 Tomoharu Eguchi
Tomoharu Eguchi Sam McClatchie
Sam McClatchie Cara Wilson
Cara Wilson Scott R. Benson
Scott R. Benson Robin A. LeRoux1
Robin A. LeRoux1 Jeffrey A. Seminoff
Jeffrey A. Seminoff- 1Marine Mammal & Turtle Division, Southwest Fisheries Science Center, National Marine Fisheries Service, La Jolla, CA, United States
- 2Fisheries Resources Division, Southwest Fisheries Science Center, National Marine Fisheries Service, La Jolla, CA, United States
- 3Environmental Research Division, Southwest Fisheries Science Center, National Marine Fisheries Service, Monterey, CA, United States
- 4Marine Mammal & Turtle Division, Southwest Fisheries Science Center, National Marine Fisheries Service, Moss Landing, CA, United States
- 5Moss Landing Marine Laboratories, Moss Landing, CA, United States
Environmental variability affects distributions of marine predators in time and space. With expected changes in the ocean climate, understanding the relationship between species distributions and the environment is essential for developing successful management regulations. Here we provide information on an ephemeral but important habitat for North Pacific loggerhead turtles (Caretta caretta) at the northeastern edge of their range. North Pacific loggerhead turtles nest on Japanese beaches and juveniles disperse throughout the North Pacific; some remain in the high seas of the central North Pacific whereas others transition to the eastern Pacific and forage near Baja California, Mexico. Loggerheads have also been reported along the United States west coast, with the majority of sightings off southern California. Here we describe their demography and distribution in the area, based on two aerial surveys (2011, 2015), at-sea sightings, and stranding records. Our aerial survey during fall 2015 determined density, abundance, and distribution of loggerheads in the area, when anomalous warming of the North Pacific and El Niño conditions co-occurred. Using line-transect analysis, we estimated ca. 15,000 loggerheads at the sea surface (CV = 21%) and more than 70,000 loggerheads when accounting for those that were submerged and not available for detection. Our survey during fall 2011 resulted in no loggerhead sightings, demonstrating a high variability of loggerhead density in the region. We encourage further research on loggerheads in the area to determine the mechanisms that promote their occurrence. These studies should include regular surveys throughout their foraging areas along the west coast of the North America as well as assessments of prey availability and local oceanographic conditions.
Introduction
Spatial distributions of marine predators are strongly affected by the environmental conditions they are exposed to via physiological constraints and shifts in prey distribution (Pörtner and Knust, 2007; Hazen et al., 2012). As ocean temperatures are expected to increase over the next decades (IPCC, 2013), distributions of marine animals will undoubtedly continue to change. To predict the expected changes of distributions, it is essential to understand the environmental conditions that influence a species’ whereabouts. Such understanding, however, requires dedicated study over large spatial and temporal scales. Here, we describe how shifting oceanographic conditions affect the distribution of a threatened marine species.
The loggerhead turtle (Caretta caretta) is found throughout temperate and tropical regions of all oceans (Dodd, 1988). In the North Pacific, loggerheads nest exclusively in Japan, where annual nesting populations are a fraction of their former levels (Kamezaki et al., 2003). Upon hatching, small loggerheads depart Japanese nesting beaches and disperse eastward following the Kuroshio and its extensions, and eventually disperse throughout the central North Pacific (CNP; Polovina et al., 2006; Kobayashi et al., 2008). Some juveniles transition to the eastern Pacific, particularly along the west coast of the Baja California Peninsula (BCP) (Bowen et al., 1995; Peckham et al., 2007), where they reside for decades until they mature (Turner Tomaszewicz et al., 2015). Other juveniles remain in oceanic habitats of the North Pacific where their movements have been studied extensively (Polovina et al., 2000; Kobayashi et al., 2008; Abecassis et al., 2013; Briscoe et al., 2016a). At the onset of maturity, loggerheads depart these areas and return to their natal nesting beaches in Japan for reproduction and remain in the western Pacific for the remainder of their lives (Resendiz et al., 1998; Nichols et al., 2000; Hatase et al., 2002).
Throughout their range, North Pacific loggerheads are impacted by incidental capture in fishing gear (i.e., bycatch; Gilman et al., 2007; Finkbeiner et al., 2011), which in some cases has reached catastrophic levels (Senko et al., 2017). Indeed, artisanal fisheries bycatch resulted in substantial juvenile mortality along the BCP throughout the 1990s and 2000s (Koch et al., 2006, 2013; Peckham et al., 2007, 2008), and entrapment in pound nets has been reported as a major source of death near the loggerhead nesting beaches in Japan (Gilman et al., 2010). In the CNP, interactions of loggerheads with the Hawaii-based longline fishery stimulated development and application of various fishing gear modifications (Gilman and Huang, 2016) and fishing regulations (Bartram et al., 2010). Further, a dynamic, realtime bycatch avoidance tool for longline fishers targeting swordfish (Xiphias gladius) was developed based on the affinity of loggerheads to a specific sea surface temperature (SST) range (TurtleWatch; Howell et al., 2008). This was among the first-ever tools to help fishers avoid areas of suspected high concentrations of protected species based on environmental conditions (e.g., Howell et al., 2015; Hazen et al., 2018).
Fisheries management to reduce loggerhead bycatch within United States waters has also been developed in the eastern Pacific. For example, a correlation between bycatch of loggerheads in the California-based drift gillnet fishery (CADGN) and El Niño conditions led the U.S. National Marine Fisheries Service to develop a time-limited spatial closure of the CADGN fishery off southern California (Federal Register, 2004, 2007) (Figure 1). The closure is triggered when the National Oceanic and Atmospheric Administration (NOAA) officially declares or predicts El Niño conditions in any given year, and since its designation it has been implemented three times so far (Federal Register, 2014, 2015, 2016).
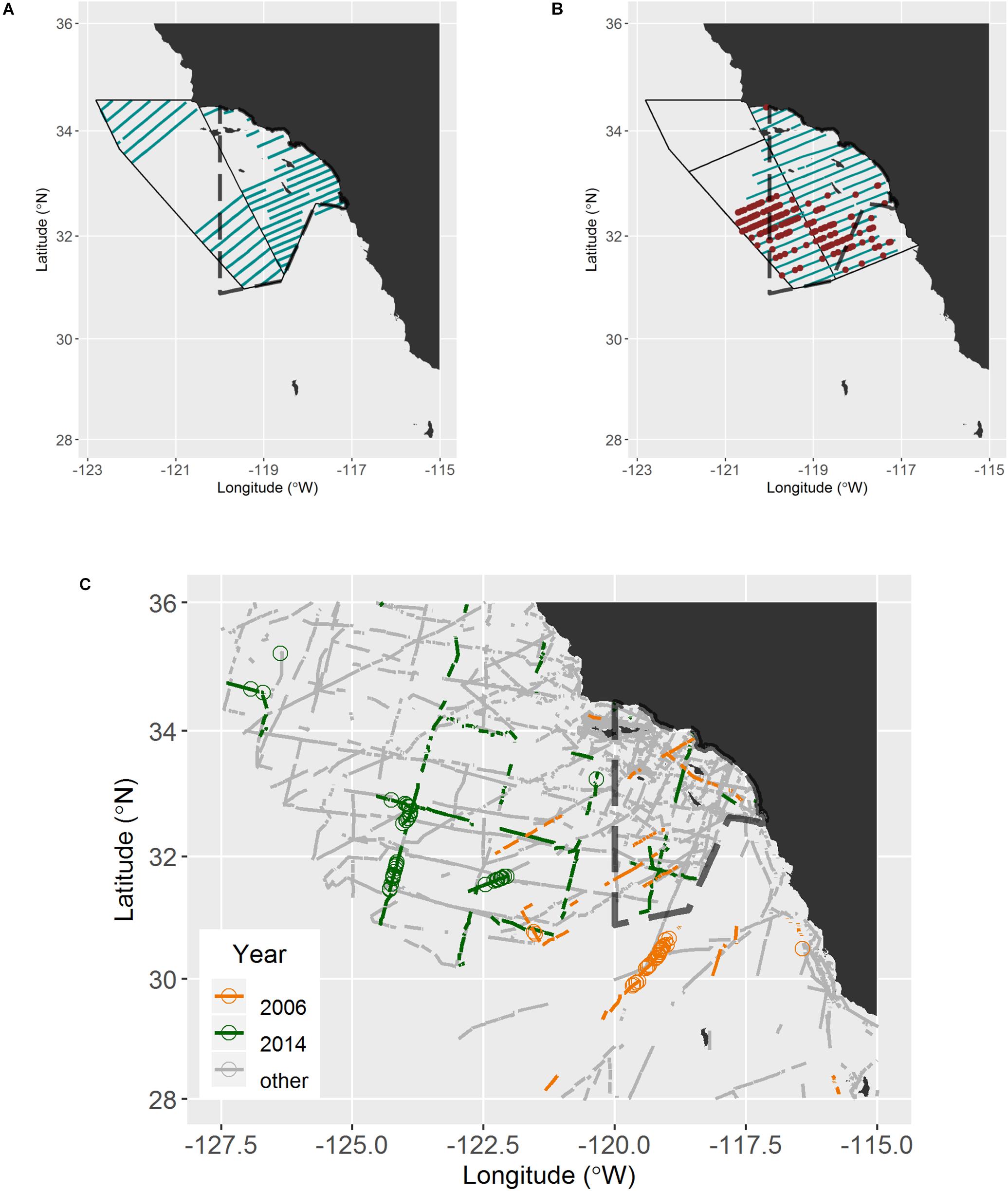
Figure 1. Survey effort and loggerhead sightings from dedicated aerial survey in 2011 (A) and 2015 (B), indicating inshore/offshore strata and truncated study area in 2015, and from marine mammal ship-board survey (C). The study area for the aerial survey is indicated by solid black lines, whereas completed track lines are indicated by dark cyan. Colored dots indicate sightings of loggerheads. The current time-area CADGN closure area (i.e., Loggerhead Turtle Management Area) is indicated with dashed lines; during closure years fisheries are prohibited in this area from 1 June to 31 August.
In the eastern Pacific, loggerhead presence has been reported from Alaska to Sinaloa, Mexico with a major foraging hotspot identified along the BCP, within the California Current Large Marine Ecosystem (CCLME; Hodge and Wing, 2000; Peckham et al., 2008; Carretta and Barlow, 2011; Carretta et al., 2017; Zavala-Norzagaray et al., 2017; Halpin et al., 2018). Occasional presence of loggerheads off southern California has been reported, with the recorded bycatch largely occurring during El Niño conditions (National Marine Fisheries Service [NMFS], 2000; Carretta et al., 2003). When the aforementioned CADGN fishery closure off southern California was established, the immediate origin of loggerheads in the region was thought to be Mexico. However, a study of body sizes and stable isotope values indicated that loggerheads caught in the CADGN fishery most likely originated from the CNP (Allen et al., 2013).
The exact path by which loggerheads arrive in the CCLME region is unknown, as are their movement patterns and residence times in the region. Environmental factors that promote the transition of loggerheads to the CCLME are likely to be associated with the North Pacific Central Water mass that has its origin in the Central Pacific Gyre. Warm, oligotrophic North Pacific Central Water lies to the west of the California Current, and there is no clear boundary between the western margin of the California Current and the edge of the Central Gyre (Lynn and Simpson, 1990). Waters of the California Current are cooler and more nutrient rich than further offshore; therefore, it is possible that when waters warm throughout the region a thermal corridor may develop that allows loggerheads to access waters off southern California, and upon arrival, encounter sufficiently warm and productive waters that enable their sustained presence for several months or more.
In this paper, we describe the presence of loggerheads off the southern United States west coast using various data sources. To determine the distribution and density of loggerheads in the region, aerial surveys were conducted during September and October of 2011 and 2015. We examined the oceanographic conditions associated with extreme inter-annual variability in turtle abundance between these 2 years. We also analyzed a variety of other datasets to enhance our understanding of loggerhead presence in relation to SST patterns in the region. To describe the demography of loggerheads in the region, size distributions were obtained from stranding and fishery observer data. These data provided valuable insight into the spatial and temporal distribution of juvenile loggerheads in the eastern North Pacific, and would serve as a baseline to compare with future distribution of loggerheads in the region as the ocean climate changes.
