 Peter J. Auster
Peter J. Auster Brooke C. Hodge3
Brooke C. Hodge3 Scott D. Kraus
Scott D. Kraus- 1Mystic Aquarium, Mystic, CT, United States
- 2Department of Marine Sciences, University of Connecticut, Groton, CT, United States
- 3Anderson Cabot Center for Ocean Life, New England Aquarium, Boston, MA, United States
The Northeast Canyons and Seamounts Marine National Monument (NECSMNM) was designated by President Barack Obama in 2016, using his authority under the Antiquities Act of 1906. The Act allows a President to proclaim as national monuments “historic landmarks, historic and prehistoric structures, and other objects of historic or scientific interest” that are “upon the lands owned or controlled” by the United States but to reserve each designation to “the smallest area compatible with the proper care and management of the objects to be protected.” Protection in general excludes commercial scale extraction and is in perpetuity. Here we present analyses of physiographic and ecological datasets that facilitated assessment of the conservation benefits of protections for a new monument. We also review and synthesize the ecological literature to describe processes that operate in continental margin and deep-sea settings, in order to demonstrate the monument area is bounded for proper management and is an object of scientific interest. Results indicate that the current monument designation is an area of high diversity and ecological connectivity across depths and along the continental margin. The monument boundaries contain hot spots (areas of high abundance and species richness) for seafloor communities (inclusive of benthic invertebrate and demersal fish) as well as marine mammals in the epipelagic. Many species are sensitive to disturbance and vulnerable to human activities (e.g., deep-sea corals and sponges) with very long recovery times and extremely low resilience. The monument contains at least nine exemplars of offshore northwest Atlantic marine wildlife communities and habitats (e.g., deep shelf invertebrates, shelf fish, deep sea corals and sponges in canyons and on seamounts, deep sea fish, chemosynthetic communities, deep sea soft sediment, shelf edge cetaceans, and seabirds). The region is relatively undisturbed and can serve as a reference site to focus future research on ecological processes in an increasingly industrialized ocean and one subject to the synergies of regional climate effects. These results suggest that there is great potential for discovery and novel research in this first Atlantic Ocean Marine National Monument.
Introduction
President Barack Obama designated the Northeast Canyons and Seamounts Marine National Monument (hereafter NECSMNM) by proclamation on 15 September, 2016 (Federal Register, 2016), using authorities delegated to the President of the United States by the Antiquities Act of 1906 (United States Congress, 1906). The Antiquities Act (hereafter “the Act”) was the first federal legislative tool for the protection of cultural and natural resources in the United States and preceded the National Park Service Act by 8 years. The Act allows a President to proclaim as national monuments “historic landmarks, historic and prehistoric structures, and other objects of historic or scientific interest” that are “upon the lands owned or controlled” by the United States but to reserve each designation to “the smallest area compatible with the proper care and management of the objects to be protected.”
Passage of the Antiquities Act was a Congressional response to public concerns related to theft and destruction of archeological sites and was ultimately designed to provide a rapid means to protect threatened federal lands and resources. Since it was signed into law by President Theodore Roosevelt, 17 of 20 Presidents have used the Act to create 158 monuments, including the Grand Canyon, Zion, Olympic, Statue of Liberty, and Stonewall (CRS, 2018).
For this monument as with earlier proclamations since President Roosevelt first used the authority in 1906, there have been conflicts between those that seek a utilitarian approach to use of public lands and waters and those that desire protections for some of the best examples of diverse natural resources across the United States (Costello, 2019).
The history of proclamations includes those that have identified particular types of objects (e.g., archeological objects) needing protection, while others have protected scenic resources (e.g., geologic features) and ecosystem or community types (e.g., coral reefs) of scientific and educational interest. The types of threats to resources within monuments are addressed within proclamations but an immediate threat of harm is not required by the Act to use this authority.
Here we address the scientific justification for designation of the NECSMNM in terms of the principal requirements of the Act. Our focus is on the rationale for “… the smallest area compatible …” and the designation protects “objects of … scientific interest.” The Proclamation states that the monument includes the waters and submerged lands in and around the deep-sea canyons Oceanographer, Lydonia, and Gilbert, and the seamounts Bear, Physalia, Retriever, and Mytilus. Further, it stipulates explicitly that the objects to be protected “… are the canyons and seamounts themselves, and the natural resources and ecosystems in and around them.” The rationale for the monument rests with the complex and precipitous geology and associated biological diversity of the region. Indeed, the proclamation emphasizes that “… the Atlantic Ocean meets the continental shelf in a region of great abundance and diversity as well as stark geological relief. The waters are home to many species of deep-sea corals, fish, whales and other marine mammals. Three submarine canyons and, beyond them, four undersea mountains lie in the waters approximately 130 miles southeast of Cape Cod. This area (the canyon and seamount area) includes unique ecological resources that have long been the subject of scientific interest.” For perspective, the canyons are deeper than the Grand Canyon (i.e., when measured from the head to the deepest channel feature the submarine canyons measure 2,272 m deep, versus 1,857 m depth of the Grand Canyon) and the seamounts taller than any mountains east of the Rocky Mountains (i.e., seamounts rise as high as 3,272 m above the surrounding seafloor while Harney Peak in South Dakota is 2,194 m). While the monument includes waters from the ocean surface to the seafloor, the shallowest seafloor depths begin just north of the canyon heads at 92 m and stretch to the abyss at 4,382 m. The shallowest seamount peak, on Bear Seamount, is at 1,110 m below sea level with the deepest peak on Mytilus Seamount at 2,389 m.
Here we summarize a series of analyses that quantified the “objects of scientific interest” and the natural resources of the region, which were used to facilitate discussions about monument boundaries and conservation benefits. In addition, we synthesize existing literature linking the scope of oceanographic drivers and species interactions to the spatial boundaries described in the proclamation. Our findings indicate that the final monument boundaries capture dominant processes and a high level of the diversity of species represented in the northeast region of the continental shelf edge and adjacent deep-sea ecosystems.
Developing Boundaries
Prior to designation, there was an initial strawman set of boundaries that were advanced by a coalition of environmental groups for a monument designation (Figure 1). This set of boundaries included a western unit inclusive of Oceanographer, Gilbert, and Lydonia submarine canyons as well as Bear, Physalia, Retriever, and Mytilus seamounts. An eastern unit included Nygren to Heezen submarine canyons. These proposed units covered an area of 16,211 km2. After a period of extended discussions with executive branch agencies, other regional congressional delegations, and the White House, the Connecticut congressional delegation led by U.S. Senator Richard Blumenthal, sent a formal request to the President to designate a continuous section of the continental margin from Oceanographer to Heezen canyons as a northern unit and separate seamounts unit to the south, proposing to protect an area of 16,006 km2 (Blumenthal et al., 2016). The final designation by President Obama, after considering feedback from the fishing industry (Mitchell, 2016), reduced the size of the proposal but included Oceanographer, Gilbert, and Lydonia canyons as a northern unit and the four seamounts as a separate southern unit, creating a monument of 12,696 km2. The designation protects an area approximately the size of the state of Connecticut and represents just 1.5% of the Atlantic region of the U.S. EEZ and 0.11% of the entirety of the U.S. EEZ.
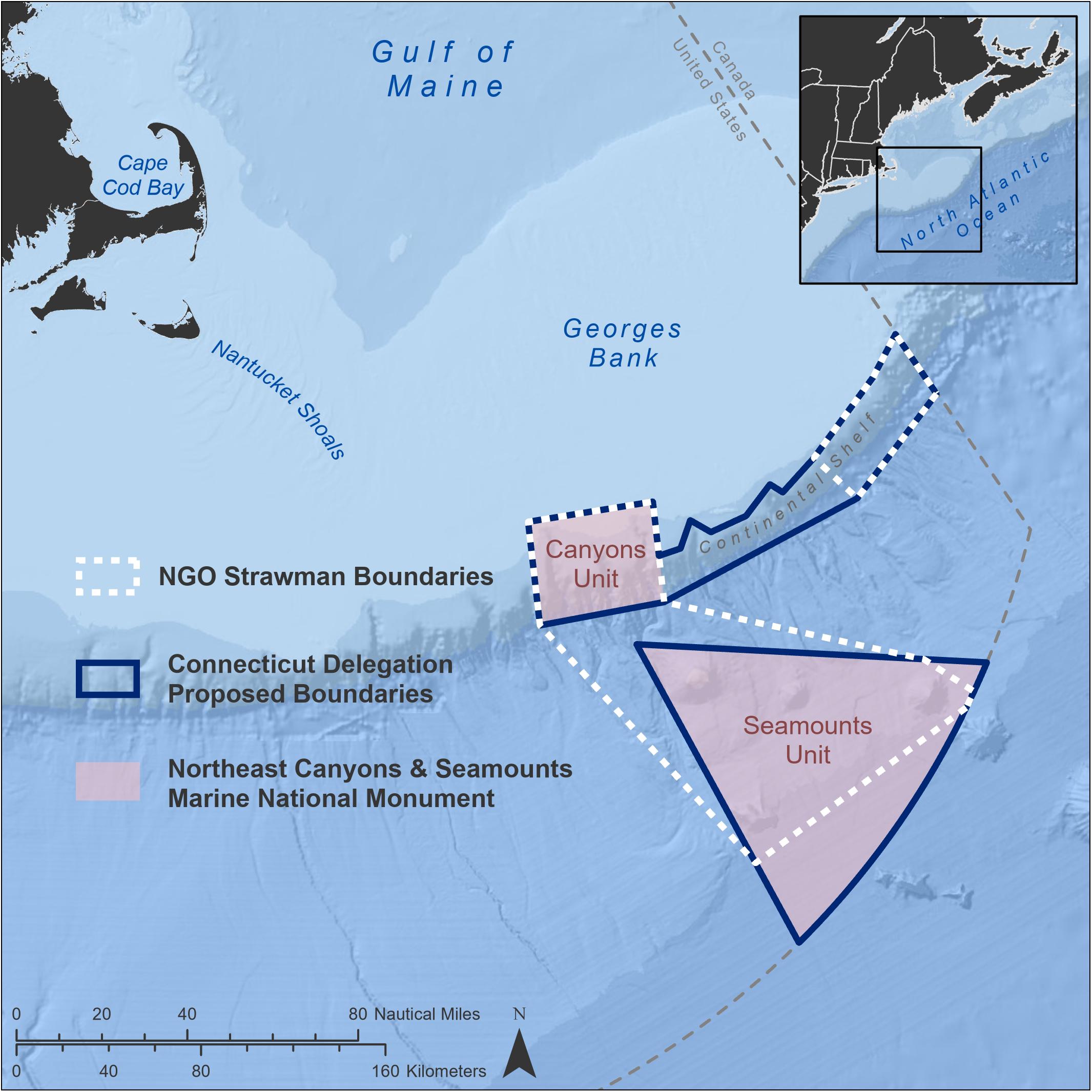
Figure 1. Changes in boundaries from conception through final proclamation of the Marine National Monument. Areas delineated above include the initial boundaries proposed by a coalition of public aquaria and environmental groups (14,834 km2), the request directly to President Obama by the Connecticut Congressional delegation (16,006 km2), and the final area designated by President Obama under the authority of the Antiquities Act (12,696 km2). The area in the final designation was significantly reduced in size from original requests as a response to fishing industry concerns and those of other stakeholders.
