 Ailén Chuchuy1†
Ailén Chuchuy1† Marcela S. Rodriguero
Marcela S. Rodriguero- 1Centro de Estudios Parasitológicos y de Vectores (CEPAVE-CCT-La Plata-CONICET-UNLP), La Plata, Argentina
- 2Instituto de Ecología, Genética y Evolución (IEGEBA), CONICET—Universidad de Buenos Aires, Buenos Aires, Argentina
The intracellular bacterium Wolbachia pipientis has emerged as a promising tool for controlling mosquito-borne diseases; however, key aspects of its biology remain insufficiently understood, particularly how Wolbachia influences vector competence for certain arboviruses. The main factors implicated are the activation of mosquito antiviral pathways and competition for cellular resources at the viral replication site. Transinfection of Wolbachia strains into vector populations has proven to be an effective strategy for controlling arboviral diseases. Here, we investigate the within-host density and tissue distribution of two naturally occurring Wolbachia strains—wAlbA and wAlbB—n Aedes albopictus from Argentina, where infection patterns diverge from those observed globally. Using quantitative PCR, we assessed symbiont density in ovarian (n = 5) and somatic tissues (n = 5) of adult females, and in adult males across different ages: 0, 5 and 14 days post-emergence (n = 5 per age group). Our results reveal superinfection in ovaries (wAlbA + wAlbB) with similar densities (median relative densitywAlbA = 3.78 and median relative densitywAlbB = 3.31), but only wAlbB was consistently detected in somatic tissues (median relative densitywAlbB = 0.41), suggesting tissue-specific distribution of strains. Additionally, wAlbB density in males remained stable throughout the adult lifespan (median relative densityTime0 = 0.83; median relative densitytime 5 = 1.98; median relative densitytime 14 = 0.66). These findings support the hypothesis that Wolbachia somatic localization is strain-specific and may be under evolutionary selection, with implications for vertical transmission and host fitness. By advancing our understanding of Wolbachia density dynamics in a natural mosquito vector population, this study contributes critical baseline data to inform and optimize Wolbachia-based biocontrol strategies in regions at risk of arboviral outbreaks. Because the wAlbB strain from Ae. albopictus is widely used in replacement techniques, any knowledge of its behavior in natural host populations is valuable.
1 Introduction
The recent emergence and re-emergence of mosquito-borne diseases (MBD) like yellow fever (YF), dengue fever (DF), chikungunya fever (CHIKF) and Zika disease (ZVD) is a cause for international concern. While DF remains the world’s most prevalent arboviral disease with tens of millions of cases annually and severe outbreaks in the Americas, YFV persists in endemic foci with high fatality among severe cases, CHIKF continues to cause regional outbreaks with occasional severe neurological sequelae, and ZVD circulates at lower levels globally but poses a persistent threat due to its teratogenic potential (1–5). Changes in climate and anthropogenic factors (e.g., land-use transformations and the large-scale movement of people, animals, and goods) are altering environmental conditions. These changes can indirectly affect the transmission and geographical distribution of MBD by facilitating the spread and redistribution of disease vectors across regions (6, 7).
Current approaches to managing MBD primarily focus on reducing populations of both immature and adult mosquitoes through insecticide application and community-based efforts to eliminate breeding habitats (8, 9). However, despite substantial resource investment, long-term reductions in mosquito densities remain challenging, with insecticide resistance being one of the main contributing factors (10), and seasonal outbreaks continue to occur (11, 12). This highlights a widely acknowledged need for innovative, cost-effective, and efficient tools to control arboviruses (13, 14). The limited success of conventional control methods has driven the exploration of innovative entomological strategies. The reproductive parasite Wolbachia pipientis Hertig, 1936 (Rickettsiales: Rickettsiaceae) (hereafter, Wolbachia), an obligatory intracellular and maternally inherited bacterium found in many arthropod species, sounds as a promising environmentally friendly weapon against MBD (15). Its ability to invade and maintain itself through manipulation of its host reproduction can be used to diminish mosquito population levels. Laying in the induced cytoplasmic incompatibility (CI) between uninfected females and infected males, both suppression and replacement with immune mosquito strategies have been proposed (16). While the concept of using the intracellular bacterium Wolbachia to manage mosquito populations was introduced over five decades ago, its potential role in dengue control has only garnered significant attention in the last ten years (17). Field studies in Australia and Indonesia have shown that releasing Wolbachia-transinfected Aedes aegypti (Linnaeus, 1762) mosquitoes can lead to a significant and lasting decrease in the DF transmission (18). Notably, a cluster randomized trial in Yogyakarta, Indonesia, demonstrated a 77% reduction in dengue cases in areas treated with Wolbachia (19). In Brazil, pilot releases in Rio de Janeiro and Niterói successfully established Wolbachia in local mosquito populations. Subsequently, Niterói expanded the intervention city-wide, using a phased strategy that included community engagement, information campaigns, mosquito releases, and field monitoring of Wolbachia prevalence (20, 21). Something similar occurred in the Colombian cities of Bello, Medellín, and Itagüí (22, 23). However, further field data are needed to evaluate Wolbachia’s effectiveness against these viruses and to assess its broader public health impact, as some investigations from Colombia are suggesting (24). Thus, it is important to study other Wolbachia strains and mosquito vectors to facilitate the introduction of this technology in other at-risk countries. The Asian tiger mosquito Aedes albopictus (Skuse, 1894) (Diptera: Culicidae) has expanded significantly over the past three decades from its native range in Southeast Asia to regions across North and South America, Southern Europe, parts of Africa, and various islands in Oceania, where it is now well established (25, 26). This mosquito is a competent vector for at least 22 arboviruses, including CHIKF and all four DF serotypes (27, 28). Although Ae. albopictus generally plays a secondary role in the transmission of DF and CHIKF compared to Ae. aegypti—partly due to its lower vector competence (29)—it has been implicated in outbreaks of these viruses in locations such as Hawaii, Mauritius, Gabon, Madagascar, and La Réunion (30–32). Moreover, autochthonous transmission of both viruses in parts of Europe by this species (33–36) highlights its growing public health relevance on a global scale. In Argentina, Ae. albopictus was first detected in 1998 (37). Its geographic distribution is restricted to the northeastern provinces of Misiones (where it was initially observed) and Corrientes (where it arrived due to a recent range expansion) (38). While it has been considered a secondary vector in Argentina (39), the wide variation in transmission efficiency observed across the Americas (40) suggests a substantial risk of CHIKF and DF becoming established and spreading throughout tropical, subtropical, and even temperate areas of the continent.
Aedes albopictus is naturally superinfected with two Wolbachia strains, identified as wAlbA and wAlbB (41), across nearly its entire global distribution (see Figure 1 in (42)). A notable exception occurs in Argentina, where individuals have been found carrying only the wAlbB strain or lacking Wolbachia infection altogether (39). The relatively low vector competence observed in Ae. albopictus populations from the Argentinean subtropics—when compared to Ae. aegypti—may be influenced by the presence of Wolbachia. This hypothesis is supported by studies on Ae. albopictus from La Réunion Island, where the endosymbiont was shown to reduce DENV-2 dissemination and salivary gland infection (43). However, given potential differences in Wolbachia strains, densities, and environmental factors between regions, further investigation is warranted to clarify its role in modulating vector competence under local conditions.
Figure 1. Map indicating the collection site for Ae. albopictus. (A) Regional map showing the relative position of Argentina within the continent. (B) Map of Argentina highlighting the sampling location in Puerto Iguazú. Maps were created using Adobe InDesign.
