 Martin Thiel1,2,3*
Martin Thiel1,2,3* Guillermo Luna-Jorquera1,2,3
Guillermo Luna-Jorquera1,2,3 Rocío Álvarez-Varas2,4Camila Gallardo1,2
Rocío Álvarez-Varas2,4Camila Gallardo1,2 Iván A. Hinojosa1,2,5
Iván A. Hinojosa1,2,5 Nicolás Luna1,2
Nicolás Luna1,2 Diego Miranda-Urbina6Naiti Morales1,2
Diego Miranda-Urbina6Naiti Morales1,2 Nicolas Ory1,2,7Aldo S. Pacheco8
Nicolas Ory1,2,7Aldo S. Pacheco8 Matías Portflitt-Toro1,2
Matías Portflitt-Toro1,2 Carlos Zavalaga9
Carlos Zavalaga9- 1Facultad de Ciencias del Mar, Universidad Católica del Norte, Coquimbo, Chile
- 2Millennium Nucleus Ecology and Sustainable Management of Oceanic Island, Coquimbo, Chile
- 3Centro de Estudios Avanzados en Zonas Áridas, Coquimbo, Chile
- 4Departamento de Ciencias Ecológicas, Facultad de Ciencias, Universidad de Chile, Santiago, Chile
- 5Facultad de Ciencias, Departamento de Ecología, Universidad Católica de la Santísima Concepción, Concepción, Chile
- 6Instituto de Ciencias Biológicas, Universidad de Talca, Talca, Chile
- 7GEOMAR Helmholtz Centre for Ocean Research Kiel, Kiel, Germany
- 8Instituto de Ciencias Naturales Alexander von Humboldt, Universidad de Antofagasta, Antofagasta, Chile
- 9Universidad Científica del Sur, Lima, Peru
Anthropogenic Marine Debris (AMD) in the SE Pacific has primarily local origins from land-based sources, including cities (coastal and inland), beach-goers, aquaculture, and fisheries. The low frequency of AMD colonized by oceanic biota (bryozoans, lepadid barnacles) suggests that most litter items from coastal waters of the Humboldt Current System (HCS) are pulled offshore into the South Pacific Subtropical Gyre (SPSG). The highest densities of floating micro- and macroplastics are reported from the SPSG. An extensive survey of photographic records, unpublished data, conference proceedings, and published studies revealed interactions with plastics for 97 species in the SE Pacific, including 20 species of fish, 5 sea turtles, 53 seabirds, and 19 marine mammals. Sea turtles are most affected by interactions with plastics, underlined by the fact that 4 of the 5 species suffer both from entanglement and ingestion. Reports gathered in this review suggest that interactions along the continental coast are mostly via entanglement. High frequencies of microplastic ingestion have been reported from planktivorous fish and seabirds inhabiting the oceanic waters and islands exposed to high densities of microplastics concentrated by oceanic currents in the SPSG. Our review also suggests that some species from the highly productive HCS face the risk of negative interactions with AMD, because food and plastic litter are concentrated in coastal front systems. In order to improve the conservation of marine vertebrates, especially of sea turtles, urgent measures of plastic reduction are needed.
Introduction
Marine plastic pollution is generating impacts on marine biota and ecosystems at many different levels (Ryan, 2016). Impacts are reported from a wide range of organisms, including microbiota, invertebrates, and vertebrates (Galloway et al., 2017; Law, 2017). An increasing number of reports document microplastic ingestion by marine invertebrates (Lusher, 2015); certain species also grow on large, floating plastic items, and can be transported to new habitats they had not previously inhabited (Kiessling et al., 2015). Interactions with vertebrates are best known, because vertebrates are larger and therefore more visible and recognizable than small marine invertebrates. Entanglement of seabirds and marine mammals in large plastic litter (nets, ropes, etc.) has been known since the early 1970s (Derraik, 2002). Similarly, ingestion of microplastics by fishes and seabirds is well known since about the same time period (Kenyon and Kridler, 1969; Carpenter et al., 1972; Ryan, 1987), and the number of affected species, such as seabirds (Wilcox et al., 2015), is continuously increasing.
The risk of interactions between marine organisms and plastics is not equal across the oceans. It depends on feeding biology and amount of plastic litter in the environment where the organisms are foraging. For example, seabird species feeding at the sea surface are more susceptible to plastic ingestion than diving species (Ryan, 1987). Species that ingest small microplastics, such as many fishes and surface-foraging seabirds might be at highest risk in areas where microplastics concentrate, such as the subtropical gyres, whereas species ingesting larger plastic items could potentially encounter these closer to the continental coasts where rivers and other human activities spill and accumulate large quantities of plastic litter (Rech et al., 2014, 2015; Di-Méglio and Campana, 2017; Fossi et al., 2017). Indeed, many reports of meso- and macroplastics ingestion include sea turtles and whales stranded on continental shores (Schuyler et al., 2014; Lusher et al., 2018). Similarly, the risk of entanglement for marine vertebrates is likely to be higher in areas with large amounts of derelict fishing gear, such as the North Pacific subtropical convergence zone (Pichel et al., 2007) or coastal areas where ghost nets accumulate (Wilcox et al., 2013).
These considerations suggest that the risk of harmful interactions with marine plastic pollution depends on (a) the biology of the species, and (b) the distribution and abundance of the different plastic types. To examine these predictions, herein we gather reports of interactions with plastic litter for marine vertebrates in the SE Pacific. We compare reports from the highly productive Humboldt Current System (HCS) with those from the oligotrophic open ocean, in particular the Easter Island ecoregion close to the South Pacific Subtropical Gyre (SPSG) accumulation zone (for methodological details see Supplement 1).
Plastic Litter in the South-East Pacific
Litter Sources and Pathways in the South-East Pacific
In the South Pacific Ocean, anthropogenic marine debris (AMD) originates from the surrounding landmasses and oceanic sources (Thiel et al., 2003, 2013; Kiessling et al., 2017). Especially in the eastern part of the S Pacific, litter pollution comes from land sources (e.g., Hidalgo-Ruz et al., 2018), beach-goers (Bravo et al., 2009), and marine activities including aquaculture (Astudillo et al., 2009; Hinojosa and Thiel, 2009), coastal (Perez-Venegas et al., 2017) and high seas fisheries (Kiessling et al., 2017). Rivers also contribute large amounts of macro- and microplastics (Rech et al., 2014, 2015).
The majority of the litter from land sources is probably trapped in coastal waters or on shores very close to its sources (Hinojosa and Thiel, 2009; Thiel et al., 2011; Rech et al., 2014). The low incidence of marine organisms growing on marine litter stranding on the continental coasts of the SE Pacific suggests that this fouling-free litter has likely passed very little time (if at all) at sea. On the other hand, strong offshore currents within the HCS are thought to move floating plastics quickly to the open ocean, where they become trapped in the SPSG (Martinez et al., 2009).
Floating Macro- and Microplastics in the SE Pacific
Given the multiple sources and transport dynamics of floating AMD in the SE Pacific, the density of macroplastics is high in immediate coastal waters, but rapidly decreases further away from the continental coast (Thiel et al., 2003; Miranda-Urbina et al., 2015). When approaching the center of the SPSG, densities of floating AMD reach very high abundances, as also reflected in recent data taken in two surveys conducted in 2015 and 2016 (Figure 1). A total of 477 items of floating AMD were observed across both surveys, the majority corresponding to 2015 when a major area including the Easter Island ecoregion was surveyed (Figure 1). Of the total floating AMD, 77% were macroplastics, most of which were large plastic fragments (95.4%). Other items included lines (17.7%), buoys (7.6%), plastic trays (4.9%), and plastic bags and nets (2.5 and 2.2%, respectively) (Figure 1). Accounting for the total number of AMD in both years, it seems that the distribution of floating AMD is comparable to patterns previously determined for the SE Pacific (see Miranda-Urbina et al., 2015). Out of the total (477 items), 8% were found in the HCS, only 3% occurred around Juan Fernandez and Desventuradas Islands, while 78 and 11% of all floating AMD were concentrated in the Oceanic and Polynesian sector, respectively. Large numbers of marine litter accumulate on beaches of Rapa Nui (Easter Island) and the uninhabited island Salas and Gómez in the Polynesian sector (Kiessling et al., 2017). Furthermore, marine litter on Salas and Gómez imposes a severe risk of entanglement for several seabird species breeding on the island (Miranda-Urbina et al., 2015).
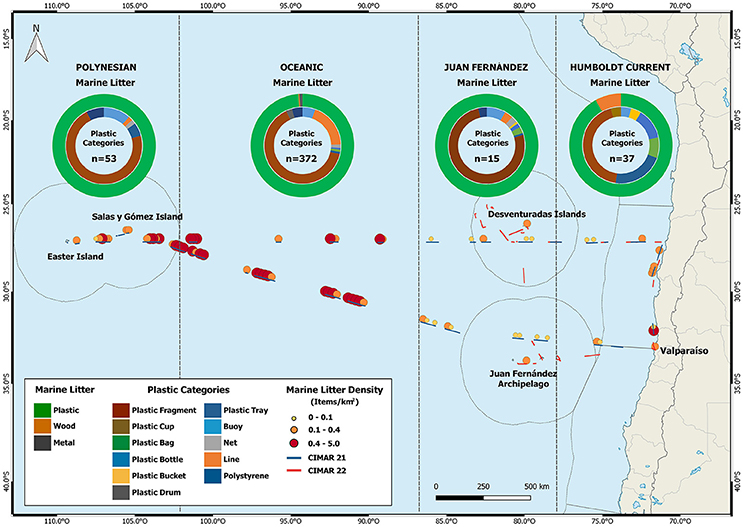
Figure 1. Types and density of marine debris in different sectors of the central SE Pacific, in 2015 (CIMAR 21) and 2016 (CIMAR 22). Open circles at the top indicate the proportions of different plastic types; numbers inside the circles represent the total of items observed in each sector. Dots show the density of marine debris from visual ship surveys (for details see Miranda-Urbina et al., 2015). Thin lines show the Exclusive Economic Zone.
