 Julian Lilkendey1,2*†
Julian Lilkendey1,2*† Timo Pisternick1,3
Timo Pisternick1,3 Sarah I. Neumann1
Sarah I. Neumann1 Danishta Dumur Neelayya3Stefanie Bröhl1
Danishta Dumur Neelayya3Stefanie Bröhl1 Yashvin Neehaul3
Yashvin Neehaul3 Nils Moosdorf1,4†
Nils Moosdorf1,4†- 1Leibniz Centre for Tropical Marine Research (ZMT), Bremen, Germany
- 2School of Science, Auckland University of Technology, Auckland, New Zealand
- 3Mauritius Oceanography Institute (MOI), Albion, Mauritius
- 4Institute of Geosciences, University of Kiel, Kiel, Germany
Fresh submarine groundwater discharge (fresh SGD), the efflux of terrestrial groundwater directly into the ocean, is a ubiquitous pathway for nutrient-rich freshwater to coastal ecosystems, altering their hydrography, hydrochemistry, and primary productivity. Yet only little is known about the effects of fresh SGD on the fitness of higher trophic levels such as teleost fish. Otolith analysis revealed that somatic growth rates were significantly higher and settlement to reef habitat took place significantly earlier in juvenile gray demoiselle Chrysiptera glauca exposed to fresh SGD as compared to strictly marine conditions. Contrary to expectations, feeding conditions were comparable in both habitats. We propose that physiologically beneficial environmental conditions brought about by the submarine influx of cold acidic freshwater enabled juvenile fish to exhibit elevated growth rates, thereby increasing their survival potential. This effect would directly link changes in groundwater on land to variations in marine primary and secondary consumer biomass at the coast.
Introduction
Submarine groundwater discharge (SGD) is an important, albeit subtle pathway for nutrient rich freshwater to coastal ecosystems (Johannes and Hearn, 1985; Lapointe and O’Connell, 1989; Slomp and Van Cappellen, 2004; Luo et al., 2018). The phenomenon referred to as SGD entails the recirculation of seawater as well as the influx of fresh terrestrial groundwater (fresh SGD) (Burnett et al., 2003; Moore, 2010) and occurs nearly ubiquitously at the shorelines (Moosdorf et al., 2015). Here we will focus on fresh SGD, since its ramifications on coastal ecology are far more incisive: globally, fresh SGD amounts to up to 10% of the gross river discharge (Taniguchi et al., 2002). The process is responsible for habitat modifications and niche partitioning in benthic communities via local changes in water temperature, salinity, and pH (Amato et al., 2016; Foley, 2018). Fresh SGD can drive a coastal ecosystem’s primary production via nutrient enrichment, which in turn leads to elevated primary and secondary consumer biomass (Dale and Miller, 2008; Waska and Kim, 2011; Encarnação et al., 2014; Hata et al., 2016; Utsunomiya et al., 2017; Lecher and Mackey, 2018; Piló et al., 2018). The assessment of factors influencing the abundance and growth of these consumers, in particular teleost fishes, is of ever-growing concern since this information is vital to predict consequences of anthropogenic actions on ecosystem functioning and productivity (Burnett et al., 2018; Shoji and Tominaga, 2018).
In marine fish early life stages rapid growth generally decreases predation mortality (Anderson, 1988; Houde, 1989). Improved feeding conditions are proposed to have positive effects on the condition and growth of teleost fishes (Jones, 1986). On settlement, juvenile reef fish suffer high rates of mortality which signifies this life history stage as an especially critical bottleneck for a fish cohort (Almany and Webster, 2006). Enhanced somatic condition, augmented growth, and earlier settlement are consequently indicators of elevated survival potential and confer higher fitness to reef fish (Booth, 1995; Suthers, 1998; Hoey and McCormick, 2004). Water temperatures, pH levels, dissolved oxygen content, and salinity divergent from the marine conditions a fish is adapted to may pose as physiological stressors, potentially negatively affecting condition and growth (Pauly, 1998; Choat and Roberson, 2002). Altered growth rates caused by the influx of fresh SGD to a coastal marine ecosystem are, therefore, highly likely to modify individual fitness and thereby population sizes in fishes.
Damselfishes are a widespread and abundant component of coastal fish communities around the world. Members of the family frequently serve as model organisms since they share life-history traits with the majority of benthic and coastal fishes and invertebrates (Simpson et al., 2016). Demersal, territorial adults produce pelagic larvae which are dispersed throughout the intertidal zone by tides and currents (Sale, 2002). Juveniles settle at first in small upper littoral rock or tide pools before dispersal onto the upper reef flat takes place (Gopinadha Pillai and Mohan, 1990).
We hypothesize that fresh SGD alters the fitness of primary and secondary consumers through changes in water quality characteristics and enhanced primary productivity in coastal marine ecosystems. We consequently aim at investigating the small-scale effects of fresh SGD on the growth rates of juvenile gray demoiselle Chrysiptera glauca in two contrasting tide pools in Mauritius to predict meso-and large-scale impacts of the process on secondary consumer biomass in coastal ecosystems. Semi-enclosed tide pools influenced by fresh groundwater seepage constitute a valuable field laboratory since these environments exhibit markedly lowered water temperatures, pH, and salinities combined with elevated nutrient loadings due to high water residence times at low tide (Foley, 2018).
Materials and Methods
Study Sites
The intertidal zone of Mauritius is characterized by volcanic rock (Montaggioni, 1982) and serves as a habitat to early life stages of various reef fishes (Sato et al., 2008). Mauritius’ lagoons are highly impacted by dissolved nutrient input from SGD (Burnett et al., 2006; Ramessur et al., 2012). The main source of anthropogenic derived nutrients in fresh SGD in Mauritius are domestic and industrial sewage as well as agricultural activities such as sugar cane farming (Ramessur, 2002). Sampling took place in two tide pools situated at the West coast of Mauritius: one characterized by visible fresh submarine groundwater seepage in Albion (375 m2, 20°12′59.5″S, 57°23′48.2″E) and another strictly marine one in Flic-en-Flac (350 m2, 20°16′17.7″S, 57°22′13.9″E) (Figure 1). Both tide pools were located in lagoons of fringing reefs and sheltered from waves by reef crests. Even though post-larvae of C. glauca are usually recruited to tidal pools all year round, there is a profusion of them during November–December (Gopinadha Pillai and Mohan, 1990). Sampling from October 2017 to January 2018 also allowed for investigating a period with high precipitation rates and strong expected effects from fresh SGD (Oehler et al., 2018) (see Supplementary Table S1).
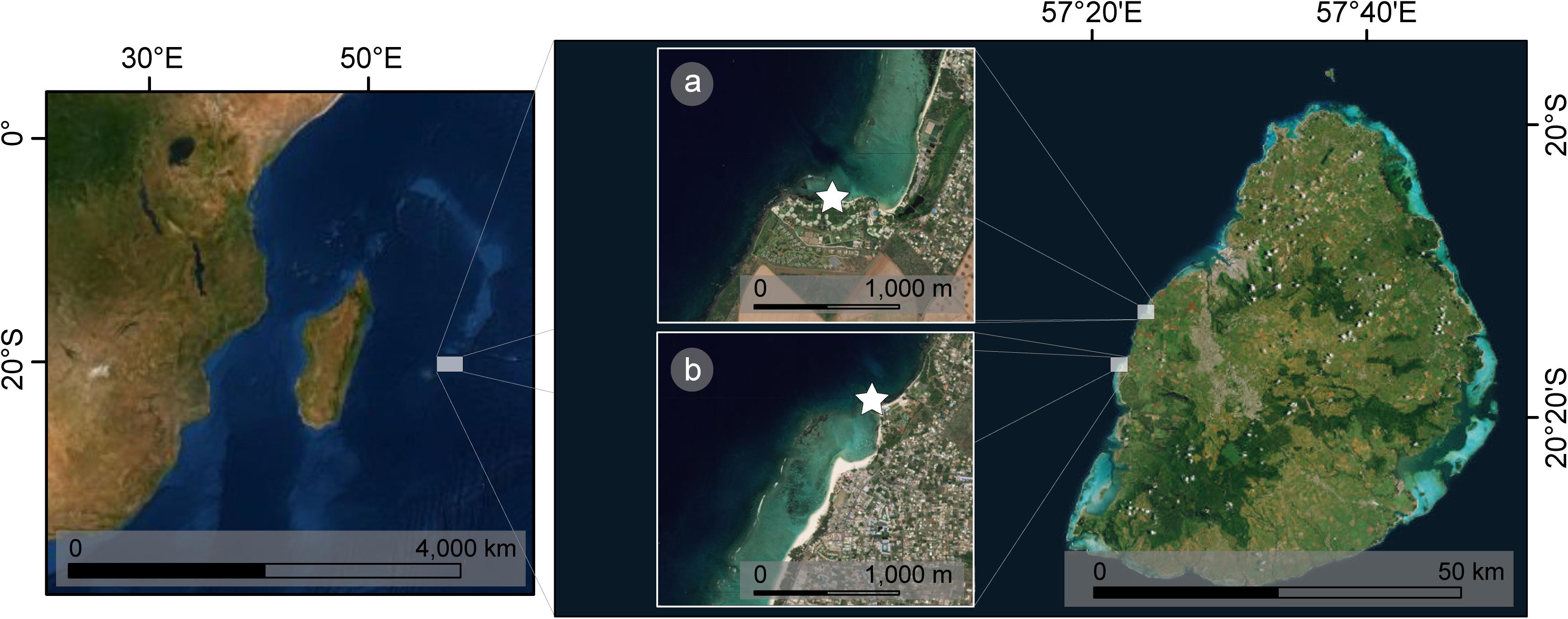
Figure 1. The island of Mauritius in the South-West Indian Ocean with the two sampling tide pools at its West coast: Stars mark the locations of (A) a tide pool influenced by submarine groundwater discharge at Albion, (B) a strictly marine tide pool at Flic-en-Flac.
