 María del Carmen Jiménez-Quiroz1*
María del Carmen Jiménez-Quiroz1* Francisco Javier Barrón-Barraza2
Francisco Javier Barrón-Barraza2 Rafael Cervantes-Duarte3
Rafael Cervantes-Duarte3 René Funes-Rodríguez3
René Funes-Rodríguez3- 1Dirección General de Investigación Pesquera del Pacífico, Instituto Nacional de Pesca y Acuacultura (INAPESCA), Secretaria de Agricultura y Desarrollo Rural (SADER), Cd de México, Mexico
- 2Centro Regional de Investigación Acuícola y Pesquera en La Paz, INAPESCA, SADER, La Paz, Mexico
- 3Instituto Politécnico Nacional, Centro Interdisciplinario de Ciencias Marinas, La Paz, Mexico
This study presents an overview of bivalve assemblages in Bahia Magdalena (BM, México) and the possible impact of environmental variability on these populations, constantly stressed by fishing. This lagoon is responsible for a high proportion of harvest of regional bivalves. First, we list the bivalve species reported in public biogeographic databases. Based on eight commercially exploited species, we described the composition of the bivalve assemblage and its biological characteristics, the history of fishery, and environmental variability in the marine area adjacent to the lagoon (1970–2019) and the habitat of bivalves (2002–2020). Sources of data were public databases and published literature. The enlisted species (n = 184) belong to six orders, and most are small and infaunal, but the structure of the assemblage is unknown. The fisheries began at different times and focused on the most valuable resources. Almost all harvest of bivalves had wide variations because of intensive fishing and a weak regulation frame. After 2015, the main resources were the Pacific wing-oyster (a new resource since 2017) and the geoduck clam due to the declining abundance of other resources (e.g., pen shells, Pacific calico scallop). There was a warming trend in the region since the 1970's, but the strongest El Niño-Southern Oscillation (ENSO) phases caused the most notable changes before 2013; after that year, a combination of large-scale phenomena increased the temperature significantly. The trend of chlorophyll-a abundance negatively correlated with temperature, but there was an almost constant supply of particulate organic matter in the interior of Bahia Magdalena (BM). After 2015, the quality of lagoon water gradually deteriorated, and in 2017 and 2019, harmful algal blooms developed, but the impact was not fully assessed. The challenges faced by the fishery are multiple (institutional weakness and regional warming); however, permanent monitoring programs of environmental conditions and critical biological variables should be implemented to design scenarios that allow fishery sustainability.
Introduction
The diversity of bivalves in the American Tropical Pacific is high, especially between the Gulf of California and Peru, with latitudinal differences (Roy et al., 2000); on the western coast of Baja California, diversity decreases, so at the 24°N parallel, around 200 species have been recorded (Coan and Valentich-Scott, 2012), composed mainly of infauna species (80%). Bivalves provide numerous ecosystem services, including particle removal, carbon fixation, sediment stabilization, and protection against storm surges, among others (Brumbaugh and Toropova, 2008). On the other hand, clams accounted for 2% of the Mexican Pacific fishing catch in 2018 (CONAPESCA home page1), and the Baja California Sur (BCS) state contributed around 70% of the total, and ≈22% of the income of the state from fishing corresponded to bivalves.
The coastal lagoon system Bahia Magdalena-Bahia Almejas (BM-BA) is the most important source of bivalve fisheries from BCS and the Mexican Pacific. It is considered a biological activity center (BAC) because of its high productivity and biodiversity (Lluch-Belda, 2000; Funes-Rodríguez et al., 2007). Fourteen species support fisheries (Baqueiro et al., 1982a); however, the most valuable are the California Venus (Chione californiensis), frilled Venus (Chione undatella), pustulose ark (Anadara tuberculosa), squalid callista (Megapitaria squalida), Pacific calico scallop (Argopecten ventricosus), pen shells (Pinna rugosa, Atrina maura, Atrina oldroydii), and the geoduck clam (Panopea globosa).
Commercial fisheries began between the 1970 and 1990's, except for the geoduck clam, which began in 2005 (Félix-Pico, 2006; Amezcua-Castro, 2014; Magadán-Revelo, 2015). Commercial harvest has had wide variations during its history; however, from 2014 to 2017, almost all species were scarce. Squalid callista and pen shell populations were classified as “overexploited” (DOF, 2017, 2018b), and the Pacific calico scallop reached a low density that the fishery was closed for several years. The primary support of the fishery was the geoduck clam, complemented with Venus clams and pustulose ark, whose value is significantly lower. In 2017, fishermen began to harvest the Pacific-wing oyster to compensate for the declining of most valuable species (Barrón-Barraza, comm. pers.).
One hypothesis to explain the decrease in catch is that the lagoon system has been undergoing significant environmental changes since 2010. The variability of Physico-chemical characteristics and the plankton communities in BM-BA depend on periodic fluctuations of the California and Mexican currents and large-scale climatic processes, such as the El Niño-Southern Oscillation (ENSO) phenomenon (Gárate-Lizárraga and Siqueiros-Beltrones, 1998; Palomares-García et al., 2003; Jiménez-Quiroz et al., 2019). Other large-scale climatic processes, such as the Pacific Decadal Oscillation (PDO) and the North Pacific Gyre Oscillation (NPGO), also affect BM, because they influence the California current and the transport of nutrients from north of the Pacific to Baja California (Di Lorenzo et al., 2008). Marine heatwaves, The Blob (2013–2015), and the 2019 and 2020 North Pacific Marine Heatwaves (NEP) have also disturbed all-region conditions. On the other hand, global temperature has been increasing since the 1970's, and the 2014–2017 span has been the warmest since the measurement of temperature began in the nineteenth century (NOAA, 2021).
The quality of water in BM, described with the trophic index (TRIX, 2015–2019), has seasonal changes. It has low values during winter and autumn, indicating oligotrophic-mesotrophic status, and eutrophic during spring and at the beginning of summer when coastal upwellings are more frequent and stronger (Cervantes-Duarte et al., 2021). However, there was a gradual deterioration in the quality of water over the last 5 years. In 2019, a harmful algal bloom (HAB) covered the adjacent marine region and the interior of the lagoon system. HABs are recurrent phenomena in the NE Pacific; on the western coast of the United States and Latin America, they are common in the El Niño years and affect ecosystems (composition and function), fisheries, and even human health (Müller et al., 2020). However, there is little information on the southern region of Baja California, including BM-BA, because of the absence of a systematic sampling program.
In this context, this study presents an overview of bivalve assemblages in BM and the possible impact of environmental variability on these populations, constantly stressed by fishing. First, we compile a list of species and their biological characteristics in biodiversity databases to show the biological richness of BM; we describe the commercially exploited biological characteristics of species to have insight into the assemblage characteristics of the bivalves. Also, we depict the history of bivalve fisheries, including the regulations of fishing authorities, to illustrate the intensity of the harvest. The environmental characteristics were described from 1970 to 2019 in the oceanic zone and from 2002 to 2020 in the interior of BM to show the variations in temperature and food availability indicators. This theme includes an analysis of the trophic state and water quality from 2015 to 2019 when high temperatures prevailed. Finally, we discussed the impact of fishing and environmental variability on commercial bivalve abundance, and pointed out the importance of monitoring the harvest, biological characteristics, and environmental variability.
Methods
Study Area
The Bahia Magdalena (BM)-Bahia Almejas (BA) system is at the coordinates 24.26–25.75°N, 111.33–112.30°W (Figure 1A) in the southwestern coast of Baja California, a transitional zone between tropical and temperate environments. BM circulation is an anti-estuary type, because it is located in a semi-arid zone and lacks permanent continental drainages. The Channels Zone (CZ; 137 km2) is constituted by several tidal channels lined by mangal, and it communicates BM with the Gulf of Ulloa at the north. BM (883 km2) has two zones according to depth and thermohaline structure: the inner is shallow (<20 m) and occupies approximately half of the surface of the lagoon; it is well-mixed, so the vertical distribution of salinity, temperature, nutrients, and chl-a is homogeneous (Cervantes-Duarte et al., 2010; Zaitsev et al., 2010). The deeper zone (>20 m) communicates with the ocean through an inlet and receives the influence of marine processes as the coastal upwelling (Zaitsev et al., 2010). BA (370 km2) communicates with BM through the La Gaviota channel; this lagoon is shallow and smaller than BM.
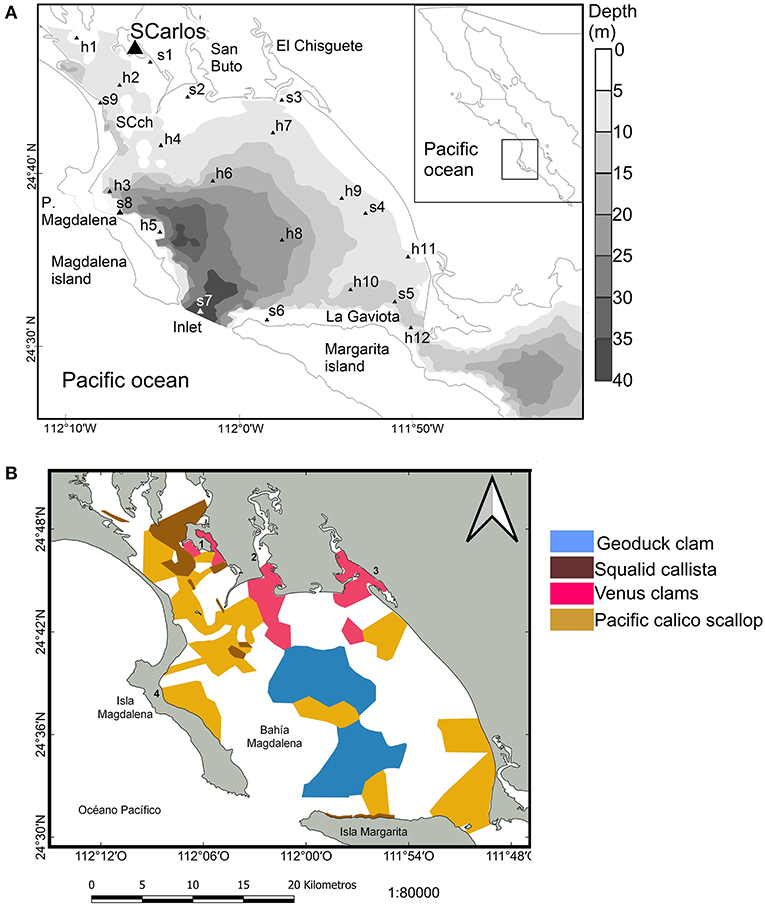
Figure 1. Bahia Magdalena (BM) (A) location and banks of commercial species (B). In (A), the sites used for hydrological and plankton sampling were from s1to s9, while for the habitat analysis made with MODIS-Aqua data, we added the sites h1–h12 for NSST and h3–h12 to chl-a and POC.
Coastal upwellings are the primary source of BM nutrients, and tidal currents advect the cold, nutrient-rich water from the ocean into the lagoon (Zaytsev et al., 2003; Cervantes-Duarte et al., 2010). During spring, upwellings are stronger, so temperature diminishes, and nutrient concentrations (nitrate and phosphate, mainly) increase; ammonium is also higher in this period because of high biological activity. The temperature is high, and upwelling strength and the advection of nutrients are lower in summer and autumn, although it may have strong upwellings in the fall (Cervantes-Duarte et al., 2013). The temperate California Current (winter-spring) and a coastal current from the Mexican tropical Pacific and the Gulf of California (Durazo, 2015; Gómez-Valdivia et al., 2015; summer-fall) influence this region periodically. Global-scale climate processes, such as the ENSO and PDO among others (Sydeman et al., 2014), enhance the tropical or temperate characteristics according to their phase.
Bivalvia Community Characteristics
The list of species was obtained from published literature (Abbott and Dance, 2000), the Global Biodiversity Information Facility (GBIF.org, 2020), the Ocean Biodiversity Information System (OBIS), and the National Commission for the Knowledge and Use of Biodiversity (CONABIO in Spanish) datasets. Information on its size, distribution, and habitat was obtained from the same databases. The length of the bivalve of 156 species obtained from the literature was classified as very small (VS), small (S), medium (M), large (L), and extra-large (XL); percentiles define the superior limit of each class: 0.25 (30 mm), 0.5 (53.4 mm), 0.75 (86 mm), 0.9 (140 mm), and >140 mm.
The biological characteristics (size, reproduction, habitat, and feed-habits) of the commercial species were obtained from several post-grade theses, articles, and other published literature. A map with locations of banks was made with data from Magadán-Revelo (2015), Amezcua-Castro (2014), and Morales-Zárate (2006).
Fisheries History and Trend
Historical time series were inferred from the Fisheries National Charter (DOF, 2018a), published literature (Félix-Pico, 2006; Amezcua-Castro, 2014, among others), the “Información Estadística por Especie y Entidad2” database available at the home page of the Fisheries and Aquaculture National Commission (CONAPESCA in Spanish), in the section from the San Carlos Fisheries Office (Oficina de Pesca de San Carlos), and the Secretary of Fisheries, Aquaculture and Agricultural Development of the state of Baja California Sur. Fishing activity is reported to CONAPESCA authorities by economic units (EUs), including fish production cooperative societies, permit holders, and rural production societies. EU number was used as an indicator of the evolution of fishing effort. On the other hand, data of harvest refer to the whole clam, including shells.
Environmental Characteristics
We analyzed three periods: the first was from 1970 to 2019 to describe the variability of marine conditions at large and local scales, although harvest data were unavailable at the beginning of the series; this span was selected to include several phases of ENSO, PDO, and NPGO. The second one was from 2002 to 2020 to describe the temperature and food availability in the habitats of bivalves, and the third included 2015–2019 to identify trophic state and water quality. The first two were analyzed with remote sensing tools and the third with in situ measurements.
The large-scale marine conditions were described with the climate indices: PDO3, ONI4 (Oceanic El Niño Index), and NPGO.5 The temperature of the marine surroundings of BM-BA was analyzed with the sea surface temperature (SST) Reynolds' reconstructed series6 of the quadrants centered at 23°N, 125°W, and 25°N, 125°W. Standardized temperature anomalies were calculated with the 1961–1990 average and standard deviation. The relationship among SST anomalies with global-scale climate indices, air temperature anomalies in Ciudad Constitución, a city 50 km close to the lagoon (altitude: 50 m), and Bakun Index (BI) anomalies, was described by multiple regression analysis. The trend of SST and residuals was described by simple regression.
The Bakun Index, calculated by the Pacific Fisheries Environmental Laboratory NOAA7 for the 1° quadrant centered in 24°N 113°W, described upwelling strength and seasonality. The trends of the monthly averages and their anomalies were described with a simple regression method. Chlorophyll-a (chl-a) concentration from September 1997 to June 2018 was obtained from the OC-CCI product (v.4) of the European Space Agency (ESA) on monthly compounds with a resolution of 0.04166° (~4 km) from ERDDAP8 database gestor. The quadrant (24.27–24.89°N; 113.02–111.02°W) encompassed 491 pixels. All monthly images were used because the percentage of the image with chla-a values ranged from 85 (September 1997) to 100% (October 2007). Standardized anomalies were calculated with September 1997 to September 2013 average and standard deviation; we did not include posterior months to avoid the bias caused by The Blob and El Niño. The trend in chl-a anomalies from 1970 to 2019 was estimated with a multiple regression equation [R2 = 0.32, F(4,245) = 29.431, p < 0.01].
Chl-a anomalies = −0.0867–0.2218 (SST anomalies) + 0.216 (NPGO) +0.0029 (BI anomalies) −0.0905 (PDO).
An unrotated principal component analysis (PCA) was performed with basin (climate indices) and local scale variable anomalies (SST, chl-a, and BI) from 1998 to 2018 to identify variables that explain the system variance. The PC1 and PC2 factor scores were standardized.
The variability of the bivalves' habitat was characterized using the night SST (NSST) and the chl-a and particulate organic carbon (POC) as food availability indicators. Values in 16 sites for NSST and in 14 sites for chl-a and POC (Figure 1A) were extracted from weekly composite Moderate Resolution Imaging Spectroradiometer (MODIS)-Aqua images from 2002 to 2020 from the Giovanni-NASA database.9 The standardized anomalies were calculated with the mean and standard deviation from the 2003 to 2013 span, and they were used in two Varimax normalized rotated PCAs. The first was done with the data of each site by variable to identify the distribution patterns (zones) according to each PC; the original data of the sites were averaged to describe the seasonal pattern (2003–2013) and time series trend (2002–2020) of each zone. The second PCA was performed with all the data to identify the most relevant variables and sites in the habitat of the bivalves. Marine (from 1970 to −2019 dataset) and BM (inlet vicinity) temperature anomalies from July 2002 to October 2019 were correlated to determine if the former is an indicator of BM variability before 2002. In this analysis, the anomalies were calculated with 10 years (July 2002–December 2013) to be comparable.
The relationship between the bivalves and the environment was analyzed using the monthly harvest data of the squalid callista and the Pacific calico scallop (July 2002–December 2020) with the NSST, chl-a, and POC measured in their habitat (Figure 1B). The data of the Pacific calico scallop (1982–2020) by annual harvest season (expressed as logarithms) were correlated with the average number of strong upwellings (IB > 100 m3 s per 100 m of coastline) by the astronomical season of the same year and the year before. We assumed that the strongest upwellings, the primary fertilization mechanism (Zaytsev et al., 2003), may indicate the introduction of larvae into the lagoon (Morales-Zárate, 2006), and that scallops reach harvest size in ~12 months.