Both wAlbA and wAlbB strains have been associated with increased resistance to arboviral infections in mosquitoes (43), with wAlbB typically reaching higher densities within the host (44). The density of the symbiont plays a critical role in the host–symbiont interaction, influencing not only the efficiency of maternal transmission but also the potential virulence of the symbiont itself (45, 46). Moreover, symbiont density has been identified as a key factor modulating immune function and antiviral responses in mosquitoes, alongside the genetic makeup of both the host and the Wolbachia strain (47, 48). Several studies have reported a positive correlation between Wolbachia density and the strength of antiviral protection in the mosquito host (47, 49, 50). Although the precise mechanisms by which Wolbachia influences vector competence remain to be fully elucidated, current evidence points to bacterial density as a major factor (47), as well as the bacterium’s localization in somatic tissues—outside the gonads—where it coexists with the pathogen (51). Given that Wolbachia strains differ in both tissue tropism and replication levels within their hosts (52–54), and that these parameters can vary between strains in superinfected individuals (46, 55–57), our study aims to quantify the density of Wolbachia strains in Ae. albopictus populations from Argentina. As a preliminary step to explore the symbiont’s potential influence on arbovirus transmission, we assessed variation in Wolbachia density according to tissue type in females. Additionally, we examined whether symbiont density changes with host age, in order to evaluate if this factor may underlie the previously reported infection polymorphism in Argentinean populations.
2 Materials and methods
2.1 Sample collection
Immature stages of Ae. albopictus were collected from a natural population at a go-kart track in Puerto Iguazú, Argentina (25°39’20”S–54°33’12”W) in February–March of 2019 (Figure 1), and reared to adulthood in the laboratory of the Centro de Investigaciones Ecológicas Subtropicales (CIES), at Puerto Iguazú. First instar larvae were separated in 1 L of dechlorinated water in a plastic flat tray with finely ground guinea pig food until pupation. Larval density was not quantified due to high and continuous larval mortality, which caused density to vary during this stage. The pupae were removed and individualized to plastic containers and provided with water and raisins in preparation for emergence. Following emergence adults were sexed and maintained in plastic vials containing cotton and moist filter paper, and fed raisins. Larvae and adults were kept in an incubator with a temperature cycle fluctuating between 21 °C and 34 °C, 60% of humidity and a photoperiod of 14:10 (L:D), simulating field conditions. The F0 generation was used in two simultaneous experiments: (i) comparison of wAlbA and wAlbB strain densities between ovaries and somatic tissues of females, and (ii) comparison of wAlbB density among mosquitos of different ages using males as a model. Both experiments were made at the CIES.
2.2 DNA extraction
DNA extraction was performed on ten adult individuals of Ae. albopictus (per experiment) using Chelex® 100 resin (Bio-Rad, US). Each mosquito was placed in an Eppendorf tube containing 100 uL of 5% Chelex solution and 2 µL of Proteinase K (20 mg/mL; Promega, US). The mixture was incubated overnight at 56 °C for a period of 18 h. Proteinase K was then inactivated at 95 °C for 10 minutes. Subsequently, the sample was centrifuged at 14,000 rpm for 5 minutes, and the supernatant was transferred to a new tube and stored at −20 °C. DNA quality was assessed using a NanoDrop spectrophotometer (Thermo Fisher Scientific, USA).
2.3 Quantitative PCR assays
Wolbachia strain densities were investigated through quantitative PCR (hereafter, qPCR). Quantification of wAlbA and wAlbB strain densities was performed using the standard curve method. The wall surface protein gene (wsp) was used with strain-specific primers (Table 1). Quantification was normalized using the actin gene of Ae. albopictus as reference gene. These normalized values were used in comparative analyses (soma vs. ovary; and 0 days old males vs. 5days old males; 5days old males vs. 14 days old males). Target genes (wsp of wAlbA and wAlbB) and the reference gene (actin) were amplified from the same genomic DNA samples.

Table 1. List of primers used in qPCR assays, including primer ID, sequences (5′–3′), melting temperature (Tm), and GC content (%).
Previously published primers (58–60) were first tested. Due to non-specific amplification in negative controls, new primers were designed and tested for specificity. Primer design was performed in Gene Runner V3.05 (61) with selection criteria as follows: amplicon size of 150–200 bp, GC content of 30–80%, and absence of secondary structures, which was checked with DNAMAN (62). The selected primers are shown in Table 1. To confirm annealing temperature and expected amplicon size (189 bp for actine gene, 194 bp for wAlbA wsp gene, and 213 bp for wAlbB wsp gene), endpoint PCR was performed under the following conditions: 94 °C for 15 s, 60 °C for 25 s, and 72 °C for 15 s (40 cycles). DNA from Ae. albopictus was used as template; DNase-free water was included as a negative control. PCRs were run on a Labnet Multigene thermal cycler (Thermo Fisher Scientific, US). Reaction volumes were 12.5 µl: 6.25 µl GoTaq Master Mix (Promega, US), 10 µM each primer (Macrogen, South Korea), 40 ng DNA, and 4.25 µl DNAse-free water. Amplicons were visualized on 2% agarose gels stained with ethidium bromide 0.4 µg/ml (Promega, US) under UV light. Two infected mosquitoes per Wolbachia strain were tested, and for the actin gene, one mosquito.
All quantitative PCR assays were conducted on a StepOne Plus instrument (Applied Biosystems, USA), using 96-well plates and MicroAmp™ adhesive seals (Thermo Fisher Scientific, USA), in a final volume of 20 μl following the design and reporting guidelines of Bustin et al. (63). Each reaction used 10 μl of a MasterMix with SYBR® Green intercalating dye (Thermo Fisher Scientific, USA), 10 uM of oligonucleotides (Macrogen, South Korea), and 40 ng of total genomic DNA template.
Cycling conditions were 95 °C for 10 min, followed by 40 cycles of 15 sec at 95 °C and 1 min at 60 °C. Each plate included target and reference genes for each sample, with five biological replicates per comparison group, three technical replicates of each biological replicate and three technical replicates of negative controls per gene. Amplification quality was assessed by inspecting the amplification and melting temperature curves (see Supplementary Material). Cq values were averaged after verifying specificity via melting curves.
PCR efficiencies were calculated from standard curves (one per gene) generated using serial dilutions of a purified-PCR product (ranging from 10ng/µl and 0.001 ng/µl), with each dilution run in triplicate. Amplification efficiency (E) was calculated using the slope (m) of the linear regression line according to Applied Biosystems (2004): E = 10 (−1/m) – 1. Slopes ranging from –3.1 to –3.6 (corresponding to 90–110% efficiency) were considered acceptable. These efficiency values were used to validate the performance of the primer pairs, but not to calculate absolute quantities. Instead, relative quantification was performed using the comparative Cq method (ΔΔCt) (64). Normalized Wolbachia density in each sample was calculated as the ratio of wsp gene concentration (wAlbA or wAlbB) to actin gene amplification. These normalized values were used in statistical analyses. All analyses were conducted in R v4.1.0 (65) using integrated RStudio v1.0.153 environment (66).
2.4 Biological assays
2.4.1 qPCR assay for comparison of bacterial density between soma and ovary
Five newly emerged females (biological replicates) were dissected immediately after emergence and sex determination to separate somatic and ovarian tissues under a light microscope. Each female was placed in a drop (ca. 20–30 ul) of PBS 1X on a microscope slide. Using fine forceps, the terminal abdominal segment was gently pulled to expose and remove the ovaries. The ovaries were rinsed in distilled water to prevent cross-contamination with somatic tissues, and the forceps were sterilized before handling the soma. Somatic and ovarian tissues were placed in separate Eppendorf tubes, and DNA was extracted as explained in subsection 2.2. DNA dilutions were prepared at a final concentration of 10 ng/µl and used in qPCR assays. The relative densities of Wolbachia strains wAlbA and wAlbB were compared between ovary and soma according to subsection 2.3. Relative densities obtained from this assay were compared using the Mann–Whitney U test using the rstatix (67) and ggpubr libraries (68). Box plots were generated using the base R function boxplot.