Object of Scientific Interest
The seamounts and canyons, due to their geographic complexity and location, influence oceanographic conditions at multiple space and time scales. The submarine topography (steep slopes, deep canyons, and tall undersea mountains; Figure 2A) influences the currents, upwellings, stratification, and mixing (Shank, 2010) that make the species and ecosystems within the monument so diverse, abundant, and unique. Indeed, the seamounts affect the Gulf Stream and deeper Atlantic western boundary currents (e.g., Ezer, 1994), producing complex currents around seamounts at the seafloor, along the continental margin, and throughout the overlying water column (Auer, 1987; Waring et al., 2001; White et al., 2007). This interaction of currents and topography influence both temperature and nutrient transport, upon which the community structure of these deep-ocean ecosystems depend (Griffin, 1999; Waring et al., 2001).
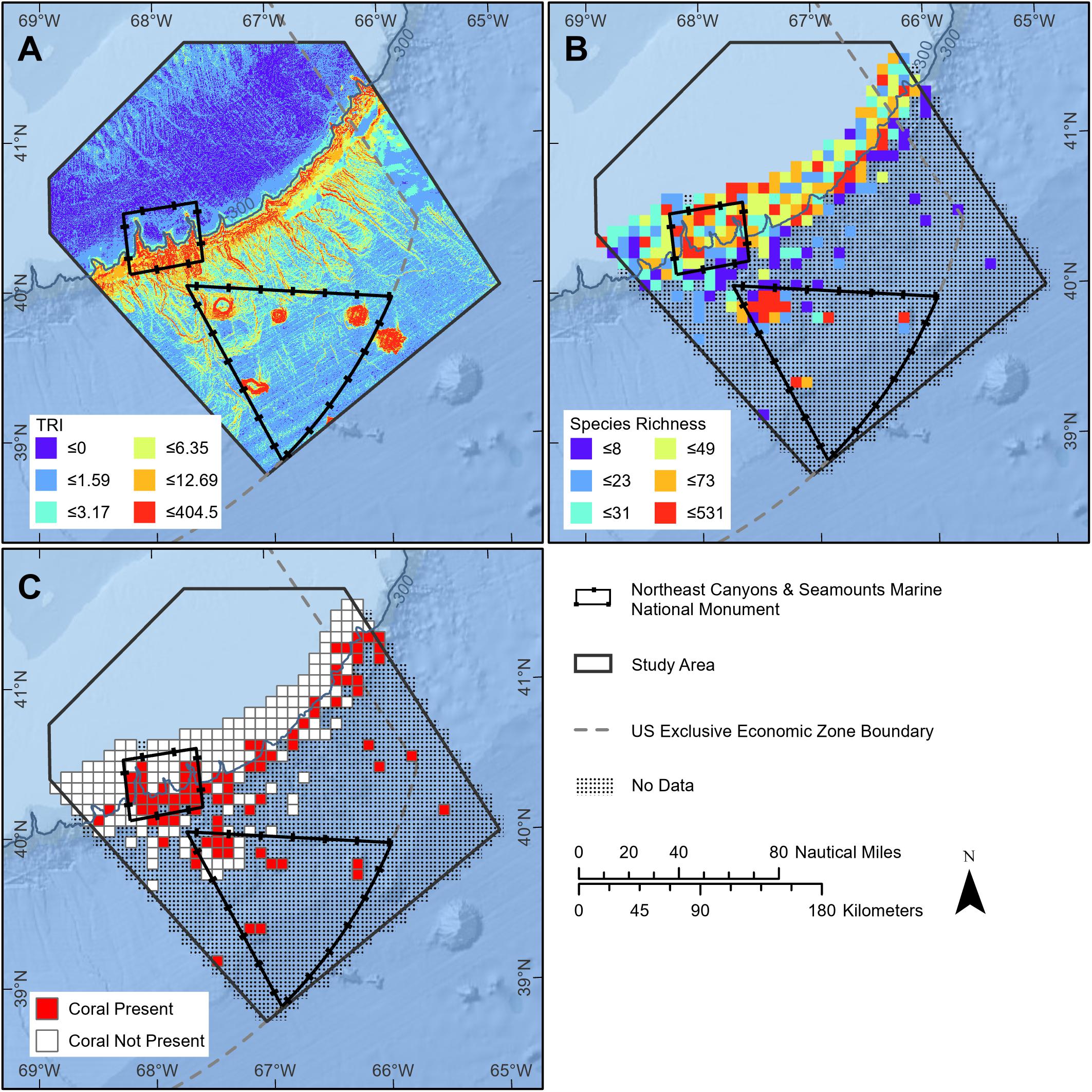
Figure 2. Attributes of the seafloor and associated species in the monument region (slides are based on Kraus et al., 2016 with designated boundary configuration). (A) Topographic roughness index based on bathymetry shows the variability in medium-scale habitat complexity with warmer colors (reds) indicating steepest terrain. (B) Species richness of seafloor fauna (invertebrates and demersal fishes) aggregated from multiple types of sampling programs. Note hotspots occur in canyons and on seamounts. (C) Grid cells with documented presence of deep-sea corals. Supplementary Appendix 1 summarizes details of the geospatial analyses presented here.
Marine species are differentially distributed across depths and substrates, similar to the various plant and animal zones observed across different altitude zones of tall mountains on land. Because the monument encompasses a diversity of topography, depths, and substrates, it protects multiple communities of organisms and their component species in a small area. For seafloor species alone, four major community types have been identified along the continental slope and rise binned by depth interval (i.e., upper slope 200–700 m, upper middle slope 700–1,200 m, transitional 1,200–1,300 m, and lower slope 1,300–>2,400 m; Hecker et al., 1980; Hecker, 1990b). At least four other ecological communities have been classified down the slopes of the seamounts grading into the abyss (i.e., <1,300 m, 1,300–2,300 m, 2,300–2,600 m, and >2,600 m; Cho, 2008). Further, the canyons and the seamounts sections of the Monument harbor significantly different marine communities, even when taking depth variation into account (Kilgour et al., 2016). In addition, there are both cold-seep chemosynthetic and deep-sea Xenophyophore communities within the boundaries of the monument. The small spatial-scale variability of substrate types and topographies contributes greatly to these overall patterns of diversity (Ryan et al., 1978; Valentine, 1987; Auster et al., 2005). Thus, within a relatively small area, the combined canyon and seamount units in the Monument capture a wide diversity of species and biological community types (Table 1). This unique combination of marine ecosystem elements, as well as the abundance and biological diversity of life at the seafloor, in the water column, and at the surface, all meet the definition of “objects of scientific interest”, as we will address below.
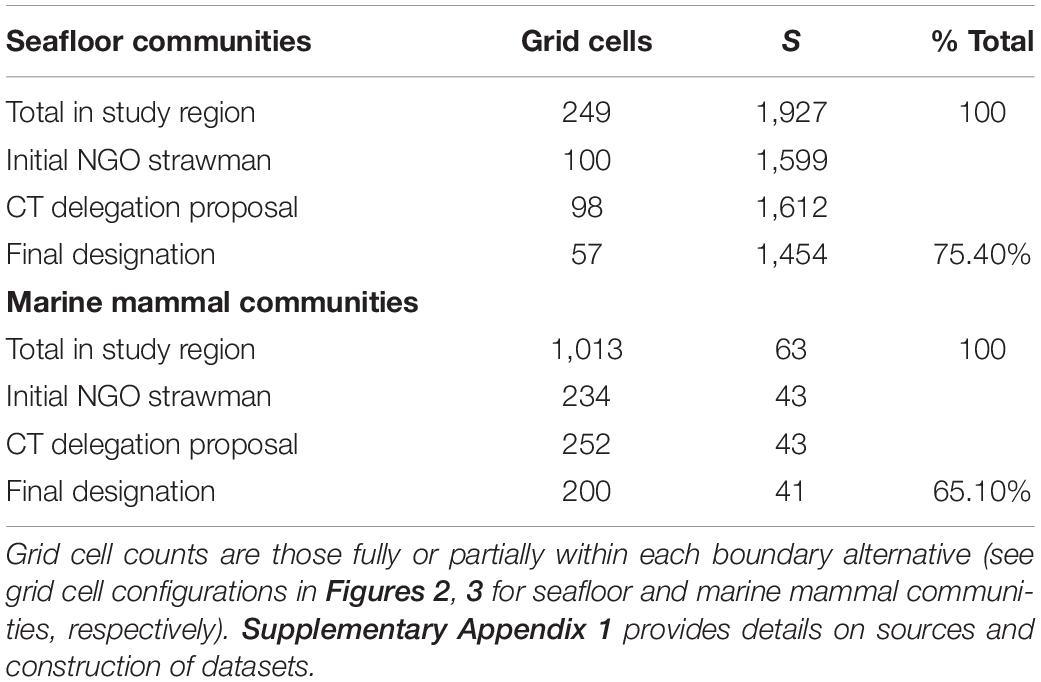
Table 1. Comparison of species richness (S) represented by each boundary alternative as illustrated in Figure 1.
