Abstract
Salt marshes are under increasing anthropogenic pressures that have been reported to affect the diet of fish (e.g., change in prey composition and availability), eventually resulting in alterations in their nursery function. Most studies in Europe are based on fish gut content analysis, which only reflect a small proportion of pressures to salt marshes, and do not necessarily reflect long-term disturbances. In this study, we investigated the impact of salt-marsh vegetation type on trophic network structures (i.e., fish diet and trophic position). Primary producers (particulate organic matter, microphytobenthos, and dominant terrestrial plants), potential aquatic and terrestrial prey, and fish of two dominant species (sea bass and thinlip mullet) were sampled during the summer of 2010 in four creeks from two sites from Western France (the Mont-Saint-Michel Bay and the Seine Estuary). Analysis was undertaken using C and N stable-isotope compositions. Tested response variables (diet and trophic position) suggested a dominant site effect and a weaker effect of surrounding vegetation type. Site effect was attributed to differences in anthropogenic nitrogen inputs (with a steep increase in the Mont-Saint-Michel Bay) and tidal regime between the two bays, with more marine signatures associated with a higher frequency and duration of tidal flooding events in the Seine Estuary. A second hypothesis is that invasive Elytrigia acuta, which has recently replaced typical salt-marsh vegetation in Mont-Saint-Michel Bay, negatively impacted the native salt-marshes nursery function by modifying the access to terrestrial prey on this site. The trophic position of the sea bass and the thinlip mullet was unchanged by local salt-marsh vegetation, and considered consistent with their diet. This study highlights the relevance of stable-isotopes analyses for assessing long-term and integrative effects of changes in vegetation resulting from human disturbances in salt marshes.
Introduction
Mostly localized in the vicinity or within estuaries, salt marshes have typical ecotone functioning with fresh and salt water influences, and are subject to anthropic pressures from both marine and terrestrial ecosystems (McLusky and Elliott, 2004). This location explains their high primary production (up to 30 tonnes of dry matter per ha per year; Lefeuvre et al., 2000). Organic matter resulting from the decomposition of plant material is exported to the sea following high tides, as an outwelling process (Odum, 1980). In unaltered European macrotidal areas, salt-marsh vegetation is entirely submerged only once or twice a month. However, marine waters drill webs of drainage creeks, allowing aquatic fauna to temporarily access salt marshes resources during high tides. The high primary production of salt marshes supports high densities of arthropods and notably decomposers such as amphipods (Schrama et al., 2012). Some of these arthropods are in turn largely consumed by spiders and insectivorous birds (Foucreau et al., 2012; Geslin et al., 2006, respectively) and, when they fall into creeks during flooding (Laffaille et al., 1998; Joyeux et al., 2017), by juvenile fish. The complex created by salt marshes and their creeks thus plays a key role for the survival of many fish species during their first year, and can be qualified as a nursery (e.g., Cattrijsse et al., 1994; Kneib, 1997, the review of Cattrijsse and Hampel, 2006; Green et al., 2009, 2012; Nunn et al., 2016; Joyeux et al., 2017). Nursery habitat value has been described by Sheaves et al. (2015) as including (1) connectivity and population dynamic (2) ecological and ecophysiological factors and (3) resource dynamics. The nursery function of salt marshes can be modified by many disturbances, especially when salt-marsh vegetation is submitted to numerous pressures (e.g., invasive plant species). Evaluating in what extent nursery function changes along with vegetation changes is thus of primary importance to sustain fish populations.
Biological invasions can have huge consequences on ecosystems structure and functioning (see Grosholz, 2002 for an extensive review). Invasive plants are reported as a disturbing factor in several salt marshes. Most of these studies have focused on Spartina spp. (Ayres et al., 2004) that can change the habitat structure of salt marshes and have cascading effects on birds (Gan et al., 2010; Ma et al., 2011), arthropods (Wu et al., 2009), benthic macrofauna (Hedge and Kriwoken, 2000; Neira et al., 2006), deposit-feeding snails (Wang et al., 2014), and global carbon and nitrogen fluxes and stocking (Cheng et al., 2008; Chen et al., 2011; Yin et al., 2015; Yang et al., 2016; Davidson et al., 2018). Other examples include Tamarix spp. (Whitcraft et al., 2008) or Phragmites australis (Silliman and Bertness, 2004). An atypical case of invasion was also reported for the nitrophilous grass Elytrigia acuta (formerly Elymus athericus), which had spectacular effects in several sites of North-West Europe (Bockelmann and Neuhaus, 1999), particularly well characterized in the bay of Mont-Saint-Michel (Valéry et al., 2017). The species occupied only 3% of the total area of salt marshes in 1984 and spread up to 45% of their area in 2013. The invasion by E. acuta has been reported to drastically modify nesting bird (Geslin et al., 2006) and arthropod (Pétillon et al., 2005) communities.
Plant composition of salt marshes is first driven by salt-marsh inundation frequency and a resulting change in soil salinity gradient (e.g., Pétillon et al., 2010), but also depends largely on management practices such as grazing (Bouchard et al., 2003). Inundation frequency and management practices affect both ongoing processes and higher trophic levels. For example, management (grazing) reduces the dominance of Atriplex portulacoides or the invasive E. acuta, with lower arthropod densities (Pétillon et al., 2005), and less insectivorous birds (Geslin et al., 2006). The impact of salt-marsh management is important on potential prey of fish with generally lower abundances in managed areas (e.g., Pétillon et al., 2007; Ford et al., 2012; van Klink et al., 2014). Indeed, salt marshes are essential habitats for particular stages of life cycles for numerous species such as fish or invertebrates (Boesch and Turner, 1984; Rountree et al., 1992; Kneib, 1997; Laffaille et al., 2001). Their nursery function seems particularly impacted by habitat disturbance, and by changes in dominant vegetation as a result of management (Dionne et al., 1999; Laffaille et al., 2000, 2005; Warren et al., 2002; Joyeux et al., 2017). Laffaille et al. (2000) found sea bass juveniles relying less on terrestrial prey of the marsh in grazed areas than in unmanaged areas.
In salt marshes, plant communities are often dominated by a single or a few plant species that are considered as keystone species affecting other species and/or matter fluxes out of proportion to their abundance and through biotic mechanisms (Power et al., 1996). In this study, we tested the broad hypothesis that changes in salt-marsh plant composition cascade into aquatic food webs, including their consumers (e.g., Laffaille et al., 2005; Mantzouki et al., 2012). More precisely, the following expectations were tested: (i) a strong impact of vegetation type and a lesser site effect on fish diet and (ii) some changes in trophic position only visible for predatory species. These hypotheses were tested by comparing the C and N stable-isotope compositions of trophic networks from invasive and natural vegetation from two bays of Western France.
Materials and Methods
Site Description
Two salt marshes located in northwest France were sampled: the Mont-Saint-Michel Bay and the Seine Estuary (Figure 1). The Mont-Saint-Michel Bay is located at the border between Brittany and Normandy at northwestern France (latitude 48°40′N, longitude 1°31′W). It is a macrotidal bay characterized by one of the highest tidal ranges in the world (15 m). The intertidal area covers 220 km2, and is composed of 180 km2 of mudflats (slikke) and 40 km2 of salt marshes (schorre). The salt marsh is drained by a web of creeks filled by tidal waters when the tidal range exceeds 11.25 m (Lefeuvre et al., 2000) for a duration of 2–3 h according to the water level. Salinity range was the same in the two sampling locations: 17.4–35.1 (mean salinity: 32.5 ± 5.7) in 2010. Contrary to the Mont-Saint-Michel Bay, the morphology of the Seine Estuary (located in Normandy: latitude 49°27′N, longitude 0°16′E) has been heavily modified by humans, to reduce tidal influence on intertidal habitats. The tidal range reaches 7 m, and the intertidal area covers 16 km2 and is composed of 4 km2 of mudflats (slikke) and 12 km2 of salt marshes (schorre). Tidal waters fill the creeks when tidal range exceeds 3 m for a period of 5 h according to the water level. The salinity range was 9.8–28.8 (mean salinity: 22.7 ± 4.4) and 3.2–25.3 (mean salinity 16.6 ± 5.8) in each sampling site. Two vegetation types were sampled in the Seine Estuary: one was dominated by Puccinellia maritima, the other by Elytrigia acuta (which, in this site, is not considered as invasive by local managers based on vegetation mapping). The sampling sites (Figure 1) were located in the middle estuary. The Seine Estuary that is characterized by a strong salinity gradient.