2.4.2 qPCR assay to evaluate the variation in bacterial density of the wAlbB strain over male longevity
Because the wMel strain may show reduced density and CI when Ae. aegypti larvae are reared at high temperatures (69–71), but the wAlbB strain proved to be much less susceptible to the effects of similar high rearing temperatures (70, 71), the latter might be well suited for population replacement in hot environments, given its ability to effectively block transmission of DF and other arboviruses (72). Thus, our efforts were primarily devoted to quantification of this strain. Additionally, attempts made to quantify the wAlbA strain yielded inconclusive results due to non-specific amplification. Thus, we proceeded to investigate only the density of the wAlbB strain.
The density of the wAlbB strain was measured in five males (biological replicates) at three time points: immediately after emergence, at 5 days post-emergence, and 14 days post-emergence. As explained before, larvae collected from the field were individually isolated in transparent plastic containers at the pupal stage. Adults were monitored daily. Five males were euthanized at each time point by cold exposure. DNA was extracted as described in subsection 2.2 and dilutions were prepared at 5 ng/µl for the actin gene and 50 ng/µl for the wsp gene of the wAlbB strain. The relative density was quantified according to subsection 2.3.
Comparisons of wAlbB density were performed between time points 0 and 1, and between 1 and 2. The results were analyzed with the Kruskal–Wallis test using libraries tidyverse (73). Box plots were generated using the base R function boxplot.
3 Results
3.1 qPCR assay for comparison of bacterial density in soma and ovary
Primer efficiency (Ef) resulted in Ef_wsp_wAlbB = 109.12% (R2 = 0.9911), Ef_wsp_wAlbA = 103.61% (R2 = 0.9884), and Ef_Actin = 117.83% (R2 = 0.9866). Although the actin primer showed an efficiency slightly exceeding the recommended range for the comparative Cq (ΔΔCt) method, no correction was applied. Given that all reactions were performed under identical conditions and involved comparative analyses within primer sets, we consider the relative quantification results to be reliable. However, this deviation is acknowledged as a limitation of the method. Melting showed three distinct peaks corresponding to each one wsp_wAlbB (87.5° C), wsp_wAlbA (80.1° C), and actin (78.6° C), confirming specific amplification for each target (see Supplementary Material).
Both wAlbA (median relative density = 3.78) and wAlbB (median relative density = 3.31) strains were detected in all five ovarian samples, indicating superinfection (Figure 2). No significant difference in density between wAlbA and wAlbB was observed in ovaries (Mann–Whitney U test, p > 0.05). In somatic tissues, wAlbB was detected in four of the five individuals (median relative density = 0.41), whereas wAlbA was not detected in any (Figure 2). A significant difference in bacterial density was observed between wAlbA and wAlbB in somatic tissues (Mann–Whitney U test, p < 0.05).
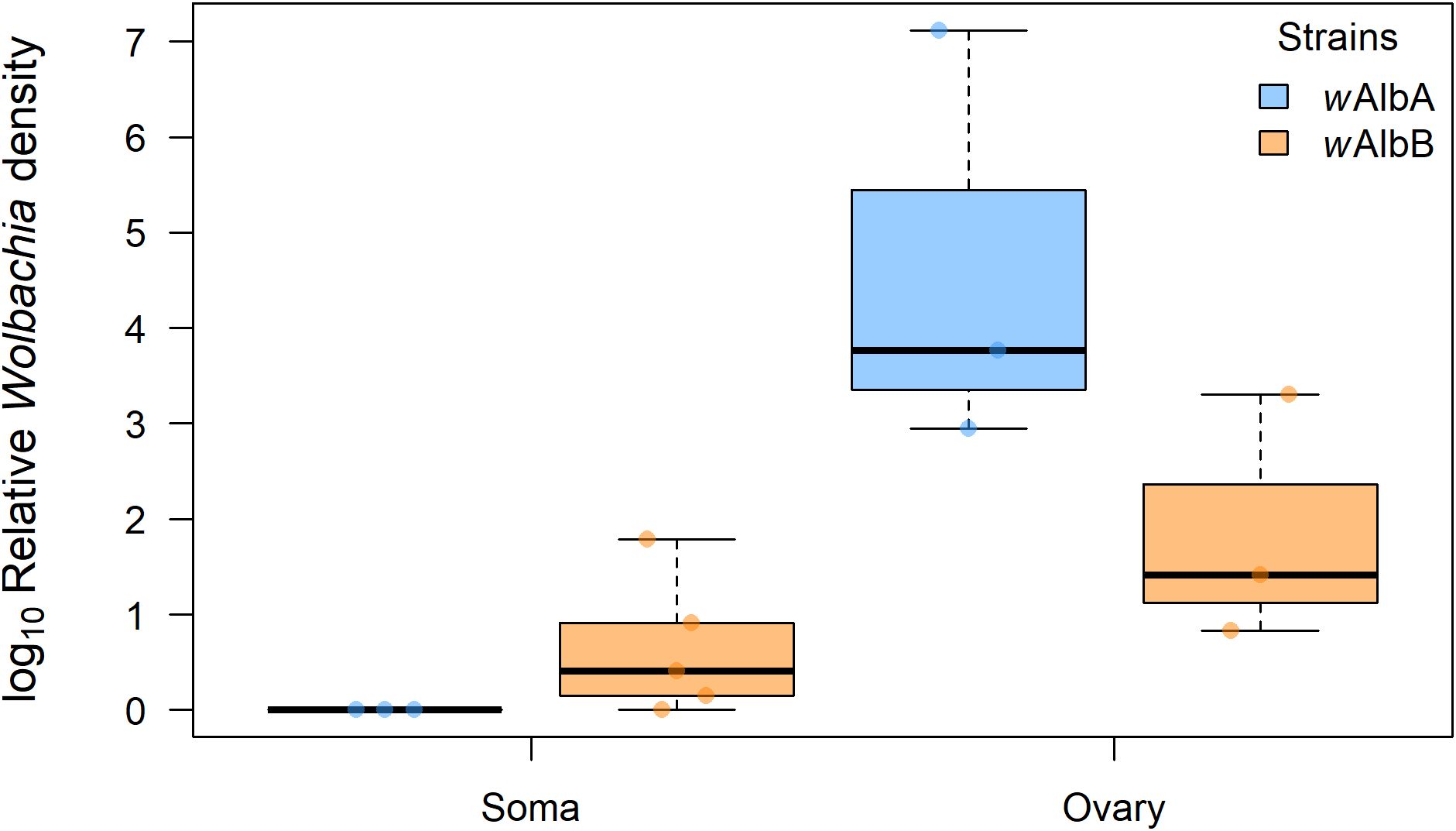
Figure 2. Relative densities of Wolbachia strains wAlbA (light blue) and wAlbB (orange) in ovaries and somatic tissues of Ae. albopictus females. Log10-transformed mean Wolbachia density is expressed as the ratio of the Wolbachia copy numbers of the gene wsp to the Ae. albopictus gene Actin, as estimated by qPCR on genomic DNA. A total of five biological replicates were used. Thick horizontal lines represent medians, box limits indicate first and third quartiles, whiskers represent interquartile range. Individual data points (jittered) are shown to illustrate the distribution of values.
3.2 qPCR assay to evaluate changes in wAlbB density over male lifespan
For the assay evaluating the dynamics of wAlbB density over male lifespan, primer efficiencies were Ef_wspAlbB = 98.76% (R2 = 0.9920) and Ef_Actin = 111.55% (R2 = 0.9960). As these values were within or near the acceptable range for the comparative Cq (ΔΔCt) method, no correction was applied. However, we acknowledge that the actin primer exceeded the ideal efficiency range, which may have introduced some slight bias in the estimation. Melting temperatures were 79.03 °C for wsp (wAlbB) and 87.63 °C for actin. Melting curves showed specific amplification (see Supplementary Material).
A total of five males were analyzed for each time point. One sample from time = 14 days post-emergence was excluded due to ≥ 35 Cq value, in accordance with our pre-established threshold. No significant differences in wAlbB density were found between time = 0 days post-emergence (median relative density = 0.83) and 5 days post-emergence (median relative density = 1.98), nor between 5 and 14 (median relative density = 0.66) (Kruskal–Wallis test, p = 0.3362) (Figure 3).