Materials and Methods
Six independent datasets were used in this study: oceanographic surveys, aerial surveys, shipboard marine mammal surveys, opportunistic sighting reports from the public and oceanographic cruises, CADGN fishery observer data, and stranding reports (Table 1). Observations from the oceanographic surveys were used to characterize the oceanographic conditions during aerial surveys in 2011 and 2015. Aerial surveys were limited to the southern-most portion of the United States west coast and followed basic line-transect survey protocols (Buckland et al., 2001). Data from the aerial surveys were used to estimate density and abundance within the respective study areas (see below for details). Sightings from ship-board marine mammal surveys in 2006 and 2014 also were used to estimate densities of loggerheads when there were sufficient sightings. Loggerhead stranding data from the United States west coast since 1980 were obtained from the NMFS West Coast Region Stranding Network. Fishery observer data since 1990 and at-sea sightings data since 1992 were obtained from NOAA Southwest Fisheries Science Center (SWFSC). These data sources were used to identify timing and location of loggerhead occurrence as well as size distributions.

Table 1. Datasets used in this study with corresponding numbers of turtle sighting locations.
Environmental Conditions
To characterize environmental conditions that might affect distributions of loggerheads in the area, we examined oceanographic conditions when aerial surveys were conducted. Data were obtained from two oceanographic surveys by California Cooperative Oceanic Fisheries Investigation (CalCOFI, Anon, 2012, 2016). The CalCOFI program survey methods and results are described in the cruise data reports (Anon, 2012, 2016; McClatchie, 2014). Based on these data reports, we examined flow regimes and water properties in the upper 50 – 100 m of the water column, comparing the flows and properties during the fall of 2011 and 2015 when the aerial surveys were conducted.
Aerial Surveys
The aerial survey study areas in 2011 and 2015 encompassed approximately 114,000 km2 and 129,000 km2, respectively, from 31.0 to 34.6°N and 123.0°W to the southern California coast (Figure 1). Given the warm water affinity of juvenile loggerheads in the CNP (mean = 16.3°C, SE = 1.6°C; Howell et al., 2008), we expected to find most loggerheads during late summer and fall when the water off southern California was likely to be warmest (Lynn and Simpson, 1987). We divided the study area into inshore and offshore strata by the approximate inshore edge of the California Current and previous studies of marine mammals in the area (Forney et al., 1995). The offshore stratum is dominated by the south-flowing California Current, whereas the inshore stratum is dominated by the summer-intensified, northward surface expression of the California Undercurrent (also known as the Southern California Countercurrent; Dailey et al., 1993; Hickey, 1993). Because we lacked information on the spatial distribution of loggerheads in the study area, we arranged survey track lines evenly within the study area. The angle of track lines from the shoreline was changed in 2015 to decrease transit time from airports and to shorten the longest track lines. In 2015, the southern boundary was extended south of the United States – Mexico border. Not all transect lines were completely surveyed due to airspace limitations, such as military activity and flight paths of airplanes using commercial and military airports. Details of the aerial survey protocol can be found in Supplementary Material.
Ship-Board Marine Mammal Surveys
We compiled sightings of loggerheads and unidentified turtles from ship-board marine-mammal surveys off the United States west coast (28–36°N, 127.5–115 °W) since 1991 (Figure 1). There were sufficient numbers of sightings in this dataset to conduct line-transect analyses for 2006 and 2014. Details of ship-board surveys can be found in Supplementary Material.
Distance Analysis
Line-transect analysis was conducted on distance data from aerial and ship-board surveys using the mrds package (v. 2.1.17; Laake et al., 2016) for R statistical software (v. 3.3.2; R Development Core Team, 2016). For each platform (airplane and ship), all sightings were pooled to estimate the detection function. For the aerial survey data, the most distant 1% of sightings were truncated (n = 3) because the observers were instructed not to look for turtles beyond 60 degrees from the track line. For the ship-board survey data, the most distant 15% of sightings were truncated (n = 18, >2269 m), as recommended by Buckland et al. (2001). Half-normal and hazard-rate key functions with cosine adjustments or Beaufort sea state as a covariate were used to model the detection function. We determined the best fitting detection function using AIC (an information criteria; Akaike, 1973). The best model, then, was used to estimate loggerhead density. The shipboard survey data were only used to estimate densities along portions of the surveyed track lines, not total abundance, because there was no predefined study area for loggerheads. For the aerial survey data, post-survey stratification was attempted to reduce the coefficient of variation (CV) of estimated density. There was no estimate of the probability of detecting a loggerhead on the transect line, g(0), because no studies had been conducted to determine the diving behavior of juvenile loggerheads in this area. Previous studies elsewhere in the North Pacific indicated loggerheads may spend 20–40% of their time at the ocean surface (Polovina et al., 2003; Howell et al., 2010), although surface time could vary significantly with water temperature, location, season, and age/size (Mansfield and Putman, 2013). To account for the uncertainty associated with a lack of g(0) for this survey, we computed two separate abundance estimates encompassing the range of likely values: a raw estimate without any g(0) correction (minimum abundance) and an estimate that assumes loggerheads in the region spend only 20% of their time at the ocean surface (maximum abundance).
Stranding Records, Fishery Observer Data, and Opportunistic Sightings
Spatial and temporal distribution of loggerheads in the eastern North Pacific was examined by compiling sighting records of loggerheads from various sources, including scientific cruises, opportunistic reports from the public, official stranding reports, and CADGN fishery observer data. Scientific cruises included marine mammal surveys (Table 2) and CalCOFI cruises. A marine turtle sightings hotline email address (the SWFSC Marine Turtle Sightings Hotline) was created on April 1, 2015 for the public to report sightings of marine turtles along the west coast of the United States.
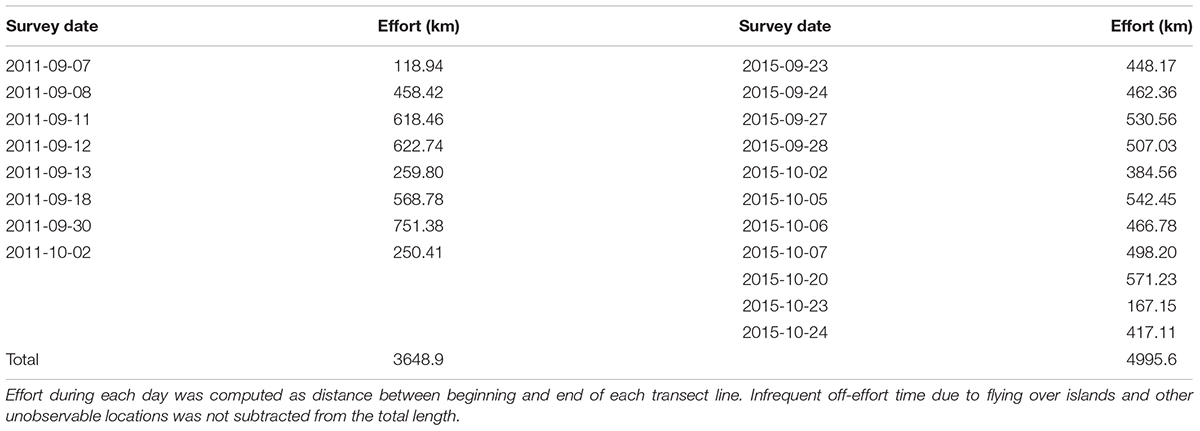
Table 2. Aerial survey effort during 2011 and 2015.
We used all sightings to determine the thermal habitat for loggerheads off southern California. We obtained sea surface temperature data (AVHRR Pathfinder version 5.3 before 2002 and Multi-scale ultra-high resolution MUR SST analysis after 2001) from NOAA1, available at the resolution of 0.0417 or 0.01 degrees, respectively, at the time and location of sightings. To take into account the spatial and temporal autocorrelation of SST within the aerial survey data, we show these distributions for different data sources, i.e., aerial survey data, CADGN observer data, opportunistic sightings from shipboard marine mammal surveys, oceanographic surveys, and the general public. For the aerial survey data, SST along the track lines and SST at loggerhead sighting locations were compared.
Size Distribution of Loggerheads in CCLME
Size distribution of loggerheads in the CCLME was characterized from stranding and CADGN fishery observer datasets. Mean size and size range were compared to those for loggerheads in BCP, Mexico based on published reports by Peckham et al. (1995–2007, 2008, n = 1918) and Turner Tomaszewicz et al. (2003–2011, 2015, n = 146). All sizes were recorded as curved carapace length (CCL). Gaussian kernel density estimates with Silverman’s rule-of-thumb bandwidth [0.9 × A ×n-1/5, where A = min(SD, IQR/1.34) and n = sample size; Silverman, 1986, p. 48] were used to smooth CCL of the four datasets. Means were compared to detect differences in CCLs among the datasets.
Results
Environmental Conditions
In fall 2014, ocean conditions along the United States west coast were significantly different from the long-term average, a trend that lasted throughout 2015 (Leising et al., 2015) due to a combination of anomalous North Pacific warming and interaction with developing El Niño conditions. These conditions coincided with the 2014 marine mammal ship survey and the 2015 aerial survey.
CalCOFI survey data indicated strong inter-annual oceanographic differences between fall 2011 and 2015, the years of the aerial surveys. The main inter-annual difference was the contrast between the cool-core eddy south of Pt. Conception and an anomalously warm and saline surface pool that appeared to be related to an intrusion of waters from the south. Near surface (10-m depth) temperatures in fall 2011 were dominated by a cool-core eddy with a core of 13–15°C water contrasting with warmer temperatures of 17.5–19.5°C further offshore (Supplementary Figure S1). Dynamic height anomalies were dominated by this eddy and by strong meandering of the currents (Supplementary Figure S2). In contrast, the dominant surface temperature feature in fall 2015 was a warm intrusion of 21.0–22.5°C water coming from south of the United States – Mexico border, clearly shown in dynamic height anomalies (Supplementary Figure S3). The intrusion warmed the coastal waters of the Southern California Bight (SCB) (Supplementary Figure S1) and mean mixed layer temperature was ∼2.5°C warmer during fall 2015 than during fall 2011. Surface water was as much as 4–5°C warmer than adjacent water and was confined to the upper 20–30 m of the water column.
Survey Effort
In 2011, approximately 3,650 km of aerial track lines were surveyed during 8 days from 7 September to 2 October and no loggerheads were observed. In 2015, approximately 5,000 km of track line were surveyed over 11 days from 23 September to 24 October and 215 loggerheads were observed (Table 2 and Figure 1). Strong wind, cloud cover, and military exercises limited our survey effort during both years, especially in the offshore strata. Consequently, the northwest portion of the study area (∼25,000 km2) was excluded from abundance estimation (Figure 1). The spatial distribution of loggerheads off southern California was more concentrated in the southern portion of the offshore area than the northern area; however, this finding may reflect the low search effort in the northern offshore area (Figure 1).
During the 2006 and 2014 ship-board marine mammal surveys, approximately 960 and 1,926 km of track line were surveyed, respectively, during adequate sea state conditions (Figure 1 and Table 3). Search effort within the polygon occurred from 7 April to 6 December 2006 and from 5 August and 9 December 2014. In 2006, turtles were seen from 8 April to 30 July, whereas in 2014 they were seen from 14 October to 7 December. Sightings occurred on less than 50% of the track lines within the polygon (Table 3). During those 2 years, most of the turtles were identified as loggerheads (n = 110), and the remainder were unidentified to species (n = 56).