Samples for the hydrological analysis were obtained in monthly campaigns (one per month) carried out at nine sites (s1–s9 in Figure 1), taken at two depths (four sites) and three depths (five sites) during neap tides from January 2015 to December 2019 (Figure 1). Samples for the chl-a analysis were filtered with GF/F fiberglass filters; the content of the filters was used to extract chl-a according to the procedure of Venrick and Hayward (1984). Concentration was estimated with the method by Jeffrey and Humphrey (1975) using a Spectronic Genesys-2 spectrophotometer. Nutrients [nitrite (NO2), nitrate (NO3), phosphate (PO4), and total phosphorus (PT)] were determined using QuikChem 8500 Series 2 Flow Injection Analysis System (Lachat Instruments, Milwaukee, WI, United States); ammonium (NH4) was determined using the method described by Strickland and Parsons (1972). The detection limit of the nutrient analytical methods was 1% precision below 10 μM.
The water quality and trophic level of BM from January 2015 to December 2019 were described with the TRIX index: TRIX = (Log 10 (chl-a × aD% O × DIN × PT) + k)/m:
where chl-a is chlorophyll-a concentration, in μg/L; aD% O is the absolute percentage deviation from oxygen saturation, 100–% OD; DIN is the concentration of dissolved inorganic nitrogen (∑N-NO2, N-NO3, N-NH4), in mg L−1; and PT is total phosphorus, in mg L−1. The constants k = 1.5 and m = 12/10 = 1.2 are scale values introduced to associate TRIX values with eutrophication levels (Barraza-Guardado et al., 2014; Pérez-Ruzafa et al., 2019). The TRIX values range from 0 to 10 and define four trophic status levels: 0–4 (oligotrophic), 4–5 (mesotrophic), 5–6 (mesotrophic to eutrophic), and 6–10 (eutrophic).
The distribution of TRIX into BM was described by hierarchical cluster analysis (CA), carried out with the unweighted pair group average method using arithmetic averages. We also performed a nonmetric two-axis multidimensional scaling analysis (MSA). The analyses showed two sets of data; the first is compound by internal sites (s1, s2, s3, s9, and s5) and the other by the sites closest to the inlet (s6, s7, and s8), so each set of data was averaged to describe its trend. All the statistical analyses were performed with Statistica v.8 (StatSoft, 2007; Statsoft, Tulsa, OK, United States).
The history of harmful algae blooms in BM-BA was obtained from Gárate-Lizárraga et al. (2016) and Jiménez-Quiroz et al. (2019). There were also systematic campaigns to collect phytoplankton that developed simultaneously to hydrological sampling in nine sites (s1–s9) from 2015 to 2018, while in 2019, it was done in June, during HAB occurrence. The sampling and processing methods are described in Jiménez-Quiroz et al. (2019).
Results
Biological Characteristics of the Mollusks From Bahia Magdalena
The GBIF, OBIS, and CONABIO databases enlist 184 species of bivalves belonging to 14 orders and 42 families (Table 1, Supplementary Material), and 78.4% of the species were of the orders Cardiida (cockles), Venerida (clams), Ostreida (oysters), Mytilida (mussels), Arcida (arks), and Pectinida (scallops). The classification by size of 156 species showed that very small bivalves (<30 mm) were a little more abundant (26.9%) than the small (23.08%), medium (25.6%), large (17.31%), and extra-large ones (7.05%).
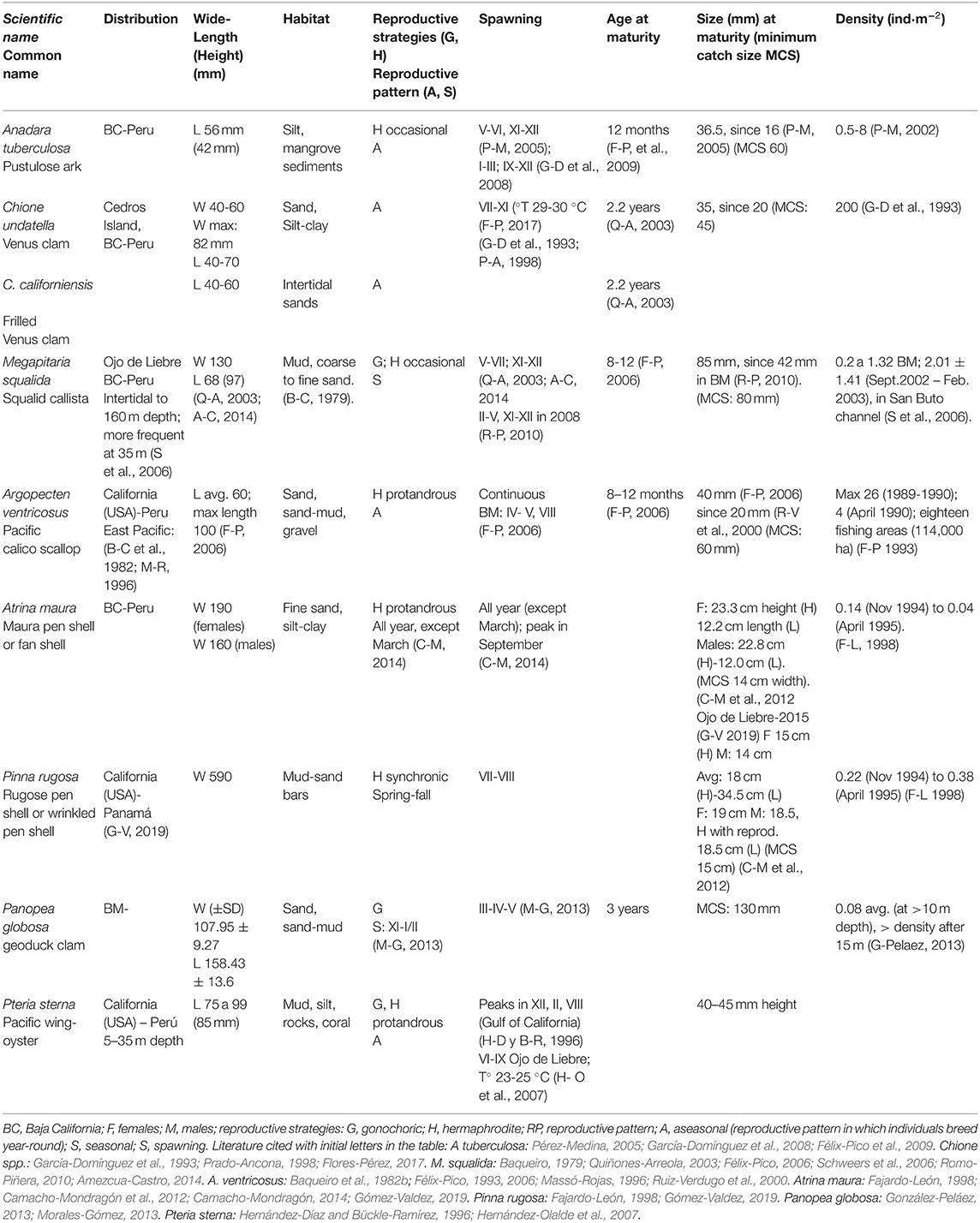
Table 1. Biological characteristics of commercial bivalves from Bahia Magdalena-Bahia Almejas (BM-BA), BCS, México.
There are no reports on the community structure of mollusks and most biological characteristics of BM-BA bivalves. However, according to the literature and databases, the feeding strategies of 127 species were suspension feeders (84%), deposit feeders (12%), and chemiosymbiosis (4%). The main ecomorphology guild was infaunal (58%), followed by epifaunal (27.17%), semi-infaunal (8.15%), endolithic, (4.34%), and 2% undefined. Most species are suspension feeders, regardless of their size and habitat (Figure 1, Supplementary Material). All the epifaunal species are suspension feeders, while among the infaunal, there are some deposit feeders, mainly of larger species than medium size. Among the smaller species, there is a greater diversity of both habitats and types of food.
The commercial bivalves, a sample of medium to extra-large species, inhabit different habitats (Table 1). Infaunal species such as Chione spp., A. tuberculosa, M. squalida, and P. globosa are buried in soft bottoms (Schweers et al., 2006; Amezcua-Castro, 2014). The smaller clams are in shallow areas (1–5 m depth) at depths lesser than 30 cm into the sediments (Chávez-Villalba et al., 2019) in the vicinity of channels (Figure 1B). Squalid callista often forms large banks with Megapitaria aurantiaca and Dosinia ponderosa (Baqueiro et al., 1982a); both are large species and commercially harvested. Geoduck clam is buried from 60 to 100 cm in the central part of BM (10–20 m depth), where the temperature ranges from 18 to 22°C because of the entrance of ocean waters through the inlet. Semi-infaunal species P. rugosa and A. maura live in the intertidal zone, while epifaunal species, such as A. ventricosus, live in a greater variety of substrates, but their banks are in protected areas (Baqueiro et al., 1982a; Massó-Rojas, 1996). Scallops appear in pulses in BM and can move great distances during their juvenile stage carried by currents and contracting their valves. One hypothesis establishes that larvae are transported by tidal currents and wind forcing currents (strong westerly winds) into the lagoon system from banks outside BM-BA. Juveniles settle in seagrass beds (Maeda-Martínez et al., 2000; Morales-Zárate, 2006).
Most species are gonochoric, but the pen shells (A. maura, P. rugosa) and A. ventricosus are functional hermaphrodites. A maura is a protandric species that presents sexual reversion after 18 cm of shell height is reached. A. tuberculosa and M. squalida are casual hermaphrodites. The age of sexual maturity is around 1 or 2 years for all the species, except geoduck (3 years), while the size of sexual maturity is variable and related to environmental conditions (Table 1). A. tuberculosa and Chione spp. spawn all year-round, but in the former, there are two peaks (black band in Figure 2A), and in the latter a long peak from July to November (black band Figure 2B); spawning peaks are related with temperature and food availability (García-Domínguez et al., 2008; Flores-Pérez, 2017). M. squalida spawning is all-year-round, with peaks in different months depending on the year (Table 1; black band Figure 2C). A. ventricosus is a semelparous species (Baqueiro and Aldana-Aranda, 2003), and its spawning is continuous, with peaks in spring (Figure 2D), correlated with temperature and the seston characteristics (Félix-Pico, 2006).
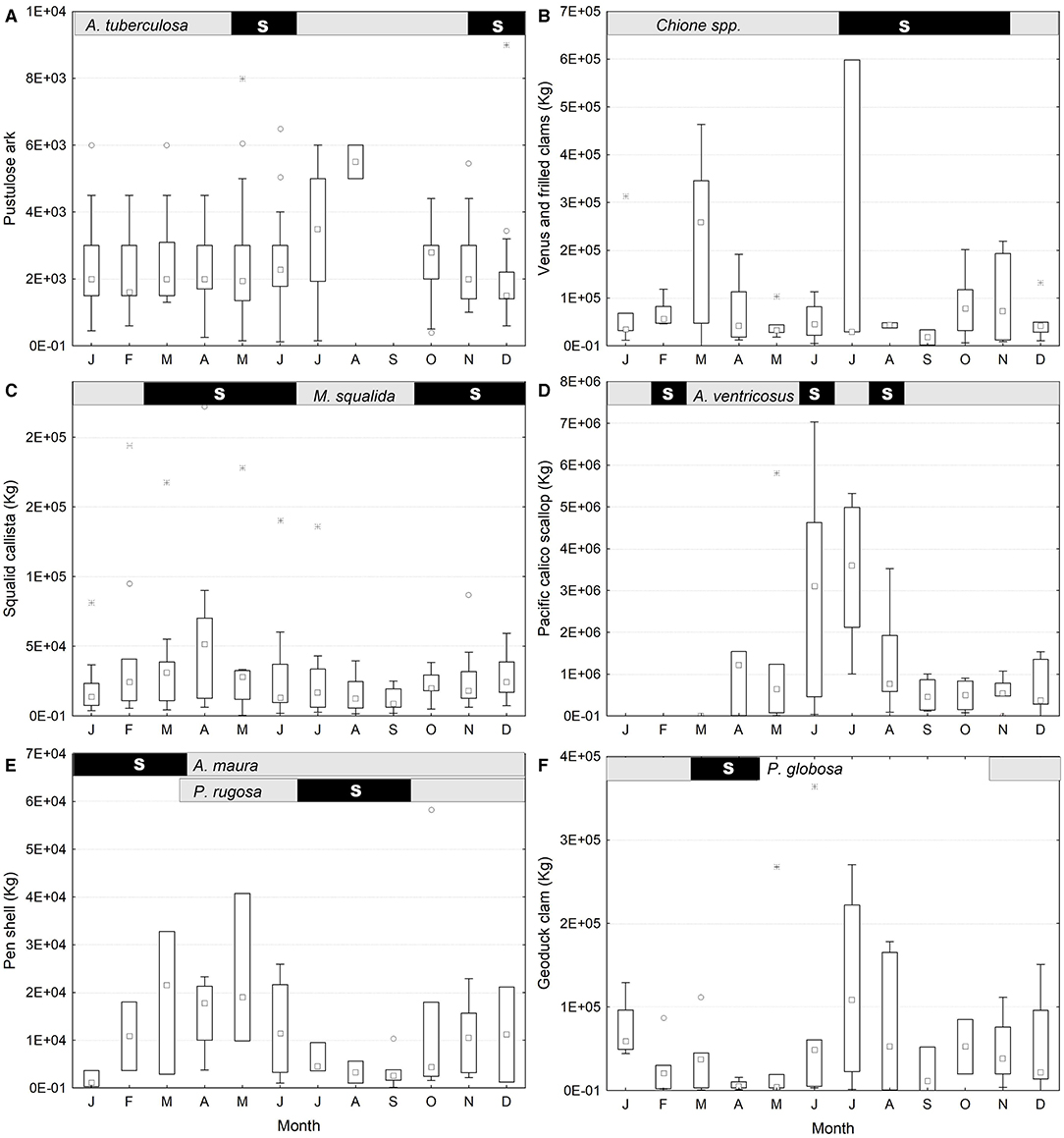
Figure 2. Seasonal variations in harvest (period: 2006–2014) and reproductive season (upper bar gray). The spawning months (black) were drawn according to *Pérez-Medina, 2005, **García-Domínguez et al., 2008 (Pustulose ark: Anadara tuberculosa); Flores-Pérez, 2017 (Venus and frilled clams: Venus spp.); Amezcua-Castro, 2014 (callista squalida: Megapitaria squalida); Félix-Pico, 2006 (Pacific calico scallop: Argopecten ventricosus); Camacho-Mondragón, 2014; Gómez-Valdivia et al., 2015 (Pen shells: Atrina maura, Pinna rugosa); Morales-Gómez, 2013 (Geoduck clam: Panopea globosa). Box graphs represent the 25–50 quartiles, whiskers the non-outlier data, and central squares the median. (A) Pustulose ark (A. tuberculosa); (B) Venus and frilled clams (Chione spp.); (C) Squalid callista (M. squalida); (D) Pacific calico scallop (A. ventricosus); (E) Pen shells (A. maura, P. rugosa); (F) Geoduck clam (P. globosa).
The reproduction of A. maura depends on temperature and food availability. Spawning is all-year-round but is higher during the cold season (Table 1; Figure 2E). Gonadal atresia is continuous after 25°C, but it also depends on the size of individuals, so it occurs in males at the smallest sizes, females in the biggest, and hermaphrodite in intermedia sizes organisms (Camacho-Mondragón, 2014). The reproductive cycle of P. globose begins in November (Figure 2F), with maturation peaks in the January–February and March–April spawning periods (Cruz-Gómez, 2012; González-Peláez, 2013). Most of the clams are 7 years old (Larios-Castro, 2020), although there are some that are older (20–47 years), according to González-Peláez (2013). Growth is very fast in the first 10 years and reaches to asymptotic phase around 20 years. The asymptotic growth of the shell is between 167.5 (González-Peláez et al., 2015) and 179.8 mm (Luquin-Covarrubias et al., 2016). Recruitment is in irregular pulses.
Clam Fisheries in Bahia Magdalena: Composition, Seasonality, and Historical Trend
All bivalve species support artisanal and recreational fisheries. According to the San Carlos Fisheries Office data (2006–2014), bivalves were the second resource in importance recorded by weight, just surpassed by sardines (73.3%), an industrial fishery; in contrast, by economic value ($), bivalves represented 25.8%, surpassing sardines by 2%, shrimp (10%), and finfish (16.1%). A field study showed that the fishery of bivalves is the third most important after shrimp and finfish fisheries (Magadán-Revelo, 2015). However, while the shrimp season lasts around 6 months (September–February) and the finfish all-year-round, with a peak from June to August (Cavieses-Nuñez, 2016), the harvest of geoduck clam occurs in periods of 15 days, distributed throughout the year, so it is much more profitable than the other two (Magadán-Revelo, 2015).
Among bivalves, the Venus and frilled Venus clams (Chione spp.) were second in importance by volume, but by price, the most valuable resources were Pacific calico scallop (A. ventricosus), pen shells (P. rugosa, A. maura), and the geoduck clam (P. globosa). Squalid callista (M. squalida) is important, because it was harvested all year (Magadán-Revelo, 2015) until recent years.
There are no fishing effort records; however, according to the Fisheries National Charter (DOF, 2018a), the number of permits (p) in BM to harvest Venus and frilled Venus clams was 9, which comprises 17 minor vessels (mv). There is no recent information on squalid callista because 50% of the number reported in the Fisheries National Charter corresponded to both BM and Ojo de Liebre lagoons (74 p:157 mv). Information is also unclear on Pacific calico scallop, but there are 144 p and 354 mv (Osuna-Castro, 2012), and 100 p (165 mv) to harvest pen shells (DOF, 2018a). The number of permits to harvest geoduck clam is 143 (166 mv), with the register currently closed (DOF, 2018a). On the other hand, the number of economic units (EU), another indicator of fishing effort, increased significantly from 2001 (77) to 2016 (165). The most significant increase occurred between 2006 and 2009. In parallel to the authorized capture, large quantities are illegally harvested by an unknown number of artisanal fishermen with very few resources, locally called “Patos buzos” (diver ducks) and “guateros,” smugglers with human and material infrastructures to efficiently harvest bivalves, mainly during the night (Magadán-Revelo, 2015).