Environmental Parameters, Substrate Compositions, and Fish Densities
Local influx of fresh SGD is known to significantly alter the water quality characteristics of coastal ecosystems (Moosdorf et al., 2015). We used in situ measurements of physico-chemical parameters to characterize the hydrography and hydrochemistry of the two study tide pools. Water temperature, pH level, oxygen content, and salinity were recorded at both locations during rising tide conditions at mid tide cycle once per month using a WTW multiprobe. Further, hobo loggers were deployed for 24 h inside both tide pools in November 2017. This allowed for the assessment of water depth, temperature, and salinity fluctuations along a full tidal cycle (see Supplementary Figure S1). Water samples for subsequent nutrient measurements were collected in replicates once per month. Per sample 50 ml of seawater were filtered (Sterile Syringe Filter, Corning, CA 0.2 μm), transferred to pre-rinsed centrifuge tubes (with a headspace remaining) and stored frozen (–20°C) in the laboratory. Nutrient analyses of water samples were performed in the laboratory of the Mauritius Oceanography Institute in Albion, Mauritius. Nitrite, nitrate, silicate, and phosphate concentrations were determined using standard methods with a discrete analyser (Systea Easychem Plus) equipped with a 5 cm reading cell.
To evaluate substrate composition, a 50 × 50 cm rectangle containing a grid of 25 10 cm × 10 cm squares was placed randomly 10 times inside each tide pool. Relative substrate cover was assessed visually for each placement. Since visual counts are a common methodology to quantify organism densities in reef environments (Halford and Thompson, 1994), we counted juveniles while wading in transects through each tide pool once per month. We tracked the waded distance via the global positioning system (Garmin GPSMAP 64s) and counted C. glauca individuals in a range of 50 cm to each side. Every sampling month we covered ca. 60 m × 1 m, equivalent to an area of ca. 60 m2. Identification from on top the water surface was possible since juveniles of C. glauca are easily distinguishable from other damselfish species by their gray color and the fluorescent V on the dorsal side of the head (Allen and Steene, 1987). The total number of individuals was set in relation to tide pool area and pooled across sampling months.
Feeding Conditions
Fish were caught with hand nets, stored in ambient seawater for 1–2 h before being euthanized by transfer to a 30% ethanol/seawater solution. In the laboratory, fish were gradually transferred to 50 and 70% ethanol/freshwater solution (Döring et al., 2018). Specimens of C. glauca were measured by standard length (distance between the tip of the snout and the posterior end of the last vertebra, SL, nearest mm), eviscerated, and weighed (EM, ±0.001 g). We identified food organisms to class level. As plants (macrophytes) were among the principal food components in the stomachs of juvenile C. glauca, numerical counts were regarded as not suitable. We, therefore, followed the recommendations from literature and used the frequency of occurrence method for stomach content analysis (Hyslop, 1980). Further, a condition index (CI) was calculated for each individual by using b of the length-weight relationship (Suthers, 1998) (Eq. 1):
Chlorophyll a concentrations were recorded once at the end of the sampling period in January 2018 using a MANTA water quality multi-probe (Eureka Environmental Engineering).
Otolith Preparation and Reading
The lapilli otoliths were dissected under a stereomicroscope, cleaned with deionized water, and stored dry in FEMA-cells (26 × 76 mm). Otoliths were fixed on glass slides using 2-component adhesive (Araldite 2020/A and Araldite 2020/B) and ground sequentially on glass plates by silicon carbide (SiC) powder with grit sizes of 400 and 800. After polishing with waterproof SiC grinding paper (grain size of 5 μm), otoliths in immersion oil were examined with a digital microscope (Keyence VHX-5000) using transmitted light at a magnification of 400×–800×. With the digital microscope it was possible to compose depth-stitchings of different focus levels. Daily deposition of increments on the lapillus otolith has been validated in a number of species from this family (Pitcher, 1988; Thresher et al., 1989; Wellington and Victor, 1989; Thorrold and Milicich, 1990) and we therefore assumed that increments on the lapilli of C. glauca were deposited daily (Wellington and Victor, 1992). The increment closest to the core of the otolith was assumed to be formed at the day of hatching, as is the case in many other species from the family Pomacentridae (Wellington and Victor, 1989). The number of increments was determined from 3 replicate increment counts. The mean count was accepted if the counts deviated by less than 10%, otherwise the otolith was rejected (n = 7) (Wilson and Meekan, 2002). Increment counts were conducted and otolith radius (OR) was measured using ImageJ 1.49 (Rasband, WS, US National Institutes of Health, Bethesda, MD)1. Peripheral otolith growth is a valuable condition index in juvenile reef fishes since it is correlated with RNA/DNA ratios, an indicator for protein biosynthesis in teleost fishes (Clemmesen and Doan, 1996; Suthers, 1998). We, therefore, digitally measured the widths of the last two complete peripheral increments of the otolith. The settlement mark was identified as the first increment of a transition zone, characterized by a rapid narrowing in increment width (Wilson and McCormick, 1997; Retzel et al., 2007) (Supplementary Figure S2).
Growth Model
The relationship between SL and juvenile age (in days) was modeled using the Gompertz growth curve (Stevenson and Campana, 1992) (Eq. 2):
Daily somatic growth rates thus are described with first derivate of Eq. 2 (Döring et al., 2018) (Eq. 3):
where SLt is the larval length at a given time t, SL0 is the fish length on the day of hatching, and k the specific growth rate. G characterizes the exponential decline of the specific growth rate.
Statistical Analyses
Inter tide pool differences in the linear relationships between the log-transformed data on SL and EM, as well as between SL and OR were tested. The assumption of parallel lines was met for the relationship between SL and EM, intercepts were consequently compared using analysis of covariance. Since no significant differences were found, the relationship between SL and EM of all sampled juveniles was described using a single power function. Further, since the assumption of parallel lines was not met, linear regression analysis was performed to describe the relationship between SL and OR for each tide pool individually (Villegas-Hernández et al., 2007). For all data the assumptions of normal distribution (Shapiro–Wilk-test, p > 0.05) and homogenous variances (Bartlett-test, p > 0.05) were tested (Howell, 2007). One-way analysis of variance (ANOVA) was employed to compare the monthly pooled density and age at settlement values between tide pools. Since data did not meet the assumption of equal variances a nonparametric Mann-Whitney-U-test was used to test for inter tide pool differences in chlorophyll a concentrations. Factorial ANOVA was employed to explore the effects of the categorical predictors tide pool and sampling month on the explanatory variables CI, mean peripheral otolith increment width, and somatic growth rates. To achieve normal distribution and variance homogeneity, data on somatic growth rates were log-transformed. All statistical analyses were carried out in JMP Pro 14.0.0 (SAS Institute Inc., Cary, NC)2. Sample sizes for all conducted statistical analyses are listed in Table 1.
