 Dalia C. Barragán-Barrera1,2*
Dalia C. Barragán-Barrera1,2* Karina Bohrer do Amaral3*
Karina Bohrer do Amaral3* Paula Alejandra Chávez-Carreño4Nohelia Farías-Curtidor1Rocío Lancheros-Neva1,5Natalia Botero-Acosta1Paula Bueno6
Paula Alejandra Chávez-Carreño4Nohelia Farías-Curtidor1Rocío Lancheros-Neva1,5Natalia Botero-Acosta1Paula Bueno6 Ignacio Benites Moreno3,7Jaime Bolaños-Jiménez8,9Laurent Bouveret10Delma Nataly Castelblanco-Martínez11,12Jolanda A. Luksenburg13,14Julie Mellinger10
Ignacio Benites Moreno3,7Jaime Bolaños-Jiménez8,9Laurent Bouveret10Delma Nataly Castelblanco-Martínez11,12Jolanda A. Luksenburg13,14Julie Mellinger10 Roosevelt Mesa-Gutiérrez15
Roosevelt Mesa-Gutiérrez15 Benjamin de Montgolfier16
Benjamin de Montgolfier16 Eric A. Ramos12,17Vincent Ridoux18
Eric A. Ramos12,17Vincent Ridoux18 Daniel M. Palacios19*
Daniel M. Palacios19*- 1Fundación Macuáticos Colombia, Medellín, Colombia
- 2Laboratorio de Ecología Molecular de Vertebrados Acuáticos, Department of Biological Sciences, Universidad de los Andes, Bogotá, Colombia
- 3Laboratório de Sistemática e Ecologia de Aves e Mamíferos Marinhos, Instituto de Biociências, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil
- 4Independent Researcher, La Paz, Mexico
- 5Parques Nacionales Naturales de Colombia, Bogotá, Colombia
- 6WWF Colombia, Bogotá, Colombia
- 7Centro de Estudos Costeiros, Limnológicos e Marinhos, Universidade Federal do Rio Grande do Sul, Imbé, Brazil
- 8Asociación Civil Sea Vida, Aragua, Venezuela
- 9Instituto de Ciencias Marinas y Pesquerías, Universidad Veracruzana, Veracruz, Mexico
- 10Observatoire des Mammifères Marins de l’Archipel Guadeloupéen, Archipelago’s Guadeloupe, France
- 11Consejo Nacional de Ciencia y Tecnología, University of Quintana Roo, Chetumal, Mexico
- 12Fundación Internacional para la Naturaleza y la Sostenibilidad, Chetumal, Mexico
- 13Institute of Environmental Sciences, Leiden University, Leiden, Netherlands
- 14Department of Environmental Science and Policy, George Mason University, Fairfax, VA, United States
- 15Duke Marine Laboratory, Nicholas School of the Environment, Duke University, Durham, NC, United States
- 16Aquasearch, Sainte-Luce, France
- 17The Graduate Center, City University of New York, New York, NY, United States
- 18Observatoire PELAGIS, Université de La Rochelle, La Rochelle, France
- 19Marine Mammal Institute and Department of Fisheries and Wildlife, Oregon State University, Newport, OR, United States
Dolphins of the genus Stenella occur in pelagic waters of both tropical and warm-temperate oceans. Three species, the Atlantic spotted dolphin (Stenella frontalis), the pantropical spotted dolphin (S. attenuata), and the spinner dolphin (S. longirostris) are abundant worldwide, but in the Caribbean Basin they have been poorly studied and information on their distribution patterns is scarce. Specifically, in Colombia’s remote Seaflower Biosphere Reserve (SFBR) S. attenuata has been reported occasionally, but S. frontalis and S. longirostris have never been recorded before. To address this information gap, an ecological niche modeling approach was used to determine the potential distribution patterns of these three dolphin species in the region. Records of these species for the Caribbean Basin were compiled, including both published and unpublished data. Environmental information, including bathymetry, bathymetric slope, distance to shore, sea surface temperature, sea surface salinity, and chlorophyll-a concentration was gathered from public databases (MARSPEC and Bio-ORACLE) in raster format. The maximum entropy algorithm (Maxent) for modeling species’ geographic distributions with presence-only data was used. After filtering the data, 210 records of S. attenuata, 204 of S. frontalis, and 80 of S. longirostris were used to run models. The best configuration for each model was chosen based on the ΔAICc criterion. For all three species, the final ecological niche models returned AUC test values higher than 0.8, indicating satisfactory model performance. The resulting potential distribution maps suggested that areas closest to continental shorelines of the Caribbean Basin and surrounding islands had the highest environmental suitability for all species (>70%). All models reported high environmental suitability for S. attenuata and S. longirostris in the SFBR, mainly in the southernmost part surrounding San Andrés and Providence Archipelago. Assessment of niche overlap from the predictions of species distributions using the similarity statistic and pairwise map overlap indicated that S. frontalis and S. longirostris had niches slightly more similar in comparison to S. attenuata. As this was a first effort to fill a gap in our understanding of the distribution of species in the genus Stenella in the Caribbean Basin, further studies are necessary using both niche modeling and biological/ecological approaches.
Introduction
The ocean comprises 70% of the planet and offers a wide range of habitats to marine life (e.g., Spalding et al., 2012; Kelley et al., 2016). These different habitats are characterized by gradients in sea surface temperature (SST), sea surface salinity (SSS), productivity, and topography at both broad and local scales (Redfern et al., 2006), playing an important role to explain distribution patterns of marine top predators such as cetaceans (e.g., Baumgartner et al., 2001; Palacios et al., 2013b). Dolphins of the genus Stenella (Gray, 1866) are common in tropical, subtropical, and temperate waters worldwide. Because of such wide distribution patterns, they are often considered “umbrella species,” since any conservation efforts made on their behalf benefit many other species within their habitat (Jefferson et al., 2008).
Although the taxonomy of Stenella remained unclear until the 1980s, the five species that make up the genus are now well characterized. The genus is considered a non-monophyletic assemblage (see Perrin et al., 2013), with some species being more related to Tursiops (Leduc et al., 1999). While the Stenella species have great dispersal capacity, they exhibit significant distribution changes in response to seasonal variation in oceanographic conditions (Reilly, 1990; Moreno et al., 2005). Additionally, some species within the genus are considered sympatric throughout their distribution, although at an individual level they appear to have distinct environmental requirements at regional scales (e.g., Baumgartner et al., 2001; Davis et al., 2002; do Amaral et al., 2015).
The Atlantic spotted dolphin (S. frontalis), endemic to the Atlantic Ocean, is distributed mainly in areas closer to continental shelves and surrounding islands (Perrin et al., 1987; Davis et al., 1998; do Amaral et al., 2015). The pantropical spotted dolphin (S. attenuata) and the spinner dolphin (S. longirostris) are more widely distributed in the Atlantic, Indian, and Pacific Oceans (Jefferson et al., 2008), and appear to have similar patterns of habitat use that favor their presence over deeper waters (Davis et al., 1998, 2002; Baumgartner et al., 2001; do Amaral et al., 2015). Although these three species have been previously reported in the Caribbean Basin (e.g., Caldwell et al., 1971; Mignucci-Giannoni, 1998; Pardo et al., 2009; Palacios et al., 2013b; Niño-Torres et al., 2015; Ramos et al., 2016), little is known about their habitat preferences and spatial distribution in coastal or oceanic areas. Hence, it is important to identify areas where these species are likely to be found in order to direct research and improve management measures.
Particularly in Colombia’s Caribbean Sea, cetacean research has been limited, with most studies focusing on specific coastal mainland locations, including Santa Marta (Pardo and Palacios, 2006; Fraija et al., 2009; Pardo et al., 2009), Bay of Cispatá (García and Trujillo, 2004), Gulf of Morrosquillo (Palacios et al., 2013b), and La Guajira (Combatt and González, 2007; Palacios et al., 2012; Farías-Curtidor et al., 2017). To date, almost no research effort has been conducted in the remote oceanic region surrounding the Archipelago of San Andrés, Providencia and Santa Catalina (Pardo et al., 2009; Palacios et al., 2013b). Located in this region, the Seaflower Biosphere Reserve (SFBR) is one of the largest marine reserves in the western hemisphere, containing the most extensive open-ocean coral reefs in the Caribbean Sea (Mow et al., 2007; Coralina-Invemar, 2012). This unique marine ecosystem is highly biodiverse (Taylor et al., 2013) and may be of importance for cetaceans. However, given the remoteness of the region and the prevalence of windy conditions, the development of systematic visual surveys necessary to assess cetacean abundance and distribution has been logistically and financially challenging.
To date, at least 22 species of marine mammals have been confirmed to occur in the Colombian Caribbean, including 18 odontocetes (Palacios et al., 2013b). Within the SFBR, pantropical spotted dolphins, bottlenose dolphins (Tursiops truncatus), false killer whales (Pseudorca crassidens), and sperm whales (Physeter macrocephalus) are all confirmed species (Pardo et al., 2009; Barragán-Barrera et al., 2017). Stranding reports in the San Andrés Archipelago also indicate the presence of pygmy or dwarf sperm whales (Kogia sp.) (Palacios et al., 2013b) and Cuvier’s beaked whales (Ziphius cavirostris) (Prieto-Rodríguez, 1988; Vidal, 1990). However, the area can potentially host several other species, including Atlantic spotted dolphins, spinner dolphins, striped dolphins (S. coeruleoalba), short-finned pilot whales (Globicephala macrorhynchus), killer whales (Orcinus orca), and even humpback whales (Megaptera novaeangliae) (Pardo et al., 2009; Palacios et al., 2013b; Bolaños-Jiménez et al., 2014).
Ecological niche modeling is a widely used tool to predict cetaceans’ distribution and understand the ecological precursors (Redfern et al., 2006; Gregr et al., 2013; Palacios et al., 2013a). These models can also be implemented to assess areas of high conservation priority and identify key areas for research on interactions between humans and wildlife (Bailey and Thompson, 2009). For the Caribbean Basin, the use of advanced geospatial analysis tools has the potential to improve the current comprehension of cetacean distribution patterns and the environmental factors influencing them. However, these analyses have never been conducted in the region, despite some studies being developed in the adjacent Gulf of Mexico (e.g., Davis et al., 1998, 2002; Baumgartner et al., 2001) and the Western South Atlantic Ocean (e.g., do Amaral et al., 2015). Since 2014, Colombia has been carrying out scientific cruises in the SFBR to assess the local biodiversity of the islands (Murillo, 2014). Despite cetaceans having been taken into account in these efforts, in practice the Colombian Program of Marine Fauna Observers has been the main source of this type of information in recent times (Palacios et al., 2013b). Research efforts could be optimized if uncertainty regarding cetacean occurrence and distribution in the Reserve is overcome.
With the aim of determining the potential distributions of spotted and spinner dolphins throughout the Caribbean Basin, and more specifically within the SFBR, we adopted an ecological niche modeling approach. Because absence data are generally not available for oceanic dolphins and the offshore limits of their distributions are unknown, we used a presence-only method to predict the potential distribution of these species (Robinson et al., 2017). The chosen method was maximum entropy (Phillips et al., 2006), which has been applied successfully on species with limited data (Wisz et al., 2008; Elith et al., 2011). Furthermore, this method has been successfully used with cetaceans (Derville et al., 2018), and has already been used to predict the potential distribution of Stenella species in the Western South Atlantic (do Amaral et al., 2015). We hypothesized that these species have different distribution patterns as a consequence of distinct environmental requirements, as has been found in adjacent regions such as the Gulf of Mexico and the Western South Atlantic. In addition to overcoming the prevailing information gaps, our results could contribute to the understanding of the effectiveness of the SFBR in protecting top predators such as dolphins, and to identify other areas that deserve protection throughout the Caribbean Basin.
Materials and Methods
Study Area
The geographical extent of the models and the background sampling was restricted to the Caribbean Basin (59°24′W–89°21′W and 7°42′N–22°41′N). The Caribbean is a semi-enclosed tropical sea adjacent to the Atlantic Ocean (Müller-Karger et al., 1989; Aguirre, 2014). Although the area is influenced by the discharge of the Amazon and Orinoco rivers, it is generally considered oligotrophic and homogeneous despite seasonal upwelling events (Müller-Karger et al., 1989; Criales-Hernández et al., 2006; Arévalo-Martínez and Franco-Herrera, 2008; Chollett et al., 2012; Rueda-Roa and Muller-Karger, 2013), and the influence of the warm Caribbean Current (Aguirre, 2014).
Within the Caribbean Basin, the SFBR is located in oceanic waters of the Colombian Exclusive Economic Zone (80°39′W–82°17′W and 11°56′N–13°55′N) (Coralina-Invemar, 2012) (Figure 1). The Reserve was created as the first Colombian Marine Protected Area and was declared as a UNESCO Biosphere Reserve in November 2000 (Taylor et al., 2013). It has an area of around 65,000 km2 and encompasses the San Andrés, Providencia and Santa Catalina Archipelago (Supplementary Figure S1) (Taylor et al., 2013). The SFBR is a highly biodiverse region, containing more than 77% of the shallow coral reefs of Colombia, as well as other coastal/marine ecosystems such as atolls, beaches, sandy bottoms, mangroves, and seagrass beds (Coralina-Invemar, 2012; Taylor et al., 2013).