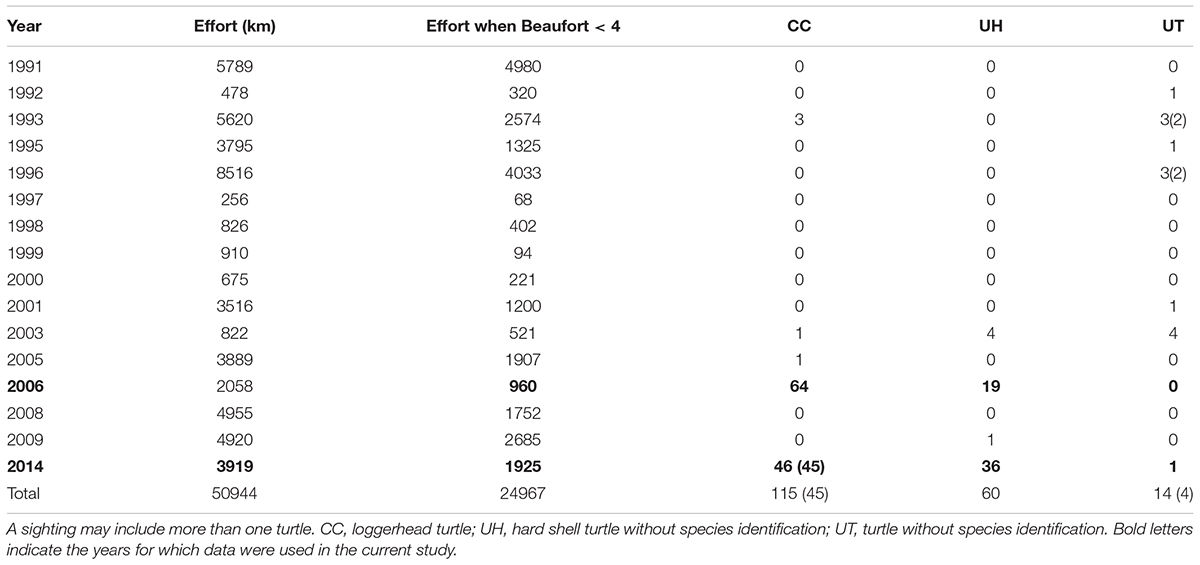
Table 3. Marine mammal survey effort (total and when Beaufort sea state < 4) and number of turtle sightings (Beaufort < 4 in parentheses when different) between 1991 and 2014 within the polygon defined by latitude 28–36°N and longitude 127.5–115°W.
Distance Analysis
For the 2015 aerial survey data, a hazard rate (HR) detection function without any adjustment terms or Beaufort sea state covariate was found to be most parsimonious (Table 4). The estimated effective strip half-width was 156.2 m (SE = 7.2; Figure 2). Encounter rate was more than three times greater in the offshore stratum than inshore stratum (Table 5). Post-stratification reduced the CV of the estimated densities (Table 5). The overall density of loggerheads in the study area was 0.15 km-2 (0.08 vs. 0.24 km-2 in the inshore and offshore strata, respectively) and the estimated total minimum abundance was 15,286 (95% CI: 9945–23,495, CV = 0.21; Table 5) during the 2015 survey period. Removing sightings that occurred when the Beaufort sea state was greater than 3 (n = 21) did not affect the selection of the best detection function or parameter estimates (results not shown), which permitted maximal spatial coverage of the study area. Applying the g(0) correction, i.e., assuming loggerheads in this area spend 20% of their time at the ocean surface, the maximum abundance was 76,430 (95%CI: 49,725–117,475).
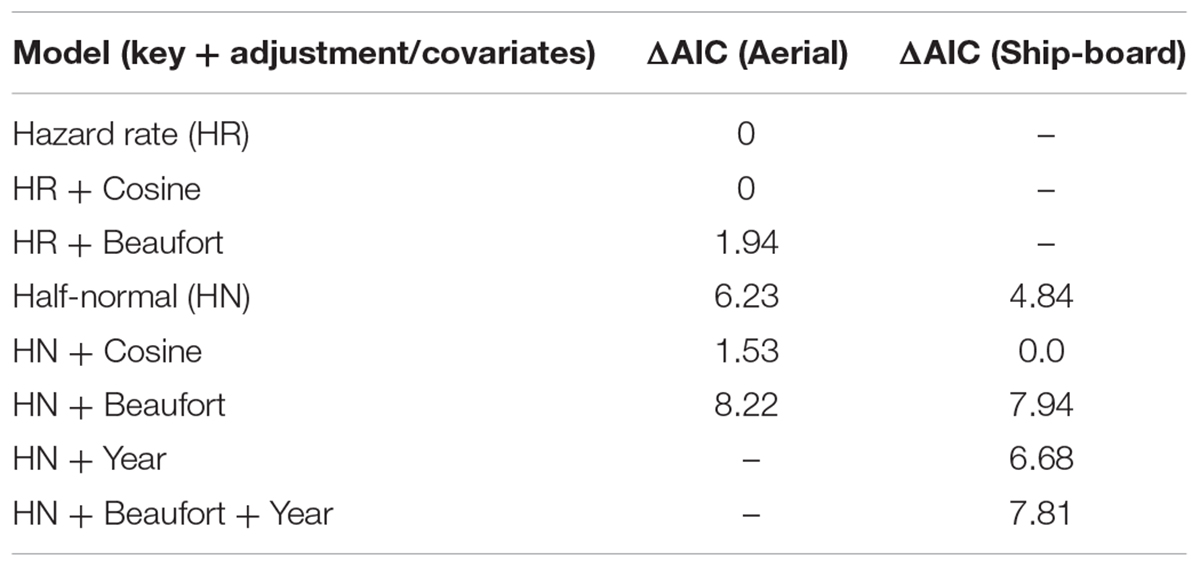
Table 4. Delta AIC values among six detection-function models that were fitted to aerial and ship-board sightings data.
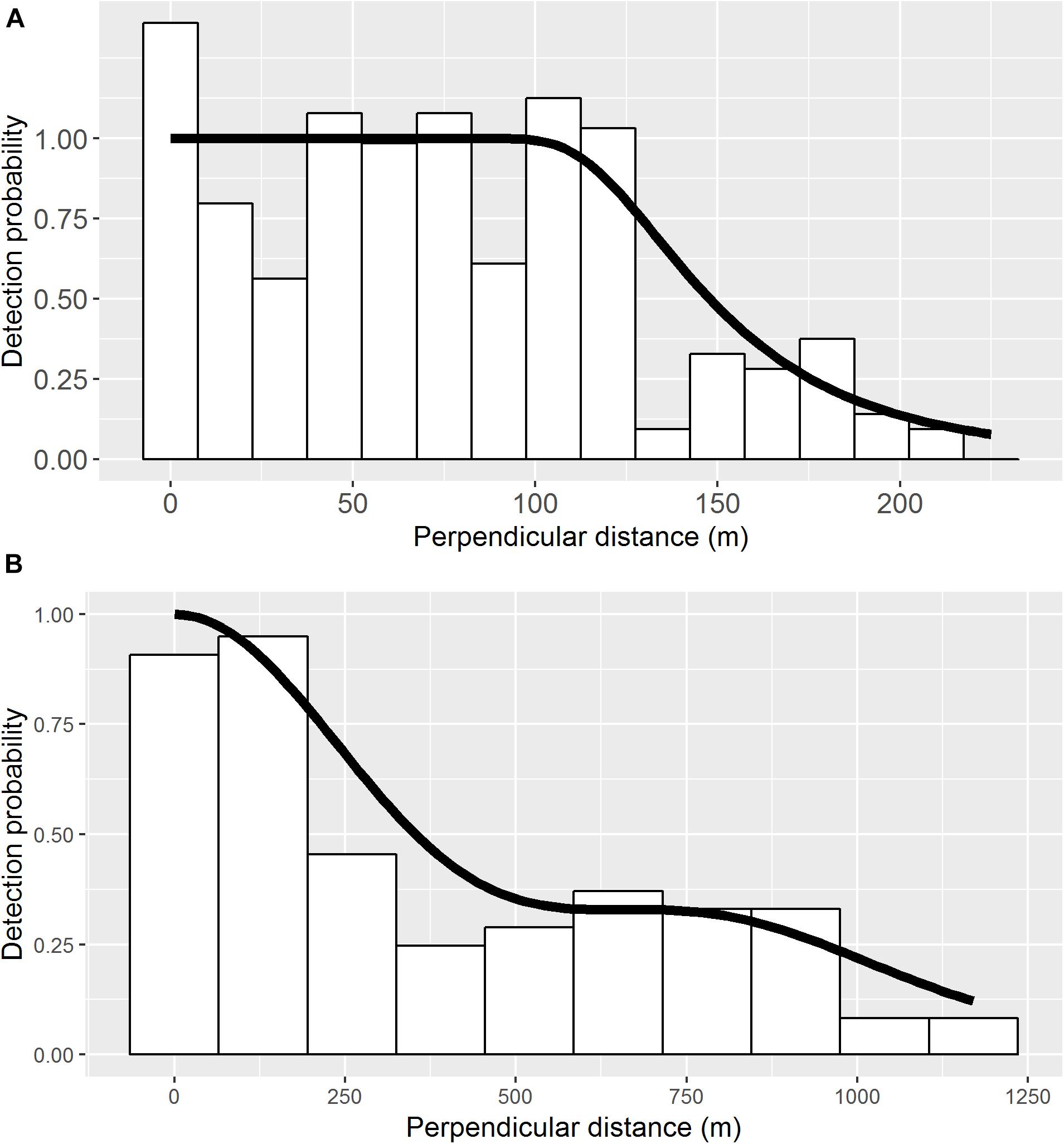
Figure 2. Perpendicular sighting distances (m) and fitted detection functions for aerial (A) and ship-board marine mammal survey (B) when detecting turtles. For the aerial survey, hazard-rate detection function without adjustment terms was the best fit, whereas half-normal with cosine adjustment terms (order 2) was the best fit for the ship-board survey based on AIC values (Table 4). The estimated effective strip half-widths were 156.1 m (SE = 7.2 m, n = 212) for aerial survey and 540.0 m (SE = 75.1 m, n = 99) for ship-board survey.
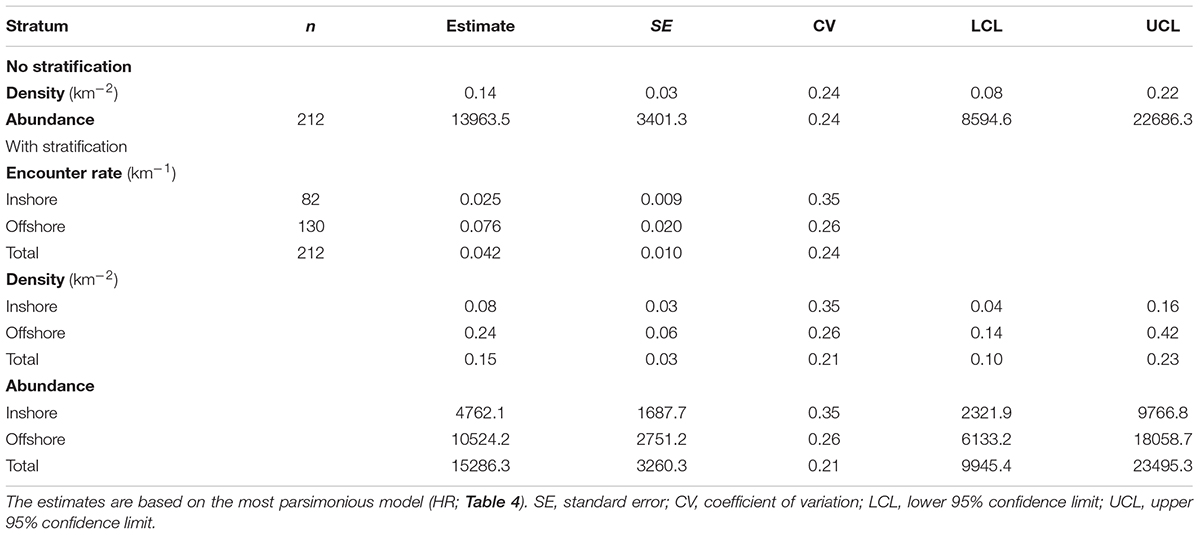
Table 5. Estimates and uncertainties of encounter rates, densities, and abundance of loggerheads in the study area from the line-transect survey conducted during fall 2015.