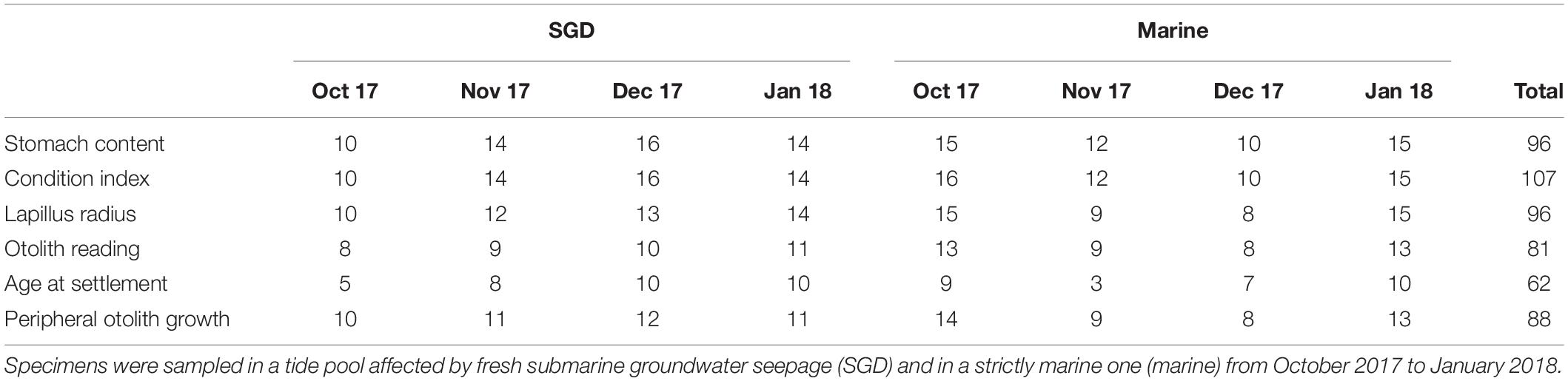
Table 1. Number of juvenile Chrysiptera glauca used for the respective analysis.
Results
Tide Pool Hydrography and Hydrochemistry
Water temperature and pH in the tide pool at Albion steadily increased from October 2017 to January 2018 and were usually markedly lower than in the tide pool at Flic-en-Flac (Figures 2A,B). Water in both tide pools was generally supersaturated with oxygen and dissolved oxygen content was higher in the tide pool at Flic-en-Flac as opposed to the one at Albion in all months except November (Figure 2C). Salinity in the tide pool at Flic-en-Flac continuously ranged around 35 while in the tide pool at Albion salinity was below 20 in most months (Figure 2D). Nitrite, Nitrate, and Silicate concentrations were generally substantially higher in the tide pool at Albion than in the one at Flic-en-Flac. All three parameters exhibited increasing trends inside both tide pools throughout the sampling period (Figures 2E–G). Phosphate concentrations, on the other hand, were very similar and exhibited decreasing trends from October to January (Figure 2H).
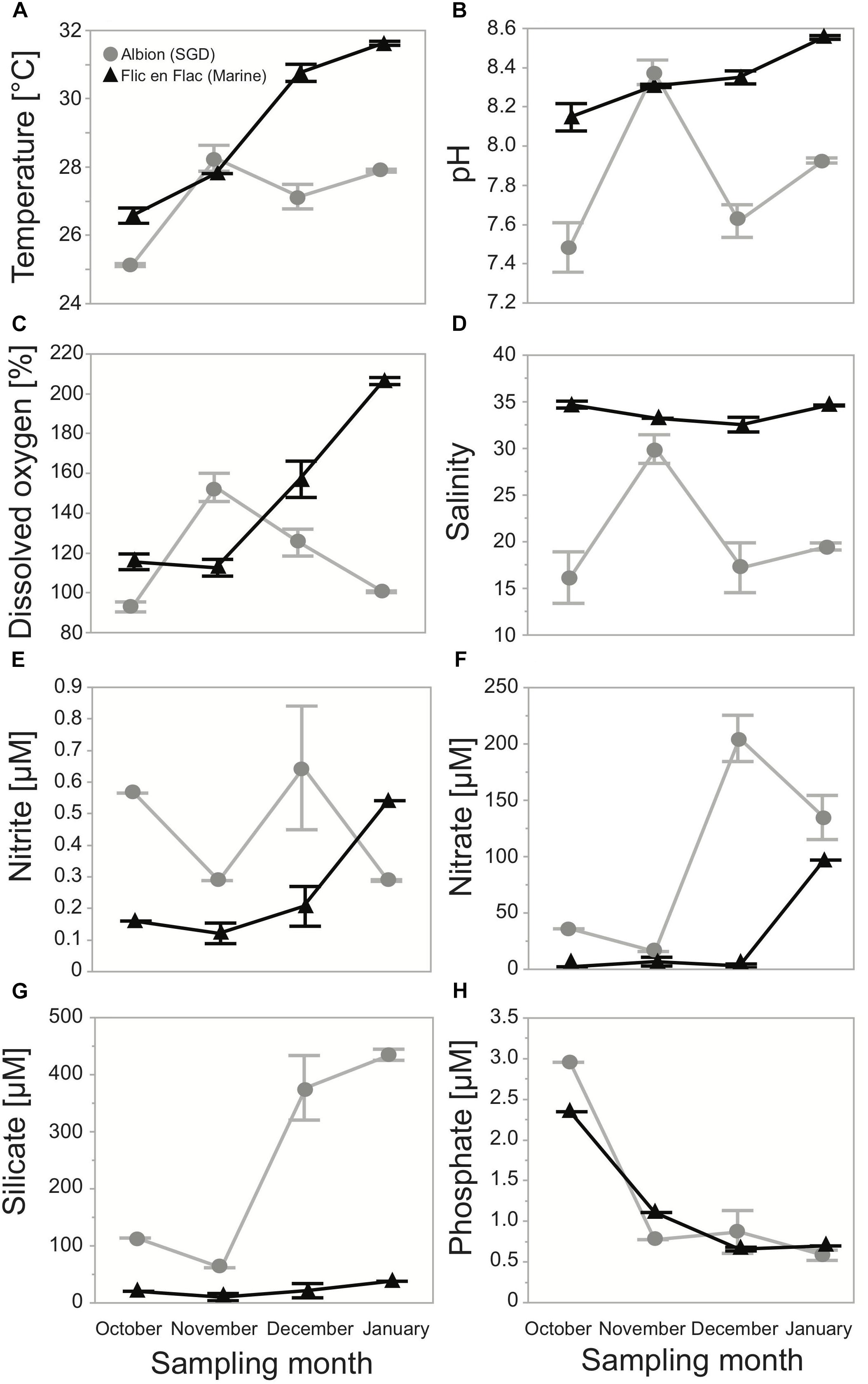
Figure 2. Monthly time series of physico-chemical parameters recorded inside a tide pool influenced by fresh SGD at Albion (SGD, gray dots) and inside a strictly marine tide pool at Flic-en-Flac (Marine, black triangles): (A) Water temperature, (B) water pH, (C) water dissolved oxygen content, (D) salinity, (E) nitrite, (F) nitrate, (G) silicate, and (H) phosphate concentrations.
In November environmental parameters were, as an exception, sampled closer to high tide, explaining the divergent salinity, oxygen, and pH levels for this month (Supplementary Figure S1). All these observations suggest a strong influx of fresh SGD into the Albion tide pool hence this tide pool will be referred to as influenced by fresh “SGD.” Contrary, the Flic-en-Flac tide pool provided strictly marine environmental conditions for the study species and this tide pool is henceforth called “marine.”
Benthic Cover and Juvenile Densities
Relative benthic cover compositions were quite different between both tide pools. While no sessile algae were recorded in the tide pool influenced by fresh SGD, the bottom of the marine tide pool was to a great extant covered by macroalgae and turfalgae (Table 2). We also found that C. glauca densities were not significantly different in the two study tide pools (ANOVA, F(1, 8) = 0.2497, p = 0.6326) (Table 2).

Table 2. Mean benthic cover composition (% ±SD) and juvenile Chrysiptera glauca densities (individuals 100 m–2 ±SD) in a tide pool affected by fresh submarine groundwater seepage (SGD) and in a strictly marine one (marine).
Feeding Conditions
Almost all juveniles examined for the present study fed on a mix of filamentous green algae, crustacean copepods, moluscs, and plant detritus. Moreover, the diet occasionally included calcifying corals, gastropods, insects/arachnids of terrestrial origin, and ichthyoplankton. Plastic particles or fibers were found in the stomachs of 16% of all fish (Table 3). Two factorial ANOVA revealed no significant differences in juvenile CI throughout tide pools and months (F(7, 104) = 1.70, p = 0.12). Mean chlorophyll a concentrations [± standard deviation (SD)] were significantly higher in the strictly marine tide pool (0.50 ± 0.20 μg/l) when compared to the tide pool influenced by SGD (0.39 ± 0.07 μg/l) (Mann–Whitney-U-test, Z = –2.23, n1 = 86 and n2 = 156, p < 0.05).