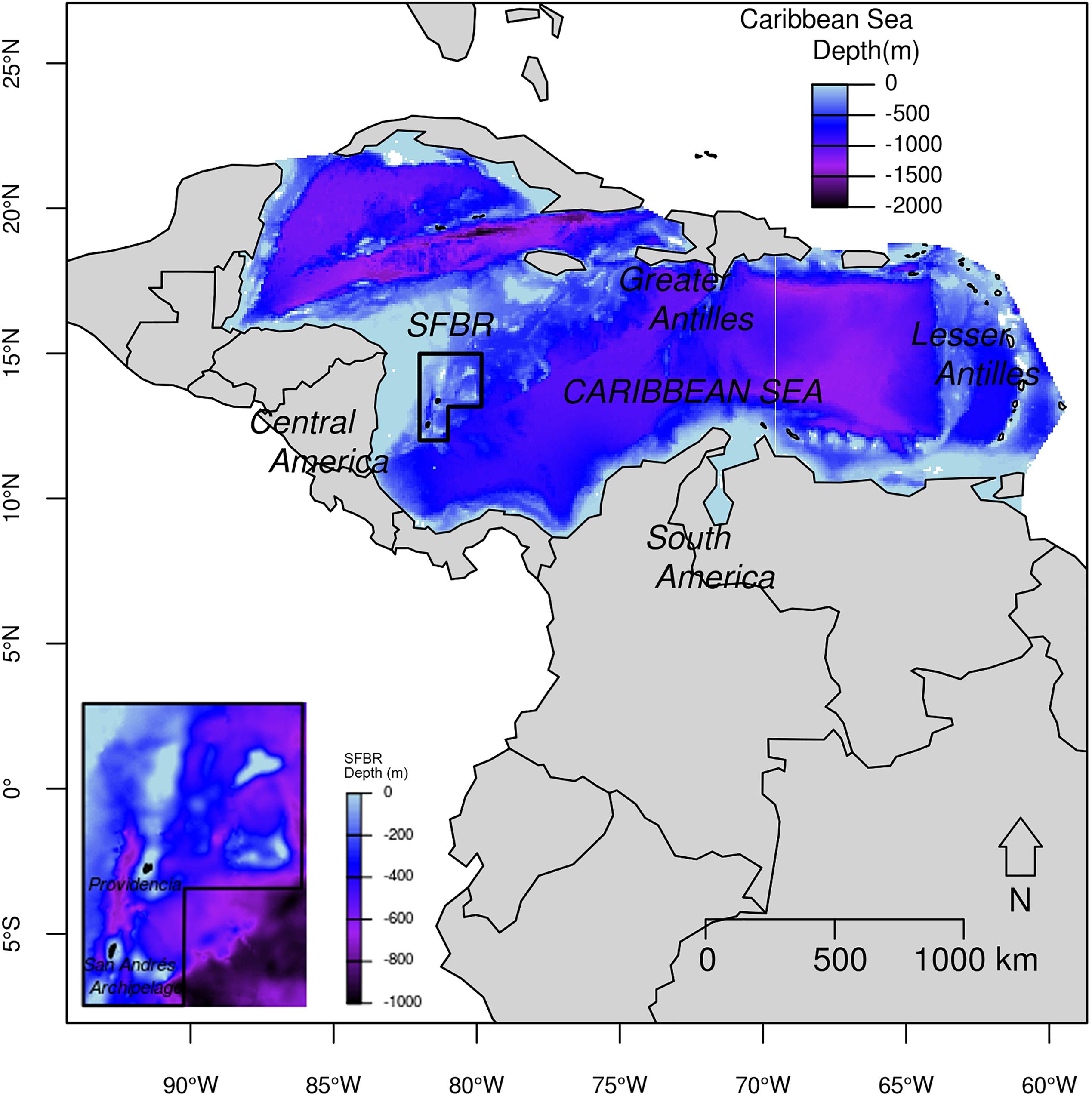
Figure 1. Study area with the location of Seaflower Biosphere Reserve (polygon in bold black line). Inset provides a zoom in the Seaflower Biosphere Reserve and indicates the main islands.
Environmental Data
Environmental layers used in the modeling were selected after reviewing previous cetacean niche modeling studies (e.g., Baumgartner et al., 2001; Cañadas et al., 2002; Davis et al., 2002; do Amaral et al., 2015; Tobeña et al., 2016; Gomez et al., 2017). The selected layers included both static and dynamic variables for all three species (Table 1). Dynamic variables used in this study included SST, chlorophyll-a concentration (Chl-a), diffuse attenuation (Da), and SSS. Static layers included bathymetry, slope and distance to shore (Table 1). Environmental layers representing SST, SSS and Chl-a were included as two different metrics: the annual mean and the annual range (the differences between annual maximum and minimum). For Da, only the annual mean was included because annual range was not available.
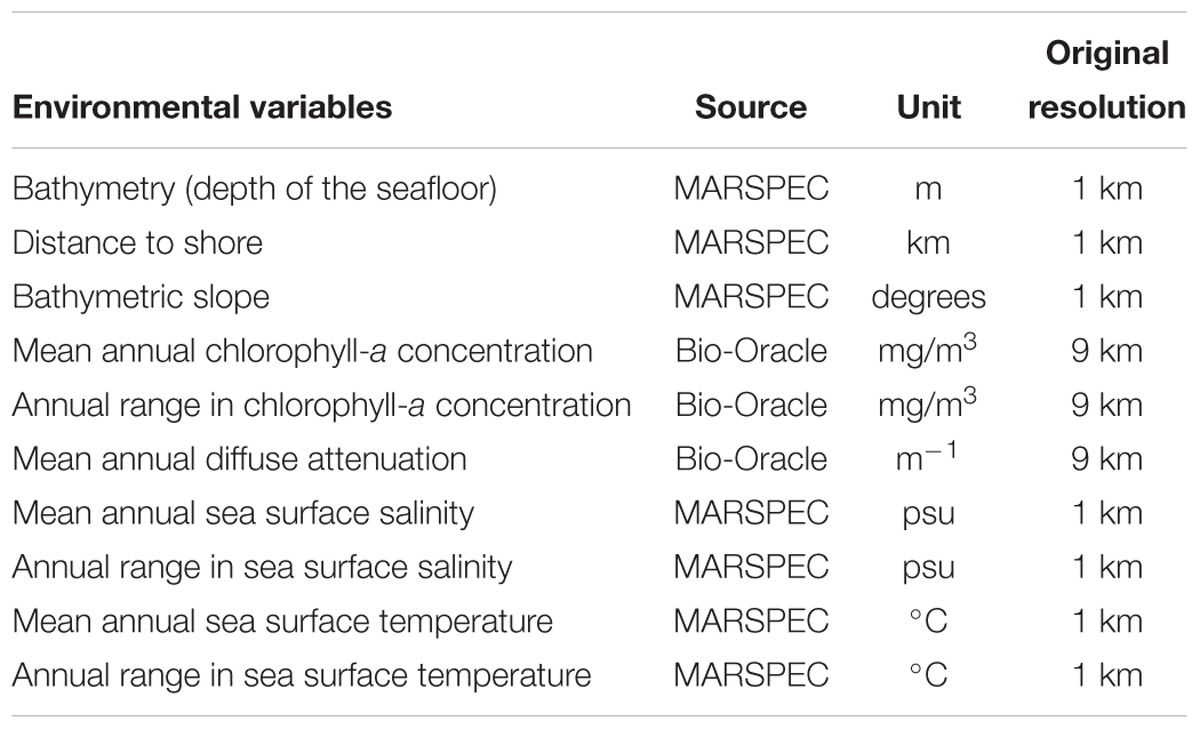
Table 1. List of environmental variables used in this study and their respective source, original grid resolution, and unit of measurement.
Raster layers of dynamic and static information data were gathered from the public database Bio-Oracle (Oceans Rasters for Analysis of Climate and Environment) (Tyberghein et al., 2012) and MARSPEC (Ocean Climate Layers from Marine Spatial Ecology) (Sbrocco and Barber, 2013). While the Bio-Oracle public database carried raster layers of Chl-a and Da at the 9.2 km resolution, the remaining layers were available in the MARSPEC public database at 1 km resolution. All layers were cropped to encompass the study area and adjusted to a common resolution of 9.2 km per grid cell using the raster package (Hijmans, 2017) in R 3.4.2 (R Core Team, 2018). The correlation among layers was investigated using the function pairs in the raster package. Only layers with correlation coefficient below 0.7 were included in the ecological niche models (Dormann et al., 2013) (Supplementary Figure S2). A principal component analysis (PCA) was performed to visualize the environmental heterogeneity across the study area using the rasterPCA function from Rstoolbox package (Leutner and Horning, 2016) (Supplementary Figure S3).
Occurrence Data
Georeferenced sighting data for Atlantic spotted dolphins, pantropical spotted dolphins, and spinner dolphins occurring between 1981 and 2016 in the Caribbean Basin, were compiled from several sources: published records, unpublished data, and the Sistema de Información Ambiental Marina (SIAM1) operated by the Instituto de Investigaciones Marinas y Costeras (INVEMAR, Colombia’s Marine and Coastal Research Institute). These occurrence data were collected through different seasons and years, during both systematic and opportunistic marine wildlife observations. Only confirmed sightings correctly identified at the species level as Stenella were included.
After scrutinizing, filtering, cleaning duplicate records and removing some of the bias by subsampling records (see Hijmans and Elith, 2017), a total of 494 records were included: 210 sightings of pantropical spotted dolphins, 204 Atlantic spotted dolphins, and 80 spinner dolphins (Table 2 and Figure 2). The dataset used for this study is presence-only, meaning that absences (such as survey track lines) were not considered (Franklin, 2009). A subset of records (20%) was used as testing data as explained below, and the remaining was used as training data.
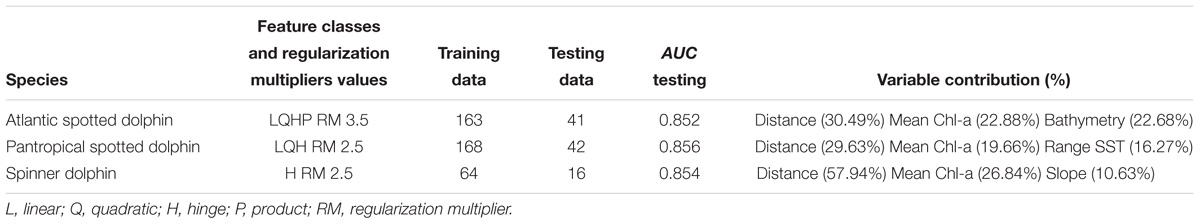
Table 2. The ecological niche models with the best configuration for each species, the number of training and testing data, the AUC testing values and the relative contributions of the major environmental variables to the Maxent models with dynamic and static variables.
