001). Pore-forming activity of type III system-secreted proteins leads to oncosis of Pseudomonas aeruginosa-infected macrophages. Mol. Microbiol. 40, 76–85. doi: 10.1046/j.1365-2958.2001.02368.x
de Bentzmann, S., and Plésiat, P. (2011). The Pseudomonas aeruginosa opportunistic pathogen and human infections. Environ. Microbiol. 13, 1655–1665. doi: 10.1111/j.1462-2920.2011.02469.x
de Lorenzo, V., Krasnogor, N., and Schmidt, M. (2021). For the sake of the bioeconomy: define what a synthetic biology chassis is! N. Biotechnol. 60, 44–51. doi: 10.1016/j.nbt.2020.08.004
Dennis, J. J., and Sokol, P. A. (1995). Electrotransformation of Pseudomonas. Methods Mol. Biol. 47, 125–133. doi: 10.1385/0-89603-310-4:125
Déziel, E., Comeau, Y., and Villemur, R. (2001). Initiation of biofilm formation by Pseudomonas aeruginosa 57RP correlates with emergence of hyperpiliated and highly adherent phenotypic variants deficient in swimming, swarming, and twitching motilities. J. Bacteriol. 183, 1195–1204. doi: 10.1128/JB.183.4.1195-1204.2001
Dietrich, G., Spreng, S., Favre, D., Viret, J. F., and Guzman, C. A. (2003). Live attenuated bacteria as vectors to deliver plasmid DNA vaccines. Curr. Opin. Mol. Ther. 5, 10–19.
Diggle, S. P., Stacey, R. E., Dodd, C., Camara, M., Williams, P., and Winzer, K. (2006). The galactophilic lectin, lecA, contributes to biofilm development in Pseudomonas aeruginosa. Environ. Microbiol. 8, 1095–1104. doi: 10.1111/j.1462-2920.2006.001001.x
Ditta, G., Stanfield, S., Corbin, D., and Helinski, D. R. (1980). Broad host range DNA cloning system for gram-negative bacteria: construction of a gene bank of Rhizobium meliloti. Proc. Natl. Acad. Sci. U.S.A. 77, 7347–7351. doi: 10.1073/pnas.77.12.7347
Feldman, M., Bryan, R., Rajan, S., Scheffler, L., Brunnert, S., Tang, H., et al. (1998). Role of flagella in pathogenesis of Pseudomonas aeruginosa pulmonary infection. Infect. Immun. 66, 43–51. doi: 10.1128/IAI.66.1.43-51.1998
Filloux, A. (2011). Protein secretion systems in Pseudomonas aeruginosa: an essay on diversity, evolution, and function. Front. Microbiol. 2:155. doi: 10.3389/fmicb.2011.00155
Fukuoka, T., Ohya, S., Narita, T., Katsuta, M., Iijima, M., Masuda, N., et al. (1993). Activity of the carbapenem panipenem and role of the OprD (D2) protein in its diffusion through the Pseudomonas aeruginosa outer membrane. Antimicrob. Agents Chemother. 37, 322–327. doi: 10.1128/AAC.37.2.322
Giraud, C., and de Bentzmann, S. (2012). Inside the complex regulation of Pseudomonas aeruginosa chaperone usher systems. Environ. Microbiol. 14, 1805–1816. doi: 10.1111/j.1462-2920.2011.02673.x
Hauser, A. R. (2009). The type III secretion system of Pseudomonas aeruginosa: infection by injection. Nat. Rev. Microbiol. 7, 654–665. doi: 10.1038/nrmicro2199
Herrero, M., de Lorenzo, V., and Timmis, K. N. (1990). Transposon vectors containing non-antibiotic resistance selection markers for cloning and stable chromosomal insertion of foreign genes in gram-negative bacteria. J. Bacteriol. 172, 6557–6567. doi: 10.1128/JB.172.11.6557-6567.1990
Hilker, R., Munder, A., Klockgether, J., Losada, P. M., Chouvarine, P., Cramer, N., et al. (2015). Interclonal gradient of virulence in the Pseudomonas aeruginosa pangenome from disease and environment. Environ. Microbiol. 17, 29–46. doi: 10.1111/1462-2920.12606
Huang, H., Shao, X., Xie, Y., Wang, T., Zhang, Y., Wang, X., et al. (2019). An integrated genomic regulatory network of virulence-related transcriptional factors in Pseudomonas aeruginosa. Nat. Commun. 10:2931. doi: 10.1038/s41467-019-10778-w
Hutchison, C. A. III, Chuang, R. Y., Noskov, V. N., Assad-Garcia, N., Deerinck, T. J., Ellisman, M. H., et al. (2016). Design and synthesis of a minimal bacterial genome. Science 351:aad6253. doi: 10.1126/science.aad6253