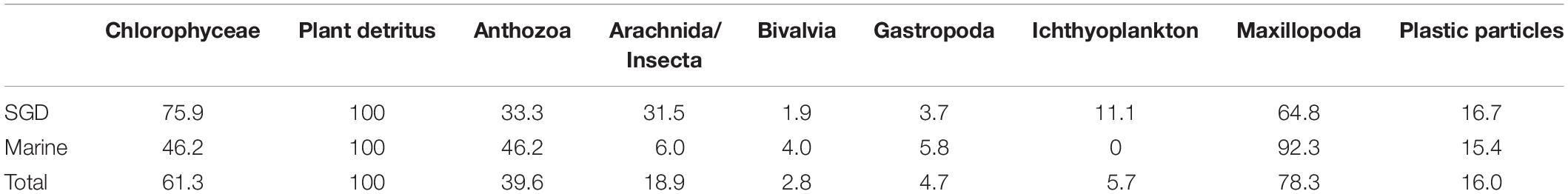
Table 3. Frequency of occurrence of food organisms (to phylogenetic class level) and particles in the stomachs of Chrysiptera glauca (as percentage of all juveniles) sampled in either a tide pool influenced by fresh submarine groundwater discharge (SGD), in a strictly marine tide pool (marine), as well as in both tide pools combined (total).
Otolith Growth, Somatic Growth, and Settlement
Since there was no observable spatial difference in the relationship between juvenile standard length (Figure 3) and eviscerated mass (EM), a common power function is given as follows: EM = 0.0019 × SL3.4507, r2 = 0.99.
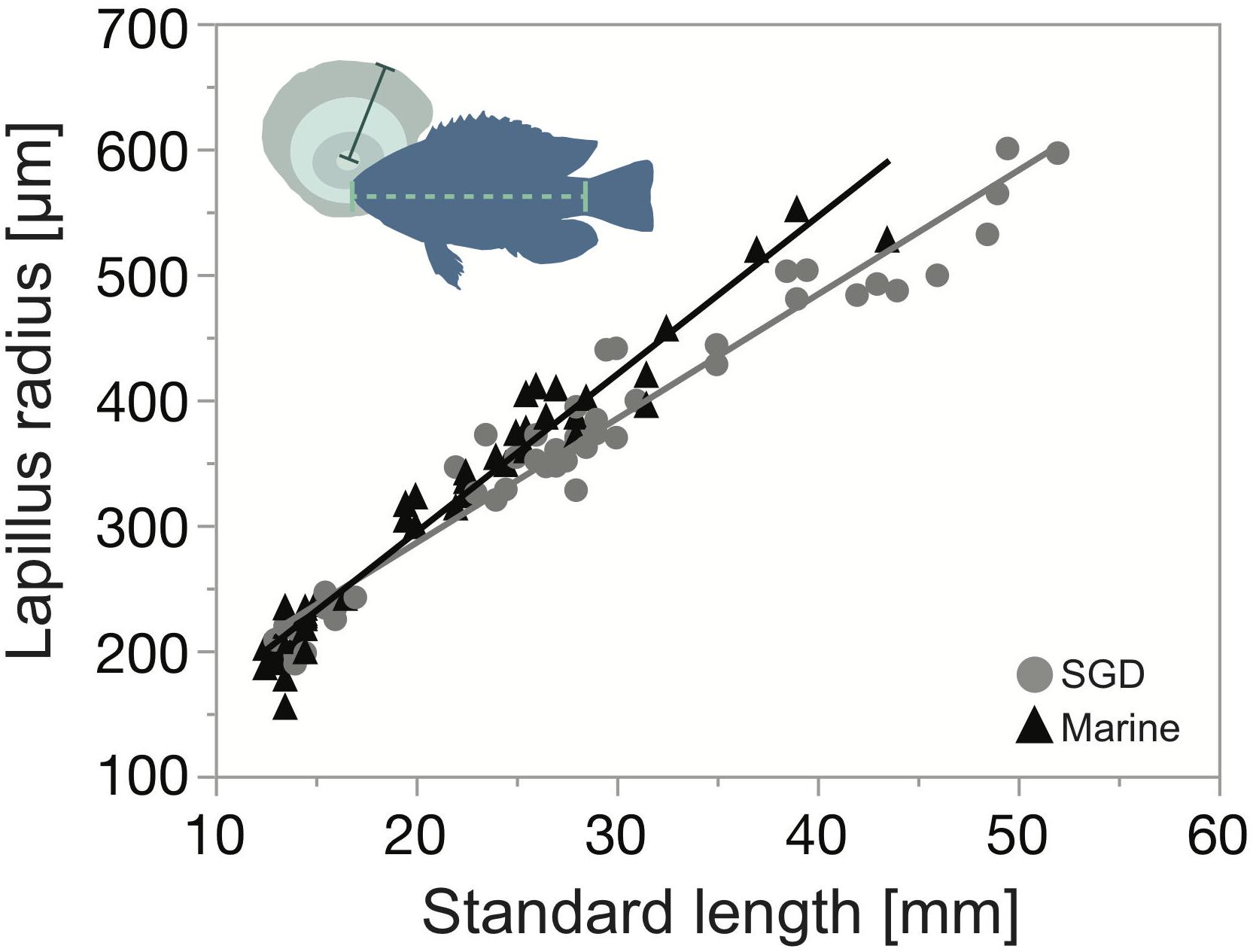
Figure 3. Relationship between lapillus (green) radius (dark green solid line) and juvenile Chrysiptera glauca (blue silhouette) standard length (light green dashed line) inside a tide pool influenced by fresh submarine groundwater seepage (SGD, gray circles), and in a strictly marine tide pool (marine, black triangles).
Two factorial ANOVA showed no significant differences in juvenile peripheral otolith growth throughout tide pools and months (F(7, 87) = 1.00, p = 0.44). The linear relationships between SL and OR showed a proportionality between otolith growth and fish somatic growth (inside the fresh SGD influenced tide pool: OR = 87.140321 + 9.9018163 × SL, r2 = 0.95, and inside the strictly marine tide pool: OR = 42.1809 + 12.571021 × SL, r2 = 0.95) (Figure 3).
For each sampling location we computed a distinctive Gompertz growth function (Figure 4): SGD tide pool: SLt = 7.26 × e2.85 (1−e(0.0099 × t)), r2 = 0.89; marine tide pool: SLt = 7.21 × e2.78 (1−e(0.0083 × t)), r2 = 0.91. The first derivative of the Gompertz growth models gave us the daily somatic growth rates, which served to back-calculate the amount of somatic tissue individuals were able to accumulate in both tide pools per day. For all months, somatic growth rates were significantly higher in juveniles sampled in the fresh SGD influenced tide pool when compared to their conspecifics sampled in the marine tide pool. Also, in both tide pools juvenile somatic growth rates were significantly higher in November and January than in October (ANOVA, F(7, 78) = 32.2585, p < 0.0001, Tukey HSD, p < 0.05).
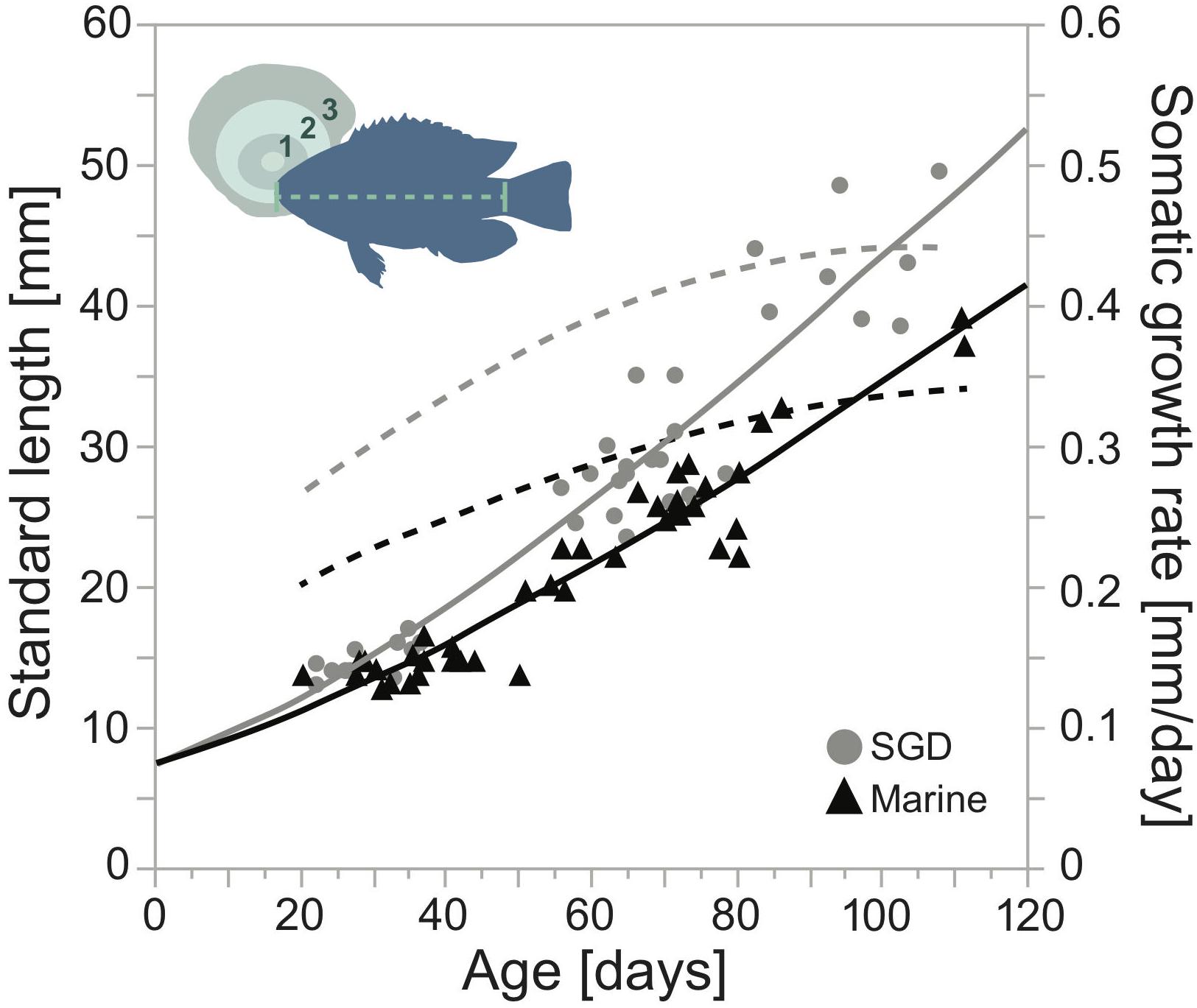
Figure 4. Relationship between standard length (light green dashed line, mm), somatic growth rates (mm per day), and age (days) as deducted from otolith (green) increment counts (dark green numbers) in juvenile Chrysiptera glauca (blue silhouette). Gray and black solid lines represent Gompertz growth curves (GGC), gray and black dashed lines represent growth rates (mm per day) calculated from first derivative of the GGC. Gray circles indicate individuals sampled inside a tide pool influenced by fresh submarine groundwater seepage (SGD), and black triangles those sampled in a strictly marine tide pool (marine).