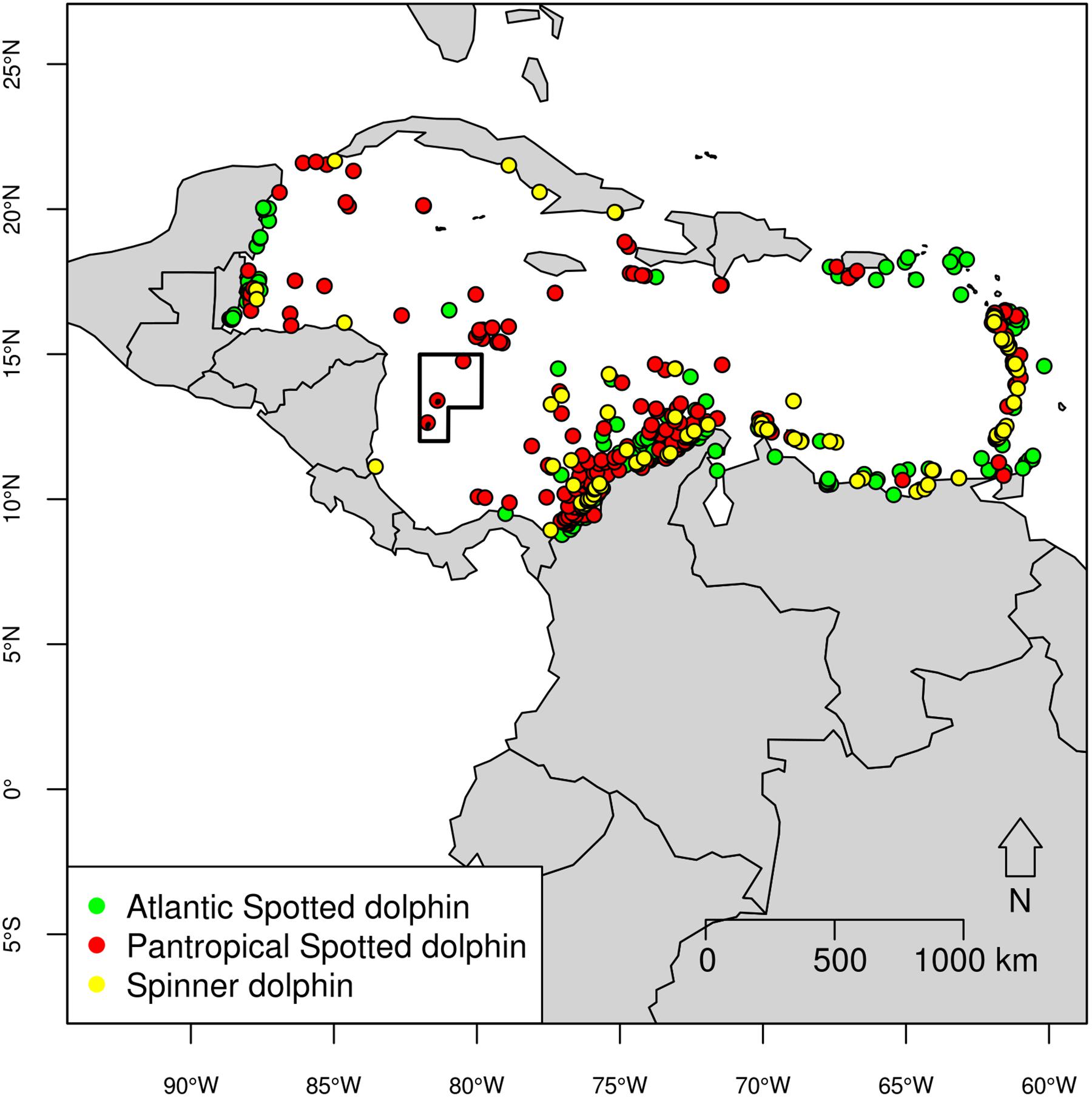
Figure 2. Species records in the Caribbean Basin used as both training and testing data in the ecological niche models for Atlantic spotted dolphin (green, n = 204), pantropical spotted dolphin (red, n = 210), three of which occurred inside the Seaflower Biosphere Reserve, and spinner dolphin (yellow, n = 80).
Ecological Niche Modeling
We used a presence-only approach to model the distribution of Stenella species in the Caribbean Basin. Specifically, we used the maximum entropy (Maxent) algorithm (Phillips et al., 2017) to model the potential distribution of the three species. Maxent estimates the geographic range of a species by finding the distribution that has the maximum entropy constrained by the environmental conditions at recorded occurrence locations (Phillips et al., 2017). Maxent predicts environmental suitability, with better conditions represented by higher values (Phillips et al., 2006). This method tends to predict the largest possible suitable area consistent with the data (Merow et al., 2013). While careful consideration of the limitations and biases in the data must be taken, Maxent performs well when compared to presence-absence models (Derville et al., 2018).
In order to build the Maxent species distribution models, we used the Maxent function available within the dismo version 1.1-4 package in R (Hijmans et al., 2017). Maxent model settings were defined through the ENMevaluate function of the ENMeval package (Muscarella et al., 2014), which provided species-specific settings to generate models, such as feature classes and standardized multiplier values. The model with the lowest value of the Akaike’s Information Criterion corrected for small samples sizes (AICc) reflects both model’s goodness-of-fit and complexity (Muscarella et al., 2014). We used the block as data partitioning method, which splits the data into four bins based on lines of latitude and longitude to divide occurrence localities as equally as possible (Muscarella et al., 2014). For model validation, we adopted the values of the area under the receiver-operator (ROC) curve (AUC) as indicators of the predictive skill of the model (Phillips et al., 2006), where AUC closest to a value of 1 would be a perfect model and AUC = 0.5 would indicate that the model performed no better than random.
Twenty percent of the sightings for each species were randomly selected as test data sets to evaluate the fit of the model and its predictive power through the evaluate function in the dismo package. Maps were exported in raster format with continuous values and were interpreted from a percentage of environmental suitability ranging from unsuitable (0%) to highly suitable (100%). For visualization purposes of the potential distributions of these three species across the study area in a binary map of presence/absence, these were built based on the threshold of equal test sensitivity and specificity.
Statistical Comparisons of Distribution Patterns
We tested the null hypothesis that each species had similar distributions of the environmental variables using the non-parametric Kruskal–Wallis test followed by the Dunn test available in the dunn package (Dinno, 2017). The range and distributional properties of the environmental layers where species were recorded were represented as boxplots.
Comparative similarity measures among Stenella species and the niche identity test implemented in the dismo package were computed. Specifically, we estimated the “D” and “I” similarity statistics of Warren et al. (2008) using the functions niche overlap from predictions of species’ distributions, where the resulting values ranges from 0 (no overlap) to 1 (the distributions are identical). We tested the null hypothesis that the ecological niche models of each species pair were identical using the function nicheEquivalency based on 499 randomizations. To visualize the degree of spatial overlap between the Stenella species distributions, we overlapped binary maps for the three pairwise combinations of species using the crop function in the raster package.
Results
Ecological Niche Modeling
Ecological niche models were built based on seven uncorrelated environmental layers: bathymetry, distance to shore, slope, Mean SSS, Mean SST and Mean Chl-a, as well as Range SST. In total, 887 records of Stenella species were compiled (Supplementary Table S1), and after data cleaning 494 were kept (Table 2 and Figure 2).
The model with the best configuration for each species is presented in Table 2. Ecological niche models incorporating dynamic and static variables returned AUC test values higher than 0.85 for all three species, indicating that these models had a good performance. In general, distance to shore and Mean Chl-a were the environmental variables that contributed the most within all models (Table 2). The potential geographic range of distribution for each species, as predicted by the best models, is shown in Figures 3–5.
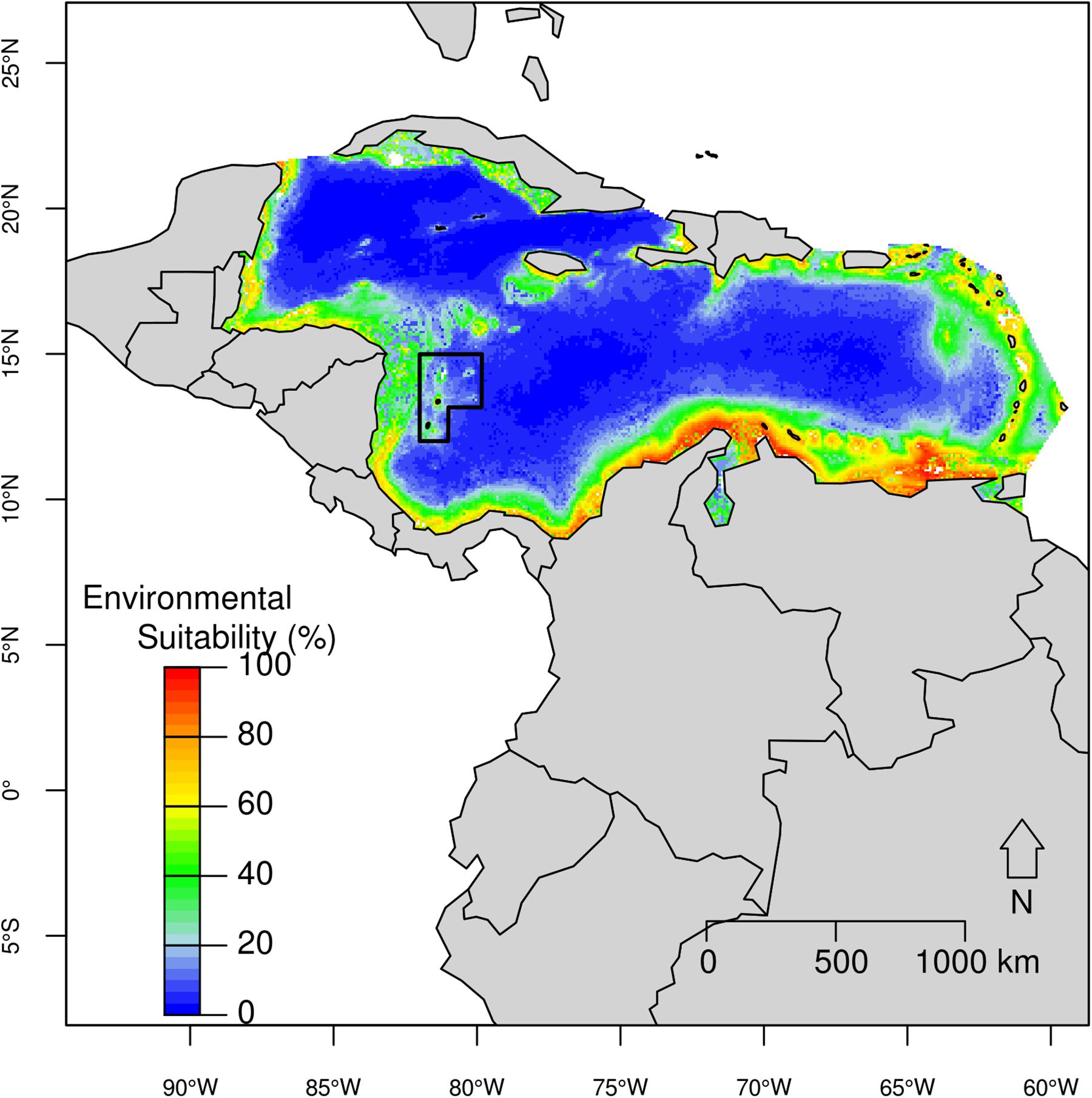
Figure 3. Predicted potential distribution for the Atlantic spotted dolphin in the Caribbean Basin from the best ecological niche model. Red colors indicate highest habitat suitability and blue lowest suitability.
Atlantic Spotted Dolphin Ecological Niche Model
The predicted potential distribution map for the Atlantic spotted dolphin indicated that the areas closest to continental shorelines had the highest environmental suitability (>70%). Specifically a high suitability area was found mainly in the coastal region off mainland Colombia and more broadly, along the southeastern Caribbean, with oceanic areas and particularly the SFBR presenting a low environmental suitability (Figure 3). High environmental suitability (higher than 80%) was observed in areas with bathymetry ranging from 9 m to 1,067 m (median = 77 m); distance to shore between 1 km and 12 km (median = 4 km); slope ranging from 1° to 113° (median = 24°); Mean SST between 25 and 27°C (median = 26.9°C); Range SST between 1.89 and 3.34°C (median = 2.55°C); Mean SSS ranging from 34.3 to 36.1 psu (median = 34.7 psu); and Mean Chl-a between 0.12 and 1.68 mg/m3 (median = 0.64 mg/m3).
Pantropical Spotted Dolphin Ecological Niche Model
An environmental suitability higher than 60% was predicted for the pantropical spotted dolphin across most of the Caribbean Basin, but mainly closer to the continent and nearby islands, including the SFBR (Figure 4). Records with suitability higher than 80% ranged in depth from 58 m to 2,215 m (median = 813 m); distance to shore between 3 km and 72 km (median = 42 km); slope ranging from 2° to 120° (median = 26°); Mean SST between 25.6 and 28.3°C; Range SST was between 1.1 and 4.1°C (median = 1.3°C); Mean SSS around 35 psu; and Mean Chl-a ranging from 0.12 to 1.12 mg/m3 (median = 0.21). In the SFBR, environmental suitability around 60% was detected within the southern portion of the Reserve.