The distribution of floating microplastics in the SE Pacific shows the typical distribution documented for other ocean basins (Eriksen et al., 2014; Law, 2017), with highest concentrations in the subtropical gyre (Eriksen et al., 2013, 2018). Microplastic densities in the HCS generally remain far below the densities reported from the gyre (Figure 2; NO, unpublished data). This distribution pattern is also well reflected in concentrations of small plastics from sandy beaches in the SE Pacific, where beaches along the HCS feature moderate densities, while sandy beaches on Rapa Nui have very high densities (Hidalgo-Ruz and Thiel, 2013). This pattern, with the abundances of microplastics increasing with distance from the continental coast (Figure 2), is also suggestive of progressive fragmentation of large plastics during their journey toward the SPSG.
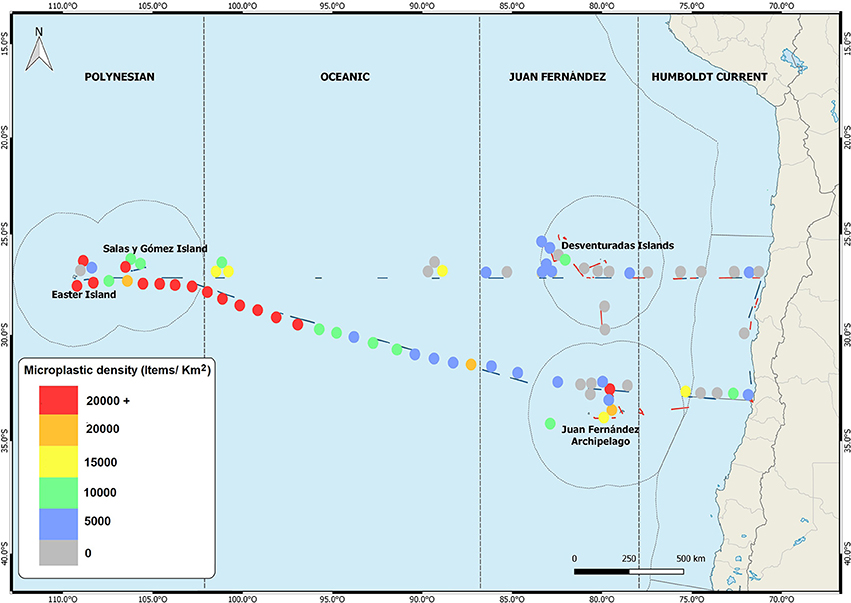
Figure 2. Density of microplastics (0.3–5 mm) in the central SE Pacific, in 2015 (CIMAR 21) and 2016 (CIMAR 22). Data from Eriksen et al. (2013, 2018) and MT, unpublished data. Thin lines show the Exclusive Economic Zone.
Interactions of Marine Vertebrates From the SE Pacific With Plastic Litter
A large number of marine vertebrate species from the SE Pacific were documented to have interacted with marine litter, including fishes, seabirds, sea turtles, and marine mammals (Thiel et al., 2011; Miranda-Urbina et al., 2015; Ory et al., 2017). In revising and compiling information from diverse sources we found that marine litter affects at least 97 different species (see Table 1 and text below for details). These reports comprise those of entanglement (including incorporation of plastics in seabird nests) and the ingestion of plastics.

Table 1. Reports on entanglement with macroplastics or plastic ingestion by marine vertebrates from open ocean (OO) and continental coastal (CC) (<5 nm from the land) waters of the southeast Pacific, based on literature, or anecdotal reports.
Fishes
The few studies documenting the interaction of microplastics with fishes in the SE Pacific revealed a high incidence of microplastics in planktivorous fish from the coast of Rapa Nui, located within the SPSG: 80% of the examined individuals of the amberstripe scad Decapterus muroadsi (Ory et al., 2017) and 14% of the flying fish Cheilopogon rapanouienesis (Chagnon et al., unpublished data) had ingested microplastics, many of which were similar in size and color to blue-pigmented planktonic organisms (copepods, crustacean larvae), which are common prey of these fish (Figure 3B). These findings suggest that visually-oriented planktivorous fish mostly ingest microplastics resembling their prey. The study of the digestive tracts of 7 planktivorous fish species from the HCS revealed that, overall, few of the fish analyzed (~2%) had ingested microplastics (Ory et al., 2018). Microplastics were also found more often (23% of all individuals analyzed) in herbivorous fish from Rapa Nui than in bottom-dwelling carnivorous and herbivorous fish species from the HCS (Table 1; NO, unpublished data). Such contrasting ingestion of microplastics well reflects the observed gradient of low microplastic densities in surface waters along the SE Pacific continental coast and high densities within the SPSG (Eriksen et al., 2013, 2014, 2018), and in particular along the Rapa Nui coast (Ory et al., 2017; Eriksen et al., 2018).

Figure 3. Examples of ingestion of plastic litter by marine species. (A) Microplastics collected from the stomach of one Christmas shearwater Puffinus nativitatis found dead on Salas y Gómez Island; Image by Matías Portflitt Toro. (B) Blue microplastics found in the stomach of amberstripe scads Decapterus muroadsi, fished at Rapa Nui; Image by Nicolas Ory. (C) Meso- and macroplastics found obstructing the cloacal opening of a green turtle Chelonia mydas found near Antofagasta. After carefully taking out these items the turtle recovered and later was liberated by the Wildlife Rescue and Rehabilitation Centre (Centro de Rescate y Rehabilitación de Fauna Silvestre) from Universidad de Antofagasta, Chile; Photo by Guerra-Correa et al. (2007). (D) Meso- and macroplastics found in the stomach of a blue shark Prionace glauca; Image courtesy of Carlos Canales-Cerro.
Apex predators play an important role in the exchange of energy between upper trophic levels in the marine environment (Markaida and Sosa-Nishizaki, 2010). However, little is known about the interaction of these species with AMD (Choy and Drazen, 2013). Plastics have been documented in the stomachs of tuna from Rapa Nui (Table 1), but little is known about plastic ingestion by other piscivorous fish from the open ocean. Publications on the diet of piscivorous fishes from the continental coast have not reported plastics (Fariña and Ojeda, 1993; Fariña et al., 2000), which indeed have not been found in the stomach contents of the investigated fish species (Jose Miguel Fariña, personal comment). Similarly, plastic ingestion by oceanic sharks seems to be very rare, estimated to occur in <1% of all analyzed stomachs (Sebastian Klarian, personal comment). Plastics found in sharks were PVC fragments in deep-sea species, and (positively buoyant) bottle and pen caps in coastal species (S. Klarian, personal comment; Figure 3D).
Reports on entangled fish are scarce, but there are three notable exceptions. In December 2016, an entangled Pacific chub Kyphosus sandwicensis was observed in a derelict fishing net found floating in the SPSG (28°23′S, 105°42′W) (Table 1). There were several living chubs swimming around the mass of net and ropes, and it is likely that the individual was trapped and died while this raft floated in the gyre. On the coast of Rapa Nui a surgeon fish Acanthurus leucopareius was entangled in a fishing line (NL, personal observation). The other case of an entangled fish is of a juvenile Galapagos shark Carcharhinus galapagensis that was documented in the waters around Rapa Nui. In June 2017 one specimen (about 160 cm in total length) of C. galapagensis was recorded swimming in shore waters with a plastic collar-like debris obstructing its gill region (Figure 4A). The animal appeared otherwise healthy, presenting an active feeding behavior. It showed initial tissue damage, which indicates that the collar recently became attached to the animal; similar records have shown that the internal projections of plastic collars have the potential to severely damage the tissue by affecting normal feeding and ventilation (Sazima et al., 2002), followed by body deformations as the animal grows (Wegner and Cartamil, 2012). The plastic ring attached to the animal was identified as the screwing part of a plastic barrel; this type of plastic debris is most likely coming from the industrial fishery that operates in the S Pacific and is an important source of recognizable plastic litter stranding on the shores of Rapa Nui (Kiessling et al., 2017).

Figure 4. Examples of species entangled with marine plastic litter. (A) Galapagos shark Carcharhinus galapagensis from coastal waters of Rapa Nui (Easter Island) entangled with a closure ring for plastic drums; Image Naiti Morales. (B) Humpback whale Megaptera novaeangliae from the Peruvian coast entangled in large fishing net; Image Aldo Pacheco. (C) Inca tern Larosterna inca found entangled in remains of fishing net near Valparaiso; Image courtesy of Fernanda Barilari. (D) Red-legged cormorant Phalacrocorax gaimardi from Taltal in nest; Image courtesy of Ivan Torres.
Continental coastal species of Chondrichthyes are also known to interact with marine litter. Oviparous species like cat sharks (Scyliorhinidae) and skates from the genus Symperygia lay egg capsules with long tendrils (Oddone and Vooren, 2002, 2008; Hernández et al., 2005; Flammang et al., 2007; Concha et al., 2013). These long tendrils are used to entangle the egg capsule to different substrata in order to maintain the vertical positioning and facilitate oxygen flow (Flammang et al., 2007). Along the central coast of Chile, dense multispecies coils of capsules are commonly found firmly attached to algae and/or plastic debris floating close to the coast or stranded on the shores after storms (MT and NM, personal observations).
Sea Turtles
Sea turtles are exposed to a variety of anthropogenic stressors, including marine plastic pollution, because of their use of diverse habitats, migratory behavior, and complex life histories (Nelms et al., 2016). Indeed, litter ingestion and entanglement in plastic debris have been recognized as serious threats to these species worldwide (Nelms et al., 2016; Clukey et al., 2017; Duncan et al., 2017). Five sea turtle species inhabit the SE Pacific (Caretta caretta, Chelonia mydas, Dermochelys coriacea, Eretmochelys imbricata, and Lepidochelys olivacea); all are listed from vulnerable to critically endangered on the IUCN Red List (IUCN, 2018) and have documented interactions with marine litter.
