 Rianna Burnham
Rianna Burnham David Duffus1
David Duffus1 Xavier Mouy
Xavier Mouy- 1Whale Research Lab, Department of Geography, University of Victoria, Victoria, BC, Canada
- 2JASCO Applied Sciences, Victoria, BC, Canada
Gray whale (Eschrichtius robustus) vocalizing behavior varies over its range, both in call type and frequency. This study adds to the growing body of passive acoustic research outside of the calving/breeding lagoons, and moves toward a description of calling behaviors throughout the whales' range. Data are presented here from acoustic surveys for two northward and one southward migration, with recordings taken off the west coast of Vancouver Island. We found gray whales to be highly sonorous, with extensive calling during the study periods of February to May for northward migrations and September to the end of January for southward travel. Low frequency moans were the most prevalent call type, with others, including knocks, up- and down sweeps, and rumbles, recorded in varying numbers. We hypothesize that calling is an aid for navigation and orientation of the herd along the migration route, in addition to holding more social functions usually assigned to baleen whale calling.
Introduction
For many whale species, calls have been correlated with either a behavioral, social or geographic context, with changes in call repertoire and call rate over a species range. Whereas, vocalizing behaviors of gray whales (Eschrichtius robustus) have been well-documented in their calving/breeding lagoons (Dahlheim et al., 1984; Dahlheim, 1987; Ollervides, 2001; Charles, 2011; López-Urbán et al., 2016), studies have only recently added to the body of knowledge from the rest of their range (see for example Moore and Ljungblad, 1984; Stafford et al., 2007; Youngson and Darling, 2016; Guazzo et al., 2017; Rannankari et al., 2018). The use of bottom-mounted passive acoustic monitoring (PAM) systems allows long duration recordings with sensitivity into the lowest frequencies, capturing their full vocal range. These acoustic survey techniques are insensitive to sea state, weather, or light conditions, and data collection that was previously difficult or impossible, particularly during migration periods, is now feasible. These data document a highly acoustic whale, with extensive calling the norm, rather than scarce, as noted in previous vessel-based studies (Rasmussen and Head, 1965; Cummings et al., 1968; Fish et al., 1974; Crane and Lashkari, 1996; Wisdom et al., 2001).
Research from breeding lagoons describes six primary call types in the frequency range of 20 Hz to 2 kHz: class 1, a series of metallic pulses or knocks, sometimes called bongo or conga calls; class 2, a longer duration, single, up or down sweeping metallic pulse; class 3, low frequency modulated moan-like pulse, in some works described as a “growl”; class 4, higher frequency modulated pulses forming grunt or rumble like calls; class 5, higher frequency “bubble blasts,” and class 6, extended sub-surface exhalations (Dahlheim et al., 1984; Dahlheim, 1987). Additional call types have been described, including a potential “motherese” used specifically by cow-calf pairs (Ollervides, 2001; Charles, 2011; López-Urbán et al., 2016), “clicks” or “pulses” sometimes suggesting echolocation potential (Asa-Dorian and Perkins, 1967; Fish et al., 1974), “rasping,” “grunts,” “chirps,” “pop,” “croak” or “bongs,” and vocalizations where more than one call type is super-positioned, for example “knock-moans” or “knock-grunts” (Eberhardt and Evans, 1962; Painter, 1963; Wenz, 1964; Gales, 1966; Hubbs, 1966; Poulter, 1968; Fish et al., 1974; Dahlheim, 1987; Wisdom et al., 2001).
On the northward migration, previous studies have found class 3 moans most frequent, in contrast to the class 1 knocking calls that dominate in breeding areas. Moans comprise up to 87% of the total calls recorded during migration (Cummings et al., 1968; Dahlheim, 1987; Crane and Lashkari, 1996; Guazzo et al., 2017), and are presumed to aid long distance herd cohesion, with broader spectrum modulated calls like class 1 and 4 used for sub- or within-group communication. At a time of negative energy balance, such as, northward migration, we assume acoustic behaviors would be limited as a conservation measure, but recent recordings taken in California show that not to be the case (Guazzo et al., 2017). Limited data is available for southward migratory behaviors, where calling behaviors may reflect an altered physiological state compared to northward travel.
Here we aim to add to the body of knowledge of gray whale acoustic behavior with long duration recordings taken during two northward and one southward migration by deploying a PAM device in the migration corridor. We look to variation in call type and rate for temporal patterning, particularly diurnal, and within- and between seasons, to refine hypotheses about potential call function.
Methods
An Autonomous Multichannel Acoustic Recorder (AMAR, JASCO G3A) fitted with a GeoSpectrum M8E-132 calibrated omnidirectional hydrophone (sensitivity −165 dB re 1 V/μPa, effective 5 Hz−150 kHz, gain of 6 dB) was deployed at a location along the gray whale migration route approximately 5 nm/9.26 km southwest of Siwash Point, Flores Island, on the west coast of Vancouver Island (49°12′37″ N, −126°14′48″ W) in 51 m of water. The AMAR recorded continuously in the low frequency ranges 0–8,000 Hz throughout each of the deployments, the duty cycle extended into the recording range into the higher frequencies, though this is not relevant to this study. The location was chosen using previous knowledge of the migration route in this area (Duffus, Pers. Obs., 1984–2017), as well as periodic vessel-based observations prior to, and during the deployment. The migration corridor is estimated to be within 8 km of the shore for breeding and non-breeding adults and juveniles, and within 5 km for cow-calf pairs, with some observations as close as 200–400 m from shore (Poole, 1984; Perryman et al., 2002; DeAngelis et al., 2010). The southward migration is similar, but with some observations of whales traveling up to 40 km offshore (Green et al., 1995; Shelden and Laake, 2002; Figure 1).
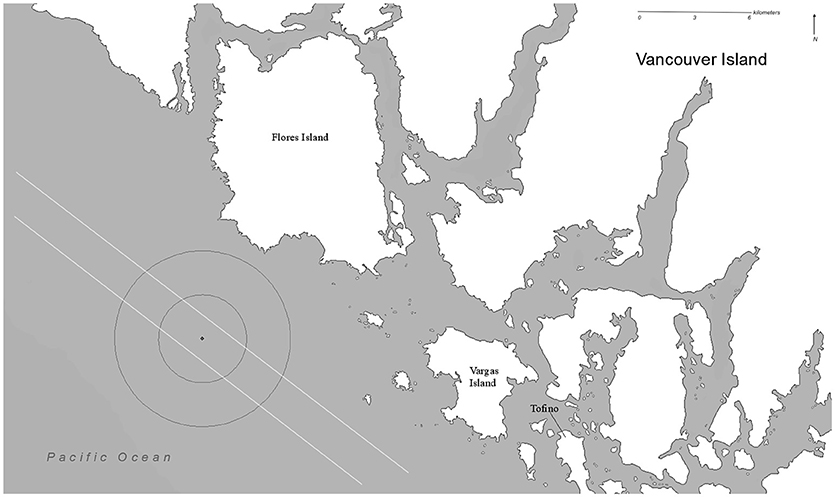
Figure 1. Location of AMAR deployment, with migratory corridor and likely range detection radii of gray whale class 3 moan calls. AMAR location is in the center of the detection circles with the smaller circle representing the range of detection 80% of the time (3 km) and the larger 10% of the time (6 km). The lines parallel to the coast line are 8 km, to indicate the migratory path of Phase A whales, and 5 km, for the cow-calves in Phase B.
