 Giacomo Milisenda1*
Giacomo Milisenda1* Germana Garofalo2
Germana Garofalo2 Fabio Fiorentino2
Fabio Fiorentino2 Francesco Colloca1,3
Francesco Colloca1,3 Francesc Maynou4
Francesc Maynou4 Alessandro Ligas5Claudia Musumeci5Luís Bentes6
Alessandro Ligas5Claudia Musumeci5Luís Bentes6 Jorge M. S. Gonçalves6
Jorge M. S. Gonçalves6 Karim Erzini6
Karim Erzini6 Tommaso Russo7
Tommaso Russo7 Lorenzo D’Andrea7
Lorenzo D’Andrea7 Sergio Vitale2
Sergio Vitale2- 1Dipartimento di Ecologia Marina Integrata, Stazione Zoologica Anton Dohrn di Napoli, Napoli, Italy
- 2National Research Council – Institute of Marine Biological Resources and Biotechnologies (CNR IRBIM), Mazara del Vallo, Italy
- 3Department of Biology and Biotechnology ‘Charles Darwin’, Sapienza University of Rome, Rome, Italy
- 4Institut de Ciències del Mar (CSIC), Barcelona, Spain
- 5Consorzio per il Centro Interuniversitario di Biologia Marina ed Ecologia Applicata G. Bacci, Livorno, Italy
- 6Centro de Ciências do Mar (CCMAR), Universidade do Algarve, Faro, Portugal
- 7Laboratory of Experimental Ecology and Aquaculture, Department of Biology, University of Rome Tor Vergata, Rome, Italy
The recent establishment of the “landing obligation” under the reformed EU Common Fishery Policy has the twofold objective of reducing the excessive practice of discarding unwanted catch at sea and encouraging more selective and sustainable fisheries. Within this context, the awareness of the spatial distribution of potential unwanted catches is important for devising management measures aimed to decrease discards. This study analyzed the distribution of Hot Spot density areas of demersal fish and crustaceans below the Minimum Conservation Reference Size (MCRS) in four different southern European seas: continental Portuguese coast, Catalan Sea, South of Sicily, Liguria and northern Tyrrhenian Seas using both bottom trawl survey data and information on the spatial distribution of commercial fisheries. Critical areas for discarding were identified as zones where the highest densities of individuals below MCRS were consistently recorded throughout a series of years. Results clearly showed a patchy distribution of undersized individuals in each investigated area, highlighting the overlap between high density patches of both discards and fishing effort. The present findings provide a relevant knowledge for supporting the application of spatial-based management actions, such as the designation of Fisheries Restricted Areas (FRAs), in order to minimize the by-catch of undersized specimens and improve the sustainability of demersal fisheries.
Introduction
Unwanted Catches (UCs) are common and critical problems in fisheries worldwide, as they can alter marine biodiversity and at the same time could worsen overexploitation of commercial species (Kelleher, 2005). UCs constitute the portion of the total catch that is returned to the sea, and typically include undersized individuals of commercial species and non-marketable species or with low market value (Alverson et al., 1994; Kelleher, 2005; Bellido et al., 2011). Recent assessment indicate as more than 40 percent of global catch is discarded outboard (Davies et al., 2009). The quantity of the UCs mainly depends on the characteristics of fishing grounds (e.g., geographic area, type of habitat, abundance of either juveniles of commercial species or non-commercial species etc.) and on the fishing gears used (Catchpole et al., 2005; Kelleher, 2005). In the Mediterranean Sea, demersal trawling is responsible for about 50 percent of the total discard produced (STECF, 2006), highlighting the low level of selectivity of this sector (FAO, 2016), while a lower discard rate characterizes other gear belonging to industrial pelagic fishing, such as the pelagic trawl or purse seine. As regards artisanal fisheries, few data have been collected to date on the discarded fraction, but the first results obtained show that this sector has a higher level of selectivity (FAO, 2019). Other factors influencing the UCs production include technical characteristics of the vessels and gear (e.g., mesh size) (Graham et al., 2008), fishing strategies (Lleonart and Maynou, 2003), fishing season and environmental conditions (Milisenda et al., 2017b). Nonetheless, “the decision of discarding part of the catch is a choice that fishermen ultimately take, and is mainly driven by economic reasons or fishing regulations” (Batsleer et al., 2013; Condie et al., 2014).
Under the current trawl net selectivity patterns used in southern European fisheries, undersized individuals of some important commercial species, such as European hake (Merluccius merluccius) and Atlantic horse mackerel (Trachurus trachurus), predominate in the discard (Milisenda et al., 2017b; Damalas et al., 2018; Despoti et al., 2020), especially during the recruitment period (FAO, 2016). The combined provisions of art. 14 on “Avoidance and minimization of unwanted catches” and art. 15 on “Landing Obligation – LO” under the reformed Common Fisheries Policy – CFP (Reg. EC n. 1380/2013) has the objective of reducing the excessive practice of discarding unwanted catch at sea. The new Common Fisheries Policy promotes the technical measures aimed at reducing and discouraging the capture of specimens below the Minimum Conservation Reference Size (MCRS, Reg. EC No 1967/2006), demanding from fishermen also to land all the catches of regulated species. Given the specific characteristics of each Mediterranean sub-basin and the objective of moving toward regional-based fisheries management in the reformed CFP, “local management plans are needed as solutions to set up the most appropriate measures at regional level” (Vitale et al., 2016). In the southern European waters, both Mediterranean and Atlantic, there are many measures, both regulatory and technical, aimed to minimize unwanted catch including, for instance the discard ban, gear selectivity improvement, fisheries restricted areas, and “monitoring and control measures” (Karp et al., 2019; Common Fisheries Policy – CFP).
Even the Marine Spatial Planning (MSP) must take into account the reduction of discarding and its spatial dynamics, i.e., where and when the natural stocks and fleets interact (Maiorano et al., 2009; Bellido et al., 2019; D’Andrea et al., 2020). One possible approach to reduce discards is to avoid areas and seasons where UCs is more likely (O’Keefe et al., 2013; Vilela and Bellido, 2015; Paradinas et al., 2016), and for this reason data on the spatial distribution of undersized organisms is an important source of information to reach lower discard production at the regional level (Vitale et al., 2016). Highlighting the distribution and persistence of areas where high abundance of organisms under the MCRS is present can support the designation of no-take zones or Fishery Restricted Areas (FRAs) and their inclusion in conservation networks (Colloca et al., 2015). The GFCM has already started working in this direction, decreeing the formation of different FRAs in the Mediterranean Sea. Three of these are located in the Strait of Sicily as part of the multiannual management plan for hake and deep-water rose shrimp fisheries (“East of Adventure Bank,” “West of Gela basin,” and “East of Malta bank”) and another one in the Adriatic Sea (“Jabuka/Pomo Pit”) as an important essential fish habitat for hake and Norway lobster (Nephrops norvegicus). Besides areas closed to trawling according to the EC Reg. 1967/2006 (e.g., 3 miles or 1.5 miles from the coast/50 m isobath), two FRAs (Fishing Restricted Areas) are present in geographical subarea 9 of Mediterranean Sea (Ligurian Sea and Northern Tyrrhenian Sea): (1) Off the coasts of Argentario promontory (50 km2, from 160 to 220 m depth); (2) in the Gulf of Gaeta (125 km2 from 100 to 200 m depth). Both FRAs are located in areas with high concentration of juveniles (nursery areas) of European hake and deep-water rose shrimp. According to the Italian Ministry (MIPAAF) Decree (22nd January 2009), fishing activities with towed gears are forbidden in the two FRAs from January 1st to June 30th. In May 2020, in the West Med Multiannual Plan (WM MAP) Spain established two FRAs located in the geographical subareas 2 and 5. In the Portuguese Atlantic area, two Fishing Restricted Areas are established. These two FRAs are limited temporal closures, from the first day of December until the last day of February for hake and from the beginning of May to the end of August for Norway lobster. The main objective of both FRAs is to protect juveniles and their spawning stock of these two species from trawling or any other fishing activities using towed gears. Moreover, in the Mediterranean Sea there is an increasing interest in spatial based measures as management tools for achieving several objectives related to fisheries sustainability, such as species conservation, reduction of trawl impact on demersal community, restoration of essential fish habitat and preservation of vulnerable marine ecosystem (Fiorentino et al., 2003; Maiorano et al., 2009; De Juan and Demestre, 2012; Russo et al., 2014b, 2019; Pipitone et al., 2014; Colloca et al., 2015). Of course, the rationale of these measures is inspired by the fact that fishing effort pattern, at the scales of both individual vessels and fleets, is highly heterogeneous in space and time (Russo et al., 2013, 2019).
The aim of this paper is to identify critical areas for the production of UCs (and therefore, implicitly and intuitively, for the occurrence of discards) in four important fishing areas of the southern European waters, including both Mediterranean Sea and Atlantic Ocean off the Portuguese coast. These areas are detected by matching hotspots of high densities of individuals below MCRS, with high levels of trawling effort. The output of this study could provide a base for supporting the application of spatial based fishery management in order to minimize UCs and definitively improve the sustainability of demersal fisheries.
Materials and Methods
Source of Data
The study was conducted in four different areas (Figure 1), three in the Mediterranean Sea: the Catalan Sea (Area 1), the Central Mediterranean (i.e., the South of Sicily) (Area 2) the Ligurian and northern Tyrrhenian Seas (Area 3), and one in the north-east Atlantic Ocean: the continental Portuguese coast (Area 4).
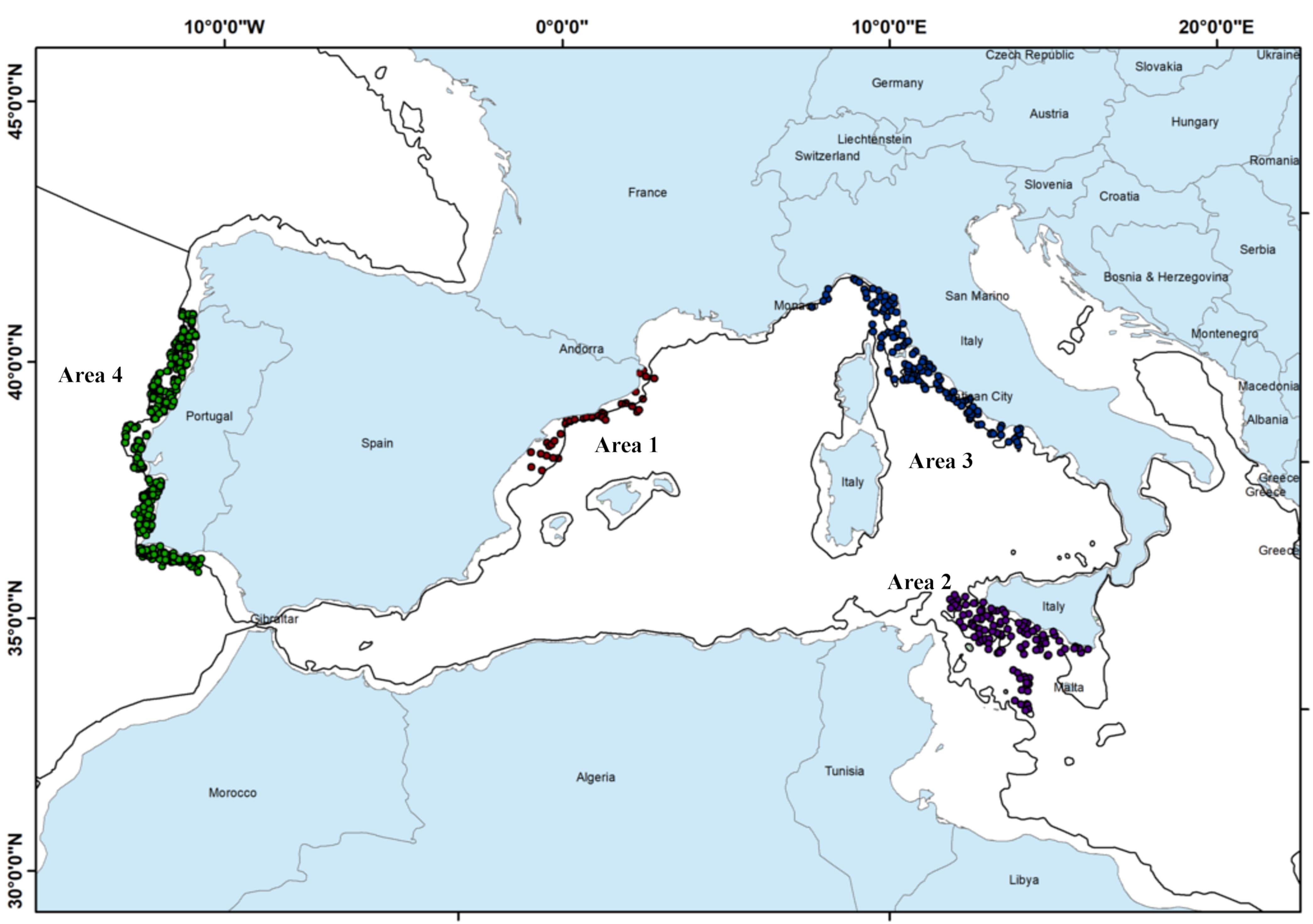
Figure 1. Location of the study areas: Area 1 – Catalan Sea, Area 2 – Strait of Sicily, Area 3 – Ligurian and northern Tyrrhenian Seas, Area 4 – Portuguese area. Dots = hauls locations; Black line = 200 m depth.