For the ship-board survey data, the half-normal key function with cosine adjustments without Beaufort sea state covariate was found to be most parsimonious based on AIC values (Table 4). Estimated effective strip half-width was 540.0 m (SE = 75.1 m, Figure 2). Estimated densities for 2006 and 2014 were 0.08 (SE = 0.04, n = 70) and 0.04 (SE = 0.01, n = 67) turtles km-2, respectively.
Stranding Data, Observer Data, and Opportunistic Sightings
From 1980 to August 2017, 75 loggerheads were reported to the NMFS West Coast Region Stranding Network, of which 22 did not have exact coordinates but were all found in the state of California. Stranding records indicated loggerheads have been reported as far north as Alaska and British Columbia, Canada. The majority of strandings, however, occurred within California (n = 62), specifically south of Pt. Conception (34.45°N; n = 37). For stranded loggerheads with size information, curved carapace lengths (CCL) ranged from 24.1 to 86.3 cm (54.5 ± 2.4, mean ± SE, n = 40). Among 75 loggerhead strandings, 40 occurred during summer months (July, August, September, and October), whereas 35 occurred from November through June.
Fishery observers aboard CADGN fishing vessels reported 17 bycaught loggerheads from 1992 to 2006, whose CCLs ranged from 22 to 59 cm (42.1 ± 0.6, mean ± SE). All bycatch events occurred south of Pt. Conception, bounded by latitudes 34.1 and 31.7°N (Figure 3). Although bycatch occurred throughout the year, these events were most frequent in August (n = 7 out of 17).
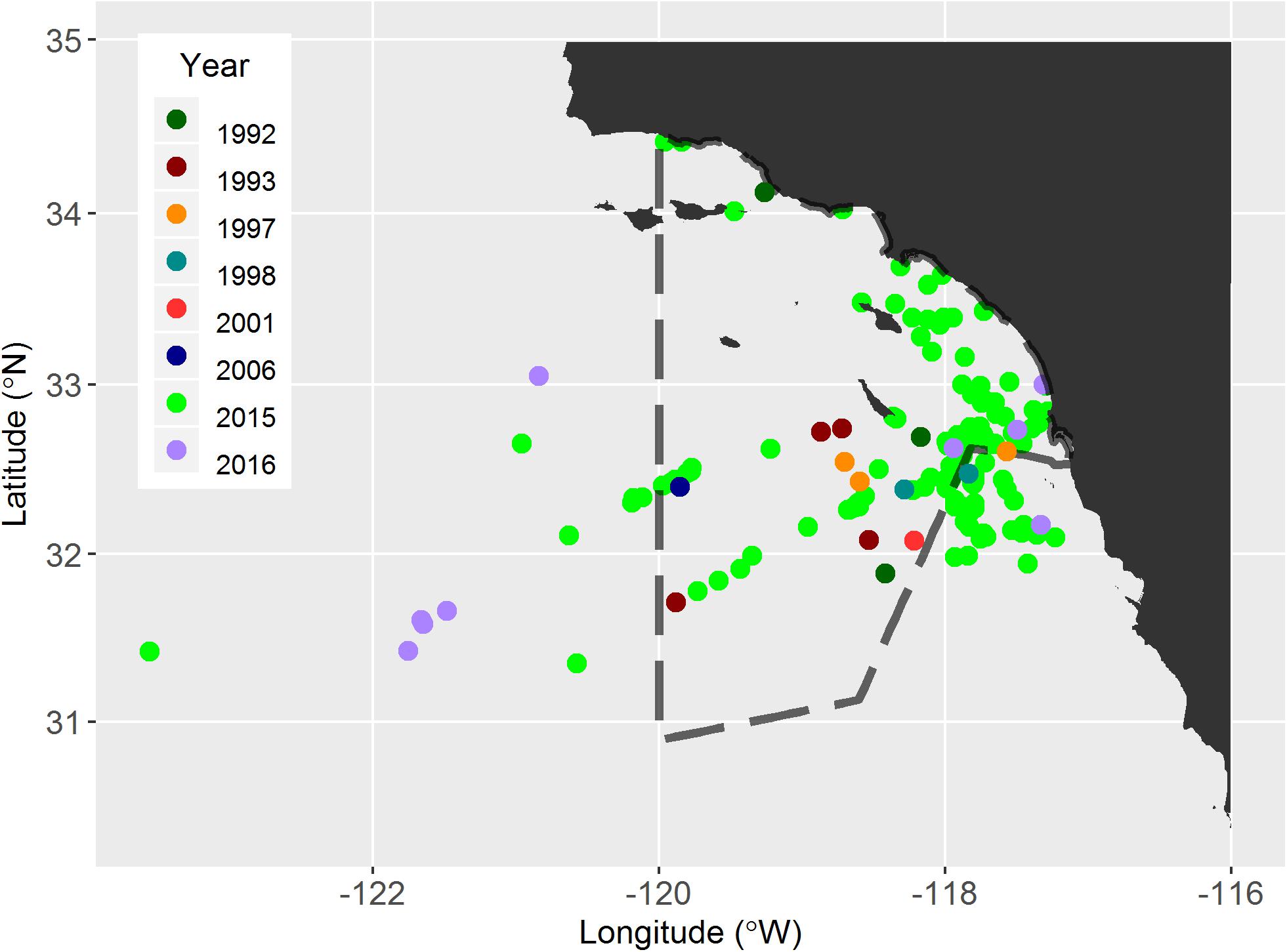
Figure 3. Location and timing of loggerheads and unidentified turtles opportunistically sighted by the public and from oceanographic cruises and bycaught in the CADGN fishery. Bycatch occurred from 1992 to 2006, whereas reported observations are in 2015 and 2016. The current time-area CADGN closure area (Loggerhead Turtle Management Area) is indicated with dashed lines.
A total of 306 hard shell turtles were reported from Marine Mammal survey cruises and opportunistic reporting, which included 125 loggerheads and 181 unidentified species (Table 1). Since its inception in April 2015 to the end of July 2017, 39 loggerheads have been reported to the SWFSC Marine Turtle Sightings Hotline by recreational boaters, surfers, sport fishing boat captains, and naturalists onboard whale watching boats. All but two sightings included latitude and longitude with the precision of at least 0.1 min.
SST at Sighting Locations
Among the 306 sightings, SST was obtained for 293 locations. Although these included unidentified hard shell turtle species, we think the majority of them were loggerheads based on their locations. Sea surface temperatures at sighting locations ranged from 14.4 to 27.5°C (Figure 4). During the aerial survey in 2015, loggerheads were found in water with SST between 20 and 21°C in a disproportionately higher proportion (49.3%) than it was available (20.9%). Another disproportionate peak of SST at turtle sightings was found at approximately 23.5°C (Figure 4). CADGN bycatch occurred in waters at the lower half of the temperature range (14.4–21.1°C). Sea surface temperature at opportunistic sighting locations indicated a bimodal distribution with peaks at approximately 19 and 21°C.
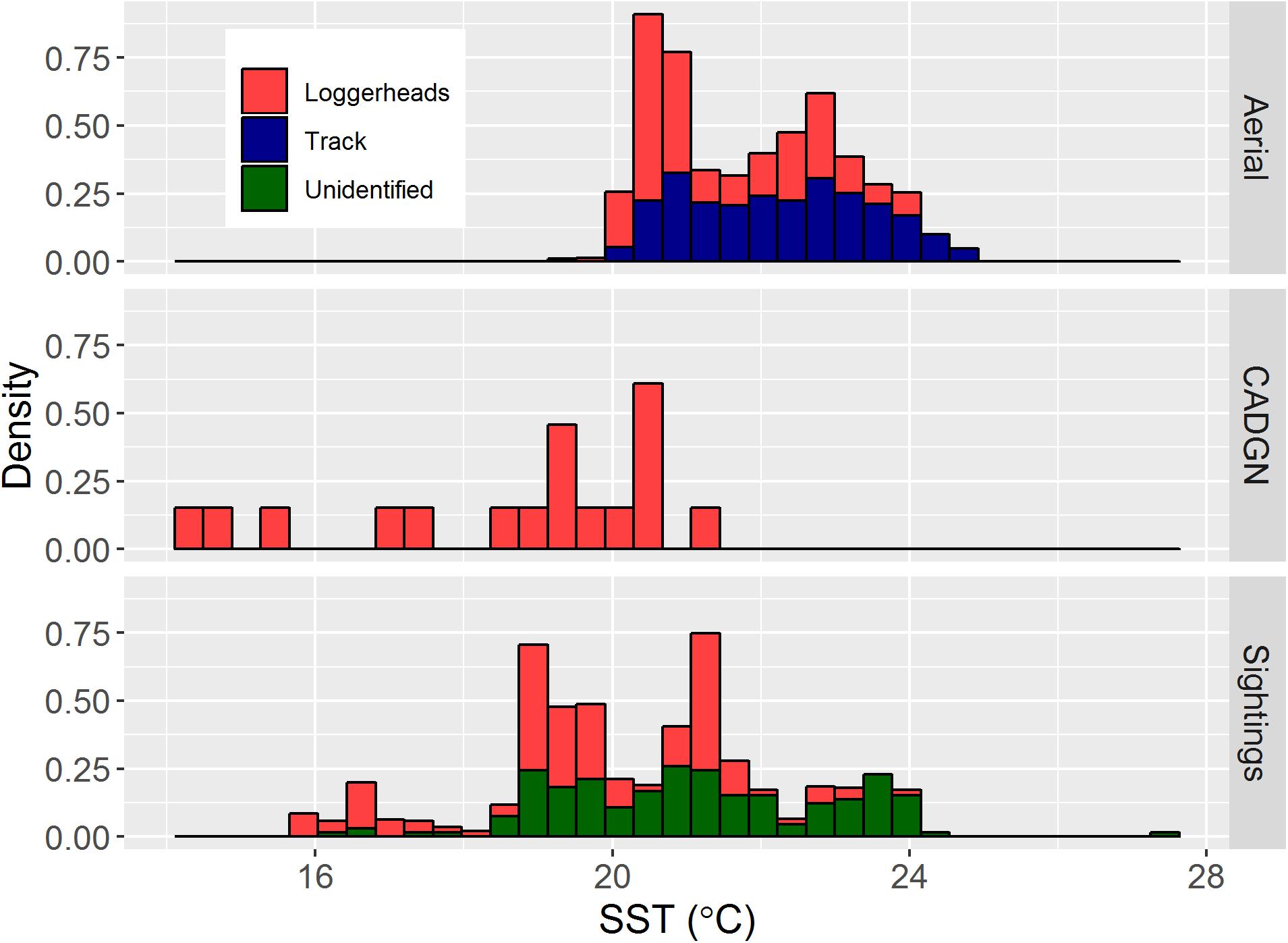
Figure 4. Distributions of SST at loggerhead sightings from various data sources. The (top) “Aerial” shows the SST at aerial survey sighting locations (n = 215) and along the track lines (split into 2 km chunks; n = 2643) during the 2015 aerial survey. The (middle) “CADGN” shows the SST at the 17 bycatch locations. The (bottom) “Sightings” shows the SST for other sighting locations including marine mammal shipboard surveys, oceanographic cruises, and from the general public for loggerheads (n = 123) and unidentified hard shell turtles (n = 170).
Discussion
Our study is the first to characterize loggerhead presence off the United States west coast using multiple lines of evidence. Clearly, the most influential data for our analyses come from the aerial surveys in 2011 and 2015, and the disparity in survey results (0 vs. 215 loggerheads) underscores the ephemeral nature of loggerheads in the area. Our analyses of ship-board survey data as well as sighting and stranding records complement our aerial surveys and further support that loggerheads are present along the southern California coast most commonly during warm water conditions.