The PAM system was deployed between February 21 and April 25, 2015 (N1) and March 7 to May 5, 2016 (N2) for the northward migration, and September 27, 2016 to January 25, 2017 (S1) for southward migration. Recordings were made constantly, covering the peak migration sightings off the west coast of Vancouver Island (Duffus, Pers. Obs, 1984–2017) and correlated to shore counts from California counting and PAM survey stations (ACS Sighting data, Guazzo et al., 2017).
Manual identification of calls was made through visual and aural examination of sound files and respective spectrograms using Raven Pro Interactive Sound Analysis Software. For the 2015 northward migration (N1) the data was split into 900 s segments, and the first 340 s of every segment was manually examined. A random selection of 200 of the 900 s segments was fully analyzed. For all deployments a minimum of every fifth day of the recording period was manually inspected, accounting for a total of 56.6% for N1, 23% for N2 and 20.7% for S1. Spectrograms were generated using a 256-point Hanning-window FFT with 50% overlap. Comparisons were made to the core call types, described by Dahlheim (1987) and subsequent others (Crane and Lashkari, 1996; Ollervides, 2001; Wisdom et al., 2001; Stafford et al., 2007; Charles, 2011; López-Urbán et al., 2016; Youngson and Darling, 2016) where appropriate. Calls were categorized as class 1–4, as described by Dahlheim (1987), possible “motherese,” or “other” if they matched the descriptions from other studies or were still believed to be gray whale sounds due to frequency (Hz) and received volume of sound relative to ambient conditions. We modified class 1 to indicate frequency modulation (1a) or not (1b), and class 2 to distinguish between upsweeps (2a) and downsweeps (2b). Sub-surface exhalation sounds for class 5 and 6 (Dahlheim, 1987) were noted, but used here only to indicate whale presence, and were not subject to further analysis. In addition, the full recordings were subject to an automated detection system (Mahoney et al., 2014; Mouy et al., 2015), to compare with the manual analysis. An estimated number of calls for the deployment was calculated based on the number of calls identified in the percentage of each deployment that was manually inspected, and extrapolating to the full deployment [(Number of calls identified/Amount of time verified) × Total recording time]. This extrapolation presumed in this case that calling was at a consistent rate with no diurnal or seasonal patterning to vocal behaviors, and did not discern the number of callers at any time. Also a correction of call number, using a comparison of the results of the automated detector and manual verification was calculated to establish a level of detection accuracy.
For each of the calls, a series of measurements were made. Calls were described by their duration (s), lower and upper frequency (Hz) extents, the frequency range (Hz) of the call harmonics, and peak frequency (Hz), where the acoustic energy of the call is greatest. Comments on call qualities were also noted, to allow for later refinement of call categories. Any calls where background noise, particularly vessel noise, masked the accurate measurement of these metrics were excluded from further analysis, as were those calls labeled “other.”
The appropriateness of the separate call categories, and subsequent subdivisions, was analyzed for each call class and sub-class comparing call metrics of frequency extents, range, peak, and duration of all calls recorded during the migration periods.
An estimate of the range and probability of call detection was made using ambient noise levels taken at each minute of the recording, source levels for moan calls as reported by Guazzo et al. (2017, 156.9 ± 11.4 dB re 1 μPa @ 1 m), and an approximation of transmission loss using the spherical spreading law (Urick, 1983). Given the low frequency nature of gray whale calls found in this and other works, attenuation was not included in the transmission loss estimation (see Supplementary Material).
External factors that may affect calling behaviors were also explored, including photoperiod, tidal cycle, ambient noise and presence of vessel noise. Also, changes in call rate and call type was explored as the season progresses. For photoperiod, a day-night comparison was used, where day was defined as 07.00–19.00, and night 19.00–07.00. To include periods of twilight this definition was adjusted so that daylight represented the hours of 09.00–17.00, night 21.00–05.00, dawn 05.00–09.00 and dusk 17.00–21.00. Tidal effects were examined by correlating to tide levels, as well as grouping calls at either low or high slack tide, which was defined as 2 h before and after the turning of the tide, and ebb and flood periods between slacks. Ambient levels were estimated by summing the 1/3 octave bands centered around 1–2,000 Hz, to capture the full frequency range of calls. Ambient condition was correlated on a minute-by-minute scale to each call identified, or converted to an hourly average to examine calling rate in changing background noise conditions. Finally, the calling rate and use of each call type was considered as the survey period progressed by correlating call number with the number of days elapsed since January 1 of that year.
Results
A total of 13,749 calls were analyzed from the northward migration from 2015 to 2016, and 3,691 were analyzed for the southward migration. The appropriateness in combining the two northward deployments was tested and deployments were not found to be significantly different. The number of whales cannot be discerned from these numbers, as call number and number of callers may not be linear (Ponce et al., 2012). All core call types were present for all deployments (Figures 2–5). Also see Supplementary Materials for example spectrograms of each call type in more detail and example audio files), but in all cases, class 3 moan calls were dominant, 83.69% for northward and 81.67% for southward travel. The number of calls identified by the auto-detector, corrected by the manually verified data is also given as a comparator for each deployment (Table 3). Number of calls, call duration, frequency (Hz) extents, and peak were compared for northward and southward movement for this study as well as to previous studies (Tables 1,2). The number of calls expected for the full deployment was extrapolated using the rate of calls per hour of the manually inspected data, under the assumption that the rate of calling was consistent in all time periods throughout the deployment (“calls/rate” in Table 3). Using a comparison of the auto-detector results and manually inspected data, estimates of a corrected call number were possible (“calls corrected” in Table 3). We compared the proportion of false positive and false negatives between the detector to the verified data (0.52–4.88% and 14.95–26.55%, respectively), and analyzed whether the number of calls correctly identified were accurate, or an over- or underestimate (50.96–60.55% and 4.16–41.12%, respectively).