Two different data sources were used to gather information on the spatial distribution of specimens below MCRS within each zone: the International Bottom Trawl Surveys (IBTS) for the Portuguese waters, and the Mediterranean International Bottom Trawl Survey (MEDITS) for the Spanish and Italian areas, respectively. The data used in the areas 2, 3, and 4 were provided directly by the partners of the present work, as they are part of the institutes in charge of data collection under the European Data Collection Framework (National Research Council – CNR, Consorzio per il Centro Interuniversitario di Biologia Marina ed Ecologia Applicata G. Bacci – CIBM, and the Centro de Ciências do Mar – CCMAR), while the data used in the area 1 was requested to the European commission, as these data were collected by the Instituto Español de Oceanografía (IEO). The IBTS time series spanned between 2002 and 2011 (661 hauls). MEDITS time series spanned between 2009 and 2014 for the Catalan Sea (202 hauls) and the Ligurian and northern Tyrrhenian Seas (720 hauls). In the South of Sicily, the time series was 2009–2013 (600 hauls).
The Portuguese IBTS (PT-IBTS) is carried out yearly in Portuguese continental waters at the beginning of the 4th quarter (October) between depths of 20–500 m. The main objective of this survey is to estimate recruitment indices of European hake and horse mackerel. It uses a standardized trawl net (Norwegian Campell Trawl 1800/96 NCT) with a mean vertical opening of 4.6 m, mean horizontal opening of 15 m, and 20 mm stretched mesh size in the cod-end. Tow duration is 30 min at a mean speed of 3.5 knots (ICES, 2017). MEDITS is carried out in EU Mediterranean waters in late spring-early summer (May–July) to gather data on the wide range of species characterizing the Mediterranean demersal communities between depths of 10 and 800 m. The survey adopts a standardized trawl net (GOC 73) with vertical opening of about 2.5 m, mean horizontal opening of 18 m and 20 mm stretched mesh size in the cod-end, and a standardized fishing duration of 30 or 60 min depending on the bathymetric layer. The net is designed to catch a wide range of species characterizing the Mediterranean demersal communities, from strictly benthic to bentho-pelagic organisms, in a large range of sizes, including juveniles (Bertrand et al., 2002).
Species Selection
The selection of the species to be analyzed was based on their contribution, in terms of abundance, to the total amount of waste potentially produced. In each study area European hake, Merluccius merluccius, deep-water rose shrimp, Parapenaeus longirostris (with the exception of Catalan Sea), and horse mackerels, Trachurus spp. were selected, while blue whiting, Micromesistius poutassou, was considered in the Portuguese area only (Table 1). UCs percentage of the aforementioned species could reach values greater than 40% in the total catch (Moranta et al., 2000; Carbonell et al., 2003; Sánchez et al., 2004, 2007; Tsagarakis et al., 2014; Milisenda et al., 2017b), and 90% of the total UCs (Despoti et al., 2020), being consequently classified as “high discard rate species” according to FAO (FAO, 2016), and largely representative of the discard produced (understood as undersize) in the investigated areas.
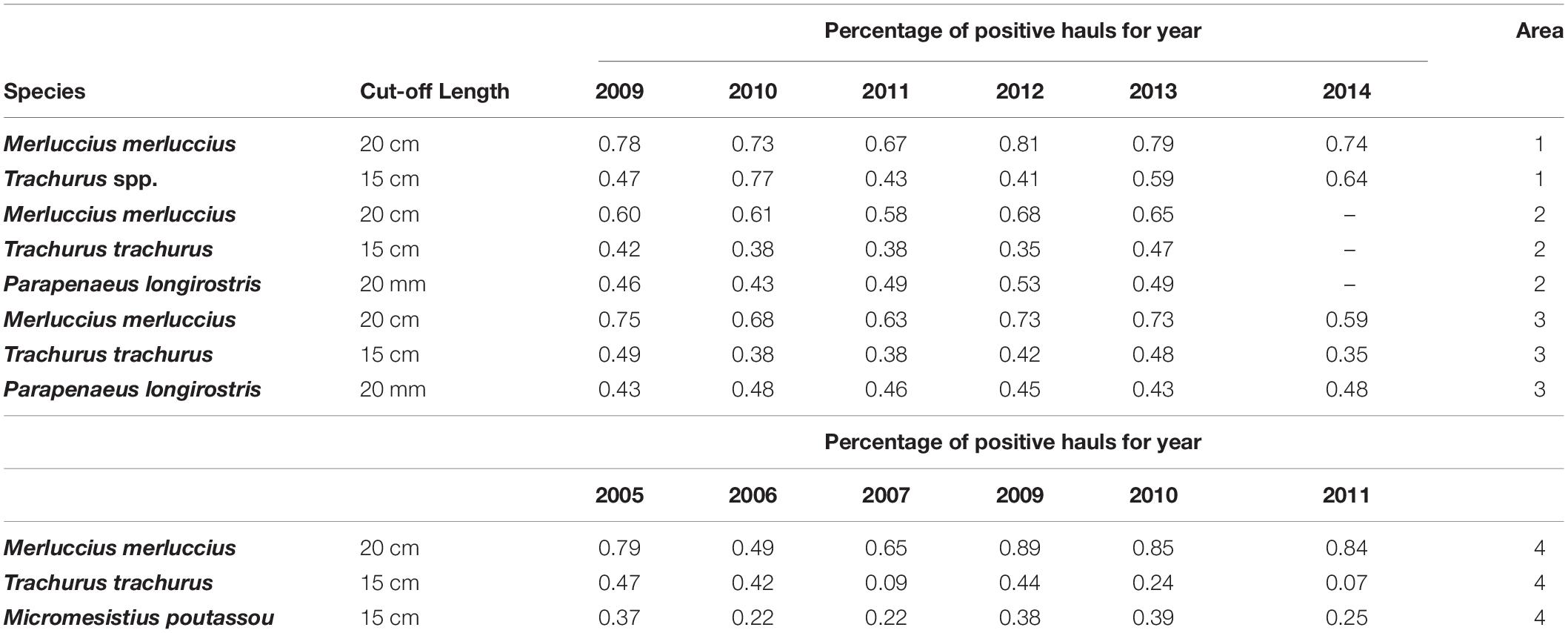
Table 1. Investigated species in each study area with the relative minimum conservation reference size (MCRS) value and the proportion of annually positive hauls (Area 1 – Catalan Sea, Area 2 – Strait of Sicily, Area 3 – Ligurian and northern Tyrrhenian Seas, and Area 4 – Continental Portuguese area).
Data Analysis
Under-Sized Catch Distribution
The first step in our analytical framework was the identification of the portion of undersized catch (USC) in the population. For this purpose, a cut-off size equal to the MCRS was used for all species (Table 1 shows the MCRS adopted by the Reg. EC No 1967/2006) except for M. poutassou for which we used the length at first maturity lacking a MCRS for the species (Silva et al., 1996). For each survey haul, the number of individuals whose length was smaller than the MCRS was standardized to the swept area (number of individuals per km2) assuming a catchability coefficient equal to 1. This resulted in a density Index of Undersized Specimens (UDI) by haul, which represented the input data for the following spatial analysis.
For each species and year, the spatial distribution of undersized specimens’ was analyzed using geostatistical methods (variogram analysis and kriging) whose fundamentals can be found in the works by Petitgas (1996) and Goovaerts (1997). The proportion of positive hauls was calculated for each year of the time series. The surveys in which the occurrence of a few positive hauls (less than 20%) precluded the structural analysis (variogram modeling) were excluded from subsequent investigations. Different exploratory plots (contour plots, scatter plots of density indices versus geographical components, normal q–q plots) were examined to check for significant deviations from stationarity and normality (the basic assumptions of linear geostatistics). A logarithmic transformation of UDIs values was applied in order to improve data normality. The successive step was to characterize the structure of spatial dependence of UDIs data through analysis of the experimental variogram, i.e., the function that summarizes the variance of all pairs of data according to distance. The experimental variograms were computed and fitted using asymptotic models such as exponential or Gaussian models (Supplementary Material 1). These models are commonly applied on fisheries data because they account for a low or medium degree of irregularity at small distances (Petitgas, 2001). Anisotropy was not analyzed because data were insufficient to characterize the possible directionality in spatial correlation. Optimal estimation of the variogram parameters (range, nugget, and sill) was obtained by minimizing the mean square error. The range provides an estimate of the maximum distance at which pairs of observations are spatially correlated. The nugget represents the amount of variance not explained by the spatial model and may arise from a combination of sampling error and small-scale variability. The sill estimates the maximum variance of the data. Once the variogram model was estimated, ordinary kriging was applied for estimating undersized specimens densities at not sampled locations and mapping their distribution (see Supplementary Material for the model parameters). Grids were constructed using 1 km × 1 km cells. Cross-validation procedures were applied to check the goodness of fit of selected variogram models and the choice of kriging parameters. In particular, the goal was to have standardized mean prediction errors near 0, and standardized root-mean-squared prediction errors near 1 (see Supplementary Material).
Density Hot Spot were identified on the annual maps of distribution using the Hot Spot Analysis tool in ArcMap. This tool identifies statistically significant spatial clusters of high values (Hot Spot) and low values (Cold Spot) applying the local Getis-Ord Gi∗ statistic (Getis and Ord, 1992; Ord and Getis, 1995). Z-score, p-value, and confidence level bin (Gi_Bin) are the final results of the analysis. Features in the ±3 bins reflect statistical significance with a 99% confidence level; features in the ±2 bins reflect a 95% confidence level; features in the ±1 bins reflect a 90% confidence level; and the clustering for features in bin 0 is not statistically significant. A 0.05 significance level was selected for spatial Hot Spot identification and the approach was applied separately in each case study and for each year of the time series in order to identify and locate spatial clusters of undersized organisms with significantly high density.
A persistent zone of occurrence of undersized organisms was defined as an area consistently occupied by the highest density of individuals below MCRS throughout the years. Hence, this zone was identified by means of GIS tools, extracting the area where the overlap of annual Hot Spot occurred over time. The overlap rate for each grid cell, defined as index of persistence (PI) (Fiorentino et al., 2003; Colloca et al., 2009), was quantified measuring the relative persistence of a cell as a potential zone where undersized organisms aggregate. This index was obtained as a ratio of the number of years that a given area was classified as a Hot Spot to the total number of years according to the formula:
where δ_ij = 1 when grid cell i is included in a Hot Spot in year j and δ_ij = 0 otherwise; n is the total number of years. The PI decreases to zero where density Hot Spots have never been observed, while it increases to 1 where density Hot Spots occur every year throughout the time series. Results were plotted in maps of persistence reporting a scale of different persistence classes.
A persistent interspecific overlap zone of undersized organisms was defined as an area consistently occupied by the highest values (greater or equal to 0.8) of persistence index of all species together. Interspecific Persistence Index (IPI) was computed as:
where K is the total number of modeled species (s) and δ is = 1 if PI is ≥0.8 in the cell i for the species s, and 0 otherwise. The index classified each cell of the study area between 0 (no interspecific spatial overlap of PI) and 1 (complete interspecific spatial overlap). Maps of IPI were constructed and visualized in ArcGIS.
Fishing Effort Distribution
The amount of fishing effort yearly deployed by the trawlers operating in each area was estimated, for the years 2012–2016, using all the available data collected by means of Automatic Identification System (AIS) and Vessel Monitoring System (VMS), the two main tracking devises applied nowadays in fisheries science. This implies that only the portions of each fleet equipped by one (or both) these tracking devices were considered in this study. While AIS data are not characterized by confidentiality issues and the identity of the fishing vessels is always associated to each AIS ping as MMSI (Maritime Mobile Service Identity) and Maritime Call sign codes, VMS data are confidential, and the access to the dataset for each area/case study was allowed by the researchers previously identified as the reference person in charge to assist the University of Rome Tor Vergata (UTV), through a one-to-one communication, during the processing of AIS/VMS data and the validation of the outcomes (Figure 2). According to the methodology (D’Andrea et al., 2020), AIS data were transferred to the reference researchers for each case study in order to allow (Figure 2): (1) The inspection of the AIS data in terms of fleet coverage; (2) The merging of AIS data with VMS data; (3) The selection of the fishing vessels to be considered for each case study. In this way, the case study reference researchers returned a list of selected vessels with their respective characteristics in terms of length-over-all (LOA, in meters) and engine power (PW in Kw).