FIGURE 1
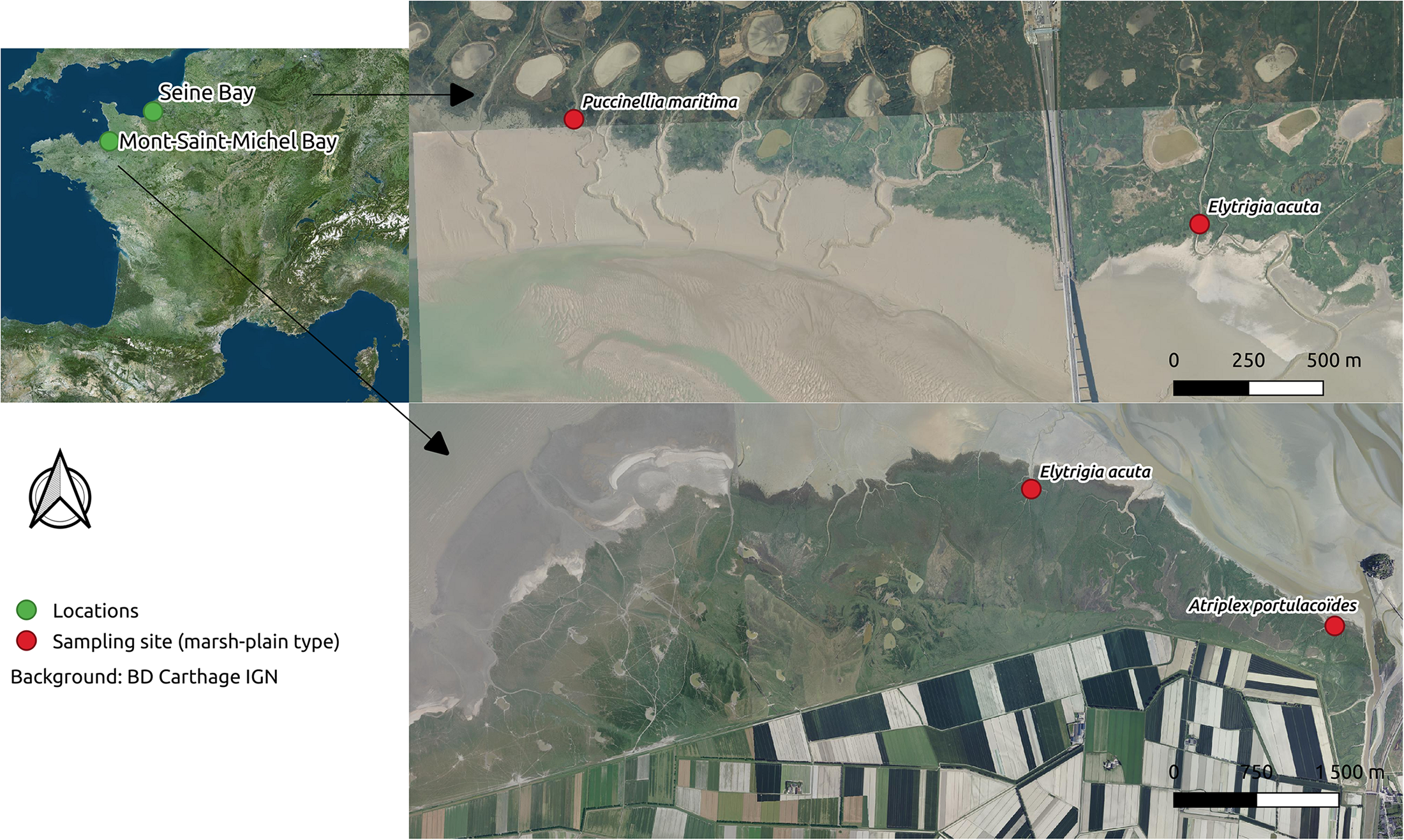
Map of the sampling sites (green dots: Mont-Saint-Michel Bay and Seine Estuary) with sampling locations (red dots with dominant plant species name, referred as marsh-plain type in the body-text).
In the Mont-Saint-Michel Bay, two vegetation types were also sampled: one dominated by E. acuta (which is considered as invasive in this site, see Valéry et al., 2017 for a review) and the other by native Atriplex portulacoides. The dominant vegetation types in which sampling took place is from here on referred to “marsh-plain type.”
Food Web Sampling and Stable Isotopes Preparation
Sampling took place during summer 2010. A fyke net (4 mm mesh size) was set across one representative creek within each site in both salt marshes to catch fish leaving sites after the ebb (see Joyeux et al., 2017 for more details on a similar design). All fish captured were identified and measured (fork length) to get a proxy of individual age. Only the most abundant species, i.e., juvenile sea bass (Dicentrarchus labrax) and juvenile thinlip mullet (Chelon ramada), were kept for isotope analyses. Size ranges were 30.8 ± 8.4 mm and 26.5 ± 5.3 mm for D. labrax, 55.60 ± 11.1 mm and 72.5 ± 12.6 mm for C. ramada, in Mont-Saint-Michel Bay and Seine Estuary, respectively. White muscle tissues were collected from each individual at the laboratory.
Potential preys were collected by hand or caught in the fyke net when sampling fish. A minimum of 10 sample individuals per site and vegetation type and per predator was collected. For potential prey species (based on previous studies using gut content identification, e.g., Laffaille et al., 2001, 2002, 2005), a minimum of six samples per site was collected. Crustaceans (Carcinus maenas, Lekanesphaera rugicauda, Mysida, Orchestia gammarellus, and Palaemonetes varians) samples were subjected to a rapid acid wash to reduce inorganic carbon content. For these species, an extra sample (not submitted to acid washing) was collected to check for the possible impact of the decarbonation treatment on δ15N values (Pinnegar and Polunin, 1999). No significant differences were found, so only decarbonated samples were used.
Particulate organic matter (POM), microphytobenthos (MPB), and terrestrial plants were sampled as different primary producers. Terrestrial plants were collected by hand. For the Mont-Saint-Michel Bay, POM came from 1 L of water collected in the creek during the ebb. The water was filtered on decarbonated (4 h at 550°C) GF-F filters (47 mm diameter Whatman). For the Seine Estuary, as analyses of field samples failed, POM data were extracted from the SOMLIT1 long-term database (9 values for 2010). MPB was extracted from the superficial layer (5 mm) of scraped muddy sediment by exposing it to light during 2 h. The sediment was then covered with a 100 μm nylon filter and sand previously sieved (63 and 250 μm) and decarbonated (5 h, 550°C). After waiting several hours for MPB to migrate to the surface through the nylon filter, the superficial layer (2 mm) was scraped and sieved through seawater on a 45 μm filter. The content of the filter was then filtered again under decarbonated GF-F filters (4 h, 550°C).
All samples (except for the filters -POM and MPB) were then frozen before being lyophilized and crushed into a fine powder, then dried for 24 h at 50°C in a drying oven. No delipidation was performed on lyophilized samples since the C/N threshold of 3.5 proposed by Post et al. (2007) for aquatic animal samples has never been reached.
Stable Isotopes Analyses
The nitrogen and carbon isotopic compositions were determined using EA- IRMS (Isoprime, Micromass, United Kingdom). The carbon and nitrogen isotope ratios are expressed in the delta notation δ13C and δ15N where: δX = [(RSample / RReference) − 1] × 1000, where X = δ13C or δ15N and R is the ratio 13C:12C or 15N:14N in the sample and in the reference material. Results are referred to Vienna Pee Dee Belemnite (VPDB) for C and to atmospheric nitrogen for N and expressed in units of ‰ ± standard deviation (sd). The analytical precision (standard deviation for repeated measurements of internal standards) was ±0.2‰ and ±0.3‰ for δ13C or δ15N, respectively.
Statistical Analyses
Isotopic signatures of fish, prey and baselines were compared between sites and marsh-plain types using multivariate analysis of variance (MANOVA) with site, marsh-plain type and site/marsh-plain type interaction as fixed factors. Normality of the δ15N and δ13C values was evaluated using Kolmogorov–Smirnov test. As both variables had a normal distribution, parametric tests were used.
Fish diet partitioning was achieved using Bayesian mixing model performed with package MixSIAR 3.1.10 (Stock and Semmens, 2013) in R 3.6.1 (R Core Team, 2020) with two tracers: δ13C and δ15N.
As a first step, models were built for each fish species with POM, MPB and terrestrial plant as potential primary producers. We used primary producers rather than potential prey as we were interested in studying the effect of terrestrial plants changes in the overall contribution of salt-marsh components. For D. labrax, we also built models with potential prey, inferred from stomach contents obtained from previous studies and stable isotope signatures. Given the close isotopic signatures of Carcinus maenas, Mysida sp., Palaemonetes varians, and Hediste diversicolor (Figure 2), the four prey were grouped altogether (further referred as “marine preys”) for diet modeling. Chelon ramada juveniles being grazers (Lebreton et al., 2011), no diet mixing model based on prey was built. For each fish species, a null model and models including different combinations of random (site) and fixed (marsh-plain type) effects were tested with the following settings: three chains with 300,000 iterations and burn-in of 200,000 to allow for adequate model convergence. Site was included as a random factor for both fish species and primary producer signatures. Model convergence was checked using Gelman-Rubin and Geweke diagnostics. Candidate models were then compared using leave-one-out cross-validation (LOO). Input values for each model were the raw isotopic values for fish and mean and standard deviation of each primary producer. To compare the relative contribution of factors included in the best model, a new model with both factors included as random variables was run. Trophic enrichment factor values were derived from Kostecki et al. (2012) and Selleslagh et al. (2015). Two sets of values were used according to fish species trophic guilds. Values for predator (D. labrax) were 2 ± 0.6% and 5.6 ± 1.5% for δ13C and δ15N, respectively. Values for grazer (C. ramada) were 1 ± 0.6% and 3.4 ± 1.5% for δ13C and δ15N, respectively (Carpentier et al., 2014; Como et al., 2018).