Chrysiptera glauca individuals sampled in the SGD influenced tide pool settled at a significantly younger age (19.9 ± 2.8 days, n = 33) than their counterparts taken from the strictly marine tide pool (22.1 ± 2.7 days, n = 29) (ANOVA, F(1, 61) = 9.67, p < 0.01).
Discussion
While the impact of fresh SGD on benthic and pelagic primary producer community structuring is rather well studied, little is known about the process’ effects on the fitness of consumers in higher trophic levels such as teleost fish (Lecher and Mackey, 2018). Determination of factors influencing the fitness of fish in coastal environments is, however, of consistently high concern to make predictions about anthropogenic impacts on marine ecosystems and, ultimately, on fisheries’ productivity. We show that somatic growth rates were significantly higher and settlement to reef habitat took place significantly earlier in juvenile gray demoiselle C. glauca exposed to fresh SGD when compared to strictly marine conditions. Since feeding conditions were comparable among tide pools, we propose that physiologically beneficial environmental conditions rather than food availability elevate the survival potential of marine fish exposed to fresh SGD.
The encountered cold, acidic, nutrient-rich fresh SGD is known to condition benthic marine macro faunal communities (Amato et al., 2016; Foley, 2018; Piló et al., 2018). The interactions between substrate for algae cultivation, macrophytic food abundance, and damselfish densities can be manifold (Wellington and Victor, 1985; Ceccarelli et al., 2005; Ceccarelli, 2007; Hoey and Bellwood, 2010) and intraspecific density dependent effects were additionally shown to modify growth in damselfish (Booth, 1995). We followed up on this relationship by visually assessing substrate cover composition and C. glauca densities within each tide pool. The strictly marine tide pool was to a great extant covered with macroalgae, while these benthic primary producers were completely absent from the tide pool influenced by SGD. Due to similar values in shelter providing rocks and in juvenile densities, it can be suspected that density dependent mortality as well as predator induced stress are comparable in both tide pools (Schmitt and Holbrook, 1999; Holbrook and Schmitt, 2002).
Feeding conditions are proposed to be directly related to somatic wealth and growth in fishes (Jones, 1986), and are thus expected to have important ramifications for juvenile survival and recruitment (Hoey and McCormick, 2004). Further, nutrient rich fresh SGD has the potential to elevate chlorophyll a levels and macrophyte abundances in coastal marine ecosystems (Machado and Imberger, 2014; Welti et al., 2015; Amato et al., 2016; Honda et al., 2018). To test for inter tide pool differences in feeding conditions, we investigated juvenile stomach contents and nutritional condition as well as chlorophyll a concentrations. In juvenile reef fishes, otolith growth rates as condition indices are regarded as superior to morphometric (e.g., length to mass) relationships (Suthers, 1998). We, therefore, analyzed somatic energy storage (i.e., CI) combined with peripheral otolith growth to assess the nutritional wealth of C. glauca individuals in the two study tide pools. Both indices showed neither significant spatial nor temporal differences. C. glauca is omnivorous and demonstrably able to utilize a variety of food sources (Hiatt and Strasburg, 1960; Gopinadha Pillai and Mohan, 1990). Stomach contents analysis showed that C. glauca’s diet is highly dependent on green algae and plant detritus. Taking into account the markedly higher benthic algae abundance and chlorophyll a levels in the marine tide pool, and that nutritional condition (CI and peripheral otolith growth) was temporally and spatially constant, feeding conditions for C. glauca could not be assumed to be more favorable in the SGD tide pool.
Back-calculation of size from otoliths assumes that there is proportionality between otolith and somatic growth rates (Vigliola et al., 2000). We verified this assumption by calculating a regression relationship between otolith radius and SL for each tide pool. Because of a lower slope inside the tide pool influenced by fresh SGD, a significant interaction between somatic growth and site was detected. An otolith growth uncoupled from somatic growth in acidic waters was previously described in juvenile ocellated wrasse Symphodus ocellatus and attributed to the “growth effect” (Di Franco et al., 2019): slower-growing individuals have a tendency to have larger otoliths than their faster-growing counterparts at the same length (Campana, 1990).
In reef fishes it is likely that linkages between early life stages occur when there is selection for a cumulative trait, e.g., body size. They can be effective not only between life history stages of the same individual, but also between generations through maternal effects, e.g., size at hatching (Leis and McCormick, 2002). In brown demoiselle Neopomacentrus filamentosus individuals that survived intense selective mortality 1–3 months after settlement were those fish that were larger at hatching and the ones that grew faster during planktonic life (Vigliola and Meekan, 2002). In our case a strong carry over effect of larval fitness at hatching on juvenile growth cannot be expected since length at hatching did not differ markedly between both sampled populations (SGD tide pool: 7.26 mm; marine tide pool: 7.21 mm).
Predatory mortality severely affects population sizes and is often highest for the youngest recruits (Hixon, 1991; Almany and Webster, 2006). A rapid increase in body length, however, generally enhances the survival chances in marine fish early life stages [growth-mortality hypothesis (Anderson, 1988; Houde, 1989)]. Somatic growth rates at a given age were in the range of previously published values for the species (Gopinadha Pillai and Mohan, 1990) and consistently higher in juveniles sampled in the SGD influenced tide pool than in juveniles taken from the marine tide pool. Previous studies in damselfish have shown that faster growing larvae settle at a younger age (Thorrold and Milicich, 1990). Settlement in C. glauca occurred inside a time frame frequently observed in damselfishes (Wellington and Victor, 1989). Further, the faster growing C. glauca individuals sampled in the SGD influenced tide pool indeed settled at a significantly younger age when compared to their slower growing counterparts taken from the strictly marine tide pool. Earlier settlement decreases the planktonic larval duration, a life history stage particularly prone to high mortality rates (Leis, 1991). Thus, the observed elevated somatic growth rates and earlier settlement can be expected to elevate survival potential and ultimately confer higher fitness to individuals sampled in the fresh SGD influenced tide pool as compared to the strictly marine tide pool (Houde, 1989; Thorrold and Milicich, 1990; Hoey and McCormick, 2004).
The interplay between food levels, metabolism, and somatic growth may be complex (Auer et al., 2015), but direct effects of environmental parameters on growth and survival in marine fish are generally regarded as stronger than the indirect ones exhibited by enhanced food availability (Houde, 2008). Additionally, we show that feeding conditions were comparable in both study tide pools. Our results, therefore, lead us to conclude that physiologically beneficial environmental conditions brought about by the submarine influx of cold acidic freshwater enabled juvenile fish to exhibit elevated growth rates in the SGD influenced tide pool.