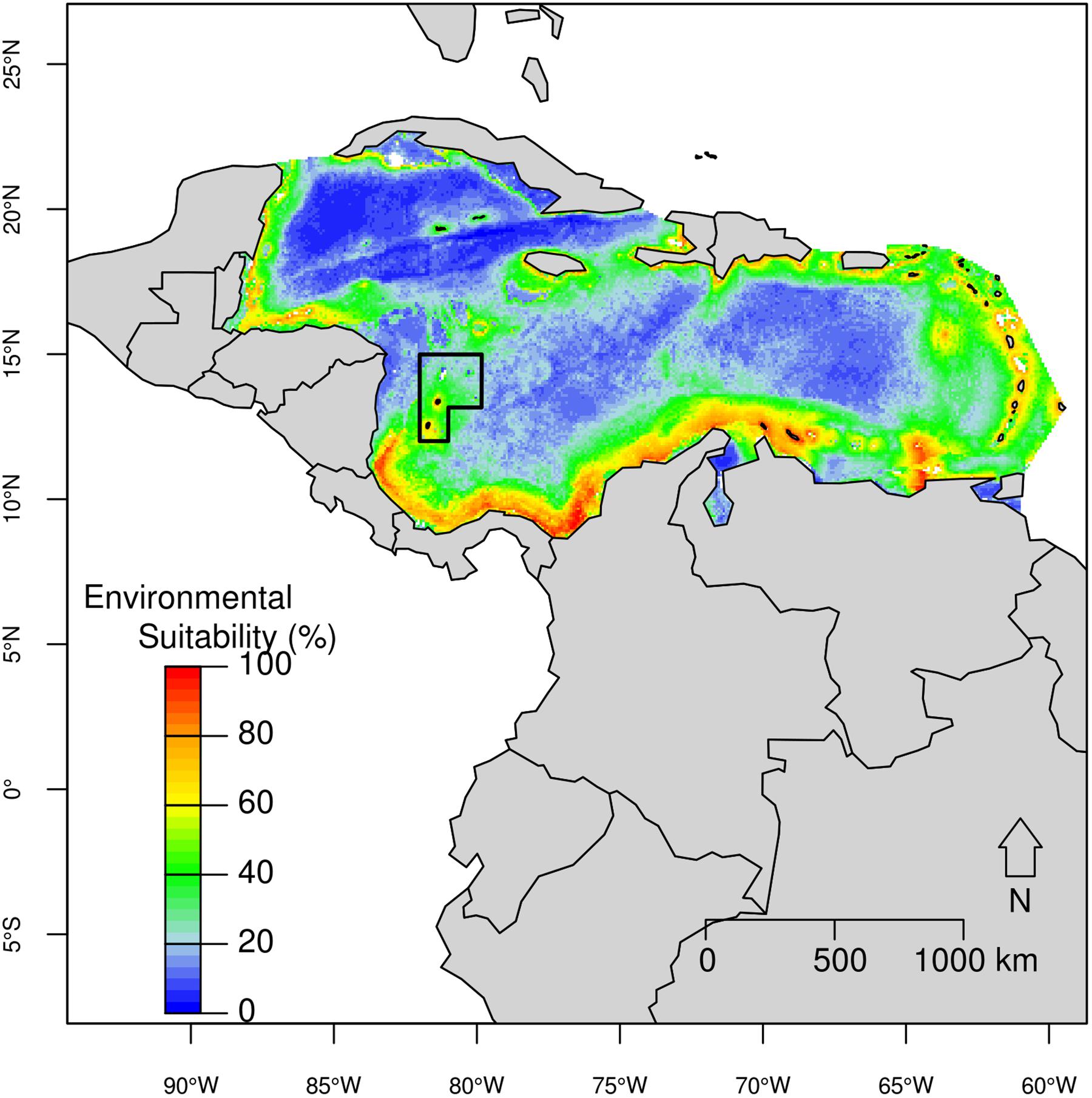
Figure 4. Predicted potential distribution for the pantropical spotted dolphin in the Caribbean Basin from the best ecological niche model. Red colors indicate highest habitat suitability and blue lowest suitability.
Spinner Dolphin Ecological Niche Model
The highest suitability (>70%) for spinner dolphins was predicted in waters surrounding islands or seamounts, and with a steeper slope (Figure 5), in almost all coastal areas in the Caribbean including surrounding islands. High environmental suitability (over 80%) corresponded to areas with bathymetry ranging from 30 m to 1,599 m (median = 620 m); distance to shore between 1 km and 9 km (median = 5 km); slope ranging from 1° to 165° (median = 106.5°); Mean SST between 25.4 and 27.7°C (median = 27.2°C); Range SST between 2.14 to 4°C (median = 2.5°C); Mean SSS ranging from 34.6 to 35.9 psu (median = 35.1 psu); and Mean Chl-a between 0.11 and 1.7 mg/m3 (median = 0.38 mg/m3). The SFBR exhibited a high habitat suitability for spinner dolphins, primarily around the insular region, where environmental suitability reached 72% (e.g., around San Andrés Island).
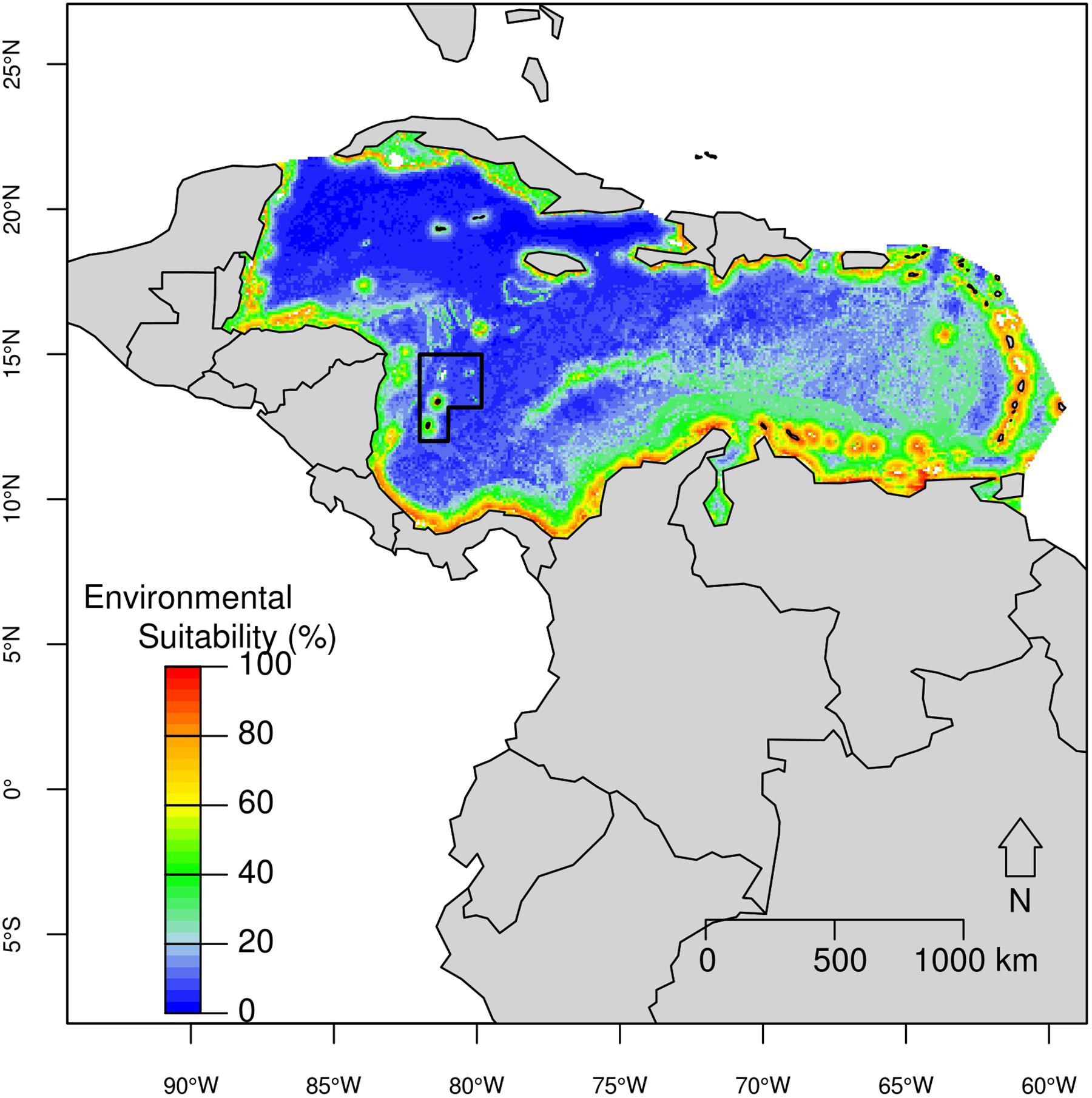
Figure 5. Predicted potential distribution for the spinner dolphin in the Caribbean Basin from the best ecological niche model. Red colors indicate highest habitat suitability and blue lowest suitability.
Niche Overlap and Identity Test
Pairwise overlap for the three Stenella species is presented in Table 3. We found high “I” and “D” similarity statistic scores for all pairwise comparisons, indicating that ecological niches were broadly similar among Atlantic spotted dolphins, pantropical spotted dolphins, and spinner dolphins in the Caribbean Basin. Niche identity tests did not reject the null hypotheses that assumed the niches were identical between pairwise Stenella species comparisons (Table 3).
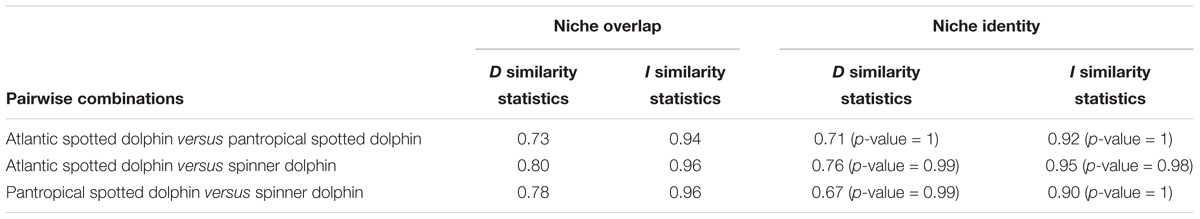
Table 3. Metrics obtained in niche overlap comparison and niche identity tests conducted pairwise between Stenella species.
However, our results indicated that while the distribution of Atlantic spotted dolphins and spinner dolphins showed the highest statistical similarity, Atlantic and pantropical spotted dolphins shared the lowest similarity values. In general, all three species coincided in some areas, according to the overlap map based on the threshold of equal test sensitivity and specificity (Table 4 and Figures 6–8). All species overlapped in their distributions in coastal areas of the southern and southeastern Caribbean off Colombia, Venezuela, and surrounding areas in the Antilles, as well as in the SFBR. Nonetheless, while the distribution of pantropical spotted dolphins included more oceanic waters around 1,000 m deep, spinner dolphins appeared to occupy areas closer to island chains, and Atlantic spotted dolphins appeared to be more restricted to coastal/neritic waters.

Table 4. Threshold of equal test sensitivity and specificity obtained for each model generated among three Stenella species in the Caribbean Basin.
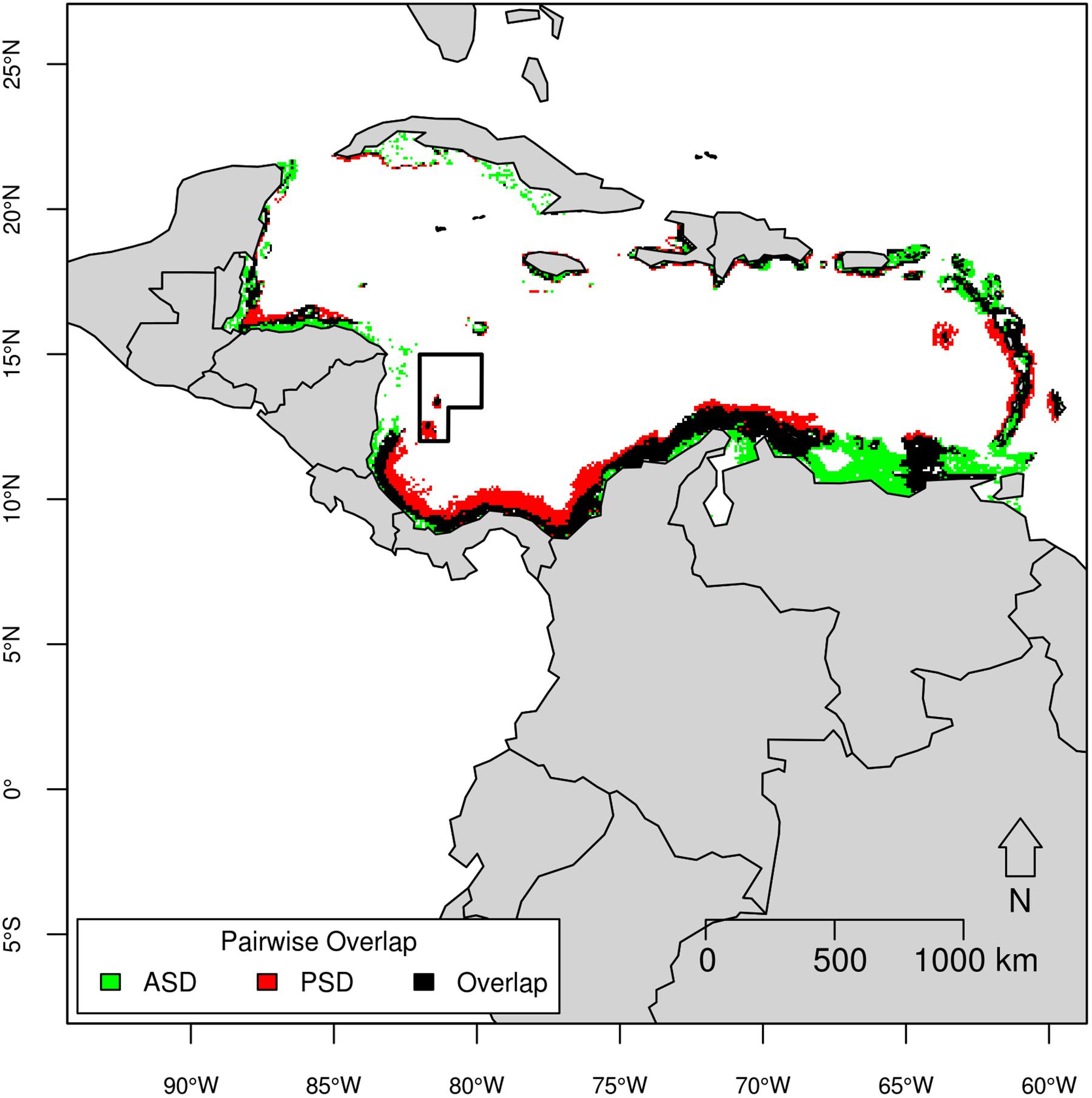
Figure 6. Pairwise overlap map of Atlantic spotted dolphin (ASD) and pantropical spotted dolphin (PSD) based on the threshold of equal test sensitivity and specificity.