FIGURE 2
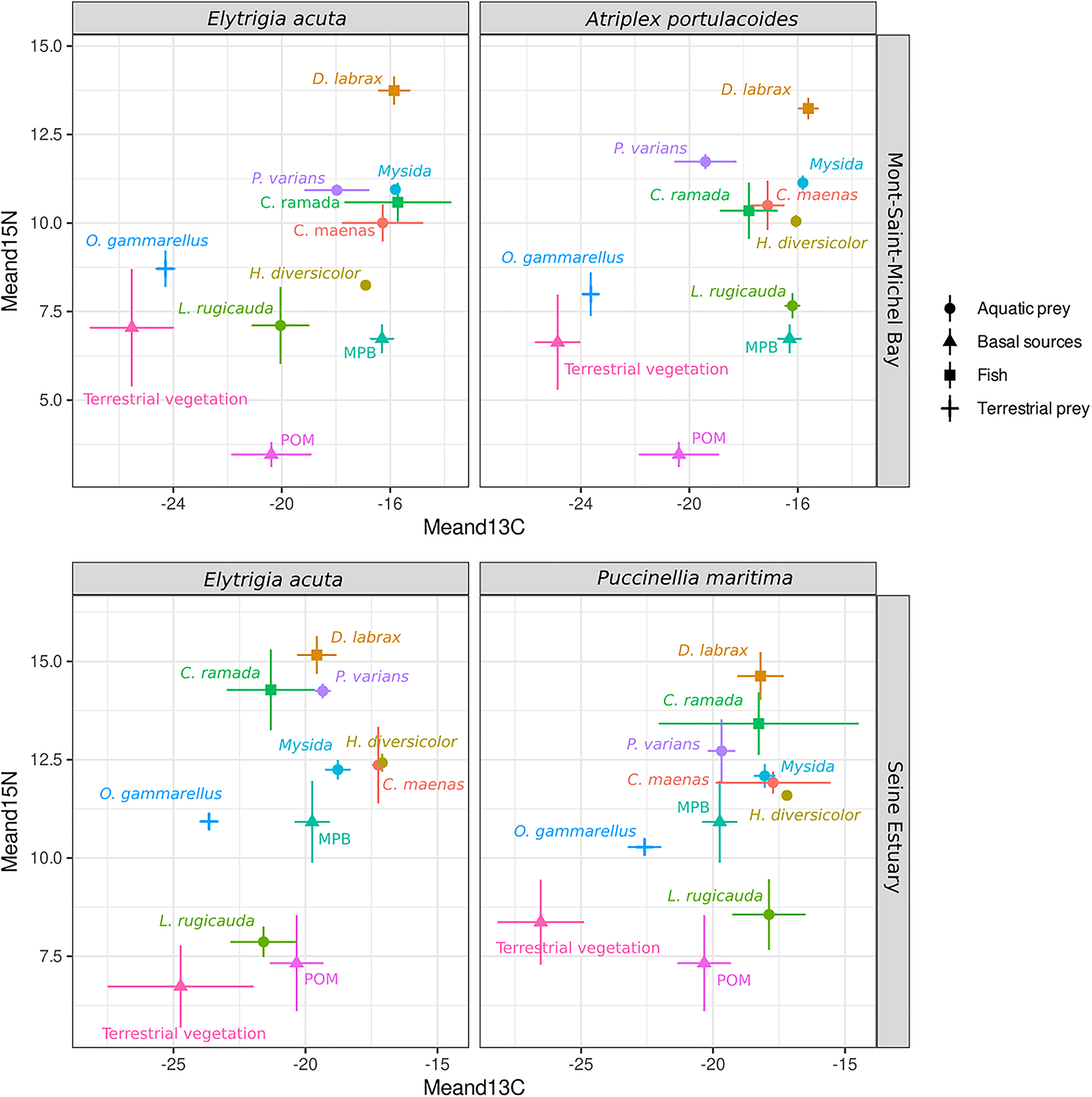
Biplots presenting the mean and standard deviation of isotopic values (δ13C and δ15N) of fish, their prey and basal resources for each site and marsh-plain type (Elytrigia acuta, Atriplex portulacoides, Puccinellia maritima). Note, the prey and basal resources in the models are without marsh-plain type effects, the values presented are means per site.
The fish trophic position was estimated using a two-sources Bayesian model. The two sources included were the ones contributing most to the fish’s diet according to the previous mixing models. Estimation was performed using the tRophicPosition 0.7.7 package (Quezada-Romegialli et al., 2019). Input values for each model were the raw isotopic values for fish and primary producers. The grouping variable was marsh-plain type.
Results
Overall Stable Isotope Composition of Salt-Marsh Food Web
The MANOVA on fish isotopic signatures (Figure 2, Table 1, and Supplementary Table 1) demonstrated a significant effect of fish species (F = 49.23, df = 4, p < 0.001), site (F = 220.86, df = 2, p < 0.001) and marsh-plain type (F = 9.07, df = 4, p < 0.001), but no interaction between site and marsh-plain type (F = 2.48, df = 2, p = 0.087). The MANOVA on primary producers (MPB, POM, and terrestrial plants) isotopic signatures (Table 1) demonstrated significant differences between primary producers (F = 11.21, df = 4, p < 0.001), sites (F = 27.32, df = 2, p < 0.001), but no effect of marsh-plain type (F = 2.03, df = 4,z p = 0.094), and no interaction between site and marsh-plain type (F = 0.05, df = 2, p = 0.943).
TABLE 1
| Marsh plain type | Species | Common name | MSMB | SE |
| E. acuta | Dicentrarchus labrax | Sea bass | 14 | 20 |
| E. acuta | Chelon ramada | Thinlip mullet | 14 | 10 |
| Native vegetation | Dicentrarchus labrax | Sea bass | 21 | 25 |
| Native vegetation | Chelon ramada | Thinlip mullet | 13 | 10 |
| Carcinus maenas | Shore crab | 7 | 7 | |
| Chorthippus albomarginatus | Lesser marsh grasshopper | 3 | 3 | |
| Hediste diversicolor | Ragworm | 6 | 5 | |
| Lekanesphaera rugicauda | Sea slater | 6 | 5 | |
| Lycosidae | Wolf spiders | 6 | 2 | |
| Microphytobenthos | 6 | 6 | ||
| Mysida | Opossum shrimp | 6 | 6 | |
| Orchestia gammarellus | Sea flea | 5 | 5 | |
| Palaemonetes varians | Common ditch shrimp | 6 | 6 | |
| Particulate organic matter | 6 | 3 | ||
| Terrestrial vegetation | 24 | 18 |
Number of samples collected (per marsh-plain type and site for predators and per site for primary producers and prey).
SE, Seine Estuary; MSMB, Mont-Saint-Michel Bay. “Native vegetation” refers to Atriplex portulacoides in the MSMB and to Puccinellia maritima in the SE.
The MANOVA on potential prey isotopic signature demonstrated a significant effect of prey species (F = 32.04, df = 42, p < 0.001), site (F = 87.06, df = 2, p < 0.001) and marsh-plain type (F = 5.10, df = 4, p < 0.001), but no interaction between site and marsh-plain type (F = 0.234, df = 2, p = 0.790).
Fish Diet Partitioning
Diets of the two fish species were better explained by models based on primary producers (MPB, POM, and terrestrial plants) including both site and marsh-plain type (Table 2). Similar results were found for D. labrax models based on potential prey (Table 3).
TABLE 2
| Fish species | Common name | Model | LOOic | Std error LOOic | Weight |
| Dicentrarchus labrax | Sea bass | Site + marsh-plain type | −126.2 | 25.9 | 1.0 |
| Site | −103.4 | 22.6 | 0.0 | ||
| Marsh-plain type | −19.0 | 25.0 | 0.0 | ||
| Null | 92.5 | 14.1 | 0.0 | ||
| Chelon ramada | Thinlip mullet | Site + marsh-plain type | 99.7 | 0.0 | 0.852 |
| Site | 103.2 | 13.8 | 0.148 | ||
| Marsh-plain type | 173.9 | 74.2 | 0.0 | ||
| Null | 195.9 | 96.2 | 0.0 |
LOOic values of each mixing model tested with primary producers (microphytobenthos, particulate organic matter, and terrestrial vegetation).
Weights closer to 1 indicate an increased probability that the model explains diet partitioning.
TABLE 3
| Model | LOOic | Std error LOOic | Weight |
| Site + marsh-plain type | 25.0 | 27.3 | 1.0 |
| Site | 59.7 | 22.7 | 0.0 |
| Marsh-plain type | 156.2 | 23.5 | 0.0 |
| Null | 278.5 | 14.1 | 0.0 |
LOOic values of each mixing model tested with potential prey as sources for Dicentrarchus labrax.
High weight indicates high probability the model is the best explaining diet partitioning.