Bivalves are collected by Hookah-diving up to 10 m deep. A fishing team has three members: a diver, a lifeline operator, and an outboard motor operator (DOF, 2018a). Gathering is done by hand, sometimes with a trident to remove sediment, substituted by a motor pump in geoduck clam harvest (DOF, 2015a). Gathering in shallow areas at low tides is common to harvest Venus clams, pustulose ark, and squalid callistas for subsistence consumption (Schweers et al., 2006).
The harvest of almost all species occurs all-year-round, but there are distinct peaks (Figure 2); some of them coincided with spawning peaks (black bands on the top of each graph). However, in almost all species, the lowest harvest occurs in September and October. In the analyzed database, the pustulose ark information was scarce; however, its harvest was higher in June–July (Figure 2A) in 1992–2002 (Félix-Pico et al., 2009). Venus and frilled Venus clam gatherings are higher in three periods (March, July, November; Figure 2B). The harvest of squalid callista peaks in April, June, and November–December (Figure 2C), although there may be local differences (Schweers et al., 2006). The peak of Pacific calico scallop harvest is from May to July (Figure 2D), with a closure time from December 15 to March 31 (DOF, 1994). The gathering of pen shells is similar to the that of squalid callista but is more frequent from March to May and October to December (Figure 2E), with low values in January and July-September; in 2021, authorities established a closure from April 1 to July 31 (DOF, 2021). The harvest of geoduck lasted 15 days per year and is closed from January 25 to April 30 (Figure 2F).
The harvest of pustulose ark and Venus clams significantly increased after 2010 (Figures 3A,B), but it was more irregular in the previous years. The intense gathering period of arks was shorter (since 2016) than that of Venus clams (2011–2020). According to comments of fishermen, both species were part of subsistence fisheries. However, the market demand for both clams has increased recently. These resources have no fishing regulations.
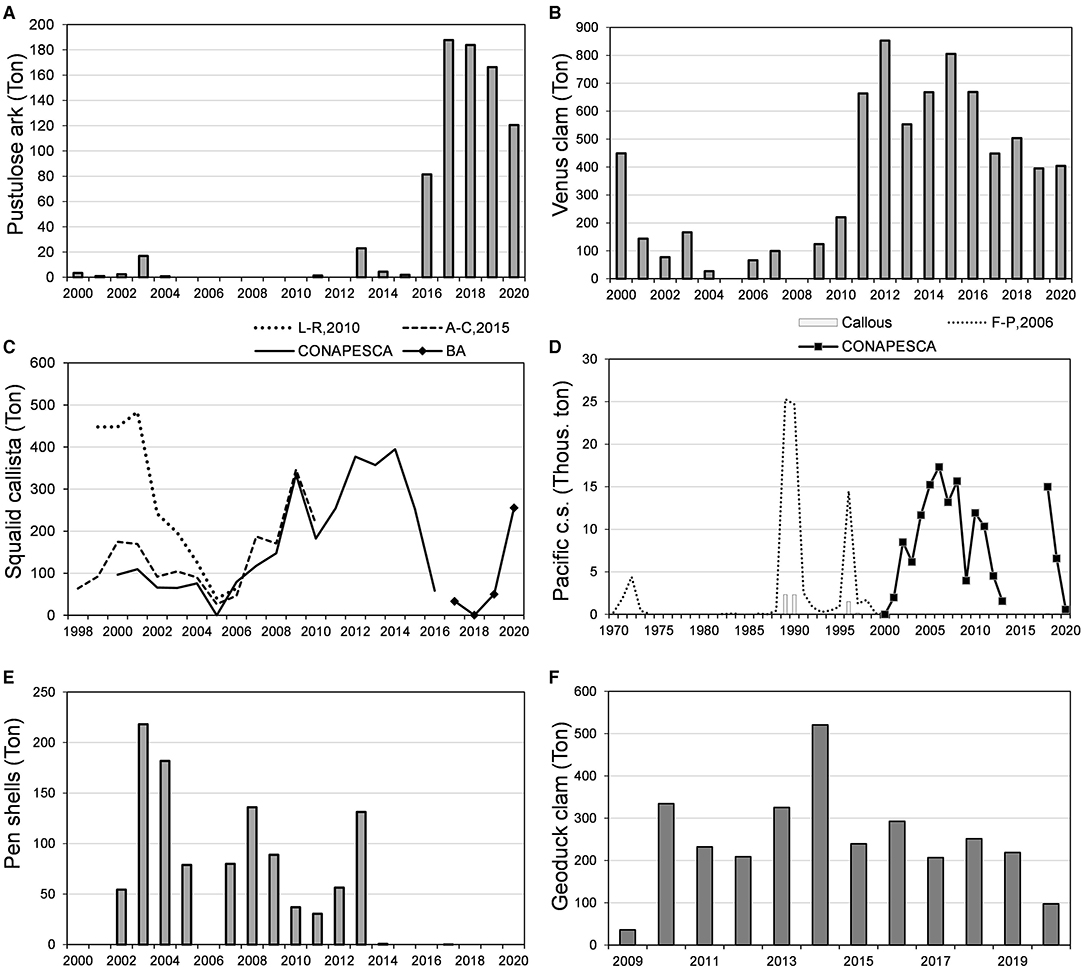
Figure 3. Historical variations in the harvest of clams in BM- Bahia Almejas (BA), recorded by CONAPESCA, and Amezcua-Castro et al. (2015), López-Rocha et al. (2010), and Félix-Pico (2006). (A) Pustulose ark; (B) Venus and frilled clams; (C) Squalid callista; (D) Pacific calico scallop (Pacific c.s.); (E) Pen shells; (F) Geoduck clam. Pacific c.s, Pacific calico scallop.
Bahia Magdalena-Bahia Almejas has been an important contributor to the production of squalid callista (Amezcua-Castro et al., 2015), and the supply in BM (82%) was higher than that in BA (10.6%) and CZ (6.7%). Maximum catches and catch per unit of effort (CPUE) occurred from 1999 to 2001 (López-Rocha et al., 2010), but Amezcua-Castro et al. (2015) and CONAPESCA reported lower values (Figure 3C). Harvest decreased until the minimum was reached in 2005 (catch) and 2006 (CPUE). In this period, the fishing effort (number of vessels) also decreased. On the contrary, from 2006 to 2014, the harvest increased, reaching around 400 tons; after 2014, it fell sharply (Figure 3C) because the average density (abundance) was close to 50% (10%) to the extent, in 2015, of signals of overexploitation of banks (DOF, 2017). In 2019 and 2020, gathering was only done in BA, but it was around 255 tons in the last year. Fishing authorities established the first regulations (quotas and minimum catch size) in 2007; however, the fishing effort increased from 2007 to 2010 (Amezcua-Castro, 2014). The fishery was closed for 2 years in 2017 (DOF, 2017), but while the closure was maintained in BM for another 2 years, it was lifted in BA in 2018.10 During the closure, illegal harvest continued, according to comments of fishermen.
The commercial harvest of Pacific calico scallop began in 1972, with a very high value. There were no records from 1975 to 1982. The 1989 and 1990 harvests were the highest in the time series (Félix-Pico, 2006; DOF, 2010). This species was the most important resource for artisanal fishermen from 1998 to 2009 (Ojeda-Ruiz de la Peña and Ramírez-Rodríguez, 2012). The catch from 1998 to 2008 was high (≈15,000 tons), decreased in 2009 (<5,000 tons), and had a slight rebound in 2010 and 2011, with an average of ≈12,000 tons (DOF, 2018a). Harvest was suspended in 2014 because of low-density values and resumed in 2018 with a volume similar to that in 2008 (Figure 3D). In 1987, there were established rules to scallop harvesting in an Agreement (DOF, 1987), including minimum catch size (60 mm), quotas, fishing equipment, extraction method, and permit conditions to harvest and culture. In 1993, authorities published fishing regulations in the NOM-004-PESC-1993 (DOF, 1993) and the seasonal closure in 1994 (DOF, 1994). In 2015, the NOM was substituted by the NOM-004-SAG/PESC-2015 (DOF, 2015b).
Atrina maura and P. rugosa sustain the fishery of pen shells fishery because A. oldroydii is depleted. Time series data show very variable harvest values, with a decreasing trend since 2003 (Figure 3E), with peaks in 2003–2004 (218–182 tons), 2008 (136 tons), and 2013 (130 tons). There were no evaluations performed in 2014 and 2015 because of low-density values reported by fishermen. In 2017, authorities classified the resource as overexploited and decreed a 2-year closure (DOF, 2018b).
The prospective gathering of P. globosa began in BM in 2002 and commercial fishing between 2004 and 2005 when the first commercial permits were issued (González-Peláez, 2013; Larios-Castro, 2020). The extraction in a regulated manner began in 2009, and there were peaks in 2010 and 2014 (Figure 3F). Harvest oscillated around 200 tons in the subsequent years, except in 2020, because of the low international demand associated with the severe acute respiratory syndrome coronavirus 2 (SARS-Cov-2) pandemic (Figure 3F); regulations include fishing methods and quotas determined by density evaluations (DOF, 2018a). On the other hand, the harvest of Pacific-wing oyster began in 2017 with no regulations; in 2020, fishermen reported 259,355 tons of this clam (US $146, 937.5), and authorities gave the first permits.
Evolution of the Climate Characteristics at a Large and Synoptic Scale From 1970 to 2019
Time series of the global-scale indices (ONI and PDO) showed that the 1980's, 1990's, and especially 2014–2018 were warming periods, and that the opposite occurred in the 1970's and the first decade of the 2000's. The impact of these phenomena on the productivity and biodiversity of marine ecosystems in Baja California, including BM-BA, is well-documented. Warming (cooling) phases diminish (increase) coastal upwellings, deepen (shoaled) thermoclines and chemoclines, modifying the composition of biological communities (Gárate-Lizárraga et al., 2001; Palomares-García et al., 2003; Jiménez-Quiroz et al., 2019). The NPGO was positive in four periods, which coincided with the negative values of PDO and La Niña. In contrast, the index was negative during El Niño and the positive phase of the PDO. It was correlated with the PDO and ONI (r = −0.54).
The trend of SST and its anomalies in BM-BA (23–25°N; 113°W) has been positive (p < 0.05), since 1970 (Figure 4A). A high percentage of the variance in SST anomalies (R2 = 0.48) was related to large-scale indices, local air temperature (AT) anomalies, and BI. The main contributors to the determination coefficient (p < 0.01) were ONI (0.32), AT (0.12), and PDO (0.039). The trend of residuals trend was also positive, and the slope (m = 0.002) indicated the warming in the region during the study period. The extreme positive anomalies (>3) coincided with the positive phase of PDO, extreme El Niño, and The Blob, and the negative (≈-2) with the negative phase of PDO and La Niña.
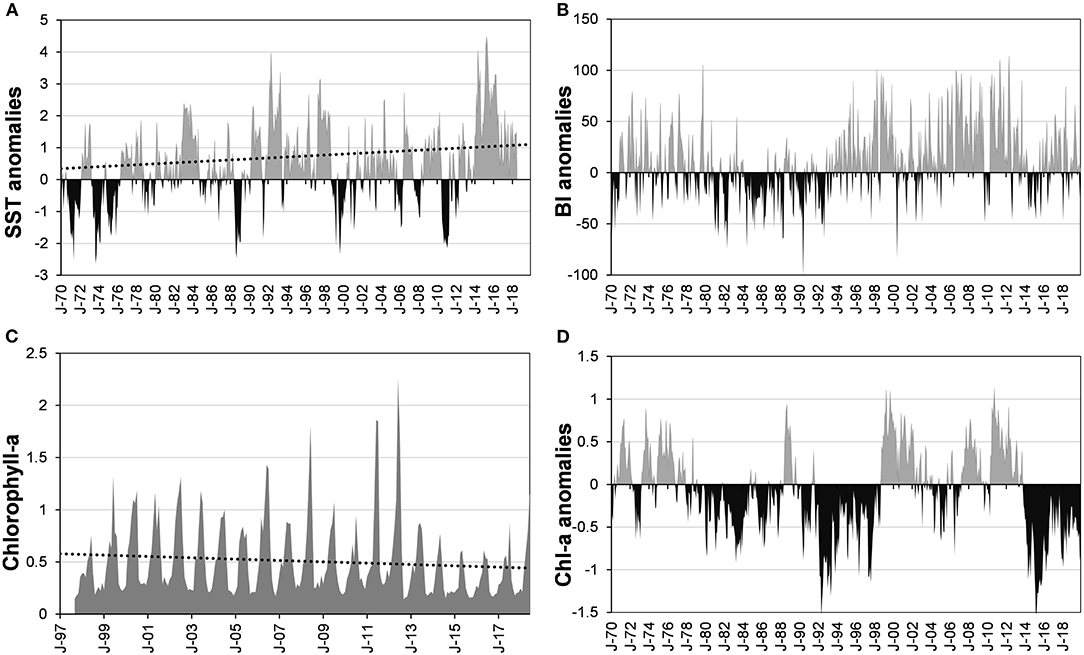
Figure 4. Time series of variables measured off BM-BA. (A) SST anomalies and its trend (y =-0.182 + 0.002x; R2 = 0.09, p < 0.01, N = 582); (B) Bakun Index (BI) anomalies; (C) chl-a 1998 to 2018 (y = 0.573–0.0003x; R2 = 0.01; p = 0.08; n = 250); (D) chl-a anomalies estimated by multiple linear regression. Source: C-CCI product (v.4) ERDDAP Database. J, January.
The frequency and strength of upwellings, the primary source of BM nutrients, showed wide variations throughout the period of study (Figure 4B). In the 1970's, the number of positive and negative monthly anomalies of the Bakun Index (BI) was similar. From 1980 to 1992, negative values were more frequent, but after that year, upwellings gradually strengthened and were very strong between 1994 and 1995, 1999 and 2000, and 2003 and 2011. Between 2013 and 2015, negative anomalies were common, while after 2016 they returned to average. Chlorophyll-a (chl-a) varied seasonally, with a peak from April to July in the marine zone off BM-BA. From 1998 to 2013, mean chl-a concentrations during those months ranged from 0.75 to 1.3 mg m−3, with peaks in 2008, 2011, and 2012 (Figure 4C). After 2013, concentrations diminished until 2016, when they increased again. The anomalies varied from −1 to +2.5, with relatively short periods with negative values from 1998 to 2012. However, in 2005 and 2009, and from 2012 to 2018, negative anomalies were frequent, and in the last period, there were 57 uninterrupted months of negative values.
The variability in chl-a concentration from 1998 to 2018 was related to local and large-scale variables (R2 = 0.32, p < 0.01). The contribution of each variable to the determination coefficient (p < 0.01) was: SST anomalies (0.24), NPGO (0.06), BI anomalies (0.014), and PDO (0.006). Temperature had a negative sign, while NPGO and BI had a positive sign. Chl-a anomaly values were negative during a more extended span rather than positive (Figure 4D) but mostly into the normal interval (−0.5 to 0). However, anomalies lower than −0.5 prevailed during the warming periods. In contrast, from 1970 to 1977 and from 1999 to 2013, chl-a anomalies were normal (0 to +0.5) or high (≈1), coinciding with La Niña and NPGO positive phase.
The first three components of the PCA (1998–2018) explained 78.9% of the variance (Table 2). The factor loadings of the first PC (49% explained variance) showed the inverse relationship of temperature at local (SST) and large-scale (ONI, PDO) with chl-a concentration and NPGO. The standardized PC1 graph shows that positive and normal values (0 to 0.5) prevailed from October 1998 to October 2013 (Figure 5A). Negative values were common between 1997 and 1998, and from March 2014 to July 2016, coinciding with the anomalous warming caused by El Niño and The Blob. This graph is very similar to chl-a anomalies, highlighting the importance of temperature in its abundance. The highest factor loadings in the PC2 (16% explained the variance) were from the NPGO (−0.66) and BI anomalies (+0.62), with chl-a closer to the first one, suggesting that these variables are positively related. The PC2-standardized graph suggests that the effect NPGOs was more significant during La Niña years and upwellings the rest of the years (Figure 5B).
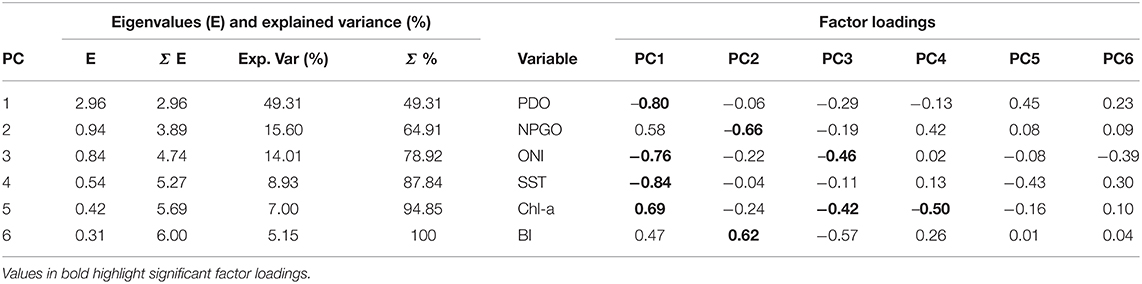
Table 2. Eigenvalues and factor loadings of principal component analysis (PCA) applied to climate indices, sea surface temperature (SST), and chlorophyll-a (chl-a) anomalies off BM-BA, and Bakun Index (BI) anomalies.