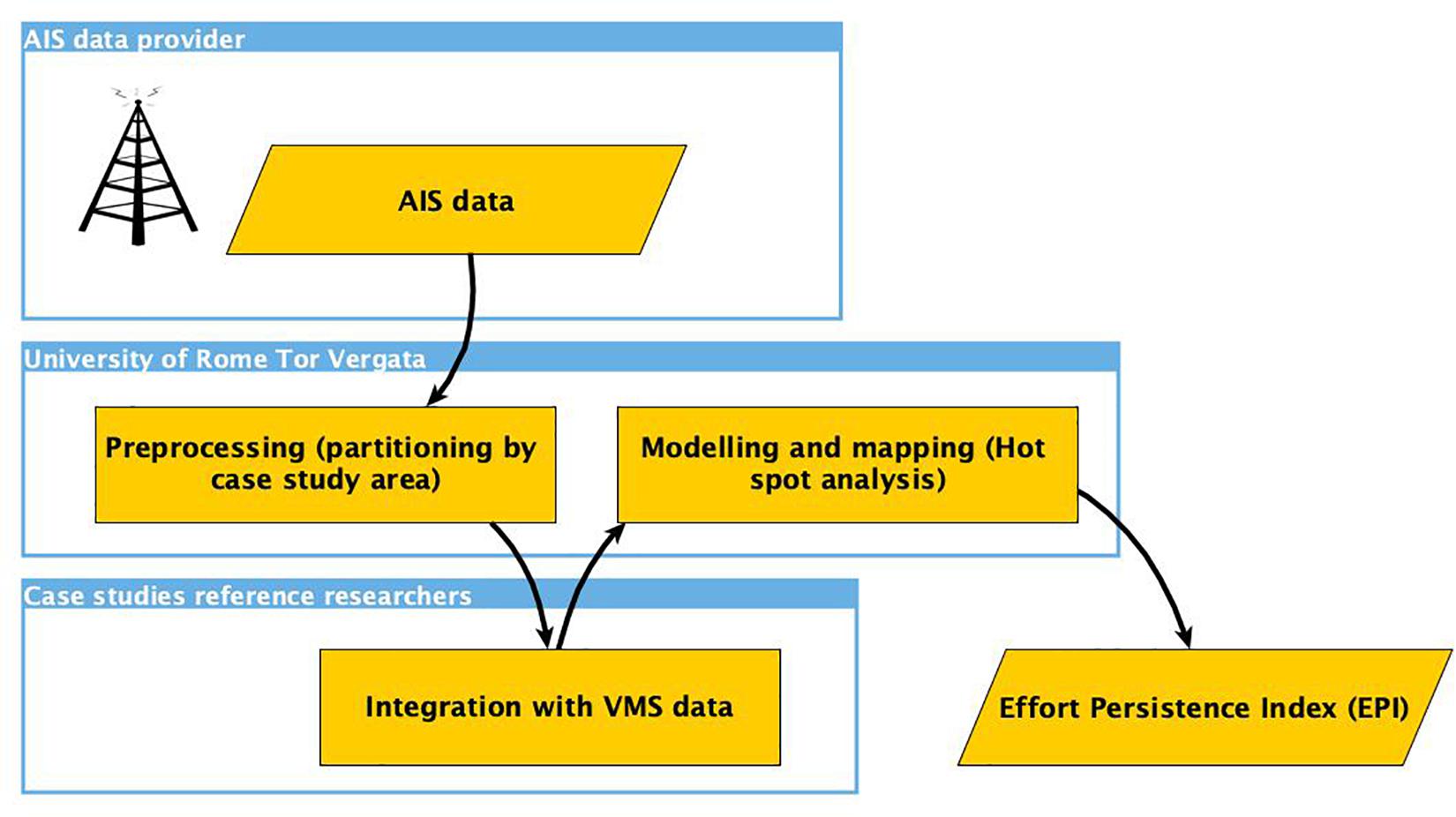
Figure 2. Representation of the processing for the integration of AIS and encrypted VMS data. The process involved UTV and the Case Studies Reference researchers through a one-to-one data flow.
AIS and VMS data were merged, at the scale of single vessels, using the procedure detailed in Russo et al. (2016). AIS/VMS data where then processed using the VMSbase platform (Russo et al., 2014a): (1) temporal frequency of pings was standardized and interpolated at 10′; (2) fishing trips by vessels were identified and (3) fishing set positions by trip (hauls) were identified using speed and depth filters. The yearly amount of effective trawling effort (ETE) was finally estimated, with respect to the cells of the same 1 km × 1 km grid described above, as the cumulated sum of the product of the number of fishing hours (FD) times the engine power (PW in Kw) by vessel. Here, FD of each vessel/year was computed using the procedure of the R package fecR (Scott et al., 2017). Basically, in case of fishing trips within which a single gear is used, each date which has a fishing activity is allocated to 1 fishing day. Fishing effort Hot Spot were then outlined on the yearly ETE maps using the Hot Spot Analysis tool in ArcMap (see above for methodological aspect).
A persistent zone of occurrence of fishing effort Hot Spot was defined as an area consistently occupied by the highest value of fishing effort throughout the years. Hence, this zone was identified by means of GIS tools, extracting the area where the overlap of annual fishing effort Hot Spot occurred over time. The overlap rate for each grid cell, defined as the Index of Effort Persistence (EPI) (Fiorentino et al., 2003; Colloca et al., 2009), was quantified measuring the relative persistence of a cell as a potential zone where trawlers work persistently. This index was obtained as a ratio of the number of years that a given area was classified as a fishing effort Hot Spot to the total number of years according to the formula:
where δ_ij = 1 when grid cell i is included in a Hot Spot in year j and δ_ij = 0 otherwise; n is the total number of years. The EPI decreases to zero where fishing effort Hot Spots have never been observed, while it increases to 1 where fishing effort Hot Spots occur every year throughout the time series. The overlap between high fishing effort persistent zone (effort persistence ≥0.8) and the persistence index of each species was calculated and expressed as square kilometers.
Results
Catalan Sea
The average percentage of positive hauls in trawl surveys carried out between 2009 and 2014 was 78% and 62% for European hake and horse mackerel, respectively (Table 1 shows the annual value of positive hauls). Hot Spot zones of undersized horse mackerel, were found in proximity of Tarragona (Figure 3A), though high values of persistence of density hot-spots can be found up to Villanova i la Geltru, where the continental shelf is narrower, between 100 and 200 m depth. A surface of 219.7 km2 was characterized by a persistence index of 1 was, while the surface increases to 891.1 km2 considering the areas with a persistence index greater between 0.8 and 1.0.
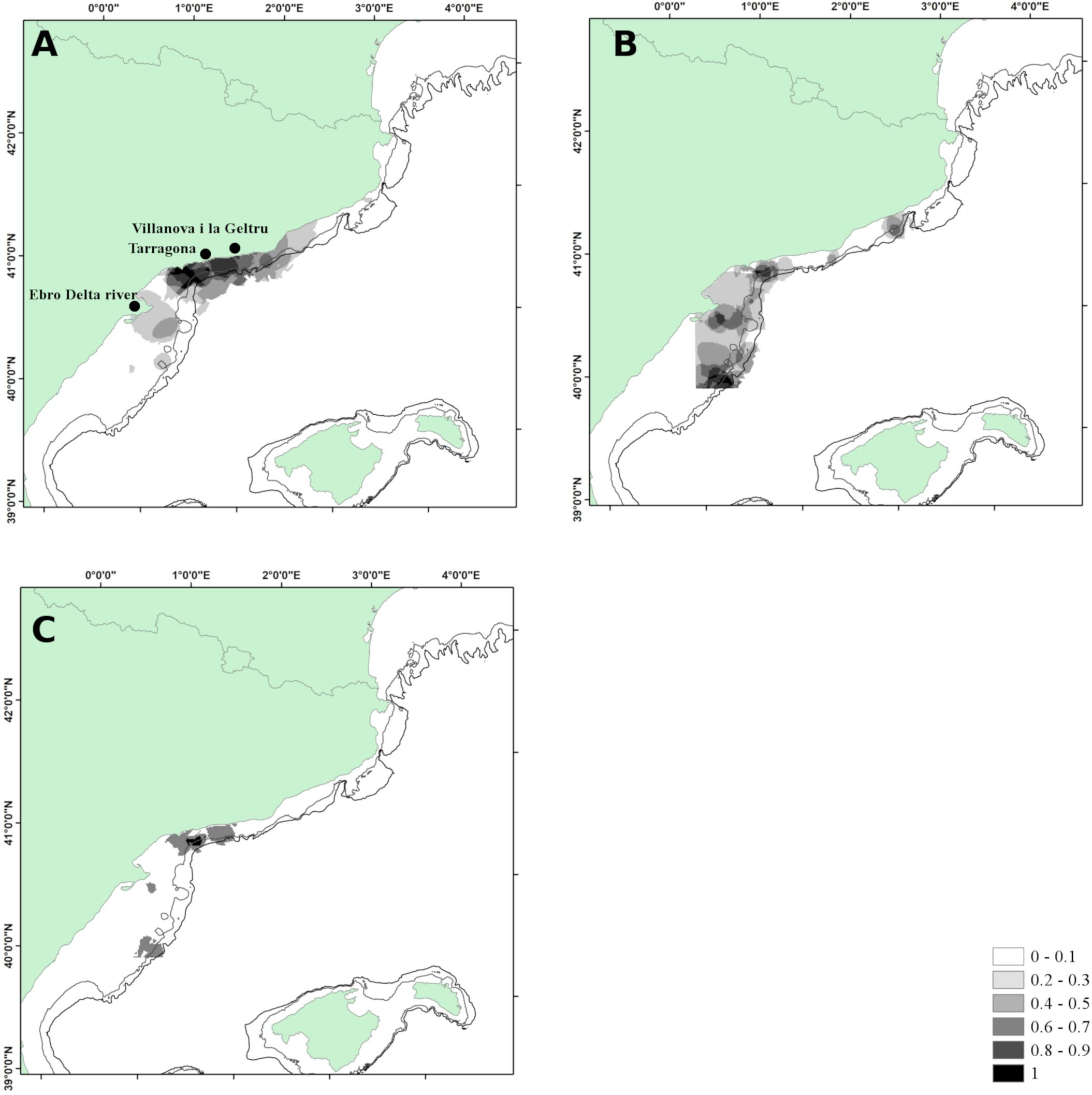
Figure 3. Area 1 – Catalan Sea. (A) Persistence index of Trachurus spp.; (B) persistence index of Merluccius merluccius; (C) interspecific persistence index (IPI). Depth contours shown are 100 and 200 m.
European hake showed a more heterogeneous distribution of undersized individuals Hot Spots than horse mackerel (Figure 3B). The area with PI greater than 0.8 covered 475.5 km2 in the south of Tarragona area, where the continental shelf reached the maximum extension. Values of PI equal to 1 covered a surface of 85.9 km2, located off Tarragona and south to the Ebro Delta.
IPI index was greater than 0.8 overlap only in the area off Tarragona, with a surface of 84.1 km2 (Figure 3C).
South of Sicily
In the Strait of Sicily the average percentage of positive hauls of European hake, horse mackerel and deep-water rose shrimp was 62, 40, and 40%, respectively (Table 1). Highly persistent density Hot Spots of small hake (PI > 0.8) were found on the eastern side of the Adventure Bank in a shelf-break area of 1575.0 km2. The surface with a PI = 1 was 293.1 km2 (Figure 4A).