Environmental Conditions
The Oceanic Niño Index (ONI), which is NOAA’s climate indicator for categorizing El Niño Southern Oscillation conditions as El Niño or La Niña, characterized fall 2011 as La Niña (anomalously cool) conditions and fall 2015 as El Niño (anomalously warm) conditions (Supplementary Figure S4). The Pacific Decadal Oscillation index (PDO; Mantua et al., 1997) also exhibited contrasting oceanographic conditions between 2011 and 2015; the PDO was negative in 2011, indicating lower than average North Pacific SST pattern, but changed to positive beginning in 2014 and extending to 2015 (Supplementary Figure S4). During 2014, the CCLME was affected by the North Pacific warm anomaly in the Gulf of Alaska (Bond et al., 2015) and by August 2014, most of the CCLME was affected by a moderate El Niño (Leising et al., 2014, 2015). In December 2014, the North Pacific and the southern California warm anomalies started (Leising et al., 2015) and the PDO reached its highest value since June 1997, indicating the abnormally high North Pacific SST condition (Supplementary Figure S4; Leising et al., 2015). Combined with other factors, such as atmospheric forcing, advection, and responses to El Niño, waters off the west coast of United States were experiencing a marine heat wave throughout 2014 and 2015 (Leising et al., 2015). The anomalously warm conditions were associated with changes in planktonic communities (Sakuma et al., 2016; Santora et al., 2017) as well as the arrival of several exotic species to the region, including pelagic red crabs, sea snakes (Pelamis platura), hammerhead sharks (Sphyrna sp.), dolphin fish (Coryphaena hippurus), wahoo (Acanthocybium solandri), manta rays (Manta hamiltoni), and pygmy killer whales (Feresa attenuata) (Barlow, 2016, Captain Paul Fischer, pers. comm., various news reports2), as well as loggerheads. During our aerial survey in 2015, we observed some of these species (hammerhead sharks, manta rays, and dolphin fish). With the predicted warming of oceans in the future (Cavole et al., 2016), the presence of these exotic species in the southern California waters may become a more common occurrence.
Abundance and Density
This study provides the first estimate of spatial and temporal variability of loggerhead density at the northeastern limit of their range in the Pacific. The extreme warm condition in 2014–2015 appeared to be a large factor in promoting their presence during this period, and the observed temporal variability in density across years indicated that this is an ephemeral habitat for the species. In contrast, the Baja California habitat in Mexico is known to be a permanent habitat for the species, apparently regardless of the presence or absence of El Niño. The estimated density of loggerheads in 2015 (0.24 km-2) was comparable to those from the Baja habitat (0.205–0.265 km-2: Seminoff et al., 2014), suggesting that when available, this northern habitat may be as important for the species as the Baja habitat.
The high productivity of the CCLME likely promotes sustained high quality habitat for juvenile North Pacific loggerheads, especially during anomalously warm periods. Based on the difference in size distributions between the central west coast of the BCP, Mexico, and southern California (Figure 5), these two locations appear to contain distinct life-stage groups. The predominant age group in the BCP habitat is large juveniles, whereas a large variability of age groups, including very small individuals, were found in southern California. Seminoff et al. (2014) reported the mean annual abundance estimate of approximately 43,000 loggerheads (approximate 95% CI – 15,000 to 100,000) in the Mexican foraging area. Simply adding the abundance estimates from these two regions results in >100,000 loggerheads within the CCLME, albeit the estimates were separated by 10 years.
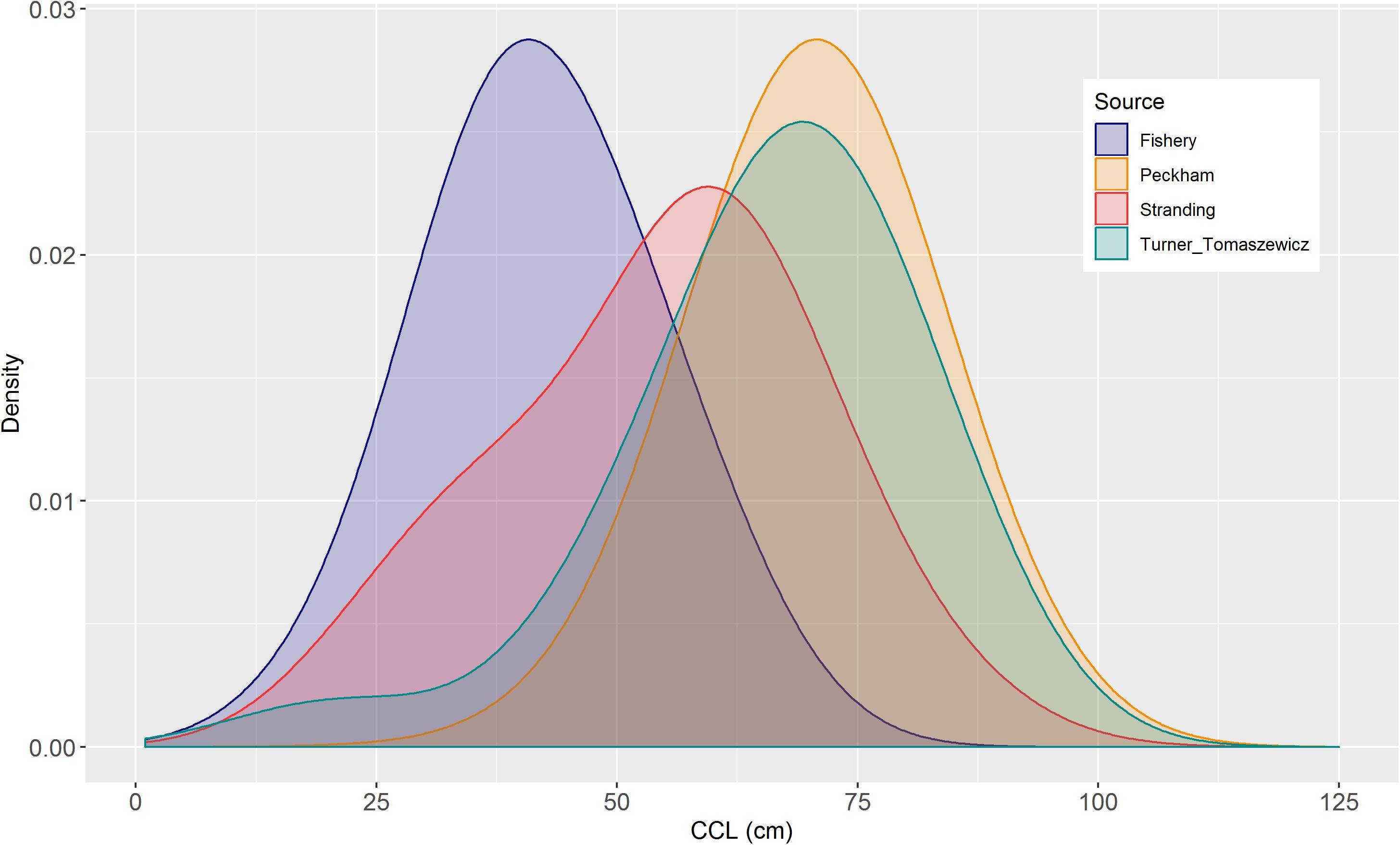
Figure 5. Size distributions of loggerheads from four data sources. Data for Peckham et al. (2008) were simulated from the means, standard deviations, and sample sizes provided in Table 4 in Peckham et al. (2008) using normal distributions.
We are unsure what proportion of all North Pacific juvenile loggerheads this number represents. The most recent estimate of annual nesting female abundance from nesting beaches in Japan is approximately 2300 (Matsuzawa, 2011). If this number is still valid, the CCLME provides foraging habitat for a significant proportion of loggerheads in this population. Based on these numbers, it is clear that the California Current foraging areas are important for North Pacific loggerheads. To further clarify the value of these areas, regular aerial surveys should be conducted to estimate loggerhead abundance and distribution.
It is also possible that some or all of the turtles found off southern California are part of a central Pacific foraging group that moves between the eastern and central Pacific as thermal corridors open (e.g., Abecassis et al., 2013). Allen et al. (2013) found that loggerheads bycaught in California-based fisheries originated from the CNP based on stable isotope analysis. However, the destination for loggerheads departing offshore areas of California are less clear and deserving of future study. Satellite telemetry efforts should be implemented to characterize the movements of individual turtles tagged within the southern California region. Further sampling and stable isotope analyses should also be conducted to help clarify if the California-CNP linkage is a consistent pattern or if perhaps some turtles found in United States west coast waters originate from Baja, Mexico.
Distributions of Loggerheads
Although less common north of southern California, loggerheads are found widely along the United States west coast. In the stranding records, a loggerhead was reported as far north as 58.5°N. Hodge and Wing (2000) described two loggerheads (one dead and one alive) along the coast of Alaska north of 57°N in the early 1990s, and more recently Halpin et al. (2018) reported a loggerhead in waters of British Columbia, Canada. Strandings of loggerheads in northern/central California, Oregon, and Washington have become more frequent during the last decade, coinciding with warmer water conditions throughout the CCLME. Eight strandings were reported since 1997 north of Pt. Conception (34.45°N). In 2017, two strandings were reported in Oregon in the first 3 months. It is well known that only a small fraction of carcasses at sea reach the shore and is reported (Epperly et al., 1996; Eguchi, 2002; Koch et al., 2013). Postmortem drifting adds another source of difficulty in interpreting stranding locations as a proxy for the distribution of live animals without considering oceanographic information, such as wind and current (Hart et al., 2006). In this study, we provided the observed latitudinal distribution of stranded loggerheads, which indicated their occurrence off southern California, but we hesitate in drawing firm conclusions about the origins of these turtles. Nevertheless, this information can be treated as the baseline to which future stranding patterns are compared as the large-scale oceanographic conditions and total abundance of North Pacific loggerheads change in the future.
The distribution of loggerheads in our study area appeared to be heterogeneous (Figure 1). Presence of loggerheads appeared to be ephemeral, where they were not present in the same location on consecutive days (Paul Fischer, the captain of F/V Outer Limits, a San Diego based sport fishing vessel, personal communication). Although the apparent absence of the turtles could have been caused by the change in sighting conditions rather than movements of turtles, this disparity is likely due, at least in part, to their high motility. Satellite telemetry of juvenile loggerheads within the SCB supports this possibility, as three small juveniles (29.0, 31.5, and 61.9 cm straight carapace length: SCL) tracked in the area had high daily vagility (7.6 ± 1.6 km d-1, mean ± SE) and moved large distances in the study area (399–730 km) during their tracking durations (SWFSC, Unpublished data). Briscoe et al. (2016b) showed that movements of telemetered juvenile loggerheads were significantly different from those of simulated drifting particles. This suggests that the movements of these three turtles was not necessarily tied to prevailing ocean currents, perhaps to stay within a preferred SST range. The ability of small turtles to stay within a desirable range of SSTs in oceanic regions has been reported elsewhere in the Pacific (Abecassis et al., 2013).
During the 2015 aerial survey, loggerheads off southern California were found in warmer waters than in the CNP (middle 50th percentiles: 18.6–22.9°C vs. 15.6–17.1°C, Howell et al., 2008). Using several data sources together, however, loggerheads off southern California were found in SST between 14.4 and 27.5°C (Figure 4), which encompassed many reported SST ranges around the globe, where means ranged from 16.3°C in the North Pacific to 28.8°C in the Gulf of Mexico (Table 6). Although SST may be a proxy for loggerhead habitat, it may not be a good predictor of loggerhead presence in all systems. Instead, affinity to a certain range of SST may be ecosystem-specific, as has been shown for other marine turtle species (e.g., leatherback turtles, Dermochelys coriacea; Benson et al., 2011). A broad analysis of relationships between presence/absence of loggerheads and environmental variables may provide insights into what variables are important in predicting presence of loggerheads.