Even in waters well saturated with oxygen, increased temperatures caused by climate change will act as stressors on low-latitude fishes, hampering growth via metabolic constraints (Rodgers et al., 2018). The submarine influx of cold freshwater, on the other hand, may act as a buffer against elevated water temperatures, thereby enhancing growth in ectothermic organisms. Further, teleost fishes almost always exhibit better growth rates in intermediary salinity conditions (Boeuf and Payan, 2001). This has been correlated with a lower standard metabolic rate and explained by a salinity dependent food conversion efficiency (Kinne, 1960; Imsland et al., 2001). Lowered pH also tends to increase the growth rates in marine fish (Di Franco et al., 2019; Jarrold and Munday, 2019). This could potentially be explained by either increased energy intake or reduced energy expenditure in acidified waters (Munday et al., 2009). First evidence of enhanced growth in temperate juvenile marbled sole Pseudopleuronectes yokohamae caused by the influx of nutrient rich fresh SGD was presented only recently by Fujita et al. (2019). The authors proposed a positive relationship between fish growth, SGD derived nutrient loadings, and elevated primary producer as well as primary consumer (i.e., prey) abundances but were unable to conclusively substantiate a connection between these trophic levels. We, on the other hand, propose that elevated somatic growth rates in juvenile fish subjected to fresh SGD are caused by a combination of the aforementioned physiological effects and not necessarily food availability.
Past studies have shown that small-scale field experiments are suitable to estimate fitness of reef fishes at large scales (Steele and Forrester, 2005). Still, it remains to be determined how much the local variations in water quality caused by fresh SGD affect the biomass of marine secondary consumers on larger geographical scales. Investigations combining otolith microchemistry (e.g., oxygen isotopy) and age reading are warranted to further assess whether even non-territorial fishes exposed to fresh SGD exhibit signs of increased survival potential (Thorrold et al., 1997; Kim and Lee, 2003). Due to elevated primary productivity, food is not necessarily a limited resource in coastal ecosystems influenced by fresh SGD (Jones, 1986; Lecher and Mackey, 2018). Thus, even small changes in ambient hydrography and hydrochemistry caused by the influx of fresh groundwater will lead to differences in growth, which - over time - will translate into significant differences in population sizes (Retzel et al., 2007), elevating the biomass available to a fishery.
A range of stressors increasingly threatens coastal marine ecosystems, yet these habitats provide livelihoods through fisheries to 260 million people (Teh and Sumaila, 2013). Anthropogenic modifications to the hydrography and hydrochemistry of coastal marine ecosystems through altered groundwater fluxes will have significant implications for a system’s carrying capacity and fisheries productivity (Burnett et al., 2018). Given the ubiquitous nature of fresh SGD (Taniguchi et al., 2002) our work highlights the need for groundwater fluxes to be included in environmental management plans. It furthermore illuminates future challenges such as balancing anthropogenic freshwater use and coastal fisheries’ productivity, particularly given the potential for climate change to aggravate freshwater scarcity.
Data Availability Statement
Biological data that support the findings of this study have been deposited in PANGAEA and are accessible at https://doi.pangaea.de/10.1594/PANGAEA.897645.
Ethics Statement
Research was carried out in accordance to Mauritian laws and regulations and under direct supervision of scientist of the Mauritius Oceanography Institute (MOI). The animal study was reviewed and approved by the MOI Board. The employed sampling strategy ensured that the fish stock at those study sites was not depleted – therefore, as a conservation effort and to minimize the impact on the fish population, the monthly sample size was kept relatively small. No non-target or by-catch specimens were collected during the study.
Author Contributions
JL and NM conceived the research and prepared the initial manuscript. JL and TP designed the sampling. JL, TP, DD, and YN conducted the field work. TP and YN measured the physico-chemical parameters. SN and SB processed the fish and prepared the otoliths for reading. SB conducted the stomach content analysis. JL performed the otolith readings and statistically analyzed the data. All authors contributed to later revisions.
Funding
This study was funded by the German Federal Ministry of Education and Research (BMBF; Grant #01LN1307A awarded to NM).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank Elisabeth Myers, Gladys M. Okemwa, and Andreas Kunzmann for help in species identification; Anishta Audit-Manna, Chetanand Samyan, and Shane Sunassee for their help in sample collection and analytical procedures; Rebecca Gorniak for her help in fish dissection and otolith preparation; Sebastian Flotow for his valuable support in otolith preparation and in digital imagery; and Werner Ekau for help regarding statistical applications. We would like to express our gratitude to Ruby M. Pillay for administrative support in conducting this study.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2019.00613/full#supplementary-material
Footnotes
References
Allen, G. R., and Steene, R. C. (1987). Reef Fishes of the Indian Ocean. Neptune City, NJ: T.F.H. Publications.
Almany, G. R., and Webster, M. S. (2006). The predation gauntlet: early post-settlement mortality in reef fishes. Coral Reefs 25, 19–22. doi: 10.1007/s00338-005-0044-y
Amato, D., Bishop, J. M., Glenn, C. R., Dulai, H., and Smith, C. M. (2016). Impact of submarine groundwater discharge on marine water quality and reef biota of Maui. PLoS One 11:e016582. doi: 10.1371/journal.pone.0165825
Anderson, J. T. (1988). A review of size dependant survival during pre-recruit stages of fishes in relation to recruitment. J. Northwest Atl. Fish. Sci. 8, 55–66. doi: 10.2960/j.v8.a6
Auer, S. K., Salin, K., Rudolf, A. M., Anderson, G. J., and Metcalfe, N. B. (2015). The optimal combination of standard metabolic rate and aerobic scope for somatic growth depends on food availability. Funct. Ecol. 29, 479–486. doi: 10.1111/1365-2435.12396
Boeuf, G., and Payan, P. (2001). How should salinity influence fish growth? Comp. Biochem. Physiol. - C Toxicol. Pharmacol. 130, 411–423. doi: 10.1016/S1532-0456(01)00268-X
Booth, D. J. (1995). Juvenile groups in a coral-reef damselfish: density-dependent effects on individual fitness and population demography. Ecology 76, 91–106. doi: 10.2307/1940634
Burnett, K. M., Wada, C. A., Taniguchi, M., Sugimoto, R., and Tahara, D. (2018). Evaluating the tradeoffs between groundwater pumping for snow-melting and nearshore fishery productivity in Obama City, Japan. Water 10:1556. doi: 10.3390/w10111556
Burnett, W. C., Aggarwal, P. K., Aureli, A., Bokuniewicz, H., Cable, J. E., Charette, M. A., et al. (2006). Quantifying submarine groundwater discharge in the coastal zone via multiple methods. Sci. Total Environ. 367, 498–543. doi: 10.1016/j.scitotenv.2006.05.009
Burnett, W. C., Bokuniewicz, H., Huettel, M., Moore, W. S., and Taniguchi, M. (2003). Groundwater and pore water inputs to the coastal zone. Biogeochemistry 66, 3–33. doi: 10.1023/b:biog.0000006066.21240.53
Campana, S. E. (1990). How reliable are growth back-calculations based on otoliths? Can. J. Fish. Aquat. Sci. 47, 2219–2227. doi: 10.1139/f90-246
Ceccarelli, D. M. (2007). Modification of benthic communities by territorial damselfish: a multi-species comparison. Coral Reefs 26, 853–866. doi: 10.1007/s00338-007-0275-1
Ceccarelli, D. M., Jones, G. P., and McCook, L. J. (2005). Effects of territorial damselfish on an algal-dominated coastal coral reef. Coral Reefs 24, 606–620. doi: 10.1007/s00338-005-0035-z
Choat, J., and Roberson, D. (2002). “Age-Based Studies,” in Coral reef fishes: dynamics and diversity in a complex ecosystem. ed P. F. Sale (San Diego, CA: Academic Press), 57–80. doi: 10.1016/b978-012615185-5/50005-0
Clemmesen, C., and Doan, T. (1996). Does otolith structure reflect the nutritional condition of a fish larva? Comparison of otolith structure and biochemical index (RNA/DNA ratio) determined on cod larvae. Mar. Ecol. Prog. Ser. 138, 33–39. doi: 10.3354/meps138033
Dale, R. K., and Miller, D. C. (2008). Hydrologic interactions of infaunal polychaetes and intertidal groundwater discharge. Mar. Ecol. Prog. Ser. 363, 205–215. doi: 10.3354/meps07455
Di Franco, A., Calo, A., Sdiri, K., Cattano, C., Milazzo, M., and Guidetti, P. (2019). Ocean acidification affects somatic and otolith growth relationship in fish: evidence from an in situ study. Biol. Lett 15:20180662. doi: 10.1098/rsbl.2018.0662
Döring, J., Neumann, S. I., Sloterdijk, H., and Ekau, W. (2018). Seasonal growth differences of larval Hyporhamphus picarti (Hemiramphidae) in the Sine Saloum estuary, Senegal. J. Appl. Ichthyol 34, 97–102. doi: 10.1111/jai.13528
Encarnação, J., Leitão, F., Range, P., Piló, D., Chícharo, M. A., and Chícharo, L. (2014). Local and temporal variations in near-shore macrobenthic communities associated with submarine groundwater discharges. Mar. Ecol 36, 926–941. doi: 10.1111/maec.12186
Foley, L. (2018). Karst-channelled intertidal submarine groundwater discharge (SGD) conditions the form of the rock pool sessile assemblage. Estuar. Coast. Shelf Sci. 213, 236–244. doi: 10.1016/J.ECSS.2018.08.014
Fujita, K., Shoji, J., Sugimoto, R., Nakajima, T., Honda, H., Takeuchi, M., et al. (2019). Increase in fish production through bottom-up trophic linkage in coastal waters induced by nutrients supplied via submarine groundwater. Front. Environ. Sci. 7:82. doi: 10.3389/fenvs.2019.00082
Gopinadha Pillai, C. S., and Mohan, M. (1990). Ecology and biology of Abudefduf glaucus (Cuvier) (Pomacentridae, Pisces) from minicoy atoll, Lakshadweep. Indian J. Fish. 37, 15–23.