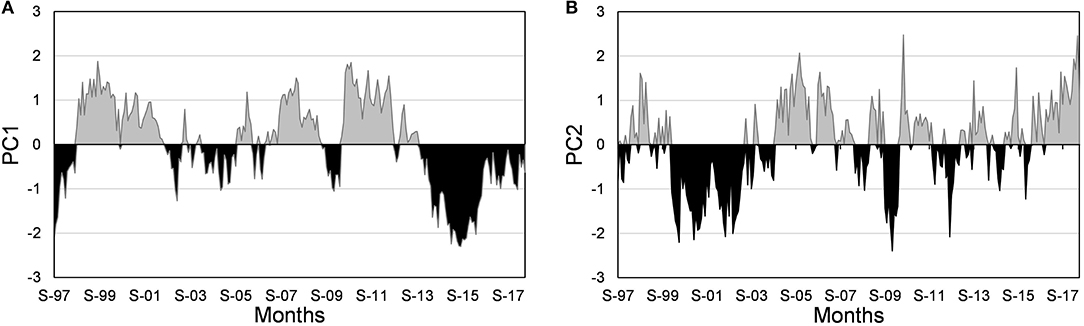
Figure 5. Factor scores standardized of PC1 (A) and PC2 (B). S, September.
Temperature and Food Availability in Habitats of Clams (2002–2019)
The PCA allowed identifying zones inside BM according to the factor loading of sites in each PC. We selected the PCs that explained ≈80% of the variance. The first five PCs of NSST explained 96.6% of the variance, and PC1 explained 89.11%. The factor loadings of the rotated PCA (Table 3) show four zones in the lagoon: the western shore-inlet (WS-I), San Carlos channel, mouth of the northeastern and southern channels (El Chisguete and La Gaviota, respectively), and eastern-southwestern shore (Figure 6A). The lowest temperatures and anomalies were recorded in 2005 and 2011, and the highest were from 2014 to 2016 (Figures 6B,C).
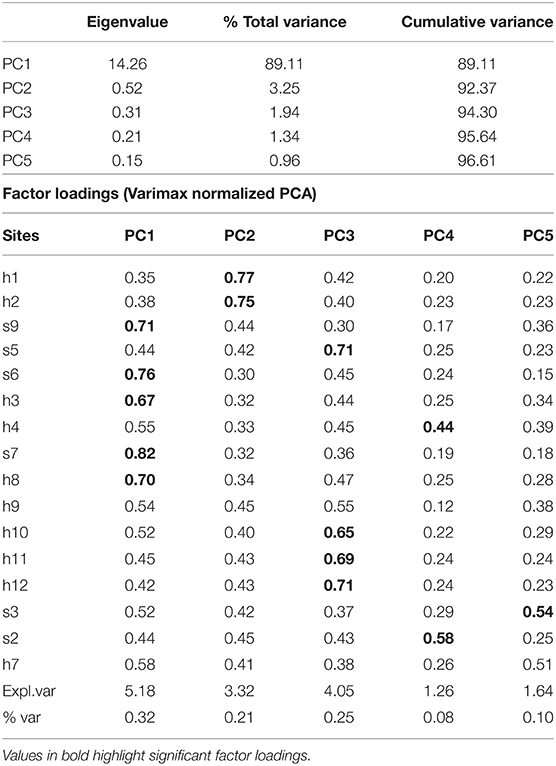
Table 3. Eigenvalues and factor loadings of the rotated PCA applied to night SST (NSST) anomalies recorded in BM.
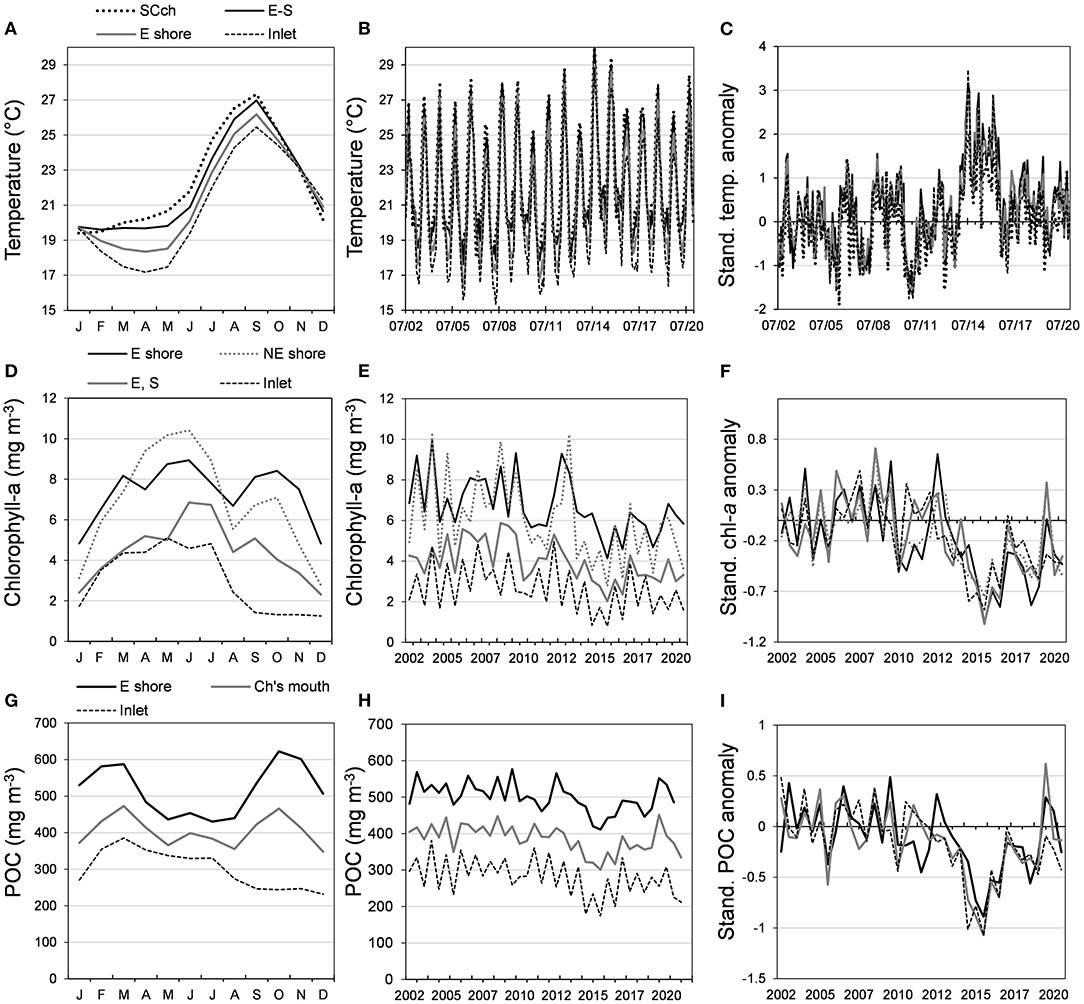
Figure 6. (A–C) Night sea surface temperature, (D–F) chl-a, and (G–I) POC. The left column shows the seasonal pattern of BM zones identified with the rotated principal component analysis (PCA); in the middle column are the time series of temperature (monthly data), chl-a (average semester data), and POC (average semester data); in the right column are standardized anomalies. Inlet includes the western shore (WS-I in the text); SCch, San Carlos channel; E, east; S, south; NE ch, northeastern channels (San Buto and El Chisguete); Ch's mouth, channels mouth (San Carlos, San Buto, El Chisguete, and La Gaviota channels).
Night sea surface temperature follows a seasonal pattern: temperate (January to June) and warm (July to November). There are differences among the zones of the lagoon (Figure 6A); the lowest NSST occurs in the inlet and its surroundings and is 3°C colder than in the channels. In summer, the opposite occurs, and the maximum difference is ≈+2°C (September). The trend of time series (Figure 6B) was slightly positive, but it was significant (p = 0.06) only in the mouth of the channels; in contrast, the positive trend of NSST anomalies was significant (p < 0.01) in all the zones, and a higher slope was recorded in the same zone. The trend from the regression residuals from the WS-I zone anomalies vs. climate indices (R2 = 0.4, p < 0.05) was significant, indicating that BM has been warming since 2002. In the same way, the correlation between SST off BM and NSST from WS-I (R = 0.78, p < 0.05) suggests that Reynolds' reconstructed series variations have been representative of BM NSST variability since 1970.
The PC1 applied to the chl-a data represented 55.57% of explained variance, and the first five, 85.59% (Table 4). The analysis showed four lagoon zones (Figure 6D) in the main body of water. The seasonal pattern has two peaks near the mouth of the northeastern channels and the eastern shore (spring and autumn), while in the inlet, the southeastern and south sites, there is only one (spring). The highest values were registered in the mouth of the El Chisguete channel and the eastern shore, while the lowest was in the surroundings of the inlet. The chl-a concentrations before 2012 were oscillating (Figure 6E), but anomalies were in the average span (−0.5 to 0.5). However, from 2013 to 2020, there were more negative anomalies with extreme values (<-1) in the second half of 2015, and in the first half of 2018, the anomalies in the following month oscillated around −0.5, indicating a chl-a deficit (Figure 6F). Positive anomalies were registered in 2019, mainly in the eastern shore and the south.
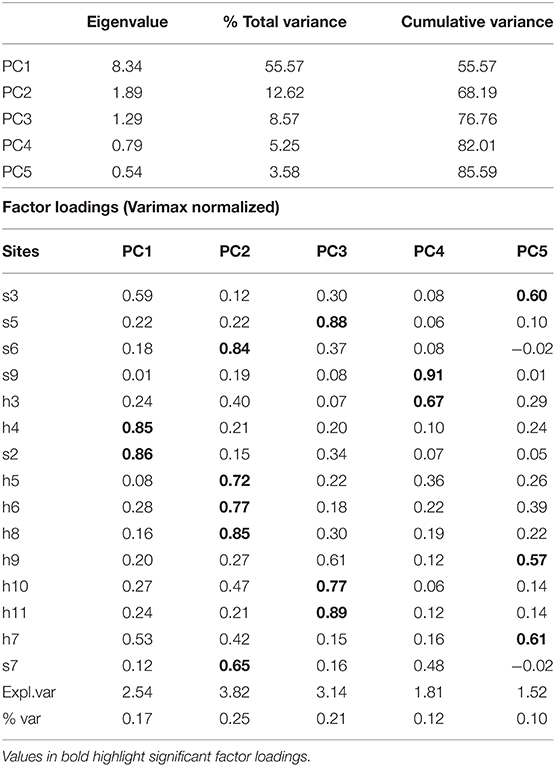
Table 4. Eigenvalues and factor loadings of the rotated PCA applied to chl-a anomalies recorded in BM.
The PC1 explained 50.85% of the variance in particulate organic carbon (POC) and the first five 80.86% (Table 5). The rotated PCA suggests three zones with increasing values: inlet, the mouth of the channel, and eastern shore. Seasonal variations, as the chl-a, follow two trends: in the channels' surroundings and the eastern shore, there are two peaks (winter and autumn) and one in the inlet (February to July); however, unlike the chl-a, the size of both peaks is similar, and the POC is abundant at the inlet for a longer span (Figure 6G). The trend from 2002 to 2020 was decreasing (Figure 6H), but the slope was smaller than chl-a, and anomalies oscillated into the normal interval after 2017 (Figure 6I), indicating a recovery of POC supply.
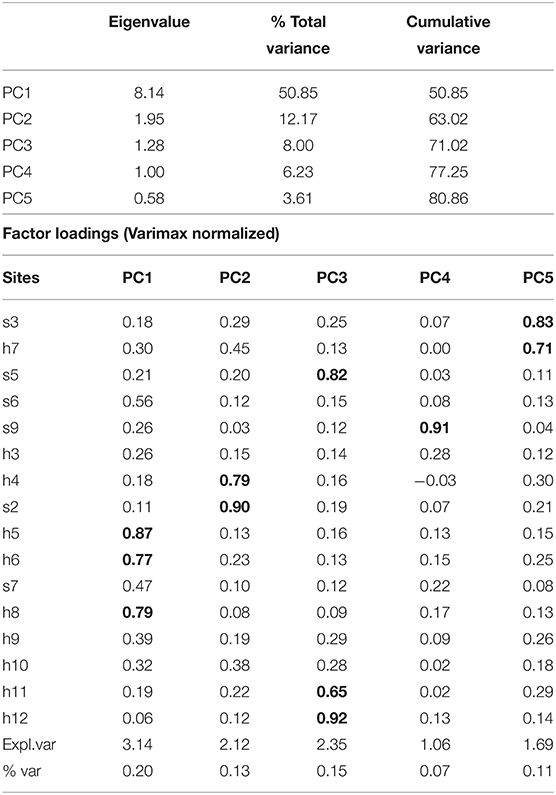
Table 5. Eigenvalues and factor loadings of the rotated PCA applied to particulate organic carbon (POC) anomalies recorded in BM.
The first 10 PCs represented 87.8% of the explained variance of all the variables and sites. In PC1 (31.8%), NSST anomalies had the highest factorial loadings, while in the rest, these corresponded to POC and chl-a measured in different zones of the lagoon: southeastern shore and the surroundings of the La Gaviota channel (PC2: 12.7%), inlet (PC3: 16.2%), eastern shore (PC4: 10.1%), south of the San Carlos channel (PC5: 3.8%). The rest of PCs represented <2% of the explained variance. The factor loadings from rotated PCA highlighted the inverse relationship between NSST and the variables related to food availability (POC and chl-a).
Relationship Between the Environment and Bivalve Populations
The regression of the squalid callista and NSST, chl-a, and POC (2002–2020) from the San Carlos and northeastern channels was significant and negative (R2 = 0.13, p < 0.05), but the explained variance was small, and the most relevant variables were NSST (R2 = 0.11) and chl-a (R2 = 0.033). Pacific calico scallop (1982–2020) was not related to these variables, and only with the average number of strong upwellings of the autumn previous to the harvest season did it show a weak relationship (R2 = 0.1, p = 0.06, n = 30).
Water Quality Inside Bahia Magdalena From 2015 to 2019
Cluster and the non-metric multidimensional analysis applied to the saturation percentage of oxygen (%sat O2) showed two sets: sites near the inlet (s6, s7, and s8) and of the interior of the lagoon (s1, s2, s3, s4, s5, and s9). This variable had a seasonal cycle (Figure 7A); however, in the inlet, it decreased significantly during spring and at the beginning of summer, reaching values ≈50% because the subsurface water upwelled. In contrast, the %sat O2 was high in the interior sites, especially in March and April, and low in September. From 2015 to 2017 (Figure 7B), the mean %sat O2 in both zones oscillated from 50 (inlet) to 70 (inner) to 100 (inlet) to 110% (inner); however, in 2018 and 2019, the percentage decreased significantly (p < 0.01). The decreasing rate, estimated by regression analysis, was greater in the interior (−1.1% per month) than in the inlet (−0.88% per month).
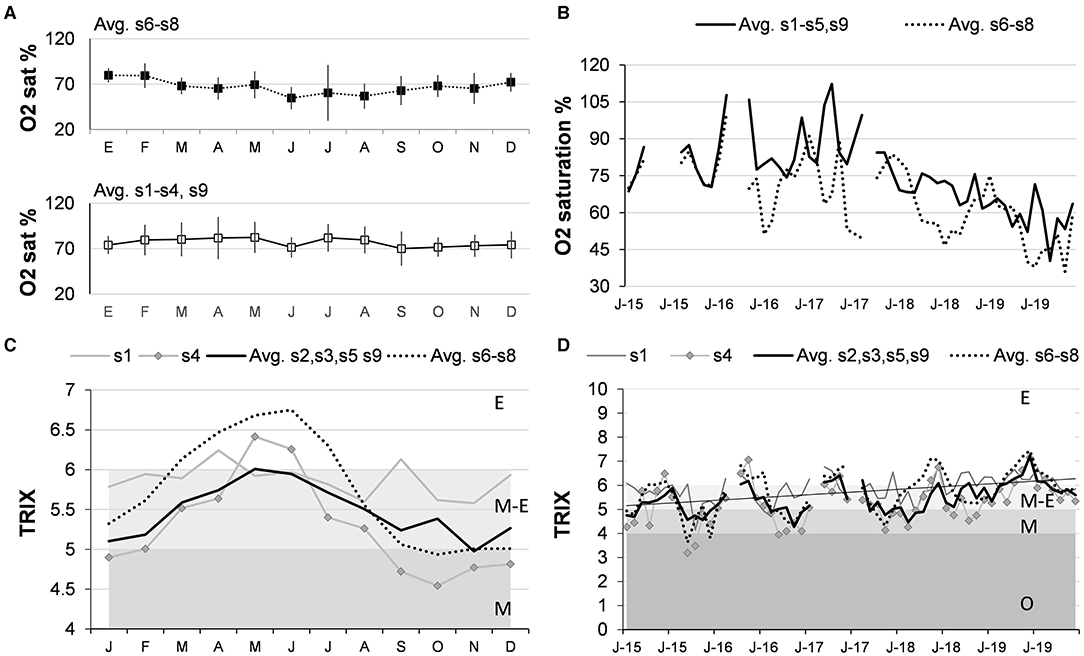
Figure 7. Oxygen saturation percentage (O2 sat%) and trophic index (TRIX). The left column (A,C) shows seasonal variability in the interior (s1, s4, avg, 2, 3, 5, 9) and the inlet (Avg. s6–s8). The right column (B,D) shows the 2015–2019 variability (J, January; J, June). Keys: E, eutrophic; M, mesotrophic; O, oligotrophic.
The results of multivariate methods applied to the TRIX trophic index were similar to %sat O2, with two sets: sites nearest to the inlet and in the interior of the lagoon (Figure 7C). The average of each set showed that TRIX presents a well-defined seasonal cycle: in the inlet, it varies between eutrophic-mesotrophic in summer and autumn and eutrophic in spring and early summer. TRIX varied between mesotrophic and eutrophic in the interior of the lagoon, with peaks at the end of summer and in the beginning of autumn. The time series showed a positive trend (p < 0.01), and the slope of regression was slightly higher in the sites of the inlet (0.018) than in the interior (0.015), showing progressive eutrophication (Figure 7D).