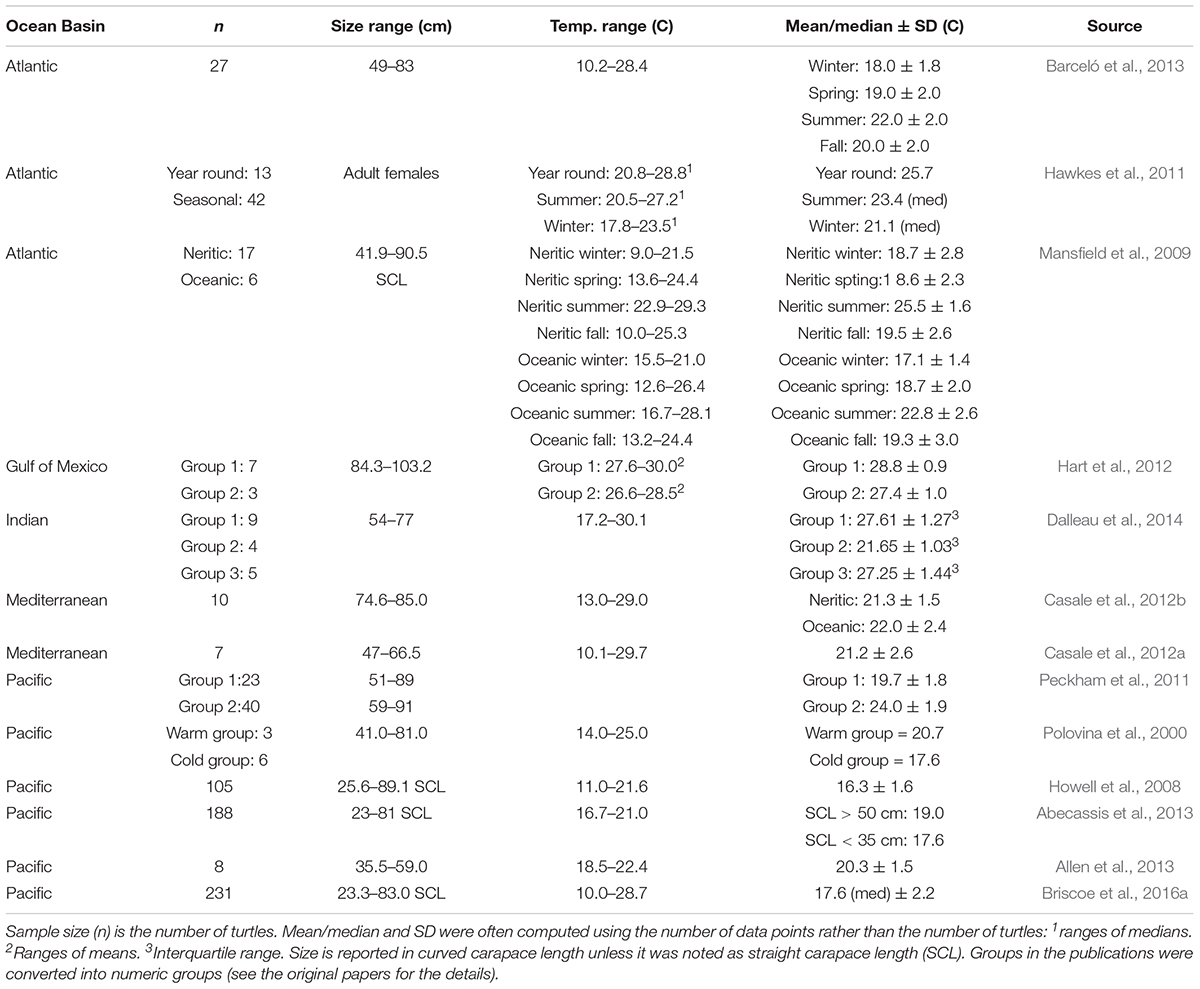
Table 6. Published information about SST and loggerhead turtle relationships.
A study in the Atlantic Ocean indicated fidelity of juvenile loggerheads to foraging sites over years and another study indicated a large proportion of transients within a foraging site (Sasso et al., 2006; Mansfield et al., 2009). The observed coexistence of transients and philopatry to foraging areas suggested that multiple foraging strategies exist within a loggerhead population. Consequently, it is possible that some juvenile loggerheads in the BCP foraging ground may move to high seas as such behavior has been observed in the Atlantic (Mansfield et al., 2009). These ‘transient’ loggerheads from nearshore BCP, as well as those from the pelagic CNP, may be found within the southern California area when environmental conditions are optimal. Particle tracking, regular surveys, stable isotope analyses, and satellite telemetry with careful examination of oceanographic conditions would help elucidate the mechanisms by which loggerheads enter the southern California waters and their movements within the North Pacific.
Effects of changing ocean climate and local ocean conditions on loggerhead distributions should be examined closely. Changes in species compositions as a response to El Niño and other local ocean conditions have been reported globally (e.g., Benson et al., 2002; Wernberg et al., 2012). The observed high density of loggerheads in 2015–2016 also was likely caused by the warming of the Eastern Pacific, as we described in the Environmental Conditions section above. As the ocean temperature is expected to increase in the coming years, occurrence of loggerheads in the area may become more common.
Size Range of Loggerheads in the CCLME
The observed size range of loggerheads indicated the presence of multiple life stages off southern California and in the CCLME in general (Figure 5). Stranding records encompassed very small individuals of the same sizes as those caught in the CADGN fishery, as well as large individuals of the same sizes as those found in the BCP foraging ground (71.8 ± 0.21, n = 1918, Peckham et al., 2008; 65.9 ± 1.23, n = 146, Turner Tomaszewicz et al., 2015). Further, we captured two juvenile loggerheads off southern California with CCLs of 22.3 and 61.9 cm in two consecutive days, the smallest of which was about 1-year-old, whereas the largest may have been 20 years or older (Turner Tomaszewicz et al., 2015). Together, these data underscore the fact that multiple cohorts of loggerheads exist in the CCLME.
Size distributions of loggerheads along the west coast of North America are comparable to those found along the west coast of South America where loggerheads originate from Australia and New Caledonia (Alfaro Shigueto et al., 2008; Boyle et al., 2009; Donoso and Dutton, 2010). The minimum size found in our study (22 cm) was also similar to that reported from the eastern South Pacific (26 cm; Alfaro-Shigueto et al., 2004). Size distribution data combined with loggerhead growth rates will provide useful information about juvenile loggerheads in each of these regions. Because El Niño events strongly affect these regions, influence of changes in SST and ocean productivity on these populations should be examined.
Conclusion
Considering all the available data sources on loggerheads in the Northeast Pacific, we found that a large number of multiple cohorts of loggerheads exist in the region. Spatial distributions of loggerheads appeared not to associate with a particular oceanographic feature, including SST or meso-scale eddies, as was suggested by studies from different regions. Consequently, generalization of findings about association of loggerheads to environmental features is not warranted. With the expected change in the ocean climate, we expect to see loggerheads and other exotic species in the region more frequently. In order to monitor the effects of changing environment on distributions of various species in the area, aerial surveys need to be conducted during thermally anomalous years as well as non-anomalous years in order to determine inter-annual variability in density, abundance, and distribution of loggerheads in the region. Multi-year dedicated surveys in the surrounding areas and concurrent telemetry studies are also needed to determine how loggerheads use this area. Long-term studies need to be conducted to help elucidate the effects of oceanographic variability and climate shifts on loggerhead distribution and movements. Detailed studies at various locations using a wide variety of research techniques are necessary to elucidate demography and ecology of loggerheads within a population.
Author Contributions
TE, JS, and SB conceived the study, secured funding, and collected data. SM and CW provided oceanographic information. RL dealt with permit, stranding data and sighting database, budget, and other administrative tasks. TE drafted the manuscript. TE, JS, SB, SM, CW, and RL edited the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Nicki Beaulieu, Joel Schumacher, Camryn Allen, Erin LaCasella, Dan Prosperi, Jim Carretta, Lisa Komoroske, Morgane Lauf, and Dan Lawson for their keen eyes on airplanes as well as other duties, related to this study, including stranding responses and data entry. We also thank pilots (Shanae Coker, Rick De Triquet, Kevin Doremus, Chris Kerns, Robert Mitchell, and Sandor Silagi) of NOAA Office of Marine and Aviation Operations for their excellent aviation skills. Commander Michelle Kemp at San Nicolas Island generously allowed us to use Outlying Field San Nicolas Island. We also appreciated those who responded to strandings, enduring nasty smells for the sake of science. Thanks also to those who reported turtle sightings to the SWFSC Sea Turtle Sighting Hotline (c3dmc2MudHVydGxlLXNpZ2h0aW5nc0Bub2FhLmdvdg==) and to Lynn Massey for entering sightings data to the database. Karin Forney and Elizabeth Becker generously shared R code to analyze sightings data. Al Jackson kindly responded to our requests for sightings data from various marine mammal surveys. Suzy Kohin shared the DGN observer data. Calandra Turner Tomaszewicz kindly shared her raw turtle size data with us. Jim Carretta, Christina Fahy, and Penny Ruvelas provided constructive comments to the manuscript. Aerial and shipboard surveys were conducted under the NMFS permit numbers 774-1714 and 14097, respectively.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2018.00452/full#supplementary-material
Footnotes
- ^ http://coastwatch.pfeg.noaa.gov
- ^Pelagic red crabs: https://www.washingtonpost.com/news/capital-weather-gang/wp/2015/06/17/red-crabs-swarm-southern-california-linked-to-warm-blob-in-pacific/?utm_term=.aeed5bd10a50, sea snake: http://www.livescience.com/53182-venomous-sea-snake-california.html, hammerhead sharks: https://losangeles.cbslocal.com/2015/09/07/why-more-hammerhead-sharks-are-popping-up-off-southern-california-coast/, dolphin fish: http://www.sandiegouniontribune.com/sports/outdoors/sdut-dorado-encinitas-oceanside-fishing-california-2015aug30-story.html, wahoo: http://www.reuters.com/article/us-california-elnino-fish-idUSKCN0T214X20151113, manta ray: http://www.ocregister.com/articles/rays-64054-manta-sighting.html.
References
Abecassis, M., Senina, I., Lehodey, P., Gaspar, P., Parker, D., Balazs, G., et al. (2013). A model of loggerhead sea turtle (Caretta caretta) habitat and movement in the oceanic North Pacific. PLoS One 8:e73274. doi: 10.1371/journal.pone.0073274
Akaike, H. (1973) “Information theory and an extension of the maximum likelihood principle,” in Proceedings of the 2nd International Symposium Information Theory, Supp. to Problems of Control and Information Theory, eds B. N. Petrov and F. Csaki (Budapest: Akademiai Kiado), 267–281.
Alfaro Shigueto, J., Mangel, J. C., Seminoff, J. A., and Dutton, P. H. (2008). Demography of loggerhead turtles Caretta caretta in the southeastern Pacific Ocean: fisheries-based observations and implications for management. End. Sp. Res. 5, 129–135. doi: 10.3354/esr00142
Alfaro-Shigueto, J., Dutton, P. H., Mangel, J., and Vega, D. (2004). First confirmed occurrence of loggerhead turtles in Peru. Mar. Turtle Newsl. 103, 7–11.
Allen, C. D., Lemons, G. E., Eguchi, T., LeRoux, R. A., Fahy, C. C., Dutton, P. H., et al. (2013). Stable isotope analysis reveals migratory origin of loggerhead turtles in the Southern California Bight. Mar. Ecol. Prog. Ser. 472, 275–285. doi: 10.3354/meps10023
ANON (2012). Physical, Chemical and Biological Data CalCOFI Cruise 1110 16 October – 1 November 2011 CC. Reference 12-05. 5 November 2012. Reference 16 – 04 25 Oct 2016. University of California, San Diego, La Jolla, CA: Scripps Institution of Oceanography.
ANON (2016). Physical, Chemical and Biological Data CalCOFI Cruise 1511 25 October – 13 November 2015 CC. Reference 16 – 04. 25 Oct 2016. University of California, San Diego, La Jolla, CA: Scripps Institution of Oceanography.
Barceló, C., Domingo, A., Miller, P., Ortega, L., Giffoni, B., Sales, G., et al. (2013). High-use areas, seasonal movements and dive patterns of juvenile loggerhead sea turtles in the Southwestern Atlantic Ocean. Mar. Eng. Pror. Ser. 479, 235–250. doi: 10.3354/meps10222
Barlow, J. (2016). Cetacean Abundance in the California Current Estimated from Ship-based Line-transect Surveys in 1991-2014. La Jolla, CA: Southwest Fisheries Science Center, Administrative Report, LJ-2016-01, 63.