Halford, A. R., and Thompson, A. A. (1994). “Visual census surveys of reef fish,” in Long-term Monitoring of the Great Barrier Reef - Standard Operational Procedure Number 3, (Townsville: Australian Institute of Marine Science).
Hata, M., Sugimoto, R., Hori, M., Tomiyama, T., and Shoji, J. (2016). Occurrence, distribution and prey items of juvenile marbled sole Pseudopleuronectes yokohamae around a submarine groundwater seepage on a tidal flat in southwestern Japan. J. Sea Res. 111, 47–53. doi: 10.1016/j.seares.2016.01.009
Hiatt, R. W., and Strasburg, D. W. (1960). Ecological relationships of the fish fauna on coral reefs of the Marshall Islands. Ecol. Monogr. 30, 65–127. doi: 10.2307/1942181
Hixon, M. A. (1991). “Predation as a process structuring coral reef fish communities,” in The Ecology of Fishes on Coral Reefs, ed. P. F. Sale (San Diego: Academic Press), 475–508. doi: 10.1016/b978-0-08-092551-6.50022-2
Hoey, A. S., and Bellwood, D. (2010). Damselfish territories as a refuge for macroalgae on coral reefs. Coral Reefs 29, 107–118. doi: 10.1007/s00338-009-0567-8
Hoey, A. S., and McCormick, M. I. (2004). Selective predation for low body condition at the larval-juvenile transition of a coral reef fish. Oecologia 139, 23–29. doi: 10.1007/s00442-004-1489-3
Holbrook, S. J., and Schmitt, R. J. (2002). Competition for shelter space causes density-dependent predation mortality in damselfishes. Ecology 83, 2855–2868. doi: 10.1890/0012-9658(2002)083%5B2855:cfsscd%5D2.0.co;2
Honda, H., Sugimoto, R., and Kobayashi, S. (2018). “Submarine groundwater discharge and its influence on primary production in japanese coasts: case study in Obama Bay,” in The Water-Energy-Food Nexus: Human-Environmental Security in the Asia-Pacific Ring of Fire, eds A. Endo, and T. Oh (Singapore: Springer), 101–115. doi: 10.1007/978-981-10-7383-0_8
Houde, E. D. (1989). Comparative growth, mortality, and energetics of marine fish larvae: temperature and implied latitudinal effects. Fish. Bull. 87, 471–495.
Houde, E. D. (2008). Emerging from Hjort’s shadow. J. Northwest Atl. Fish. Sci. 41, 53–70. doi: 10.2960/J.v41.m634
Hyslop, E. J. (1980). Stomach contents analysis - a review of methods and their application. J. Fish Biol. 17, 411–429. doi: 10.1111/j.1095-8649.1980.tb02775.x
Imsland, A. K., Foss, A., Gunnarsson, S., Berntssen, M. H., FitzGerald, R., Bonga, S. W., et al. (2001). The interaction of temperature and salinity on growth and food conversion in juvenile turbot (Scophthalmus maximus). Aquaculture 198, 353–367. doi: 10.1016/s0044-8486(01)00507-5
Jarrold, M. D., and Munday, P. L. (2019). Diel CO2 cycles and parental effects have similar benefits to growth of a coral reef fish under ocean acidification. Biol. Lett. 15:20180724. doi: 10.1098/rsbl.2018.0724
Johannes, R. E., and Hearn, C. (1985). The effect of submarine groundwater discharge on nutrient and salinity regimes in a coastal lagoon off Perth. Western Australia. Estuar. Coast. Shelf Sci. 21, 789–800. doi: 10.1016/0272-7714(85)90073-3
Jones, G. P. (1986). Food availability affects growth in a coral reef fish. Oecologia 70, 136–139. doi: 10.1007/BF00377123
Kim, G., and Lee, K.-K. (2003). Large submarine groundwater discharge (SGD) from a volcanic island. Geophys. Res. Lett. 30, 1–4. doi: 10.1029/2003gl018378
Kinne, O. (1960). Growth, food intake, and food conversion in a euryplastic fish exposed to different temperatures and salinities. Physiol. Zool. 33, 288–317. doi: 10.1086/664584
Lapointe, B. E., and O’Connell, J. (1989). Nutrient-enhanced growth of Cladophora prolifera in Harrington Sound, Bermuda: eutrophication of a confined, phosphorus-limited marine ecosystem. Estuar. Coast. Shelf Sci. 28, 347–360. doi: 10.1016/0272-7714(89)90084-X
Lecher, A. L., and Mackey, K. R. M. (2018). Synthesizing the effects of submarine groundwater discharge on marine biota. Hydrobiologia 5:60. doi: 10.3390/hydrology5040060
Leis, J. M. (1991). “The pelagic stage of reef fishes: the larval biology of coral fishes,” in The Ecology of Fishes on Coral Reefs, ed. P. F. Sale (San Diego: Academic Press), 277–278.
Leis, J. M., and McCormick, M. I. (2002). “The biology, behavior, and ecology of the pelagic, larval stage of coral reef fishes,” in Coral Reef Fishes: Dynamic and Diversity in a Complex Ecosystem, ed. P. F. Sale (San Diego, CA: Elsevier), 171–199. doi: 10.1016/b978-012615185-5/50011-6
Luo, X., Jiao, J. J., Liu, Y., Zhang, X., Liang, W., and Tang, D. (2018). Evaluation of water residence time, submarine groundwater discharge, and maximum new production supported by groundwater borne nutrients in a coastal upwelling shelf system. J. Geophys. Res. Ocean. 123, 631–655. doi: 10.1002/2017JC013398
Machado, D. A., and Imberger, J. (2014). Modeling the impact of natural and anthropogenic nutrient sources on phytoplankton dynamics in a shallow coastal domain. Western Australia. Environ. Fluid Mech. 14, 87–111. doi: 10.1007/s10652-013-9296-1
Montaggioni, L. F. (1982). Pleistocene marine depositional environments from Mauritius island, Indian Ocean. Geobios 15, 161–179. doi: 10.1016/s0016-6995(82)80018-1
Moore, W. S. (2010). The effect of submarine groundwater discharge on the ocean. Ann. Rev. Mar. Sci. 2, 59–88. doi: 10.1146/annurev-marine-120308
Moosdorf, N., Stieglitz, T., Waska, H., Dürr, H. H., and Hartmann, J. (2015). Submarine groundwater discharge from tropical islands: a review. Grundwasser 20, 53–67. doi: 10.1007/s00767-014-0275-3
Munday, P. L., Crawley, N. E., and Nilsson, G. E. (2009). Interacting effects of elevated temperature and ocean acidification on the aerobic performance of coral reef fishes. Mar. Ecol. Prog. Ser. 388, 235–242. doi: 10.3354/meps08137
Oehler, T., Eiche, E., Putra, D., Adyasari, D., Hennig, H., Mallast, U., et al. (2018). Timing of land-ocean groundwater nutrient fluxes from a tropical karstic region (southern Java, Indonesia). J. Hydrol. 565, 662–671. doi: 10.5194/hess-2017-621
Pauly, D. (1998). Tropical fishes: patterns and propensities. J. Fish Biol. 53, 1–17. doi: 10.1006/jfbi.1998.0810
Piló, D., Barbosa, A. B., Teodósio, M. A., Encarnação, J., Leitão, F., Range, P., et al. (2018). Are submarine groundwater discharges affecting the structure and physiological status of rocky intertidal communities? Mar. Environ. Res. 136, 158–173. doi: 10.1016/j.marenvres.2018.02.013
Pitcher, C. R. (1988). Validation of a technique for reconstructing daily patterns in the recruitment of coral reef damselfish. Coral Reefs 7, 105–111. doi: 10.1007/BF00300969
Ramessur, R. T. (2002). Anthropogenic-driven changes with focus on the coastal zone of Mauritius, south-western Indian Ocean. J. Mater. Cycles Waste Manag. 3, 99–106. doi: 10.1007/s10113-002-0045-0
Ramessur, R. T., Boodhoo, K., Balgobin, J., Povinec, P., and Burnett, W. C. (2012). Dissolved nutrients from submarine groundwater in Flic en Flac lagoon, Mauritius. West. Indian Ocean J. Mar. Sci. 10, 121–127.