Algal Blooms
There are few records of algal blooms into or off BM-BA. Nineteen ABs observed at discontinuous moments from 1931 to 2014 were formed by numerous diatoms species but dominated by a few (Gárate-Lizárraga et al., 2016). The most abundant and frequent causal species were Eucampia zodiacus, Leptocylindrus danicus, and several species of the Chaetoceros genus, among which Chaetoceros compressus stands out. Dinoflagellate blooms were less common; dominant species were Prorocentrum triestinum, Prorocentrum koreanum, Akashiwo sanguinea, Noctiluca scintillans, and Peridinium quadridentatum, which were observed mainly in summer and in the channel zone, in the northern extreme of the lagoon system.
From 2015 to 2019, seven blooms with densities >5 × 103 cells L−1 were identified and composed mainly of diatoms. The dominant species were the chain-forming diatoms E. zodiacus, Rhizosolenia setigera, Guinardia striata, Skeletonema sp, and Dactyliosolen fragilissimus; also, there were unidentified nanoplankton cells. There were also high densities of the thecate dinoflagellates P. quadridentatum and Prorocentrum gracile and specimen cf. Gyrodinium estuariale.
In May and June 2017, a HAB occurred, possibly caused by D. fragilissimus, P. gracile, Pseudo-nitzschia spp. (delicatissima complex) and an unidentified species cf. G. estuariale (Barón-Campis, pers. comm.). As a consequence, during June, by caution, the harvest season for Panopea globosa was closed by fishing authorities, and also there were reports of gill damages in some yellowtail amberjack juveniles in a culture near the inlet.
In 2019, a very extensive HAB occurred off the western coast of Baja California, from the San Ignacio lagoon to BM, with reports of demersal species mortality on the north shore of Isla Magdalena (Gabriel Jiménez, pers. comm.). In May, the chain-forming diatom E. zodiacus and the dinoflagellate Noctiluca sp. were the most abundant species, while in June, the diatoms E. zodiacus, Coscinodiscus wailesii, and several unidentified species of nanoplankton (<20 microns) were the most abundant species. In that month, there were also dinoflagellates with toxic potential but in low quantities. In July, nanoplankton species were the most abundant (≈1.6 million cells l−1), although there were also the diatoms E. zodiacus and Thalassiosira spp. Several species with toxic potential were recorded (Pseudo-nitzschia cf. delicatissima, Karlodinium cf veneficum); however, their identification was not definitive, and an analysis of toxins could not be performed, but there were no reports of fish damage. On the other hand, there were unpleasant odors, and the color of the water was reddish-brown for several weeks between May and July.
Discussion
This section is organized around three issues: (1) studies on bivalve assemblages and hypotheses on their characteristics from commercial species; (2) fishing and its problems; (3) environmental variations and hypotheses on their impact on commercially collected bivalves.
The Community of Bivalves
The services of bivalves in the ecosystem are multiple (Brumbaugh and Toropova, 2008), and due to their great abundance, they are an important trophic link in marine ecosystems (Arapov et al., 2010; Palomares-Sala, 2020). However, the study and management of bivalves and their habitat for purposes beyond harvesting and culture has not yet become widespread (Northern Economics Inc., 2009), as to what happened in Mexico and BM, where despite their economic value, there are no studies on community structure and function and the little information available is recent and is in postgraduate theses. There are probably around 200 species, most of which are very-small and small infaunal, which can explain the little information that exists. The bivalves from medium to extra-large sizes are intensively harvested because the fishing effort invested is lesser than shrimp and finfish fisheries.
The almost yearly monitoring studies made by authorities are focused on abundance measures done at the solicitude of fishermen, which requires many human and material resources, limiting the number of banks and stocks that can be assessed. On the other hand, these data are not available in public databases. Another indicator of this variable is the CPUE. Nevertheless, effort data are minimal, and in most cases, only are available the number of permits or vessels, so it is not possible to calculate the CPUE, and catch is not a reliable indicator of abundance, because it is biased to market demand and more valuable resource abundance (Amezcua-Castro et al., 2015). This lack of information limits the analysis of commercially exploited stocks and their ecosystem services (Amezcua-Castro, 2020).
The trophic and abiotic conditions in each zone of the lagoon probably influence the structure of bivalve assemblages and their reproductive and feeding strategies. The characteristics of mollusk assemblages in other coastal lagoons, both seasonal and spatial, are related to freshwater inputs, temperature, dissolved oxygen, salinity, and submerged vegetation (De Jesús-Carrillo et al., 2020), with a higher richness species and abundance where conditions were less extreme (Koutsoubas et al., 2000). In BM, in the northeastern shallow zone (including the San Carlos channel) and southern channels and its vicinity, the temperature is high, chl-a and POC are abundant, and the water column is well-mixed and oxygenated (Cervantes-Duarte et al., 2021); into the channels, there are also macroalgae and seagrasses prairies during spring (Hernández-Carmona et al., 2007). These conditions are likely to favor rich species assemblages for infauna and other species such as scallops, which can move when conditions are stressful. Something similar is likely to occur on the shore of the central body of water, while in the deepest part, where oxygen and temperature are comparatively low, the assemblages of bivalves are probably poorer.
The biology of the commercially exploited bivalves suggests that in other species, reproduction would be aseasonal, with peaks associated with food availability and temperature (Félix-Pico, 2006). In the San Carlos channel and the northern and northeastern tidal channels, where the temperature is high and mesotrophic conditions prevail all year, species such as A. tuberculosa, Chione spp., M. squalida spawn in two periods (spring and autumn), and their infaunal habits probably allow for survival despite stressful conditions in the shallower areas. In contrast, the largest species inhabit the body of water, and, although they may have aseasonal reproductive cycles (pen shells), tend to have a period in which spawning is high. P. globosa and A. maura reproduction peaks coincide with algae spring blooms and eutrophic conditions caused by coastal upwellings. P. rugosa reproduction, on the other hand, is high during summer, when POC is also high. However, they are distributed in different zones: geoduck clams are in the deepest zones, close to the inlet, where the concentrations of chl-a and POC are high during spring, while pen shells live in the intertidal-subtidal zones of the channels and the eastern shore (Fajardo-León, 1998), where food availability is high almost all year. The scallop A. ventricosus is widely distributed, which means that it tolerates a wide range of environmental conditions or that it moves to more favorable zones.
Fisheries Impact on Populations
The harvest of bivalves is almost all-year-round, depending on the abundance of the bivalve in question and other most valuable resources and market demand. For example, the March and April high catches respond to high shellfish demand during the Holy Week. In contrast, the low September values are usually associated with the beginning of the shrimp fishing season (September-February/March). The trend of the historical time series responds to a similar dynamic, so the wide variations of the squalid callista depend on market demand and the abundance of the Pacific calico scallop, which has a better price (Amezcua-Castro et al., 2015).
Almost all commercial species have been intensively harvested, giving place to cycles of abundance and overexploitation along with its history. For example, due to the intense fishing that has followed each pulse of the abundance of Pacific calico scallop, they have been classified as unprofitable (DOF, 1987) or even overexploited (Massó-Rojas, 1996; Magadán-Revelo, 2015; Narchi et al., 2018). Squalid callista and pen shells also have received this label at different times (Fajardo-León, 1998; López-Rocha et al., 2010; Narchi et al., 2018). These wide variations occurred because fishermen focus on daily subsistence without considering the long-term consequences of immoderate harvest.
After 2014, harvest declined abruptly in almost all species. In the case of the scallops, some media indicate a 7-year closure, but Magadán-Revelo (2015) pointed out that the resource disappeared in 2013, and that the fishery also did. The resource reappeared in 2018 when authorities extended quotas again. Pen shell abundance has been very low since 2015, and in 2017, the authorities classified the resource as overexploited (DOF, 2018b; Narchi et al., 2018) and decreed a 2-year closure (DOF, 2018b). The harvest of squalid callista was very intense from 2012 to 2014, probably due to low the abundance of scallops, disappearance of pen shells, and lack of regulations; the results of our analysis s suggest that after 2014 high temperature and low food availability also contributed to the diminished abundance. In 2015, the resource was labeled as “overexploited” because of its low density values. In 2017, the authorities decreed 2 years of closure in BM-BA, extended 1 year more in BM. In 2016, the harvest of Venus and frilled Venus clams was very high due to the scarcity of other bivalves and the appearance of new buyers from Tijuana and Guadalajara metropolises. In 2017, the harvest of both diminished. The main target of fisheries has been the geoduck clam since 2009, and CONAPESCA inspectors and non-governmental organizations supervised the harvest of legal fishermen (Magadán-Revelo, 2015). However, illegal fishing by guateros is very intense.
In 2017, the intense harvest of Pacific-wing oysters began to offset the depletion of the most valuable resources. Its callus resembles pen shells, and it has a reasonable price. A few fishing permits were extended in 2020, but there are no other regulations. Abundance evaluations have not been made, although fishermen have requested them (Guzmán 2021, pers. comm.). This oyster is an example of what happens when new resources are discovered; fishermen start the exploitation without any control, which decreases their abundance and price (Narchi et al., 2018). Meanwhile, other important species recover, or the season of others begins, so fishing is redirected, giving rise to cycles. On the other hand, the impact of these cycles on populations and the ecosystem is not evaluated.
Regulation of the harvest of bivalves combines minimum shell size, closure seasons to protect reproduction, density minimum, and quotas. National Fisheries Charter (DOF, 2018a) establishes that fishermen registration of scallops, squalid callista, pen shells, and geoduck clam is closed in BM, and that fishing efforts must not increase. Quotas are established yearly after technical surveys from the National Institute of Aquaculture and Fisheries (INAPESCA in Spanish), the assessor of CONAPESCA, and collaborations with fishermen. In recent years, budget cuts and the SARS-Covid-2 pandemic have limited these activities.
In all the species, the implementation of fishing regulations has been reactive to “intensive exploitation” as it indicates the first Agreement decreed to regulate the Pacific calico scallop populations (DOF, 1987). On the other hand, regulations usually are delayed. The Norma Oficial Mexicana that established the closure to protect the span of Pacific calico scallop reproduction was published in 1994 (DOF, 1994), after the very intense harvest of 1989–1990, while the closure for pen shells is published in 2021, after the depletion of the resource (DOF, 2021). The closure for other species, such as the geoduck clam, whose harvest began in 2009, is under review. This mismatch must significantly impact populations, because even though they reproduce all-year-round in BM-BA, harvest and spawning peaks coincide frequently, limiting the recovery of the populations. A strategy to alleviate this problem is establishing annual quotas, subject to stock density assessments; however, biological evaluations of population conditions are scarce or limited (Amezcua-Castro, 2020). Therefore, it is necessary to design policies to protect populations and diminish the gap between legal regulations and fishing activities.
Illegal fishing is a big challenge, because guateros have the infrastructure and personnel to harvest very efficiently, and the inspection and surveillance by fishery authorities have been very limited. There have been community participation attempts at surveillance, but they have failed due to lack of long-term budget. Another problem is the traceability of illegal harvest, as it easily integrates with the legal catch (Magadán-Revelo, 2015). It is almost impossible to evaluate the impact of illegal fishing, but the most obvious is the diminishing of the price in the short term and the resilience of stocks in the long term.
Artisanal fishing is an escape valve to unemployment, favoring illegality. There were 74,286 small vessels registered in 2017 in Mexico, but the total is probably double this number (Inteligencia Pública EDF de México, 2019). A similar situation probably occurs in BM-BA. On the other hand, the fishing authority has been substantially weakened in recent years. The budget of the Mexican government assigned to CONAPESCA had a negative trend, going from 1,102.6 million pesos in 2015 (DOF, 2014) to 584.3 million pesos in 2021 (DOF, 2020). Something similar has happened with INAPESCA, the technical advisor of the authorities, whose budget decreased from 721 million pesos in 2015 to 455.5 million pesos in 2021, significantly affecting the fishery resources monitoring and surveillance programs.
From 2019 to 2020, 262 complaints of illegal fishing11 in the northwestern Mexican region were registered, of which 51 were from the municipality of Comondú, where the BM-BA lagoon system is located. Until 2020, fisheries inspection and surveillance were the responsibilities of CONAPESCA, but they were transferred to the Marine Secretary (SeMar) on April 1 to increase the coverage of these actions. CONAPESCA had problems developing these activities because of the SARS-Covid-2 pandemic and internal corruption (Walter Hubbard declaration; July 8, 2020).12 Although the consequences of these measures will be seen in the future, fishermen from Puerto San Carlos express their annoyance by saying that illegal fishing continued during the first months of 2021 and fear that it will increase even though SEMAR is already in charge (Barrón, pers. comm.).
Despite efforts of the government, the Organization for Economic Co-operation and Development (OECD) has pointed out the existence of legal gaps and the indefiniteness of legal guidelines to confront overexploitation and illegal fishing (Martínez-Martínez and González-Laxe, 2016). OECD also highlights the absence of a clear sectoral strategy to increase the profitability of fishing activity in the short term. Therefore, it recommends adopting agreements to move toward decentralized institutional schemes that empower government states without weakening federal institutions or risking marine and fishing resources.
Several lines of action must be considered in the development strategy for the fishing activity, such as creating conditions to strengthen the target market of products and improving prices by adding value to final products. They have to consider the needs of the producers, such as constructing their own offices, maintenance workshops, storing goods, searching for dock concessions, and renewing the fishing fleet. These actions require financing, which has been practically nil in the fishing sector (Olmos-Martínez, 2012).
The fishery presents significant problems associated with illegal harvesting, a lack of regulatory measures enacted on time, and inefficient monitoring and surveillance. We can suppose that fisheries have survived thanks to the high recovery capacity of the populations of bivalves because of their short life span, aseasonal reproductive cycles, and the high food availability in BM. A different situation is the geoduck clam, whose biological characteristics could lead to depletion without adequate regulation and surveillance (Amezcua-Castro, 2020).
Environment and the Bivalve Populations
In this section, we discuss BM environmental variability, some factors that could influence it, and the impact on the physiology of bivalves. From 1970 to 2019 off BM, the evolution of environmental conditions was characterized by warming and diminishing productivity at a synoptic scale, with wide variations caused by equatorial and northern Pacific climatic oscillations. In the 1980 and 1990's, the warming periods, associated with the very strong El Niño phenomena, were followed by very intense La Niña, which reactivated productivity. However, after 2013, temperature anomalies were very high because of the co-occurrence of The Blob and the 2015–2016 ENSO. The last one, a combination of Eastern Pacific and Central Pacific El Niño, was characterized by a second warming point located in front of Baja California (Kug et al., 2021). During this span, the Pacific Meridional Mode (PMM) reached record positive values, and subtropical Pacific temperature anomalies were high before, during, and after El Niño. High temperature anomalies also may be associated with the positive phase of PDO, warming due to anthropogenic forcing (Kug et al., 2021), and the heatwaves NEPH 19 a, b, and NEPH20, the second-largest heatwave since 1982. After 2016, the La Niña phenomenon and NPGO have been very weak, limiting upwellings and nutrient-rich water mass advection from the northern Pacific.
Temperature and chl-a variations at the local scale were similar to the synoptic scale, suggesting a longer trend of warming and food availability diminishing into the lagoon than the 2002–2020 data indicated. During warm periods, upwelling strength and nutrient entrance to the lagoon diminish, altering chl-a seasonal patterns (Jiménez-Quiroz et al., 2019). The decrease in chl-a associated with lower phytoplanktonic biomass is mitigated by the organic material supplied from other plants, as the POC recovery after 2017 indicated. The persistence of low chl-a values can also be due to changes in the dominant phytoplankton fraction from micro to nanoplankton. On a longer scale, our results suggest that variations of the California Current (related to NPGO) also affect chl-a concentrations, probably because the nutrients transported by this current are minor.
Although TRIX index values are an instantaneous picture of the trophic state of the lagoon, it highlights some remarkable characteristics: the broad seasonal changes in the inlet surroundings and the more constant values in the eastern shore, the channels, and their vicinity. In the body of water, the seasonality of upwelling defines TRIX variations, while in the channels, other mechanisms contribute to nutrient input as water column mixing (Cervantes-Duarte et al., 2021). These seasonal and spatial patterns could define the characteristics of bivalve assemblages, as we have discussed previously. TRIX also suggest a progressive deterioration of water quality; in the interior of the lagoon, it can be associated with human activities in the San Carlos port, which include fishing, industrial and wastewater discharges, and cargo, among others (Cervantes-Duarte et al., 2021). In the body of water, the positive trend of eutrophication coincided with the diminishing of oxygen saturation, especially near the inlet after 2017. The lowest values coincided with extensive algal blooms in June 2017 and May-June 2019, and with high ammonia contents indicating poor water quality. In 2019, those values prevailed until August, probably because of microalgae decomposition after HAB ended. The difference in the slope of time series trend from each zone of the lagoon suggests that the eutrophication was due to marine rather than inner processes.
The 2019 HAB extended from the San Ignacio lagoon to BM from May to the 1st days of July. In BM, it consisted mainly of the diatoms E. zodiacus, Thalassiosira sp. and C. wailesii, the dinoflagellate Noctiluca sp., and nanoplankton cells. E. zodiacus is large and chain-forming; it is common in BM and can absorb nutrients quickly (Nishikawa et al., 2009). This species is holoplanktonic, so its abundance increases faster than that of meroplanktonic species, such as Skeletonema spp., Chaetoceros spp., and Thalassiosira spp, other common BM species (Naito et al., 2011). E. zodiacus also reached high densities in April 2015 (>500 × 103 cells L−1) after a period of strong upwelling (Jiménez-Quiroz et al., 2019). In May, Noctiluca sp. was observed, whose abundance has been positively correlated with ammonium and phosphates (Genitsaris et al., 2019). This species consumes diatoms, such as Thalassiosira sp. (Tsai et al., 2018). In June, in addition to E. zodiacus, the density of C. wailesii was also very high; it is an invasive species that produces mucilage, which causes oxygen and nutrient depletion (Talgatti et al., 2010). In July, in addition to E. zodiacus, there were very high densities of nanoplankton cells, and Thalassiosira spp. species with harmful potential were identified (Baron-Campis, comm. pers.), but apparently, there were no reports of intoxications.