Jeannot, K., Bolard, A., and Plesiat, P. (2017). Resistance to polymyxins in Gram-negative organisms. Int. J. Antimicrob. Agents 49, 526–535. doi: 10.1016/j.ijantimicag.2016.11.029
Juan, C., Macia, M. D., Gutierrez, O., Vidal, C., Perez, J. L., and Oliver, A. (2005). Molecular mechanisms of beta-lactam resistance mediated by ampC hyperproduction in Pseudomonas aeruginosa clinical strains. Antimicrob. Agents Chemother. 49, 4733–4738. doi: 10.1128/AAC.49.11.4733-4738.2005
Juan, C., Moya, B., Perez, J. L., and Oliver, A. (2006). Stepwise upregulation of the Pseudomonas aeruginosa chromosomal cephalosporinase conferring high-level beta-lactam resistance involves three AmpD homologues. Antimicrob. Agents Chemother. 50, 1780–1787. doi: 10.1128/AAC.50.5.1780-1787.2006
Kamei, A., Coutinho-Sledge, Y. S., Goldberg, J. B., Priebe, G. P., and Pier, G. B. (2011). Mucosal vaccination with a multivalent, live-attenuated vaccine induces multifactorial immunity against Pseudomonas aeruginosa acute lung infection. Infect. Immun. 79, 1289–1299. doi: 10.1128/IAI.01139-10
Kaniga, K., Delor, I., and Cornelis, G. R. (1991). A wide-host-range suicide vector for improving reverse genetics in gram-negative bacteria: inactivation of the blaA gene of Yersinia enterocolitica. Gene 109, 137–141. doi: 10.1016/0378-1119(91)90599-7
Kazmierczak, B. I., Schniederberend, M., and Jain, R. (2015). Cross-regulation of Pseudomonas motility systems: the intimate relationship between flagella, pili and virulence. Curr. Opin. Microbiol. 28, 78–82. doi: 10.1016/j.mib.2015.07.017
King, J. D., Kocincova, D., Westman, E. L., and Lam, J. S. (2009). Review: lipopolysaccharide biosynthesis in Pseudomonas aeruginosa. Innate Immun. 15, 261–312. doi: 10.1177/1753425909106436
Kohler, T., Curty, L. K., Barja, F., van Delden, C., and Pechere, J. C. (2000). Swarming of Pseudomonas aeruginosa is dependent on cell-to-cell signaling and requires flagella and pili. J. Bacteriol. 182, 5990–5996. doi: 10.1128/JB.182.21.5990-5996.2000
Kostylev, M., Kim, D. Y., Smalley, N. E., Salukhe, I., Greenberg, E. P., and Dandekar, A. A. (2019). Evolution of the Pseudomonas aeruginosa quorum-sensing hierarchy. Proc. Natl. Acad. Sci. U.S.A. 116, 7027–7032. doi: 10.1073/pnas.1819796116
Kurokawa, M., Seno, S., Matsuda, H., and Ying, B. W. (2016). Correlation between genome reduction and bacterial growth. DNA Res. 23, 517–525. doi: 10.1093/dnares/dsw035
Lacks, S., and Greenberg, B. (1977). Complementary specificity of restriction endonucleases of Diplococcus pneumoniae with respect to DNA methylation. J. Mol. Biol. 114, 153–168. doi: 10.1016/0022-2836(77)90289-3
Li, X. Z., Plésiat, P., and Nikaido, H. (2015). The challenge of efflux-mediated antibiotic resistance in gram-negative bacteria. Clin. Microbiol. Rev. 28, 337–418. doi: 10.1128/CMR.00117-14
Lieder, S., Níkel, P. I., de Lorenzo, V., and Takors, R. (2015). Genome reduction boosts heterologous gene expression in Pseudomonas putida. Microb. Cell Fact. 14:23. doi: 10.1186/s12934-015-0207-7
Lister, P. D., Wolter, D. J., and Hanson, N. D. (2009). Antibacterial-resistant Pseudomonas aeruginosa: clinical impact and complex regulation of chromosomally encoded resistance mechanisms. Clin. Microbiol. Rev. 22, 582–610. doi: 10.1128/CMR.00040-09
Manoil, C., and Beckwith, J. (1985). TnphoA: a transposon probe for protein export signals. Proc. Natl. Acad. Sci. U.S.A. 82, 8129–8133. doi: 10.1073/pnas.82.23.8129
Martinez-Garcia, E., and de Lorenzo, V. (2019). Pseudomonas putida in the quest of programmable chemistry. Curr. Opin. Biotechnol. 59, 111–121. doi: 10.1016/j.copbio.2019.03.012