Figure 2. Spectrogram of class 1 calls set in full frequency range of calls (0–2,000 Hz). (Left) 1a, frequency modulated calls; (Right) 1b, unmodulated calls. Spectrogram was generated using a 256-point Hanning-window FFT with 50% overlap. For more call focused spectrograms and audio file of these calls see Supplementary Materials.
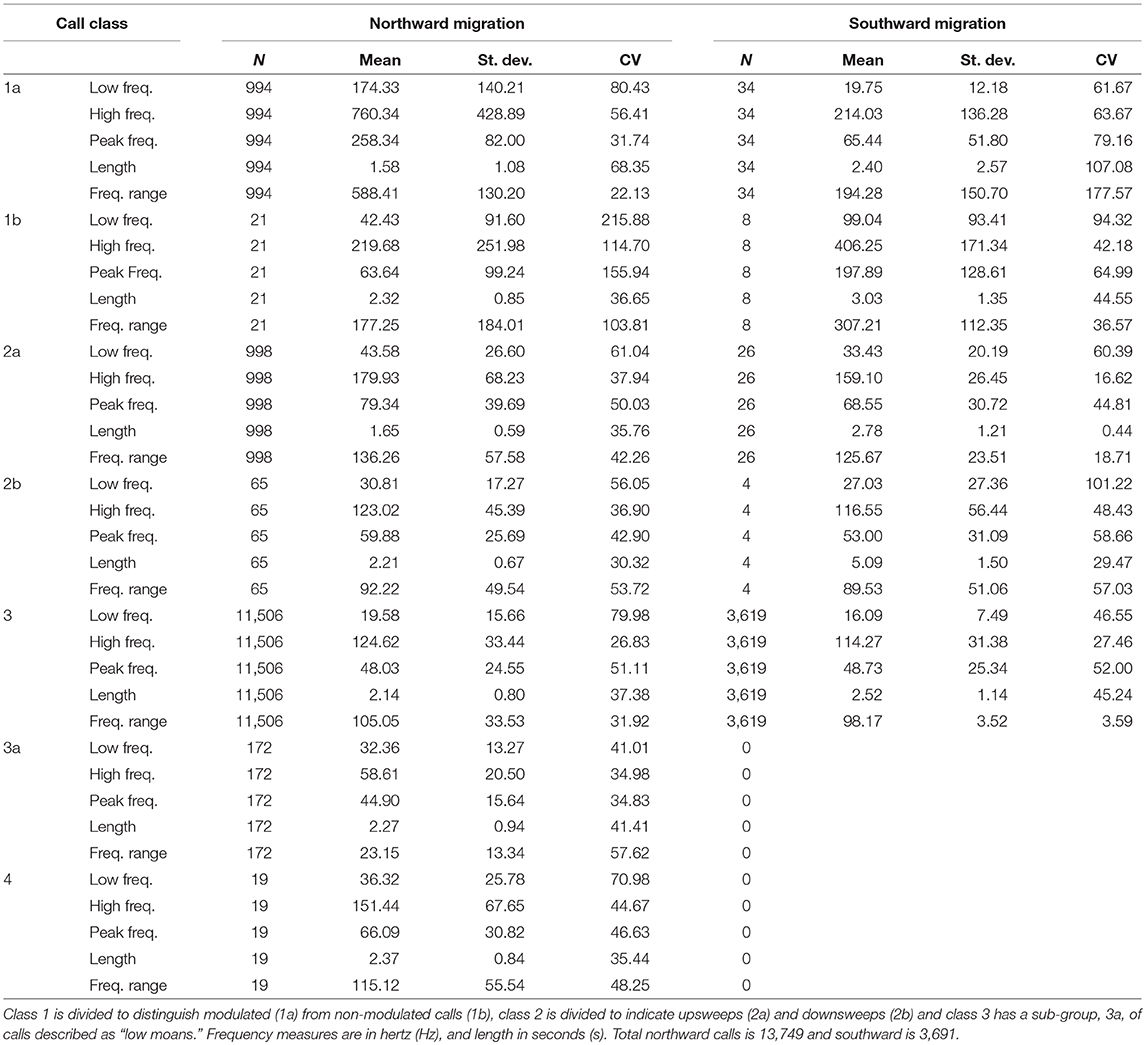
Table 1. Number of calls (N) and mean, standard deviation (St. Dev.) and coefficient of variation (CV) for each call metric by call type.
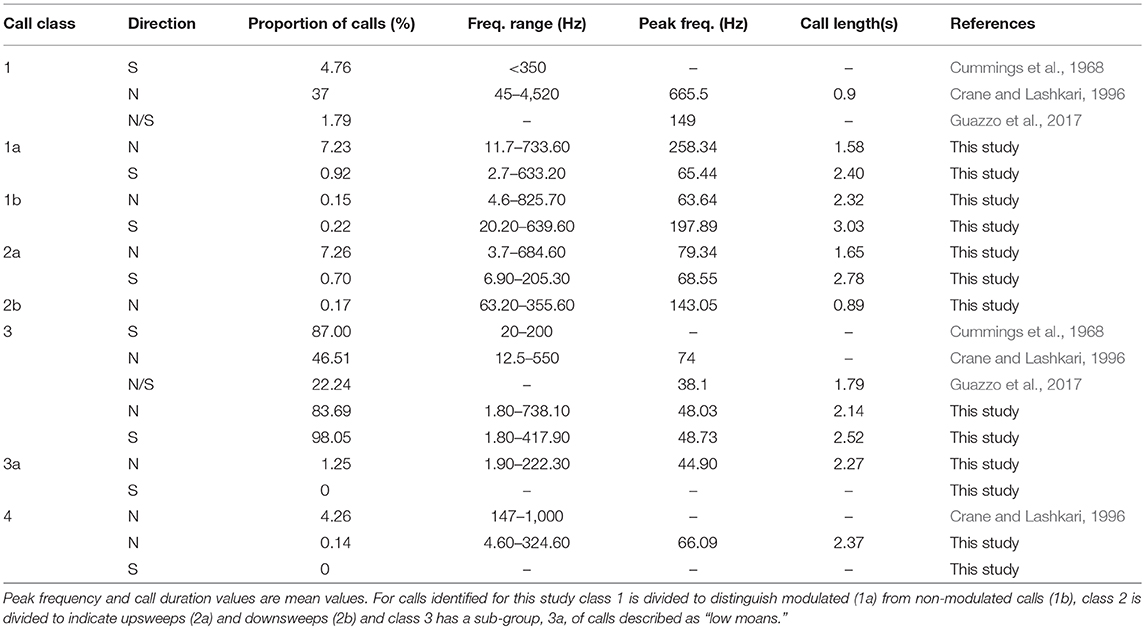
Table 2. Call proportion and descriptors for core call types for this and previous PAM of gray whale calls during migrating periods.