The posterior estimates of diet proportions demonstrated that MPB and POM were the main sources of organic matter contributing to D. labrax diet (Figure 3). Site and marsh-plain type equally contributed to variations in D. labrax diet [site median standard deviation = 8.17 (95% CI = 1.65–17.86), marsh-plain type median standard deviation: 6.21 (95% CI = 1.16–16.24)]. For sampling locations from the Mont-Saint-Michel Bay, no terrestrial primary producer contribution was observed in D. labrax diet, whichever marsh-plain type the individuals were sampled in (Figure 3 and Supplementary Table 2). In the Seine Estuary, the contribution of terrestrial primary producers to D. labrax diet was 0.274 (95% CI = 0.190–0.346) for individuals collected in E. acuta-dominated marsh-plain areas, and 0.049 (95% CI = 0.009–0.098) in P. maritima areas. MPB was the main source of aquatic primary producers for D. labrax (>0.975) in the Mont-Saint-Michel Bay, whichever marsh-plain type the individuals were sampled in (Figure 3 and Supplementary Table 2). In the Seine Estuary, POM and MPB both contributed to D. labrax diet in similar proportions (0.427 and 0.525), except in E. acuta areas where POM was lower (0.185 and 0.541) (Figure 3 and Supplementary Table 2).
FIGURE 3
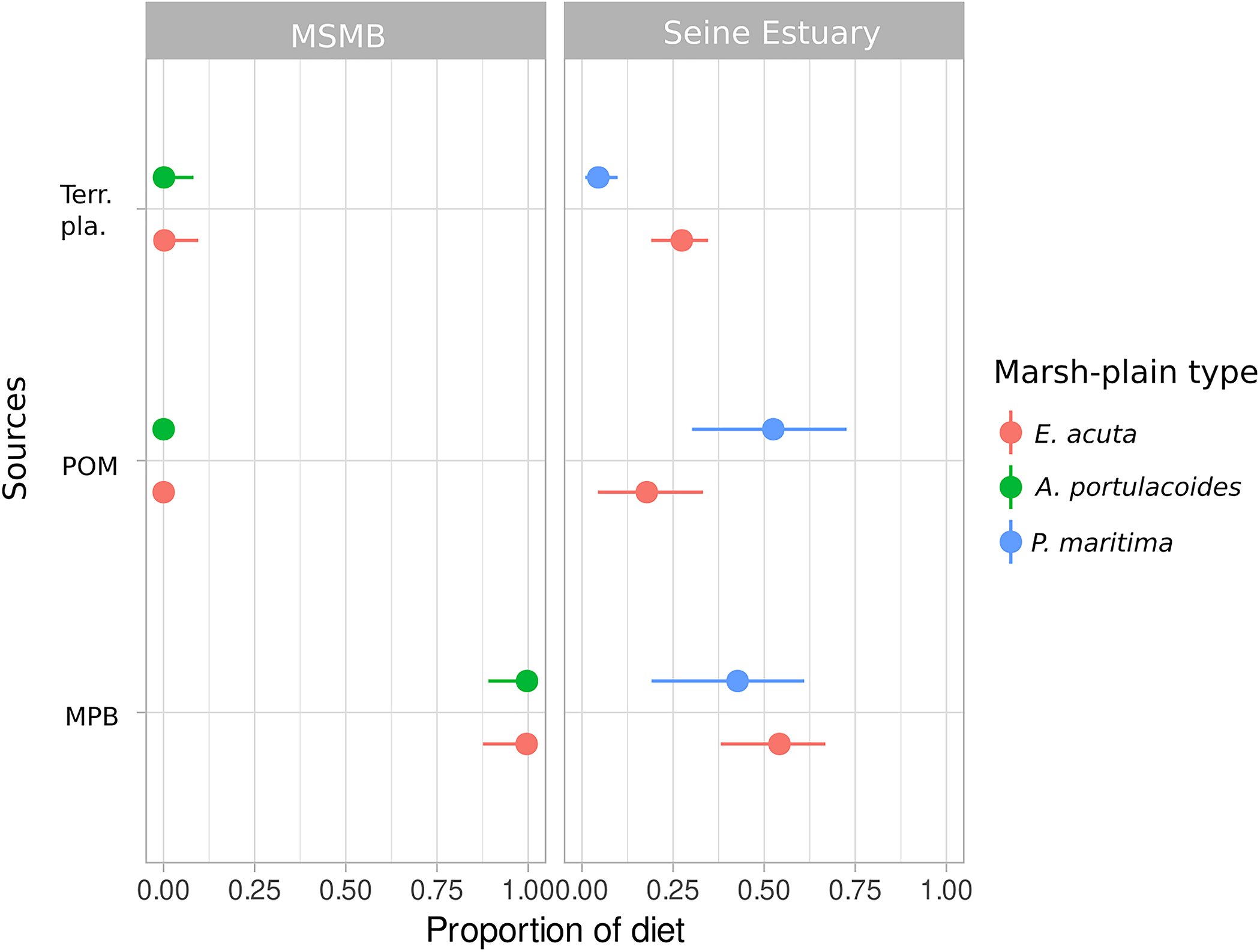
Posterior median (points) with 90% (horizontal lines) confidence intervals for contribution of each primary producer (MPB, microphytobenthos; POM, particulate organic matter; Terr. pla., terrestrial vegetation) according to marsh-plain type in the sampling area for D. labrax. MSMB, Mont-Saint-Michel Bay.
The posterior estimates of diet proportions based on prey signatures showed that marine species (Carcinus maenas, Mysida sp., Palaemonetes varians, and Hediste diversicolor) were the main prey of D. labrax in the Mont-Saint-Michel Bay (contribution >0.5 in all cases, Figure 4 and Supplementary Table 3). Nevertheless the contribution of Lekanesphaera rugicauda and Orchestia gammarellus were also important, especially in E. acuta-dominated areas of the Seine Estuary (Figure 4 and Supplementary Table 3).
FIGURE 4
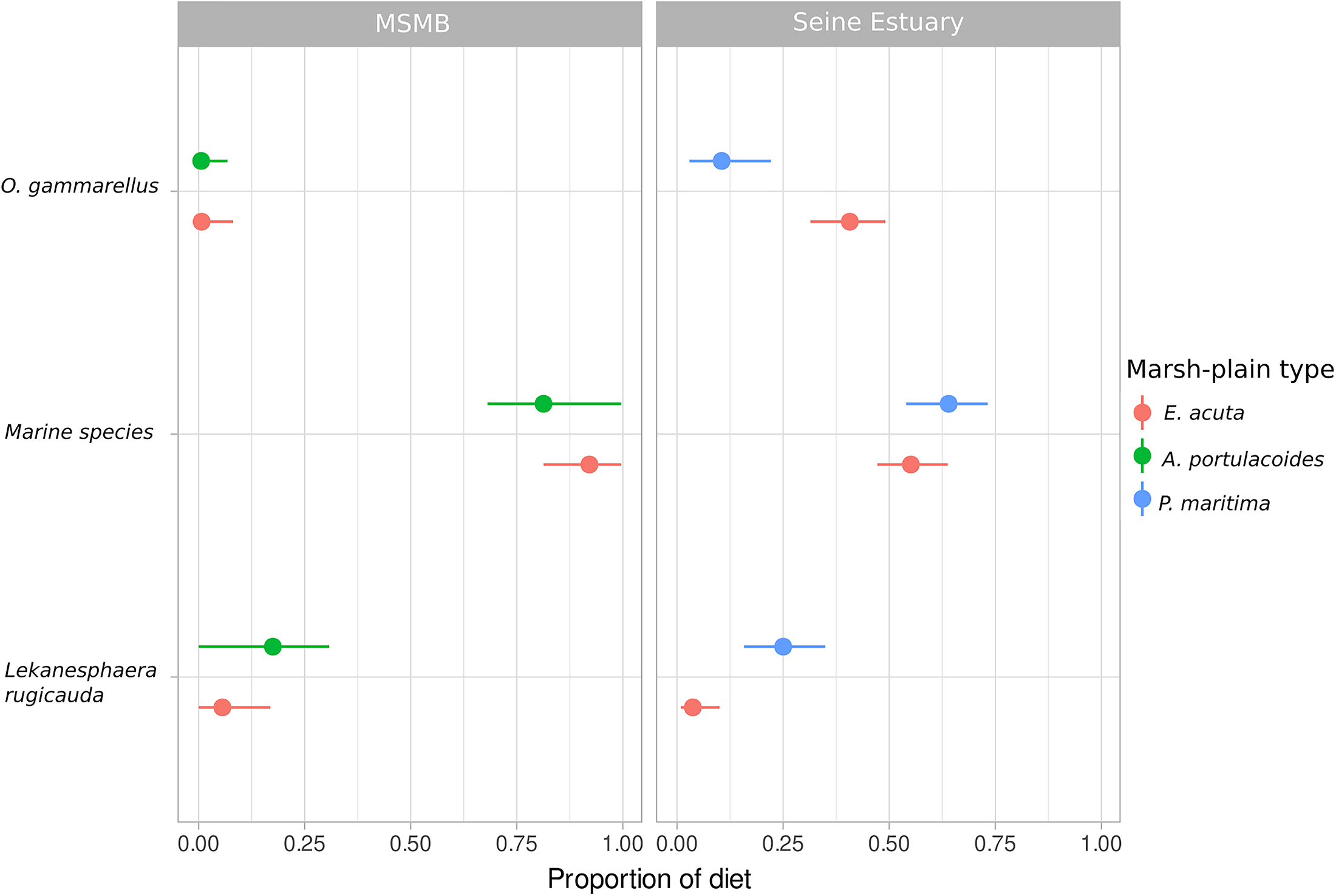
Posterior median (points) with 90% (horizontal lines) confidence intervals for the contribution of each prey according to marsh-plain type in the sampling area for Dicentrarchus labrax. MSMB, Mont-Saint-Michel Bay. Marine species refers to the group: Carcinus maenas, Mysida sp., Palaemonetes varians, and Hediste diversicolor.