As a result of the complex interactions between topography and oceanography, the canyons and seamounts within the Monument are biodiversity hotspots (Kelly et al., 2010; Kilgour et al., 2016) and protect outstanding examples of our marine biological heritage that are scientific objects of national significance. Hotspots have been identified for seafloor diversity (including deep sea corals, sponges, deep sea fish, and cold-seep chemosynthetic species; Kraus et al., 2016; Figures 2B,C). Many species function as ecosystem engineers within canyon and seamount communities [e.g., American lobster Homarus americanus and tilefish Lopholatilus chamaeleonticeps burrow into canyon walls and produce small-scale habitat complexity; deep-sea corals support species-specific commensals; both types of interactions (i.e., species and habitat) enhance local biological diversity; Cooper et al., 1987a; Auster et al., 2005; Watling et al., 2011]. Whales, dolphins, seabirds, sea turtles, and pelagic fishes (tunas, billfish, and sharks) exhibit high abundance and diversity along the continental shelf edge (CeTAP, 1982; Powers, 1984; Hain et al., 1985; Kenney and Winn, 1986, 1987; Palka, 2012; Kraus et al., 2016). Further, analyses of whale and dolphin sighting data demonstrate both abundance and diversity hot-spots of whales and dolphins in the monument area (Figures 3A–D).
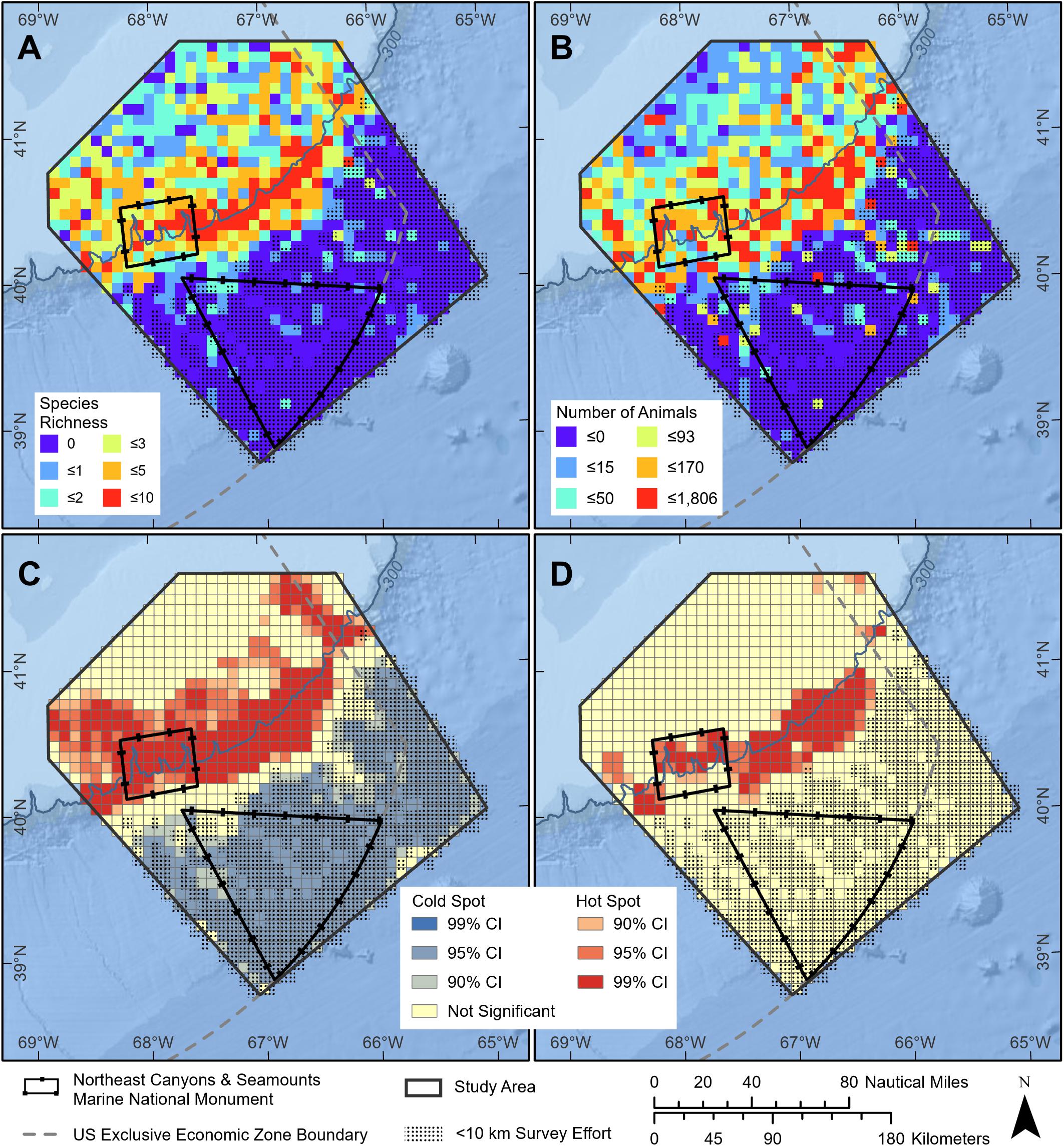
Figure 3. Patterns in the distribution of marine mammal species in the monument region (slides are based on Kraus et al., 2016 with designated boundary configuration). (A,B) Marine mammal distribution showing species richness and total number of individuals for all marine mammals as a heat map. (C,D) Hot spot statistical analysis of species richness and numbers of marine mammals, with hot spots based on significantly higher values than surrounding areas. Note hot spots over canyon heads. Low values over seamounts is due in part to low survey effort. Supplementary Appendix 1 summarizes details of the geospatial analyses presented here.
The outer shelf around the canyon heads is a vital initial link to the ecological processes that occur within canyons and contributes to the patterns of diversity and productivity. For example, dense near-bottom swarms of krill have been observed during daytime in canyon heads at 300–400 m that rise in the water column toward the surface at night (Greene et al., 1988). These dense swarms exceeded 1,000 animals per cubic meter in a layer up to 50 m above the seafloor and occur due to a combination of “topographic blockage” at the shallowest heads of the canyons and surrounding deep shelf (ca. 200 m) during downward migration at dawn, and funneling produced by the canyon head morphology (Greene et al., 1988; Hobson et al., 1989). Such swarms can function as trophic subsidies to a diversity of large zooplanktivores because krill are energy rich, large in size, and at high densities are easy to prey on. Dense aggregations of fish and squid that feed on krill and small fishes have been observed in these areas, which explains the high density and diversity of marine mammals in canyon environments (Moors-Murphy, 2014).
The occurrence of marine mammals in the monument area is remarkably high in terms of both abundance and diversity (CeTAP, 1982; Kenney and Winn, 1986, 1987; Payne and Heinemann, 1993; Palka, 2012; Kraus et al., 2016) hosting at least 10 dolphin species, seven large whale species, and six medium whale species. In this latter group, the monument is home to the extreme deep diving species on the planet, the beaked whales (Waring et al., 2001). At least three species of beaked whale reside in the monument region, all capable of diving to nearly 1,900 m based on tagging studies elsewhere, and staying submerged for over an hour. Evidence of beaked whale predation, based on characteristic gouge marks in the seafloor from whale beaks manipulating prey, was found in Gilbert and Lydonia Canyons with a maximum depth of 2,745 m, nearly 900 m deeper than any beaked whale previously recorded (Auster and Watling, 2010, but see Schorr et al., 2014). While there is undoubtedly migration along the shelf edge by marine mammals, whenever systematic surveys have been done in the Monument, abundance has been high (CeTAP, 1982; Palka, 2012). A report posted on 12 August, 2016 from an Atlantic Marine Assessment Program for Protected Species (AMAPPS) cruise aboard the NOAA Ship Henry Bigelow states “… the scientific party encountered the most animals ever in a single day” in Oceanographer and Lydonia canyons. Sightings included sperm whales in deep waters near Oceanographer Canyon and in the canyon head region between Oceanographer and Lydonia Canyons (100–200 m) “2,500 common dolphins, 120 fin whales, 50 humpback whales, 60 Risso’s dolphins, 70 pilot whales, 80 bottlenose dolphins, 100 striped dolphins, with a few beaked whales and ocean sunfish when the ship was in the deeper waters” (NEFSC, 2016).
Many acoustic and sightings surveys have consistently verified this high level of abundance and diversity. It appears that the monument area has an unusually high abundance of beaked whales, deep divers that appear to be taking advantage of the deep water canyons and oceanography to forage (Auster and Watling, 2010; Cholewiak et al., 2013; Wenzel et al., 2013; Stanistreet et al., 2017; DeAngelis et al., 2018). The monument also continues to be year-round habitat for sperm whales (Stanistreet et al., 2018) as well as a variety of other species of dolphins and whales. Indeed, a recent New England Aquarium aerial survey on 28 June, 2019 observed 1,252 marine mammals, including 1,036 common dolphins (Delphinus delphis), 192 Risso’s dolphins (Grampus griseus), and four Cuvier’s beaked whales (Ziphius cavirostris). An earlier survey on 6 September, 2018 observed 335 common dolphins, 234 Risso’s dolphins, 30 bottlenose dolphins, two Cuvier’s beaked whales, three ocean sunfish (Mola mola), and a rare giant manta ray (Manta birostris). On 23 April, 2018, the day’s sightings included 169 bottlenose dolphins (Tursiops truncatus), 44 Risso’s dolphins, 13 Sowerby’s beaked whales (Mesoplodon bidens), 57 pilot whales (Globicephala spp.), four sperm whales (Physeter macrocephalus), and 44 other dolphins whose species could not be determined (see text footnote 1)1. These sightings and publications demonstrate that over at least 40 years, the monument area has been home to consistently high levels of marine mammal abundance and diversity, apparently due to the dynamic and productive oceanographic and bathymetric features that enhance trophic interactions in a relatively small area (Griffin, 1997).