The green turtle (C. mydas) is the species most commonly mentioned to have ingested plastic items, with a frequency ranging from 28% in the Ecuadorian part of the HCS (Alemán, 2014) to 56 and 91% in Peru (Alfaro-Shigueto et al., 2005; Jiménez et al., 2017). The olive ridley turtle (L. olivacea) also has a high incidence of plastic ingestion, reaching up to 43% in Ecuador (Alemán, 2014), but this species has a lower incidence in other parts of the HCS (8%), both in Peru and southern Chile (de Paz et al., 2005; Brito et al., 2007). Furthermore, specific cases of plastic ingestion have been reported for leatherback turtles (D. coriacea) from the HCS in southern Peru and central Chile (Brito, 2001; IMARPE, 2011) and a hawksbill turtle (E. imbricata) in Rapa Nui (Brain et al., 2015).
Items most commonly found in stomachs or intestines of sea turtles are plastic pieces of intermediate size, including plastic bags, monofilament nylon, rope, and fishing nets (Brito, 2001; Guerra-Correa et al., 2007; IMARPE, 2011; Jiménez et al., 2017; Figure 3C). Several authors suggested that plastic ingestion has been the cause of death of stranded turtles in Ecuador and Chile (Brito et al., 2007; Silva et al., 2007; Alemán, 2014).
Even though many studies have focused on evaluating sea turtle bycatch rates (in active fishing gear) in the Pacific Ocean (Wallace et al., 2010), to date almost no reports exist on sea turtle entanglements (derelict fishing gear) in the region (Nelms et al., 2016). In fact, to our knowledge no peer-reviewed articles have informed incidents of entanglements from the SE Pacific. In Rapa Nui, a case of C. caretta with fishing line in both anterior flippers caused their amputation and subsequent death a few hours later (RAV, personal observation). In addition, several cases of entanglements of green and olive ridley turtles have been informed from Ecuador (Rubén Alemán, personal communication).
Seabirds
Many different seabird species have been reported to be entangled in marine debris or have ingested plastic (Table 1). Interestingly, for most fish-feeding species from the HCS, the incidence of individuals with microplastics in their stomachs is low, although these species face other threats to their conservation (Luna-Jorquera et al., 2012). We found 6 species in which plastic litter has been found in their stomachs, 3 of them (Pelecanoides garnotii, P. urinatrix, Phalacrocorax bougainvillii, and Spheniscus humboldti) being true diving species, and one a plunge diver (Pelecanus thagus). One species with relatively high frequency of plastic ingestion is the kelp gull Larus dominicanus, which is commonly observed feeding in fishing ports, at garbage containers, and on waste disposal facilities.
In addition to the low number of continental species with plastic in their stomach, it seems that the number of affected individuals per species is relatively low: 10 out of 450 examined individuals (2.2%) S. humboldti, 4 out of 103 (3.9%) Pelecanoides garnotii, and 12 out of 363 studied pellets (3.3%) of Phalacrocorax bougainvillii (CZ, unpublished data). The diet of the Humboldt penguin S. humboldti has also been examined in another study, but the authors did not mention any plastic items in the species' stomach contents (Herling et al., 2005). The tendency of low incidence of plastic ingestion in seabirds inhabiting the HCS is supported by several other studies. Jahncke et al. (1997) and García-Godos and Goya (2006) studied the diet of P. bougainvillii, Sula variegata, and Pelecanoides garnotii, and did not find any plastic artifacts in any of the examined individuals (I. García-Godos, personal communication).
In contrast to continental seabirds, oceanic species are severely affected (see Table 1). Of the 37 seabird species from the SE Pacific with reported ingestion of plastic, 31 are oceanic and the majority are Procellariiformes, which retain particles for several months before the particles are evacuated from the digestive system by regurgitation (Terepocki et al., 2017; Figure 3A). The high incidence of microplastic ingestion in the species listed in Table 1 is very likely due to the high concentration of AMD observed in the Oceanic and Polynesian sectors of the SE Pacific (see Figure 2). Several harmful effects have been reported due to microplastic ingestion, ranging from stomach ulcers, intestinal obstruction, reduced body condition, and increased contaminant load (Derraik, 2002; Lavers et al., 2014).
Body injuries, severe negative effects on behavior, and even mortality, are typical consequences of seabird entanglement in floating or stranded marine litter. As opposed to the observed tendency of plastic ingestion, entanglement mostly affects continental species. Seventeen seabird species inhabiting the HCS are affected by entanglement, which most of the time occurs during foraging activities at sea, when seabirds are trapped in discarded fishing lines, derelict fishing nets (Figure 4C) (Thiel et al., 2011), and single-use plastic bags on beaches (see Supplements 2, 3). We have also observed that lines and rope fragments discarded by aquaculture activities affect coastal species, such as gulls and cormorants (Figure 4C) (MPT and GLJ, own unpublished data). Entanglement in derelict fishing nets is also affecting diving waterbirds, including grebes (Podilymbus podiceps, Photo PP01 in Supplement 3 by P. Valencia, and Podiceps occipitalis, Portflitt-Toro et al., 2016). During the winter season, grebes are often observed foraging in nearshore waters in the bays of the Coastal System of Coquimbo, where fishermen use gillnets to capture pelagic fishes.
A handful of seabird species are affected by both entanglement and ingestion (Larus dominicanus and Phalacrocorax bougainvilli), but the negative effects of marine litter are also reaching the nests of several species. The incidence of anthropogenic marine litter for nest construction is not well known for the SE Pacific, but gulls from the Peruvian coast have been reported to use plastic in their nests (Stucchi and Figueroa, 2006). Our revision revealed that 12 species are using litter for nest construction, comprising both continental and oceanic seabirds; at least 10 of those 12 species are also exposed to organism entanglement and plastic ingestion at sea. Species like cormorants (Phalacrocorax spp.), frigatebirds (Fregata minor), and albatrosses (Thalassarche spp.) are actively selecting and transporting marine litter to their nests, while other species inhabiting the oceanic Salas and Gómez Island are affected by plastic litter accumulated by oceanic currents near their foraging grounds. Abundant incorporation of plastic litter in nests (see Figure 4D) calls for further research due to (i) the risk of entanglement of adults and chicks, and (ii) the potential effects of anthropogenic litter on the thermal properties of the nest both during nesting and chick-rearing phases.
Marine Mammals
There is no published information regarding the ingestion of marine debris for marine mammals in the SE Pacific. Overall, few studies examined the stomach contents of marine mammals. In a study on the diet of the South American sea lion Otaria flavescens, George-Nascimento et al. (1985) did not report any plastic items in stomach contents. Feces from the South American sea otter Lontra felina contained a diverse range of prey remains from fishes, crustaceans, and molluscs, but no plastic items were listed (Medina-Vogel et al., 2004; Córdova et al., 2009). A study of four delphinid species captured along the central coast of Peru reported prey items in stomach contents based on fish otoliths and cephalopod beaks, but no marine debris was documented (García-Godos et al., 2007). In southern Chile, an examination of seven stranded false killer whale carcasses reported only empty stomachs (Haro et al., 2015). Similarly, a study on the diet of long-finned pilot whales from southern Chile (Mansilla et al., 2012) found no plastic particles or marine litter (Carlos Olavarría, personal communication). However, the case of a Risso's dolphin Grampus griseus with a plastic bag in its mouth (Photo by M. Bravo) in the Coastal System of Coquimbo, in northern-central Chile, shows that plastic ingestion by cetaceans should not be ruled out.
Reports on the entanglement of marine mammals along the SE Pacific are much more common than reports about plastic ingestion. Throughout the HCS, fisheries are very intense at both artisanal and industrial levels (Thiel et al., 2007; Alfaro-Shigueto et al., 2010), and many of the observed entanglements are likely to have occurred with active fishing gear. An important number of pinnipeds, large baleen whales, and Odontocetes in general (dolphins, porpoises, and toothed whales) have been reported entangled in fishing gear throughout neritic and oceanic waters off Peru and Chile (Table 1). Entanglements may occur when large gillnets are deployed at the bottom or drifting near the sea surface, depending on the fishery (Aguayo-Lobo, 1999; Alfaro-Shigueto et al., 2010). Marine mammals swim or dive through the mesh and become entangled. Small animals, such as sea otters and sea lions, may die shortly after entanglement, as these species may not have the necessary strength to escape from nets and being air-breathers will consequently drown. This is probably also the case for dolphins such as the dusky dolphin (Lagenorhynchus obscurus), bottlenose dolphin (Tursiops truncatus), long-beaked common dolphin (Delphinus capensis), and the Burmeister porpoise (Phocoena spinipinnis) in Peru. In addition to the aforementioned species, in Chile the southern right whale dolphin (Lissodelphis peronii), Commerson's dolphin (Cephalorhynchus commersonii), Chilean dolphin (Cephalorhynchus eutropia), and Peale's dolphin (Lagenorhynchus australis) also suffer mortality from entanglements (Table 1).
Large baleen whales get into nets but may keep moving, subsequently carrying large pieces of entangled nets on their body. Along the HCS of Peru, the humpback whale Megaptera novaeangliae is the most affected large whale species. Campbell et al. (2017) reported that 51% of stranded whales off Peru were humpback whales and all individuals showed evidence of entanglement (Figure 4B). Observations made during daily sightings of humpback whales throughout their breeding migration (mid July to late October, Guidino et al., 2014) in northern Peru (~4°S) suggest that in the HCS, humpback whales frequently get entangled with drifting gillnets (ASP, unpublished data). As the stranding data suggest (Campbell et al., 2017), the fate of entangled whales is often death, unless the net is quickly removed from the whale.
Along the HCS of Chile, artificial coastal structures such as breakwaters and harbors appear to accumulate more marine debris than the natural rocky intertidal shore (Aguilera et al., 2016). This accumulation could be a problem for the marine otter (Lontra felina), a small mustelid endemic along the Pacific coast of South America. This species uses natural rocks or artificial constructions as habitat. At the northern-most location of its distributional range in northern Peru, the marine otter has been observed resting in an artificial cave full of marine debris (see Figure 3 in Alfaro-Shigueto et al., 2011). While this case does not constitute a direct interaction with marine plastic litter, the risk for entanglement or ingestion is evident.