Martinez-Garcia, E., Nikel, P. I., Aparicio, T., and de Lorenzo, V. (2014). Pseudomonas 2.0: genetic upgrading of P. putida KT2440 as an enhanced host for heterologous gene expression. Microb. Cell Fact. 13:159. doi: 10.1186/s12934-014-0159-3
McCarthy, K. (2015). Pseudomonas aeruginosa: evolution of antimicrobial resistance and implications for therapy. Semin. Respir. Crit. Care Med. 36, 44–55. doi: 10.1055/s-0034-1396907
Mielko, K. A., Jablonski, S. J., Milczewska, J., Sands, D., Lukaszewicz, M., and Mlynarz, P. (2019). Metabolomic studies of Pseudomonas aeruginosa. World J. Microbiol. Biotechnol. 35:178. doi: 10.1007/s11274-019-2739-1
Miyata, S., Casey, M., Frank, D. W., Ausubel, F. M., and Drenkard, E. (2003). Use of the Galleria mellonella caterpillar as a model host to study the role of the type III secretion system in Pseudomonas aeruginosa pathogenesis. Infect. Immun. 71, 2404–2413. doi: 10.1128/IAI.71.5.2404-2413.2003
Molina, G., Pimentel, M. R., and Pastore, G. M. (2013). Pseudomonas: a promising biocatalyst for the bioconversion of terpenes. Appl. Microbiol. Biotechnol. 97, 1851–1864. doi: 10.1007/s00253-013-4701-8
Moradali, M. F., Ghods, S., and Rehm, B. H. (2017). Pseudomonas aeruginosa lifestyle: a paradigm for adaptation, survival, and persistence. Front. Cell. Infect. Microbiol. 7:39. doi: 10.3389/fcimb.2017.00039
Newman, J. R., and Fuqua, C. (1999). Broad-host-range expression vectors that carry the L-arabinose-inducible Escherichia coli araBAD promoter and the araC regulator. Gene 227, 197–203. doi: 10.1016/S0378-1119(98)00601-5
Poblete-Castro, I., Becker, J., Dohnt, K., dos Santos, V. M., and Wittmann, C. (2012). Industrial biotechnology of Pseudomonas putida and related species. Appl. Microbiol. Biotechnol. 93, 2279–2290. doi: 10.1007/s00253-012-3928-0
Poole, K. (2004). Efflux-mediated multiresistance in gram-negative bacteria. Clin. Microbiol. Infect. 10, 12–26. doi: 10.1111/j.1469-0691.2004.00763.x
Poole, K. (2014). Stress responses as determinants of antimicrobial resistance in Pseudomonas aeruginosa: multidrug efflux and more. Can. J. Microbiol. 60, 783–791. doi: 10.1139/cjm-2014-0666
Poon, K. K., Westman, E. L., Vinogradov, E., Jin, S., and Lam, J. S. (2008). Functional characterization of MigA and WapR: putative rhamnosyltransferases involved in outer core oligosaccharide biosynthesis of Pseudomonas aeruginosa. J. Bacteriol. 190, 1857–1865. doi: 10.1128/JB.01546-07
Priebe, G. P., Brinig, M. M., Hatano, K., Grout, M., Coleman, F. T., Pier, G. B., et al. (2002). Construction and characterization of a live, attenuated aroA deletion mutant of Pseudomonas aeruginosa as a candidate intranasal vaccine. Infect. Immun. 70, 1507–1517. doi: 10.1128/IAI.70.3.1507-1517.2002
Priebe, G. P., Meluleni, G. J., Coleman, F. T., Goldberg, J. B., and Pier, G. B. (2003). Protection against fatal Pseudomonas aeruginosa pneumonia in mice after nasal immunization with a live, attenuated aroA deletion mutant. Infect. Immun. 71, 1453–1461. doi: 10.1128/IAI.71.3.1453-1461.2003
Rada, B., and Leto, T. L. (2013). Pyocyanin effects on respiratory epithelium: relevance in Pseudomonas aeruginosa airway infections. Trends Microbiol. 21, 73–81. doi: 10.1016/j.tim.2012.10.004
Ropy, A., Cabot, G., Sanchez-Diener, I., Aguilera, C., Moya, B., Ayala, J. A., et al. (2015). Role of Pseudomonas aeruginosa low-molecular-mass penicillin-binding proteins in AmpC expression, beta-lactam resistance, and peptidoglycan structure. Antimicrob. Agents Chemother. 59, 3925–3934. doi: 10.1128/AAC.05150-14
Schuster, M., and Greenberg, E. P. (2006). A network of networks: quorum-sensing gene regulation in Pseudomonas aeruginosa. Int. J. Med. Microbiol. 296, 73–81. doi: 10.1016/j.ijmm.2006.01.036