The shallowest areas in the Monument (the western upper canyon regions and the epi-pelagic zones over the seamounts) are critical to protecting the ecosystem linkages that both transport nutrients to the surface through predator–prey interactions, and organic carbon to deep sea ecosystems (corals and benthic communities) through plankton and fecal detritus, downwelling materials, down-slope currents, and animal migration and mortality (e.g., Youngbluth et al., 1989; Hecker, 1990a; van Oevelen et al., 2009; Soetaert et al., 2016). These processes influencing carbon flux and sequestration in the ocean are enhanced in canyon and seamount landscapes, by linking shallow seafloor and pelagic areas to the deep sea. Carbon sequestration through this oceanographic “pump” is of global importance, and the implications for the monument are that the extraordinary diversity of species, including corals at depth, produce a wide array of ecological benefits in these undisturbed communities.
Mid-water (mesopelagic) fish are the most abundant group of vertebrate animals on our planet (Irigoien et al., 2014) and also play a significant role in the transport of carbon from surface waters into the deep sea (Davison et al., 2013; St. John et al., 2016). These highly abundant animals undertake large daily vertical migrations through the water column where gut carbonates, released through digestion as particulates at the surface, interact with dissolved CO2 that subsequently rapidly sinks into the deep ocean. Protected areas like the monument contribute to the health of such fish populations and enhance resilience to the stresses from acidification and temperature due to climate change. Roberts et al. (2017) hypothesized that “mesopelagic fish may drive an upward alkalinity pump that is currently acting to counter surface ocean acidification.” Currently there is increased interest in exploiting deep sea mesopelagic fish species, and the Monument will provide significant ecological benefits not generally considered when setting reference targets for sustainable fishing.
At the upper trophic levels, the high density and diversity of shelf-edge marine mammals that feed on deep sea squid and fish, transfer deep-sea productivity to the surface through defecation, and ultimately back to the seafloor through fecal-detritus as well as via the deposition of dead animals (Smith and Baco, 2003; Roman and McCarthy, 2010; Schmitz et al., 2014). Because the Monument is an area of consistently high numbers of marine mammals (Waring, 1998; Kraus et al., 2016; NEFSC, 2016), this nutrient cycling may be critical for local ecosystem functioning (Lavery et al., 2014; Roman et al., 2014; Doughty et al., 2016). Further, the oceanographic and biological characteristics of the Monument create essential feeding and navigation waystations for marine mammals, seabirds, sea turtles, tunas, sharks, and swordfish (Holland and Grubbs, 2007; Kaschner, 2007; Litvinov, 2007; Santos et al., 2007; Thompson, 2007; Wenzel et al., 2013).
Past studies and current interest demonstrate that the monument has value for future scientific inquiry. The monument has had limited historic and contemporary disturbance from oceanic industries, is the only Atlantic Marine Monument, and encompasses a complete section of “shelf edge to deep sea” marine habitat off the continental United States. This fully protected area will serve an important role to understanding the effects of climate change (e.g., Roberts et al., 2017), by providing an undisturbed reference region in contrast to areas subject to commercial scale fisheries and other human activities. Predicted benefits include the conservation of genetic diversity, the enhanced resilience of fish, mammal, and invertebrate populations impacted by fishing and acidification, the protection of apex predators, the enhancement of commercially valuable fisheries through spillover (to recolonize habitats and communities affected by fisheries; Auster and Shackell, 2000; Sackett et al., 2017), and the sequestration of carbon. The Monument will be a critical research site with limited human impacts, a problem associated with studies in other deep-sea areas (NEFMC, 2017). Since the waters of New England and the Canadian Maritimes are warming faster than any other region of the Atlantic Ocean (Mills et al., 2013), this relatively pristine system will serve as a laboratory to assess the impacts of climate change alone and in comparison to areas with direct human impacts that lack protection (e.g., from fishing and the potential future impacts from mining methane hydrates and manganese crusts; ISA, 2008; Hand, 2014; WOR, 2014).
The Canyons and Seamounts Monument includes species and habitat types not found in any other Sanctuary, National Park or monument. Recent research conducted within the monument region shows it has an extraordinarily high potential for new scientific discoveries. Recent discoveries have included new species, genetic variability and hidden genetic structure within species, and range extensions of species known from elsewhere. Given the limited exploration work in this area, such discoveries are likely to continue, greatly increasing our understanding of marine deep sea biology (e.g., Mills, 2003; Moore et al., 2003; Cairns, 2006; France, 2007; Packer et al., 2007; Thoma et al., 2009; Cho and Shank, 2010; Quattrini et al., 2015; Coykendall et al., 2016), with positive implications for medical, aquaculture, and marine technology industries. Finally, the monument includes exemplars (Figure 4) of a diversity of Atlantic marine ecological communities and habitats, some of which are known to be unique (e.g., deep sea coral-sponge, canyon-head, cold seep, and xenophyophore communities) but are only poorly studied (see NOAA, 2013; Skarke et al., 2014; Quattrini et al., 2015).

Figure 4. Images of some exemplar communities and habitats. (From top, left to right) (A) Glacier rafted boulders at the head of Oceanographer Canyon. Golden tilefish using crevice for shelter. (B) American lobster burrowed into clay wall at the head of Gilbert Canyon. (C) Octocorals, including a recently described species Thouarella (Euthouarella) grasshoffi (Cairns, 2006) in Oceanographer Canyon. (D) Dense krill and amphipods over seafloor with offshore hake Merluccius albidus preying upon the aggregation in Lydonia Canyon. (E) Dense forest of Paramuricea placomous along outcrop in Oceanographer Canyon. (F) Cold seep with bacterial mat and chemosynthetic mussels (from Nygren Canyon and representative of seeps located with multibeam in Oceanographer and Lydonia Canyons by Skarke et al., 2014). (G) Jasonisis sp. on the flank of Mytilus Seamount. (H) A diverse coral assemblage on a manganese encrusted outcrop on Retriever Seamount, including stony corals, octocorals, precious corals, black corals, and bamboo corals. (I) Dense sponges (Euplectellidae) on Retriever Seamount. [Images (A,B,D) from UConn; (C) DeepEast Expedition NOAA/WHOI/UConn/UMaine; (E–I) Multiple NOAA Ocean Exploration Program expeditions to canyons and seamounts region].
Research in this region historically has involved many national and international investigators and has included studies on a broad range of ecological processes and taxonomic groups, from marine microbes to the great whales. The potential for ongoing research partnerships and collaborations is high, and such activities – particularly studies that collect long-term time series data – were given renewed momentum by designation of the Monument. For example, time series studies of canyon head communities by Cooper et al. (1987b) in the early 1980s resulted in a 5-year data set from submersible-based photo transects that can serve as a baseline to measure change over a 30 year time frame to the present. Previous transect data collected by DSV Alvin and the ROV Deep Discoverer in the canyons and seamounts of the Monument will also serve as a foundation for time series studies. Kilgour et al. (2016) demonstrated that benthic marine ecosystems below fishing depths in Oceanographer Canyon were remarkably stable using image data from dive transects from five different years between 1978 and 2013 (Figure 5). Such long-term studies are critical to understanding the nature and trends in oceanic ecosystem changes, with implications for both the conservation of biodiversity and the health of economically valuable offshore fisheries in the Atlantic.

Figure 5. Exemplars of imagery from submersible dives in the same area of Oceanographer Canyon at ca. 1,100 m depth in 1978, 1980, 2001, 2005, and 2013 facilitated an examination of community structure and found high similarity in community composition over time (Kilgour et al., 2016), demonstrating a high degree of stability of undisturbed habitats. (Images from 1978 to 2001 from DSV Alvin dives; 2013 from NOAA Okeanos Program dive).
Studies in and around the marine monument area have been using internationally known collections as reference materials and as repositories for new biological specimens. Primary institutions include Smithsonian Institution, Yale Peabody Museum, and Harvard Museum of Comparative Zoology. These institutions have been collecting specimens from Ocean Exploration and Census of Marine Life trawl-dredge and submersible expeditions since 2001, and have active curators and associated scientists with scholarly interests in this region.
Monument Size and Boundary Considerations
The scientific case that the existing boundaries for the NECSMNM encompass the smallest area for proper care and management of the objects of scientific interest is strong. The patterns and processes that produce and influence the Monument’s biological productivity, diversity, and ecosystems, indicate that the pelagic and abyssal elements of the Monument include a unique combination of ocean systems, all critically important and wholly appropriate to the governing mandate of the Monuments Act within a remarkable small area.
Exploiting fish populations in ever deeper waters over the course of time has been an enduring pattern of commercial fisheries (Morato et al., 2006; Watson et al., 2015) and is in part mediated by factors such as fuel costs, available biomass, value of landed catch, as well as subsidies provided by governments and private enterprises (Norse et al., 2012). While the direct effects of disturbance by fishing are well known in terms of type and direction of impacts (Auster and Langton, 1999; Koslow et al., 2016), our understanding of how such effects cascade through communities and ecosystems is only currently emerging. Bailey et al. (2009) demonstrated that the effects of exploitation of deep-sea fish populations extends beyond the depth of directed fishing, as species with populations only partially within the depths of those fisheries responded and declined despite occurring in a depth refugia. This pattern included species targeted by fisheries as well as populations of by-catch species. Other impacts from fishing are long lasting, on the scale of decades to millennia for the longest-lived habitat-forming species such as corals, with low overall ecological resilience (Koslow et al., 2001; Clark and Koslow, 2007; Waller et al., 2007). Based on the long-lived, slow reproduction, life histories of many deep-sea fish species, and empirical observations of exploited populations, most deep-sea taxa are easily overexploited, have very low ecological resilience, and could rapidly reach threatened or endangered status (Devine et al., 2006; Baker et al., 2009). In the current case, the monument as bounded protects a slice of the continental margin and seamounts region from such vertically cascading impacts and serves as a hedge against unintended impacts from fishing activities along the remainder of the continental margin.