Overview of Plastic Ingestion and Entanglement by Marine Vertebrates From the SE Pacific
This first review of interactions with AMD of marine vertebrates from the SE Pacific reported a total of 97 species (Table 1). Seabirds represent nearly 55% of the total, followed by fishes with 21%, marine mammals with 19%, and sea turtles with 5%. Considering the number of species per taxonomic group, the type of interaction (ingestion or entanglement), and environment (continental or oceanic), an overall pattern is emerging. For fishes, more species with plastic ingestion were documented along the continental coast than in the open ocean, and few oceanic species become entangled near oceanic islands (Rapa Nui and Salas and Gómez Island). However, it needs to be taken into account that the simple species list presented in Table 1 only shows whether a species had ingested microplastics or not, which does not allow for inferences on the individual risk of ingestion. For example, in the case of the Peruvian anchoveta Engraulis ringens from the HCS, of 116 studied individuals only one (<1%) had one microplastic in its stomach (Ory et al., 2018), whereas of 20 amberstrip scads Decapterus muroadsi from Rapa Nui 16 individuals (80%) had ingested on average 2.5 microplastics per individual (Ory et al., 2017), underscoring that the risk of microplastic ingestion is much higher for oceanic planktivorous species than for species from the HCS. In contrast to the general pattern documented for fishes, the incidence of microplastic ingestion in seabirds is much higher for oceanic species than for those from the HCS, but the latter are suffering intense entanglement (Table 2). Marine mammals are scarce in oceanic waters, so marine mammals are principally affected by entanglement with AMD floating in the productive waters of the HCS (Table 2). Regarding sea turtles, none of the species from the SE Pacific are safe from interactions with AMD, ingestion or entanglement, and thus are equally threatened in coastal and oceanic waters. This overall pattern agrees well with the spatial distribution of micro- and macroplastics in the SE Pacific (see Figures 1, 2). Future systematic studies in the SE Pacific should provide more information about the actual number of species affected, which will also help authorities to improve efforts for efficient solutions.
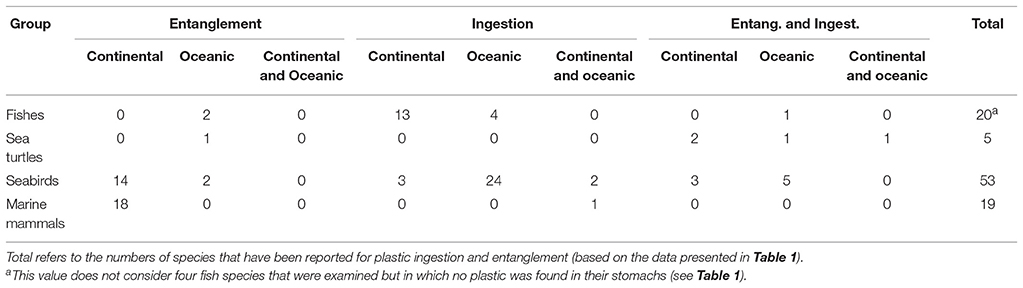
Table 2. Number of species of marine vertebrates for which ingestion and entanglement has been documented.
Other Interactions
Similar to other oceans (Carson, 2013), in the SE Pacific litter is frequently bitten by large marine organisms, which are thought to be vertebrates (Eriksen et al., 2017), but might also originate from large invertebrates with powerful jaws, in particular squids and/or cuttlefish. On Rapa Nui, up to 10% of stranded AMD can have bitemarks (MT, unpublished data).
Discussion
Ingestion of Plastics
The results from this review indicate that microplastic ingestion is uncommon along the Pacific coast of South America. Neither fishes nor seabirds from the continental coast had high frequencies of microplastic ingestion, and information on the diet of marine mammals in the SE Pacific is very limited and thus does not allow inference on the risk of microplastic ingestion. An exception from this pattern seems to be the relatively high incidences of plastic ingestion in sea turtles in the HCS, but most of those plastics are of larger sizes and can be characterized as meso- and macroplastics. Also, intertidal habitats, such as beaches, tidepools, estuarine saltmarshes, and especially the seashores in the fjords of southern Chile require future research attention, because intense microplastic pollution may cause localized impacts in species from shore habitats (see e.g., Mizraji et al., 2017).
The low incidences of microplastic ingestion in most marine vertebrates from the HCS could be resulting from low concentrations of microplastics in coastal waters or due to specific foraging behaviors, or a combination of both. A data comparison from the SPSG and from other parts of the world can shed some light on these questions. For example, some planktivorous fish species from the SPSG had a very high frequency of microplastic ingestion (Ory et al., 2017), and other species had ingested microplastics more frequently than any of the planktivorous species from the HCS (Chagnon et al., unpublished data; Ory et al., 2018). For seabirds, the pattern was similar: microplastic ingestion was much more common in oceanic species than in species from the HCS (Tables 1, 2), regardless of feeding types (planktivorous, invertivorous, and piscivorous). This suggests that the observed pattern might not be due to differences in foraging and feeding behaviors, but rather to differences in microplastic abundances. Nevertheless, plastic ingestion was also documented in a number of species from the HCS, which might be consequence of their biology. For example, incidence of plastics in the digestive system of different seabird species is related to their foraging behavior (e.g., surface feeders, pursuit feeders, among others) and the digestive system morphology (i.e., muscular gizzard in petrels) (Ainley et al., 1990; Spear et al., 1995; Roman et al., 2016). This latter aspect causes indigestible items to become trapped in the bird's gut system (Ainley et al., 1990).
Other factors such as the geographic distribution may impact the amount of plastic ingested, as found by Spear et al. (1995), who reported that seabirds foraging predominantly in the North Pacific had higher incidence of ingestion than species from the South Pacific. However, the paper suggested that this pattern may be biased by the lack of studies from the SE Pacific. This is very similar for marine mammals, in particular dolphins and whales. Since the end of whaling in the region during the early 80's, the scientific examination of body parts (including stomachs) of large baleen and sperm whale carcasses have considerably ceased in the region. Scientific treatment of stranded animals is limited due to the difficult logistics needed to examine cases of cetacean strandings in remote areas (e.g., Haro et al., 2015; Häussermann et al., 2017). In this regard, there is an urgent need for the implementation of effective action plans for the scientific treatment of stranded whales and dolphins, if we aim to understand the impacts of the ingestion of marine litter in megafauna. This is particularly important since researchers are adopting non-invasive methods (e.g., stable isotopes, DNA analysis) for both live (Haro et al., 2016) and stranded charismatic animals. At present, it is difficult to understand the magnitude of the plastic ingestion problem for marine mammals in the SE Pacific. However, plastic ingestion by diverse cetacean species has been reported in several coastal areas elsewhere (Baulch and Perry, 2014; Lusher et al., 2018), often with lethal consequences (see Jacobsen et al., 2010; de Stephanis et al., 2013; for cases on sperm whales). Microplastics of several polymer types have been documented for the first time in a humpback whale stranded on the coast of the Netherlands (Besseling et al., 2015). In the SE Pacific, the problem is likely being underestimated.
The relatively high frequency of bitemarks in plastics stranded on Rapa Nui shores indicates that some species directly bite into floating plastics. It is currently not well known which species engage in this behavior and why (Carson, 2013), but most of the bitemarks found on plastics from Rapa Nui resemble those of the green turtle Chelonia mydas (Eriksen et al., 2017), a species commonly reported to ingest larger plastic pieces (see above). The potential risk of plastic ingestion as a result of biting into floating plastic litter is reason for concern.
Entanglement
Entanglement reports of fishes are very rare, while they are common for seabirds, marine mammals, and sea turtles. The lack of reports from fishes might be due to the fact that mortality at sea would immediately cause sinking, whereas seabird, mammal, and sea turtle carcasses float at the sea surface; not surprisingly many reports of entanglement come from dead animals (see above).
From all the records of top fish predators interacting with plastic debris around the world, carcharhinid sharks seem to be most at risk of entanglement (Laist, 1997; Sazima et al., 2002; Ceccarelli, 2009), probably due the high abundance and species diversity in this group (Compagno, 1984). Cliff et al. (2002) reported an increase over time in entanglement of a carcharhinid species from South Africa, which furthermore underscores that species from this group are at highest risk of negative interactions with floating litter. Oceanic shark species are also likely to be impacted by plastic debris, but the limited number of studies on oceanic sharks underscores that more research is required to determine the full extent of this problem.
Seabird entanglement is common in the world's oceans (Kühn et al., 2015). Herein entanglement was reported mostly for species from the HCS, and observations included fisheries litter (nets, lines) and consumer plastics (mostly plastic bags). While interactions with fisheries items are likely to happen at sea (e.g., Moore et al., 2009), entanglement with consumer plastics may occur on the shore or at waste disposal facilities, as highlighted by the frequent observations of kelp gulls L. dominicanus with plastic bag entanglement (Table 1). Our data suggest that entanglement is more common in species from the continental coast than in oceanic species. The fact that Procellariiformes (which are mostly oceanic species) have the lowest frequencies of entanglement (Kühn et al., 2015) seems to support this pattern. However, herein we observed plastic litter in the nests of several oceanic species (Table 1), which underscores the imminent risk of entanglement for these species.
Although sea turtle entanglement in AMD has been recognized as a cause of mortality globally, there are quantitative knowledge gaps on rates and population implications (Duncan et al., 2017). Recent studies report entanglements across all species, life stages, and ocean basins, with higher vulnerability in pelagic juveniles (Nelms et al., 2016; Duncan et al., 2017). However, reports of entanglement incidents in the scientific literature are scarce and it is likely that many individual cases are never published, and therefore these data may be highly underestimated (Nelms et al., 2016). Duncan et al. (2017) reported that derelict fishing gear contributed globally to the majority of entanglements, while debris from land-based sources contributed to a lesser extent. Entanglements are a greater threat to sea turtles than climate change and direct exploitation, but less of a threat than plastic ingestion and bycatch (active gear) (Duncan et al., 2017).