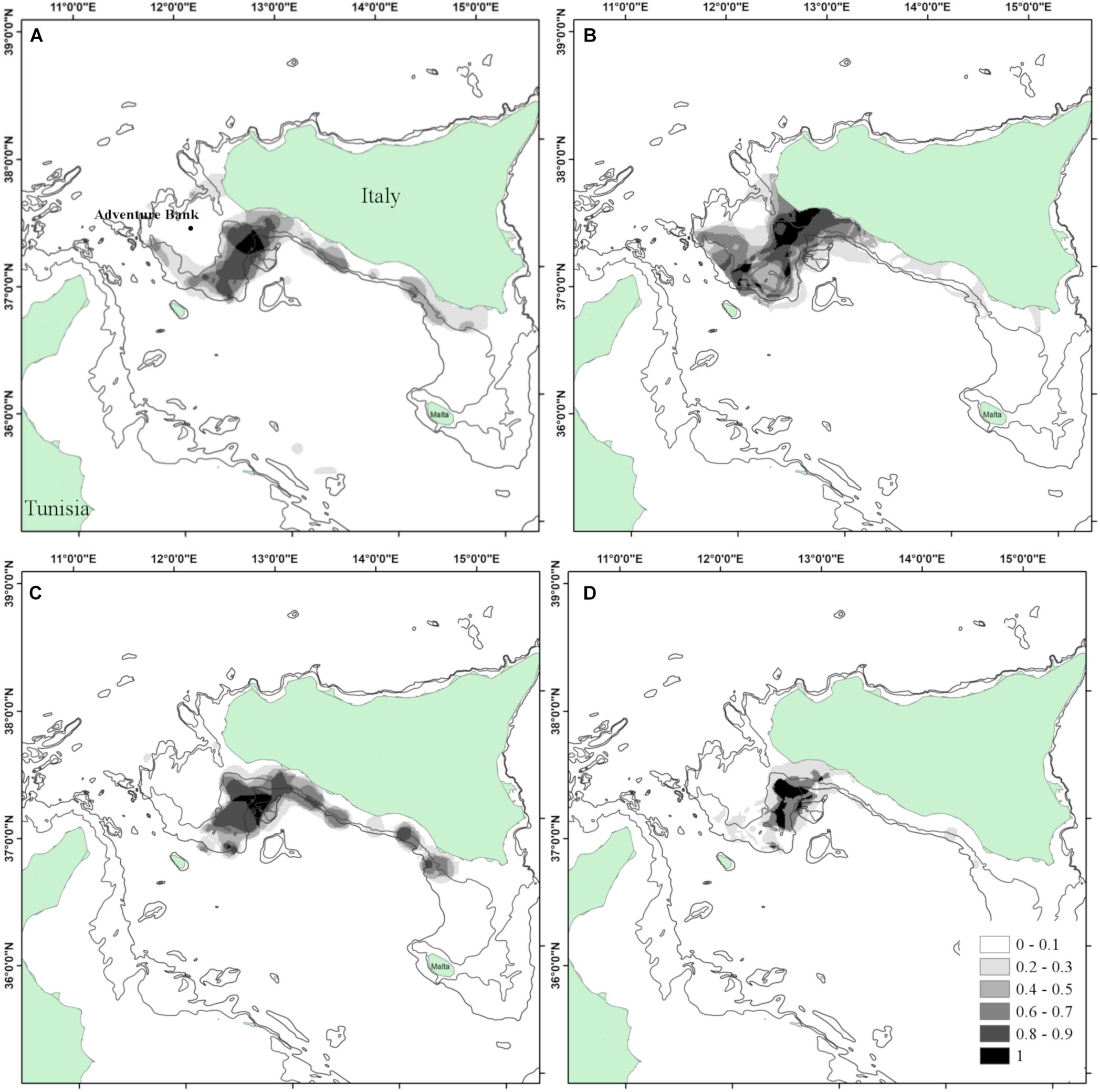
Figure 4. Area 2 – Strait of Sicily. (A) Persistence index of M. merluccius; (B) persistence index of Trachurus trachurus; (C) persistence index of Parapenaeus longirostris; (D) Interspecific persistence index (IPI). Depth contours shown are 100 and 200 m.
Small individuals of horse mackerel aggregated in a wide area (2,261 km2) over the eastern side of the Adventure Bank between 100 and 200 m depth. The core area of aggregation of juveniles (characterized by a with PI = 1) covered a surface of 741.2 km2 (Figure 4B).
As for the other species, the highest PI values of the Hot Spots of deep-water rose shrimp juveniles were found in the eastern sector of Adventure Bank on the upper part of the continental slope between 150 and 250 m depth (367.5 km2). PI values higher than 0.8 were, however, found all along the south Sicilian coasts, mostly between 100 and 250 m, covering a surface of 1977.4 km2 (Figure 4C).
Interspecific co-occurrence of highly persistent Hot Spot (IPI > 0.8) covered a wide area of 519.68 Km2 along the eastern deep shelf (100–200 m) of the Adventure Bank (Figure 4D).
Ligurian and Northern Tyrrhenian Seas
As the Ligurian and northern Tyrrhenian Seas concerns, the average percentage of positive hauls was 68% for undersized European hake, 42% for horse mackerel and 45% for deep-water rose shrimp. Persistence analysis of European hake highlighted the presence of four main zones with an index of 1, three north of Elba Island and one to the south, all located in the deeper part of the continental shelf and covering a total surface of 630.0 km2. The zone with a PI greater than 0.8 was 1,859 km2 (Figure 5A).
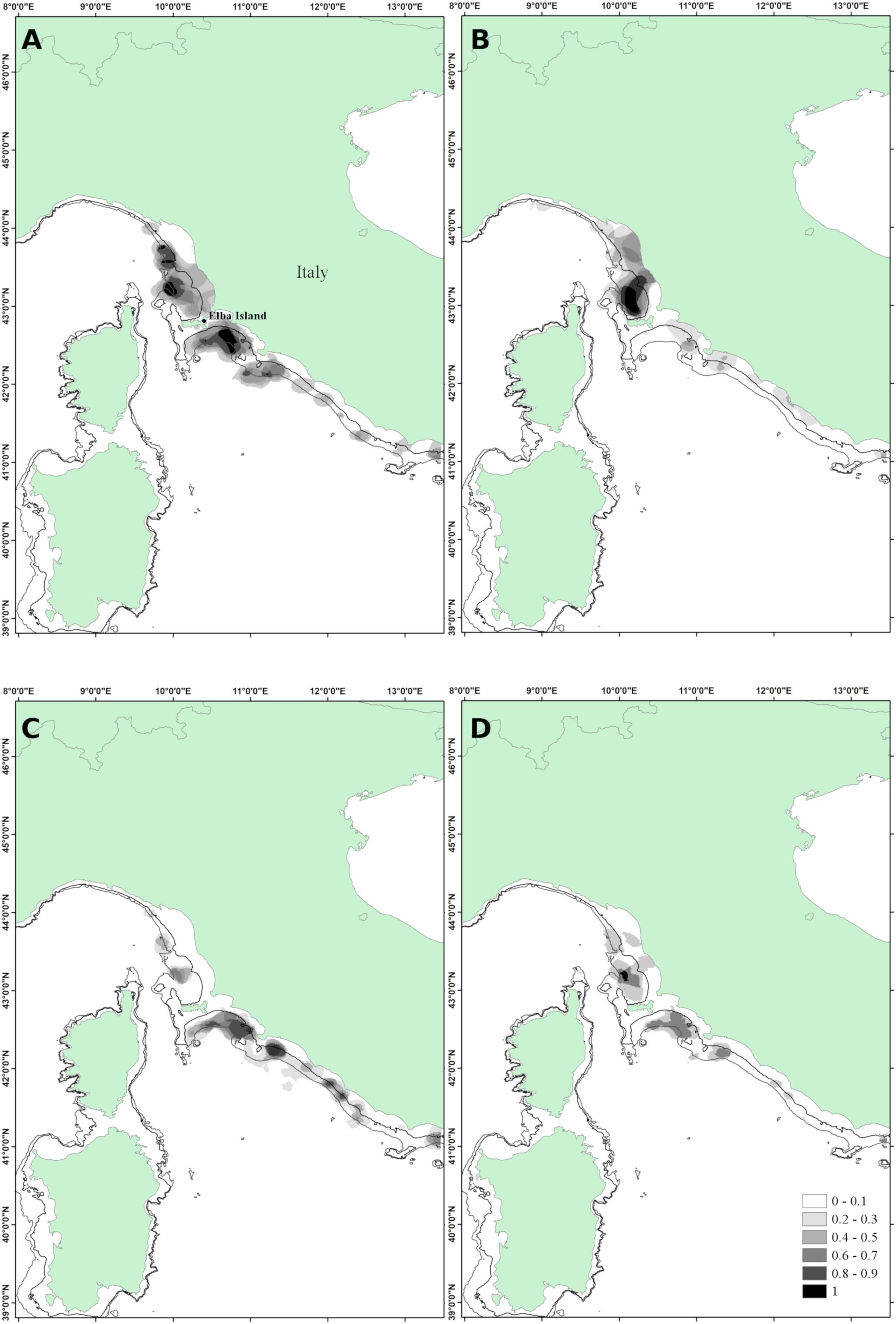
Figure 5. Area 3 – Ligurian and northern Tyrrhenian Seas. (A) Persistence index of Merluccius merluccius; (B) persistence index of T. trachurus; (C) persistence index of Parapenaeus longirostris; (D) interspecific persistence index (IPI). Depth contours shown are 100 and 200 m.
Horse mackerel showed the highest persistence index (PI > 0.8) close to the north side of Elba Island (951.5 km2) between 100 and 200 m depth. The surface featured by a PI equal to 1 was 393 km2 (Figure 5B).
Small-sized specimens of deep-water rose shrimp aggregated in several patches mostly along the deep shelf of the central-north Tyrrhenian Sea. Areas with PI > 0.8 widespread over 930 km2 (Figure 5C).
The area where the undersized organisms of the three species co-occur persistently (IPI > 0.8) covered 1453.3 km2, decreasing to 81.2 km2 of areas with IPI = 1 (Figure 5D).
Continental Portuguese Waters
The analysis of potential discard areas along the continental Portuguese coast was based on 71, 30, and 23% of positive hauls for European hake, blue whiting and horse mackerel, respectively (Table 1). The highest PI values (>0.8) for European hake were found mainly in front of Lisbon and Sines, less than 200 m depth covering an area of 3489.0 km2 (Figure 6A).
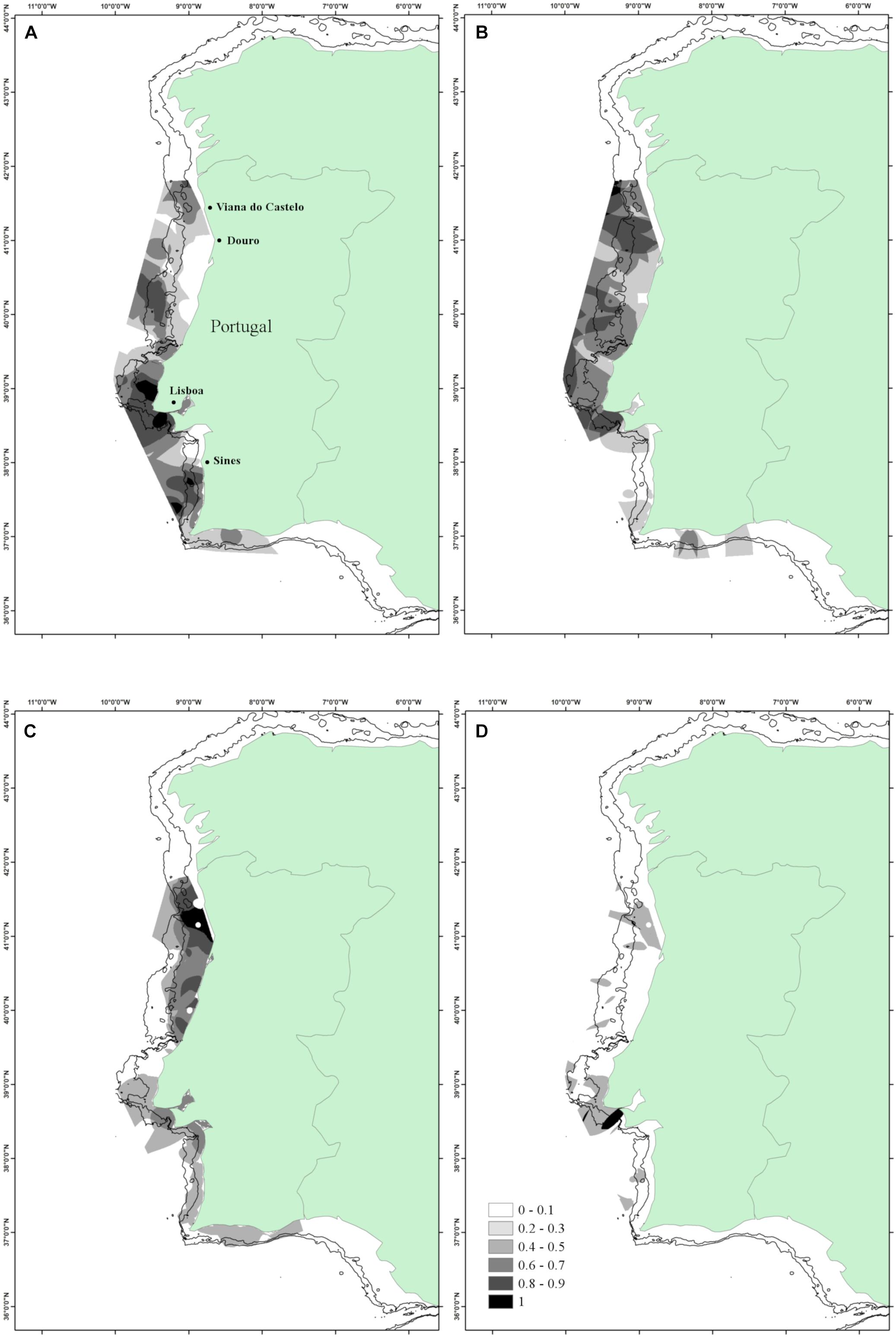
Figure 6. Area 4 – Portuguese area. (A) Persistence index of M. merluccius; (B) persistence index of Micromesistius poutassou; (C) persistence index of T. trachurus; (D) interspecific persistence index (IPI). Depth contours shown are 100 and 200 m.
Blue whiting aggregated in small patches along the north Portuguese coast, from Lisbon to Viana do Castelo, covering an area of 1933.5 km2 (Figure 6B).
Horse mackerel was the less abundant species showing the most persistent area in front of the Douro estuary (Porto) with an extension of 1513.0 km2, less than 100 m depth (Figure 6C).