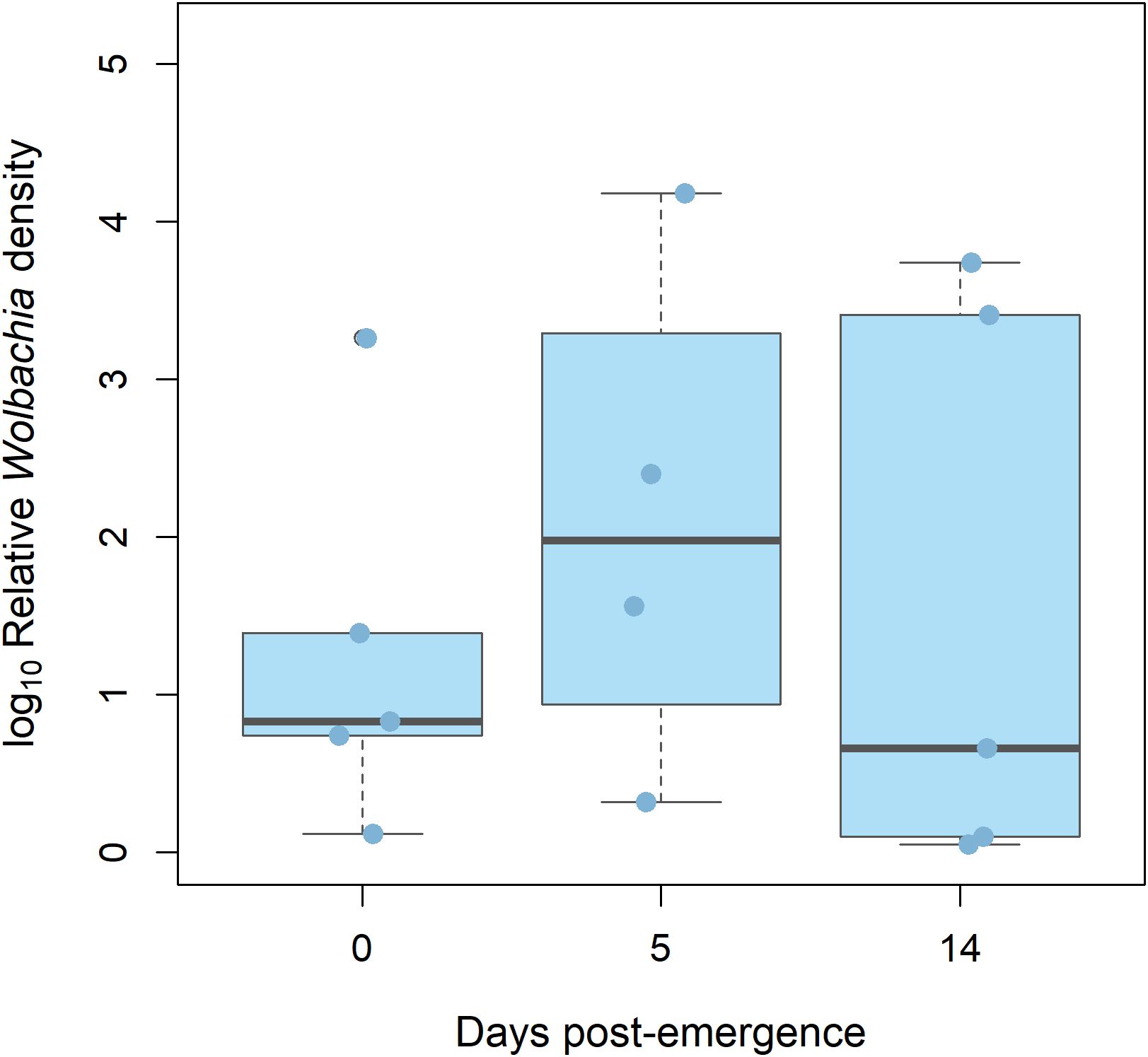
Figure 3. Relative densities of Wolbachia strain wAlbB (light blue) in 0, 5, and 14 days-old Ae. Albopictus males. Log10-transformed mean Wolbachia density is expressed as the ratio of the Wolbachia copy numbers of the gene wsp to the Ae. albopictus gene Actin, as estimated by qPCR on genomic DNA. A total of five biological replicates were used. Thick horizontal lines represent medians, box limits indicate first and third quartiles, whiskers represent interquartile range. Individual data points (jittered) are shown to illustrate the distribution of values.
Amplification of wAlbA was excluded due to poor melting curves, non-specific amplification, and high Cq values. One actin negative control showed a left-shifted curve, likely due to primer dimers, while the other showed no amplification. No such issues were observed in test samples.
4 Discussion
Bacterial density plays a critical role in all the phenotypes induced by Wolbachia in their hosts (74). Recent studies have reinforced earlier observations suggesting that, beyond its localization in the germline, somatic localization is a conserved feature of Wolbachia infection, indicating that it is not incidental but rather a key aspect of Wolbachia biology (51).
In this study, we examined the density of both wAlbA and wAlbB strains in somatic and ovarian tissues of Ae. albopictus. Both tissues were infected, with somatic tissues harboring only wAlbB, while ovaries displayed a double infection. The negative result in both tissues of a single sample may indicate that the infection is not fixed in this host population or could be due to a technical limitation. Increasing the sample size may help clarify this issue. Nevertheless, our finding is consistent with Dobson et al. (52), who reported that Ae. albopictus individuals infected solely with wAlbA lacked Wolbachia in somatic tissues, suggesting that wAlbA is restricted to reproductive tissues. Conversely, Zouache et al. (75) found both strains (wAlbA and wAlbB) in somatic tissues (salivary glands and gut) and ovaries of Ae. albopictus from Réunion Island. They also reported higher Wolbachia densities in ovaries than in somatic tissues, but did not observe significant differences between the two strains in either tissue type—a result that aligns with our findings in ovarian tissues.
Several studies have shown that the two strains differ significantly in their within-host densities, with wAlbB often reaching higher levels (44). This disparity may reflect differences in replication rates, with the Wolbachia strains from supergroup A (e.g., wAlbA) generally showing slower proliferation (56, 76). Moreover, each Wolbachia strain replicates independently, meaning that the growth rate of one strain is unaffected by the presence of the other. One possible explanation is that supergroup B strains may penetrate host cells more efficiently and replicate more rapidly in reproductive tissues than supergroup A strains (77). Possible cellular and molecular mechanisms that mediate differential tissue localization include variation in the expression of host receptors or cellular factors required for bacterial entry and replication, differential activation of immune pathways, or competition between strains for colonization niches (51, 75, 78, 79). Similar patterns of strain-specific tissue localization have been reported in other Aedes species, supporting the hypothesis that these differences are biologically driven rather than artifacts of our sampling approach (80).
The presence of Wolbachia in specific somatic tissues suggests that somatic tropism is a trait under selection, not a random byproduct of infection. Somatic localization may be maintained evolutionarily because it facilitates horizontal transmission within and between species, contributing to genetic diversity (51). Additionally, it may confer advantageous phenotypes on the host that promote vertical transmission through the germline (51), potentially increasing host fecundity or improving the vertical transmission efficiency of Wolbachia (52). From the host perspective, somatic infection may contribute to antiviral protection by interfering with viral replication in tissues critical for vector competence, thereby potentially increasing host survival and reproductive success (81). These interactions suggest that tissue-specific localization could be shaped by mutual evolutionary benefits. Given the relevance of somatic localization for biocontrol applications, it is important to rule out the possibility that the target Wolbachia genes are actually nuclear insertions of bacterial DNA fragments, as these would fail to generate the desired effects (82, 83).