Incidences of entanglement were reported for many species of marine mammals from the SE Pacific (Table 1), mostly with items of fisheries origin. Similar to other areas of the southern hemisphere (e.g., Page et al., 2004), sea lions seem to be most at risk of entanglement with derelict fishing gear as indicated by several independent observations recorded herein (Table 1). However, our review also reveals that many whale species from the HCS become entangled in (active?) fishing gear. Some efforts are being conducted to mitigate the entanglement problem. In Peru, the implementation of acoustic alarms (pingers) has proven to have dissuasive effects, thus reducing dolphin entanglements (Mangel et al., 2013). The use of modified long lines in the Patagonian toothfish (Dissostichus eleginoides) fishery in southern Chile reduces the interaction of killer and sperm whales with active fishing gear (Moreno et al., 2008). Yet, these efforts are only localized and eventually should be implemented at the whole scale of the respective fisheries.
Differential Risk of Marine Litter Interactions Across the Oceanic Gradient
Several studies highlight that risk of both ingestion and entanglement is highest where main foraging grounds overlap with accumulation areas of floating AMD (Wilcox et al., 2013; Fossi et al., 2017). Our study showed that some species foraging in the highly productive HCS frequently interact with marine plastics, which at first glance might be surprising given that litter densities are substantially lower than in the open ocean. However, many of these species feed in areas where hydrographic features, e.g., frontal systems or meso-scale gyres, concentrate food and also floating plastics (Pichel et al., 2007). Interestingly, one of the first direct observations of this phenomenon comes from the HCS off the central coast of Chile, where Bourne and Clark (1984) observed planktivorous seabirds feeding in a coastal front that also had concentrated large amounts of floating plastics. The high incidences of entanglement and also plastic ingestion, especially by sea turtles and some seabird species from the HCS, likely occurred in these temporary hotspots. These interactions are common in the productive upwelling systems of the eastern boundary currents (for overview see Scales et al., 2014), and cause high risk for marine vertebrates despite the fact that densities of floating litter are lower than in the subtropical gyres.
In the open ocean, especially in the oligotrophic subtropical gyres, marine productivity is low, and often concentrated above seamounts or near oceanic islands. If these islands are located within the range of the litter accumulation zones of the subtropical gyres, some species are at high risk of negative interactions with floating plastics (Figure 5). Our review showed that planktivorous fish and seabirds living on the oceanic islands in the vicinity of the SPSG have high incidences of microplastic ingestion, possibly due to the extraordinarily high densities of floating microplastics in this region (Figure 5). The limited number of entanglement reports from this area is likely a combination of lower densities of marine vertebrates in the subtropical gyres (see also Titmus and Hyrenbach, 2011) and the limited number of observers, compared to the continental coasts.
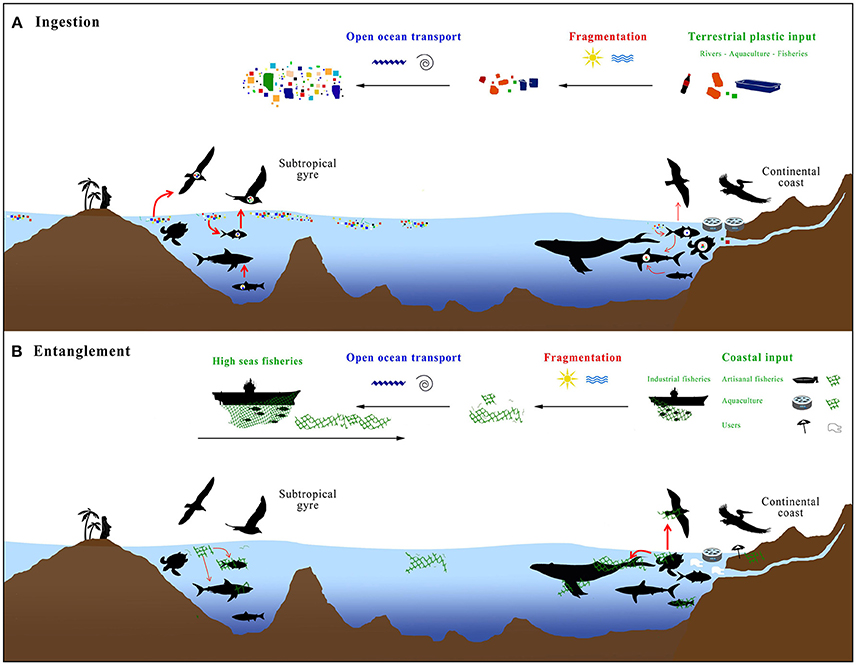
Figure 5. Conceptual model of (A) ingestion, and (B) entanglement by marine vertebrates with anthropogenic marine plastics, highlighting the litter sources and abiotic processes (upper part of figures) and the interactions with marine invertebrates (bottom part of figures).
Conclusions and Outlook
Herein interactions with marine plastic litter were documented for a total of 97 species of marine vertebrates. The risk of microplastic ingestion seems to be high in nearshore waters (including tidepools), decreases above the continental shelf of the eastern boundary currents, but again reaches very high probabilities in oceanic waters associated with the gyre accumulation zones, especially for fishes and seabirds (Figure 5). The current interaction records suggest that marine vertebrate species living in the productive waters of the HCS are at higher risk of facing entanglement than species from the open ocean, albeit several oceanic species have also been observed to be entangled in marine plastics, mostly from high seas fisheries (Figure 5).
Further systematic research on the ingestion and entanglement rates in marine vertebrates and their impacts on populations from the SE Pacific is required. Investigations to determine hotspots of marine plastic pollution will also enable prioritizing resources and to focus and steer conservation measures. Detailed stranding data and a centralized regional database are recommendable for a better documentation of negative interactions of marine vertebrates with plastic litter. Education, community involvement, together with effective measures to reduce the amounts of plastic litter entering the ocean, are essential to reduce the impact on marine vertebrates, particularly the highly threatened sea turtles.
Ethics Statement
The paper is based mostly on previously published information and does not include any sampling and laboratory analysis of marine organisms. Therefore, no ethics approval was required.
Author Contributions
MT and GL-J designed the review, coordinated the team, and led the writing. RÁ-V, CG, IH, NL, DM-U, NM, NO, ASP, MP-T, and CZ contributed the observations and helped with the writing. IH and MP-T coordinated the quest for entanglement observations from citizen scientists. CG, DM-U, and MP-T prepared the figures.
Funding
This project received support from the Chilean Millennium Initiative to ESMOI (Millenium Nucleous Ecology and Sustainable Management of Oceanic Islands). Sampling of floating plastics in the South Pacific was funded by the Comité Oceanográfico Nacional (CONA) from Chile, project codes CONA C21-15-029 and CONA C22-16-11 to MT; and project codes CONA C21-15-113 and CONA C22-16-06 to GL-J. NO received support through a postdoctoral FONDECYT grant (No. 3150636) and NM received support through the CONICYT Beca Doctorado Nacional N°21151143, and NL received support from Beca Magister Nacional CONICYT N°22161894 from the Chilean Ministry of Education. Shark interactions with plastics were observed during the project entitled The lost fish from Easter Island funded by Save Our Seas Foundation Small Grant N°361.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank all citizen science observers for contributing their photographic records about entanglement. We are especially grateful to all our colleagues for freely sharing their insights, unpublished observations, and images that significantly enhanced the scientific value of this contribution. Two reviewers provided many helpful suggestions and Annie Mejaes kindly revised the English of the final manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2018.00238/full#supplementary-material
References
Aguayo-Lobo, A. (1999). The cetaceans and their conservation perspectives. Estud. Oceanol. 18, 35–43.
Aguilera, M. A., Broitman, B. R., and Thiel, M. (2016). Artificial breakwaters as garbage bins: structural complexity enhances anthropogenic litter accumulation in marine intertidal habitats. Environ. Pollut. 214, 737–747. doi: 10.1016/j.envpol.2016.04.058
Ainley, D. G., Spear, L. B., and Ribic, C. A. (1990). “The incidence of plastic in the diets of pelagic seabirds in the eastern equatorial pacific region,” in Proceedings of the Workshop on the Fate and Impact of Marine Debris, eds R. S. Shomura and M. L. Godfrey (Honolulu, HI: U.S. Dep. Commer., NOAA Tech. Memo. NMFS), 653–665.
Alemán, R. (2014). Informe de Actividades realizadas en Conservación de Tortugas Marinas durante el año 2013. Informe Técnico MAE-PN-PNMRA- 2014-N°010, 9.
Alfaro-Shigueto, J., Mangel, J. C., Pajuelo, M., Dutton, P. H., Seminoff, J. A., and Godley, B. J. (2010). Where small can have a large impact: structure and characterization of small-scale fisheries in Peru. Fish. Res. 106, 8–17. doi: 10.1016/j.fishres.2010.06.004
Alfaro-Shigueto, J., Montes, D., Acleto, C., Zuñiga, R., and Huamán, P. (2005). “Diet analysis from green turtle Chelonia mydas agassizii from central Peruvian Coast,” in Proceedings of the Twenty-first Annual Symposium on Sea Turtle Biology and Conservation, Vol. 368, eds M. Coyne and R. Clark (Philadelphia, PA: NOAA Technical Memorandum NMFS–SEFSC-528), 11.
Alfaro-Shigueto, J., Valqui, J., and Mangel, J. C. (2011). New record of the marine otter Lontra felina (Molina, 1782) north to its current distribution. Ecol. Apli. 10, 87–91. doi: 10.21704/rea.v10i1-2.417
Arce, P., Daigre, M., and Simeone, A. (2014). “Uso diferencial de basura para la construcción de nidos en aves marinas de una colonia en chile central,” in XI Congreso Chileno de Ornitología (La Serena).