The posterior estimates of overall diet proportions showed that C. ramada’s main source of carbon was MPB in both sites and for all marsh-plain types (Figure 5 and Supplementary Table 2). In the Mont-Saint-Michel Bay, terrestrial plants also contributed to C. ramada diet in areas dominated by A. portulacoides (up to 0.148, see Figure 5 and Supplementary Table 2).
FIGURE 5
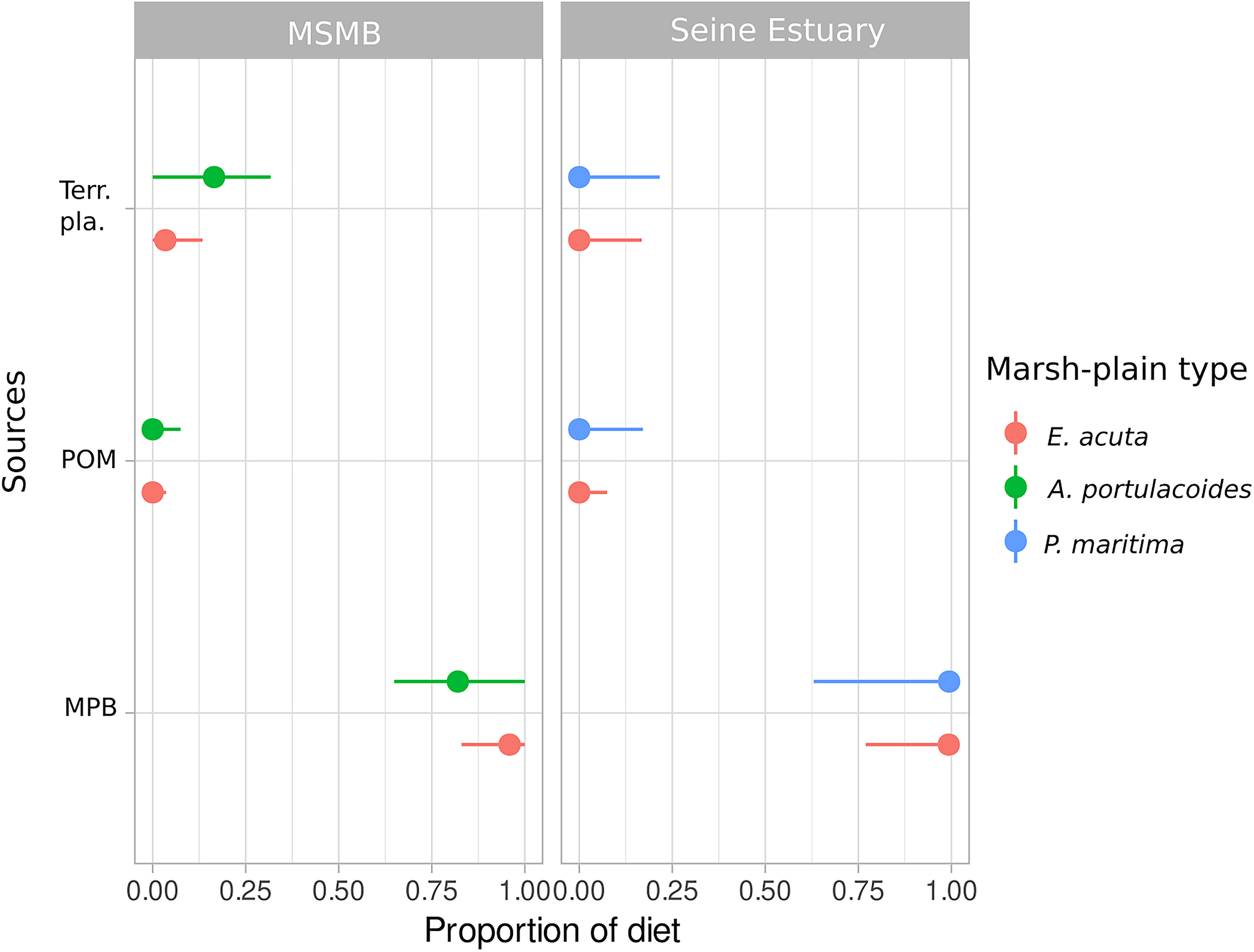
Posterior median (points) with 90% (horizontal lines) confidence intervals for contribution of each primary producer (MPB, microphytobenthos; POM, particulate organic matter; Terr. pla., terrestrial plants) according to marsh-plain type in the sampling area for Chelon ramada. MSMB, Mont-Saint-Michel Bay.
Fish Trophic Position
Fish trophic position did not vary with marsh-plain types, since probabilities of significant differences of posterior estimates were consistently inferior to 0.5 (Supplementary Tables 4, 5 and Figure 6). On the other hand, fish trophic position varied systematically with fish species, with high probabilities of difference observed when comparing sites and fish species trophic position between each other (indicating that fish species have different trophic position and that the latter is varying with site) (Supplementary Tables 4, 5 and Figure 6).
FIGURE 6
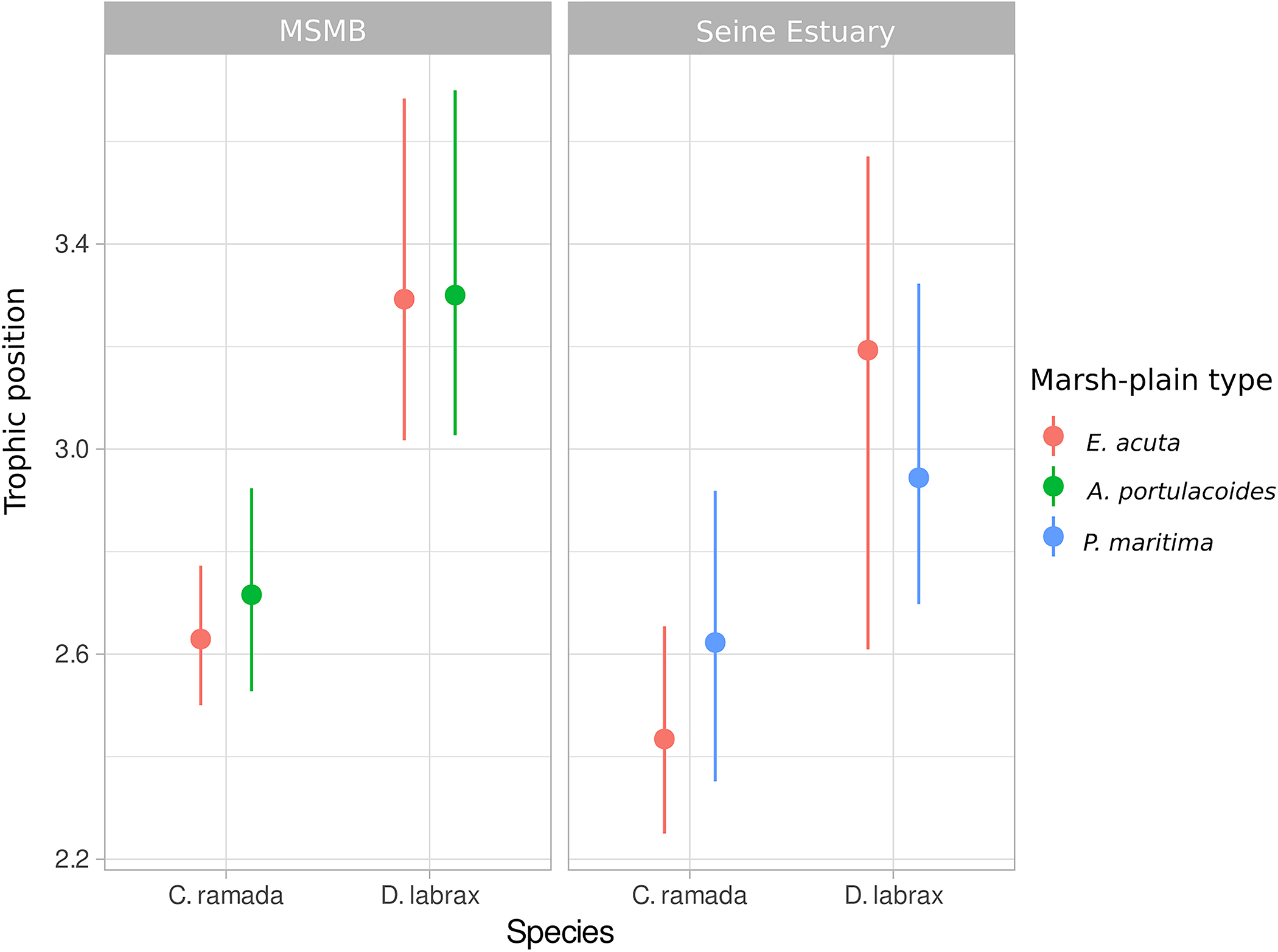
Median and 95% CI of posterior trophic position for each fish species according to marsh-plain type. MSMB, Mont-Saint-Michel Bay, C. ramada, Chelon ramada; D. labrax, Dicentrarchus labrax.