Differently from the other geographical regions, we did not find IPI values greater than 0.8 for the three species considered. The main overlapping was found between European hake and blue whiting in an area of 456.1 km2 off Lisbon (Figure 6D).
Fishing Effort Analysis
Figure 7 shows the areas characterized by high values of persistence of fishing effort (EPI) in the four areas. As can be seen from this analysis, the EPIs are mainly located on the 100-m bathymetry. Table 2 shows the values expressed in Km2 of the spatial extension of greatest persistence index (PI = 0.8–1) of the different species analyzed, and the overlap expressed in Km2 of the portion of these PI areas that falls within the areas of high persistence of fishing effort (EPI = 0.8–1). Horse mackerel is the species that has the highest values of spatial overlap between the PI and EPI indices, especially in area 1, where about 226 Km2 of PI zones fall within the EPI zones (Table 2) and in area 2, where about 1,070 Km2 of PI zones fall within the EPI areas. In area 3, even though horse mackerel does not represent the species with highest overlap values, there is still around 150 Km2 of PI zones falling within the EPI zones. European hake has the highest overlap values in area 4, followed by those observed in area 2. The spatial distribution of the PI areas of deep-water rose shrimp has the highest overlap with the EPI areas in area 3, which share approximately 630 Km2 of space, followed by those present in area 2.
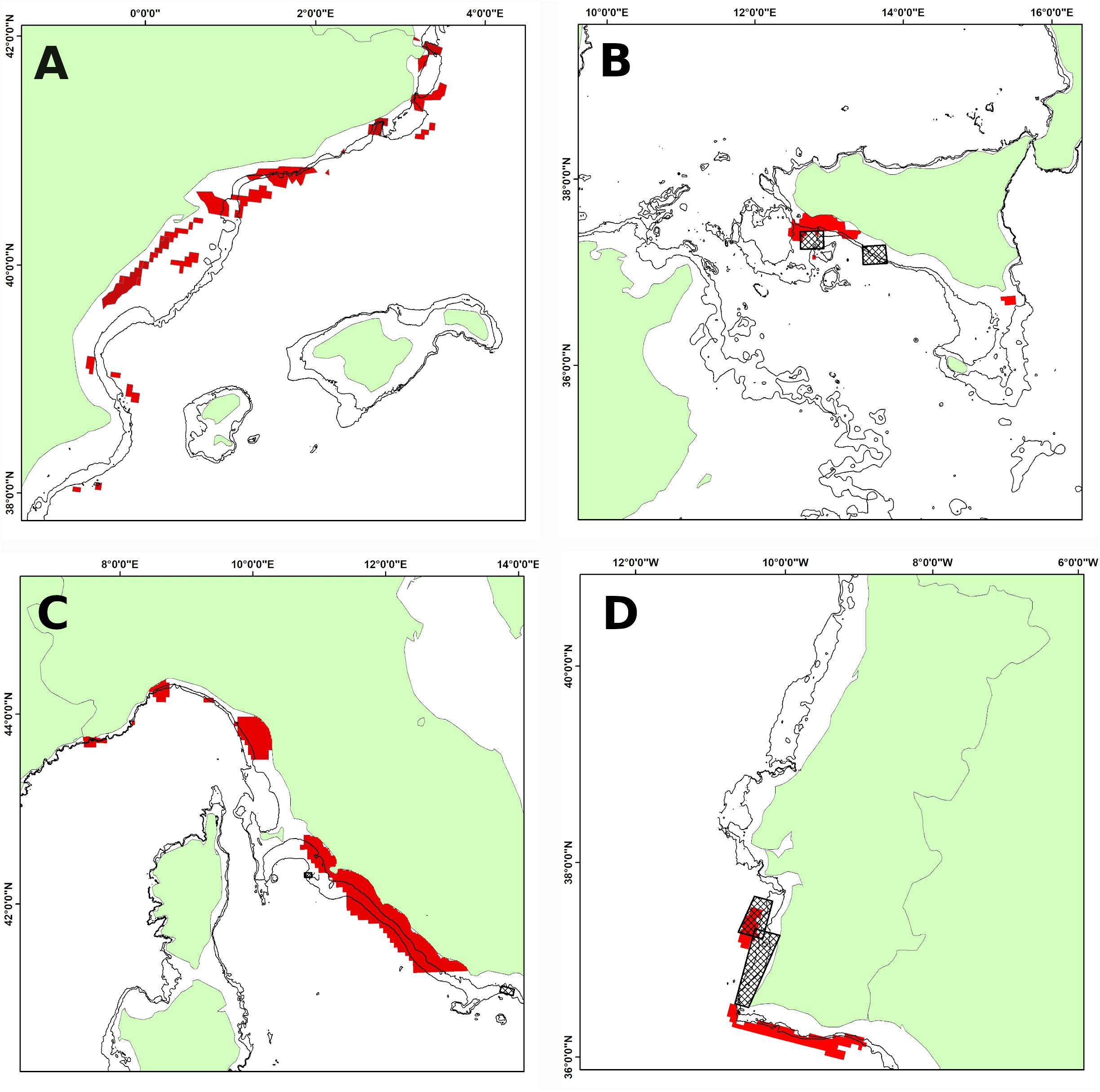
Figure 7. Zones with high persistence of fishing effort (EPI ≥ 0.8). Depth contours shown are 100 and 200 m. (A) Area 1; (B) Area 2; (C) Area 3; (D) Area 4. The square boxes indicate the fishing restricted areas.
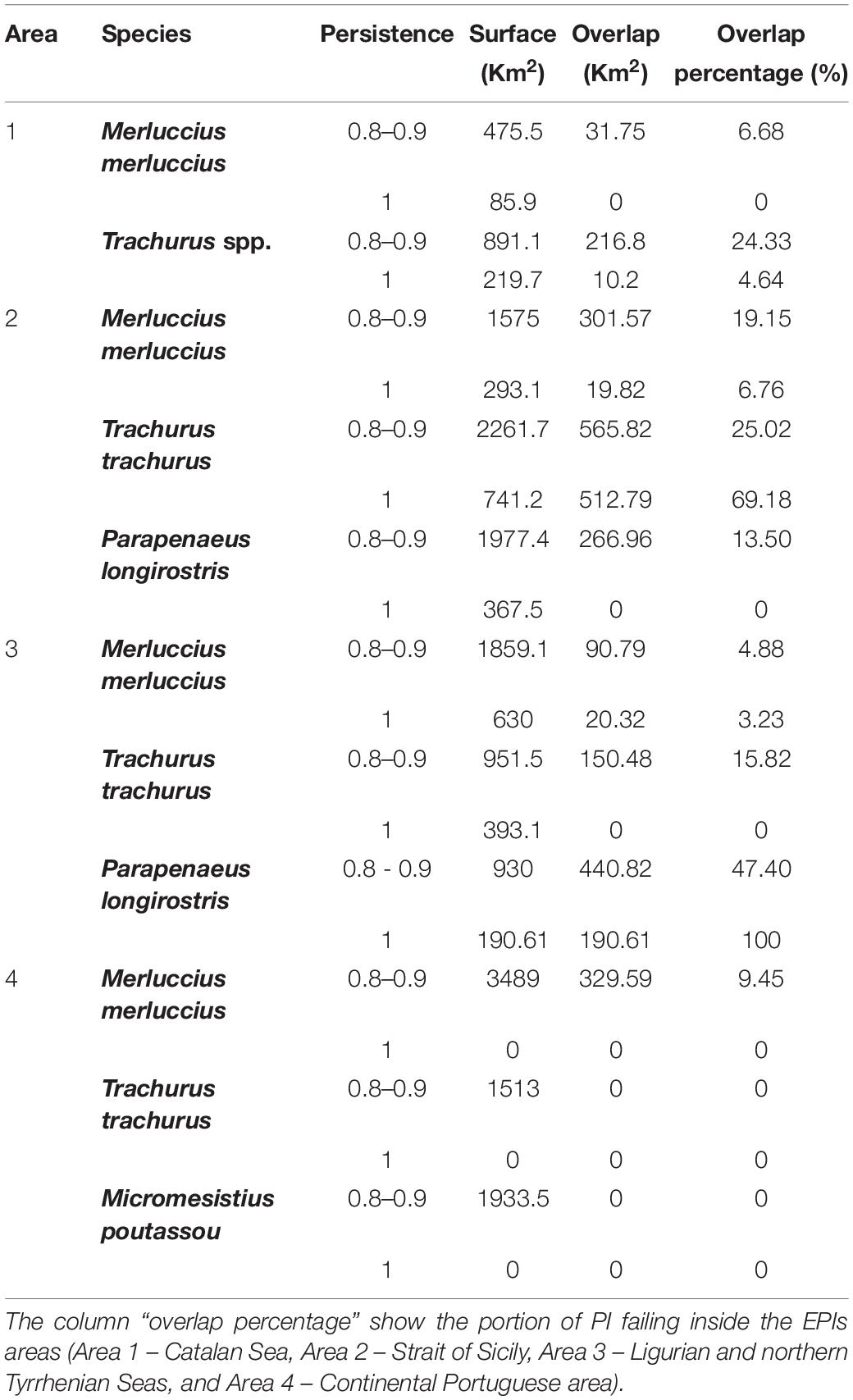
Table 2. Spatial extension of persistence index (PI) expressed as Km2 (Surface), and their overlap with persistence areas of fishing effort expressed as Km2 (Overlap).
Discussion
The present study provides a large scale assessment of the distribution and temporal persistence of areas with high density of individuals below MCRS of important commercial demersal species, based on a time series of bottom trawl surveys data. The study shows also the overlap of hot spots of undersized individuals with the areas that are persistently used by commercial trawlers. The most important result of the analysis is the first evaluation of the importance of different areas in the Mediterranean Sea and the Atlantic waters off the Continental Portuguese coast as potential source of discards for species subjected to the landing obligation in bottom trawling, due to MCRS in the Mediterranean or TAC for the Atlantic area. These results provide a relevant knowledge-base for the development of a spatial planning approach to fisheries management in order to mitigate negative effects of discarding.
Although our data comes from scientific surveys conducted in a given period of the year, previous studies have shown that the aggregation areas of juveniles of the species included in the study remain substantially unchanged in the different periods of the year (Abella et al., 2008; Colloca et al., 2009; Fortibuoni et al., 2010; Garofalo et al., 2011). The data on the spatial and temporal distribution of the trawl fleet was obtained starting from daily data, therefore the hotspots of the highly-impacted fishing areas obtained (fishing effort Hot Spots) represent a good approximation of the average annual distribution, and the overlap zones between the PI indices and EPI a good indicator of reality. Moreover, we are aware on the possible error that could come from the temporal mismatch of the survey and the VMS/AIS data. However, to minimize this error we have considered the persistence over time of hotspots distribution of the fishing fleet. The persistent areas, calculated using the VME/AIS 2012–2016 time series (5 years) should well represent the areas “persistently” most impacted by trawling in the various areas. From a biological point of view, the fishing fleet is distributed according to the distribution of the fishery resource, which in turn is distributed according to environmental characteristics that hardly change drastically over the time span used in this study, therefore also the distribution of the fishing fleet could be considered stable in the considered time. Furthermore, we are aware that the species considered in the present study do not cover all the species regulated by the MCRS or the TAC, but since they represent in some cases up to 90 percent of the under-sized fraction discarded (Despoti et al., 2020), our results may be considered representative of a generalizable trend.
We found a quite high stability of the density hot spots of undersized juveniles as already observed for the nursery areas of these species in the North Mediterranean Sea (Colloca et al., 2015). This can be due to the combined effect of connectivity between spawning and recruitment areas, and the role of habitats for juvenile growth and survival, and effects of fishing pressure (Lortie et al., 2004; Guisan and Thuiller, 2005; Ferrier and Guisan, 2006; Algar et al., 2009; Wisz et al., 2013). Based on the frequency of occurrence (Table 1), European hake, is the species showing the widest distribution of individuals below MCRS in all the four case study areas, thus making it one of the mostly discarded species by the Mediterranean and Portuguese trawl fisheries (Borges et al., 2001; FAO, 2016; Milisenda et al., 2017b). As showed in Table 2, this species is the only species whose areas of persistence (IP) fall within the EPI areas in all four areas considered. European hake recruits and juveniles are abundant on the continental shelf, and in particular at the shelf break (Colloca et al., 2003, 2015). “The shelf break is a high-energy zone as a result of the interplay of tides, storm waves, breaking internal waves and bottom turbulence produced by fronts separating shelf and slope waters (Herman, 1972; Vanney and Stanley, 1983). The high production of plankton at the shelf break makes it an important feeding ground for fish (Williams et al., 2001) and other organisms” (e.g., cephalopods, crustaceans, macro-epibenthos, etc.) (Colloca et al., 2003). Results also show persistence areas with high abundance of deep-water rose shrimp, below the MCRS on the continental shelf, in proximity of the shelf break.