Statistical Comparisons of Environmental Variables
The median values of the environmental variables for each species indicated significant interspecific differences (Table 5 and Figure 9). The null hypothesis of equal medians for bathymetry, distance to shore, slope, Mean Chl-a, Mean SST and Range SST was rejected (Kruskal–Wallis test, p < 0.05).
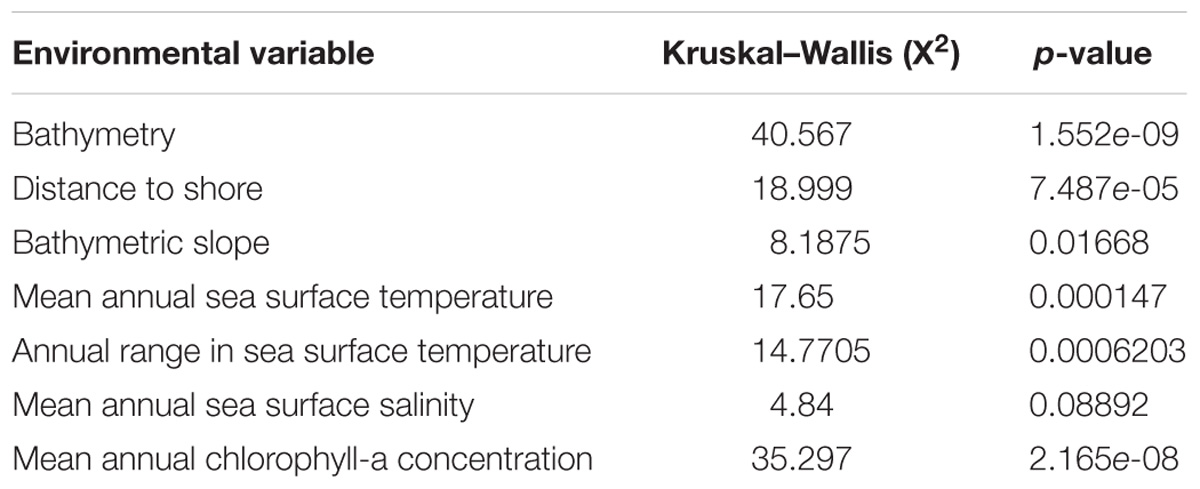
Table 5. Results of the Kruskal–Wallis test for comparing environmental variables between Atlantic spotted dolphins, pantropical spotted dolphins and spinner dolphins.
Pantropical spotted dolphins had the highest median bathymetry, distance to shore, Mean SST, and the lowest median Mean Chl-a. In other words, pantropical spotted dolphins occupied areas of low phytoplankton biomass, and deeper (up to about 1,000 m) and warmer waters. Atlantic spotted dolphins and pantropical spotted dolphins differed in relation to bathymetry, slope, distance to shore, Range SST and Mean Chl-a, with the former occurring in shallow waters closer to the shore, with lower slope values and Mean SST but higher productivity. Likewise, spinner dolphins occurred in areas surrounding the coast, with more phytoplankton biomass, and in shallower waters, although in regions characterized by a steeper slope.
Discussion
This study established a series of potential “baseline” ecological niche models for Atlantic spotted dolphins, pantropical spotted dolphins and spinner dolphins. These models were used for assessing their distribution patterns in the Caribbean Basin, particularly in the SFBR. In this study, we provide the first characterization of the environmental niches of three Stenella species and their potential distributional patterns in the Caribbean Basin. Our findings show that distance to coastline and Mean Chl-a play an important role that influences the potential distribution of these three species in the region.
Species of Stenella are widely distributed in the Caribbean Basin, yet their patterns have been poorly studied. Small delphinids of the genus Stenella carry out long-range movements of up to 2,500 km (Reilly, 1990), presumably associated with prey availability dynamics (Davis et al., 2002). In fact, ecological niches for dolphins appear to be defined by variables that affect distribution and abundance of resources they rely upon (Baumgartner et al., 2001; MacLeod et al., 2008). The availability of dolphin prey depends mainly on primary productivity, which is highly variable in the oligotrophic waters of the Caribbean Basin (Corredor, 1979; Müller-Karger and Aparicio, 1994; Aguirre, 2014). In the first part of the year, primary productivity concentrates along the continental coasts of Colombia and Venezuela, while shifting to Caribbean islands such as Puerto Rico, the Virgin Islands and the Antilles in the second part (Müller-Karger et al., 1989; Müller-Karger and Aparicio, 1994). This variability enables Stenella species, such as pantropical spotted dolphins and spinner dolphins, to exploit oceanic and deeper-water environments given their long-distance movement capabilities.
Coastal-marine ecosystems such as coral reefs and seagrass beds host a high biodiversity and productivity that provide food resources for local and oceanic biological communities. The SFBR appears to be an important area for dolphins in the Caribbean Basin not only because it contains the largest open-ocean coral reefs in the Caribbean, but because it also holds extensive and healthy seagrass beds (Taylor et al., 2013). Consequently, the overlap in the ecological niche of pantropical spotted dolphins and spinner dolphins in the area is not surprising. In contrast, Atlantic spotted dolphins primarily inhabit coastal environments throughout most of their range, a distribution pattern similar to the one previously described in the Western South Atlantic Ocean by do Amaral et al. (2015). Productive coral reefs and seagrass beds are also found along the Colombian mainland coast, notably in Morrosquillo Gulf, the Rosario and San Bernardo Archipelago, Santa Marta, and the Tayrona region (Garzón-Ferreira et al., 2001).
Water temperature has been shown to be an important attribute of cetaceans’ ecological niches (MacLeod, 2009). In the Caribbean Basin, the southern region along the coast of Colombia and Venezuela exhibits low SST associated with coastal upwelling, mainly off La Guajira region (Colombia) in the central Caribbean and around Margarita Island (Venezuela) in the eastern Caribbean (Gordon, 1967; Müller-Karger et al., 1989; Andrade et al., 2003; Andrade and Barton, 2005; Lonin et al., 2010; Paramo et al., 2011). This upwelling generates a high primary productivity (Corredor, 1979; Andrade et al., 2003; Andrade and Barton, 2005; Criales-Hernández et al., 2006; Arévalo-Martínez and Franco-Herrera, 2008; Lonin et al., 2010) that has been related to the occurrence of other dolphin species in the area (Farías-Curtidor et al., 2017). Considering that low SST was an important descriptor of habitat suitability for Atlantic spotted dolphins, it is not surprising that they were preferentially found in Caribbean environments defined by shallow waters with lower temperatures and elevated primary productivity.
Bathymetry is another important predictor of Atlantic spotted dolphin habitats in the Caribbean Basin. Similar to what has been reported for the Gulf of México and the Western South Atlantic Ocean, this species showed a preference for shallower waters along the Colombian Caribbean (Davis et al., 1998; do Amaral et al., 2015). Atlantic spotted dolphins feed preferentially on coastal prey (Perrin et al., 1994), so their presence in shallow waters is frequent (Davis et al., 1998, 2002). These coastal habits could suggest that this species is not very cosmopolitan in the Atlantic Ocean, showing restricted movements across northern and southern portions of this ocean. Genetic studies have shown that individuals from the Colombian Caribbean are closely related to those population units from locations as far as west and east north Atlantic (Mesa-Gutiérrez et al., 2015); while populations from Azores and Madeira, which show high connectivity between them (Quérouil et al., 2010), resulted closely related to those from the Western North Atlantic and Brazil (Caballero et al., 2013; Mesa-Gutiérrez et al., 2015). Additionally, Atlantic spotted dolphins along the Brazilian coast also show genetic (Caballero et al., 2013) and morphological divergence (Moreno et al., 2005) associated with differences in environmental conditions between southern and northern Brazilian populations (do Amaral et al., 2015). Furthermore, Atlantic spotted dolphin populations along the east coast of the United States and Gulf of Mexico show significant population structure due mainly to an extensive continental shelf that promotes differential habitat structuring and hence, the aggregation of resources in specific zones to be exploited (Adams and Rosel, 2006). All these evidences suggest that environmental conditions associated with coastal habitats could drive differentiation between Atlantic spotted dolphins across the Atlantic Ocean although more studies are needed to support this assumption.
Similar to Atlantic spotted dolphins, spinner dolphins occur in productive regions surrounding coastlines but with steeper slopes, mainly near islands (Figure 7). Indeed, the niche for these two species overlapped in much of coastal areas around the Caribbean Basin, except in waters surrounding islands such as in the SFBR, which is characterized by a pronounced slope (mean = 4.5°). The slope had a considerable contribution for the spinner dolphin model, preferred by this species due to the vertical and horizontal migration of mesopelagic fish considered an important part of their diet (Benoit-Bird and Au, 2003; Thorne et al., 2012).
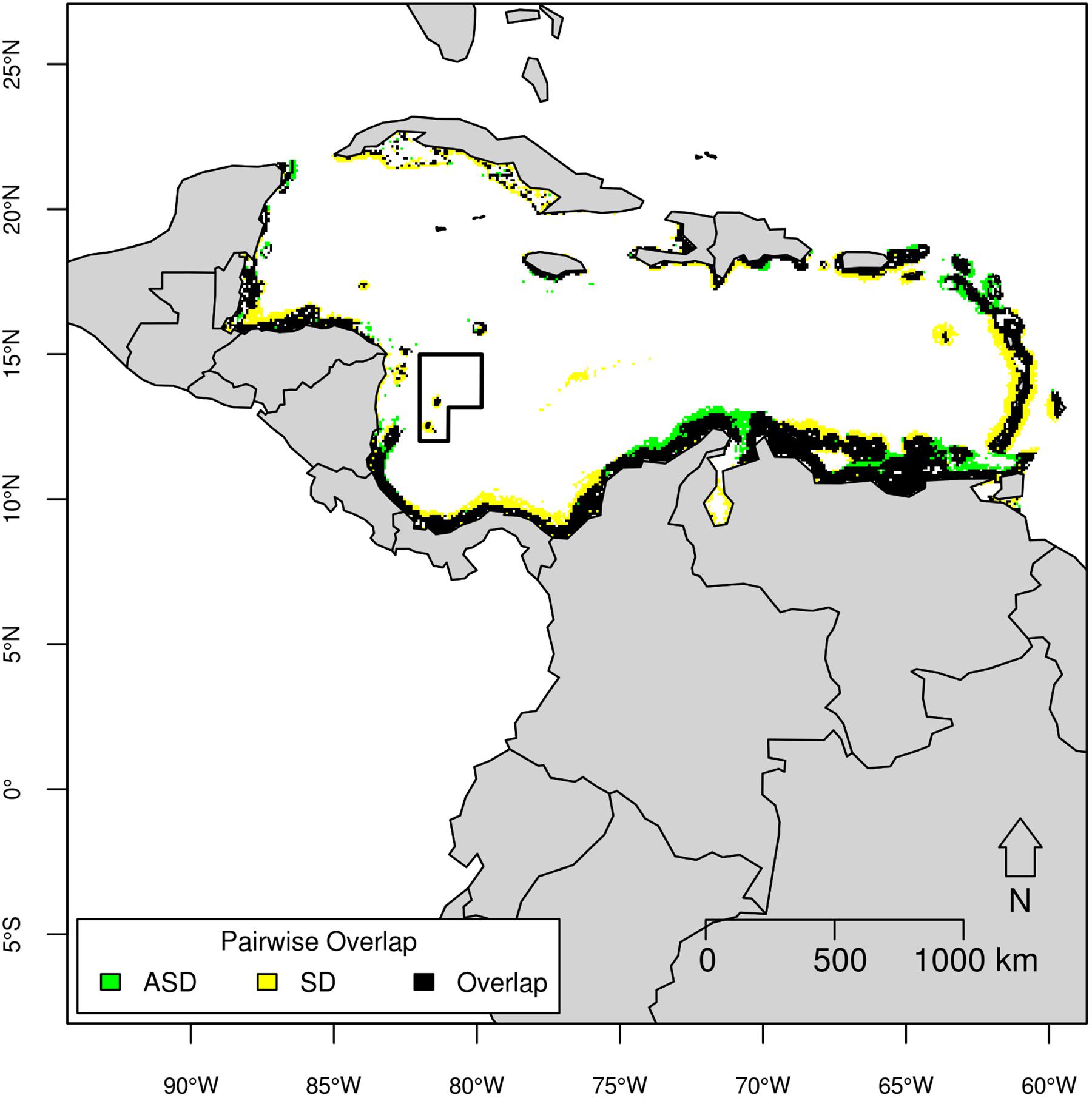
Figure 7. Pairwise overlap map of Atlantic spotted dolphin (ASD) and spinner dolphin (SD) based on the threshold of equal test sensitivity and specificity.