The HAB was composed of opportunistic organisms with a high capacity to absorb nutrients, even when these are scarce. It was not possible to gather more samples, but taxonomic composition differences among May, June, and July suggest a succession of species. On the other hand, in June, we recorded the highest concentrations of nitrates, phosphates, and silicates in the 2015–2019 period due to intense upwellings throughout the southwestern coast of Baja California. In turn, the death of these organisms decreased oxygen concentration and increased ammonia. There are no evaluations of the impact of the HAB in wildlife or organic matter accumulation in BM, even though it caused mortalities in demersal fish from the San Juanico lagoon to the northern shore of Magdalena Island. It is possible, in most BM sites, that topographical relief, semidiurnal mixed tides amplitude, and hydrodynamics imposed by wind and tide contributed to avoiding anoxia.
The impact of warming and habitat deterioration in the bivalve community of BM-BA is unknown. The increase in temperature observed between 2014 and 2016 (NSST anomalies >1) was probably more stressful for bivalve populations than in previous years when anomalies were close to the unity. The optimal and lethal temperature information on Baja California bivalves is scarce. For the scallops A. ventricosus and Nodipecten subnodosus, the parameters are 20 and ≈28°C, while for the pen shell A. maura, the parameters are 29 and 33.2°C, respectively (Sicard et al., 2006). Geoduck clam (P. globosa) lethal temperature is 32°C in the laboratory (González-Peláez et al., 2013). Mean temperatures in BM are usually below these values; however, after 2014, they were close to 30°C during several months (Jiménez-Quiroz et al., 2019; Cervantes-Duarte et al., 2021). Over time, the increase in temperature allows for the acclimatization of organisms, but there is no information on the resilience of BM bivalves to anomalous warming periods. Hermaphroditism prevalence and reproduction to a smaller than average size, strategies that favor survival under stressful conditions (Romo-Piñera, 2010; Breton et al., 2019), could be indicators of the magnitude of these conditions.
The abundance of bivalves is very variable, and in addition to fishing pressure, it depends on environmental conditions, which have not been fully understood, as is the case of the Pacific calico scallop. The pulses of abundance of scallops (Morales-Zárate, 2006) have coincided with cold periods, as the La Niña phenomenon of 1989–1990, 2006–2008, and 2009–2010. In these spans, positive chl-a anomalies were frequent, indicating high food concentrations. A circulation model suggests that moderate to strong westerly winds (3–5 m s−1) advect particles (as larvae) from the ocean off BM into the lagoon (Morales-Zárate, 2006), which probably were more common during the La Niña in 1989–1990 and in 1995–1996 when low temperatures and strong westerly winds coincided. After these years, the harvest of scallops was very high. In this context, Ruiz-Verdugo et al. (2016) suggested that warm temperatures after 2015 would affect their abundance, but in 2018, the harvest was similar to that of 2008, after the strong 2007–2008 La Niña, which caused negative temperature anomalies from the summer of 2007 to the spring of 2008.
After the very strong 2015-2016 El Niño, there were two consecutive weak La Niña phenomena. In 2016, when the El Niño phenomenon was decaying, the strength of upwellings returned to normal after it became very weak from 2014 to 2015. In 2017, upwellings were more frequent and stronger since March, and they were particularly strong during the autumn, even though temperature anomalies were positive in almost the entire BM. According to the results from the yearly time series (1982–2020) analysis, autumn upwellings could contribute to the abundance in the following year, and probably, they could be a gross index of it. Therefore, it is necessary to deepen the study on this topic and the environmental effects on the Pacific calico scallop abundance pulses and identify the location of banks off BM.
There is evidence that high temperatures affect reproduction, condition state, intra- and inter-specific relationships (depredation, parasitism), among other aspects of the biology of bivalves. Scallops and squalid callista reproduce when temperature is ≈20°C (Félix-Pico, 2006; Romo-Piñera, 2010). High temperatures produce gonadic atresia in pen shells from BM (>27.5°C) and in these species and Spondylus crassisquama from the Ojo de Liebre lagoon (temperature anomalies >2°C) (Camacho-Mondragón, 2009, 2014; Del Rio, 2018; Gómez-Valdez, 2019), suggesting that it could be a more spread strategy to survive anomalous warming events. Similarly, there were a higher number of hermaphrodites in the Ojo de Liebre lagoon and of M. squalida and A. tuberculosa in BM during warm periods (Romo-Piñera, 2010). Larval development can also be affected by temperature. The larvae growth and survival of P. globosa were better at 22 and 26°C than at 18 and 30°C (Buitimea-Álvarez, 2011). In Pteria sterna larvae, fixation occurs between 21 and 22°C, and massive fixation occurs between 27 and 28°C (McAnally-Salas and Valenzuela-Espinoza, 1990). Thus, higher temperatures that have been prevailing in the last years have probably been affecting the reproduction and recruitment of bivalves.
Metabolism rate is directly related to temperature weakening the organism when outside their optimal range; high values increase the susceptibility of organisms to diseases and parasites. In the Ojo de Liebre lagoon, large quantities of the nematode Echinocephalus pseudouncinatus have been observed in the populations of A ventricosus (Gómez del Prado-Rosas, 1984), A. maura, and P. rugosa (Gómez-Valdez, 2019). The prevalence of the parasite in pen shells was very high in 2015, favored by the elevated temperature and low chl-a concentrations (Gómez-Valdez, 2019). In BM-BA, the crustacean Tumidotheres margarita was in 31% of Pacific calico scallop adults during the spawning season; other organisms observed in this species may limit its growth and reproduction and leave it vulnerable to predators (Félix-Pico, 2006).
The acidification caused by excess CO2 and warming of water have a chronic but non-lethal effect on juvenile Arpecten purpuratus (Lagos et al., 2016). During warm periods (such as El Niño), individuals can increase their calcification rate and reach a larger shell size; however, they would be thinner, more fragile, and vulnerable to predators and parasites. On the other hand, in Pinna nobilis juveniles, acidification influences growth and partially, mortality, while the increase in temperature significantly increases mortality rates and oxygen consumption (Basso et al., 2015). The accumulation of organic matters in decomposition, as what occurs after an extensive HAB at the end of spring (e.g., 2019), could cause acidification and affect the physiology of some species despite present hydrodynamic conditions.
Bivalves feed on phytoplankton, dissolved organic matter, and bacteria [Sastry taken from Amezcua-Castro (2014) and Góngora-Gómez et al. (2016)]. The seasonality of chl-a; POC showed continuous food availability in BM; in spring and sometimes, in autumn, nutrients supplied by upwellings cause algal blooms (ABs), composed mainly of diatoms and dinoflagellates (Gárate-Lizárraga and Siqueiros-Beltrones, 1998; Jiménez-Quiroz et al., 2019). In addition to ABs, other vegetal organic matter inputs in the interior of the lagoon during summer and fall (Jiménez-Quiroz et al., 2021), including benthic diatoms (Gárate-Lizárraga et al., 2001), are responsible for POC abundance, which explained the recovery of this variable after 2017. An indicator of food availability is the average oocyte size. The size of M. squalida, A. maura, and A. ventricosus oocytes is bigger in BM than in La Paz and other Baja California locations (Romo-Piñera, 2010; Amezcua-Castro, 2014; Camacho-Mondragón, 2014), probably because of its high chl-a and POC concentration. These studies were conducted before the anomalous warming, and it would be recommendable to compare their results with other more recent ones.
Characteristics of phytoplankton assemblage change over the year in BM. Spring communities are formed by micro-phytoplankton dominated by diatoms, while dinoflagellates became dominant in summer. However, nanoplankton became the main fraction during warmer periods in 1986 (Gárate-Lizárraga and Siqueiros-Beltrones, 1998), 1988–1989 (Gárate-Lizárraga et al., 2001), 2015 (Jiménez-Quiroz et al., 2019), and the 2019 summer. Dominant diatom species, such as E. zodiacus, Skeletonema sp, G. striata, D. fragilissimus, and R. setigera have mechanisms to avoid zooplankton depredation, such as chain formation (Bjærke et al., 2015) and aldehyde excretion (Miralto et al., 1999), among others. It would be essential to determine how these phytoplankton strategies affect the larvae in bivalves, especially during HABs. In other cases (e.g., D. fragilissimus), high cell densities may harm the respiration of scallops either by gill clogging or by oxygen depletion (Lorrain et al., 2000).
There are no studies on the gut content of BM bivalves, but in some Ostreidae species, it is composed of benthic diatom characteristics of their habitats (Hernández-Almeida et al., 2019). Bivalves can discriminate between different microalgae species and ingest certain species to a greater extent; however, this procedure is not 100% effective, so they ingest small quantities of less desirable particles (Davenport et al., 2011). On the other hand, factors that influence the selection vary between bivalve species, and the variation in the size of phytoplankton could influence the rate of filtration and decrease the condition index of the organisms (Rosa et al., 2018). Global warming is expected to modify the phytoplankton phenology and species composition. In BM, nanoplankton forms have increased in the warmest months and years, and the decrease in average size and fewer large diatoms could produce impoverished conditions for feeding zooplankton and a less efficient transfer of energy to the rest of the trophic web (Basu and Mackey, 2018). It may affect the feeding of veliger larvae, because the ingestion of larger cells is more advantageous for a veliger larva than smaller cells (Raby et al., 1997).
There is no information on food quality changes related to the composition of phytoplankton assemblages in BM-BA. In Bahía Concepción (Gulf of California), the carbohydrate/protein ratio and chl-a concentration of the particulate organic matter during the cold season have better nutritional quality, probably associated with diatom blooms typical of this season (Verdugo-Díaz, 1997). In this regard, it should be mentioned that the changes in the structure of phytoplankton assemblages (cell size, taxonomic composition, among others) during the warming period of 2014–2016 probably caused the deficit in chl-a and POC.
Final Comments and Conclusion
There are no studies on the structure of bivalve communities or their “ecological role” within BA-BM; the biological characteristics of the commercially exploited species and the high availability of food have favored their survival despite being intensively harvested. During the period in which commercial fishing has developed, environmental conditions have varied following the synoptic and global-scale climatic processes. The long-term trend suggests that the region has gradually warmed, and that the biomass of the phytoplankton has decreased probably because of variations in the size of the dominant fraction;. However, inside BM, there is a constant contribution of POC by other plants and phytoplankton, so the availability of food is almost constant.
Since 2010, there have been periods of excessive warming caused by a conjunction of oceanic and atmospheric phenomena, which has not been recorded previously. Bivalves have physiological mechanisms to respond and adapt when these changes are gradual; however, anomalous warming events may limit the resilience of populations. Fishing has been poorly monitored and regulated because of the weakness of government institutions. The SARS-Covid-2 pandemic has highlighted this situation. On the other hand, bivalves are part of the self-purification mechanism of bodies of water; the decrease in the population abundance of the larger species has probably negatively affected this capacity. These mollusks cover the entire BM-BA lagoon, and it is essential to determine the impact of their extraction on the resilience of the lagoon.
The management of bivalve fisheries must consider constant monitoring of environmental variables, both in situ and with remote sensing tools. Some authors have pointed out the importance of adjusting catch quotas to prevailing environmental conditions, since they are currently based on percentages of the abundance recorded during assessments. Therefore, it must also monitor key biological variables of sensitive species (pen shells, geoduck). Currently, the ability to predict weather conditions (e.g., El Niño) can only be done a few months in advance. However, scenarios could be developed that would allow fishermen to adapt to these changes to maintain the sustainability of the fisheries.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
MJ-Q and FB-B designed the study, carried out statistical analysis, and interpretation of the results. RC-D designed the sampling field, carried out the calculation, and interpretation of the TRIX index. RF-R contributed to the writing and interpretation of the results. All authors contributed to the article and approved the submitted version.
Funding
This study was carried out with funding from the National Institute of Fisheries and Aquaculture (INAPESCA-SADER) and the National Polytechnic Institute-CICIMAR.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We deeply appreciate the collaboration of Sofía Alida Barón Campis in the identification of phytoplankton, as well as Don Luis Muñoz in the field activities; Felipe de Jesús García Romero and Luis Ricardo Amaro in the field and laboratory work; and Raul Martell Dubois in the MODIS-Aqua data acquisition. Finally, a recognition to Luis Daniel Magadán-Revelo for allowing us to use the bivalve bank distribution map included in his Master's thesis. On the other hand, the data of the NSST, chl-a, and POC used in this study were produced with the Giovanni online data system, and developed and maintained by the NASA GES DISC.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2021.682148/full#supplementary-material
Footnotes
1. ^https://www.conapesca.gob.mx/work/sites/cona/dgppe/anuarios/BASE_DE_DATOS_DE_PRODUCCION_ANUARIO_2018.zip
2. ^https://www.conapesca.gob.mx/wb/cona/informacion_estadistica_por_especie_y_entidad
3. ^https://psl.noaa.gov/pdo/ Using EOF from 1920 to 2014 for N Pacific; https://psl.noaa.gov/pdo/
4. ^http://www.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/detrend.nino34.ascii.txt
5. ^http://www.o3d.org/npgo/npgo.php
6. ^https://podaac-tools.jpl.nasa.gov/drive/files/allData/ersst/L4/ncei/v4/monthly/netcdf
7. ^https://oceanwatch.pfeg.noaa.gov/products/PFELData/upwell/monthly/upindex.mon
8. ^Dataset ID: pmlEsaCCI42OceanColorMonthly; https://coastwatch.pfeg.noaa.gov/erddap/index.html
9. ^https://giovanni.gsfc.nasa.gov/giovanni/
10. ^No. RJL/INAPESCA/DGAIPP/1673/2018, November 22nd of 2018. www.cofemersimir.gob.mx/expediente/22992/mir/46626/anexo/4977137+&cd=1&hl=es-419&ct=clnk&gl=mx.
11. ^https://pescandodatos.causanatura.org/combate-a-la-pesca-ilegal, 2021.
12. ^Ernesto Méndez Conapesca no puede con inspección y vigilancia de mares mexicanos. https://www.excelsior.com.mx/nacional/conapesca-no-puede-con-inspeccion-y-vigilancia-de-mares-mexicanos/1392883.
References
Amezcua-Castro, S. (2014). Uso de áreas de pesca de almeja chocolata (Megapitaria squalida) en Bahía Magdalena-Almejas, Baja California Sur, México. (dissertation/ master's thesis), IPN-CICIMAR, La Paz, México. Available online at: https://www.repositoriodigital.ipn.mx/bitstream/123456789/19981/1/amezcuaca1.pdf (accessed November 20, 2020).
Amezcua-Castro, S. (2020). Evaluación de la población de almeja de sifón (Panopea globosa) en Bahía Magdalena, Baja California Sur, México. (dissertation/ PhD thesis), IPN-CICIMAR, La Paz, México. Available online at: https://repositoriodigital.ipn.mx/bitstream/123456789/26264/1/amezcuaca2.pdf (accessed February 02, 2021).
Amezcua-Castro, S., Ramírez-Rodríguez, M., Medina-Gómez, S. P., and Gutiérrez-González, J. L. (2015). Tendencias de producción de almeja chocolata Megapitaria squalida en Bahía Magdalena-Almejas, Baja California Sur, México. Ciencia Pesquera 23, 3–10.
Arapov, J., Ezgeta–Balić, D., Peharda, M., and Ninčević Gladan, Ž. (2010). Bivalve feeding - how and what they eat? Ribarstvo 68, 105–116.
Baqueiro, C. E. (1979). Sobre la distribución de Megaptairia aurantiaca (Sow., 1831), M. squalida (Sow., 1835) y Dosinia ponderosa (Gray, 1838) en relación a la granulometría del sedimento (Bivalvia: Veneridae). An. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. Mex. 1, 25–32. Available online at: http://biblioweb.tic.unam.mx/cienciasdelmar/centro/1979-1/articulo54.html (accessed November 06, 2020).
Baqueiro, C. E., and Aldana-Aranda, D. (2003). Patrones en la biología poblacional de moluscos de importancia comercial en México. Rev. Biol. Trop. 51, 97–102. doi: 10.15517/rbt.v51i4
Baqueiro, C. E., Massó, R. J. A., and Guajardo-B, H. (1982a). Distribución y abundancia de moluscos de importancia comercial en Baja California Sur. Serie de Divulgación no. 11. 15BN-968-817-016-X. Secretaria de Pesca, México, 29. Available online at: https://www.inapesca.gob.mx/portal/Publicaciones/Series/1970s-INP-Serie-Divulgacion/SD-11-Baqueiro-etal-1982-moluscos-BCS.pdf?download (accessed November 05, 2020).
Baqueiro, C. E., Muciño, M., and Merino, R. (1982b). Análisis de una población de pata de mula Anadara tuberculosa sujeta a explotación intensiva en la Bahía de La Paz, Baja California Sur, México. Ciencia Pesquera 3, 75–82.