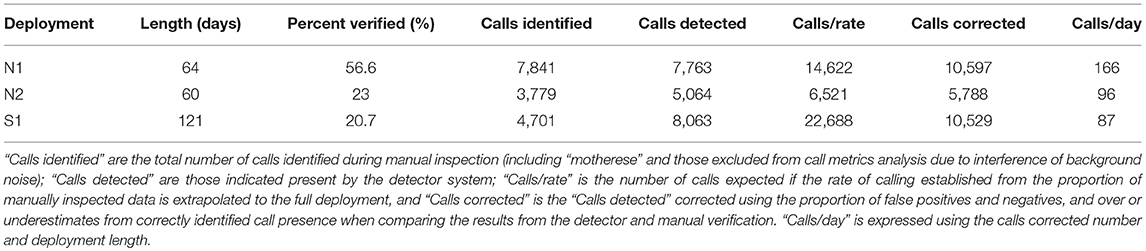
Table 3. Call numbers from manual verification and auto-detector.
The distinction of call classes was tested by comparing the means for each call metric by class (T-test, Tables 1–5 in Supplementary Materials). Overall, significant differences in composition of call types were found for calls employed during migration. A pair-wise comparison of migration calls (both north and southward) found significant differences between call classes consistently, with class 4 the most like other call types (T-test, Tables 1–5 in Supplementary Materials). In particular the appropriateness of sub-division was explored with calls 1a and 1b (Figure 2), 2a and 2b (Figure 3), and 3 and 3a (Figure 4), all being significantly different in all call metrics. Subdivision on class 1 calls was based on presence of frequency modulation of call, for class 2 it was whether the call swept up or down in frequency, and for class 3 the category of “low moan” (class 3a) was used for moan calls focused in the lower frequencies and did not show call harmonics above the fundamental frequency (Figure 4; Tables 1–5 in Supplementary Materials).
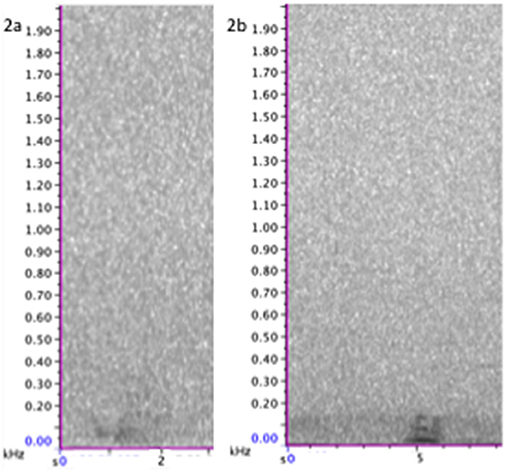
Figure 3. Spectrogram of class 2 calls set in full frequency range of calls (0–2,000 Hz). (Left) 2a, upsweeping call; (Right) 2b, downsweeping call. Spectrogram was generated using a 256-point Hanning-window FFT with 50% overlap. For more call focused spectrograms and audio file of these calls see Supplementary Materials.
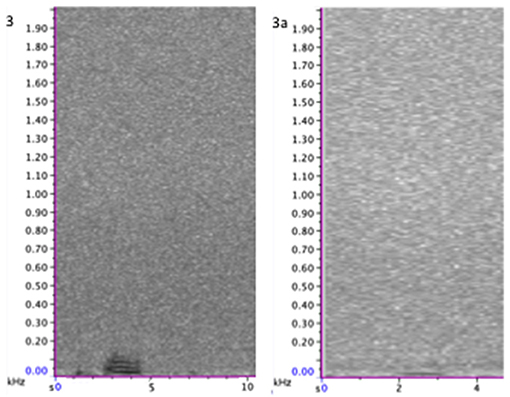
Figure 4. Spectrogram of class 3 calls set in full frequency range of calls (0–2,000 Hz). (Left) 3, moan call; (Right) 3a, low moan, with no harmonics. Spectrogram was generated using a 256-point Hanning-window FFT with 50% overlap. For more call focused spectrograms and audio file of these calls see Supplementary Materials.
Calls identified as possible “motherese,” first described in breeding lagoons by Ollervides (2001) and Charles (2011), were noted for both northward and southward migrations, but comprised only 0.28% and 0.13% of calls, respectively. Therefore, they are not included in the more detailed call analysis. Class 5 and 6 exhalations were noted, with increased prevalence in the latter part of northward migrations, with the opposite true for southward recordings.
There were proportionately more calls at night than during the day, and at night compared to both day and twilight periods (dusk and dawn; Table 4). Call rate differences between day and night periods, or day-night and twilight periods was tested (Table 4). For northward migration, there is a significant difference when all call types are pooled (Mann-Whitney U, p > 0.000), and between day-night calling behaviors in class 1 knock calls (Mann-Whitney U, class 1a p = 0.048; class 1b p = 0.012) and class 3 moans (Mann-Whitney U, p = 0.002). For southward migration the trend is not statistically significant when comparing day and night call behaviors, although the use of upsweeps is significantly different (Mann-Whitney U, p = 0.521, Table 4). Light conditions seem to have a significant influence on calling for southward travel when considering periods of twilight, particularly for class 3 moan calls (Kruskall-Wallis, p < 0.001). When considered by call type and travel direction, the mean number of calls per hour for class 3 calls are the only call type to differ significantly on northward migrations when comparing photoperiods (Welches t-test: t(734.008) = −3.495, p = 0.001, Table 4) and class 1 calls (both Student t-test: 1a, t(613) = −0.103, p = 0.918; 1b, t(613) = 0.181, p = 0.856, Table 4) were not significantly different between day and night periods in their rate of use (mean number of calls/hour) on the southward migration, with all other call classes showing significant differences in response to photoperiod (T-test, Table 4).

Table 4. Mean number of calls per hour across different light conditions.
A similar analysis of call rate correlated to tide cycle was performed, grouping calls to those at either low or high slack tide, which was 2 h before and after the turning of the tide, and ebb and flood periods between slacks. This showed no significant results for any call class in any deployment. Therefore, tides are deemed ineffective to calling in this setting.
Correlating call rate and metrics with ambient noise produced a negative correlation between frequency extent, frequency range, peak frequency and duration with increased ambient noise (Table 5). However, the rate of employment of each call type was not significantly altered in the presence of vessels. Call parameters for classes 2a, 3a, and 4 were correlated to changes in ambient noise levels significantly to the p = 0.05 level if not p < 0.001 for northward migration (Spearman's rho, Table 5), with no significant correlation consistently seen for any call type on the southward migration (Spearman's rho, Table 5). Most of the correlations are weak, only a few are moderately strong. Rises in ambient noise levels are coupled with lower frequency calls, but the relationship again is not strong (Table 5).