The commercial fishing industry has voiced concerns about economic losses from exclusion of seafloor fisheries prosecuted around and in the heads of the submarine canyons, and with pelagic fisheries in surface waters throughout. Arguments have focused on the value of catch lost within monument boundaries, the authority of the President to use the Antiquities Act to designate monuments in the U.S. Exclusive Economic Zone, and the primacy of federal fisheries laws to manage fisheries throughout the region (Eilperin, 2016; White, 2016). Analyses of reported landings in the regional squid-mackerel-butterfish fishery, the principal seafloor fishery that included monument waters, demonstrate there has been an increase in catch since designation while the catch of highly migratory species has remained stable (i.e., comparing 3 years pre- to three post-designation; Eilperin, 2018; NRDC, 2019). Noteworthy is that allowable catch levels are not affected by the monument designation, just that fishing for these species must take place elsewhere. Challenging the authority of the President to designate monuments in the EEZ and the primacy of federal fisheries law (i.e., Magnuson Act and amendments) for fisheries have played out in Federal courts (Fedderly, 2018). Decisions upheld Presidential authority to designate monuments in waters controlled by the United States and that federal laws do not impart primacy of one over another, and can be applied to meet disparate societal goals (Ryan, 2019).
If the monument boundaries were to be moved deeper, away from canyon heads into deeper regions of the pelagic zone, the effects of fishing on target and by-catch species will cascade through deeper portions of the monument. This is due to impacts at the level of populations that extend beyond fishing depths (Bailey et al., 2009), as well as through changes in predator–prey interactions, and shifts in rates of carbon transport from surface and mid-water regions to the deep sea. A monument boundary adjustment to deeper waters would also eliminate the principal way that the general public would experience the monument. The ability to visit the monument to observe the high diversity of marine mammals and seabirds in high density, in the absence of commercial activities would be eliminated. Recreational fisherman would also suffer the same degradation of experience. Indeed, the same important policy goal was applied in Glacier Bay National Park when commercial fishing activities were phased out to maximize the wilderness experience of visitors (Sloan, 2002; Sen, 2010). (Note the monument proclamation contains a seven-year sunset provision solely for offshore lobster and red crab trap fisheries because of the perceived difficulties of shifting fixed fishing gear to new grounds).
The boundaries of the Monument will encompass the only entanglement-free and by catch-free zone off the east coast of the United States after the sunset period for trap fisheries expires in 2023. All bottom contact gear threatens long-lived species such as deep-sea corals and other fragile structure-forming species. Fixed gear (traps, gillnets, and both bottom and pelagic long-line gear) with buoy and submerged lines (and associated traps, mesh, or hooks) are all identified as significant mortality risk to all marine mammals (Reeves et al., 2013; Lewison et al., 2014; Werner et al., 2015), sea turtles (Lewison et al., 2004; Finkbeiner et al., 2011), and seabirds (Winter et al., 2011; Hatch, 2017). Mobile gear including deep and mid-water trawls are known to catch and kill numerous species of dolphins (Rossman, 2010). Baited hooks also directly capture non-target species, including pilot whales and beaked whales, sea turtles, seabirds, and sharks (Northridge, 1996; Moore et al., 2009). Both short- and long-finned pilot whales are a primary component of marine mammal by catch in the pelagic long-line fishery in the monument region (Garrison and Stokes, 2014). The Western North Atlantic stock of long-finned pilot whales has a potential biological removal of only 35 individuals and therefore is extremely vulnerable to even low levels of mortality (Garrison and Stokes, 2014). Incidental catch also threatens leatherback and loggerhead sea turtle populations in the Monument, both of which are ESA listed species, and have been observed in large numbers among the canyons in the Monument (Northeast Ocean Data Working Group. Data Explorer2). The exclusion of commercial scale fishing in the monument therefore contributes to minimizing exposure of these vulnerable species to fishing gear in an area where intense feeding and species interactions are occurring.
The current boundaries also define an area that is protected from any activities related to the future extraction of oil, gas, methane hydrates and manganese crusts, resources under scrutiny for exploitation along the entire east coast and on the high seas (WOR, 2014; Snow, 2017). The impacts from such activities will be extreme and the monument represents a reference site to compare with impacted areas.
The Proclamation (No. 9496, 81 Fed. Reg 65,163) plainly states, “[t]hese canyons and seamounts, and the ecosystems they compose, have long been of intense scientific interest… [T]he waters and submerged lands in and around the deep-sea canyons. and the seamounts. contain objects of scientific and historic interest. These objects are the canyons and seamounts themselves, and the natural resources and ecosystems in and around them.” The final boundaries delineated in the proclamation of the monument considered the effects of designation on stakeholder communities, given the reduced size of the area compared to the original strawman advanced by public aquaria and environmental organizations, and the actual proposal submitted by the CT Congressional delegation to the President.
Conclusion
In conclusion, the canyons, seamounts and the natural resources and ecosystems within and in the surrounding waters are objects of scientific interest that are conserved with the existing boundaries of the NECSMNM. The Monument’s boundaries capture the local scale processes that sustain the natural resources, ecosystems, and patterns of biological diversity we observe. The monument encompasses an incredibly diverse range of overlapping and interacting species, communities, and habitats all of which are vulnerable and sensitive to human disturbance. By eliminating all commercial extractive activities as described in the 2016 Proclamation, the unique biological and scientific features of this wild ocean setting, not only provides an area to study undisturbed oceanic wildlife but is a gift to the American public, indeed people around the world, as a place to marvel at a wonder of our natural marine heritage.
UPDATE: A proclamation signed by President Trump on 5 June, 2020 revises the earlier designation and eliminates protections from commercial fishing in the NECSMNM (Holden, 2020). Legal challenges by environmental groups were filed on 17 June, 2020 in Federal court to challenge use of the Antiquities Act to reverse protections enacted by the original proclamation (Frazin, 2020).
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation, to any qualified researcher.
Ethics Statement
Ethical review and approval was not required for the animal study because this study used archived data throughout. All fieldwork reported here is exempt from IACUC protocols as projects were observational in nature. The marine mammal surveys were all conducted under multiple marine mammal permits from NOAA to multiple investigators contributing to the data base.
Author Contributions
PA and SK contributed to project design and identification of datasets. BH and MM organized data and conducted geospatial analyses as well as prepared manuscript figures. PA, BH, MM, and SK contributed to writing and reviewing the manuscript. All authors contributed to the article and approved the submitted version.
Funding
Funding for retrospective analysis and ongoing surveys was provided by the Natural Resources Defense Council, Conservation Law Foundation, Waitt Foundation, and the National Ocean Protection Coalition.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This article is based on a public comment letter sent to Secretary of the Interior Ryan Zinke from PA and SK regarding a review of Marine National Monuments pursuant to Executive Orders 13792 and 13795 from President Donald Trump. We thank Jon Witman and Ester Quintana for helpful discussions and comments. The comments of two reviewers further improved the article. This article represents the views of the authors and not our respective institutions or funders.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2020.00566/full#supplementary-material
Footnotes
- ^ https://www.andersoncabotcenterforoceanlife.org/blog/scientists-see-over-1200-animals-in-marine-monument-in-just-over-one-hour/
- ^ Available at: http://www.northeastoceandata.org/data-explorer/
References
Auer, S. J. (1987). Five-year climatological survey of the Gulf Stream System and its associated rings. J. Geophys. Res. 92, 11709–11726.
Auster, P. J., and Langton, R. W. (1999). The effects of fishing on fish habitat. Am. Fish. Soc. Symp. 22, 150–187.
Auster, P. J., Moore, J., Heinonen, K., and Watling, L. (2005). “A habitat classification scheme for seamount landscapes: assessing the functional role of deepwater corals as fish habitat,” in Cold-water Corals and Ecosystems, eds A. Freiwald and J. M. Roberts (Berlin: Springer-Verlag), 761–769. doi: 10.1007/3-540-27673-4_40
Auster, P. J., and Shackell, N. L. (1997). “Fishery reserves,” in Northwest Atlantic Groundfish: Perspectives on a Fishery Collapse, eds J. G. Boreman, B. S. Nakashima, H. W. Powles, J. A. Wilson, and R. L. Kendall (Bethesda, MA: American Fisheries Society), 159–166.
Auster, P. J., and Shackell, N. L. (2000). Marine protected areas for the temperate and boreal Northwest Atlantic: the potential for sustainable fisheries and conservation of biodiversity. Northeastern Nat. 7, 419–434.
Auster, P. J., and Watling, L. (2010). Beaked whale foraging areas inferred by gouges in the seafloor. Mar. Mamm. Sci. 26, 226–233. doi: 10.1111/j.1748-7692.2009.00325.x
Bailey, D. M., Collins, M. A., Gordon, J. D. M., Zuur, A. F., and Priede, I. G. (2009). Long-term changes in deep-water fish populations in the North East Atlantic: a deeper-reaching effect of fisheries? Proc. R. Soc. B 275, 1965–1969. doi: 10.1098/rspb.2009.0098
Baker, K. D., Devine, J. A., and Haedrich, R. L. (2009). Deep-sea fishes in Canada’s Atlantic: population declines and predicted recovery times. Environ. Biol. Fish. 85, 79–88. doi: 10.1007/s10641-009-9465-8
Blumenthal, R., Murphy, C. S., DeLauro, R. L., Larson, J. B., Courtney, J., Himes, J., et al. (2016). Letter to the President of the United States Barack Obama regarding designation of a national monument. Available online at: https://www.blumenthal.senate.gov/newsroom/press/release/connecticut-delegation-urges-president-obama-to-designate-new-england-coral-canyons-and-seamounts-as-first-ever-atlantic-marine-national-monument (accessed 2 August 2016).
Brown, S. K. R., Zwanenburg, K., and Branton, R. (2005). ECNASAP - East Coast North America Strategic Assessment. Version 1 In OBIS Canada Digital Collections. Dartmouth, NS: Bedford Institute of Oceanography.