The main persistence areas of small-sized horse mackerel were found in shallower waters (∼100 m) than those of the other two species. As observed in previous studies the deep shelf habitats seem play a key role for the recruitment of horse mackerel (Milisenda et al., 2017a; Rumolo et al., 2017). This pattern of distribution in areas around 100 m depth makes juveniles of horse mackerel the ones with the highest spatial extension values of overlap between IP and EPI, compared to other species (Table 2). In fact, as shown in Figure 7, the areas of temporal persistence of the commercial fishing effort are mainly distributed along the 100-m bathymetry, acting mainly on the IP areas of the T. trachurus (Areas 1, 2, and 3), and of P. longirostris (Areas 2 and 3).
According to Colloca et al. (2015), the three main oceanographic mechanisms proposed by Bakun (1996) to explain the main spatial patterns of fishery resources, i.e., (1) “nutrient enrichment” (e.g., upwelling, water mixing), (2) “concentration” (convergence, frontal formation, and water column stability) and (3) “retention within (or drift toward) appropriate habitats,” enable the establishment and co-occurrence of nursery areas of different species. An exhaustive literature exists on the spatial distribution of juveniles of main commercial species in the Mediterranean Sea (Fiorentino et al., 2003; Maynou et al., 2003; Colloca et al., 2009, 2015; Fortibuoni et al., 2010; Garofalo et al., 2010, 2011; Giannoulaki et al., 2013; Druon et al., 2015; Ligas et al., 2015). However, these studies often differed in the criteria used to identify juveniles (e.g., young of the year, first cohort of the 0 age group immature specimens) and their main purposes were not always linked to the management of local fisheries. In our study we propose to use MCRS of target commercial species to explicitly identify stable concentration areas of undersized individuals in a clear management framework aimed at minimizing the discards of trawl fisheries also considering the interspecific spatial overlap.
Our results show the presence of four important areas, one in each case study, where individuals of important commercial species below the MCRS permanently coexist. These areas are located in front of Tarragona (Catalan Sea), in the eastern part of Adventure bank (Strait of Sicily), around Elba Island (Ligurian and northern Tyrrhenian Seas) and off Lisbon (Continental Portuguese coast), mainly between 100 and 200 m depth. The eastern part of Adventure Bank is an important area where nurseries of several commercial species are located (Fiorentino et al., 2003; Fortibuoni et al., 2010; Garofalo et al., 2011; Colloca et al., 2015; Di Lorenzo et al., 2018). Here, the surface current Atlantic Ionian Stream, generates some semi-permanent mesoscale features (e.g., upwelling, eddies, and fronts) (Robinson et al., 1999; Lermusiaux and Robinson, 2001). In particular, the eastern area of Adventure Bank is characterized by large cyclonic vortex, which produces an upwelling at its center, that in turn increases primary and secondary production (Agostini and Andrew, 2002). This area is exploited by the trawlers of Sciacca and Mazara del Vallo, being the most important fishing fleet operating in the area. Our results showed the importance of the bottoms between 100 and 200 m depth as those where undersized European hake, deep-water rose shrimp and horse mackerel aggregate. These results are in line with what was found by Gristina et al. (2013) in the Strait of Sicily. Most juveniles concentrate in the circalittoral and in particular on the Coastal Terrigenous Mud biocenosis. It is worth noting that three fisheries restricted areas (FRA) for the protection of part of the nurseries of European hake and deep-water rose shrimp have been established in 2016 (FAO, 2016) and are in force since July 2019 in the northern part of the Strait of Sicily.
In the Catalan Sea, important stable aggregations of undersized specimens were found in our study south of Tarragona and off the Ebro delta. These areas are coincident with the main nurseries of hake reported by Maynou et al. (2003) and are likely related to two factors that enhance local productivity, especially in the deeper continental shelf, close to the shelf break. First, the Ebro River outflow in combination with the local upwelling generated by the south flowing coastal current along the shelf break (Liguro-Provençal-Catalan or Northern current, Millot, 1999) increases biological productivity in the area off the Ebro delta, especially along its northern margin. In contrast, the highly stratified summer water column, characterized by limited vertical movement, high depletion of surface nutrients, and with a primary production restricted to the deep chlorophyll maximum (DCM, usually located between 60 and 80 m depth), at the deepest levels of the photic zone (Estrada et al., 1993; Maynou et al., 2020). The presence of the shelf-slope front combined with the seasonal cycle is the second factor contributing to enhancing the productivity in the southern part of Catalonia, particularly at the DCM level (Estrada and Salat, 1989). Despite the concentration of biological productivity in the southern part of Catalonia, the fishing activity is based on 20 harbors of similar importance, spaced regularly along the coast. Harbors in the northernmost part, near the French border, exploit the highly productive fishing grounds near the Gulf of Lions.
The largest IPI area was found in the Tyrrhenian seas. “The circulation in the Tyrrhenian Sea is organized in a series of cyclonic (anti-clockwise) and anticyclonic (clockwise) gyres determined by the wind” (Artale et al., 1994; Gasparini et al., 2005). These vertical movements and water turbulences enriches the upper layer with nutrients, giving the Tyrrhenian Sea a relatively high concentration of nutrients within the Mediterranean basin (Nair et al., 1992; Nezlin et al., 2004). These favorable trophic conditions allow the presence in these areas of the largest hake nursery in the western Mediterranean basin (Orsi Relini et al., 2002; Abella et al., 2008; Colloca et al., 2009; Ligas et al., 2015; Cantafaro et al., 2017). In the northern part of Elba Island, the wide shelf is characterized by soft bottoms proper to trawling, favoring the development of the fleet of Viareggio, which is the largest one in the Ligurian Sea. South of Elba Island, the shelf is slightly narrower and fishing activity is concentrated on the continental slope. Important trawling fleets operate in this area (Porto Santo Stefano, Porto Ercole, and Castiglione della Pescaia), targeting European hake, red mullet (Mullus barbatus), horned octopus (Eledone cirrhosa), deep-water pink shrimp, Norway lobster and giant red shrimp (Aristaeomorpha foliacea).
The west coast of Portugal is an important persistent nursery area of the species studied in this work, mainly in the northern and central part of the coast, near the outflow of major rivers. Three factors influence overall productivity in this area: this coast is characterized by an upwelling phenomenon during the summer months providing nutrients (Peliz and Fiúza, 1999; Edwards et al., 2005) and increasing overall productivity (Loureiro et al., 2005); in autumn, there is an increase of pigment concentration in the northern part of the western coast (Peliz and Fiúza, 1999). One of the most important factors influencing nursery areas seems to be the water outflows from the two most important rivers along Portugal’s coastline (Tejo and Douro), which contribute with nutrients for the high productivity areas reported near the river mouths (Peliz and Fiúza, 1999). According to Peliz and Fiúza (1999), the persistence of a strong pigment front near the shelf break zone establishes the limits between the rich coastal area and the oligotrophic oceanic waters. Relvas et al. (2007) also indicates that the establishment of convergence zones in the shelf break is significant for the retention of larvae, a factor that appears to limit the distribution of the nursery areas. Part of the nursery area for M. merluccius, located in the south part of Portugal, is already protected by a fishing regulated area created by the Regulation Council 850/98. This area excludes trawl activities from December to the end of February. The southern part of Portugal is explored by the southern fleet, located mainly in Portimão and Olhão fishing harbors and dominated by crustacean trawlers. Several other important fishing harbors, from Sesimbra to Matosinhos, explore the central and north fishing grounds operating in the persistence areas, however, the trawl fleet has a larger mobility being able to spread their activity at national level.
The safeguard of the areas with a high presence of young organisms is a priority for the management of Southern European fisheries resources because juvenile fish are very vulnerable to a fine-mesh trawl fishery (Caddy, 1993), particularly when they assemble in nursery areas, and a decrease in fishing mortality of immature fish is fundamental for more sustainable fisheries (Beverton and Holt, 1957). The effects of nurseries protection were assessed in a simple way by estimating the fraction of recruits in the stock that benefit directly by the closure (Colloca et al., 2013). Modeling the spatial distribution of specimens below MCRS could offer fine scale management possibilities to mitigate discarding practices (Jennings et al., 2012). Our results provide a significant contribution for developing functional marine spatial plans (MSP), identifying the principal vulnerable areas, where individuals of important commercial species below the MCRS permanently coexist (Bastardie et al., 2017).
Moreover, one common reason for failure of the closure areas strategy management is the shift of fishing effort from the closure into the neighboring area still open to fishing (Agardy et al., 2011; O’Keefe et al., 2013), leading an increase of by-catch in these surrounding areas, undermining or completely nullifying any bycatch reduction achieved by the removal of effort within the closure (Hiddink et al., 2006; Powers and Abeare, 2009). Therefore, fisheries managers should not only consider the by-catch reduction within a closure area, but also the potential by-catch production outside the closure area (Wilen et al., 2002; Halpern et al., 2004; Powers and Abeare, 2009). In this direction, the combined approach used in this study, combining fisheries independent (Scientific surveys) and fisheries dependent data (distribution of commercial fishing activity), can represent a winning solution. In fact, from the scientific surveys we can obtain the distribution of organisms under MCRS in the space, we can highlight the areas most affected by fishing, and identify the areas where fishing pressure should not be concentrated (e.g., by means of spatial closure). Furthermore, we are aware that the trawl net used in scientific surveys has a smaller cod-end mesh size compared to the commercial trawl nets, therefore it is capable of capturing also the smallest fraction of the population. For this reason, we used survey data to identify the “potential” discard areas (areas with high density of small-sized individuals) and not the undersized organisms that will be caught by commercial fishing. In addition, the results of this work may contribute to an augmented compliance with the landing obligation implemented by the new CFP, and a reduction in the fishing mortality of juveniles that is currently weakening the productivity of Mediterranean demersal stocks (Colloca et al., 2013), by highlighting the “potential” discard areas in the four studied regions.
In conclusion, the strength of our results is to make available for the existing (e.g., WestMed Multi Annual Plan of the European Union) and future management plans the spatial distribution of areas where the probability to catch organisms under the legal minimum size is significantly high. In the current scenario of the new CFP, the creation of management measures to make trawl fisheries more selective is mandatory, with knowledge about the key areas where high density of individuals under MRCS are usually present critical. In this way, instead of applying strict measures covering the whole fishing grounds, an adaptive spatial management based on temporal or permanent area closure can be enforced to reduce the catch of undersized specimens and protect the habitats that are important for recruitment processes. Therefore, considering that uncertainty pervades fishery management, mainly in a multispecies fishery where it seems unfeasible to optimize the yield for all stocks at the same time, specific actions like the Hot Spots conservation of individuals below MCRS could represent an action toward sustainable fisheries.
Data Availability Statement
The datasets presented in this article are not readily available because some of the original raw data are protected by confidentiality. Requests to access the datasets should be directed to GM, Z2lhY29tby5taWxpc2VuZGFAc3pu.
Author Contributions
GM created the original outline for the manuscript. GM, GG, TR, and LD’A conducted the data analysis. FF, FC, FM, AL, CM, LB, JG, KE, and SV provided input into drafting the manuscript. All authors participated in the editing and final preparation of the manuscript.
Funding
This work was funded by the European Commission’s Horizon 2020 Research and Innovation Program under Grant Agreement No. 634495 for the project Science, Technology, and Society Initiative to minimize Unwanted Catches in European Fisheries (MINOUW). This study received Portuguese national funds from FCT-Foundation for Science and Technology through project UIDB/04326/2020.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Dr. M. Bosch-Belmar for her comments and suggestions which greatly improved the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2021.610241/full#supplementary-material
References
Abella, A., Fiorentino, F., Mannini, A., and Orsi Relini, L. (2008). Exploring relationships between recruitment of European hake (Merluccius merluccius L. 1758) and environmental factors in the Ligurian Sea and the Strait of Sicily (Central Mediterranean). J. Mar. Syst. 71, 279–293. doi: 10.1016/j.jmarsys.2007.05.010
Agardy, T., di Sciara, G. N., and Christie, P. (2011). Mind the gap: addressing the shortcomings of marine protected areas through large scale marine spatial planning. Mar. Policy 35, 226–232. doi: 10.1016/j.marpol.2010.10.006
Agostini, V. N., and Andrew, B. (2002). Ocean triads’ in the Mediterranean Sea: physical mechanisms potentially structuring reproductive habitat suitability (with example application to European anchovy, Engraulis encrasicolus). Fish. Oceanogr. 11, 129–142. doi: 10.1046/j.1365-2419.2002.00201.x
Algar, A. C., Kharouba, H. M., Young, E. R., and Kerr, J. T. (2009). Predicting the future of species diversity: macroecological theory, climate change, and direct tests of alternative forecasting methods. Ecography 32, 22–33. doi: 10.1111/j.1600-0587.2009.05832.x
Alverson, D. L., Freeberg, M. H., Murawski, S. A., and Pope, J. G. (1994). A Global Assessment of Fisheries Bycatch and Discards. FAO Fisheries Technical Paper. No. 339. Rome: FAO, 233.