Discussion
To the best of our knowledge, this study is the first to investigate the effect of terrestrial vegetation type on salt-marsh nursery function for juvenile fish using stable isotopes. We found a clear difference in the contribution of main primary producers to sea bass (D. labrax) and thinlip mullet (C. ramada) diets. Nevertheless, vegetation type had an influence on the contribution of primary producers and prey only to D. labrax diet and only in the Seine Estuary. In the Seine Estuary, however, terrestrial primary producers had a significant contribution to D. labrax diet. The most striking and unexpected result of our study was the very low contribution of terrestrial carbon to D. labrax diet provided by salt marshes in the Mont-Saint-Michel Bay (MSMB) which strongly contrasts with the conclusions of previous studies (e.g., Laffaille et al., 2001, 2005). Interestingly, we could not find any difference of trophic position between the different marsh-plain types (i.e., types of vegetation surrounding the creek) nor for D. labrax nor for C. ramada.
Results of Bayesian mixing models suggest that the contribution of primary producers to juvenile D. labrax did not vary with marsh-plain type where fish are foraging [i.e., >90% microphytobenthos (MPB)] in the MSMB. MPB was the dominant source in the Seine Estuary but other primary producers (POM and terrestrial vegetation) significantly contributed to D. labrax diet, in proportions differing according to the marsh-plain type. The mixing model based on prey revealed that marine species (Carcinus maenas, Mysida, Palaemonetes varians, and Hediste diversicolor) appeared to be the main prey of juvenile D. labrax in both sites, but O. gammarellus and Lekanesphaera rugicauda significantly contributed to D. labrax diet in areas dominated, respectively, by E. acuta and P. maritima in the Seine Estuary.
From these results, one can state that in the Seine Estuary, terrestrial primary producers have a significant contribution to the diet of D. labrax juveniles, of up to 30% in areas dominated by E. acuta. Conversely, the terrestrial contribution of salt-marsh sources to D. labrax diet was almost inexistent in the MSMB. In the MSMB, Laffaille et al. (2001) identified Mysida as the main prey of D. labrax juveniles entering salt marshes but O. gammarellus was the main prey in the stomach of individuals leaving salt marshes during ebb. In Laffaille et al. (2005) demonstrated, based on stomach content analyses, that in the MSMB, D. labrax juveniles were consuming less O. gammarellus in areas invaded by E. acuta. In the same study, they estimated the numerical contribution of O. gammarellus to the diet of D. labrax juveniles as 58% (73% in biomass) in uninvaded areas and 28% (40% in biomass) in invaded areas. They hypothesized that such change in diet was due to a decrease in accessibility of terrestrial prey (here O. gammarellus) because of changes in vegetation structure (increased stem density may have been a physical obstruction for fish preying on amphipods). Laffaille et al. (2005) indeed observed a reduced prey availability due to the E. acuta invasion, but considered it was still high enough to not act a limiting factor on fish feeding behavior.
Our results suggest that, about 15 years after Laffaille et al. (2005) study, the contribution of terrestrial primary producers to D. labrax diet decreased to close to zero in the MSMB, both in creeks invaded by E. acuta and natural ones. We also found no significant difference in O. gammarellus contribution between marsh-plain types in the Seine Estuary, suggesting that vegetation is not the main factor explaining the contribution of terrestrial prey to D. labrax diet.
We hypothesize that, in the MSMB, the access to prey was altered by changes in halophylic vegetation, resulting from increasing nitrogen inputs (the main factor explaining the spread of E. acuta in the Saint-Michel Bay: Valéry et al., 2017). Nelson et al. (2019) demonstrated that the nutrient enrichment of waters entering salt marshes triggers a cascade of effects, resulting in geomorphological changes (creek deepening and enlargement with cliff-like banks: Deegan et al., 2012) and decoupling the high-marsh ecosystem from the creek ecosystem. Similarly to our results on D. labrax in the MSMB, Nelson et al. (2019) observed a lower contribution of terrestrial prey to mummichog (Fundulus heteroclitus) diet. Our hypothesis is reinforced by the situation in the Seine Estuary where E. acuta is not invasive and where D. labrax juveniles appear more dependent on salt-marsh sources (O. gammarellus contributed up to 40% in diet partitioning). In the Seine Estuary, the contribution of terrestrial prey to D. labrax diet varied with vegetation type. We are aware that our interpretation implies that site fidelity exists for juveniles of D. labrax. Despite several studies demonstrating that D. labrax are site (and even creek) dependent (Vinagre et al., 2011; Green et al., 2012; Joyeux et al., 2017), evidence is still lacking, and should be the focus of further studies.
Differences in the contributions of terrestrial primary producers between the two sites may be the result of different frequencies of access to salt marshes for fish. The two sites strongly differ in tidal amplitude (15 m in the MSMB vs. 7 m in the Seine Estuary) and high water slack duration (zero in the MSMB vs. ±3 h in the Seine Estuary because of the existence in the channel of two asynchronic tides entering the estuary). Because of this differing tidal amplitude, the percent of tides during which fish are able to enter into salt-marsh creeks are estimated to be 43% per year in the MSMB (Laffaille et al., 2001) and up to 60% in the Seine Estuary (Duhamel comm.pers.). Finally, the huge difference in the total surface of sites, with 220, 180, and 40 km2 in MSMB and 16, 4, and 12 km2 in the Seine Estuary for intertidal area, mudflats and salt marsh areas, respectively, could also lead to contrasting accessibility and attractivity for fish. One can thus hypothesize that such a difference in the ability for fish to exploit salt-marsh creeks for feeding, would have a significant effect on their isotopic signature.
As expected, juveniles of C. ramada derived most of their carbon from benthic primary producers, and particularly MPB. Similar results were reported by Lebreton et al. (2011) using stable isotopes. These authors confirmed that C. ramada juveniles exploiting salt marshes are limno-benthophagous. Nevertheless, the uncertainty regarding the contribution of each primary producer was very high in our study. Such a result suggests that this species probably uses several salt marshes with different isotopic signatures to forage. This hypothesis tends to be confirmed by the telemetry results obtained by Trancart et al. (2012) and by several studies on other species highlighting the existence of individual variations in fish movement behavior that can mediate the direction and scale of food web subsidies (e.g., Lesser et al., 2020; Rezek et al., 2020).
The trophic position of the two studied fish species was not different according to the marsh-plain type. This result is not surprising as we found a very weak contribution of terrestrial primary producers to fish diet in the MSMB. In the Seine Estuary, despite the higher contribution of terrestrial primary producers to fish diet, the trophic position of fish did not vary with vegetation type.
We indeed found a strong, unexpected site effect on all characteristics of the food web. All best diet partitioning models included site and in a lesser extent vegetation type as a predictive variable. Nevertheless, the large uncertainty on the contribution of the two variables renders any comparison hard. A long-term study combining stable isotopes and stomach content identification approaches could probably confirm our findings. Nevertheless, diet and food availability have been largely studied over the last two decades notably in MSMB (Laffaille et al., 2001, 2005; Carpentier unpublished data from 2015 to 2017), limiting potential errors in preys identification through isotopes. The contribution of terrestrial sources to predatory fish diet was much higher in the Seine Estuary. At a larger spatial scale, a recent review by Ziegler et al. (2019) emphasized the key role of tidal amplitude in structuring food webs. We believe that a similar process (i.e., tides regulate the access to salt marshes for transient estuarine fishes) can be observed within the same biogeographic area. We finally hypothesize that the importance of nursery function of macrotidal salt marshes in Western Europe would be more diverse than expected given the large inter-site variability and overall low terrestrial contribution to fish diet we found in this study.
Statements
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://gitlab.com/DenisLafage/marsh_veget_foodweb.
Author contributions
AC and JP conceived and designed the experiment. AC, SD, and SL conducted the field and lab work. DL performed the statistical analyses. AC, DL, and JP wrote the manuscript draft. All authors contributed critically to the drafts and gave final approval for publication.