Barraza-Guardado, R. H., Martínez-Córdova, L. R., Enríquez-Ocaña, L. F., Martínez-Porchas, M., Miranda-Baeza, A., and Porchas-Cornejo, M. A. (2014). Effect of shrimp farm effluent on water and sediment quality parameters off the coast of Sonora, Mexico. Cienc. Mar. 40, 221–235. doi: 10.7773/cm.v40i4.2424
Basso, L., Hendriks, I. E., and Duarte, C. M. (2015). Juvenile pen shells (Pinna nobilis) tolerate acidification but are vulnerable to warming. Estuaries Coast. 38:1976–1985. doi: 10.1007/s12237-015-9948-0
Basu, S., and Mackey, K. R. M. (2018). Phytoplankton as key mediators of the biological carbon pump: Their responses to a changing climate. Sustainability 10:869. doi: 10.3390/su10030869
Bjærke, O., Jonsson, P. R., Alam, A. S., and Selander, E. (2015). Is chain length in phytoplankton regulated to evade predation? J. Plankton Res. 37, 1110–1119. doi: 10.1093/plankt/fbv076
Breton, S., Capt, C., Guerra, D., and Stewart, D. (2019). “Sex-determining mechanisms in bivalves,” in Transitions Between Sexual Systems: Understanding the Mechanisms of and Pathways Between Dioecy, Hermaphroditism, and Other Sexual Systems, ed J. L. Leonard (Springer), 165–192. ISBN, 978-3-319-94137-0 (hc); 978-3-319-94139-4 (eb). 2018.
Brumbaugh, R. D., and Toropova, C. (2008). Economic valuation of ecosystem services: a new impetus for shellfish restoration? Basin Coasts News 2, 8–15. Available online at: https://www.conservationgateway.org/Documents/Brumbaugh-ToropovaBasinsCoast.pdf
Buitimea-Álvarez, M. I. (2011). Efecto de la salinidad y temperatura sobre el desarrollo larval de Panopea spp. (dissertation/bachelor's tesis), Universidad Autónoma de Baja California, Ensenada, Mexico.
Camacho-Mondragón, M. A. (2009). Efecto de los fenómenos de “El Niño” y “La Niña” en la reproducción de hacha china Atrina maura en la Ensenada de la Paz, BCS. México. (dissertation/ master's thesis), IPN-CICIMAR, La Paz, Mexico.
Camacho-Mondragón, M. A. (2014). Organización de la gónada, gametogénesis, sexualidad y variaciones estacionales y geográficas de las tácticas reproductivas del hacha china Atrina maura (Sowerby, 1835) (Bivalvia: Pinnidae). (dissertation/ doctoral thesis). IPN-CICIMAR, La Paz, México. Available online at: https://www.repositoriodigital.ipn.mx/handle/123456789/21725 (accessed January 28, 2021).
Camacho-Mondragón, M. A., Arellano-Martínez, M., and Ceballos-Vázquez, B. P. (2012). Particular features of gonadal maturation and size at first maturity in Atrina maura (Bivalvia: Pinnidae). Scientia Marina 76, 539–548. doi: 10.3989/scimar.03522.05A
Cavieses-Nuñez, R. A. (2016). Modelos predictivos multiespecíficos para la pesca ribereña: pesquería de escama en la región de Bahía Magdalena Almejas. (dissertation/ master's thesis). UABCS-Ciencias del Mar y de la Tierra, La Paz, México. Available online at: http://rep.uabcs.mx:80/handle/23080/207 (accessed November 05, 2020).
Cervantes-Duarte, R., Jiménez-Quiroz, M. C., Funes-Rodríguez, R., Hernández-Trujillo, S., González-Armas, R., and Anaya-Godinez, E. (2021). Interannual variability in the trophic status and water quality of Bahía Magdalena, Mexico, during the 2015–2018 period: TRIX. Reg. Stud. Mar. Sci. 42:101638. doi: 10.1016/j.rsma.2021.101638
Cervantes-Duarte, R., López-López, S., González-Rodríguez, E., and Futema-Jiménez, S. (2010). Ciclo estacional de nutrientes, temperatura, salinidad y clorofila a en Bahía Magdalena, BCS, México (2006-2007). Oceanides 25:87. doi: 10.37543/oceanides.v25i2.87
Cervantes-Duarte, R., Prego, R., López-López, S., Aguirre-Bahena, F., and Ospina-Alvarez, N. (2013). Annual patterns of nutrients and chlorophyll in a subtropical coastal lagoon under the upwelling influence (SW of Baja-California Peninsula). Estuar. Coast. Shelf Sci. 120, 54–63. doi: 10.1016/j.ecss.2013.01.020
Chávez-Villalba, J., Arreola-Lizárraga, J. A., and Castillo-Durán, A. (2019). Condition index in three size classes of the smooth venus clam Chionista fluctifraga from a commercial fishery in Sonora, Mexico. Acta Universitaria 29:e1836. doi: 10.15174/au.2019.1836
Coan, E., and Valentich-Scott, P. (2012). Bivalve seashells of tropical West America. Marine Bivalve Mollusks from Baja California to Northern Perú. Santa Barbara: Santa Barbara Museum of Natural History.
Cruz-Gómez, L. M. (2012). Ciclo anual de la condición nutricional en la almeja generosa, P. globosa, de Bahía Magdalena, B. C. S. México. (dissertation/ master's thesis), IPN-CICIMAR, La Paz, Mexico.
Davenport, J., Ezgeta-Balić, D., Peharda, M., Skejić, S., Nincević-Gladan, Z., and Matijević, S. (2011). Size-differential feeding in Pinna nobilis L. (Mollusca: Bivalvia): exploitation of detritus, phytoplankton, and zooplankton. Estuar. Coast. Shelf Sci. 92, 246–254 doi: 10.1016/j.ecss.2010.12.033
De Jesús-Carrillo, R. M., Ocaña, F. A., Hernández-Ávila, I., Mendoza-Carranza, M., Sánchez, A. J., and Barba-Macías, E. (2020). Mollusk distribution in four habitats along a salinity gradient in a coastal lagoon from the Gulf of Mexico. J. Natural Hist. 54, 19–20. doi: 10.1080/00222933.2020.1785030
Del Rio, J. (2018). Biología reproductiva de Spondylus crassisquama en Laguna Ojo de Liebre, Baja Calfornia Sur, México. (dissertation/ master's thesis), IPN-CICIMAR, La Paz, Mexico.
Di Lorenzo, E., Schneider, N., Cobb, K. M., Franks, P. J. S., Chhak, K., Miller, A. J., et al. (2008). North Pacific Gyre Oscillation links ocean climate and ecosystem change. Geophys. Res. Lett. 35:L08607, doi: 10.1029/2007GL032838
DOF (1987). ACUERDO que regula la explotación de almeja catarina (Argopecten circularis) en aguas litorales del Estado de Baja California Sur. 1 de julio 1987. Available online at: http://dof.gob.mx/nota_detalle.php?codigo=4663930&fecha=01/07/1987&print=true (accessed February 10, 2021).
DOF (1993). NORMA Oficial Mexicana 004-PESC-1993, para regular el aprovechamiento de la almeja catarina, en aguas de jurisdicción federal de los estados de Baja California y Baja California Sur. 21 de diciembre 1993. Available online at: http://www.dof.gob.mx/nota_detalle.php?codigo=4779595&fecha=01/09/1993&print=true (accessed November 17, 2020).
DOF (1994). AVISO por el que se da a conocer el establecimiento de épocas y zonas de veda para la pesca de diferentes especies de la fauna acuática en aguas de jurisdicción federal de los Estados Unidos Mexicanos. 16 de marzo a 1994. Available online at: http://www.dof.gob.mx/nota_detalle.php?codigo=4678590&fecha=16/03/1994 (accessed November 17, 2020).
DOF (2010). ACUERDO mediante el cual se da a conocer la actualización de la Carta Nacional Pesquera (Continúa en la Tercera Sección). SEGOB. Available online at: http://dof.gob.mx/nota_detalle.php?codigo=5169418&fecha=02/12/2010 (accessed November 11, 2020).
DOF (2014). PRESUPUESTO de Egresos de la Federación para el Ejercicio Fiscal 2015. Available online at: http://www.dof.gob.mx/nota_detalle.php?codigo=5374053&fecha=03/12/2014 (accesed November 17, 2020).
DOF (2015a). NORMA Oficial Mexicana NOM-014-SAG/PESC-2015, Especificaciones para regular el aprovechamiento de almeja generosa (Panopea generosa y Panopea globosa) en aguas de jurisdicción federal del litoral del Océano Pacífico y Golfo de California. Available online at: http://www.dof.gob.mx/nota_detalle.php?codigo=5398036&fecha=25/06/2015 (accessed February 01, 2021).
DOF (2015b). NORMA Oficial Mexicana NOM-004-SAG/PESC-2015, Especificaciones para el aprovechamiento de la almeja catarina (Argopecten circularis) en aguas de jurisdicción federal de los Estados Unidos Mexicanos. Available online at: https://www.dof.gob.mx/nota_detalle.php?codigo=5410616&fecha=06/10/2015#:~:text=NORMA%20Oficial%20Mexicana%20NOM%2D004,de%20los%20Estados%20Unidos%20Mexicanos (accessed November 17, 2020).
DOF (2017). Veda temporal de dos años, contados a partir de la entrada en vigor del presente Acuerdo para M. squalida en el sistema Lagunar Bahía Magdalena-Almejas en el Estado de Baja California Sur. Available online at: https://www.dof.gob.mx/nota_detalle.php?codigo=5508553&fecha=21/12/2017 (accessed November 17, 2020).
DOF (2018a). ACUERDO por el que se da a conocer la actualización de la Carta Nacional Pesquera. (Continúa en la Tercera Sección). SEGOB. Available online at: http://dof.gob.mx/nota_detalle.php?codigo=5525712&fecha=11/06/2018 (accessed November 07, 2020).
DOF (2018b). ACUERDO por el que se establece veda temporal para la captura de callo de hacha (Pinna rugosa y Atrina maura), en aguas marinas de jurisdicción federal de la Bahía Magdalena, en el Estado de Baja California Sur. Available online at: http://www.dof.gob.mx/nota_detalle.php?codigo=5525397&fecha=07/06/2018 (accessed January 25, 2021).
DOF (2020). PRESUPUESTO de Egresos de la Federación para el Ejercicio Fiscal 2021. Available online at: https://www.dof.gob.mx/2020/SHCP/PEF_2021.pdf (accessed June 28, 2021).
DOF (2021). ACUERDO por el que se establece veda temporal para el aprovechamiento de callo de hacha (Pinna rugosa y Atrina maura) en aguas marinas de jurisdicción federal de Bahía Magdalena-Almejas, Baja California Sur. Available online at: http://www.dof.gob.mx/nota_detalle.php?codigo=5611232&fecha=10/02/2021&print=true (accessed February 11, 2021).
Durazo, R. (2015). Seasonality of the transitional region of the California Current System off Baja California. J. Geophys. Res. Oceans 120, 1173–1196. doi: 10.1002/2014JC010405
Fajardo-León, M. C. (1998). Abundancia y estructura poblacional de callo de hacha (Pinna rugosa, Atrina maura y Atrina sp) en bahía Magdalena, B. C. S., México. Abril de 1995. Boletín Pesquero CRIP-LA PAZ 9, 30–37.
Félix-Pico, E. (2006). “Mexico,” in Scallop Fisheries and Aquaculture, eds S. E. Shumway and G. J. Parsons (New York, NY: Elsevier), 1336–1380. doi: 10.1016/S0167-9309(06)80056-6
Félix-Pico, E. F. (1993). Estudio biológico de la almeja catarina, Argopecten circularis (Sowerby, 1835) en Bahía Magdalena, B.C.S., México. (dissertation/ master's thesis), IPN-CICIMAR, La Paz, México. Available online at: https://repositoriodigital.ipn.mx/handle/123456789/15252 (accessed March 15, 2021).
Félix-Pico, E. F., Ramírez-Rodríguez, M., and Holguín-Quiñones, O. (2009). Growth and fisheries of the black ark Anadara tuberculosa, a bivalve mollusk, in Bahia Magdalena, Baja California Sur, Mexico. North Am. J. Fish. Manag. 29, 231–236 doi: 10.1577/M06-050.1
Flores-Pérez, A. (2017). Biología reproductiva de la almeja roñosa, Chione californiensis (Broderip, 1735) en bahía Magdalena, B.C.S., México. (dissertation/ master's thesis), IPN-CICIMAR, La Paz, Mexico.
Funes-Rodríguez, R., Gómez-Gutiérrez, J., and Palomares-García, R. (2007). Estudios Ecológicos en Bahía Magdalena. Centro Interdisciplinario de Ciencias Marinas. (La Paz: Instituto Politécnico Nacional), 311.
Gárate-Lizárraga, I., and Siqueiros-Beltrones, D. A. (1998). Time variation in phytoplankton assemblages in a subtropical lagoon system after the 1982–1983 “El Niño” event (1984 to 1986). Pac. Sci. 52, 79–97.
Gárate-Lizárraga, I., Verdugo-Díaz, G., and Okolodkov, Y. B. (2016). “Florecimientos algales nocivos en la costa occidental de Baja California Sur,” in Florecimientos Algales Nocivos en México, eds E. García-Mendoza, S. I. Quijano-Scheggia, A. Olivos-Ortiz, and E. J. Núñez-Vázquez (Ensenada: CICESE), 44–59.
Gárate-Lizárraga, I., Verdugo-Díaz, G., and Siqueiros-Beltrones, D. A. (2001). Variations in phytoplankton assemblages during 1988-1989 in a subtropical lagoon system on the west coast of Mexico. Oceanides 1–16.
García-Domínguez, F., García-Melgar, G., and González-Ramírez, P. (1993). Ciclo reproductivo de la almeja roñosa Chione californiensis (Broderip, 1835) en Bahía Magdalena, B.C.S., Mexico. Cienc. Mar. 19, 5–28. doi: 10.7773/cm.v19i1.923
García-Domínguez, F. A., De Haro-Hernández, A., García-Cuellar, A., Villalejo-Fuerte, M., and Rodríguez-Astudillo, S. (2008). Ciclo reproductivo de Anadara tuberculosa (Sowerby, 1833) (Arcidae) en Bahía Magdalena, México. Rev. Biol. Mar. Oceanogr. 43, 143–152. doi: 10.4067/S0718-19572008000100015
Genitsaris, S., Stefanidou, N., Sommer, U., and Moustaka-Gouni, M. (2019). Phytoplankton blooms, red tides and mucilaginous aggregates in the urban Thessaloniki Bay, Eastern Mediterranean. Diversity 11:136. doi: 10.3390/d11080136
Gómez del Prado-Rosas, M. C. (1984). Presencia de Echinocephalus pseudoncinatus (Nematoda: Gnathostomidae) en Heterodontus francisci (Pices: Elasmobranchii), en Baja California Sur, México. Zoo 55, 13–28.
Gómez-Valdez, M. M. (2019). Características de la biología reproductiva de dos especies de callos de hacha de importancia comercial en Laguna Ojo de Liebre, Baja California Sur: Atrina maura y Pinna rugosa. (dissertation/ doctoral thesis), Centro de Investigaciones Biológicas del Noroeste, La Paz, México. Available online at: https://cibnor.repositorioinstitucional.mx/jspui/handle/1001/1745 (accessed January 28, 2021).
Gómez-Valdivia, F., Parés-Sierra, A., and Flores-Morales, A. L. (2015). The Mexican Coastal Current: a subsurface seasonal bridge that connects the tropical and subtropical Northeastern Pacific. Continent. Shelf Res. 110, 100–107. doi: 10.1016/j.csr.2015.10.010
Góngora-Gómez, A. M., García-Ulloa, M., Hernández-Sepúlveda, J. A., Domínguez-Orozco, A. L, and Sainz-Hernández, J.C. (2016). Crecimiento y supervivencia del callo de hacha Atrina maura (Bivalvia: Pinnidae) cultivado en la costa sureste del Golfo de California, México.Growth and survival of pen shell Atrina maura (Bivalvia: Pinnidae) cultured in the southeastern coast of the Gulf of California, Mexico. Rev. Biol. Mar. Oceanogr. 51, 665–673. doi: 10.4067/S0718-19572016000300017
González-Peláez, S. S. (2013). Almeja generosa de Bahía Magdalena: dinámica poblacional y consideraciones para el manejo. (dissertation/ doctoral thesis), Centro de Investigaciones Biológicas del Noroeste, La Paz, Mexico.
González-Peláez, S. S., Leyva-Valencia, I., Pérez-Valencia, S., and Lluch-Cota, D. B. (2013). Distribution limits of the geoduck clams Panopeagenerosa and P. globosa on the Pacific coast of Mexico. Malacologia 56, 85–94. doi: 10.4002/040.056.0206
González-Peláez, S. S., Morales-Bojórquez, E., Lluch-Cota, D. B., Lluch-Cota, S. E., and Bautista-Romero, J. J. (2015). Modeling geoduck growth: multimodel inference in Panopea globosa from the southwestern Baja California Peninsula, Mexico. J. Shellfish Res. 34, 101–112. doi: 10.2983/035.034.0113
Hernández-Almeida, O. U., Estrada-Gutiérrez, K. M., Siqueiros-Beltrones, D. A., and Inda-Díaz, E. A. (2019). Composición de especies de diatomeas en la dieta in situ del ostión de placer Crassostrea corteziensis en un sistema estuarino. Hidrobiológica 29, 109–127. doi: 10.24275/uam/izt/dcbs/hidro/2020v29n3/Hernandez
Hernández-Carmona, G., Serviere-Zaragoza, E., Riosmena-Rodríguez, R., and Sánchez-Rodríguez, I. (2007). “Flora marina del sistema lagunar de Bahía Magdalena-Bahía Almejas,” in Estudios Ecológicos en Bahía Magdalena. Centro Interdisciplinario de Ciencias Marinas, eds R. Funes-Rodríguez, J. Gómez-Gutiérrez, and R. Palomares-García (La Paz: Instituto Politécnico Nacional), 311.