Astudillo, J., Bravo, M., Dumont, C., and Thiel, M. (2009). Detached aquaculture buoys in the SE Pacific: potential dispersal vehicles for associated organisms. Aquat. Biol. 5, 219–231. doi: 10.3354/ab00151
Baulch, S., and Perry, C. (2014). Evaluating the impacts of marine debris on cetaceans. Mar. Pollut. Bull. 80, 210–221. doi: 10.1016/j.marpolbul.2013.12.050
Besseling, E., Foekema, E. M., Van Franeker, J. A., Leopold, M. F., Kühn, S., Bravo Rebolledo, E. L., et al. (2015). Microplastic in a macro filter feeder: humpback whale Megaptera novaeangliae. Mar. Pollut. Bull. 95, 248–252. doi: 10.1016/j.marpolbul.2015.04.007
Bourne, W. R. P., and Clark, G. C. (1984). The occurrence of birds and garbage at the Humboldt front off Valparaiso, Chile. Mar. Pollut. Bull. 15, 343–344. doi: 10.1016/0025-326X(84)90493-4
Brain, M. J., Medrano, C., Brito-Carrasco, B., Del Río, J., and Álvarez-Varas, R. (2015). “First rehabilitation case of hawksbill turtle (Eretmochelys imbricata) stranded in Easter Island, Chile: the need to allocate resources to increase the chances of success,” in Book of Abstracts of the 35th Annual Symposium on Sea Turtle Biology and Conservation, eds Y. Kaska, B. Sonmez, O. Turkecan, and C. Sezgin (Dalaman), 214.
Bravo, M., de Los Angeles Gallardo, M., Luna-Jorquera, G., Núñez, P., Vásquez, N., and Thiel, M. (2009). Anthropogenic debris on beaches in the SE Pacific (Chile): results from a national survey supported by volunteers. Mar. Pollut. Bull. 58, 1718–1726. doi: 10.1016/j.marpolbul.2009.06.017
Brito, C., Brito, J. L., Zúñiga, M., Campos, M., and Toro, S. (2007). “Tortugas marinas en el centro de rescate y rehabilitación de fauna silvestre del museo de San Antonio,” in VII Simposio Sobre Medio Ambiente: Estado Actual y Perspectivas de la Investigación y Conservación de las Tortugas Marinas en las Costas del Pacífico Sur Oriental. (Antofagasta), 96.
Brito, J. L. (2001). “Informe preliminar de tortugas marinas en Chile: su situación actual,” in Taller Nacional de Trabajo Para Definir las Líneas de Acción Prioritarias de un Programa Para la Conservación de las Tortugas Marinas (Valparaíso), 95.
Campbell, E., Alfaro-Shigueto, J., and Mangel, J. C. (2017). “Whale entanglements in Peru: frequency and cost,” in 22nd Biennial Conference on the Biology of Marine Mammals (Halifax).
Cañoles, R., Fuentes-Garrido, M., Vanerio, M., Suazo, G., Yates, O., and Cabezas, L. A. (2017). “Análisis de contenido estomacal en siete especies de aves marinas (Procellariiformes),” in XII Congreso Chileno de Ornitología (Santa Cruz).
Carpenter, E. J., Anderson, S. J., Harvey, G. R., Miklas, H. P., and Peck, B. B. (1972). Polystyrene spherules in coastal waters. Science 178, 749–750. doi: 10.1126/science.178.4062.749
Carson, H. S. (2013). The incidence of plastic ingestion by fishes: From the prey's perspective. Mar. Pollut. Bull. 74, 170–174. doi: 10.1016/j.marpolbul.2013.07.008
Ceccarelli, D. M. (2009). Impacts of Plastic debris on Australian Marine Wildlife. Report by C&R Consulting for the Department of the Environment, Water, Heritage and the Arts. Canberra, ACT, Australia.
Choy, C. A., and Drazen, J. C. (2013). Plastic for dinner? Observations of frequent debris ingestion by pelagic predatory fishes from the central North Pacific. Mar. Ecol. Prog. Ser. 485, 155–163. doi: 10.3354/meps10342
Cliff, G., Dudley, S. F., Ryan, P. G., and Singleton, N. (2002). Large sharks and plastic debris in KwaZulu-Natal, South Africa. Mar. Freshw. Res. 53, 575–581. doi: 10.1071/MF01146
Clukey, K. E., Lepczyk, C. A., Balazs, G. H., Work, T. M., and Lynch, J. M. (2017). Investigation of plastic debris ingestion by four species of sea turtles collected as bycatch in pelagic Pacific longline fisheries. Mar. Pollut. Bull. 120, 117–125. doi: 10.1016/j.marpolbul.2017.04.064
Compagno, J. V. L. (1984). FAO Species Catalogue Vol. 4, part 2 Sharks of the World: An Annotated and Illustrated Catalogue of Shark Species Known to Date. Rome: Food and Agriculture Organization of the United Nations.
Concha, F., Morales, N., and Larraguibel, J. (2013). Egg capsules of the Filetail fanskate Sympterygia lima (Poeppig 1835) (Rajiformes, Arhynchobatidae) from the southeastern Pacific Ocean, with observations on captive egg-laying. Ichthyol. Res. 60, 203–208. doi: 10.1007/s10228-012-0333-8
Córdova, O., Rau, J. R., Suazo, C. G., and Arriagada, A. (2009). Estudio comparativo de la ecología alimentaria del depredador de alto nivel trófico Lontra felina (Molina, 1782) (Carnivora: Mustelidae) en Chile. Rev. Biol. Mar. Oceanog. 44, 429–438. doi: 10.4067/S0718-19572009000200016
de Paz, N., Reyes, J., and Echegaray, M. (2005). “Capture and trade of marine turtles at San Andres, Southern Peru,” in Proceedings of the Twenty-First Annual Symposium on Sea Turtle Biology and Conservation, Vol. 368, eds M. Coyne and R. Clark (Philadelphia, PA: NOAA Technical Memorandum NMFS–SEFSC-528), 52.
Derraik, J. G. (2002). The pollution of the marine environment by plastic debris: a review. Mar. Pollut. Bull. 44, 842–852. doi: 10.1016/S0025-326X(02)00220-5
de Stephanis, R., Giménez, K., Carpinelli, E., Gutierrez-Exposito, C., and Cañadas, A. (2013). As main meal for sperm whales: plastic debris. Mar. Pollut. Bull. 69, 206–214. doi: 10.1016/j.marpolbul.2013.01.033
Di-Méglio, N., and Campana, I. (2017). Floating macro-litter along the Mediterranean French coast: composition, density, distribution and overlap with cetacean range. Mar. Pollut. Bull. 118, 155–166. doi: 10.1016/j.marpolbul.2017.02.026
Duncan, E. M., Boterelli, L. R., Broderick, A. C., Galloway, T., Lindeque, P. K., Nuno, A., et al. (2017). A global review of marine turtle entanglement in anthropogenic debris: a baseline for further action. Endang. Species. Res. 34, 431–448. doi: 10.3354/esr00865
Eriksen, M., Lebreton, L. C., Carson, H. S., Thiel, M., Moore, C. J., Borerro, J. C., et al. (2014). Plastic pollution in the world's oceans: more than 5 trillion plastic pieces weighing over 250,000 tons afloat at sea. PLoS ONE 9:e111913. doi: 10.1371/journal.pone.0111913
Eriksen, M., Liboiron, M., Kiessling, T., Charron, L., Alling, A., Lebreton, L., et al. (2018). Microplastic sampling with the AVANI trawl compared to two neuston trawls in the Bay of Bengal and South Pacific. Environ. Pollut. 232, 430–439. doi: 10.1016/j.envpol.2017.09.058
Eriksen, M., Maximenko, N., Thiel, M., Cummins, A., Lattin, G., Wilson, S., et al. (2013). Plastic pollution in the South Pacific subtropical gyre. Mar. Pollut. Bull. 68, 71–76. doi: 10.1016/j.marpolbul.2012.12.021
Eriksen, M., Thiel, M., and Lebreton, L. (2017). “Nature of plastic marine pollution in the subtropical gyres,” in Hazardous Chemicals Associated with Plastics in the Environment, eds H. Takada, and H. K. Karapanagioti (Heidelberg: Springer Verlag), 1–28.
Fariña, J. M., Aldana, M., Ogalde, F., and Ojeda, F. P. (2000). Trophic ecology of Girella laevifrons (Pisces: Kyphosidae) in rocky intertidal zones of northern Chile affected an non-affected by copper mine tailings. Rev. Chil. Hist. Nat. 73, 139–149. doi: 10.4067/S0716-078X2000000100013
Fariña, J. M., and Ojeda, F. P. (1993). Abundance, activity, and trophic patterns of the redspotted catshark, Schroederichthys chilensis, on the Pacific temperate coast of Chile. Copeia 2, 545–549. doi: 10.2307/1447159
Fernández, N., Muñoz, S., Ardiles, K., Torres, I., Figueroa, R. A., and González-Acuña, D. (2011). “Uso de residuos plásticos en la construcción de nidos por el Lile (Phalacrocorax gaimardi) en el Norte de Chile,” in X Congreso Chileno de Ornitología, (Santiago).
Flammang, B. E., Ebert, D. A., and Cailliet, G. M. (2007). Egg cases of the genus Apristurus (Chondrichthyes: Scyliorhinidae): phylogenetic and ecological implications. Zoology 110, 308–317. doi: 10.1016/j.zool.2007.03.001
Fossi, M. C., Romeo, T., Baini, M., Panti, C., Marsili, L., Campani, T., et al. (2017). Plastic debris occurrence, convergence areas and fin whales feeding ground in the Mediterranean marine protected area Pelagos sanctuary: a modelling approach. Front. Mar. Sci. 4:167. doi: 10.3389/fmars.2017.00167
Galloway, T. S., Cole, M., and Lewis, C. (2017). Interactions of microplastic debris throughout the marine ecosystem. Nat. Ecol. Evol. 1:0116. doi: 10.1038/s41559-017-0116
García-Godos, I., and Goya, E. (2006). Diet of the peruvian diving petrel Pelecanoides garnotii at La Vieja island, Peru, 1997 – 2000: potential fishery interactions and conservation implications. Mar. Ornithol. 34, 33–41.
García-Godos, I., Goya, E., and Jahncke, J. (2002). The diet of Markham's storm petrel Oceanodroma markhami on the central coast of Peru. Mar. Ornithol. 30, 77–83.