Cairns, S. D. (2006). Studies on western Atlantic Octocorallia (Coelenterata: Anthozoa). Part 6: the genera Primnoella Gray, 1858; Thouarella Gray, 1870; Dasystenella Versluys, 1906. Proc. Biol. Soc. Washington 119, 161–194. doi: 10.2988/0006-324x(2006)119[161:sowaoc]2.0.co;2
CeTAP (1982). A Characterization of Marine Mammals and Turtles in the Mid- and North-Atlantic Areas of the U.S. Outer Continental Shelf. Final Report No. AA551-CT8-48 of Cetacean and Turtle Assessment Program (CETAP). Washington, D.C: U.S. Department of the Interior.
Chen, J., Yang, S., Li, H. W., Zhang, B., and Lv, J. (2013). Research on geographical environment unit division based on the method of natural breaks (Jenks). Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. XL-4/W3, 47–50. doi: 10.5194/isprsarchives-xl-4-w3-47-2013
Cho, W., and Shank, T. M. (2010). Incongruent patterns of genetic connectivity among four ophiuroid species with differing coral host specificity on North Atlantic seamounts. Mar. Ecol. 31, (Suppl. 1), 121–143. doi: 10.1111/j.1439-0485.2010.00395.x
Cho, W. W. (2008). Faunal Biogeography Community Structure and Genetic Connectivity of North Atlantic Seamounts. Ph.D. Dissertation, Woods Hole Oceanographic Institution, Woods Hole, MA.
Cholewiak, D. M., Baumann-Pickering, S., and Van Parijs, S. (2013). Description of sounds associated with Sowerby’s beaked whales (Mesoplodon bidens) in the western North Atlantic Ocean. J. Acoust. Soc. Am. 134, 3905–3912. doi: 10.1121/1.4823843
Clark, M. R., and Koslow, J. A. (2007). “Impacts of fisheries on seamounts,” in Seamounts: Ecology, Conservation and Fisheries, eds T. J. Pitcher, T. Morato, and P. J. B. Hart (Oxford: Blackwell), 413–441. doi: 10.1002/9780470691953.ch19
Cooper, R. A., Valentine, P., Uzmann, J. R., and Slater, R. A. (1987a). “Submarine canyons,” in Georges Bank, ed. R. H. Backus (Cambridge, MA: MIT Press), 52–63.
Cooper, R. A., Shepard, A., Valentine, P., Uzmann, J. R., and Hulbert, A. (1987b). Pre- and post-drilling benchmarks and monitoring data of ocean floor fauna, habitats, and contaminant loads on Georges Bank and its submarine canyons. NOAA Symp. Ser. Undersea Res. 2, 17–48.
Costello, T. C. (2019). Are Marine National Monuments situated on lands owned or controlled by the Government of the United States. Ocean Coast. Law J. 24, 219–249.
Coykendall, D. K., Hererra, S., Nizinski, M. S., Waller, R. G., Brooke, S., et al. (2016). “Contrasting? patterns of population genetic connectivity in octocorals from the northern Atlantic Ocean,” in Proceedings of the Oral Presentation. 6th International Symposium Deep Sea Corals, Boston.
Davison, P. C., Checkley, D. M., Koslow, J. A., and Barlow, J. (2013). Carbon export mediated by mesopelagic fishes in the northeast Pacific Ocean. Prog. Oceanogr. 116, 14–30. doi: 10.1016/j.pocean.2013.05.013
CRS. (2018). National monuments and the antiquities act. Congressional Research Service. Report R41330. Ver. 30. Available online at: http://crsreports.congress.gov (accessed 30 November 2018).
DeAngelis, A., Stanistreet, J., Baumann-Pickering, S., and Cholewiak, D. (2018). A description of echolocation clicks recorded in the presence of True’s beaked whale (Mesoplodon mirus). J. Acoust. Soc. Am. 144, 2691–2700. doi: 10.1121/1.5067379
Devine, J. A., Baker, K. D., and Haedrich, R. L. (2006). Fisheries: deep-sea fishes qualify as endangered. Nature 439:29. doi: 10.1038/439029a
Doughty, C. E., Roman, J., Faurby, S., Wolf, A., Haque, A., Bakker, E. S., et al. (2016). Global nutrient transport in a world of giants. Proc. Natl. Acad. Sci. U.S.A. 113, 868–873. doi: 10.1073/pnas.1502549112
Eilperin, J. (2016). Obama Designates the First-Ever Marine Monument Off the East Coast, in New England. Washington Post. Available at https://www.washingtonpost.com/news/energy-environment/wp/2016/09/15/obama-to-designate-the-first-ever-marine-monument-off-the-east-coast-in-new-england/ (accessed September 15, 2016).
Eilperin, J. (2018). Trump Administration Officials Dismissed Benefits of National Monuments. Washington Post. Available at https://www.washingtonpost.com/national/health-science/trump-administration-officials-dismissed-benefits-of-national-monuments/2018/07/23/5b8b1666-8b9a-11e8-a345-a1bf7847b375_story.html (accessed July 23, 2018).
Ezer, T. (1994). On the Interaction between the Gulf Stream and the New England Seamount Chain. Journal of Physical Oceanography 24, 191–204. doi: 10.1175/1520-0485(1994)024<0191:otibtg>2.0.co;2
Fedderly, E. (2018). Fishing Groups Lose Legal Battle Over Marine Monument. Pasadena, CA: Courthouse News Service.
Federal Register (2016). Proclamation 9496 of September 15, 2016 from President Barack Obama. Northeast Canyons and Seamounts Marine National Monument. Federal Register 81 FR 65159. Washington, DC: Federal Register, 65159–65167.
Finkbeiner, E. M., Wallace, B. P., Moore, J. E., Lewison, R. L., Crowder, L. B., and Read, A. J. (2011). Cumulative estimates of sea turtle bycatch and mortality in USA fisheries between 1990 and 2007. Biol. Conserv. 144, 2719–2727. doi: 10.1016/j.biocon.2011.07.033
France, S. C. (2007). Genetic analysis of bamboo corals (Cnidaria: Octocorallia: Isididae): does lack of colony branching distinguish Lepidisis from Keratoisis? Bull. Mar. Sci. 81, 323–333.
Frazin, R. (2020). Trump Administration Sued Over Marine Monument Rollback. The Hill. Available online at: https://thehill.com/policy/energy-environment/503142-trump-administration-sued-over-marine-monument-rollback (accessed June 17, 2020).
Garrison, L. P., and Stokes, L. (2014). Estimated Bycatch of Marine Mammals and Sea Turtles in the U.S. Atlantic pelagic longline fleet during 2013. NOAA Technical Memorandum NMFS-SEFSC-667. Silver Spring, MD: NOAA.
Getis, A., and Ord, J. K. (1992). The analysis of spatial association by use of distance statistics. Geograph. Anal. 24, 189–206. doi: 10.1111/j.1538-4632.1992.tb00261.x
Greene, C. H., Wiebe, P. H., Burczynski, J., and Youngbluth, M. J. (1988). Acoustical detection of high-density krill demersal layers in the submarine canyons off Georges Bank. Science 241, 359–361. doi: 10.1126/science.241.4863.359
Griffin, R. B. (1997). Relationships between odontocete distributions and zooplankton community structure along the southern edge of Georges Bank. J. Northw. Atl. Fish. Sci. 22, 27–36. doi: 10.2960/j.v22.a2
Griffin, R. B. (1999). Sperm whale distributions and community ecology associated with a warm core ring off Georges Bank. Mar. Mamm. Sci. 15, 33–51. doi: 10.1111/j.1748-7692.1999.tb00780.x
Hain, J. W., Hyman, M. A. M., Kenney, R. D., and Winn, H. E. (1985). The role of cetaceans in the shelf-edge of the northeastern United States. Mar. Fish. Rev. 47, 13–17.
Hatch, J. M. (2017). Comprehensive Estimates of Seabird-Fishery Interactions for the US Northeast and Mid-Atlantic, Aquatic Conservation: Marine and Freshwater Ecosystems. 1–12. Available online at: http://onlinelibrary.wiley.com/doi/10.1002/aqc.2812/abstract (accessed 15 December 2018).
Hecker, B. (1990a). Photographic evidence for the rapid flux of particles to the sea floor and their transport down the continental slope. Deep Sea Res. 37, 1773–1782. doi: 10.1016/0198-0149(90)90077-9
Hecker, B. (1990b). Variation in megafaunal assemblages on the continental margin south of New England. Deep Sea Res. 37, 37–57. doi: 10.1016/0198-0149(90)90028-t
Hecker, B., Blechschmidt, G., and abd Gibson, P. (1980). Epifaunal Zonation and Community Structure in Three Mid- and North Atlantic Canyons Final Report: Canyon Assessment Study in the Mid- and North Atlantic Areas of the US Outer Continental Shelf. Washington, DC: U.S. Department of the Interior.
Hobson, E. S., Greene, C. H., and Wiebe, P. H. (1989). Predation on ocean krill. Science 243, 237–239.
Holden, E. (2020). ‘Selling Off the Future’: Trump Allows Fishing in Marine Monument. Available online at: https://www.theguardian.com/environment/2020/jun/06/donald-trump-commercial-fishing-marine-monument (accessed June 6, 2020).
Holland, K. N., and Grubbs, R. D. (2007). “Fish visitors to seamounts:tunas and billfish at seamounts,” in Seamounts: Ecology, Conservation and Fisheries, eds T. J. Pitcher, T. Morato, and P. J. B. Hart (Oxford: Blackwell), 189–201. doi: 10.1002/9780470691953.ch10a
Irigoien, X., Klevjer, T. A., Røstad, A., Martinez, U., Boyra, G., Acuña, J. L., et al. (2014). Large mesopelagic fishes biomass and trophic efficiency in the open ocean. Nat. Commun. 5:3271.