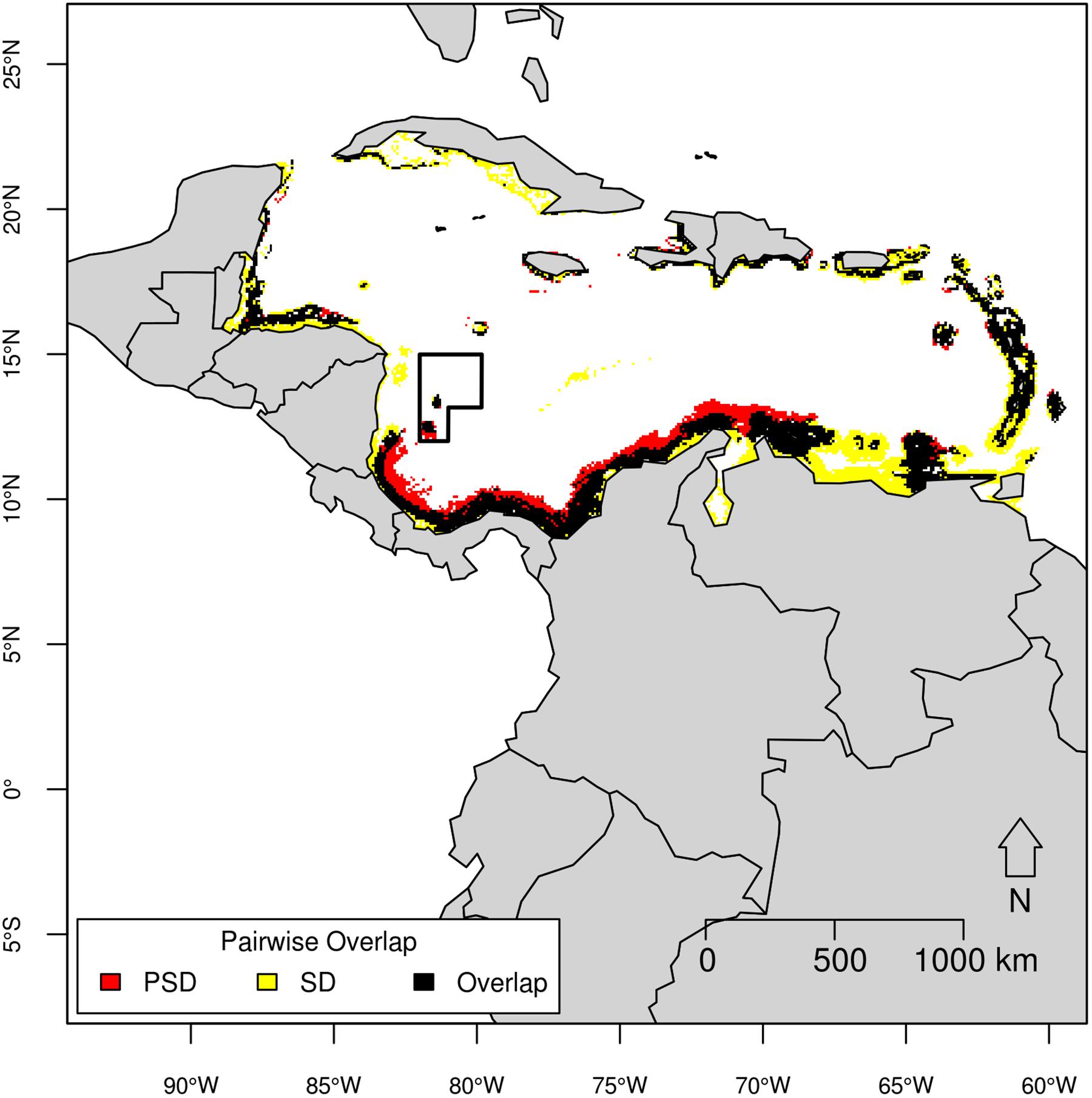
Figure 8. Pairwise overlap map of pantropical spotted dolphin (PSD) and spinner dolphin (SD) based on the threshold of equal test sensitivity and specificity.
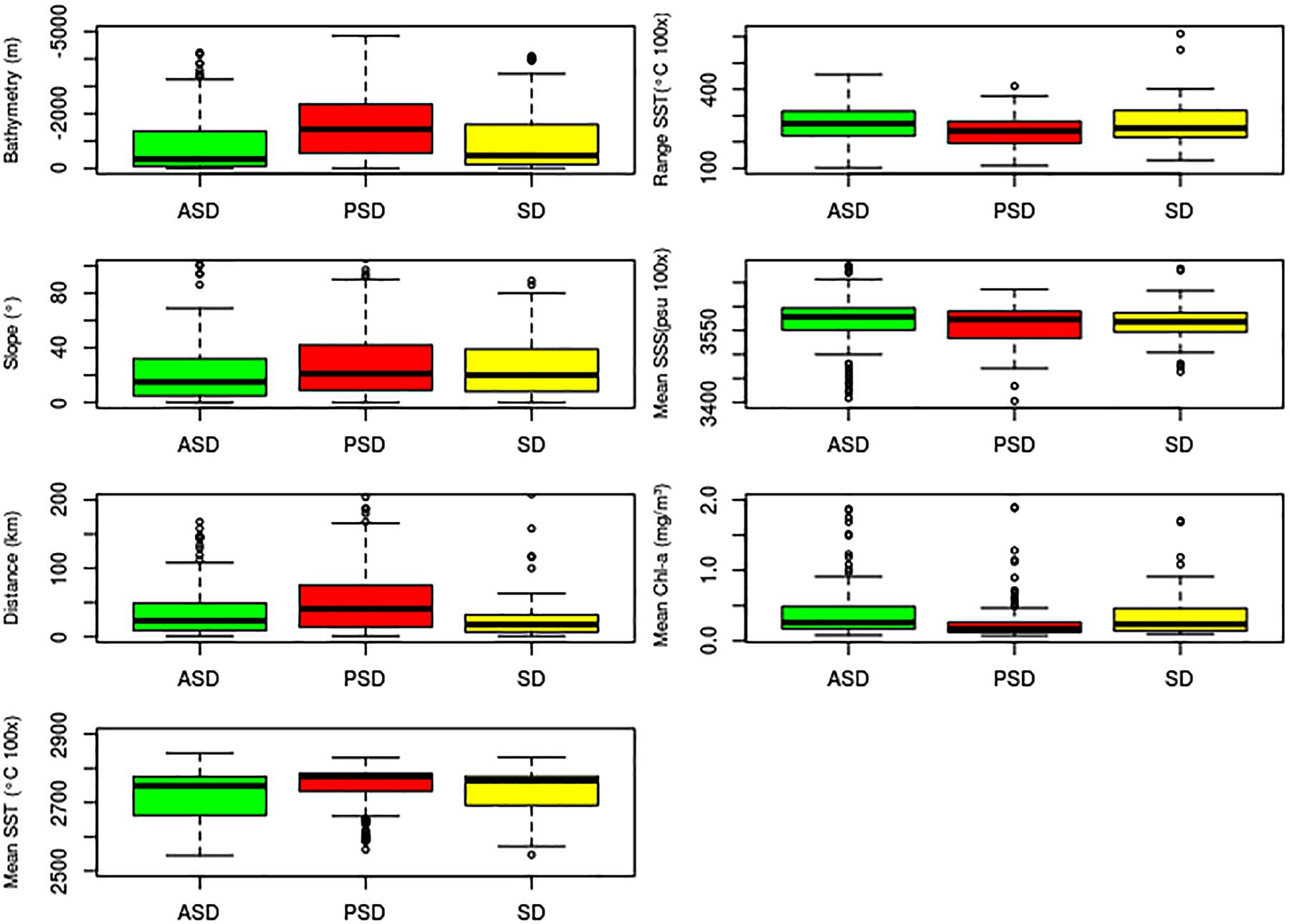
Figure 9. Box plots of values extracted from environmental layers for Atlantic spotted dolphins (ASD), pantropical spotted dolphins (PSD) and spinner dolphins (SD) in the Caribbean Basin. The thick horizontal line inside each box represents the median; the upper and lower borders of the box are the 25th and 75th percentiles; the 5th and 95th percentiles are represented by the error bars; small circles are outliers.
Pantropical spotted dolphins also appeared to prefer waters around islands, but are present in deeper waters, as reported in the Gulf of Mexico (Davis et al., 1998) and in the Western South Atlantic (do Amaral et al., 2015). SST had a considerable contribution to the species’ model, indicating that offshore pantropical spotted dolphins are usually found in warm tropical waters, as reported previously in the Western South Atlantic (do Amaral et al., 2015).
Although the Stenella species analyzed here showed differences in their environmental requirements in the Caribbean Basin, in general both spotted and spinner dolphins occupied similar habitats in the region. It has been suggested that Atlantic spotted dolphins and pantropical spotted dolphins are sympatric in the Atlantic Ocean (Perrin and Hohn, 1994). Likewise, associations between pantropical spotted dolphins and spinner dolphins have been documented in the Atlantic (Silva et al., 2005). However, do Amaral et al. (2015) suggested that Stenella dolphins, particularly the three species considered for the current study, could exhibit niche partitioning and spatial segregation in the Western South Atlantic. Davis et al. (1998) also reported differences in distribution of these three Stenella species along the Gulf of Mexico. These differences have been related not only to oceanographic conditions (do Amaral et al., 2015), but also to prey distribution and feeding preferences among Stenella species (Davis et al., 2002). However, in areas where nutrients are scarce, feeding preferences facilitate that small cetaceans share their habitat (Davis et al., 2002). In the Caribbean Basin, Atlantic spotted, pantropical and spinner dolphins seem to overlap much more in their distributions, particularly in high-productivity areas.
These findings generate special concern regarding climate change due to the effects on marine and coastal productive ecosystems such as coral reefs. Particularly, these ecosystems have been heavily affected in the Caribbean Basin (Connell, 1997; Pandolfi et al., 2003), allowing the increase in macroalgae coverage (Mumby et al., 2007) and the consequent decreases in local biodiversity. Similarly, prey distribution could be affected due to increases in water temperature (Learmonth et al., 2006; MacLeod, 2009). These shifts in prey availability can affect distributional patterns on dolphins in the Caribbean Basin (Learmonth et al., 2006; MacLeod, 2009; Ramos et al., 2016), which could force them to spend more time traveling in oceanic waters looking for resources.
While oceanic areas in the Caribbean Basin appear to be potentially important for Stenella dolphins, most studies have taken place in coastal waters. Therefore, the distribution patterns suggested in this study, based only on sightings, do not predict suitable habitat beyond the known records of Stenella in the Caribbean Basin. Although efforts have increased in the region to advance marine mammal research, there is still a large knowledge gap to improve the management of such species. Areas of very low coverage (like the center of Western Caribbean where no research has been done) could contribute to the habitat close to shore and of specific depths being deemed highly suitable. This is the case for the SFBR, which is an oceanic area poorly studied with regarding marine mammals. This Reserve holds high local biodiversity related to coastal marine ecosystems that are potentially important for dolphins’ distribution as foraging hotspots. The SFBR appears to be more suitable in its southern portion for pantropical and spinner dolphins, mainly around the islands, but their ecological and oceanographic conditions could allow the occurrence of other small cetacean species. Future research should focus on this protected area, to assess if it should be considered as a priority area for cetacean species in the Caribbean Basin.