Artale, V., Astraldi, M., Buffoni, G., and Gasparini, G. P. (1994). Seasonal variability of gyre-scale circulation in the northern Tyrrhenian Sea. J. Geophys. Res. 99, 14127–14137. doi: 10.1029/94JC00284
Bakun, A. (1996). Patterns in the Ocean: Ocean Processes and Marine Population Dynamics. La Paz: California Sea Grant College System, 323.
Bastardie, F., Angelini, S., Bolognini, L., Fuga, F., Manfredi, C., Martinelli, M., et al. (2017). Spatial planning for fisheries in the Northern Adriatic: working toward viable and sustainable fishing. Ecosphere 8:e01696. doi: 10.1002/ecs2.1696
Batsleer, J., Poos, J. J., Marchal, P., Vermard, Y., and Rijnsdrop, A. D. (2013). Mixed fisheries management: protecting the weakest link. Mar. Ecol. Prog. Ser. 479, 177–190. doi: 10.3354/meps10203
Bellido, J. M., Paradinas, I., Vilela, R., Bas, G., and Pennino, M. G. (2019). “A marine spatial planning approach to minimize discards: challenges and opportunities of the landing obligation in European waters,” in The European Landing Obligation, eds S. Uhlmann, C. Ulrich, and S. Kennelly (Cham: Springer).
Bellido, J. M., Santos, M. B., Pennino, M. G., Valeiras, X., and Pierce, G. J. (2011). Fishery discards and bycatch: Solutions for an ecosystem approach to fisheries management? Hydrobiologia 670, 317–333. doi: 10.1007/s10750-011-0721-5
Bertrand, J. A., Gil De Sola, L., Papaconstantinou, C., Relini, G., and Souplet, A. (2002). The general specifications of the MEDITS surveys. Sci. Mar. 66, 9–17. doi: 10.3989/scimar.2002.66s2169
Beverton, R. J. H., and Holt, S. J. (1957). On the Dynamics of Exploited Fish Populations. Fish and Fisheries Series, Vol. 11. London: Chapman and Hall.
Borges, T. C., Erzini, K., Bentes, L., Costa, M. E., Gonçalves, J. M. S., Lino, P. G., et al. (2001). By-catch and discarding practices in five Algarve (southern Portugal) métiers. J. Appl. Ichthyol. 17, 104–114. doi: 10.1111/j.1439-0426.2001.00283.x
Caddy, J. F. (1993). Some future perspectives for assessment and management of Mediterranean fisheries. Sci. Mar. 57, 121–130.
Cantafaro, A., Ardizzone, G., Enea, M., Ligas, A., and Colloca, F. (2017). Assessing the importance of nursery areas of European hake (Merluccius merluccius) using a body condition index. Ecol. Indic. 81, 383–389. doi: 10.1016/j.ecolind.2017.06.012
Carbonell, A., Pomar, G., Gaza, M., Torres, A., García, M., Pereda, P., et al. (2003). The deep-water red shrimp fishery in the Spanish Mediterranean Sea. J. Northwest Atl. Fish. Sci. 31:353.
Catchpole, T. L., Frid, C. L. J., and Gray, T. S. (2005). Discarding in the English north-east coast Nephrops norvegicus fishery: the role of social and environmental factors. Fish. Res. 72, 45–54. doi: 10.1016/j.fishres.2004.10.012
Colloca, F., Bartolino, V., Lasinio, G. J., Sartor, P., and Ardizzone, G. (2009). Identifying fish nurseries using density and persistence measures. Mar. Ecol. Prog. Ser. 381, 287–296. doi: 10.3354/meps07942
Colloca, F., Cardinale, M., Belluscio, A., and Ardizzone, G. (2003). Pattern of distribution and diversity of demersal assemblages in the central Mediterranean sea. Estuar. Coast. Shelf Sci. 56, 469–480. doi: 10.1016/S0272-7714(02)00196-8
Colloca, F., Cardinale, M., Maynou, F., Giannoulaki, M., Scarcella, G., Jenko, K., et al. (2013). Rebuilding Mediterranean fisheries: a new paradigm for ecological sustainability. Fish Fish. 14, 89–109. doi: 10.1111/j.1467-2979.2011.00453.x
Colloca, F., Garofalo, G., Bitetto, I., Facchini, M. T., Grati, F., Martiradonna, A., et al. (2015). The seascape of Demersal fish nursery areas in the North Mediterranean sea, a first step towards the implementation of spatial planning for trawl fisheries. PLoS One 10:e0119590. doi: 10.1371/journal.pone.0119590
Condie, H. M., Catchpole, T. L., and Grant, A. (2014). The short-term impacts of implementing catch quotas and a discard ban on English North Sea otter trawlers. ICES J. Mar. Sci. 71, 1266–1276. doi: 10.1093/icesjms/fst187
Damalas, D., Ligas, A., Tsagarakis, K., Vassilopoulou, V., Stergiou, K., Kallianiotis, A., et al. (2018). The “discard problem” in Mediterranean fisheries, in the face of the European Union landing obligation: the case of bottom trawl fishery and implications for management. Mediterr. Mar. Sci. 19, 459–476. doi: 10.12681/mms.14195
D’Andrea, L., Campos, A., Erzin, K., Fonseca, P., Franceschini, S., Kavadas, S., et al. (2020). The MINOUWApp: a web-based tool in support of by-catch and discards management. Environ. Monit. Assess. 192, 754. doi: 10.1007/s10661-020-08704-5
Davies, R. W. D., Cripps, S. J., Nickson, A., and Porter, G. (2009). Defining and estimating global marine fisheries bycatch. Mar. Policy 33, 661–672. doi: 10.1016/j.marpol.2009.01.003
De Juan, S., and Demestre, M. (2012). A Trawl Disturbance Indicator to quantify large scale fishing impact on benthic ecosystems. Ecol. Indic. 18, 183–190. doi: 10.1016/j.ecolind.2011.11.020
Despoti, S., Milisenda, G., Ligas, A., Bentes, L., Maynou, F., Vitale, S., et al. (2020). Marine spatial closures as a supplementary tool to reduce discards in bottom trawl fisheries: examples from southern European waters. Fish. Res. 232, 105714. doi: 10.1016/j.fishres.2020.105714
Di Lorenzo, M., Sinerchia, M., and Colloca, F. (2018). The North sector of the Strait of Sicily: a priority area for conservation in the Mediterranean Sea. Hydrobiologia 821, 235–253. doi: 10.1007/s10750-017-3389-7
Druon, J.-N., Fiorentino, F., Murenu, M., Knittweis, L., Colloca, F., Osio, C., et al. (2015). Modelling of European hake nurseries in the Mediterranean Sea: an ecological niche approach. Prog. Oceanogr. 130, 188–204. doi: 10.1016/j.pocean.2014.11.005
Edwards, V., Icely, J., Newton, A., and Webster, R. (2005). The yield of chlorophyll from nitrogen: a comparison between the shallow Ria Formosa lagoon and the deep oceanic conditions at Sagres along the southern coast of Portugal. Estuar. Coast. Shelf Sci. 62, 391–403. doi: 10.1016/j.ecss.2004.09.004
Estrada, M., Marrase, C., Latasa, M., Berdalet, E., Delgado, M., and Riera, T. (1993). Variability of deep chlorophyll maximum characteristics in the northwestern Mediterranean. Mar. Ecol. Prog. Ser. 92, 289–300. doi: 10.3354/meps092289
Estrada, M., and Salat, J. (1989). Phytoplankton assemblages of dep and surface water layers in a Mediterranean frontal zone. Sci. Mar. 53, 203–214.
FAO (2019). Monitoring Discards in Mediterranean and Black Sea Fisheries: Methodology for Data Collection. FAO Fisheries and Aquaculture Technical Paper No. 639. Rome: FAO.
Ferrier, S., and Guisan, A. (2006). Spatial modelling of biodiversity at the community level. J. Appl. Ecol. 43, 393–404. doi: 10.1111/j.1365-2664.2006.01149.x
Fiorentino, F., Garofalo, G., De Santi, A., Bono, G., Giusto, G. B., and Norrito, G. (2003). Spatio-temporal distribution of recruits (0 group) of Merluccius merluccius and Phycis blennoides (Pisces, Gadiformes) in the Strait of Sicily (Central Mediterranean). Hydrobiologia 503, 223–236. doi: 10.1023/B:HYDR.0000008485.85468.ef
Fortibuoni, T., Bahri, T., Camilleri, M., Garofalo, G., Gristina, M., and Fiorentino, F. (2010). Nursery and spawning areas of deep-water rose shrimp, Parapenaeus longirostris (Decapoda: Penaeidae), in the Strait of Sicily (Central Mediterranean Sea). J. Crustac. Biol. 30, 167–174. doi: 10.1651/09-3167.1
Garofalo, G., Ceriola, L., Gristina, M., Fiorentino, F., and Pace, R. (2010). Nurseries, spawning grounds and recruitment of Octopus vulgaris in the Strait of Sicily, central Mediterranean Sea. ICES J. Mar. Sci. 67, 1363–1371. doi: 10.1093/icesjms/fsq101
Garofalo, G., Fortibuoni, T., Gristina, M., Sinopoli, M., and Fiorentino, F. (2011). Persistence and co-occurrence of demersal nurseries in the Strait of Sicily (central Mediterranean): implications for fishery management. J. Sea Res. 66, 29–38. doi: 10.1016/j.seares.2011.04.008
Gasparini, G. P., Ortona, A., Budillon, G., Astraldi, M., and Sansone, E. (2005). The effect of the Eastern Mediterranean Transient on the hydrographic characteristics in the Strait of Sicily and in the Tyrrhenian Sea. Deep Sea Res. I Oceanogr. Res. Pap. 52, 915–935. doi: 10.1016/j.dsr.2005.01.001
Getis, A., and Ord, J. K. (1992). The analysis of spatial association by use of distance statistics. Geogr. Anal. 24, 189–206. doi: 10.1111/j.1538-4632.1992.tb00261.x
Giannoulaki, M., Belluscio, A., Colloca, F., Fraschetti, S., Scardi, M., Smith, C., et al. (eds) (2013). Mediterranean Sensitive Habitats. DG MARE Specific Contract SI2.600741, Final Report. (Heraklion: Hellenic Centre for Marine Research), 557.
Goovaerts, P. (1997). Geostatistics for Natural Resources Evaluation. (New York, NY: Oxford University Press), 483.
Graham, N., Döring, R., Curtis, H., Moreau, K., van Helmond, E., Schon, P. J., et al. (2008). Scientific, Technical and Economic Committee for Fisheries. Report of the SGMOS-08-01 Working group on the Reduction of Discarding Practices. Luxembourg: Publications Office of the European Union.
Gristina, M., Fiorentino, F., Gancitano, V., Massi, D., Mirto, S., and Garofalo, G. (2013). The role of juveniles in structuring demersal assemblages in trawled fishing grounds. Estuar. Coast. Shelf Sci. 133, 78–87. doi: 10.1016/j.ecss.2013.08.014
Guisan, A., and Thuiller, W. (2005). Predicting species distribution: offering more than simple habitat models. Ecol. Lett. 8, 993–1009. doi: 10.1111/j.1461-0248.2005.00792.x
Halpern, B. S., Gaines, S. D., and Warner, R. R. (2004). Confounding effects of the export of production and the displacement of fishing effort from marine reserves. Ecol. Appl. 14, 1248–1256. doi: 10.1890/03-5136
Herman, Y. (1972). “Quaternary eastern Mediterranean sediments: micropaleontology and climatic records,” in The Mediterranean Sea: A Natural Sedimentation Laboratory, ed. D. J. Stanley (Stroudsburg, PA: Dowden, Butchinson and Ross).
Hiddink, J. G., Hutton, T., Jennings, S., and Kaiser, M. J. (2006). Predicting the effects of area closures and fishing effort restrictions on the production, biomass, and species richness of benthic invertebrate communities. ICES J. Mar. Sci. 63, 822–830. doi: 10.1016/j.icesjms.2006.02.006
ICES (2017). Manual of the IBTS North Eastern Atlantic Surveys. Series of ICES Survey Protocols SISP 15. (Copenhagen: ICES). doi: 10.17895/ices.pub.3519
Jennings, S., Lee, J., and Hiddink, J. G. (2012). Assessing fishery footprints and the trade-offs between landings value, habitat sensitivity, and fishing impacts to inform marine spatial planning and an ecosystem approach. ICES J. Mar. Sci. 69, 1053–1063. doi: 10.1093/icesjms/fss050
Karp, W. A., Breen, M., Borges, L., Fitzpatrick, M., Kennelly, S. J., Kolding, J., et al. (2019). “Strategies used throughout the world to manage fisheries discards – lessons for implementation of the EU landing obligation,” in The European Landing Obligation, eds S. Uhlmann, C. Ulrich, and S. Kennelly (Cham: Springer).