Silby, M. W., Winstanley, C., Godfrey, S. A., Levy, S. B., and Jackson, R. W. (2011). Pseudomonas genomes: diverse and adaptable. FEMS Microbiol. Rev. 35, 652–680. doi: 10.1111/j.1574-6976.2011.00269.x
Stotland, P. K., Radzioch, D., and Stevenson, M. M. (2000). Mouse models of chronic lung infection with Pseudomonas aeruginosa: models for the study of cystic fibrosis. Pediatr. Pulmonol. 30, 413–424. doi: 10.1002/1099-0496(200011)30:5<413::AID-PPUL8>3.0.CO;2-9
Stover, C. K., Pham, X. Q., Erwin, A. L., Mizoguchi, S. D., Warrener, P., Hickey, M. J., et al. (2000). Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nature 406, 959–964. doi: 10.1038/35023079
Tsai, C. J., Loh, J. M., and Proft, T. (2016). Galleria mellonella infection models for the study of bacterial diseases and for antimicrobial drug testing. Virulence 7, 214–229. doi: 10.1080/21505594.2015.1135289
Tsuchiya, K., Cao, Y. Y., Kurokawa, M., Ashino, K., Yomo, T., and Ying, B. W. (2018). A decay effect of the growth rate associated with genome reduction in Escherichia coli. BMC Microbiol. 18:101. doi: 10.1186/s12866-018-1242-4
Valentine, M. E., Kirby, B. D., Withers, T. R., Johnson, S. L., Long, T. E., Hao, Y., et al. (2020). Generation of a highly attenuated strain of Pseudomonas aeruginosa for commercial production of alginate. Microb. Biotechnol. 13, 162–175. doi: 10.1111/1751-7915.13411
Vallet, I., Olson, J. W., Lory, S., Lazdunski, A., and Filloux, A. (2001). The chaperone/usher pathways of Pseudomonas aeruginosa: identification of fimbrial gene clusters (cup) and their involvement in biofilm formation. Proc. Natl. Acad. Sci. U.S.A. 98, 6911–6916. doi: 10.1073/pnas.111551898
Vettoretti, L., Plesiat, P., Muller, C., El Garch, F., Phan, G., Attree, I., et al. (2009). Efflux unbalance in Pseudomonas aeruginosa isolates from cystic fibrosis patients. Antimicrob. Agents Chemother. 53, 1987–1997. doi: 10.1128/AAC.01024-08
Weimer, A., Kohlstedt, M., Volke, D. C., Nikel, P. I., and Wittmann, C. (2020). Industrial biotechnology of Pseudomonas putida: advances and prospects. Appl. Microbiol. Biotechnol. 104, 7745–7766. doi: 10.1007/s00253-020-10811-9
Wilder, C. N., Diggle, S. P., and Schuster, M. (2011). Cooperation and cheating in Pseudomonas aeruginosa: the roles of the las, rhl and pqs quorum-sensing systems. ISME J. 5, 1332–1343. doi: 10.1038/ismej.2011.13
Yonezawa, M., Takahata, M., Matsubara, N., Watanabe, Y., and Narita, H. (1995). DNA gyrase gyrA mutations in quinolone-resistant clinical isolates of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 39, 1970–1972. doi: 10.1128/AAC.39.9.1970
Zaidi, T. S., Priebe, G. P., and Pier, G. B. (2006). A live-attenuated Pseudomonas aeruginosa vaccine elicits outer membrane protein-specific active and passive protection against corneal infection. Infect. Immun. 74, 975–983. doi: 10.1128/IAI.74.2.975-983.2006
Keywords: Pseudomonas aeruginosa, genome reduction, virulence, antibiotic resistance, biotechnology
Citation: Grosjean M, Guénard S, Giraud C, Muller C, Plésiat P and Juarez P (2021) Targeted Genome Reduction of Pseudomonas aeruginosa Strain PAO1 Led to the Development of Hypovirulent and Hypersusceptible rDNA Hosts. Front. Bioeng. Biotechnol. 9:640450. doi: 10.3389/fbioe.2021.640450
Received: 11 December 2020; Accepted: 18 February 2021;
Published: 11 March 2021.
Edited by:
Jose Ruben Morones-Ramirez, Autonomous University of Nuevo León, MexicoReviewed by:
Thomas Dandekar, Julius Maximilian University of Würzburg, GermanyLuis G. C. Pacheco, Federal University of Bahia, Brazil
Copyright © 2021 Grosjean, Guénard, Giraud, Muller, Plésiat and Juarez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paulo Juarez, cGF1bG8uanVhcmV6QHNtYWx0aXMuZnI=