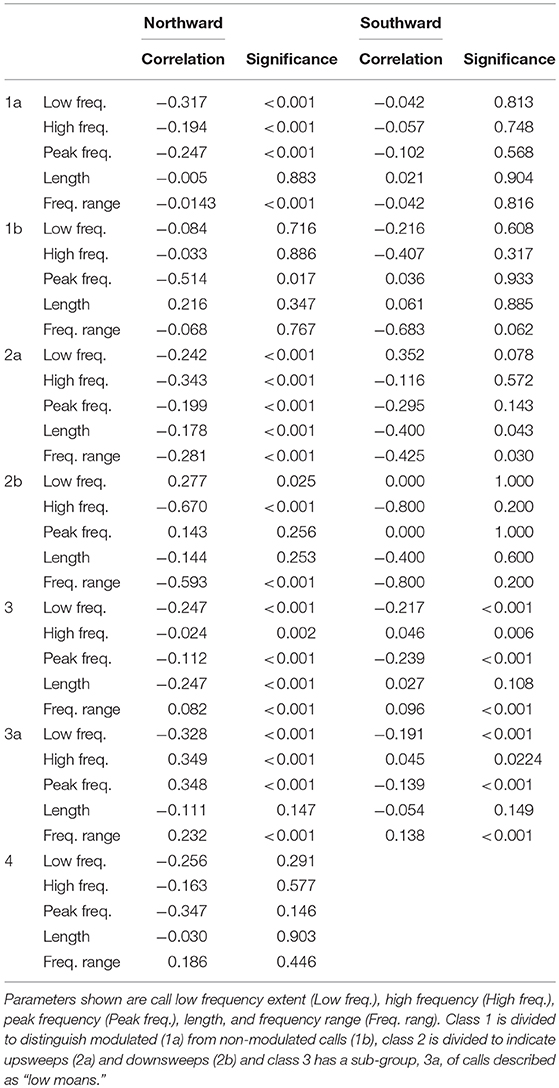
Table 5. Correlation, using Spearman's rho, between ambient condition and call metric for all call types during northward and southward migration.
In general, all call types showed a negative correlation with the seasonal progression of time, with the four core call types significant at the p = 0.001 level (Spearman's rho, Table 6). For northward migration classes all core call types show significant negative correlation with time, except upsweeps (class 2a, rs = 0.182, p < 0.000) and low moans (class 3a, rs = 0.068, p = 0.661). For whales migrating south, only moan call use is significantly correlated with time, and negatively (rs = −0.210, p < 0.000, Table 6).
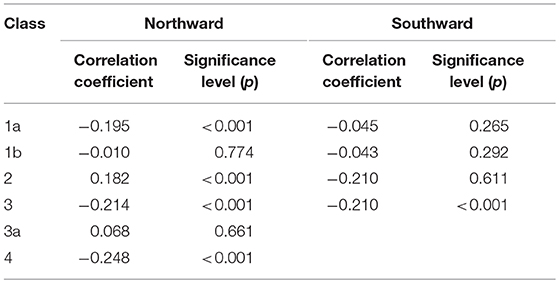
Table 6. Correlation coefficients and significance of Spearmans correlation between calling rate (calls/h) and year day (number of days elapsed since January 1), by call type and swimming direction.
The estimated detection probability of moan calls, the most prevalent call, by the recorder is shown graphically (see Figure 5, Supplementary Materials) and spatially in relation to the hydrophone deployment site (Figure 1). Moans with the highest source levels are estimated to be detectable up to 3 km from the recorder 80% of the time and up to 6 km, 10% of the time, encompassing the migration corridor and coastal and offshore waters that may be part of it. Recordings are unlikely to come from coastal feeding regions.
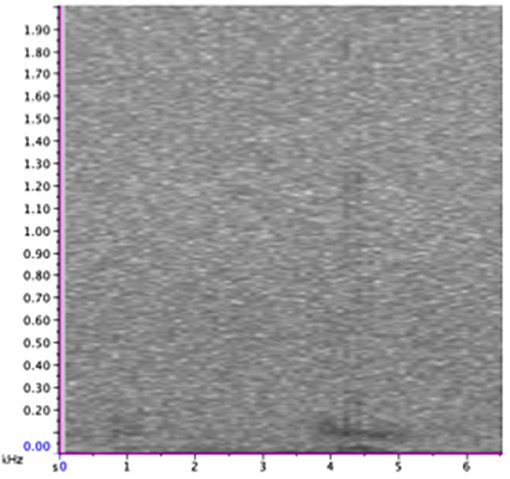
Figure 5. Spectrogram of class 4 call set in full frequency range of calls (0–2,000 Hz). Spectrogram was generated using a 256-point Hanning-window FFT with 50% overlap. For more call focused spectrograms and audio file of these calls see Supplementary Materials.
Discussion
These recordings are from a relatively uniform and stereotyped behavior state dominated by linear, constant swimming. Migration is a continuous, protracted trail of whales, whose movements are consistent in direction and average speed (Perryman et al., 2002; Mate and Urbán-Ramirez, 2003; Guazzo et al., 2017). The timing and location of the deployments, and likely detection range of the recorder, largely excludes vocal behaviors associated with foraging and weaning (see Burnham, 2015). Although courtship and mating behaviors have been noted on the southbound migration (Gilmore, 1960) and observed during northward migrations in Clayoquot Sound (Burnham/Duffus, Pers. Obs., 2015), these are thought to be minor components of the whale's behavior.
Gray whales call frequently during migration, which is in agreement with a recent study by Guazzo et al. (2017) on northward migrating whales in Californian waters, and demonstrate greater vocal activity than that suggested by earlier work (Rasmussen and Head, 1965; Dahlheim, 1987; Crane and Lashkari, 1996). The average rate of calling concurs with recent reports for migration (Guazzo et al., 2017), and bottom mounted hydrophone recordings made in the breeding lagoons (5,982 calls from 3 weeks, López-Urbán et al., 2016) and compares consistently to the corrected auto-detection data (Table 3).