Acknowledgments
We would like to thank the Programme Seine-Aval that financed this study through the DEFHFIS program and the Agence de l’Eau Loire-Bretagne (PEPPS project) and the Région Bretagne (SAD). We would also like to thank Charlène Puzin for her help in the field and samples preparation, the Plateforme de Spectrométrie Isotopique of the LIENSs laboratory of La Rochelle for the isotopic analyses and especially Gaël Guillou and Benoit Lebreton, and Sophie Elliott for editing the English.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2021.669759/full#supplementary-material
Footnotes
References
1
AyresD. R.SmithD. L.ZarembaK.KlohrS.StrongD. R. (2004). Spread of exotic cordgrasses and hybrids (Spartina sp.) in the tidal marshes of San Francisco Bay, California, USA.Biol. Invasions6221–231. 10.1023/B:BINV.0000022140.07404.b7
2
BockelmannA. C.NeuhausR. (1999). Competitive exclusion of Elymus athericus from a high-stress habitat in a European saltmarsh.J. Ecol.87503–513.
3
BoeschD. F.TurnerR. E. (1984). Dependence of fishery species on salt marshes: the role of food and refuge.Estuaries7:460. 10.2307/1351627.
4
BouchardV.TessierM.DigaireF.VivierJ.-P.ValeryL.GloaguenJ.-C.et al (2003). Sheep grazing as management tool in Western European saltmarshes.C. R. Biol.326148–157. 10.1016/S1631-0691(03)00052-0
5
CarpentierA.ComoS.DupuyC.LefrançoisC.FeunteunE. (2014). Feeding ecology of Liza spp. in a tidal flat: evidence of the importance of primary production (biofilm) and associated meiofauna.J. Sea Res.9286–91. 10.1016/j.seares.2013.10.007
6
CattrijsseA.HampelH. (2006). European intertidal marshes: a review of their habitat functioning and value for aquatic organisms.Mar. Ecol. Prog. Ser.324293–307. 10.3354/meps324293
7
CattrijsseA.MakwaiaE. S.DankwaH. R.HamerlynckO. (1994). Nekton communities of an intertidal creek of a European estuarine brackish marsh.Mar. Ecol. Prog. Ser.109195–208.
8
ChenI.-C.HillJ. K.OhlemüllerR.RoyD. B.ThomasC. D. (2011). Rapid range shifts of species associated with high levels of climate warming.Science3331024–1026. 10.1126/science.1206432
9
ChengX.ChenJ.LuoY.HendersonR.AnS.ZhangQ.et al (2008). Assessing the effects of short-term Spartina alterniflora invasion on labile and recalcitrant C and N pools by means of soil fractionation and stable C and N isotopes.Geoderma145177–184. 10.1016/j.geoderma.2008.02.013
10
ComoS.CarpentierA.RossiF.DupuyC.RichardP.FeunteunE.et al (2018). Stable isotopes as tracers can reveal resource allocation in juvenile golden gray mullets (Liza aurata, Risso, 1810).J. Exp. Mar. Biol. Ecol.50372–79. 10.1016/j.jembe.2018.01.009
11
DavidsonI. C.CottG. M.DevaneyJ. L.SimkaninC. (2018). Differential effects of biological invasions on coastal blue carbon: a global review and meta-analysis.Glob. Change Biol.245218–5230. 10.1111/gcb.14426
12
DeeganL. A.JohnsonD. S.WarrenR. S.PetersonB. J.FleegerJ. W.FagherazziS.et al (2012). Coastal eutrophication as a driver of salt marsh loss.Nature490388–392. 10.1038/nature11533
13
DionneM.ShortF. T.BurdickD. M. (1999). “Fish utilization of restored, created and reference salt-marsh habitat in the Gulf of Maine”American Fisheries Society SymposiumBethesda, MD: The American Fisheries Society.
14
FordH.GarbuttA.JonesL.JonesD. L. (2012). Grazing management in saltmarsh ecosystems drives invertebrate diversity, abundance and functional group structure.Insect Conserv. Divers.6189–200. 10.1111/j.1752-4598.2012.00202.x
15
FoucreauN.RenaultD.LuganR.HidalgoK.PétillonJ. (2012). Effects of diet and salinity on the survival, egg laying and metabolic fingerprints of the ground-dwelling spider Arctosa fulvolineata (Araneae, Lycosidae).Comp. Biochem. Physiol. A Mol. Integr. Physiol.163388–395.
16
GanX.ChoiC.WangY.MaZ.ChenJ.LiB. (2010). Alteration of habitat structure and food resources by invasive smooth cordgrass affects habitat use by wintering saltmarsh birds at Chongming Dongtan, East China.The Auk127317–327. 10.1525/auk.2009.09147
17
GeslinT.EybertM.RadureauA. (2006). Influence of natural and anthropic perturbations on the distribution of salt marsh breeding birds in the Mont Saint-Michel Bay.Cah. Biol. Mar.4723–30.
18
GreenB. C.SmithD. J.EarleyS. E.HepburnL. J.UnderwoodG. J. C. (2009). Seasonal changes in community composition and trophic structure of fish populations of five salt marshes along the Essex coastline, United Kingdom.Estuar. Coast. Shelf Sci.85247–256. 10.1016/j.ecss.2009.08.008
19
GreenB. C.SmithD. J.GreyJ.UnderwoodG. J. C. (2012). High site fidelity and low site connectivity in temperate salt marsh fish populations: a stable isotope approach.Oecologia168245–255. 10.1007/s00442-011-2077-y
20
GrosholzE. (2002). Ecological and evolutionary consequences of coastal invasions.Trends Ecol. Evol.1722–27. 10.1016/S0169-5347(01)02358-8
21
HedgeP.KriwokenL. K. (2000). Evidence for effects of Spartina anglica invasion on benthic macrofauna in Little Swanport estuary, Tasmania.Austral Ecol.25150–159. 10.1046/j.1442-9993.2000.01016.x
22
JoyeuxE.CarpentierA.CorreF.HaieS.PétillonJ. (2017). Impact of salt-marsh management on fish nursery function in the bay of Aiguillon (French Atlantic coast), with a focus on European sea bass diet.J. Coast. Conserv.21435–444. 10.1007/s11852-017-0501-0
23
KneibR. T. (1997). Early life stages of resident nekton in intertidal marshes.Estuaries20214–230. 10.2307/1352732
24
KosteckiC.RousselJ. M.DesroyN.RousselG.LanshereJ.Le BrisH.et al (2012). Trophic ecology of juvenile flatfish in a coastal nursery ground: contributions of intertidal primary production and freshwater particulate organic matter.Mar. Ecol. Prog. Ser.449221–232.
25
LaffailleP.BrosseS.FeunteunE.BaisezA.LefeuvreJ.-C. (1998). “Role of fish communities in particulate organic matter fluxes between salt marshes and coastal marine waters in the Mont Saint-Michel Bay,” in Oceans, Rivers and Lakes: Energy and Substance Transfers at Interfaces, edsAmiardJ.-C.Le RouzicB.BerthetB.BertruG. (Dordrecht: Springer Netherlands), 121–133.
26
LaffailleP.FeunteunE.LefeuvreJ.-C. (2000). Composition of fish communities in a European macrotidal salt marsh (the Mont Saint-Michel Bay, France).Estuar. Coast. Shelf Sci.51429–438. 10.1006/ecss.2000.0675
27
LaffailleP.FeunteunE.LefebvreC.RadureauA.SaganG.LefeuvreJ.-C. (2002). Can thin-lipped mullet directly exploit the primary and detritic production of European macrotidal salt marshes?Estuar. Coast. Shelf Sci.54729–736. 10.1006/ecss.2001.0855
28
LaffailleP.LefeuvreJ.-C.SchrickeM.-T.FeunteunE. (2001). Feeding ecology of 0-group sea bass, Dicentrarchus labrax, in salt marshes of Mont Saint Michel Bay (France).Estuaries24116–125. 10.2307/1352818
29
LaffailleP.PétillonJ.ParlierE.ValéryL.YsnelF.RadureauA.et al (2005). Does the invasive plant Elymus athericus modify fish diet in tidal salt marshes?Estuar. Coast. Shelf Sci.65739–746. 10.1016/j.ecss.2005.07.023
30
LebretonB.RichardP.ParlierE. P.GuillouG.BlanchardG. F. (2011). Trophic ecology of mullets during their spring migration in a European saltmarsh: a stable isotope study.Estuar. Coast. Shelf Sci.91502–510. 10.1016/j.ecss.2010.12.001
31
LefeuvreJ.-C.BouchardV.FeunteunE.GrareS.LaffailleP.RadureauA. (2000). European salt marshes diversity and functioning: the case study of the Mont Saint-Michel bay, France.Wetl. Ecol. Manag.8147–161.
32
LesserJ. S.JamesW. R.StallingsC. D.WilsonR. M.NelsonJ. A. (2020). Trophic niche size and overlap decreases with increasing ecosystem productivity.Oikos1291303–1313. 10.1111/oik.07026
33
MaZ.GanX.CaiY.ChenJ.LiB. (2011). Effects of exotic Spartina alterniflora on the habitat patch associations of breeding saltmarsh birds at Chongming Dongtan in the Yangtze River estuary, China.Biol. Invasions131673–1686. 10.1007/s10530-010-9924-3
34
MantzoukiE.YsnelF.CarpentierA.PétillonJ. (2012). Accuracy of pitfall traps for monitoring populations of the amphipod Orchestia gammarella (Pallas 1766) in saltmarshes.Estuar. Coast. Shelf Sci.113314–316. 10.1016/j.ecss.2012.07.022
35
McLuskyD. S.ElliottM. (2004). The Estuarine Ecosystem: Ecology, Threats and Management.Oxford: OUP.