Barlow, J., and Forney, K. A. (2007). Abundance and population density of cetaceans in the California current ecosystem. Fish Bull. 105, 509–526.
Bartram, P. K., John Kaneko, J., and Kucey-Nakamura, K. (2010). Sea turtle bycatch to fish catch ratios for differentiating Hawaii longline-caught seafood products. Mar. Policy 34, 145–149. doi: 10.1016/j.marpol.2009.05.006
Benson, S. R., Croll, D. A., Marinovic, B. B., Chavez, F. P., and Harvey, J. T. (2002). Changes in the cetacean assemblage of a coastal upwelling ecosystem during El Niño 1997–98 and La Niña 1999. Progr. Oceanogr. 54, 279–291. doi: 10.1016/S0079-6611(02)00054-X
Benson, S. R., Eguchi, T., Foley, D. G., Forney, K. A., Bailey, H., Hitipeuw, C., et al. (2011). Large-scale movements and high-use areas of western Pacific leatherback turtles, Dermochelys coriacea. Ecosphere 2:84. doi: 10.1890/ES11-00053.1
Benson, S. R., Forney, K. A., Harvey, J. T., Carretta, J. V., and Dutton, P. H. (2007). Abundance, distribution, and habitat of leatherback turtles (Dermochelys coriacea) off California, 1990 - 2003. Fish. Bull. 105, 337–347.
Bond, N. A., Cronin, M. F., Freeland, H., and Mantua, N. (2015). Causes and impacts of the 2014 anomaly in the NE Pacific. Geophys. Res. Lett. 42, 3414–3420. doi: 10.1002/2015GL063306
Bowen, B. W., Abreu-Grobois, F. A., Balazs, G. H., Kamezaki, N., Limpus, C. J., and Ferl, R. J. (1995). Trans-Pacific migrations of the loggerhead turtle (Caretta caretta) demonstrated with mitochondrial DNA markers. Proc. Natl. Acad. Sci. U.S.A. 92, 3731–3734. doi: 10.1073/pnas.92.9.3731
Boyle, M. C., FitzSimmons, N. N., Limpus, C. J., Kelez, S., Velez-Zuazo, X., and Waycott, M. (2009). Evidence for transoceanic migrations by loggerhead sea turtles in the southern Pacific Ocean. Proc. R. Soc. B Biol. Sci. 276, 1993–1999. doi: 10.1098/rspb.2008.1931
Briscoe, D. K., Parker, D. M., Bograd, S., Hazen, E., Scales, K., Balazs, G. H., et al. (2016a). Multi-year tracking reveals extensive pelagic phase of juvenile loggerhead sea turtles in the North Pacific. Mov. Ecol. 4:23. doi: 10.1186/s40462-016-0087-4
Briscoe, D. K., Parker, D. M., Balazs, G. H., Kurita, M., Saito, T., Okamoto, H., et al. (2016b). Active dispersal in loggerhead sea turtles (Caretta caretta) during the “lost years”. Proc. R. Soc. B Biol. Sci. 283:20160690. doi: 10.1098/rspb.2016.0690
Buckland, S. T., Anderson, D. R., Burnham, K. P., Laake, J. L., Borchers, D. L., and Thomas, L. (2001). Introduction to Distance Sampling. New York, NY: Oxford University Press.
Carretta, J. V., and Barlow, J. (2011). Long-term effectiveness, failure rates, and “dinner bell” properties of acoustic pingers in a gillnet fishery. Mar. Technol. Soc. J. 45, 7–19. doi: 10.4031/MTSJ.45.5.3
Carretta, J. V., Moore, J. E., and Forney, K. A. (2017). Regression Tree and Ratio Estimates of Marine Mammal, Sea Turtle, and Seabird Bycatch in the California Drift Gillnet Fishery: 1990-2015. Seattle, WA: NOAA Technical Memorandum, NOAA-TM-NMFS-SWFSC-568. 83. doi: 10.7289/V5/TM-SWFSC-568
Carretta, J. V., Price, T., Petersen, D., and Read, R. (2003). Estimates of marine mammal, sea turtle, and seabird mortality in the California drift gillnet fishery for swordfish and thresher shark, 1996 - 2002. Mar. Fish. Rev. 66, 21–30.
Casale, P., Affronte, M., Scaravelli, D., Lazar, B., Vallini, C., and Luschi, P. (2012a). Foraging grounds, movement patterns and habitat connectivity of juvenile loggerhead turtles (Caretta caretta) tracked from the Adriatic Sea. Mar. Biol. 159, 1527–1535. doi: 10.1007/s00227-012-1937-2
Casale, P., Freggi, D., Cinà, A., and Rocco, M. (2012b). Spatio-temporal distribution and migration of adult male loggerhead sea turtles (Caretta caretta) in the Mediterranean sea: further evidence of the importance of neritic habitats off North Africa. Mar. Biol. 160, 703–718. doi: 10.1007/s00227-012-2125-0
Cavole, L. M., Demko, A. M., Diner, R. E., Giddings, A., Kosester, I., Pagniello, C. M. L. S., et al. (2016). Biological impacts of the 2013-2015 warm water anomaly in the Northeast Pacific: winners, losers, and the future. Oceanography 29, 273–285. doi: 10.5670/oceanog.2016.32
Dailey, M. D., Anderson, J. W., Reish, D. J., and Gorsline, D. S. (1993). “The Southern California bight: background and setting,” in Ecology of the Southern California Bight: A Synthesis and Interpretation, eds M. D. Dailey, D. J. Reish, and J. W. Anderson (Los Angeles, CA: University of California Press), 1–18.
Dalleau, M., Benhamou, S., Sudre, J., Ciccione, S., and Bourjea, J. (2014). The spatial ecology of juvenile loggerhead turtles (Caretta caretta) in the Indian Ocean sheds light on the lost years mystery. Mar. Biol. 161, 1835–1849. doi: 10.1007/s00227-014-2465-z
Dodd, C. K. Jr. (1988). Synopsis of the biological data on the loggerhead sea turtle Caretta caretta (Linnaeus 1758). U.S. Fish Wildlife Ser. Biol. Rep. 88:110.
Donoso, M., and Dutton, P. H. (2010). Sea turtle bycatch in the Chilean pelagic longline fishery in the southeastern Pacific: opportunities for conservation. Biol. Cons. 143, 2672–2684. doi: 10.1016/j.biocon.2010.07.011
Eguchi, T. (2002). A method for calculating the effect of a die-off From stranding data. Mar. Mamm. Sci. 18, 698–709. doi: 10.1111/j.1748-7692.2002.tb01067.x
Epperly, S. P., Braun, J., Chester, A. J., Cross, F. A., Merriner, J. V., Tester, P. A., et al. (1996). Beach strandings as an indicator of at-sea mortality of sea turtles. Bull. Mar. Sci. 59, 289–297.
Federal Register (2004). Fisheries off West Coast States and in the Western Pacific; Highly Migratory Species Fisheries; Final Rule. Available at: https://www.gpo.gov/fdsys/pkg/FR-2004-04-07/pdf/04-7247.pdf (accessed November 17, 2017).
Federal Register (2007). Fisheries off West Coast States; Highly Migratory Species Fisheries; Final Rule. Available at: https://www.gpo.gov/fdsys/pkg/FR-2007-06-08/pdf/E7-11124.pdf (accessed November 17, 2017).
Federal Register (2014). Fisheries Off West Coast States; The Highly Migratory Species Fishery; Closure. Available at: https://www.gpo.gov/fdsys/pkg/FR-2014-07-25/pdf/2014-17644.pdf (accessed November 17, 2017).
Federal Register (2015). Fisheries off West Coast States; The Highly Migratory Species Fishery; Closure. Available at: https://www.gpo.gov/fdsys/pkg/FR-2015-06-03/pdf/2015-13601.pdf (accessed November 17, 2017).
Federal Register (2016). Fisheries Off West Coast States; The Highly Migratory Species Fishery; Closure. Available at: https://www.gpo.gov/fdsys/pkg/FR-2016-06-03/pdf/2016-13137.pdf (accessed November 17, 2017).
Finkbeiner, E. M., Wallace, B. P., Moore, J. E., Lewison, R. L., Crowder, L. B., and Read, A. J. (2011). Cumulative estimates of sea turtle bycatch and mortality in USA fisheries between 1990 and 2007. Biol. Conserv. 144, 2719–2727. doi: 10.1016/j.biocon.2011.07.033
Forney, K. A., Barlow, J., and Carretta, J. V. (1995). The abundance of cetaceans in California waters. Part II: aerial surveys in winter and spring of 1991 and 1992. Fish Bull. 93, 15–26.
Gilman, E., Gearhart, J., Price, B., Eckert, S., Milliken, H., Wang, J., et al. (2010). Mitigating sea turtle by-catch in coastal passive net fisheries. Fish Fish. 11, 57–88. doi: 10.1111/j.1467-2979.2009.00342.x
Gilman, E., and Huang, H. W. (2016). Review of effects of pelagic longline hook and bait type on sea turtle catch rate, anatomical hooking position and at-vessel mortality rate. Rev. Fish Biol. Fish. 27, 43–52. doi: 10.1007/s11160-016-9447-9
Gilman, E., Kobayashi, D., Swenarton, T., Brothers, N., Dalzell, P., and Kinan-Kelly, I. (2007). Reducing sea turtle interactions in the Hawaii-based longline swordfish fishery. Biol. Conserv. 139, 19–28. doi: 10.1016/j.biocon.2007.06.002
Halpin, L. R., Seminoff, J. A., and Hanke, G. F. (2018). First photographic evidence of a loggerhead sea turtle (Caretta caretta) in British Columbia. Northwest. Natur. 99, 73–75. doi: 10.1898/NWN17-26.1
Hart, K. M., Lamont, M. M., Fujisaki, I., Tucker, A. D., and Carthy, R. R. (2012). Common coastal foraging areas for loggerheads in the Gulf of Mexico: opportunities for marine conservation. Biol. Conserv. 145, 185–194. doi: 10.1016/j.biocon.2011.10.030
Hart, K. M., Mooreside, P., and Crowder, L. B. (2006). Interpreting the spatio-temporal patterns of sea turtle strandings: going with the flow. Biol. Conserv. 129, 283–290. doi: 10.1016/j.biocon.2005.10.047
Hatase, H., Takai, N., Matsuzawa, Y., Sakamoto, W., Omuta, K., Goto, K., et al. (2002). Size-related differences in feeding habitat use of adult female loggerhead turtles Caretta caretta around Japan determined by stable isotope analyses and satellite telemetry. Mar. Ecol. Prog. Ser. 233, 273–281. doi: 10.3354/meps233273
Hawkes, L. A., Witt, M. J., Broderick, A. C., Coker, J. W., Coyne, M. S., Dodd, M., et al. (2011). Home on the range: spatial ecology of loggerhead turtles in Atlantic waters of the USA. Divers Distrib. 17, 624–640. doi: 10.1111/j.1472-4642.2011.00768.x
Hazen, E. L., Jorgensen, S., Rykaczewski, R. R., Bograd, S. J., Foley, D. G., Jonsen, I. D., et al. (2012). Predicted habitat shifts of Pacific top predators in a changing climate. Nat. Clim. Change 3, 234–238. doi: 10.1038/nclimate1686
Hazen, E. L., Scales, K. L., Maxwell, S. M., Briscoe, D. K., Welch, H., Bograd, S. J., et al. (2018). A dynamic ocean management tool to reduce bycatch and support sustainable fisheries. Sci. Adv. 4:eaar3001.
Hickey, B. M. (1993). “Physical oceanography,” in Ecology of the Southern California Bight, eds M. D. Dailey, D. J. Reish, and J. W. Anderson (Los Angeles, CA: University of California Press), 19–70.
Hodge, R. P., and Wing, B. L. (2000). Occurrences of marine turtles in Alaska waters: 1960-1998. Herpetol. Rev. 31, 148–151.