We also investigated the dynamics of wAlbB density over the adult lifespan of male mosquitoes. Our results showed no significant variation in wAlbB density throughout adult male aging. This finding contrasts with that of Tortosa et al. (84), who observed a positive correlation between wAlbB density and age in males from Corsica and Réunion Island, but a negative correlation in males from Greece. They concluded that the association between wAlbB density and age may vary depending on the population of origin. In our study, however, these two variables appeared to be independent. Since both experimental designs were fairly similar, it is likely that the divergent results stem from intrinsic population factors. These may include differences in the genetic background of the host (such as variation in nuclear–Wolbachia interactions), local environmental adaptation, and historical selective pressures acting on both host and symbiont. For instance, Mejia et al. (85) showed that relative Wolbachia densities can be predictable across tissues and generations, but still vary depending on population origin. Furthermore, host genetic background has been shown to influence Wolbachia-mediated phenotypes: for example, wMel introgressed into different Ae. aegypti genetic backgrounds in Brazil and Vietnam produced differences in both mean and variance of dengue virus susceptibility (86). These studies support the idea that intrinsic variation among mosquito populations can significantly modulate Wolbachia dynamics and associated phenotypes.
Although this study did not quantify Wolbachia density in females, it is well established that bacterial density plays a crucial role in infection stability and the manifestation of Wolbachia-induced phenotypes in the host (74). In females, Wolbachia density may affect vector competence, while in males it could influence the strength of CI. This may help explain the low hatching rate observed in the Argentine Ae. albopictus population (see 39). Low bacterial densities could impair vertical transmission, leading to uninfected individuals, or in the case of co-infection, result in the stochastic loss of one of the strains. Such dynamics could underlie the infection polymorphism for Wolbachia infection observed in this host population (39). On the other hand, understanding strain density in females is particularly important, as they are the vectors. It is essential to assess density across the lifespan of adult females to determine whether antiviral activity remains constant and can be sustained throughout their life. This is especially relevant for a sustainable Wolbachia-based control strategy, in which the symbiont must be maintained across generations and vertical transmission reliably ensured.
We acknowledge that the relatively small sample size in our study is a limitation. Small sample sizes can reduce statistical power, making it more difficult to detect subtle differences and potentially increasing the likelihood of Type II errors. In addition, some data points were excluded due to technical issues in qPCR (e.g., failed amplification or outlier Cq values, since we discarded results with Cq ≥ 35), which further reduced the number of observations. We were also unable to include amplification of positive controls or sequencing of PCR products, which, to a certain extent, might limit the certainty of target specificity in our qPCR assays, although we partially verified the specificity of the reaction using melting curves. Despite these limitations, the observed trends were consistent across the analyzed samples, and the results provide valuable preliminary insights into tissue-specific Wolbachia densities.
In summary, the density of Wolbachia plays a critical role in shaping host–symbiont evolutionary interactions and enhancing the effectiveness of this bacterium as a biological control agent against insect pests and vector-borne diseases (16). Based on the results of this study, wAlbB emerges as a strong candidate for transinfection of native Ae. aegypti populations because of its somatic localization, which may enhance antiviral protection. If the high wAlbB density is consistently maintained throughout the female lifespan, we can expect both robust antiviral protection and reliable vertical transmission, thereby ensuring the sustainability of replacement biocontrol techniques.
Further experiments involving females—the sex that acts as arbovirus vectors—should assess Wolbachia strain density across the entire lifespan, which is relevant for antiviral activity; examine the dependence of bacterial density on temperature, as high temperatures can impair Wolbachia performance; and evaluate the influence of blood feeding on Wolbachia density, since arboviruses are acquired through blood meals, and it would be interesting to determine whether blood feeding promotes bacterial replication. Additionally, the main practical value of this work lies in the methodological information it provides for studying this strain, since we had to design several primer pairs and optimize real-time PCR conditions for multiple assays.
Given that the densities of wAlbA and wAlbB have been shown to differ across populations (87), it would be valuable to expand surveys to other locations of Argentina and explore how these variations correlate with superinfection patterns. Considering that wAlbB has demonstrated a strong potential to reduce dengue incidence in high-transmission areas (88, 89), further insights into this strain are of significant interest and importance. In order to validate the use of this strain in field settings in Argentina, the next steps would involve rearing native Ae. aegypti, as populations in Argentina are peculiar and replacement by foreign populations may be hindered by local adaptation and competition (90, 91), transinfecting them with the native wAlbB strain (the focus of our study), and conducting pre-release surveys.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
AC: Conceptualization, Validation, Writing – review & editing, Investigation, Visualization, Formal Analysis, Software. MR: Conceptualization, Writing – review & editing, Writing – original draft, Methodology, Supervision. MM: Resources, Funding acquisition, Project administration, Conceptualization, Supervision, Methodology, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This contribution was supported by grants from Agencia Nacional de Promoción Científica y Tecnológica (PICT 2015-0665) and Consejo Nacional de Investigaciones Científicas y Técnicas (PIP 584) to MM.
Acknowledgments
Thanks are due to the Centro de Investigaciones Ecológicas Subtropicales (CIES) for logistical support in Puerto Iguazú, to Martín Montes, Paula Marcotegui, Jorge Barneche, and Gastón Cavallo for their invaluable help during field trips, to Darío Balcazar, Pablo Giringhelli, and Alejandra Scannapieco for their help with molecular techniques, to Graciela Minardi and Lucía Babino for their valuable statistical advice, to Laura Morote for designing Figure 1, and the reviewers who provided invaluable feedback on the manuscript. AC was awarded doctoral scholarships from the Agencia Nacional de Promoción Científica y Tecnológica and Consejo Nacional de Investigaciones Científicas y Técnicas. MSR and MVM are members of the Research Career of the Consejo Nacional de Investigaciones Científicas y Técnicas, Argentina.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling editor ACdlF declared a shared parent affiliation with the author MR at the time of review.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/finsc.2025.1655459/full#supplementary-material
References
1. Haider N, Hasan MN, Onyango J, Billah M, Khan S, Papakonstantinou D, et al. Global Dengue epidemic worsens with record 14 Million cases and 9,000 deaths reported in 2024. Int J Infect Dis. (2025) 158:107940. doi: 10.1016/j.ijid.2025.107940
2. Mehta R, Gerardin P, de Brito CAA, Soares CN, Ferreira MLB, and Solomon T. The neurological complications of chikungunya virus: A systematic review. Rev Med Virol. (2018) 28:e1978. doi: 10.1002/rmv.1978
3. de Lima Cavalcanti TYV, Pereira MR, de Paula SO, and Franca RFDO. A review on chikungunya virus epidemiology, pathogenesis and current vaccine development. Viruses. (2022) 14:969. doi: 10.3390/v14050969
4. Servadio JL, Muñoz-Zanzi C, and Convertino M. Estimating case fatality risk of severe Yellow Fever cases: systematic literature review and meta-analysis. BMC Infect Dis. (2021) 21:819. doi: 10.1186/s12879-021-06535-4
5. Gilbert RK, Petersen LR, Honein MA, Moore CA, and Rasmussen SA. Zika virus as a cause of birth defects: Were the teratogenic effects of Zika virus missed for decades? Birth Defects Res. (2023) 115:265–74. doi: 10.1002/bdr2.2134
6. Bartlow AW, Manore C, Xu C, Kaufeld KA, Del Valle S, Ziemann A, et al. Forecasting zoonotic infectious disease response to climate change: mosquito vectors and a changing environment. Vet Sci. (2019) 6:40. doi: 10.3390/vetsci6020040
7. de Souza WM and Weaver SC. Effects of climate change and human activities on vector-borne diseases. Nat Rev Microbiol. (2024) 22:476–91. doi: 10.1038/s41579-024-01026-0
8. Shroff S, Mir S, Naik B, Baitharu I, and Behera AK. Chemical methods for control of mosquito vector. In: Barik TK, editor. Molecular Identification of Mosquito Vectors and Their Management. Springer Singapore, Singapore (2021). p. 35–50.