36
NeiraC.GrosholzE. D.LevinL. A.BlakeR. (2006). Mechanisms generating modification of benthos following tidal flat invasion by a Spartina hybrid.Ecol. Appl.161391–1404.
37
NelsonJ. A.JohnsonD. S.DeeganL. A.SpivakA. C.SommerN. R. (2019). Feedbacks between nutrient enrichment and geomorphology alter bottom-up control on food webs.Ecosystems22229–242. 10.1007/s10021-018-0265-x
38
NunnA. D.Clifton-DeyD.CowxI. G. (2016). Managed realignment for habitat compensation: use of a new intertidal habitat by fishes.Ecol. Eng.8771–79. 10.1016/j.ecoleng.2015.11.009
39
OdumE. P. (1980). “The status of three ecosystem-level hypotheses regarding salt marsh esturaries: tidal subsidy, outwelling, and detritus-based food chains,” in Estuarine Perspectives, ed.KennedyV. S. (Amsterdam: Elsevier), 485–495. 10.1016/B978-0-12-404060-1.50045-9
40
PétillonJ.ErfanzadehR.GarbuttA.HoffmannM.MaelfaitJ.-P.HoffmannM. (2010). Inundation frequency determines the post-pioneer successional pathway in a newly created salt marsh.Wetlands301097–1105. 10.1007/s13157-010-0115-x
41
PétillonJ.GeorgesA.CanardA.YsnelF. (2007). Impact of cutting and sheep grazing on ground–active spiders and carabids in intertidal salt marshes (Western France).Anim. Biodivers. Conserv.30201–209.
42
PétillonJ.YsnelF.CanardA.LefeuvreJ. (2005). Impact of an invasive plant (Elymus athericus) on the conservation value of tidal salt marshes in western France and implications for management: responses of spider populations.Biol. Conserv.126103–117. 10.1016/j.biocon.2005.05.003
43
PinnegarJ. K.PoluninN. V. C. (1999). Differential fractionation of δ13C and δ15N among fish tissues: implications for the study of trophic interactions.Funct. Ecol.13225–231. 10.1046/j.1365-2435.1999.00301.x
44
PostD. M.LaymanC. A.ArringtonD. A.TakimotoG.QuattrochiJ.MontañaC. G. (2007). Getting to the fat of the matter: models, methods and assumptions for dealing with lipids in stable isotope analyses.Oecologia152179–189. 10.1007/s00442-006-0630-x
45
PowerM. E.TilmanD.EstesJ. A.MengeB. A.BondW. J.MillsL. S.et al (1996). Challenges in the quest for keystones: identifying keystone species is difficult—but essential to understanding how loss of species will affect ecosystems.BioScience46609–620. 10.2307/1312990
46
Quezada-RomegialliC.JacksonA. L.HarrodC. (2019). tRophicPosition: Bayesian Trophic Position Calculation with Stable Isotopes.Available online at:https://CRAN.R-project.org/package=tRophicPosition(January 27, 2020).
47
R Core Team (2020). R: A Language and Environment for Statistical Computing.Vienna: R Foundation for Statistical Computing.
48
RezekR. J.MassieJ. A.NelsonJ. A.SantosR. O.ViaderoN. M.BoucekR. E.et al (2020). Individual consumer movement mediates food web coupling across a coastal ecosystem.Ecosphere11:e03305. 10.1002/ecs2.3305
49
RountreeR. A.AbleK. W. (1992). Foraging habits, growth, and temporal patterns of salt-marsh creek habitat use by young-of-year summer flounder in New Jersey.Trans. Am. Fish. Soc.121765–776. 10.1577/1548-8659(1992)121<0765:FHGATP>2.3.CO;2
50
SchramaM.BergM. P.OlffH. (2012). Ecosystem assembly rules: the interplay of green and brown webs during salt marsh succession.Ecology932353–2364. 10.1890/11-1102.1
51
SelleslaghJ.BlanchetH.BacheletG.LobryJ. (2015). Feeding habitats, connectivity and origin of organic matter supporting fish populations in an estuary with a reduced intertidal area assessed by stable isotope analysis.Estuaries Coasts381431–1447. 10.1007/s12237-014-9911-5
52
SheavesM.BakerR.NagelkerkenI.ConnollyR. M. (2015). True value of estuarine and coastal nurseries for fish: incorporating complexity and dynamics.Estuaries Coasts38401–414. 10.1007/s12237-014-9846-x
53
SillimanB. R.BertnessM. D. (2004). Shoreline development drives invasion of Phragmites australis and the loss of plant diversity on new england salt marshes.Conserv. Biol.181424–1434. 10.1111/j.1523-1739.2004.00112.x
54
StockB. C.SemmensB. X. (2013). MixSIAR GUI User Manual. Version 3.1.10.5281/zenodo.56159
55
TrancartT.LambertP.RochardE.DaveratF.CoustillasJ.RoqueploC. (2012). Alternative flood tide transport tactics in catadromous species: Anguilla anguilla, Liza ramada and Platichthys flesus.Estuar. Coast. Shelf Sci.99191–198. 10.1016/j.ecss.2011.12.032
56
ValéryL.RadureauA.LefeuvreJ.-C. (2017). Spread of the native grass Elymus athericus in salt marshes of Mont-Saint-Michel bay as an unusual case of coastal eutrophication.J. Coast. Conserv.21421–433. 10.1007/s11852-016-0450-z
57
van KlinkR.van der PlasF.van NoordwijkC. G. E.WallisDeVriesM. F.OlffH. (2014). Effects of large herbivores on grassland arthropod diversity.Biol. Rev.90347–366. 10.1111/brv.12113
58
VinagreC.SalgadoJ.CabralH. N.CostaM. J. (2011). Food web structure and habitat connectivity in fish estuarine nurseries—impact of river flow.Estuaries Coasts34663–674. 10.1007/s12237-010-9315-0
59
WangM.GaoX.WangW. (2014). Differences in burrow morphology of crabs between Spartina alterniflora marsh and mangrove habitats.Ecol. Eng.69213–219. 10.1016/j.ecoleng.2014.03.096
60
WarrenR. S.FellP. E.RozsaR.BrawleyA. H.OrstedA. C.OlsonE. T.et al (2002). Salt marsh restoration in Connecticut: 20 years of science and management.Restor. Ecol.10497–513. 10.1046/j.1526-100X.2002.01031.x
61
WhitcraftC. R.LevinL. A.TalleyD.CrooksJ. A. (2008). Utilization of invasive tamarisk by salt marsh consumers.Oecologia158259–272. 10.1007/s00442-008-1144-5
62
WuY.-T.WangC.-H.ZhangX.-D.ZhaoB.JiangL.-F.ChenJ.-K.et al (2009). Effects of saltmarsh invasion by Spartina alterniflora on arthropod community structure and diets.Biol. Invasions11635–649. 10.1007/s10530-008-9279-1
63
YangG.-Q.ChenY.-M.WangJ.-P.GuoC.ZhaoL.WangX.-Y.et al (2016). Development of a universal and simplified ddRAD library preparation approach for SNP discovery and genotyping in angiosperm plants.Plant Methods12:39. 10.1186/s13007-016-0139-1
64
YinS.AnS.DengQ.ZhangJ.JiH.ChengX. (2015). Spartina alterniflora invasions impact CH4 and N2O fluxes from a salt marsh in eastern China.Ecol. Eng.81192–199. 10.1016/j.ecoleng.2015.04.044
65
ZieglerS. L.AbleK. W.FodrieF. J. (2019). Dietary shifts across biogeographic scales alter spatial subsidy dynamics.Ecosphere10:e02980. 10.1002/ecs2.2980
Summary
Keywords
diet partitioning, trophic position, fish nursery, Dicentrarchus labrax, Chelon ramada, salt-marsh plants
Citation
Lafage D, Carpentier A, Duhamel S, Dupuy C, Feunteun E, Lesourd S and Pétillon J (2021) Site Characteristics More Than Vegetation Type Influence Food Web Structure of Intertidal Salt Marshes. Front. Mar. Sci. 8:669759. doi: 10.3389/fmars.2021.669759
Received
19 February 2021
Accepted
06 April 2021
Published
07 May 2021
Volume
8 - 2021
Edited by
Dongyan Liu, East China Normal University, China
Reviewed by
Zhenming Ge, East China Normal University, China; Chang-Keun Kang, Gwangju Institute of Science and Technology, South Korea
Updates
Copyright
© 2021 Lafage, Carpentier, Duhamel, Dupuy, Feunteun, Lesourd and Pétillon.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Denis Lafage, lafage.denis@gmail.com
†These authors have contributed equally to this work
This article was submitted to Marine Ecosystem Ecology, a section of the journal Frontiers in Marine Science
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.