Howell, E. A., Dutton, P. H., Polovina, J. J., Bailey, H., Parker, D. M., and Balazs, G. H. (2010). Oceanographic influences on the dive behavior of juvenile loggerhead turtles (Caretta caretta) in the North Pacific Ocean. Mar. Biol. 157, 1011–1026. doi: 10.1007/s00227-009-1381-0
Howell, E. A., Hoover, A., Benson, S. R., Bailey, H., Polovina, J. J., Seminoff, J. A., et al. (2015). Enhancing the TurtleWatch product for leatherback sea turtles, a dynamic habitat model for ecosystem-based management. Fish. Oceanogr. 24, 57–68. doi: 10.1111/fog.12092
Howell, E. A., Kobayashi, D. R., Parker, D. M., Balazs, G. H., and Polovina, J. J. (2008). TurtleWatch: a tool to aid in the bycatch reduction of loggerhead turtles Caretta caretta in the Hawaii-based pelagic longline fishery. End. Sp. Res. 5, 267–278. doi: 10.3354/esr00096
IPCC (2013). Climate Change 2013. Available at: https://www.ipcc.ch/
Jackson, A., Gerrodette, T., Chivers, S., Lynn, M., Rankin, S., and Mesnick, S. (2008). Marine Mammal Data Collected During a Survey in the Eastern Tropical Pacific Ocean Aboard NOAA Ships David Starr Jordan and McArthur II, July 28 – December 7, 2006. La Jolla, CA: U.S. Dep. Commer. NOAA Technical Memorandum NMFS-SWFSC-421, 45.
Kamezaki, N., Matsuzawa, Y., Abe, O., Asakawa, H., Fujii, T., Goto, K., et al. (2003) “Loggerhead turtles nesting in Japan,” in Loggerhead Sea Turtles, eds A. B. Bolten and B. E. Witherington (Washington, DC: Smithsonian Books), 210–217.
Kobayashi, D. R., Polovina, J. J., Parker, D. M., Kamezaki, N., Cheng, I.-J., Uchida, I., et al. (2008). Pelagic habitat characterization of loggerhead sea turtles, Caretta caretta, in the North Pacific Ocean (1997–2006): insights from satellite tag tracking and remotely sensed data. J. Exp. Mar. Biol. Ecol. 356, 96–114. doi: 10.1016/j.jembe.2007.12.019
Koch, V., Nichols, W. J., Peckham, H., and de la Toba, V. (2006). Estimates of sea turtle mortality from poaching and bycatch in Bahía Magdalena, Baja California Sur, Mexico. Biol. Conserv. 128, 327–334. doi: 10.1016/j.biocon.2005.09.038
Koch, V., Peckham, H., Mancini, A., and Eguchi, T. (2013). Estimating at-sea mortality of marine turtles from stranding frequencies and drifter experiments. PLoS One 8:e56776. doi: 10.1371/journal.pone.0056776
Laake, J., Borchers, D., Thomas, L., Miller, D., and Bishop, J. (2016). mrds: Mark-Recapture Distance Sampling. R Package Version 2.1.17. Available at: https://CRAN.R-project.org/package=mrds
Leising, A. W., Schroeder, I. D., Bograd, S. J., Abell, J., Durazo, R., Barceló, C., et al. (2015). State of the California Current 2014 – 15: impacts of the warm-water “Blob”. CalCOFI Rep. 56, 31–68.
Leising, A. W., Schroeder, I. D., Bograd, S. J., Bjorkstedt, E. P., Field, J., Sakuma, K., et al. (2014). State of the California Current 2013 – 14: El Niño looming. CalCOFI Rep. 55, 51–87.
Lerczak, J. A., and Hobbs, R. C. (1998). Calculating sighting distances from angular readings during shipboard, aerial, and shore-based marine mammal surveys. Mar. Mamm. Sci. 14, 590–599. doi: 10.1111/j.1748-7692.1998.tb00745.x
Lynn, R. J., and Simpson, J. J. (1987). The California current system: the seasonal variability of its physical characteristics. J. Geophys. Res. 92, 947–912. doi: 10.1029/JC092iC12p12947
Lynn, R. J., and Simpson, J. J. (1990). The flow of the undercurrent over the continental borderland off southern California. J. Geophys. Res. 95, 12995–13008. doi: 10.1029/JC095iC08p12995
Mansfield, K. L., and Putman, N. (2013). “Oceanic habits and habitats, Caretta caretta,” in Biology of Sea Turtles, Vol. 3, eds J. Wyneken, K. J. Lohmann and J. A. Musick (Boca Raton, FL: CRC Press), 189–210.
Mansfield, K. L., Saba, V. S., Keinath, J. A., and Musick, J. A. (2009). Satellite tracking reveals a dichotomy in migration strategies among juvenile loggerhead turtles in the Northwest Atlantic. Mar. Biol. 156, 2555–2570. doi: 10.1007/s00227-009-1279-x
Mantua, N. J., Hare, S. R., Zhang, Y., Wallace, J. M., and Francis, R. C. (1997). A Pacific interdecadal climate oscillation with impacts on salmon production. Bull. Am. Meteorol. Soc. 78, 1069–1079. doi: 10.1175/1520-0477(1997)078<1069:APICOW>2.0.CO;2
Matsuzawa, Y. (2011). Nesting Beach Management in Japan to Conserve Eggs and Pre-Emergent Hatchlings of the North Pacific Loggerhead Sea Turtle. Honolulu, HI: Report to NOAA Western Pacific Regional Fishery Management Councilo.
McClatchie, S. (2014). Regional Fisheries Oceanography of the California Current System: The CalCOFI Program. New York, NY: Springer. doi: 10.1007/978-94-007-7223-6
National Marine Fisheries Service [NMFS] (2000). Biological Opinion. Endangered Species Act Section 7 Consultation on Authorization to Take Listed Marine Mammals Inciden- Tal To Commercial Fishing Operations Under Section 101(a)(5)(E) of the Marine Mammal Protection Act for the California/Oregon Drift Gillnet Fishery. Available at: http://swr.nmfs.noaa.gov/bo/lb/Drift_gillnet_BO_2000.pdf
Nichols, W. J., Resendiz, A., Seminoff, J. A., and Resendiz, B. (2000). Transpacific migration of a loggerhead turtle monitored by satellite telemetry. Bull. Mar. Sci. 67, 937–947.
Peckham, S. H., Maldonado Diaz, D., Walli, A., Ruiz, G., Crowder, L. B., and Nichols, W. J. (2007). Small-scale fisheries bycatch jeopardizes endangered Pacific loggerhead turtles. PLoS One 2:e1041. doi: 10.1371/journal.pone.0001041
Peckham, S. H., Maldonado-Diaz, D., Tremblay, Y., Ochoa, R., Polovina, J., Balazs, G., et al. (2011). Demographic implications of alternative foraging strategies in juvenile loggerhead turtles Caretta caretta of the North Pacific Ocean. Mar. Ecol. Prog. Ser. 425, 269–280. doi: 10.3354/meps08995
Peckham, S. H., Maldonado-Diaz, D., Koch, V., Mancini, A., Gaos, A., Tinker, M. T., et al. (2008). High mortality of loggerhead turtles due to bycatch, human consumption and strandings at Baja California Sur, Mexico, 2003 to 2007. End. Sp. Res. 5, 171–183. doi: 10.3354/esr00123
Polovina, J. J., Howell, E., Parker D. M., and Balazs, G. H. (2003). Dive-depth distribution of loggerhead (Carretta carretta) and olive ridley (Lepidochelys olivacea) sea turtles in the central North Pacific: Might deep longline sets catch fewer turtles? Fish. Bull. 101, 189–193.
Polovina, J., Uchida, I., Balazs, G., Howell, E. A., Parker, D., and Dutton, P. (2006). The Kuroshio extension current bifurcation region: a pelagic hotspot for juvenile loggerhead sea turtles. Deep Res. II 53, 326–339. doi: 10.1016/j.dsr2.2006.01.006
Polovina, J. J., Kobayashi, D. R., Parker, D. M., Seki, M. P., and Balazs, G. H. (2000). Turtles on the edge: movement of loggerhead turtles (Caretta caretta) along oceanic fronts, spanning longline fishing grounds in the central North Pacific, 1997-1998. Fish. Oceanogr. 9, 71–82. doi: 10.1046/j.1365-2419.2000.00123.x
Pörtner, H. O., and Knust, R. (2007). Climate change affects marine fishes through the oxygen limitatoin of thermal tolerance. Science 315, 95–97. doi: 10.1126/science.1135471
R Development Core Team (2016) R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Resendiz, A., Resendiz, B., Nichols, W. J., Seminoff, J. A., and Kamezaki, N. (1998). First confirmed east-west transpacific movement of a loggerhead sea turtle, Caretta caretta, released in Baja California, Mexico. Pacific Sci. 52, 151–153.
Sakuma, K. M., Field, J. C., Mantua, N. J., Ralston, S., Marinovic, B. B., and Carrion, C. N. (2016). Anomalous epipelagic micronekton assemblage patterns in the neritic waters of the California current in spring 2015 during a period of extreme ocean conditions. CalCOFI Rep. 57, 163–183.
Santora, J. A., Hazen, E. L., Schroeder, I. D., Bograd, S. J., Sakuma, K. M., and Field, J. C. (2017). Impacts of ocean climate variability on biodiversity of pelagic forage species in an upwelling ecosystem. Mar. Ecol. Prog. Ser. 580, 205–220. doi: 10.3354/meps12278
Sasso, C. R., Braun-Mcneill, J., Avens, L., and Epperly, S. P. (2006). Effects of transients on estimating survival and population growth in juvenile loggerhead turtles. Mar. Ecol. Prog. Ser. 324, 287–292. doi: 10.3354/meps324287
Seminoff, J. A., Eguchi, T., Carretta, J., Allen, C. D., Prosperi, D., Rangel, R., et al. (2014). Loggerhead sea turtle abundance at a foraging hotspot in the eastern Pacific Ocean: implications for at-sea conservation. End. Sp. Res. 24, 207–220. doi: 10.3354/esr00601
Senko, J., Jenkins, L. D., and Peckham, S. H. (2017). At loggerheads over international bycatch: initial effects of a unilaterally imposed bycatch reduction policy. Mar. Policy 76, 200–209. doi: 10.1016/j.marpol.2016.11.017
Silverman, B. W. (1986). Density Estimation. London: Chapman and Hall. doi: 10.1007/978-1-4899-3324-9
Turner Tomaszewicz, C. N., Seminoff, J. A., Avens, L., Goshe, L. R., Peckham, S. H., Rguez-Baron, J. M., et al. (2015). Age and residency duration of loggerhead turtles at a North Pacific bycatch hotspot using skeletochronology. Biol. Conserv. 186, 134–142. doi: 10.1016/j.biocon.2015.03.015
Wernberg, T., Smale, D. A., Tuya, F., Thomsen, M. S., Langlois, T. J., de Bettignies, T., et al. (2012). An extreme climatic event alters marine ecosystem structure in a global biodiversity hotspot. Nat. Clim. Change 3, 78–82. doi: 10.1038/nclimate1627
Keywords: distance sampling, Eastern Pacific, El Niño Southern Oscillation, line transect, stranding
Citation: Eguchi T, McClatchie S, Wilson C, Benson SR, LeRoux RA and Seminoff JA (2018) Loggerhead Turtles (Caretta caretta) in the California Current: Abundance, Distribution, and Anomalous Warming of the North Pacific. Front. Mar. Sci. 5:452. doi: 10.3389/fmars.2018.00452
Received: 19 June 2018; Accepted: 12 November 2018;
Published: 06 December 2018.
Edited by:
Jeremy Kiszka, Florida International University, United StatesReviewed by:
Gail Schofield, Queen Mary University of London, United KingdomJonathan D. R. Houghton, Queen’s University Belfast, United Kingdom
Copyright © 2018 Eguchi, McClatchie, Wilson, Benson, LeRoux and Seminoff. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tomoharu Eguchi, dG9tby5lZ3VjaGlAbm9hYS5nb3Y=