9. Yulfi H, Panggabean M, Darlan DM, Siregar IS, and Rozi MF. Community-based intervention in mosquito control strategy: A systematic review. Narra J. (2025) 5:e1015. doi: 10.52225/narra.v5i1.1015
10. Bharati M and Saha D. Insecticide resistance status and biochemical mechanisms involved in Aedes mosquitoes: A scoping review. Asian Pac J Trop Med. (2021) 14:52–63. doi: 10.4103/1995-7645.306737
11. Medeiros EA. Challenges in controlling the dengue epidemic in Brazil. Acta Paul Enferm. (2024) 37:eEDT012. doi: 10.37689/acta-ape/2024EDT012
12. Ferguson NM. Challenges and opportunities in controlling mosquito-borne infections. Nature. (2018) 559:490–7. doi: 10.1038/s41586-018-0318-5
13. Olliaro P, Fouque F, Kroeger A, Bowman L, Velayudhan R, Santelli AC, et al. Improved tools and strategies for the prevention and control of arboviral diseases: A research-to-policy forum. PloS Negl Trop Dis. (2018) 12:e0005967. doi: 10.1371/journal.pntd.0005967
14. Thompson R, Martin Del Campo J, and Constenla D. A review of the economic evidence of Aedes-borne arboviruses and Aedes-borne arboviral disease prevention and control strategies. Expert Rev Vaccines. (2020) 19:143–62. doi: 10.1080/14760584.2020.1733419
15. Iturbe-Ormaetxe I, Walker T, and O'Neill SL. Wolbachia and the biological control of mosquito-borne disease. EMBO Rep. (2011) 12:508–18. doi: 10.1038/embor.2011.84
16. Hoffmann AA, Ross PA, and Rašić G. Wolbachia strains for disease control: ecological and evolutionary considerations. Evol Appl. (2015) 8:751–68. doi: 10.1111/eva.12286
17. Caragata EP, Dutra HL, Sucupira PH, Ferreira AG, and Moreira LA. Wolbachia as translational science: controlling mosquito-borne pathogens. Trends Parasitol. (2021) 37:1050–67. doi: 10.1016/j.pt.2021.06.007
18. Indriani C, Tantowijoyo W, Rancès E, Andari B, Prabowo E, Yusdi D, et al. Reduced dengue incidence following deployments of Wolbachia- infected Aedes aEgypti in Yogyakarta, Indonesia: A quasi-experimental trial using controlled interrupted time series analysis. Gates Open Res. (2020) 4:50. doi: 10.12688/gatesopenres.13122.1
19. Utarini A, Indriani C, Ahmad RA, Tantowijoyo W, Arguni E, Ansari MR, et al. Efficacy of Wolbachia-infected mosquito deployments for the control of dengue. New Engl J Med. (2021) 384:2177–86. doi: 10.1056/NEJMoa2030243
20. Durovni B, Saraceni V, Eppinghaus A, Riback TI, Moreira LA, Jewell NP, et al. The impact of large-scale deployment of Wolbachia mosquitoes on dengue and other Aedes-borne diseases in Rio de Janeiro and Niterói, Brazil: study protocol for a controlled interrupted time series analysis using routine disease surveillance data. F1000Res. (2020) 8:1328. doi: 10.12688/f1000research.19859.2
21. Pinto SB, Riback TI, Sylvestre G, Costa G, Peixoto J, Dias FB, et al. Effectiveness of Wolbachia-infected mosquito deployments in reducing the incidence of dengue and other Aedes-borne diseases in Niterói, Brazil: A quasi-experimental study. PloS Negl Trop D. (2021) 15:e0009556. doi: 10.1371/journal.pntd.0009556
22. Velez ID, Tanamas SK, Arbelaez MP, Kutcher SC, Duque SL, Uribe A, et al. Reduced dengue incidence following city-wide wMel Wolbachia mosquito releases throughout three Colombian cities: Interrupted time series analysis and a prospective case-control study. PloS Negl Trop D. (2023) 17:e0011713. doi: 10.1371/journal.pntd.0011713
23. Velez ID, Uribe A, Barajas J, Uribe S, Angel S, Suaza-Vasco JD, et al. Large-scale releases and establishment of wMel Wolbachia in Aedes aEgypti mosquitoes throughout the Cities of Bello, Medellín and Itagüí, Colombia. PloS Negl Trop D. (2023) 17:e0011642. doi: 10.1371/journal.pntd.0011642
24. Calle-Tobón A, Rojo-Ospina R, Zuluaga S, Giraldo-Muñoz JF, and Cadavid JM. Evaluation of Wolbachia infection in Aedes aEgypti suggests low prevalence and highly heterogeneous distribution in Medellin, Colombia. Acta Trop. (2024) 260:107423. doi: 10.1016/j.actatropica.2024.107423
25. Battaglia V, Agostini V, Moroni E, Colombo G, Lombardo G, Rambaldi Migliore N, et al. The worldwide spread of Aedes albopictus: New insights from mitogenomes. Front Genet. (2022) 13:931163. doi: 10.3389/fgene.2022.931163
26. Kraemer MU, Sinka ME, Duda KA, Mylne AQ, Shearer FM, Barker CM, et al. The global distribution of the arbovirus vectors Aedes aEgypti and Ae. albopictus elife. (2015) 4:e08347. doi: 10.7554/eLife.08347
27. Gratz NG. Critical review of the vector status of. Aedes albopictus Med Vet Entomol. (2004) 18:215–27. doi: 10.1111/j.0269-283X.2004.00513.x
28. Paupy C, Delatte H, Bagny L, Corbel V, and Fontenille D. Aedes albopictus, an arbovirus vector: from the darkness to the light. Microbes Infect. (2009) 11:1177–85. doi: 10.1016/j.micinf.2009.05.005
29. Lambrechts L, Scott TW, and Gubler DJ. Consequences of the expanding global distribution of Aedes albopictus for dengue virus transmission. PloS Negl Trop Dis. (2010) 4:e646. doi: 10.1371/journal.pntd.0000646
30. Rezza G. Aedes albopictus and the reemergence of Dengue. BMC Public Health. (2012) 12:72. doi: 10.1186/1471-2458-12-72
31. Rodriguez-Morales AJ, Montenegro-Idrogo JJ, Celis-Salinas JC, Angerami R, Villamil-Gómez WE, Sarute N, et al. Unraveling the unparalleled 2024 epidemic of Dengue in the Americas. Rev Chil Infectol. (2024) 41:421–8. doi: 10.4067/s0716-10182024000300133
32. López MS, Lovino MA, Gómez AA, Rodríguez ST, Radosevich AL, Müller GV, et al. Climate extremes, average conditions and temperature variability as drivers of dengue epidemics in a temperate city of Argentina. J Clim Change Health. (2025) 22:100426. doi: 10.1016/j.joclim.2025.100426
33. Liu Q, Shen H, Gu L, Yuan H, and Zhu W. Chikungunya virus in Europe: A retrospective epidemiology study from 2007 to 2023. PloS Negl Trop D. (2025) 19:e0012904. doi: 10.1371/journal.pntd.0012904
34. Brem J, Elankeswaran B, Erne D, Hedrich N, Lovey T, Marzetta V, et al. Dengue “homegrown” in Europe (2022 to 2023). New Microbes New Infect. (2023) 56:101205. doi: 10.1016/j.nmni.2023.101205
35. Medić A, Savić V, Klobučar A, Bogdanić M, Curman Posavec M, Nonković D, et al. Epidemiological and entomological study after the possible re-emergence of dengue fever in Croatia, 2024. Microorganisms. (2025) . 13:565. doi: 10.3390/microorganisms13030565
36. Fleischmann WA, Nurjadi D, and Velavan TP. Addressing the rise of autochthonous vector-borne diseases in a warming Europe. Int J Infect Dis. (2024) 149:107275. doi: 10.1016/j.ijid.2024.107275
37. Rossi GC, Pascual NT, and Krsticevic FJ. First record of Aedes albopictus (Skuse) from Argentina. J Am Mosq Control Assoc-Mosq News. (1999) 15:422–2.