The metrics of call duration and harmonics gives a rough approximation of the shape of the call and acoustic energy projected (Figures 2–5, Supplementary Materials for spectrograms). The calls identified in this study are in accordance with other PAM studies of gray whale migration. The calls are, however, typically longer and, in some cases, lower in frequency than those previously reported (Tables 1,2), suggesting a modification based on geographical location or perhaps water propagation properties. Comparison of call types over time showed a general decrease in frequencies employed this study and previous (Rasmussen and Head, 1965; Dahlheim, 1987; Crane and Lashkari, 1996), and even between 2015 and 2016 for the northward migration, with moan calls consistently employing the lowest frequencies in frequency extents and peak frequency. This may represent a more general response to changes in soundscape, with blue (Balaenoptera musculus, McDonald et al., 2006) and fin whales (Balaenoptera physalus, Bradbury and Vehrencamp, 1998) also noted to modify call structure over time to overcome anthropogenic additions to the ambient condition. The employment of “low moans” (Figure 4) by gray whales may also be part of the adaptation.
We employ a finer sub-division of classes than previous works, and present the description of a “low moan” call type, referred to herein as class 3a (Figure 4, also see Figures 3, 6,7 in Supplementary Materials). This study considers the use of all core call classes outlined by Dahlheim (1987), not seen in other previous studies, with manual inspection of the data allowing for a more nuanced consideration of vocal behaviors including these types of call subdivision, also seen in classes 1 and 2 (Figures 2,3; Figures 1,2 in Supplementary Materials). The “low moan” calls show similarity to the fundamental frequency of a moan call (Figure 4), but do not contain call harmonics in higher frequencies. A crude comparison of the received amplitude of calls supports the subdivision of moan and low-moan, rather than the call being a result of information and harmonics loss in transmission and propagation. The low moans may represent, for example, variation in the size or state of the individual animal calling compared to moan calls, or an altered application of moan calls, discussed in more detail as part of another analysis (Burnham, in review).
Class 3 moan calls dominate migration vocalizing. In each deployment, both north and southward, moan calls exceeded 80% of all calls manually verified. The rate of calling for northward migration was almost double that of southward, from manually verified data (NM = 11.62 calls/h, SM = 6.11 calls/h), and in the corrected auto-detector calling rate per day (Table 3). This rate of use of moans during migration is similar to other studies and contrasts to calling patterns noted for breeding areas (Dahlheim, 1987; Crane and Lashkari, 1996). Also, moans show much less variation than the other call types (Table 1). A low-centered peak frequency and highly controlled call duration and frequency range, as well as the prominence of this call during migration, suggests this call type is linked to traveling behaviors. Moan calls may be a means to maintain herd cohesion as the whales migrate over larger spaces, whereas other call classes such as, class 1 knocks, with their higher peak frequency and variability, are for within-group communication or possibly courtship behaviors (see Crane and Lashkari, 1996; Youngson and Darling, 2016). Deployments for both north and southward migration were timed to capture the peak of the migrations past the recording site, substantiated by shore counts and data from the timing of whales entering and leaving the breeding lagoons (see LSIESP., 2014, 2015; ACS Sighting data; Guazzo et al., 2017). The discrepancy between call rates for north vs. south movement (Tables 1,3) suggests then that vocalizations may be a more necessary component of northward migration, with the number of whales, and so potential callers, passing the recorder during migration periods presumed to be similar for north- and southward travel. Call number per day, particularly the number of moan calls per day, was found to be a good representation of the relative number of individuals passing the recorder, using shore counts as comparison to the acoustic data. Therefore, the progression and peaks of migration are able to be discerned from the number of calls detected but not the number of whales (see Burnham, in review). Calling rate may be expected to be greater for southward migrations, as whales have restored their energy reserves and may also employ social calling for mate attraction, but this was not found. The function of calls, especially moans, may be nuanced to identify topographical/bathymetric features for navigation and a means to orientate less experienced whales to the migration route, as well as productive feeding areas or safe weaning sites as part of an overall cultural transmission of information (Calambokidis et al., 2010; Lang et al., 2011; Scordino et al., 2011). Our recording site is directly offshore from a heavily used summer foraging site and cow-calf requiem. Examination of potential modifications to calling behaviors in reference to this, from whales traversing waters in or adjacent to feeding and weaning sites, is outside the scope of this study and would require additional test sites.
Variation in calling, both in the rate and the composition of calls, may be traits specific to individual whale behavior. The inherent rate of calling by an individual in the context of its social or behavioral setting is not known for gray whales, however we can look at general patterns throughout a population using this migration data. There is variability in calling rate, call type, and within call classes, in the frequency extents, peak frequency and duration, throughout all deployments. Delivery of a call may be varied to amalgamate information on the caller identity and their physiological or emotional state, social or behavioral context of the call, as well as the message for the intended receiver. Changes in call features in relation to size of individuals signaling is not considered here. Also distance from hydrophone of the signaling whale, and how that effects the received call formant, is unknown. Calls, by type, are treated in this analysis equally whether they were received from Phase A or B on northward migration, or during the day or night which may have altered propagation distances (Figure 1; Poole, 1984; Perryman et al., 1999, 2002; DeAngelis et al., 2010). However, the call classes described here are an average of thousands of calls made by numerous individuals, possibly fulfilling multiple functions.
A preliminary analysis to tie the variation to an environmental context found calling to be greater at night than during the day, as suggested by Guazzo et al. (2017) and Rannankari et al. (2018). This suggests that visual cues may supplement acoustics during the day, particularly for fine scale navigational or orientation cues (see Torres, 2017). Diurnal changes in behavior have been suggested for southward travel (Perryman et al., 1999), but not for northward migration (Perryman et al., 2002; Mate and Urbán-Ramirez, 2003). Perryman et al. (1999) suggested that whales were socializing more during the day from observations made on southward migrating whales, which resulted in a slower swimming speed, however this was not supported by any acoustic alterations.
As well as photoperiod, a change in both calling rate and composition was seen in relation to ambient conditions and the presence of vessels. The acoustic reaction by whales to changes in the soundscape and presence of human-derived noise sources has not been studied in detail for baleen whale species. Gray whales are subjected to constant additions to natural background sound levels through much of their migration, following or cutting across shipping lanes and commercial fishing areas. Their potential adaptations and thresholds for call modifications in the presence of noise are yet to be fully examined, and are outside the scope of this work (see Burnham, in review).
Studies of gray whales acoustics have relied on categories described by Dahlheim (1987), as we have done here. All core call types were found, somewhat correlated to the seasonal passage of time (Table 6). Those call types proposed for within-group communication such as, class 1 knocks and class 2 upsweeps have a significant positive correlation with time through the northward migration, with a higher occurrence in the latter part of the recordings. The reverse trend was seen for southward migrations, with moan calls decreasing significantly with time. This may be indicative of a switch from concentrated traveling behaviors to more social or prey searching/foraging behaviors in late April and May, with the opposite true for the southward migration, with some individuals still test-foraging prey patches as they return to the calving/breeding lagoons. A small proportion of calls were classified as potential “motherese” calls (Ollervides, 2001; Charles, 2011) through the manual verification process, but not further considered for analysis. They were present in the latter part of the northward migration (late March-April), within the second phase of the migration. For the recordings of southward migration they are present only in September and October. Similarly increased class 5 and 6 sub-surface exhalations during these periods suggest a shift shoreward during the second phase of the migration, particularly toward the end of April onwards for northward movements and a more offshore migration route taken after the early part of deployment recordings for southward migrations.