Jegalian, K. (1999). Plan would protect New England Coast. Science 284, 237–237. doi: 10.1126/science.284.5412.237
Kaschner, K. (2007). “Air-breathing visitors to seamounts: marine mammals,” in Seamounts: Ecology, Conservation and Fisheries, eds T. J. Pitcher, T. Morato, and P. J. B. Hart (Oxford: Blackwell), 230–238. doi: 10.1002/9780470691953.ch12a
Kelly, N. E., Shea, E. K., Metaxas, A., Haedrich, R. L., and Auster, P. (2010). Biodiversity of the deep-sea continental margin bordering the Gulf of Maine (NW Atlantic): relationships among sub-regions and to shelf systems. PLoS One 5:e13832. doi: 10.1371/journal.pone.0013832
Kenney, R. D., and Winn, H. E. (1986). Cetacean high-use habitats of the northeast United States continental shelf. Fish. Bull. U.S. 84, 345–357.
Kenney, R. D., and Winn, H. E. (1987). Cetacean biomass densities near submarine canyons compared to adjacent shelf/slope areas. Continent. Shelf Res. 7, 107–114. doi: 10.1016/0278-4343(87)90073-2
Kilgour, M. J., Auster, P. J., Packer, D., and Watling, L. (2016). “Variation in seafloor communities across the western new england seamounts and adjacent submarine canyons: implications for conservation,” in Proceedings of the International Deep-Sea Coral Symposium, Boston.
Koslow, J. A., Auster, P., Bergstad, O. A., Roberts, J. M., Rogers, A., Vecchione, M., et al. (2016). “Biological communities on seamounts and other submarine features potentially threatened by disturbance,” in The First Global Integrated Marine Assessment, World Ocean Assessment I, Chap. 51, eds L. Inniss and A. Simcock (New York, NY: Springer), 1–26.
Koslow, J. A., Gowlett-Holmes, K., Lowry, J., O’Hara, T., Poore, G., and Williams, A. (2001). The seamount benthic macrofauna off southern Tasmania: community structure and impacts of trawling. Mar. Ecol. Prog. Ser. 213, 111–125. doi: 10.3354/meps213111
Kraus, S. D., Auster, P. J., Witman, J. D., Wikgren, B., McKee, M. P., and Lamb, R. W. (2016). Scientific assessment of a proposed marine national monument off the northeast United States. Science Briefing for Press and Interested Parties. Final version 31 March 2016. Available online at: https://www.researchgate.net/profile/Peter_Auster/publication/299559963
Lavery, T. J., Roudnew, B., Seymour, J., Mitchell, J. G., Smetacek, V., and Nicol, S. (2014). Whales sustain fisheries: blue whales stimulate primary production in the Southern Ocean. Mar. Mamm. Sci. 30, 888–904. doi: 10.1111/mms.12108
Lewison, R. L., Crowder, L. B., Wallace, B. P., Moore, J. E., Cox, T., Zydelis, R., et al. (2014). Global patterns of marine mammal, seabird, and sea turtle bycatch reveal taxa-specific and cumulative megafauna hotspots. Proc. Natl. Acad. Sci. U.S.A. 111, 5271–5276. doi: 10.1073/pnas.1318960111
Lewison, R. L., Freeman, S. A., and Crowder, L. B. (2004). Quantifying the effects of fisheries on threatened species: the impact of pelagic longlines on loggerhead and leatherback sea turtles. Ecol. Lett. 7, 221–231. doi: 10.1111/j.1461-0248.2004.00573.x
Litvinov, F. (2007). “Fish visitors to seamounts: aggregations of large pelagic sharks above seamounts,” in Seamounts: Ecology, Conservation and Fisheries, eds T. J. Pitcher, T. Morato, and P. J. B. Hart (Oxford: Blackwell), 202–206. doi: 10.1002/9780470691953.ch10b
McMurray, J. (2016). On the Proposed New England Marine Monument. Marine Fish Conservation Network. Available online at: https://conservefish.org/2016/06/28/proposed-new-england-marine-monument/ (accessed June 28, 2016).
Mills, K. E., Pershing, A. J., Brown, C. J., Chen, Y., Chiang, F. S., Holland, D. S., et al. (2013). Fisheries management in a changing climate: lessons from the 2012 ocean heat wave in the Northwest Atlantic. Oceanography 26, 191–195.
Mills, S. (2003). Seamount Coral Communities. NOAA Ocean Explorer Mountains in the Sea. Available at http://oceanexplorer.noaa.gov/explorations/03mountains/background/larvae/larvae.html (accessed 20 June 2018).
Mitchell, J. (2016). Opinion. Marine Monument Designation: New Bedford Mayor Jon Mitchell — Restore Access to Process Where Fisheries are Concerned. SouthCoast Today. Available online at: https://www.southcoasttoday.com/opinion/20160826/marine-monument-designation-new-bedford-mayor-jon-mitchell—restore-access-to-process-where-fisheries-are-concerned (accessed 26 August 2016).
Moore, J. A., Vecchione, M., Collette, B. B., Gibbons, R., Hartel, K. E., Galbraith, J. K., et al. (2003). Biodiversity of Bear Seamount, New England seamount chain: results of exploratory trawling. J. Northwest Atlant. Fish. Sci. 31, 363–372. doi: 10.2960/j.v31.a28
Moore, J. E., Wallace, B. P., Lewison, R. L., Žydelis, R., Cox, T. M., and Crowder, L. B. (2009). A review of marine mammal, sea turtle and seabird bycatch in USA fisheries and the role of policy in shaping management. Mar. Policy 33, 435–451. doi: 10.1016/j.marpol.2008.09.003
Moors-Murphy, H. B. (2014). Submarine canyons as important habitat for cetaceans, with special reference to the Gully: a review. Deep Sea Res. II Top. Stud. Oceanogr. 104, 6–19. doi: 10.1016/j.dsr2.2013.12.016
Morato, T., Watson, R., Pitcher, T. J., and Pauly, D. (2006). Fishing down the deep. Fish Fish. 7, 24–34. doi: 10.1111/j.1467-2979.2006.00205.x
NEFMC (2017). Draft Omnibus Deep-Sea Coral Amendment Including a Draft Environmental Assessment. Newburyport, MA: New England Fishery Management Council.
NEFMC (2019). Section 6.2 Coral species of the New England region, In Omnibus Deep-Sea Coral Amendment Including a Final Environmental Assessment. Newburyport, MA: New England Fishery Management Council.
NEFSC (2010). Northeast Fisheries Science Center Benthic Database. Woods Hole, MA: Northeast Fisheries Science Center.
NEFSC (2016). Field Fresh: NEFSC Science in Motion website. A Record Day! Posted August 12, 2016. Available online at: https://nefsc.wordpress.com/category/amapps/ (accessed 27 June, 2017).
NOAA (2013). Chemosynthetic Communities and Gas Hydrates at Cold Seeps South of Nantucket. Available online at: http://oceanexplorer.noaa.gov/okeanos/explorations/ex1304/logs/july12/july12.html (accessed July 12, 2013).
Norse, E. A., Brooke, S., Cheung, W. W., Clark, M. R., Ekeland, I., Froese, R., et al. (2012). Sustainability of deep-sea fisheries. Mar. Policy 36, 307–320.
Northridge, S. (1996). Estimation of Cetacean Mortality in the US Atlantic Swordfish and Tuna Driftnet and Pair Trawl Fisheries. Silver Spring, MD: NMFS.
NRDC (2019). The Northeast Canyons and Seamounts Marine National Monument: Impacts On The Commercial Fisheries (Updated September 2019). New York, NY: Natural Resources Defense Council.
Packer, D. B., Boelke, D., Guida, V., and McGee, L. (2007). “State of deep coral ecosystems in the Northeastern US region: maine to cape hatteras,” in The State of Deep Coral Ecosystems of the United States, Chap. 5, eds S. E. Lumsden, T. F. Hourigan, A. W. Bruckner, and G. Dorr (Silver Spring, MD: NOAA).
Palka, D. (2012). Cetacean Abundance Estimates in US Northwestern Atlantic Ocean Waters from Summer 2011 Line Transect Survey. US Dept Commer, Northeast Fish Sci Cent Ref Doc. Woods Hole, MA: National Marine Fisheries Service, 12–29.
Payne, P. M., and Heinemann, D. W. (1993). The distribution of pilot whales (Globicephala sp.) in shelf/shelf edge and slope waters of the northeastern United States, 1978-1988. Rep. Int. Whaling Commiss. 14, 51–68.
Powers, K. D. (1984). Pelagic Distributions of Marine Birds Off the Northeastern United States. Silver Spring, MD: NOAA.
Quattrini, A. M., Nizinski, M. S., Chaytor, J. D., Demopoulos, A. W. J., Roark, E. B., France, S. C., et al. (2015). Exploration of the canyon incised continental margin of the northeastern United States reveals dynamic habitats and diverse communities. PLoS One 10:e0139904. doi: 10.1371/journal.pone.0139904
Reeves, R. R., McClellan, K., and Werner, T. B. (2013). Marine mammal bycatch in gillnet and other entangling net fisheries, 1990 to 2011. Endang. Spec. Res. 20, 71–97. doi: 10.3354/esr00481
Riley, S. J., DeGloria, S. D., and Elliot, R. (1999). A terrain ruggedness index that quantifies topographic heterogeneity. Int. J. Sci. 5, 23–27.
Roberts, C. M., O’Leary, B. C., McCauley, D. J., Cury, P. M., Duarte, C. M., Lubchenco, J., et al. (2017). Marine reserves can mitigate and promote adaptation to climate change. Proc. Natl. Acad. Sci. U.S.A. 114, 6167–6175. doi: 10.1073/pnas.1701262114
Roman, J., Estes, J. A., Morissette, L., Smith, C., Costa, D., McCarthy, J., et al. (2014). Whales as marine ecosystem engineers. Front. Ecol. Environ. 12, 377–385. doi: 10.1890/130220
Roman, J., and McCarthy, J. J. (2010). The whale pump: marine mammals enhance primary productivity in a coastal basin. PLoS One 5:e13255. doi: 10.1371/journal.pone.0013255
Rossman, M. C. (2010). Estimated bycatch of small cetaceans in northeast US bottom trawl fishing gear during 2000-2005. J. Northwest Atlantic Fish. Sci. 42, 77–101. doi: 10.2960/j.v42.m650
Ryan, T. (2019). Atlantic Ocean’s First Marine Monument Survives Court Challenge. Available online at: https://www.courthousenews.com/atlantic-oceans-first-marine-monument-survives-court-challenge/ (accessed 28 Dec 2019).