38. Goenaga S, Chuchuy A, Micieli MV, Natalini B, Kuruc J, and Kowalewski M. Expansion of the distribution of Aedes albopictus (Diptera: Culicidae): new records in northern Argentina and their implications from an epidemiological perspective. J Med Entomol. (2020) 57:1310–3. doi: 10.1093/jme/tjaa009
39. Chuchuy A, Rodriguero MS, Ferrari W, Ciota AT, Kramer LD, and Micieli MV. Biological characterization of Aedes albopictus (Diptera: Culicidae) in Argentina: implications for arbovirus transmission. Sci Rep-UK. (2018) . 8:5041. doi: 10.1038/s41598-018-23401-7
40. Vega-Rúa A, Zouache K, Girod R, Failloux A-B, and Lourenço-de-Oliveira R. High level of vector competence of Aedes aEgypti and Aedes albopictus from ten American countries as a crucial factor in the spread of Chikungunya Virus. J Virol. (2014) 88:6294–306. doi: 10.1128/JVI.00370-14
41. Sinkins SP, Braig HR, and O’Neill SL. Wolbachia superinfections and the expression of cytoplasmic incompatibility. P Roy Soc Lond B Bio. (1995) 261:325–30. doi: 10.1098/rspb.1995.0154
42. Bourtzis K, Dobson SL, Xi Z, Rasgon JL, Calvitti M, Moreira LA, et al. Harnessing mosquito-Wolbachia symbiosis for vector and disease control. Acta Trop. (2014) 132:S150–63. doi: 10.1016/j.actatropica.2013.11.004
43. Mousson L, Zouache K, Arias-Goeta C, Raquin V, Mavingui P, and Failloux AB. The native Wolbachia symbionts limit transmission of Dengue Virus in Aedes albopictus. PloS Negl Trop D. (2012) 6:e1989. doi: 10.1371/journal.pntd.0001989
44. Wiwatanaratanabutr I and Kittayapong P. Effects of crowding and temperature on Wolbachia infection density among life cycle stages of Aedes albopictus. J Invertebr Pathol. (2009) 102:220–4. doi: 10.1016/j.jip.2009.08.009
45. Mouton L, Henri H, Bouletreau M, and Vavre F. Strain-specific regulation of intracellular Wolbachia density in multiply infected insects. Mol Ecol. (2003) 12:3459–65. doi: 10.1046/j.1365-294X.2003.02015.x
46. Mouton L, Henri H, Charif D, Bouletreau M, and Vavre F. Interaction between host genotype and environmental conditions affects bacterial density in Wolbachia symbiosis. Biol Letters. (2007) 3:210–3. doi: 10.1098/rsbl.2006.0590
47. Lu P, Bian G, Pan X, and Xi Z. Wolbachia induces density-dependent inhibition to Dengue Virus in mosquito cells. PloS Negl Trop D. (2012) 6:1–8. doi: 10.1371/journal.pntd.0001754
48. Sinkins SP. Wolbachia and cytoplasmic incompatibility in mosquitoes. Insect Biochem Mol Biol. (2004) 34:723–9. doi: 10.1016/j.ibmb.2004.03.025
49. Osborne SE, Iturbe-Ormaetxe I, Brownlie JC, O’Neill SL, and Johnson KN. Antiviral protection and the importance of Wolbachia density and tissue tropism in Drosophila simulans. Appl Environ Microb. (2012) 78:6922–9. doi: 10.1128/AEM.01727-12
50. Osborne SE, Leong YS, O’Neill SL, and Johnson KN. Variation in antiviral protection mediated by different Wolbachia strains in Drosophila simulans. PloS Pathog. (2009) 5:e1000656. doi: 10.1371/journal.ppat.1000656
51. Pietri JE, DeBruhl H, and Sullivan W. The rich somatic life of. Wolbachia MicrobiologyOpen. (2016) 5:923–36. doi: 10.1002/mbo3.390
52. Dobson SL, Bourtzis K, Braig HR, Jones BF, Zhou W, Rousset F, et al. Wolbachia infections are distributed throughout insect somatic and germ line tissues. Insect Biochem Mol Biol. (1999) 29:153–60. doi: 10.1016/S0965-1748(98)00119-2
53. Dutton TJ and Sinkins SP. Strain-specific quantification of Wolbachia density in Aedes albopictus and effects of larval rearing conditions. Insect Mol Biol. (2004) 13:317–22. doi: 10.1111/j.0962-1075.2004.00490.x
54. Miller WJ and Riegler M. Evolutionary dynamics of wAu-like Wolbachia variants in neotropical Drosophila spp. Appl Environ Microb. (2006) 72:826–35. doi: 10.1128/AEM.72.1.826-835.2006
55. Ijichi N, Kondo N, Matsumoto R, Shimada M, Ishikawa H, and Fukatsu T. Internal spatiotemporal population dynamics of infection with three wolbachia strains in the adzuki bean beetle, callosobruchus chinensis (Coleoptera: bruchidae). Appl Environ Microb. (2002) 68:4074–80. doi: 10.1128/AEM.68.8.4074
56. Ikeda T, Ishikawa H, and Sasaki T. Regulation of Wolbachia density in the Mediterranean flour moth, Ephestia kuehniella, and the Almond Moth, Cadra cautella. Zool Sci. (2003) 20:153–7. doi: 10.2108/zsj.20.153
57. Mouton L, Dedeine F, Henri H, Boulétreau M, Profizi N, and Vavre F. Virulence, multiple infections and regulation of symbiotic population in the Wolbachia-Asobara tabida symbiosis. Genetics. (2004) 168:181–9. doi: 10.1534/genetics.104.026716
58. Ahantarig A, Trinachartvanit W, and Kittayapong P. Relative Wolbachia density of field-collected Aedes albopictus mosquitoes in Thailand. J Vector Ecology. (2008) 33:173–7. doi: 10.3376/1081-1710(2008)33[173:RWDOFA]2.0.CO;2
59. Raquin V, Moro CV, Saucereau Y, Tran FH, Potier P, and Mavingui P. Native Wolbachia from Aedes albopictus nlocks Chikungunya Virus infection in cellulo. PloS One. (2015) 10:1–19. doi: 10.1371/journal.pone.0125066
60. Zhou W, Rousset F, and O’Neill S. Phylogeny and PCR-based classification of Wolbachia strains using wsp gene sequences. Proc R Soc London Ser B: Biol Sci. (1998) 265:509–15. doi: 10.1098/rspb.1998.0324
61. Spruyt M and Buquicchio F. Gene runner version 3.05 (1994). Available online at: http://www.generunner.net/ (Accessed April 10, 2021).
62. Woffelman C. DNAMAN for Windows, version 2.6. Lynon Biosoft: Institute of Molecular Plant Science, Leiden University, the Netherlands (1994).
63. Bustin SA, Benes V, Garson JA, Hellemans J, Huggett J, Kubista M, et al. The MIQE Guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin Chem. (2009) 55:611 622. doi: 10.1373/clinchem.2008.112797
64. Pfaffl MW. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. (2001) 29:e45–5. doi: 10.1093/nar/29.9.e45
65. R Core Team. R: a language and environment for statistical computing. Vienna, Austría: R Foundation for Statistical Computing (2019). Available online at: https://www.R-project.org/ (Accesed July 5, 2023).
66. RStudio Team. RStudio: Integrated Development Environment for R. PBC, Boston, MA: RStudio. (2020). Available online at: http://www.rstudio.com/ (Accesed July 5, 2023).
67. Kassambara A. rstatix: Pipe-Friendly Framework for Basic Statistical Tests. R package version 0.7.2 (2023). Available online at: https://CRAN.R-project.org/package=rstatix (Accesed July 5, 2023).
68. Kassambara A. ggpubr: 'ggplot2' Based Publication Ready Plots. R package version 0.6.0 (2023). Available online at: https://CRAN.R-project.org/package=ggpubr (Accesed July 5, 2023).