García-Godos, I., Van Waerebeek, K., Reyes, J. C., Alfaro-Shigueto, J., and Arias-Schreiber, M. (2007). Prey occurrence in the stomach contents of four small cetacean species in Peru. Lat. J. Aqua. Mamm. 6, 171–183. doi: 10.5597/lajam00122
George-Nascimento, M., Bustamante, R., and Oyarzun, C. (1985). Feeding ecology of the South American sea lion Otaria flavescens: food contents and food selectivity. Mar. Ecol. Prog. Ser. 21, 135–143. doi: 10.3354/meps021135
Guerra-Correa, C., Valenzuela, A., Retamal, L. M., and Malinarich, A. (2007). “Influencia de los desechos plásticos en la sobrevivencia de tortugas: el caso de Chelonia mydas en Antofagasta,” in VII Simposio Sobre Medio Ambiente: Estado Actual y Perspectivas de la Investigación y Conservación de las Tortugas Marinas en las Costas del Pacífico Sur Oriental (Antofagasta), 96.
Guidino, C., Llapapasca, M. A., Silva, S., Alcorta, B., and Pacheco, A. S. (2014). Patterns of spatial and temporal distribution of humpback whales at the southern limit of the Southeast Pacific breeding area. PLoS ONE 9:e112627. doi: 10.1371/journal.pone.0112627
Haro, D., Aguayo-Lobo, A., Blank, O., Cifuentes, C., Dougnac, C., Arredondo, C., et al. (2015). A new mass stranding of false killer whale, Pseudorca crassidens in the Strait of Magellan Chile. Rev. Biol. Mar. Oceanogr. 50, 149–155. doi: 10.4067/S0718-19572015000100013
Haro, D., Riccialdelli, L., Acevedo, J., Aguayo-Lobo, A., and Montiel, A. (2016). Trophic ecology of humpback whales (Megaptera novaeangliae) in the Magellan strait as indicated by carbon and nitrogen stable isotopes. Aquat. Mamm. 42, 233–244. doi: 10.1578/AM.42.2.2016.233
Häussermann, V., Gutstein, C., Bedington, M., Cassis, D., Olavarria, C., Dale, A., et al. (2017). Largest baleen whale mass mortality during strong El Niño event is likely related to harmful toxic algal bloom. PeerJ 5:e3123. doi: 10.7717/peerj.3123
Herling, C., Culik, B., and Hennicke, J. (2005). Diet of the Humboldt penguin (Spheniscus humboldti) in northern and southern Chile. Mar. Biol. 147, 13–25. doi: 10.1007/s00227-004-1547-8
Hernández, S., Lamilla, J., Dupré, E., and Stotz, W. (2005). Desarrollo embrionario de la pintarroja común Schroederichthys chilensis (Guichenot, 1848) (Chondrichthyes: Scyliorhinidae). Gayana 69, 184–190. doi: 10.4067/S0717-65382005000100025
Hidalgo-Ruz, V., Honorato-Zimmer, D., Gatta-Rosemary, M., Nuñez, P., Hinojosa, I. A., and Thiel, M. (2018). Spatio-temporal variation of anthropogenic marine debris on Chilean beaches. Mar. Pollut. Bull. 126, 1516–1524. doi: 10.1016/j.marpolbul.2017.11.014
Hidalgo-Ruz, V., and Thiel, M. (2013). Distribution and abundance of small plastic debris on beaches in the SE Pacific (Chile): a study supported by a citizen science project. Mar. Environ. Res. 87–88, 12–18. doi: 10.1016/j.marenvres.2013.02.015
Hinojosa, I. A., and Thiel, M. (2009). Floating marine debris in fjords, gulfs and channels of southern Chile. Mar. Pollut. Bull. 58, 341–350. doi: 10.1016/j.marpolbul.2008.10.020
IMARPE (2011). Informe Nacional sobre la Conservación de las Tortugas Marinas en el Perú. Callao: Instituto del Mar del Perú.
IUCN (2018). The IUCN Red List of Threatened Species. Available online at: http://www.iucnredlist.org/
Jacobsen, J. K., Massey, L., and Gulland, F. (2010). Fatal ingestion of floating net debris by two sperm whales (Physeter macrocephalus). Mar. Pollut. Bull. 60, 765–767. doi: 10.1016/j.marpolbul.2010.03.008
Jahncke, J., García-Godos, A., and Goya, E. (1997). Dieta del Guanay Leucocarbo bougainvillii, del Piquero Peruano Sula variegata y otras aves de la costa peruana, Abril y Mayo de 1997. Inf. del Inst. del Mar. Perú 126, 75–86.
Jiménez, A., Pingo, S., Alfaro-Shigueto, J., Mangel, J. C., and Hooker, Y. (2017). Feeding ecology of the green turtle Chelonia mydas in northern Peru. Lat. Am. J. Aquat. Res. 45, 585–596. doi: 10.3856/vol45-issue3-fulltext-8
Kenyon, K. W., and Kridler, E. (1969). Laysan albatrosses swallow indigestible matter. The Auk 86, 339–343. doi: 10.2307/4083505
Kiessling, T., Gutow, L., and Thiel, M. (2015). “Marine litter as habitat and dispersal vector,” in Marine Anthropogenic Litter, eds M. Bergmann, L. Gutow, and M. Klages (Cham: Springer), 1–447.
Kiessling, T., Salas, S., Mutafoglu, K., and Thiel, M. (2017). Who cares about dirty beaches? Evaluating environmental awareness and action on coastal litter in Chile. Ocean Coast. Manag. 137, 82–95. doi: 10.1016/j.ocecoaman.2016.11.029
Kühn, S., Rebolledo, E. L. B., and van Franeker, J. A. (2015). “Deleterious effects of litter on marine life,” in Marine Anthropogenic Litter, eds M. Bergmann, L. Gutow, and M. Klages (Cham: Springer), 75–116.
Laist, D. W. (1997). “Impacts of marine debris: entanglement of marine life in marine debris including a comprehensive list of species with entanglement and ingestion records,” in Marine Debris. Springer Series on Environmental Management, eds J. M Coe and D. B. Rogers (New York, NY: Springer), 99–139.
Lavers, J. L., Bond, A. L., and Hutton, I. (2014). Plastic ingestion by flesh-footed shearwaters (Puffinus carneipes): implications for fledgling body condition and the accumulation of plastic-derived chemicals. Environ. Pollut. 187, 124–129. doi: 10.1016/j.envpol.2013.12.020
Law, K. L. (2017). Plastics in the marine environment. Annu. Rev. Mar. Sci. 9, 205–229. doi: 10.1146/annurev-marine-010816-060409
Ludynia, K., Garthe, S., and Luna-Jorquera, G. (2005). Seasonal and regional variation in the diet of the kelp gull in northern Chile. Waterbirds 28, 359–365. doi: 10.1675/1524-4695(2005)028[0359:SARVIT]2.0.CO;2
Luna-Jorquera, G., Fernández, C. E., and Rivadeneira, M. M. (2012). Determinants of the diversity of plants, birds and mammals of coastal islands of the Humboldt current systems: implications for conservation. Biodivers. Conserv. 21, 13–32. doi: 10.1007/s10531-011-0157-2
Lusher, A. (2015). “Microplastics in the marine environment: distribution, interactions and effects,” in Marine Anthropogenic Litter, eds M. Bergmann, L. Gutow, and M. Klages (Cham: Springer), 1–447.
Lusher, A. L., Hernandez-Milian, G., Berrow, S., Rogan, E., and O'Connor, I. (2018). Incidence of marine debris in cetaceans stranded and bycaught in Ireland: recent findings and a review of historical knowledge. Environ. Pollut. 232, 467–476. doi: 10.1016/j.envpol.2017.09.070
Mangel, J. C., Alfaro-Shigueto, J., Witt, M. J., Hodgson, D. J., and Godley, B. J. (2013). Using pingers to reduce bycatch of small cetaceans in Peru's small-scale driftnet fishery. Oryx 47, 595–606. doi: 10.1017/S0030605312000658
Mansilla, L., Olavarría, C., and Vega, M. A. (2012). Stomach contents of long-finned pilot whales (Globicephala melas) from southern Chile. Pol. Biol. 35, 1929–1933. doi: 10.1007/s00300-012-1222-3
Markaida, U., and Sosa-Nishizaki, O. (2010). Food and feeding habits of the blue shark Prionace glauca caught off Ensenada, Baja California, Mexico, with a review on its feeding. J. Mar. Biol. Assoc. UK. 90, 977–994. doi: 10.1017/S0025315409991597
Martinez, E., Maamaatuaiahutapu, K., and Taillandier, V. (2009). Floating marine debris surface drift: convergence and accumulation toward the South Pacific subtropical gyre. Mar. Pollut. Bull. 58, 1347–1355. doi: 10.1016/j.marpolbul.2009.04.022
Medina-Vogel, G., Rodriguez, C. D., Alvarez, R. E., and Bartheld, V. (2004). Feeding ecology of the marine otter (Lutra felina) in a rocky seashore of the south of Chile. Mar. Mamm. Scien. 20, 134–144. doi: 10.1111/j.1748-7692.2004.tb01144.x
Miranda-Urbina, D., Thiel, M., and Luna-Jorquera, G. (2015). Litter and seabirds found across a longitudinal gradient in the South Pacific Ocean. Mar. Pollut. Bull. 96, 235–244. doi: 10.1016/j.marpolbul.2015.05.021
Mizraji, R., Ahrendt, C., Perez-Venegas, D., Vargas, J., Pulgar, J., Aldana, M., et al. (2017). Is the feeding type related with the content of microplastics in intertidal fish gut? Mar. Pollut. Bull. 116, 498–500. doi: 10.1016/j.marpolbul.2017.01.008
Moore, E., Lyday, S., Roletto, J., Litle, K., Parrish, J. K., Nevins, H., et al. (2009). Entanglements of marine mammals and seabirds in central California and the north-west coast of the United States 2001–2005. Mar. Pollut. Bull. 58, 1045–1051. doi: 10.1016/j.marpolbul.2009.02.006
Moreno, C. A., Castro, R., Mújica, L. J., and Reyes, P. (2008). Significant conservation benefits obtained from the use of a new fishing gear in the Chilean Patagonian toothfish fishery. CCAMLR Sci. 15, 79–91.