Ryan, W. B. F., Cita, M. B., Miller, E. L., Hanselman, D., Nesteroff, W. D., Hecker, B., et al. (1978). Bedrock geology in New England submarine canyons. Oceanol. Acta 1, 233–254.
Sackett, D. C., Kelley, C. D., and Drazen, J. C. (2017). Spilling over deepwater boundaries: evidence of spillover from two deepwater restricted fishing area in Hawaii. Mar. Ecol. Prog. Ser. 568, 175–190. doi: 10.3354/meps12049
Santos, M. A., Bolten, A. B., Martins, H. R., Riewald, B., and Bjorndal, K. A. (2007). “Air-breathing visitors to seamounts: sea turtles,” in Seamounts: Ecology, Conservation and Fisheries, eds T. J. Pitcher, T. Morato, and P. J. B. Hart (Oxford: Blackwell), 239–244. doi: 10.1002/9780470691953.ch12b
Schmitz, O. J., Raymond, P. A., Estes, J. A., Kurz, W. A., Holtgrieve, G. W., Ritchie, M. E., et al. (2014). Animating the carbon cycle. Ecosystems 17:344. doi: 10.1007/s10021-013-9715-7
Schorr, G. S., Falcone, E. A., Moretti, D. J., and Andrews, R. D. (2014). First long-term behavioral records from Cuvier’s beaked whales (Ziphius cavirostris) reveal record-breaking dives. PLoS One 9:e92633. doi: 10.1371/journal.pone.0092633
Sen, S. (2010). Developing a framework for displaced fishing effort programs in marine protected areas. Mar. Policy 34, 1171–1177. doi: 10.1016/j.marpol.2010.03.017
Shank, T. M. (2010). Seamounts: deep-ocean laboratories of faunal connectivity, evolution, and endemism. Oceanography 23, 108–122. doi: 10.5670/oceanog.2010.65
Skahill, P. (2016). Could New England Host the Atlantic Ocean’s First-Ever Marine National Monument? Washington, DC: National Public Radio.
Skarke, A., Ruppel, C., Kodis, M., Brothers, D., and Lobecker, E. (2014). Widespread methane leakage from the sea floor on the northern US Atlantic margin. Nat. Geosci. 7, 657–661. doi: 10.1038/ngeo2232
Sloan, N. A. (2002). History and application of the wilderness concept in marine conservation. Conserv. Biol. 16, 294–305. doi: 10.1046/j.1523-1739.2002.00071.x
Smith, C. R., and Baco, A. R. (2003). “Ecology of whale falls at the deep-sea floor,” in Oceanography and Marine Biology: An Annual Review, eds R. N. Gibson and R. J. A. Atkinson (Boca Raton, FL:CRC Press), 311–354.
Snow, N. (2017). BOEM will resume evaluating requests to study Atlantic OCS potential. Oil Gas J. Available online at: https://www.ogj.com/general-interest/article/17290071/boem-will-resume-evaluating-requests-to-study-atlantic-ocs-potential (accessed 11 May 2017).
Soetaert, K., Mohn, C., Rengstorf, A., Grehan, A., and van Oevelen, D. (2016). Ecosystem engineering creates a direct nutritional link between 600-m deep cold-water coral mounds and surface productivity. Sci. Rep. 6:35057. doi: 10.1038/srep35057
Speer, L., Chasis, S., Azimi, S., and Luijten, J. (2001). Priority Areas for Protection in the Mid-Atlantic: Findings of NRDC’s Marine Habitat Workshop. New York, NY: Natural Resources Defense Council.
St John, M. A., Borja, A., Chust, G., Heath, M., Grigorov, I., Mariani, P., et al. (2016). A dark hole in our understanding of marine ecosystems and their services: perspectives from the mesopelagic community. Front. Mar. Sci. 3:31. doi: 10.3389/fmars.2016.00031
Stanistreet, J. E., Nowacek, D. P., Baumann-Pickering, S., Bell, J. T., Cholewiak, D. M., Hildebrand, J. A., et al. (2017). Using passive acoustic monitoring to document the distribution of beaked whale species in the western North Atlantic Ocean. Can. J. Fish. Aquatic Sci. 74, 2098–2109. doi: 10.1139/cjfas-2016-0503
Stanistreet, J. E., Nowacek, D. P., Bell, J. T., Cholewiak, D. M., Hildebrand, J. A., Hodge, L. E. W., et al. (2018). Spatial and seasonal patterns in acoustic detections of sperm whales Physeter macrocephalus along the continental slope in the western North Atlantic Ocean. Endang. Spec. Res. 35, 1–13. doi: 10.3354/esr00867
Thoma, J. N., Pante, E., Brugler, M. R., and France, S. C. (2009). Deep-sea octocorals and antipatharians show no evidence of seamount-scale endemism in the NW Atlantic. Mar. Ecol. Prog. Ser. 397, 25–35. doi: 10.3354/meps08318
Thompson, D. R. (2007). “Air-breathing visitors to seamounts: importance of seamounts to seabirds,” in Seamounts: Ecology, Conservation and Fisheries, eds T. J. Pitcher, T. Morato, and P. J. B. Hart (Oxford: Blackwell), 245–251. doi: 10.1002/9780470691953.ch12c
United States Congress (1906). An Act for the preservation of American Antiquities. Washington, DC: United States Congress.
Valentine, P. C. (1987). The shelf-slope transition; canyon and upper slope sedimentary processes on the southern margin of Georges Bank. USGS Bull. 1782, 4–29.
van Oevelen, D., Duineveld, G., Lavaleye, M., Mienis, F., Soetaert, K., and Heip, C. H. (2009). The cold-water coral community as hotspot of carbon cycling on continental margins: a food-web analysis from Rockall Bank (northeast Atlantic). Limnol. Oceanogr. 54, 1829–1844. doi: 10.4319/lo.2009.54.6.1829
Waller, R., Watling, L., Auster, P., and Shank, T. (2007). Anthropogenic impacts on the Corner Rise Seamounts, NW Atlantic Ocean. J. Mar. Biol. Assoc. U.K. 87, 1075–1076. doi: 10.1017/s0025315407057785
Waring, G. T. (1998). Results of the 1991 R/V Chapman Marine Mammal Sighting Survey. Northeast Fish. Sci. Cent. Ref Doc. 98-09. Woods Hole, MA: National Marine Fisheries Service.
Waring, G. T., Hamazaki, T., Sheehan, D., Wood, G., and Baker, S. (2001). Characterization of beaked whale (Ziphiidae) and sperm whale (Physeter macrocephalus) summer habitat in shelf-edge and deeper waters off the northeast US. Mar. Mamm. Sci. 17, 703–717. doi: 10.1111/j.1748-7692.2001.tb01294.x
Watling, L., France, S. C., Pante, E., and Simpson, A. (2011). Biology of deep-water octocorals. Adv. Mar. Biol. 60, 41–122. doi: 10.1016/b978-0-12-385529-9.00002-0
Watson, R. A., Nowara, G. B., Hartmann, K., Green, B. S., Tracey, S. R., and Carter, C. G. (2015). Marine foods sourced from farther as their use of global ocean primary production increases. Nat. Commun. 6:7365. doi: 10.1038/ncomms8365
Wenzel, F. W., Polloni, P. T., Craddock, J. E., Gannon, D. P., Nicolas, J. R., Read, A. J., et al. (2013). Food habits of Sowerby’s beaked whales (Mesoplodon bidens) taken in the pelagic drift gillnet fishery of the western North Atlantic. Fish. Bull. 111, 381–389. doi: 10.7755/fb.111.4.7
Werner, T. B., Northridge, S., McClellan Press, K., and Young, N. (2015). Mitigating bycatch and depredation of marine mammals in longline fisheries. ICES J. Mar. Sci. 72, 1576–1586. doi: 10.1093/icesjms/fsv092
White, C. (2016). Fishermen Considering Legal Steps to Alter Marine Monument in Northeast US SeafoodSource. Available online at: https://www.seafoodsource.com/news/supply-trade/fishermen-pursuing-legal-steps-to-alter-marine-monument-in-northeast-u-s (accessed September 21, 2016).
White, M. I, Bashmachnikov, J., and Martins, A. (2007). “Physical processes and seamount productivity,” in Seamounts: Ecology, Fisheries & Conservation, ed. P. Pitcher (Oxford: Blackwell Publishing), 65–84.
Winter, A. L., Jiao, Y., and Browder, J. A. (2011). Modeling low rates of seabird bycatch in the U.S. Atlantic longline fishery. Waterbirds 34, 289–303. doi: 10.1675/063.034.0304
Keywords: marine reserve, marine protected area, benthic, marine mammal, diversity, hotspot, seafloor complexity, geospatial
Citation: Auster PJ, Hodge BC, McKee MP and Kraus SD (2020) A Scientific Basis for Designation of the Northeast Canyons and Seamounts Marine National Monument. Front. Mar. Sci. 7:566. doi: 10.3389/fmars.2020.00566
Received: 05 May 2020; Accepted: 19 June 2020;
Published: 10 July 2020.
Edited by:
Rob Harcourt, Macquarie University, AustraliaReviewed by:
Elizabeth M. De Santo, Franklin & Marshall College, United StatesMiriam C. Balgos, Global Ocean Forum, United States
Copyright © 2020 Auster, Hodge, McKee and Kraus. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Peter J. Auster, cGF1c3RlckBteXN0aWNhcXVhcml1bS5vcmc=; cGV0ZXIuYXVzdGVyQHVjb25uLmVkdQ==