Kelleher, K. (2005). Discards in the World’s Marine Fisheries. An Update. FAO Technical Paper 470. (Rome: FAO), 152. doi: 10.1136/ebmed-2011-0012
Lermusiaux, P. F. J., and Robinson, A. R. (2001). Features of dominant mesoscale variability, circulation patterns and dynamics in the strait of sicily. Deep sea Res. I Oceanogr. Res. Pap. 48, 1953–1997. doi: 10.1016/S0967-0637(00)00114-X
Ligas, A., Colloca, F., Lundy, M. G., Mannini, A., Sartor, P., Sbrana, M., et al. (2015). Modeling the growth of recruits of European hake (Merluccius merluccius) in the northwestern Mediterranean Sea with generalized additive models. Fish. Bull. 113, 69–81. doi: 10.7755/FB.113.1.7
Lleonart, J., and Maynou, F. (2003). Fish stock assessments in the Mediterranean: state of the art. Sci. Mar. 67, 37–49. doi: 10.3989/scimar.2003.67s137
Lortie, C. J., Brooker, R. W., Choler, P., Kikvidze, Z., Michalet, R., Pugnaire, F. I., et al. (2004). Rethinking plant community theory. Oikos 107, 433–438. doi: 10.1111/j.0030-1299.2004.13250.x
Loureiro, S., Newton, A., and Icely, J. D. (2005). Microplankton composition, production and upwelling dynamics in Sagres (SW Portugal) during the summer of 2001. Sci. Mar. 69, 323–341. doi: 10.3989/scimar.2005.69n3323
Maiorano, L., Bartolino, V., Colloca, F., Abella, A., Belluscio, A., Carpentieri, P., et al. (2009). Systematic conservation planning in the Mediterranean: a flexible tool for the identification of no-take marine protected areas. ICES J. Mar. Sci. 66, 137–146. doi: 10.1093/icesjms/fsn148
Maynou, F., Lleonart, J., and Cartes, J. E. (2003). Seasonal and spatial variability of hake (Merluccius merluccius L.) recruitment in the NW Mediterranean. Fish. Res. 60, 65–78. doi: 10.1016/S0165-7836(02)00062-0
Maynou, F., Sabatés, A., and Raya, V. (2020). Changes in the spawning habitat of two small pelagic fish in the Northwestern Mediterranean. Fish. Oceanogr. 29, 201–213. doi: 10.1111/fog.12464
Milisenda, G., Garofalo, G., Fezzani, S., Rjeibi, O., Jarboui, O., Chemmam, B., et al. (2017a). Erratum to: Biomass HotSpot distribution model and spatial interaction of two exploited species of horse mackerel in the south-central Mediterranean Sea. Hydrobiologia 821:151. doi: 10.1007/s10750-017-3378-x
Milisenda, G., Vitale, S., Massi, D., Enea, M., Gancitano, V., Giusto, G. B., et al. (2017b). Spatio-temporal composition of discard associated with the deep water rose shrimp fisheries (Parapenaeus longirostris. Lucas 1846) in the south-central Mediterranean Sea. Mediterr. Mar. Sci. 18, 53–63. doi: 10.12681/mms.1787
Millot, C. (1999). Circulation in the Western Mediterranean Sea. J. Mar. Syst. 20, 423–442. doi: 10.1016/S0924-7963(98)00078-5
Moranta, J., Massutí, E., and Morales-Nin, B. (2000). Fish catch composition of the deep-sea decapod crustacean fisheries in the Balearic Islands (western Mediterranean). Fish. Res. 45, 253–264. doi: 10.1016/S0165-7836(99)00119-8
Nair, R., Cattini, E., Rossi, G., and Gasparini, G. P. (1992). Upwelling in the Northern Tyrrhenian Sea: Some Physical and Chemical Characteristics. Available online at: http://ciesm.org/online/archives/abstracts/pdf/33/PG_0202.pdf (accessed September 24, 2020).
Nezlin, N. P., Lacroix, G., Kostianoy, A. G., and Djenidi, S. (2004). Remotely sensed seasonal dynamics of phytoplankton in the Ligurian Sea in 1997–1999. J. Geophys. Res. 109:C07013. doi: 10.1029/2000JC000628
O’Keefe, C. E., Cadrin, S. X., and Stokesbury, K. D. E. (2013). Evaluating effectiveness of time/area closures, quotas/caps and fleet communications to reduce fisheries bycatch. ICES J. Mar. Sci. 71, 1286–1297. doi: 10.1093/icesjms/fst063
Ord, J. K., and Getis, A. (1995). Local spatial autocorrelation statistics: distributional issues and an application. Geogr. Anal. 27, 286–306. doi: 10.1111/j.1538-4632.1995.tb00912.x
Orsi Relini, L., Papaconstantinou, C., Jukic-Peladic, S., Souplet, A., Gil de Sola Simarro, L., Piccinetti, C., et al. (2002). Distribution of the Mediterranean hake populations (Merluccius merluccius smiridus Rafinesque, 1810) (Osteichthyes : Gadiformes) based on six years monitoring by trawl-surveys: some implications for management. Sci. Mar. 66, 21–38. doi: 10.3989/scimar.2002.66s221
Paradinas, I., Martín, M., Pennino, M. G., López-Quílez, A., Conesa, D., Barreda, D., et al. (2016). Identifying the best fishing-suitable areas under the new European discard ban. ICES J. Mar. Sci. 73, 2479–2487. doi: 10.1093/icesjms/fsw114
Peliz, A. J., and Fiúza, A. F. G. (1999). Temporal and spatial variability of CZCS-derived phytoplankton pigment concentrations off the western Iberian Peninsula. Int. J. Remote Sens. 20, 1363–1403. doi: 10.1080/014311699212786
Petitgas, P. (2001). Geostatistics in fisheries survey design and stock assessment: models, variances and applications. Fish Fish. 2, 231–249. doi: 10.1046/j.1467-2960.2001.00047.x
Petitgas, P. (1996). “Geostatistics and their applications to fisheries survey data BT,” in Computers in Fisheries Research,” eds B. A. Megrey and E. Moksness (Dordrecht: Springer), 113–142 doi: 10.1007/978-94-015-8598-9_5
Pipitone, C., Badalamenti, F., Vega Fernández, T., and D’Anna, G. (2014). Spatial management of fisheries in the mediterranean sea: problematic issues and a few success stories. Adv. Mar. Biol. 69, 371–402. doi: 10.1016/b978-0-12-800214-8.00010-4
Powers, J. E., and Abeare, S. M. (2009). Fishing effort redistribution in response to area closures. Fish. Res. 99, 216–225. doi: 10.1016/j.fishres.2009.06.011
Relvas, P., Barton, E. D., Dubert, J., Oliveira, P. B., Peliz, Á., da Silva, J. C. B., et al. (2007). Physical oceanography of the western Iberia ecosystem: latest views and challenges. Prog. Oceanogr. 74, 149–173. doi: 10.1016/j.pocean.2007.04.021
Robinson, A. R., Sellschopp, J., Warn-Varnas, A., Leslie, W. G., Lozano, C. J., Haley, P. J., et al. (1999). The Atlantic Ionian stream. J. Mar. Syst. 20, 129–156. doi: 10.1016/S0924-7963(98)00079-7
Rumolo, P., Basilone, G., Fanelli, E., Barra, M., Calabrò, M., Genovese, S., et al. (2017). Linking spatial distribution and feeding behavior of Atlantic horse mackerel (Trachurus trachurus) in the Strait of Sicily (Central Mediterranean Sea). J. Sea Res. 121, 47–58. doi: 10.1016/j.seares.2017.01.002
Russo, T., Carpentieri, P., D’Andrea, L., De Angelis, P., Fiorentino, F., Franceschini, S., et al. (2019). Trends in effort and yield of trawl fisheries: a case study from the Mediterranean Sea. Front. Mar. Sci. 6:153. doi: 10.3389/fmars.2019.00153
Russo, T., Parisi, A., and Cataudella, S. (2013). Spatial indicators of fishing pressure: preliminary analyses and possible developments. Ecol. Indic. 26, 141–153. doi: 10.1016/j.ecolind.2012.11.002
Russo, T., D’Andrea, L., Parisi, A., and Cataudella, S. (2014a). VMSbase: an R-package for VMS and logbook data management and analysis in fisheries ecology. PLoS One 9:e100195. doi: 10.1371/journal.pone.0100195
Russo, T., D’Andrea, L., Parisi, A., Martinelli, M., Belardinelli, A., Boccoli, F., et al. (2016). Assessing the fishing footprint using data integrated from different tracking devices: issues and opportunities. Ecol. Indic. 69, 818–827. doi: 10.1016/j.ecolind.2016.04.043
Russo, T., Parisi, A., Garofalo, G., Gristina, M., Cataudella, S., and Fiorentino, F. (2014b). SMART: a spatially explicit bio-economic model for assessing and managing demersal fisheries, with an application to Italian trawlers in the Strait of Sicily. PLoS One 9:e86222. doi: 10.1371/journal.pone.0086222
Sánchez, P., Demestre, M., and Martín, P. (2004). Characterisation of the discards generated by bottom trawling in the northwestern Mediterranean. Fish. Res. 67, 71–80. doi: 10.1016/j.fishres.2003.08.004
Sánchez, P., Sartor, P., Recasens, L., Ligas, A., Martin, J., and De Ranieri, S. (2007). Trawl catch composition during different fishing intensity periods in two Mediterranean demersal fishing grounds. Sci. Mar. 71, 765–773. doi: 10.3989/scimar.2007.71n4765
Scott, F., Prista, N., and Reilly, T. (2017). fecR: Fishing Effort Calculator in R. R package version 0.0.1., Available online at: https://CRAN.R-project.org/package=fecR (accessed September 24, 2020).
Silva, A., Pestana, G., Dias, C., and Godinho, S. (1996). Preliminary Results on the Distribution and Spawning of Blue Whiting, Micromesistius poutassou, off the Portuguese Coast. Lisbon: ICES.
STECF (2006). 23rd Report of the Scientific, Technical and Economic Committee for Fisheries (Second Plenary Meeting). Barza d’ Ispra. Luxembourg: Publications Office of the European Union.
Tsagarakis, K., Palialexis, A., and Vassilopoulou, V. (2014). Mediterranean fishery discards: review of the existing knowledge. ICES J. Mar. Sci. 71, 1219–1234. doi: 10.1093/icesjms/fst074
Vanney, J., and Stanley, D. (1983). “Shelf-Break physiography: an overview,” in Society of Economic Paleontologist and Mineralogist. eds D. J. Stanley and G. T. Moore (Tulsa, Ok: Special Publication).
Vilela, R., and Bellido, J. M. (2015). Fishing suitability maps: helping fishermen reduce discards. Can. J. Fish. Aquat. Sci. 72, 1191–1201. doi: 10.1139/cjfas-2013-0522
Vitale, S., Milisenda, G., Garofalo, G., Tserpes, G., Giannoulaki, M., Ligas, A., et al. (2016). Ecological, Socioeconomic and Technical Characteristics of Discarding Fisheries. Deliverable 1.2 GIS Maps (juveniles). Deliverables Published by the MINOUW Project. Available online at: http://minouw-project.eu/publications-and-papers/
Wilen, J. E., Smith, M. D., Lockwood, D., and Botsford, L. W. (2002). Avoiding surprises: incorporating fisherman behavior into management models. Bull. Mar. Sci. 70, 553–575.
Williams, A., Koslow, J. A., and Last, P. R. (2001). Diversity, density and community structure of the demersal fish fauna of the continental slope off Western Australia (20 to 35°S). Mar. Ecol. Prog. Ser. 212, 247–263. doi: 10.3354/meps212247
Wisz, M. S., Pottier, J., Kissling, W. D., Pellissier, L., Lenoir, J., and Damgaard, C. F. (2013). The role of biotic interactions in shaping distributions and realised assemblages of species: implications for species distribution modelling. Biol. Rev. Camb. Philos. Soc. 88, 15–30. doi: 10.1111/j.1469-185X.2012.00235.x
Keywords: minimum conservation reference size, marine spatial planning, landing obligation, EU common fishery policy, discard reduction, geostatistics, trawl fishery
Citation: Milisenda G, Garofalo G, Fiorentino F, Colloca F, Maynou F, Ligas A, Musumeci C, Bentes L, Gonçalves JMS, Erzini K, Russo T, D’Andrea L and Vitale S (2021) Identifying Persistent Hot Spot Areas of Undersized Fish and Crustaceans in Southern European Waters: Implication for Fishery Management Under the Discard Ban Regulation. Front. Mar. Sci. 8:610241. doi: 10.3389/fmars.2021.610241
Received: 25 September 2020; Accepted: 14 January 2021;
Published: 05 February 2021.
Edited by:
Wen-Cheng Wang, National Taiwan Normal University, TaiwanReviewed by:
Maria Grazia Pennino, Spanish Institute of Oceanography, SpainAthanassios C. Tsikliras, Aristotle University of Thessaloniki, Greece
Copyright © 2021 Milisenda, Garofalo, Fiorentino, Colloca, Maynou, Ligas, Musumeci, Bentes, Gonçalves, Erzini, Russo, D’Andrea and Vitale. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Giacomo Milisenda, Z2lhY29tby5taWxpc2VuZGFAc3puLml0; Z2lhY29tby5taWxpc2VuZGFAZ21haWwuY29t