Nelms, S. E., Duncan, E. M., Broderick, A. C., Galloway, T. S., Godfrey, M. H., Hamann, M., et al. (2016). Plastic and marine turtles: a review and call for research. ICES J. Mar. Sci. 73, 165–181. doi: 10.1093/icesjms/fsv165
Oddone, M. C., and Vooren, C. M. (2002). Egg cases and size at hatching of Sympterygia acuta in the southwestern Atlantic. J. Fish Biol. 61, 858–861. doi: 10.1111/j.1095-8649.2002.tb00919.x
Oddone, M. C., and Vooren, C. M. (2008). Comparative morphology and identification of egg capsules of skate species of the genera Atlantoraja Menni, 1972, Rioraja Whitley, 1939 and Sympterygia Müller & Henle, 1837. Arquivos de Ciências do Mar 41, 5–13.
Ory, N., Chagnon, C., Felix, F., Fernández, C., Ferreira, J. L., Gallardo, C., et al. (2018). Low prevalence of microplastic contamination in planktivorous fish species from the southeast Pacific Ocean. Mar. Pollut. Bull. 127, 211–216. doi: 10.1016/j.marpolbul.2017.12.016
Ory, N. C., Sobral, P., Ferreira, J. L., and Thiel, M. (2017). Amberstripe scad Decapterus muroadsi (Carangidae) fish ingest blue microplastics resembling their copepod prey along the coast of Rapa Nui (Easter Island) in the South Pacific subtropical gyre. Scien. Tot. Environ. 586, 430–437. doi: 10.1016/j.scitotenv.2017.01.175
Page, B., McKenzie, J., McIntosh, R., Baylis, A., Morrissey, A., Calvert, N., et al. (2004). Entanglement of Australian sea lions and New Zealand fur seals in lost fishing gear and other marine debris before and after government and industry attempts to reduce the problem. Mar. Pollut. Bull. 49, 33–42. doi: 10.1016/j.marpolbul.2004.01.006
Perez-Venegas, D., Pavés, H., Pulgar, J., Ahrendt, C., Seguel, M., and Galbán-Malagón, C. J. (2017). Coastal debris survey in a Remote Island of the Chilean Northern Patagonia. Mar. Pollut. Bull. 125, 530–534. doi: 10.1016/j.marpolbul.2017.09.026
Pichel, W. G., Churnside, J. H., Veenstra, T. S., Foley, D. G., Friedman, K. S., Brainard, R. E., et al. (2007). Marine debris collects within the North Pacific subtropical convergence zone. Mar. Pollut. Bull. 54, 1207–1211. doi: 10.1016/j.marpolbul.2007.04.010
Portflitt-Toro, M., Miranda-Urbina, D., and Luna-Jorquera, D. (2016). Captura incidental del blanquillo Podiceps occipitalis en una red de enmalle en Coquimbo, Norte de Chile. Rev. Chil. Ornitol. 22, 212–214.
Rech, S., Macaya-Caquilpán, V., Pantoja, J. F., Rivadeneira, M. M., Jofre Madariaga, D., and Thiel, M. (2014). Rivers as a source of marine litter - a study from the SE Pacific. Mar. Pollut. Bull. 82, 66–75. doi: 10.1016/j.marpolbul.2014.03.019
Rech, S., Macaya-Caquilpán, V., Pantoja, J. F., Rivadeneira, M. M., Campodónico, C. K., and Thiel, M. (2015). Sampling of riverine litter with citizen scientists – findings and recommendations. Environ. Monit. Assess. 187:335. doi: 10.1007/s10661-015-4473-y
Roman, L., Schuyler, Q. A., Hardesty, B. D., and Townsend, K. A. (2016). Anthropogenic debris ingestion by avifauna in Eastern Australia. PLoS ONE 11:e0158343. doi: 10.1371/journal.pone.0158343
Ryan, P. G. (1987). The incidence and characteristics of plastic particles ingested by seabirds. Mar. Environ. Res. 23, 175–206. doi: 10.1016/0141-1136(87)90028-6
Ryan, P. G. (2016) “Ingestion of Plastics by Marine Organisms,” in Hazardous Chemicals Associated with Plastics in the Marine Environment. Handbook of Environmental Chemistry, eds H. Takada, and H. K. Karapanagioti (Berlin: Springer), 1–32.
Sazima, I., Gadig, O. B., Namora, R. C., and Motta, F. S. (2002). Plastic debris collars on juvenile carcharhinid sharks (Rhizoprionodon lalandii) in southwest Atlantic. Mar. Pollut. Bull. 44, 1149–1151. doi: 10.1016/S0025-326X(02)00141-8
Scales, K. L., Miller, P. I., Hawkes, L. A., Ingram, S. N., Sims, D. W., and Votier, S. C. (2014). On the Front Line: frontal zones as priority at-sea conservation areas for mobile marine vertebrates. J. Appl. Ecol. 51, 1575–1583. doi: 10.1111/1365-2664.12330
Schuyler, Q., Hardesty, B. D., Wilcox, C., and Townsend, K. (2014). Global analysis of anthropogenic debris ingestion by sea turtles. Conserv. Biol. 28, 129–139. doi: 10.1111/cobi.12126
Silva, A., Retamal, L. M., and Guerra-Correa, C. (2007). “Registro de tortugas marinas ingresadas al centro de rescate y rehabilitación de fauna silvestre,” in VII Simposio sobre Medio Ambiente: Estado Actual y Perspectivas de la Investigación y Conservación de las Tortugas Marinas en las Costas del Pacífico Sur Oriental (Antofagasta), 96.
Spear, L., Ainley, D., and Ribic, C. (1995). Incidence of plastic in seabirds from the Tropical Pacific 1984-91: relation with distribution of species, sex, age, season, year and body weight. Mar. Environ. Res. 40, 123–146. doi: 10.1016/0141-1136(94)00140-K
Stucchi, M., and Figueroa, J. (2006). La avifauna de las islas Lobos de Afuera y algunos alcances sobre su biodiversidad. Reporte de investigación N° 2, p.88, Asociación Ucumari, Lima, Perú.
Terepocki, A. K., Brush, A. T., Kleine, L. U., Shugart, G. W., and Hodum, P. (2017). Size and dynamics of microplastic in gastrointestinal tracts of northern fulmars (Fulmarus glacialis) and sooty shearwaters (Ardenna grisea). Mar. Poll. Bull. 116, 143–150. doi: 10.1016/j.marpolbul.2016.12.064
Thiel, M., Bravo, M., Hinojosa, I. A., Luna, G., Miranda, L., Núñez, P., et al. (2011). Anthropogenic litter in the SE Pacific: an overview of the problem and possible solutions. J. Integr. Coast. Zone Manage. 11, 115–134. doi: 10.5894/rgci207
Thiel, M., Hinojosa, I. A., Miranda, L., Pantoja, J., Rivadeneira, M. M., and Vasquez, N. (2013). Anthropogenic marine debris in the coastal environment: a multi-year comparison between coastal waters and local beaches. Mar. Pollut. Bull. 71, 307–316. doi: 10.1016/j.marpolbul.2013.01.005
Thiel, M., Hinojosa, I., Vásquez, N., and Macaya, E. (2003). Floating marine debris in coastal waters of the SE-Pacific (Chile). Mar. Pollut. Bull. 46, 224–231. doi: 10.1016/S0025-326X(02)00365-X
Thiel, M., Macaya, E. C., Acuña, E., Arntz, W. E., Bastias, H., Brokordt, K., et al. (2007). The Humboldt Current System of northern and central Chile: oceanographic processes, ecological interactions and socioeconomic feedback. Ocean. Mar. Biol. Annu. Rev. 45, 195–344. doi: 10.1201/9781420050943.ch6
Titmus, A. J., and Hyrenbach, K. D. (2011). Habitat associations of floating debris and marine birds in the North East Pacific Ocean at coarse and meso-spatial scales. Mar. Pollut. Bull. 62, 2496–2506. doi: 10.1016/j.marpolbul.2011.08.007
Wallace, B. P., Lewison, R. L., McDonald, S. L., McDonald, R. K., Kot, C. Y., Kelez, S., et al. (2010). Global patterns of marine turtle bycatch. Conserv. Lett. 3, 131–142. doi: 10.1111/j.1755-263X.2010.00105.x
Wegner, N. C., and Cartamil, D. P. (2012). Effects of prolonged entanglement in discarded fishing gear with substantive biofouling on the health and behavior of an adult shortfin mako shark, Isurus oxyrinchus. Mar. Pollut. Bull. 64, 391–394. doi: 10.1016/j.marpolbul.2011
Wilcox, C., Hardesty, B. D., Sharples, R., Griffin, D. A., Lawson, T. J., and Gunn, R. (2013). Ghostnet impacts on globally threatened turtles, a spatial risk analysis for northern Australia. Conserv. Lett. 6, 247–254. doi: 10.1111/conl.12001
Keywords: anthropogenic marine debris, impacts, biota-litter interactions, entanglement, microplastic ingestion
Citation: Thiel M, Luna-Jorquera G, Álvarez-Varas R, Gallardo C, Hinojosa IA, Luna N, Miranda-Urbina D, Morales N, Ory N, Pacheco AS, Portflitt-Toro M and Zavalaga C (2018) Impacts of Marine Plastic Pollution From Continental Coasts to Subtropical Gyres—Fish, Seabirds, and Other Vertebrates in the SE Pacific. Front. Mar. Sci. 5:238. doi: 10.3389/fmars.2018.00238
Received: 01 February 2018; Accepted: 20 June 2018;
Published: 24 July 2018.
Edited by:
Francois Galgani, Institut Français de Recherche pour l'Exploitation de la Mer (IFREMER), FranceReviewed by:
André Ricardo Araújo Lima, Universidade Federal de Pernambuco, BrazilHans Uwe Dahms, Kaohsiung Medical University, Taiwan
Copyright © 2018 Thiel, Luna-Jorquera, Álvarez-Varas, Gallardo, Hinojosa, Luna, Miranda-Urbina, Morales, Ory, Pacheco, Portflitt-Toro and Zavalaga. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Martin Thiel, dGhpZWxAdWNuLmNs