Author Contributions
DB-B, PC-C, NF-C, RL-N, PB, JB-J, LB, DC-M, JL, JM, RM-G, BdM, ER, VR, and DP contributed to the acquisition of field data. DB-B, KBA, and NF-C organized the data and compiled the literature. DB-B and KBA conceived and designed the experiments, analyzed the data, and wrote the manuscript. KBA performed the experiments. DB-B, KBA, PC-C, NB-A, IM, JB-J, DC-M, JL, RM-G, BdM, ER, and DP contributed to the manuscript writing. All authors have approved the final version of the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the Colombian Program of Marine Fauna Observers, and acknowledge the collaboration of all the observers, captains, and crew members of seismic vessels. Special thanks to Comisión Colombiana del Océano, Armada Nacional de Colombia, ARC Providencia crew, and Universidad de los Andes for their logistic support during the Scientific Seaflower Expedition Cayo Serrana Island 2016. DB-B received a small grant provided by Cetacean Society International to participate in this expedition (2015). Fieldwork in Colombia was also supported by the Society for Marine Mammalogy (DB-B, 2014), the “Proyecto Semilla – 2013-2 Call for Funding of Research Category: Master and Doctoral students, project ‘Genetic structure and diversity of bottlenose dolphins Tursiops truncatus (Montagu, 1821) (Cetacea: Delphinidae) in La Guajira, Colombian Caribbean”’ from Universidad de los Andes (DB-B, 2014), the “Proyecto Semilla – 2015-1 Call for Funding of Research Category: Master and Doctoral students, project ‘Occurrence, distribution and preliminary genetic status of delphinids in La Guajira, Colombian Caribbean”’ from Universidad de los Andes (DB-B, 2015), and the Rufford Small Grant Foundation (NF-C, 2016). KBA received a Ph.D. degree scholarships from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) (process 1395438). Finally, we really appreciate the comments that reviewers have provided to the manuscript, which have been valuable and useful to improve the final version.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2019.00010/full#supplementary-material
Footnotes
References
Adams, L. D., and Rosel, P. E. (2006). Population differentiation of the Atlantic spotted dolphin (Stenella frontalis) in the western North Atlantic, including the Gulf of Mexico. Mar. Biol. 148, 671–681. doi: 10.1007/s00227-005-0094-2
Andrade, C., and Barton, D. (2005). The guajira upwelling system. Cont. Shelf Res. 25, 1003–1022. doi: 10.1016/j.csr.2004.12.012
Andrade, C. A., Barton, E. D., and Mooers, C. N. K. (2003). Evidence for an Eastward Flow along the Central and South American Caribbean Coast. J. Geophys. Res. 108, C6–C3185. doi: 10.1029/2002JC001549
Arévalo-Martínez, D. L., and Franco-Herrera, A. (2008). Características oceanográficas de la surgencia frente a la Ensenada de Gaira, Departamento de Magdalena, época seca menor de 2006. Bol. Invest. Mar. Cost. 37, 131–162.
Bailey, H., and Thompson, P. M. (2009). Using marine mammal habitat modelling to identify priority conservation zones within a marine protected area. Mar. Ecol. Prog. Ser. 378, 279–287. doi: 10.3354/meps07887
Barragán-Barrera, D. C., Farías-Curtidor, N., Do Amaral, K., Lancheros-Neva, R., Botero-Acosta, N., and Bueno, P. (2017). “Registro del delfín manchado pantropical (Stenella attenuata) en la Isla Cayo Serrana y su distribución potencial en el Caribe,” in Oral presentation from the Coloquio de Resultados Expedición Científica Seaflower 2016, (Isla Cayo Serrana).
Baumgartner, M. F., Mullin, K. D., May, L. N., and Leming, T. D. (2001). Cetacean habitats in the northern Gulf of Mexico. Fish. Bull. 99, 219–239.
Benoit-Bird, K. J., and Au, W. W. L. (2003). Prey dynamics affect foraging by a pelagic predator (Stenella longirostris) over a range of spatial and temporal scales. Behav. Ecol. Sociobiol. 53, 364–373.
Bolaños-Jiménez, J., Mignucci-Giannoni, A., Blumenthal, J., Bogomolni, A., Casas, J. J., Henríquez, A., et al. (2014). Distribution, feeding habits and morphology of killer whales Orcinus orca in the Caribbean Sea. Mammal Rev. 44, 177–189. doi: 10.1111/mam.12021
Caballero, S., Santos, M. C. O., Sanches, A., and Mignucci-Giannoni, A. A. (2013). Initial description of the phylogeography, population structure and genetic diversity of Atlantic spotted dolphins from Brazil and the Caribbean, inferred from analyses of mitochondrial and nuclear DNA. Biochem. Syst. Ecol. 48, 263–270. doi: 10.1016/j.bse.2012.12.016
Caldwell, D. K., Caldwell, M. C., Rathjen, W. F., and Sullivan, J. R. (1971). Cetaceans from the Lesser Antillean island of St. Vincent. Fish. Bull. 69, 303–312.
Cañadas, A., Sagarminaga, R., and García-Tiscar, S. (2002). Cetacean distribution related with depth and slope in the Mediterranean waters off southern Spain. Deep Sea Res. Part I Oceanogr. Res. Pap. 49, 2053–2073. doi: 10.1016/S0967-0637(02)00123-1
Chollett, I., Müller-Karger, F. E., Heron, S. F., Skirving, W., and Mumby, P. J. (2012). Seasonal and spatial heterogeneity of recent sea surface temperature trends in the Caribbean Sea and southeast Gulf of Mexico. Mar. Pollut. Bull. 64, 956–965. doi: 10.1016/j.marpolbul.2012.02.016
Combatt, J. A., and González, E. (2007). Ocurrencia y distribución del delfín nariz de botella Tursiops truncatus (Montagu, 1821) en las costas de Dibulla, Baja Guajira, durante el período de agosto a diciembre de 2005. dissertation/bachelor’s thesis, Universidad de Bogotá Jorge Tadeo Lozano, Santa Marta.
Connell, J. H. (1997). Disturbance and recovery of coral assemblages. Coral Reefs. 16, S101–S113. doi: 10.1007/s003380050246
Coralina-Invemar (2012). “Atlas de la Reserva de Biósfera Seaflower,” in Archipiélago de San Andrés, Providencia y Santa Catalina. Santa Marta: Instituto de Investigaciones Marinas y Costeras “José Benito Vives De Andréis” -INVEMAR- y Corporación para el Desarrollo Sostenible del Archipiélago de San Andrés, eds D. Gómez-López I, C. Segura-Quintero, P. C. Sierra-Correa, and J. Garay-Tinoco (Coralina: Providencia y Santa Catalina -CORALINA).
Corredor, J. E. (1979). Phytoplankton response to low level nutrient enrichment through upwelling in the Colombian Caribbean Basin. Deep Sea Res. Part A Oceanogr. Res. Pap. 26, 731–741. doi: 10.1016/0198-0149(79)90010-4
Criales-Hernández, M. C., García, C. B., and Wolff, M. (2006). Flujos de biomasa y estructura de un ecosistema de surgencia tropical en La Guajira, Caribe colombiano. Rev. Biol. Trop. 54, 1257–1282. doi: 10.15517/rbt.v54i4.14399
Davis, R. W., Fargion, G. S., May, N., Lemig, T. D., Baumgartner, M., Evans, W. E., et al. (1998). Physical habitat of the cetaceans along the continental slope in the north central and western Gulf of Mexico. Mar. Mamm. Sci. 14, 490–507. doi: 10.1111/j.1748-7692.1998.tb00738.x
Davis, R. W., Ortega-Ortiz, J. G., Ribic, C. A., Evans, W. E., Biggs, D. C., Ressler, P. H., et al. (2002). Cetacean habitat in the northern oceanic Gulf of Mexico. Deep Sea Res. Part I Oceanogr. Res. Pap. 49, 121–142. doi: 10.1016/S0967-0637(01)00035-8
Derville, S., Torres, L., Iovan, C., and Garrigue, C. (2018). Finding the right fit: comparative cetacean distribution models using multiple data sources and statistical approaches. Divers. Distrib. 24, 1657–1673. doi: 10.1111/ddi.12782
Dinno, A. (2017). Dunn.test: Dunn’s Test of Multiple Comparisons Using Rank Sums. R package version 1.3.5. Available at: https://CRAN.R-project.org/package=dunn.test
do Amaral, K. B., Alvares, D. J., Heinzelmann, L., Borges-Martins, M., Siciliano, S., and Moreno, I. B. (2015). Ecological niche modeling of Stenella dolphins (Cetartiodactyla: delphinidae) in the southwestern Atlantic Ocean. J. Exp. Mar. Bio. Ecol. 472, 166–179. doi: 10.1016/j.jembe.2015.07.013
Dormann, C. F., Elith, J., Bacher, S., Buchmann, C., Carl, G., Carré, G., et al. (2013). Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36, 27–46. doi: 10.1111/j.1600-0587.2012.07348.x
Elith, J., Phillips, S. J., Hastie, T., Dudík, M., Chee, Y. E., and Yates, C. J. (2011). A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 17, 43–57. doi: 10.1111/j.1472-4642.2010.00725.x
Farías-Curtidor, N., Barragán-Barrera, D. C., Chávez-Carreño, P. A., Jiménez-Pinedo, C., Palacios, D., Caicedo, D., et al. (2017). Range extension for the common dolphin (Delphinus sp.) to the Colombian Caribbean, with taxonomic implications from genetic barcoding analysis. PLoS One 12:e0171000. doi: 10.1371/journal.pone.0171000
Fraija, N., Flórez-González, L., and Jáuregui, A. (2009). Cetacean occurrence in the Santa Marta region, Colombian Caribbean. Lat. Am. J. Aquat. Mamm. 7, 69–73.
Franklin, J. (2009). Mapping Species Distributions: Spatial Inference and Prediction. Cambridge: Cambridge University Press.
García, C., and Trujillo, F. (2004). Preliminary observations on habitat use patterns of the Marine Tucuxi, Sotalia fluviatilis, in Cispatá Bay, Colombian Caribbean Coast. Lat. Am. J. Aquat. Mamm. 3, 53–59. doi: 10.5597/lajam00048
Garzón-Ferreira, J., Rodríguez-Ramírez, A., Bejarano-Chavarro, S., Navas-Camacho y, R., and Reyes-Nivia, C. (2001). “Estado de los arrecifes coralinos en Colombia,” in INVEMAR Informe del Estado de los Ambientes Marinos y Costeros en Colombia, eds G. H. Ospina-Salazar and A. Acero (Santa Marta: Serie de publicaciones periódicas INVEMAR), 29–40.
Gomez, C., Lawson, J., Kouwenberg, A.-L., Moors-Murphy, H., Buren, A., Fuentes-Yaco, C., et al. (2017). Predicted distribution of whales at risk: identifying priority areas to enhance cetacean monitoring in the Northwest Atlantic Ocean. Endang. Species Res. 32, 437–458. doi: 10.3354/esr00823
Gordon, A. L. (1967). Circulation of the Caribbean Sea. J. Geophys. Res. 72, 6207–6223. doi: 10.1029/JZ072i024p06207
Gregr, E. J., Baumgartner, M. F., Laidre, K. L., and Palacios, D. M. (2013). Marine mammal habitat models come of age: the emergence of ecological and management relevance. Endanger. Species Res. 22, 205–212. doi: 10.3354/esr00476
Hijmans, R. (2017). raster: Geographic Data Analysis and Modeling. R package version 2.6-7. Available at: https://CRAN.R-project.org/package=raster
Hijmans, R., and Elith, J. (2017). Species Distribution Modeling With R. Available at: https://cran.r-project.org/web/packages/dismo/vignettes/sdm.pdf
Hijmans, R. J., Phillips, S., Leathwick, J. R., and Elith, J. (2017). dismo: Species Distribution Modeling. R package version 1.1-4. Available at: https://CRAN.R-project.org/package=dismo
Jefferson, T. A., Webber, M. A., and Pitman, R. L. (2008). Marine Mammals of the World: A Comprehensive Guide to Their Identification. Oxford: Elsevier, Academic Press.
Kelley, J., Brown, A., Therkildsen, N., and Foote, A. (2016). The life aquatic: advances in marine vertebrate genomics. Nat. Rev. Genet. 17, 523–534. doi: 10.1038/nrg.2016.66
Learmonth, J. A., MacLeod, C. D., Santos, M. B., Pierce, G. J., Crick, H. Q. P., and Robinson, R. A. (2006). Potential effects of climate change on marine mammals. Oceanogr. Mar. Biol. 44, 431–464. doi: 10.1201/9781420006391.ch8
Leduc, R. G., Perrin, W. F., and Dizon, A. E. (1999). Phylogenetic relationships among the delphinid cetaceans based on full cytochrome b sequences. Mar. Mamm. Sci. 15, 619–648. doi: 10.1111/j.1748-7692.1999.tb00833.x
Leutner, B., and Horning, N. (2016). Tools for Remote Sensing Data Analysis. The Comprehensive R Archive Network. Available at: https://cran.r-project.org/web/packages/RStoolbox/RStoolbox.pdf
Lonin, S. A., Hernández, J. L., and Palacios, D. M. (2010). Atmospheric events disrupting coastal upwelling in the southwestern Caribbean. J. Geophys. Res. 115:C06030. doi: 10.1029/2008JC005100
MacLeod, C. D. (2009). Global climate change, range changes and potential implications for the conservation of marine cetaceans: a review and synthesis. Endanger. Species Res. 7, 125–136. doi: 10.3354/esr00197
MacLeod, C. D., Mandleberg, L., Schweder, C., Bannon, S. M., and Pierce, G. J. (2008). A comparison of approaches for modelling the occurrence of marine animals. Hydrobiologia 612, 21–32. doi: 10.1016/j.prevetmed.2012.11.005
Merow, C., Smith, M. J., and Silander, J. A. Jr. (2013). A practical guide to MaxEnt for modeling species’ distributions: what it does, and why inputs and settings mater. Ecography 36, 1058–1069. doi: 10.1111/j.1600-0587.2013.07872.x
Mesa-Gutiérrez, R., Barragán-Barrera, D. C., Chávez-Carreño, P., Farías-Curtidor, N., and Caballero, S. (2015). “Population structure of the Atlantic spotted dolphin (Stenella frontalis) in La Guajira, Colombian Caribbean,” in Abstract retrieved from the 21st Biennial Conference on The Biology of Marine Mammals, (San Francisco, CA).