This study adds to the body of knowledge of gray whale acoustics use by contributing long-duration multi-year data from both north- and southward migrations. It hints at the need for more studies like this one, and more detailed analysis of the extensive vocal component of gray whales' ecology. Analysis of calling behavior should extend past notation of presence, with results of PAM recordings complementing and enhancing studies on whales using and migrating in highly ensonified waters. Recognition of the reliance on the acoustic modality by gray whales suggests the addition of human-derived noise in underwater soundscapes may be a much more serious concern than has been previously assessed. Repeated and long term behavioral baseline studies such as this one may be able to give us an important cue to changes, if we can link them to altered ecosystem dynamics that migratory whales integrate over their extensive ranges and perceptual fields.
Data Availability Statement
The datasets for this manuscript are not publicly available as it is still in use for an uncompleted doctoral program. Requests to access the datasets should be directed to RB, YnVybmhhbXJAdXZpYy5jYQ==. Supporting call classification spectrograms and example audio (.wav) files of calls are available in the Supplementary Files.
Author Contributions
RB and DD led the conception and design of the work. XM provided technical expertise with AMAR system. RB analyzed and interpreted the data and drafted the work. XM completed the analysis of detection range and provided results from the automatic detector. All authors revised, provided review and revisions and final approval of the work.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Research was conducted in the traditional territory of the Ahousaht First Nation, with support from the community and the late Chief Earl George. Particular thanks to Hughie and Keith Clarke and families for their support, logistical and otherwise. Also H.R. Stevenson for help with deployment and retrieval of the AMAR system. Funding for this work was supplied in part by the Marine Environmental Observations and Response Network's (MEOPAR) WHaLE Project. JASCO Applied Sciences provided technical support for both deployment of acoustic instruments and data analysis.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2018.00329/full#supplementary-material
References
Asa-Dorian, P. V., and Perkins, P. J. (1967). The controversial production of sound by the california gray whale Eschrichtius Gibbosus. Norw. Whaling Gaz. 56, 74–77.
Bradbury, J. W., and Vehrencamp, S. L. (1998). Principles of Animal Communication. Sunderland, MA: Sinauer Associates.
Burnham, R. E. (2015). Reproductive strategies conferring species dominance in marine mysid (Peracarida, Mysida) species in coastal waters off Vancouver Island, BC. Crustaceana 88, 1421–1438. doi: 10.1163/15685403-00003502
Calambokidis, J., Laake, J. L., and Klimek, A. (2010). “Abundance and population structure of seasonal gray whales in the Pacific Northwest, 1998–2008,” in Paper SC/62/BRG32 submitted to the International Whaling Commission Scientific Committee, Agadir, May 2010. Available from Cascadia Research, 218 W. Fourth Avenue, Olympia, WA. 50pp.
Charles, S. M. (2011). Social Context of Gray Whale Eschrichtius robustus Sound Activity. Galveston, TX: Maters Thesis, Texas A and M University.
Crane, N. L., and Lashkari, K. (1996). Sound production of gray whales, Eschrichtius robustus, along their migration route: a new approach to signal analysis. J. Acoust. Soc. Am. 100, 1878–1886. doi: 10.1121/1.416006
Cummings, W. C., Thompson, P. O., and Cook, R. (1968). Underwater sounds of migrating gray whales, Eschrichtius glaucus (Cope). J. Acoust. Soc. Am. 44, 1278–1281. doi: 10.1121/1.1911259
Dahlheim, M. E. (1987). Bio-Acoustics of the Gray Whale (Eschrichtius robustus). Vancouver, BC: PhD Thesis, University of British Columbia.
Dahlheim, M. E., Fisher, H. D., and Schempp, J. D. (1984). “Sound production by the gray whale and ambient noise levels in Laguna San Ignacio, Baja California Sur, Mexico,” in The Gray Whale, Eschrichtius robustus, eds M. L. Jones, S. L. Swartz, and S. Leatherwood (Orlando, FL: Academic Press), 511–541.
DeAngelis, M., Saez, L., MacNeil, J., Mate, B., Moore, T., Weller, D., et al. (2010). “Spatio-temporal modeling of the eastern Pacific gray whale's (Eschrichtius robustus) migration through California, Oregon, and Washington,” in Poster Presented at the 19th Biennial Conference on the Biology of Marine Mammals, Tampa, FL. 27 November−02 December 2011.
Eberhardt, R. L., and Evans, W. E. (1962). Sound activity of the california gray whale, Eschrichtius glaucus. J. Audio Eng. Soc. 10, 324–328.
Fish, J. F., Sumich, J. L., and Lingle, G. L. (1974). Sounds produced by the gray whale, Eschrichtius robustus. Mar. Fish. Rev. 36, 38–35.
Gales, R. S. (1966). “Pickup, analysis, and interpretation of underwater acoustics data,” in Whales, Dolphins and Porpoises, ed K. S. Norris (Berkeley; Los Angeles, CA: UniversityCalifornia Press), 435–444.
Gilmore, R. M. (1960). A census of the California Gray Whale. La Jolla, CA: U.S. Fish and Wildlife Service Special Scientific Report (Fisheries) 342:1–30.
Green, G. A., Brueggeman, J. J., Grotefendt, R. A., and Bowlby, C. E. (1995). Offshore distances of gray whales migrating along the Oregon and Washington coasts, 1990. Northwest Sci. 69, 223–227.
Guazzo, R. A., Helbe, T. A., D'Spain, G. L., Weller, D. W., Wiggins, S. M., and Hildebrand, J. A. (2017). Migratory behaviour of eastern North Pacific gray whales tracked using a hydrophone array. PLoS ONE 12:e0185585. doi: 10.1371/journal.pone.0185585
Hubbs, C. L. (1966). “Comments,” in Whales, Dolphins and Porpoises, ed K. S. Norris (Berkeley; Los Angeles, CA: University California Press), 444.
Lang, A. R., Taylor, B. L., Calambokidis, J. C., Pease, V. L., Klimek, A., Scordino, J., et al. (2011). “Assessment of stock structure among gray whales utilizing feeding grounds in the Eastern North Pacific,” in Paper SC/M11/AWMP4 Submitted to the International Whaling Commission Scientific Committee (Cambridge, UK), 22pp.