69. Ant TH, Herd CS, Geoghegan V, Hoffman AA, and Sinkins SP. The Wolbachia strain wAu provides highly efficient virus transmission blocking in Aedes aEgypti. PloS Pathog. (2018) 14:e1006815. doi: 10.1371/journal.ppat.1006815
70. Ross PA, Wiwatanaratanabutr I, Axford JK, White VL, Endersby-Harshman NM, and Hoffman AA. Wolbachia infections in Aedes aEgypti differ markedly in their response to cyclical heat stress. PloS Pathog. (2017) 13:e1006006. doi: 10.1371/journal.ppat.1006006
71. Ulrich JN, Beier JC, Devine GJ, and Hugo LE. Heat sensitivity of wMel Wolbachia during Aedes aEgypti development. PloS Negl Trop Dis. (2016) 10:e0004873. doi: 10.1371/journal.pntd.0004873
72. Bian G, Xu Y, Lu P, and Xi Z. The endosymbiotic bacterium Wolbachia induces resistance to dengue virus in Aedes aEgypti. PloS Pathog. (2010) 6:e1000833. doi: 10.1371/journal.ppat.1000833
73. Wickham H. tidyverse: Easily Install and Load the 'Tidyverse'. R package version 2.0.0 (2023). Available online at: https://CRAN.R-project.org/package=tidyverse (Accesed July 5, 2023).
74. López-Madrigal S and Duarte EH. Titer regulation in arthropod-Wolbachia symbioses. FEMS Microbiol Lett. (2019) 366:1–19. doi: 10.1093/femsle/fnz232
75. Zouache K, Voronin D, Tran-Van V, Mousson L, Failloux AB, and Mavingui P. Persistent Wolbachia and cultivable bacteria infection in the reproductive and somatic tissues of the mosquito vector Aedes albopictus. PloS One. (2009) 4:e6388. doi: 10.1371/journal.pone.0006388
76. Wiwatanaratanabutr S and Kittayapong P. Effects of temephos and temperature on Wolbachia load and life history traits of Aedes albopictus. Med Vet Entomol. (2006) 20:300–7. doi: 10.1111/j.1365-2915.2006.00640.x
77. Kittayapong P, Baisley KJ, Sharpe RG, Baimai V, and O’Neill SL. Maternal transmission efficiency of Wolbachia superinfections in Aedes albopictus populations in Thailand. Am J Trop Med Hyg. (2002) 66:103–7. doi: 10.4269/ajtmh.2002.66.103
78. Minwuyelet A, Petronio GP, Yewhalaw D, Sciarretta A, Magnifico I, Nicolosi D, et al. Symbiotic Wolbachia in mosquitoes and its role in reducing the transmission of mosquito-borne diseases: updates and prospects. Front Microbiol. (2023) 14:1267832. doi: 10.3389/fmicb.2023.1267832
79. Porter J and Sullivan W. The cellular lives of Wolbachia. Nat Rev Microbiol. (2023) 21:750–66. doi: 10.1038/s41579-023-00918-x
80. Skelton E, Rancès E, Frentiu FD, Kusmintarsih ES, Iturbe-Ormaetxe I, Caragata EP, et al. A native Wolbachia endosymbiont does not limit dengue virus infection in the mosquito Aedes notoscriptus (Diptera: Culicidae). J Med Entomol. (2016) 53:401–8. doi: 10.1093/jme/tjv235
81. Barbar T. Exploring extrinsic and biological factors shaping Aedes albopictus vector competence: Assessing the role of Wolbachia in intestinal homeostasis maintenance. Saint-Denis, Reunion: Université de la Réunion (2024).
82. Klasson L, Kambris Z, Cook PE, Walker T, and Sinkins SP. Horizontal gene transfer between Wolbachia and the mosquito Aedes aEgypti. BMC Genomics. (2009) 10:1–9. doi: 10.1186/1471-2164-10-33
83. Woolfit M, Iturbe-Ormaetxe I, McGraw EA, and O’Neill SL. An ancient horizontal gene transfer between mosquito and the endosymbiotic bacterium. Wolbachia pipientis Mol Biol Evol. (2009) 26:367–74. doi: 10.1093/molbev/msn253
84. Tortosa P, Charlat S, Labbé P, Dehecq JS, Barré H, and Weill M. Wolbachia age-sex-specific density in Aedes albopictus: a host evolutionary response to cytoplasmic incompatibility? PloS One. (2010) 5:e9700. doi: 10.1371/journal.pone.0009700
85. Mejia AJ, Dutra HLC, Jones MJ, Perera R, and McGraw EA. Cross-tissue and generation predictability of relative Wolbachia densities in the mosquito Aedes aEgypti. Parasit Vectors. (2022) 15:128. doi: 10.6084/m9.figshare.15129894
86. King JG, Souto-Maior C, Sartori LM, Maciel-de-Freitas R, and Gomes MGM. Variation in Wolbachia effects on Aedes mosquitoes as a determinant of invasiveness and vectorial capacity. Nat Commun. (2018) 9:1483. doi: 10.1038/s41467-018-03981-8
87. Yang Q, Chung J, Robinson KL, Schmidt TL, Ross PA, Liang J, et al. Sex-specific distribution and classification of Wolbachia infections and mitochondrial DNA haplogroups in Aedes albopictus from the Indo-Pacific. PloS Negl Trop D. (2022) 16:e0010139. doi: 10.1371/journal.pntd.0010139
88. Hoffmann AA, Ahmad NW, Keong WM, Ling CY, Ahmad NA, Golding N, et al. Introduction of Aedes aEgypti mosquitoes carrying wAlbB Wolbachia sharply decreases dengue incidence in disease hotspots. Iscience. (2024) 27:108942. doi: 10.26188/24314689.v1
89. Mancini MV, Murdochy SM, Bilgo E, Ant TH, Gingell D, Gnambani EJ, et al. Wolbachia strain wAlbB shows favourable characteristics for dengue control use in Aedes aEgypti from Burkina Faso. Environ Microbiol. (2024) 26:e16588. doi: 10.1111/1462-2920.16588
90. Gloria-Soria A, Ayala D, Bheecarry A, Calderon-Arguedas O, Chadee DD, Chiappero M, et al. Global genetic diversity of. Aedes aEgypti Mol Ecol. (2016) 25:5377–95. doi: 10.1111/mec.13866
91. Gomez Palacio A, Morinaga G, Rurner PE, Micieli MV, Elnour MAB, Salim B, et al. Robustness in population-structure and demographic-inference results derived from the Aedes aEgypti genotyping chip and whole-genome sequencing data. G3: Genes Genomes Genet. (2024) 14:jkae082. doi: 10.1093/g3journal/jkae082
Keywords: Asian tiger mosquito, Wolbachia, quantitative PCR, Arbovirus, biocontrol, longevity, bacterial density
Citation: Chuchuy A, Rodriguero MS and Micieli MV (2025) Strain‐specific quantification of Wolbachia density in subtropical Argentinean Aedes albopictus: effects of tissue location and longevity. Front. Insect Sci. 5:1655459. doi: 10.3389/finsc.2025.1655459
Received: 27 June 2025; Accepted: 15 September 2025;
Published: 30 September 2025.
Edited by:
Ana Laura Carbajal-de-la-Fuente, National Scientific and Technical Research Council (CONICET), ArgentinaReviewed by:
Jeronimo Augusto Alencar, Oswaldo Cruz Foundation (Fiocruz), BrazilTiago Melo, National Health Institute Doutor Ricardo Jorge (INSA), Portugal
Copyright © 2025 Chuchuy, Rodriguero and Micieli. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marcela S. Rodriguero, cm9kcmlndWVyb0BlZ2UuZmNlbi51YmEuYXI=
†Present address: Ailén Chuchuy, Facultad de Turismo y Urbanismo—Universidad Nacional de San Luis, San Luis, Argentina