Mignucci-Giannoni, A. A. (1998). Zoogeography of cetaceans off Puerto Rico and the Virgin Islands. Caribb. J. Sci. 34, 173–190.
Moreno, B. I, Zerbini, A. N., Danilewicz, D., Santos, M. C. O., Simões-Lopes, P. C., Laílson-Brito, J. Jr., et al. (2005). Distribution and habitat characteristics of dolphins of the genus Stenella (Cetacea: delphinidae) in the southwest Atlantic Ocean. Mar. Ecol. Prog. Ser. 300, 229–240. doi: 10.3354/meps300229
Mow, J. M., Taylor, E., Howard, M., Bained, M., Connolly, E., and Chiquillo, M. (2007). Collaborative planning and management of the San Andres Archipelago’s coastal and marine resources: a short communication on the evolution of the Seaflower marine protected area. Ocean Coast. Manage. 50, 209–222. doi: 10.1016/j.ocecoaman.2006.09.001
Müller-Karger, F. E., and Aparicio, R. (1994). Mesoscale processes affecting phytoplankton abundance in the southern Caribbean Sea. Cont. Shelf Res. 14, 199–221. doi: 10.1016/0278-4343(94)90013-2
Müller-Karger, F. E., McClain, C. R., Fisher, T. R., Esaias, W. E., and Varela, R. (1989). Pigment distribution in the Caribbean Sea: observations from space. Prog. Oceanogr. 23, 23–64. doi: 10.1016/0079-6611(89)90024-4
Mumby, P. J., Hastings, A., and Edwards, H. J. (2007). Thresholds and the resilience of Caribbean coral reefs. Nature 450, 98–101. doi: 10.1038/nature06252
Murillo, C. I. (2014). “Reserva de Biósfera Seaflower,” in Construyendo País Marítimo, eds C. G. Linares, N. J. Machuca and S. H. Reyes (Bogotá, CO: Comisión Colombiana del Océano), 138–145.
Muscarella, R., Galante, P. J., Soley-Guardia, M., Boria, R. A., Kass, J. M., Uriarte, M., et al. (2014). ENMeval: an R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods Ecol. Evol. 5, 1198–1205. doi: 10.1111/2041-210X.12261
Niño-Torres, C. A., García-Rivas, M., del, C., CastelblancoMartínez, D. N., Padilla-Saldiar, J. A., Blanco-Parra, M., et al. (2015). Aquatic mammals from the Mexican Caribbean: a review. Hidrobiologica 25, 143–154. doi: 10.1007/978-1-4614-6898-1_3
Palacios, D. M., Farías-Curtidor, N., Jiménez-Pinedo, C., Castellanos, L., Gärtner, A., Gómez-Salazar, C., et al. (2012). “Range extension for the long-beaked common dolphin (Delphinus capensis) to the Colombian Caribbean,” in Paper SC/64/SM20 Presented to the IWC Scientific Committee Annual Meeting, (Panama), 6.
Palacios, D. M., Baumgartner, M. F., Laidre, K. L., and Gregr, E. J. (2013a). Beyond correlation: integrating environmentally and behaviourally mediated processes in models of marine mammal distributions. Endanger. Species Res. 22, 191–203. doi: 10.3354/esr00558
Palacios, D. M., Gärtner, A., Caicedo, D., Farías, N., Jiménez-Pinedo, C., Curcio-Valencia, J., et al. (2013b). “Mamíferos acuáticos de la región Caribe colombiana,” in Diagnóstico del Estado de conocimiento y conservación de los mamíferos acuáticos en Colombia, eds F. Trujillo, A. Gärtner, D. Caicedo, and M. C. Diazgranados (Bogotá, CO: Ministerio de Ambiente y Desarrollo Sostenible, Fundación Omacha, Conservación Internacional, and WWF), 94–127.
Pandolfi, J. M., Bradbury, R. H., Sala, E., Hughes, T. P., Bjorndal, K. A., Cooke, R. G., et al. (2003). Global trajectories of the long-term decline of coral reef ecosystems. Science 301, 955–958. doi: 10.1126/science.1085706
Paramo, J., Correa, M., and Núñez, S. (2011). Evidencias de desacople físico-biológico en el sistema de surgencia en La Guajira, Caribe colombiano. Rev. Biol. Mar. Oceanogr. 46, 421–430. doi: 10.4067/S0718-19572011000300011
Pardo, M., and Palacios, D. (2006). Cetacean occurrence in the Santa Marta Region, Colombian Caribbean, 2004-2005. Lat. Am. J. Aquat. Mamm. 5, 129–134. doi: 10.5597/lajam00105
Pardo, M. A., Mejía-Fajardo, A., Beltrán-Pedreros, S., Trujillo, F., Kerr, I., and Palacios, D. M. (2009). Odontocete sightings collected during offshore cruises in the western and Southwestern Caribbean Sea. Lat. Am. J. Aquat. Mamm. 7, 57–62. doi: 10.5597/lajam00135
Perrin, W. F., Caldwell, D. K., and Caldwell, M. C. (1994). “Atlantic spotted dolphin Stenella frontalis (G. Cuvier, 1829),” in Handbook of Marine Mammals, eds S. H. Ridgway and R. Harrison (San Diego, CA: Academic Press),173–190.
Perrin, W. F., and Hohn, A. A. (1994). “Pantropical spotted dolphin Stenella attenuata,” in The First Book of Dolphins, eds S. H. Ridgway and R. Harrison (San Diego, CA: Academic Press), 71–98.
Perrin, W. F., Mitchell, E. D., Mead, J. G., Caldwell, D. K., Caldwell, M. C., Bree, P. J., et al. (1987). Revision of the spotted dolphins, Stenella spp. Mar. Mamm. Sci. 3, 99–170. doi: 10.1111/j.1748-7692.1987.tb00158.x
Perrin, W. F., Rosel, P. E., and Cipriano, F. (2013). How to contend with paraphyly in the taxonomy of the delphinine cetaceans? Mar. Mamm. Sci. 29, 567–588. doi: 10.1111/mms.12051
Phillips, S. J., Anderson, R. P., Dudík, M., Schapire, R. E., and Blair, M. E. (2017). Opening the black box: an open-source release of Maxent. Ecography 40, 887–893. doi: 10.1111/ecog.03049
Phillips, S. J., Anderson, R. P., and Schapire, R. E. (2006). Maximum entropy modeling of species geographic distributions. Ecol. Model. 190, 231–259. doi: 10.1016/j.ecolmodel.2005.03.026
Prieto-Rodríguez, M. (1988). Reporte de algunos cetáceos del Caribe Colombiano. Bol. Fac. Biol. Mar. UJTL 8, 30–40.
Quérouil, S., Freitas, L., Cascão, I., Alves, F., Dinis, A., Almeida, J. R., et al. (2010). Molecular insight into the population structure of common and spotted dolphins inhabiting the pelagic waters of the Northeast Atlantic. Mar. Biol. 157, 2567–2580. doi: 10.1007/s00227-010-1519-0
Ramos, E. A., Castelblanco-Martínez, D. N., Niño-Torres, C. A., Jenko, K., and Gomez, N. A. (2016). A review of the aquatic mammals of Belize. Aquat. Mamm. 42, 476–493. doi: 10.1578/AM.42.4.2016.476
R Core Team (2018). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Available at: https://www.R-project.org/
Redfern, J., Ferguson, M., Becker, E., Hyrenbach, K., Good, C. P., Barlow, J., et al. (2006). Techniques for cetacean–habitat modeling. Mar. Ecol. Prog. Ser. 310, 271–295. doi: 10.3354/meps310271
Reilly, S. B. (1990). Seasonal changes in distribution and habitat differences among dolphins in the eastern tropical Pacific. Mar. Ecol. Prog. Ser. 66, 1–11. doi: 10.3354/meps066001
Robinson, N. M., Nelson, W. A., Costello, M. J., Sutherland, J. E., and Lundquist, C. J. (2017). A systematic review of marine-based Species Distribution Models (SDMs) with recommendations for best practice. Front. Mar. Sci. 4:421. doi: 10.3389/fmars.2017.00421
Rueda-Roa, D. T., and Muller-Karger, F. E. (2013). The southern Caribbean upwelling system: sea surface temperature, wind forcing and chlorophyll concentration patterns. Deep Sea Res. Part I Oceanogr. Res. Pap. 78, 102–114. doi: 10.1016/j.dsr.2013.04.008
Sbrocco, E., and Barber, P. H. (2013). MARSPEC: ocean climate layers for marine spatial ecology. Ecology 94, 979–979. doi: 10.1890/12-1358.1
Silva, J. M. Jr., Silva, F. J. L., and Sazima, I. (2005). Rest, nurture, sex, release, and play: diurnal underwater behaviour of the spinner dolphin at Fernando de Noronha Archipelago,SW Atlantic. J. Ichthyol. Aquat. Biol. 9, 161–176.
Spalding, M., Agostini, V., Rice, J., and Grant, S. (2012). Pelagic provinces of the world: a biogeographic classification of the world’s surface pelagic waters. Ocean Coast. Manage. 60, 19–30. doi: 10.1016/j.ocecoaman.2011.12.016
Taylor, E., Baine, M., Killmer, A., and Howard, M. (2013). Seaflower marine protected area: governance for sustainable development. Mar. Policy 41, 57–64. doi: 10.1016/j.marpol.2012.12.023
Thorne, L. H., Johnston, D. W., Urban, D. L., Tyne, J., Bejder, L., Baird, R. W., et al. (2012). Predictive modeling of spinner dolphin Stenella longirostris resting habitat in the Main Hawaiian Islands. PLoS One 7:e43167. doi: 10.1371/journal.pone.0043167
Tobeña, M., Prieto, R., Machete, M., and Silva, M. A. (2016). Modeling the potential distribution and richness of cetaceans in the Azores from Fisheries Observer Program data. Front. Mar. Sci. 3:202. doi: 10.3389/fmars.2016.00202
Tyberghein, L., Verbruggen, H., Pauly, K., Troupin, C., Mineur, F., and De Clerck, O. (2012). Bio-ORACLE: a global environmental dataset for marine species distribution modelling. Glob. Ecol. Biogeogr. 21, 272–281. doi: 10.1111/j.1466-8238.2011.00656.x
Warren, D. L., Glor, R. E., Turelli, M., and Funk, D. (2008). Environmental niche equivalency versus conservatism: quantitative approaches to niche evolution. Evolution 62, 2868–2883. doi: 10.1111/j.1558-5646.2008.00482.x
Keywords: Stenella, potential distribution area, Caribbean Basin, Seaflower Biosphere Reserve, Maxent, ecological niche modeling, niche overlap
Citation: Barragán-Barrera DC, do Amaral KB, Chávez-Carreño PA, Farías-Curtidor N, Lancheros-Neva R, Botero-Acosta N, Bueno P, Moreno IB, Bolaños-Jiménez J, Bouveret L, Castelblanco-Martínez DN, Luksenburg JA, Mellinger J, Mesa-Gutiérrez R, de Montgolfier B, Ramos EA, Ridoux V and Palacios DM (2019) Ecological Niche Modeling of Three Species of Stenella Dolphins in the Caribbean Basin, With Application to the Seaflower Biosphere Reserve. Front. Mar. Sci. 6:10. doi: 10.3389/fmars.2019.00010
Received: 31 July 2018; Accepted: 11 January 2019;
Published: 12 February 2019.
Edited by:
Santiago Herrera, Lehigh University, United StatesReviewed by:
Lindsay Porter, University of St Andrews, United KingdomSamuel Georgian, Marine Conservation Institute, United States
Copyright © 2019 Barragán-Barrera, do Amaral, Chávez-Carreño, Farías-Curtidor, Lancheros-Neva, Botero-Acosta, Bueno, Moreno, Bolaños-Jiménez, Bouveret, Castelblanco-Martínez, Luksenburg, Mellinger, Mesa-Gutiérrez, de Montgolfier, Ramos, Ridoux and Palacios. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dalia C. Barragán-Barrera, ZGFsaWFjLmJhcnJhZ2FuYmFycmVyYUBnbWFpbC5jb20= Karina Bohrer do Amaral, a2FyaW5hYm9ocmVyZG9hbWFyYWxAZ21haWwuY29t Daniel M. Palacios, ZGFuaWVsLnBhbGFjaW9zQG9yZWdvbnN0YXRlLmVkdQ==