López-Urbán, A.A., Thode, C., Durán, Urbán, J., and Swartz, S. (2016). Two new grey whale call types detected on bioacoustic tags. J. Mar. Biol. Assoc. U.K. 98, 1169–1175. doi: 10.1017/S0025315416001697
LSIESP. (2014). Laguna San Ignacio Ecosystem Science Program. 2014 Gray Whale Research Report, Available online at: https://www.sanignaciograywhales.org/wp-content/uploads/2015/03/lsiesp-2014-grey-whale-report-web-8-jun-20141.pdf.
LSIESP. (2015). Laguna San Ignacio Ecosystem Science Program. Report of Research and Activities for 2015, Available online at: http://www.sanignaciograywhales.org/wp-content/uploads/2015/08/LSIESP-2015-FieldReport-WEB-30-JUNE-2015.pdf.
Mahoney, J., Hillis, C., Mouy, X., Urazghildiiev, I., and Dakin, T. (2014). “AMARs on VENUS: autonomous multichannel acoustic recorders on the VENUS ocean observervatory,” in Proceedings of the IEEE Xplore Conference, Valencia, Spain, November 2–5.
Mate, B. R., and Urbán-Ramirez, J. (2003). A note on the route and speed of a gray whale on its northern migration from Mexico to central California, tracked by satellite-monitored radio tag. J. Cet. Res. Manage. 5, 155–158.
McDonald, M. A., Mesnick, S. L., and Hildebrand, J. A. (2006). Biogeographic characterisation of blue whale song worldwide: using song to identify populations. J. Cetacean Res. Manage. 8:55.
Moore, S. E., and Ljungblad, D. K. (1984). “Gray whales in the Beaufort, Chukchi, and Bering Seas: distribution and sound production,” in The Gray Whale, Eschrichtius robustus, eds M. L. Jones, S. L. Swartz, and S. Leatherwood (Orlando, FL: Academic Press), 543–559.
Mouy, X., Ford, J., Pilkington, J., Kanes, K., Riera, A., Dakin, T., et al. (2015). “Automatic marine mammal monitoring off British-Columbia, Canada,” in 7th International DCLDE Workshop, La Jolla, CA, July 13–16.
Ollervides, F. J. (2001). Gray Whales and Boat Traffic: Movement, Vocal, and Behavioral Responses in Bahia Magdalena, Mexico, Galveston, TX: PhD Thesis, Texas A and M University.
Painter, D. W. II. (1963). Ambient noise in a coastal lagoon. J. Acoust. Soc. Am. 35, 1458–1459. doi: 10.1121/1.1918729
Perryman, W. L., Donahue, M. A., Laake, J. L., and Martin, T. E. (1999). Diel variation in migration rates of eastern Pacific gray whales measured with thermal imaging sensors. Mar. Mamm. Sci. 15, 426–445. doi: 10.1111/j.1748-7692.1999.tb00811.x
Perryman, W. L., Donahue, M. A., Perkins, P. C., and Reilly, S. B. (2002). Gray whale calf production 1994–2000: Are observed fluctuations related to changes in seasonal ice cover? Mar. Mamm. Sci. 18, 121–144. doi: 10.1111/j.1748-7692.2002.tb01023.x
Ponce, D., Thode, A. M., Guerra, M., Urban, J. R., and Swartz, S. (2012). Relationship between visual counts and call detection rates of gray whales (Eschrichtius robustus) in Laguna San Ignacio, Mexico. J. Acoust. Soc. Am. 131, 2700–2713. doi: 10.1121/1.3689851
Poole, M. M. (1984). “Migration corridors of gray whales along the central California coast, 1980–1982,” in The Gray Whale, Eschrichtius robustus, eds M. L. Jones, S. L. Swartz, and S. Leatherwood (Orlando, FL: Academic Press), 389–407.
Poulter, T. C. (1968). Voclaization of the gray whales in Laguna Ojo de Liebre (Scammon's Lagoons) Baja California, Mexico. Nor Hvalfangst-Tid. Norw. Whaling Gaz. 57, 53–62.
Rannankari, L., Burnham, R. E., and Duffus, D. A. (2018). Diurnal and seasonal acoustic trends in northward migrating eastern Pacific gray whales (Eschrichtius robustus). Aquat. Mamm. 44, 1–6. doi: 10.1578/AM.44.1.2018.1
Rasmussen, R. A., and Head, N. E. (1965). The quiet gray whale (Eschrichtius glaucus). Deep Sea Res. Oceanogr. Abstracts 12, 869–877. doi: 10.1016/0011-7471(65)90809-0
Scordino, J., Bickham, J., Brandon, J., and Akmajian, A. (2011). “What is the PCFG? A review of available information,” in Paper SC/63/AWMP1 submitted to the International Whaling Commission Scientific Committee (Cambridge, UK). 15pp.
Shelden, K. E. W., and Laake, J. L. (2002). Comparison of the offshore distribution of southbound migrating gray whales from aerial survey data collected off Granite Canyon, California, 1979–96. J. Cet. Res. Manage. 4, 53–56.
Stafford, K. M., Mellinger, D. K., Moore, S. E., and Fox, C. G. (2007). Seasonal variability and detection range modeling of baleen whale calls in the Gulf of Alaska, 1999–2002. J. Acoust. Soc. Am.122, 3378–3390. doi: 10.1121/1.2799905
Torres, L. (2017). A sense of scale: Foraging cetaceans' use of scale-dependent multimodal sensory systems. Mar. Mamm. Sci. 33, 1170–1193. doi: 10.1111/mms.12426
Wenz, G. M. (1964). “Curious noises and the sonic environment in the ocean,” in Marine Bioacoustics, ed W. N. Tavolga (New York, NY: Pergamon Press), 101–123.
Wisdom, S., Bowles, A. E., and Anderson, K. E. (2001). Development of behavior and sound repertoire of a rehabilitating gray whale calf. Aquat. Mamm. 27, 239–255.
Keywords: gray whales, acoustics, vocalizations, passive acoustic monitoring, migration
Citation: Burnham R, Duffus D and Mouy X (2018) Gray Whale (Eschrictius robustus) Call Types Recorded During Migration off the West Coast of Vancouver Island. Front. Mar. Sci. 5:329. doi: 10.3389/fmars.2018.00329
Received: 04 June 2018; Accepted: 28 August 2018;
Published: 18 September 2018.
Edited by:
Lyne Morissette, M–Expertise Marine, CanadaReviewed by:
Olivier Adam, Sorbonne Universités, FranceEduardo Mercado, University at Buffalo, United States
Copyright © 2018 Burnham, Duffus and Mouy. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rianna Burnham, YnVybmhhbXJAdXZpYy5jYQ==