Retzel, A., Hansen, A. D., and Grønkjær, P. (2007). Otolith growth of Springer’s demoiselle, Chrysiptera springeri (Pomacentridae, Allen & Lubbock), on a protected and non-protected coral reef. J. Appl. Ichthyol. 23, 568–572. doi: 10.1111/j.1439-0426.2007.00865.x
Rodgers, G., Rummer, J., Johnson, L., and McCormick, M. (2018). Impacts of increased ocean temperatures on a low-latitude coral reef fish – Processes related to oxygen uptake and delivery. J. Therm. Biol 79, 95–102. doi: 10.1016/j.jtherbio.2018.12.008
Sale, P. F. (ed.) (2002). Coral Reef Fishes - Dynamics and Diversity in a Complex Ecosystem. Amsterdam: Academic Press.
Sato, N., Asahida, T., Terashima, H., Hurbungs, M. D., and Ida, H. (2008). Species composition and dynamics of larval and juvenile fishes in the surf zone of Mauritius. Environ. Biol. Fish. 81, 229–238. doi: 10.1007/s10641-007-9194-9
Schmitt, R. J., and Holbrook, S. J. (1999). Mortality of juvenile damselfish: implications for assessing processes that determine abundance. Ecology 80, 35–50. doi: 10.1890/0012-9658(1999)080[0035:MOJDIF]2.0.CO;2
Shoji, J., and Tominaga, O. (2018). “Relationships between submarine groundwater discharge and coastal fisheries as a water-food nexus,” in The Water-Energy-Food Nexus, eds A. Endo, and T. Oh (Singapore: Springer Nature), 117–131. doi: 10.1007/978-981-10-7383-0_9
Simpson, S. D., Radford, A. N., Nedelec, S. L., Ferrari, M. C. O., Chivers, D. P., McCormick, M. I., et al. (2016). Anthropogenic noise increases fish mortality by predation. Nat. Commun. 7, 1–7. doi: 10.1038/ncomms10544
Slomp, C. P., and Van Cappellen, P. (2004). Nutrient inputs to the coastal ocean through submarine groundwater discharge: controls and potential impact. J. Hydrol. 295, 64–86. doi: 10.1016/j.jhydrol.2004.02.018
Steele, M. A., and Forrester, G. E. (2005). Small-scale field experiments accurately scale up to predict density dependence in reef fish populations at large scales. PNAS 102, 13513–13516. doi: 10.1073/pnas.0504306102
Stevenson, D. K., and Campana, S. E. (1992). “Analysis of otolith microstructure data,” in Otolith Microstructure Examination and Analysis, eds D. Stevenson, and S. Campana (Ottawa: Canada Communication Group - Publishing), 126.
Suthers, I. M. (1998). Bigger? Fatter? Or is faster growth better? Considerations on condition in larval and juvenile coral-reef fish. Aust. J. Ecol. 23, 265–273. doi: 10.1111/j.1442-9993.1998.tb00730.x
Taniguchi, M., Burnett, W. C., Cable, J. E., and Turner, J. V. (2002). Investigation of submarine groundwater discharge. Hydrol. Process. 16, 2115–2129. doi: 10.1002/hyp.1145
Teh, L. C. L., and Sumaila, U. R. (2013). Contribution of marine fisheries to worldwide employment. Fish Fish. 14, 77–88. doi: 10.1111/j.1467-2979.2011.00450.x
Thorrold, S. R., Campana, S. E., Jones, C. M., and Swart, P. K. (1997). Factors determining δ13 C and δ18 O fractionation in aragonitic otoliths of marine fish. Geochim. Cosmochim. Acta 61, 2909–2919. doi: 10.1016/S0016-7037(97)00141-5
Thorrold, S. R., and Milicich, M. J. (1990). Comparison of larval duration and pre- and post-settlement growth in two species of damselfish, Chromis atripectoralis and Pomacentrus coelestis (Pisces: Pomacentridae), from the Great Barrier Reef. Mar. Biol. 105, 375–384. doi: 10.1007/BF01316308
Thresher, R. E., Colin, P. L., and Bell, L. J. (1989). Planktonic duration, distribution and population structure of Western and Central Pacific damselfishes (Pomacentridae). Copeia 2, 420–434.
Utsunomiya, T., Hata, M., Sugimoto, R., Honda, H., Kobayashi, S., Miyata, Y., et al. (2017). Higher species richness and abundance of fish and benthic invertebrates around submarine groundwater discharge in Obama Bay, Japan. J. Hydrol. Reg. Stud. 11, 139–146. doi: 10.1016/j.ejrh.2015.11.012
Vigliola, L., Harmelin-Vivien, M., and Meekan, M. G. (2000). Comparison of techniques of back-calculation of growth and settlement marks from the otoliths of three species of Diplodus from the Mediterranean Sea. Can. J. Fish. Aquat. Sci. 1299, 1291–1299. doi: 10.1139/f00-055
Vigliola, L., and Meekan, M. G. (2002). Size at hatching and planktonic growth determine post-settlement survivorship of a coral reef fish. Oecologia 131, 89–93. doi: 10.1007/s00442-001-0866-4
Villegas-Hernández, H., González-Salas, C., Aguilar-Perera, A., and López-Gómez, M. J. (2007). Settlement dynamics of the coral reef fish Stegastes partitus, inferred from otolith shape and microstructure analysis. Aquat. Biol. 1, 249–258. doi: 10.3354/ab00026
Waska, H., and Kim, G. (2011). Submarine groundwater discharge (SGD) as a main nutrient source for benthic and water-column primary production in a large intertidal environment of the Yellow Sea. J. Sea Res. 65, 103–113. doi: 10.1016/j.seares.2010.08.001
Wellington, G. M., and Victor, B. C. (1985). El Niño mass coral mortality: a test of resource limitation in a coral reef damselfish population. Oecologia 68, 15–19. doi: 10.1007/BF00379466
Wellington, G. M., and Victor, B. C. (1989). Planktonic larval duration of one hundred species of Pacific and Atlantic damselfishes (Pomacentridae). Mar. Biol. 101, 557–567. doi: 10.1007/BF00541659
Wellington, G. M., and Victor, B. C. (1992). Regional differences in duration of the planktonic larval stage of reef fishes in the eastern Pacific Ocean. Mar. Biol. 113, 491–498. doi: 10.1007/bf00349176
Welti, N., Gale, D., Hayes, M., Kumar, A., Gasparon, M., Gibbes, B., et al. (2015). Intertidal diatom communities reflect patchiness in groundwater discharge. Estuar. Coast. Shelf Sci. 163, 116–124. doi: 10.1016/j.ecss.2015.06.006
Wilson, D. T., and McCormick, M. I. (1997). Spatial and temporal validation of settlement marks in the otoliths of tropical reef fishes. Mar. Ecol. Prog. Ser. 153, 259–271. doi: 10.3354/meps153259
Keywords: coral reef, submarine groundwater discharge, damselfish, fitness, survival potential, feeding habits, otolith age reading, growth effect
Citation: Lilkendey J, Pisternick T, Neumann SI, Dumur Neelayya D, Bröhl S, Neehaul Y and Moosdorf N (2019) Fresh Submarine Groundwater Discharge Augments Growth in a Reef Fish. Front. Mar. Sci. 6:613. doi: 10.3389/fmars.2019.00613
Received: 20 July 2019; Accepted: 18 September 2019;
Published: 11 October 2019.
Edited by:
Esteban Avigliano, National Council for Scientific and Technical Research (CONICET), ArgentinaReviewed by:
Ana Laura Ibañez, Universidad Autónoma Metropolitana, MexicoMarcelo Soeth, Federal University of Paraná, Brazil
Copyright © 2019 Lilkendey, Pisternick, Neumann, Dumur Neelayya, Bröhl, Neehaul and Moosdorf. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Julian Lilkendey, anVsaWFuLmxpbGtlbmRleUBpY2xvdWQuY29t
†ORCID: Julian Lilkendey, orcid.org/0000-0003-3165-1079 Nils Moosdorf, orcid.org/0000-0003-2822-8261