Hernández-Díaz, A., and Bückle-Ramírez, L. F. (1996). Gonadal cycle of Pteria sterna (Gould 1851) (Mollusca, Bivalvia) in Baja California, Mexico. Cienc. Mar. 22, 495–509. doi: 10.7773/cm.v22i4.868
Hernández-Olalde, L., García-Domínguez, F., Arellano-Martínez, M., and Ceballos-Vázquez, B. P. (2007). Reproductive cycle of the pearl oyster Pteria sterna (Pteriidae) in the Ojo de Liebre lagoon, B.C.S., México. J. Shellfish Res. 26, 543–548. doi: 10.2983/0730-8000(2007)26[543:RCOTPO]2.0.CO;2
Inteligencia Pública EDF de México (2019). Impacto Social de la Pesca Ribereña en México: Propuestas para impulsar el bienestar social en el sector pesquero. CDMX: EDF de México. Available online at: https://mexico.edf.org/sites/mexico.edf.org/files/ImpactoSocialdelaPescaenMexico.pdf (accessed November 05, 2020).
Jeffrey, S. W., and Humphrey, G. F. (1975). New spectrophotometric equations for determining chlorophylls a, b, c and c2 in higher plants, algae and natural phytoplankton. Biochem. Physiol. Pflanz. 167, 191–194. doi: 10.1016/S0015-3796(17)30778-3
Jiménez-Quiroz, M. C., Cervantes-Duarte, R., Funes-Rodríguez, R., Barón-Campis, S. A., García-Romero, F-de-J., Hernández-Trujillo, S., et al. (2019). Impact of “The Blob” and “El Niño” in the SW Baja California Peninsula: Plankton and Environmental Variability of Bahia Magdalena. Front. Mar. Sci. 6:25. doi: 10.3389/fmars.2019.00025
Jiménez-Quiroz, M. C., Martell-Dubois, R., Cervantes-Duarte, R., and Cerdeira-Estrada, S. (2021). Seasonal pattern of the chlorophyll-a in a coastal lagoon from the southern Baja California (Mexico), described with in situ observations and MODIS-Aqua imagery. Oceanologia 63, 329–342. doi: 10.1016/j.oceano.2021.03.003
Koutsoubas, D., Arvanitidis, C., Dounas, C., and Drummond, L. (2000). Community structure and dynamics of the molluscan fauna in a Mediterranean lagoon (Gialova lagoon, SW Greece). Belgian J. Zool. 130, 131–138.
Kug, J. S., Vialard, J., Ham, Y.-G., Yu, J.-Y., and Lengaigne, M. (2021). “ENSO Remote forcing: influence of climate variability outside the tropical Pacific,” in El Niño Southern Oscillation in a Changing Climate, eds M. J. McPhaden, A. Santoso, and W. Cai (Hoboken, NJ: John Wiley & Sons), 528. doi: 10.1002/9781119548164.ch11
Lagos, N. A., Benítez, S., Duarte, C., Lardies, M. A., Broitman, B. R., Tapia, C., et al. (2016). Effects of temperature and ocean acidification on shell characteristics of Argopecten purpuratus: implications for scallop aquaculture in an upwelling-influenced area. Aquacult. Environ. Interact. 8, 357–370. doi: 10.3354/aei00183
Larios-Castro, E. (2020). Escenarios de manejo pesquero de la almeja de sifón (Panopea globosa) en Bahía Magdalena, Baja California Sur, México (dissertation/master's thesis). Centro de Investigaciones Biológicas del Noroeste, La Paz, México.
Lluch-Belda, D. (2000). “Centros de actividad biológica en la costa occidental de Baja California,” in BAC: Centros de Actividad Biológica del Pacífico mexicano, eds D. Lluch-Belda, J. Elorduy-Garay, S. E. Lluch-Cota and G. Ponce-Díaz (La Paz: Centro de Investigaciones Biológicas del Noroeste), 367.
López-Rocha, J. A., Ceballos-Vázquez, B. P., García-Domínguez, F. A., Arellano-Martínez, M., Villalejo-Fuerte, M. A., and Romo-Piñera, K. (2010). The squalid callista Megapitaria squalida (Bivalvia: Veneridae) fishery in Baja California Sur, Mexico. Hidrobiológica 20, 230–237. Available online at: https://hidrobiologica.izt.uam.mx/index.php/revHidro/article/view/818
Lorrain, A., Paulet, Y.-M., Chauvaud, L., Savoye, N., Nezan, E., and Guerin, L. (2000). Growth anomalies in Pecten maximus from coastal waters (Bay of Brest, France): relationship with diatom blooms. J. Mar. Biol. Assoc. 80, 667–673. doi: 10.1017/S0025315400002496
Luquin-Covarrubias, M. A., Morales-Bojórquez, E., González-Peláez, S. S., Hidalgo-de-la-Toba, J. A., and Lluch-Cota, D. B. (2016). Modeling of growth depensation of geoduck clam Panopea globosa based on a multimodel inference approach. J. Shellfish Res. 35, 379–387. doi: 10.2983/035.035.0212
Maeda-Martínez, A. N., Sicard, M. T., Carvalho, L., Lluch-Cota, S. E, and Lluch-Cota, D. B. (2000). “Las poblaciones de almeja catarina Argopecten ventricosus en el centro de actividad biológica de Bahía Magdalena, México,” in BAC: Centros de Actividad Biológica del Pacífico Mexicano, eds D. Lluch-Belda, J. Elorduy-Garay, S. E. Lluch-Cota and G. Ponce-Díaz (La Paz: Centro de Investigaciones Biológicas del Noroeste), 219–228.
Magadán-Revelo, L. D. (2015). Evaluación del cooperativismo pesquero artesanal en Bahía Magdalena, BCS y su incorporación a un sistema de información geográfica. (dissertation/ master's thesis), Colegio de Postgraduados, Texcoco, Mexico.
Martínez-Martínez, S. T., and González-Laxe, F. (2016). La construcción de la política pesquera en México: Una mirada desde el campo geográfico. Atlantic Rev. Econ. 2, 1–27.
Massó-Rojas, J. A. (1996). “Pesquería de almeja catarina,” in Estudio potencial pesquero y acuícola de Baja California Sur, eds M. Casas and G. Ponce (La Paz: SEMARNAP), 71–85.
McAnally-Salas, L., and Valenzuela-Espinoza, E. (1990). Growth and survival of larvae of the pearl oyster Pteria sterna under laboratory conditions. Cienc. Mar. 16:4. doi: 10.7773/cm.v16i4.712
Miralto, A., Barone, G., Romano, G., Poulet, S. A., Russo, G. L., Buttino, I., et al. (1999). The insidious effect of diatoms on copepod reproduction. Nature 402, 173–176. doi: 10.1038/46023
Morales-Gómez, A. A. (2013). Algunos aspectos reproductivos de la almeja gigante Panopea generosa de Bahía Magdalena Baja California Sur, México. (dissertation/ master's thesis), UNAM- FES-Zaragoza. Available online at: https://repositorio.unam.mx/contenidos/310649 (accessed November 05, 2020).
Morales-Zárate, M. V. (2006). Patrones de reclutamiento y deriva larvaria de la almeja catarina Argopecten ventricosus en el complejo lagunar de Bahía Magdalena-Almejas, B.C.S. México. (dissertation/ doctoral thesis), Centro de Investigaciones Biológicas del Noroeste, La Paz, México. Available online at: http://cibnor.repositorioinstitucional.mx/jspui/handle/1001/397 (accessed November 05, 2020).
Müller, M. N., Mardones, J. I., and Dorantes-Aranda, J. J. (2020). Editorial: harmful algal blooms (HABs) in Latin America. Front. Mar. Sci. 7:34. doi: 10.3389/fmars.2020.00034
Naito, K., Tanabe, A., Itakura, S., Yamaguchi, M., and Imai, I. (2011). Evaluation of major nutrients regulating the growth of diatoms in Harima Nada, the Seto Inland Sea, Japan Bull. Fish. Sci. Hokkaido Univ. 61, 5–12. Available online at: http://hdl.handle.net/2115/47535
Narchi, E., Dominguez, W., and Domínguez-Armenta, D. (2018). El ocaso de la abundancia: pescadores y pesquerías en Bahía Magdalena en el siglo XXI. Relaciones Estudios de Historia y Sociedad 39, 167–198. doi: 10.24901/rehs.v39i153.394
Nishikawa, T., Tarutani, K., and Yamamoto, T. (2009). Nitrate and phosphate uptake kinetics of the harmful diatom Eucampia zodiacus Ehrenberg, a causative organism in the bleaching of aquacultured Porphyra thalli. Harmful Algae 9, 563–567. doi: 10.1016/j.hal.2010.04.007
NOAA (2021). NOAA National Centers for Environmental Information, State of the Climate: Global Climate Report for Annual 2020. Available online at: https://www.ncdc.noaa.gov/sotc/global/202013 (accessed March 15, 2021).
Northern Economics Inc. (2009). Valuation of ecosystem services from shellfish restoration, enhancement and management: a review of the literature. Prepared Pacific Shellfish Inst. 58. Available online at: http://www.pacshell.org/pdf/ShellfishEcoServices.pdf (accessed January 28, 2021).
Ojeda-Ruiz de la Peña, M. A., and Ramírez-Rodríguez, M. (2012). Interacciones de pesquerías ribereñas en Bahía Magdalena-Almejas, Baja California Sur. Región Sociedad 24, 189–204. doi: 10.22198/rys.2012.53.a167
Olmos-Martínez, E. (2012). Evaluación de la política pública para el sector pesquero en el noroeste mexicano: análisis económico y prospective. Polis Revista de la Universidad Bolivariana 11, 423–440. doi: 10.4067/S0718-65682012000200020
Osuna-Castro (2012). Administración de la pesquería de almeja catarina en el complejo lagunar Bahía Magdalena-Almejas, Baja California Sur, México. (dissertation/ master's thesis), El Colegio de la Frontera Norte-CICESE, Tijuana, Mexico. Available online at: https://www.colef.mx/posgrado/tesis/2010919/ (accessed November 05, 2020).
Palomares-García, R., Martínez-López, A., De Silva-Dávila, R., Funes-Rodríguez, R., Carballido- Carranza, M. A., Avendaño-Ibarra, R., et al. (2003). Biological effects of El Niño 1997-98 on a shallow subtropical ecosystem: Bahía Magdalena, Mexico. Geofísica Internacional 42, 455–466. Available online at: https://www.redalyc.org/pdf/568/56842317.pdf
Palomares-Sala, I. (2020). Efecto de la temperatura en las tasas de filtración de la nacra (Pinna nobilis). (dissertation/ master's thesis), Universitat Politècnica De València, Escola Politecnica Superior De Gandia, Gandia, Spain.
Pérez-Medina, D. R. (2005). Biología reproductiva de Anadara tuberculosa (Bivalvia: Arcidae) en el estero Santo Domingo BCS, México. (dissertation/ master's thesis), IPN-CICIMAR, La Paz, México. Available online at: https://www.repositoriodigital.ipn.mx/bitstream/123456789/14383/1/perezm1.pdf (accessed January 27, 2021).
Pérez-Ruzafa, A., Campillo, S., Fernández-Palacios, J. M., García-Lacunza, A., García-Oliva, M., Ibañez, H., et al. (2019). Long-term dynamic in nutrients, chlorophyll a, and water quality parameters in a coastal lagoon during a process of eutrophication for decades, a sudden break and a relatively rapid recovery. Front. Mar. Sci. 6:26. doi: 10.3389/fmars.2019.00026
Prado-Ancona, J. D. (1998). Ciclo reproductivo de tres poblaciones de Chione californiensis Broderip, 1835 (Bivalvia: veneridae) en Baja California Sur, México. (dissertation/ master's thesis), IPN-CICIMAR, La Paz, México. Available online at: http://repositoriodigital.ipn.mx/handle/123456789/15025 (accessed March 16, 2021).
Quiñones-Arreola, M. F. (2003). Comparación del patrón reproductivo del Megapitaria squalida (Sowerby, 1835) en la laguna Ojo de Liebre, océano Pacífico y en bahía Juncalito, Golfo de California, B.C.S., México. (dissertation/ master's thesis), IPN-CICIMAR, La Paz, México. Available online at: http://repositoriodigital.ipn.mx/handle/123456789/14987 (accessed March 16, 2021).
Raby, D., Mingelbier, M., Dodson, J. J., Klein, B., Lagadeuc, Y., and Legendre, L. (1997). Food-particle size and selection by bivalve larvae in a temperate embayment. Mar. Biol. 127, 665–672. doi: 10.1007/s002270050057
Romo-Piñera, A. K. (2010). Estrategia reproductiva de Megapitaria squalida (Sowerby, 1835) en dos zonas de Baja California Sur, México. (dissertation/ doctoral thesis), IPN-CICIMAR, La Paz, México. Available online at: http://repositoriodigital.ipn.mx/handle/123456789/14109 (accessed November 20, 2020).
Rosa, M., Ward, J. E., and Shumway, S. E. (2018). Selective capture and ingestion of particles by suspension-feeding bivalve mollusks: a review. J. Shellfish Res. 37, 727–746. doi: 10.2983/035.037.0405
Roy, K., Jablonski, D., and Valentine, J. W. (2000). Dissecting latitudinal diversity gradients: functional groups and clades of marine bivalves. Proc. Royal Soc. Lond. Ser. B 267, 293–299. doi: 10.1098/rspb.2000.0999
Ruiz-Verdugo, C. A., Koch, V., Félix-Pico, E., Beltrán-Lugo, A. I., Cáceres-Martínez, C., Mazón-Suástegui, J. M., et al. (2016). “Scallop fisheries and aquaculture in Mexico,” in Scallops: Biology, Ecology, Aquaculture, and Fisheries. 3rd Edn, eds S. E. Shumway and G. J. Parsons (New York, NY: Developments in Aquaculture and Fisheries Science), 40.
Ruiz-Verdugo, C. A., Ramírez, J. L., Allen, S. K., and Ibarra, A. M. (2000). Triploid catarina scallop Argopecten ventricosus/Sowerby II, 1842: growth, gametogenesis, and suppression of functional hermaphroditism. Aquaculture 5, 13–32. doi: 10.1016/S0044-8486(99)00369-5
Schweers, T., Wolff, M., Koch, V., and Sinsel-Duarte, F. (2006). Population dynamics of Megapitaria squalida (Bivalvia: Veneridae) at Magdalena Bay, Baja California Sur, México. Rev. Biol. Trop. 54, 1003–1017. doi: 10.15517/rbt.v54i3.13973
Sicard, M. T., Maeda-Martinez, A. N., Lluch-Cota, S. E., Lodeiros, C., Roldan-Carrillo, L. M., and Mendoza-Alfaro, R. (2006). Frequent monitoring of temperature: an essential requirement for site selection in bivalve aquaculture in tropical–temperate transition zones. Aquacult. Res. 37, 1040–1049. doi: 10.1111/j.1365-2109.2006.01527.x
Strickland, J. D. H., and Parsons, T. R. (1972). A Practical Handbook of Seawater Analysis. Ottawa, ON: Fisheries Res. Board of Canada.
Sydeman, W. J., Thompson, S. A., García-Reyes, M., Kahru, M., Peterson, W. T., and Largier, J. L. (2014). Multivariate ocean-climate indicators (MOCI) for the central California Current: environmental change, 1990–2010. Progr. Oceanogr. 120, 352–369. doi: 10.1016/j.pocean.2013.10.017
Talgatti, D., Laudares-Silva, R., Garcia, M., Vettorato, B., and Simonassi, J. C. (2010). Considerations about Coscinodiscus wailesii (Diatomeae) on the Santa Catarina island coast, Brazil. Brazil. J. Oceanogr. 58, 353–358 doi: 10.1590/S1679-87592010000400009
Tsai, S. F., Wu, L. Y., Chou, W. C., and Chiang, K. P. (2018). The dynamics of a dominant dinoflagellate, Noctiluca scintillans, in the subtropical coastal waters of the Matsu archipelago. Mar. Pollut. Bull. 127, 553–558. doi: 10.1016/j.marpolbul.2017.12.041
Venrick, E. L., and Hayward, T. L. (1984). Determining chlorophyll on the 1984 CALCOFI surveys. CalCOFI Rep. XXV, 74–79.
Verdugo-Díaz, G. (1997). Cambios estacionales del fitoplancton y de la composición bioquímica del material orgánico particulado en Bahía Concepción, B.C.S. (dissertation/ master's thesis), IPN-CICIMAR, La Paz, México. Available online at: http://repositoriodigital.ipn.mx/handle/123456789/14822 (accessed February 10, 2021).
Zaitsev, O., Montante, O. S., and Saldívar-Reyes, M. (2010). Variaciones estacionales de la estructura termohalina en el sistema lagunar de Bahía Magdalena-Bahía Almejas y el mar adyacente. Cienc. Mar. 36, 413–432. doi: 10.7773/cm.v36i4.1668
Keywords: fisheries' history, overexploitation, anomalous warming, HABs, fisheries regulations
Citation: Jiménez-Quiroz MdC, Barrón-Barraza FJ, Cervantes-Duarte R and Funes-Rodríguez R (2021) Environmental Considerations for the Management of the Bivalve Fisheries of Bahía Magdalena (Mexico), a Coastal Lagoon at the Southern End of the California Current. Front. Mar. Sci. 8:682148. doi: 10.3389/fmars.2021.682148
Received: 17 March 2021; Accepted: 31 August 2021;
Published: 14 October 2021.
Edited by:
Cesar A. Salinas-Zavala, Centro de Investigación Biológica del Noroeste (CIBNOR), MexicoReviewed by:
Monica Cristina Rodriguez Palacio, Metropolitan Autonomous University, MexicoDaniel B. Lluch-Cota, Centro de Investigaciones Biológicas del Noroeste S.C., Mexico
Copyright © 2021 Jiménez-Quiroz, Barrón-Barraza, Cervantes-Duarte and Funes-Rodríguez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: María del Carmen Jiménez-Quiroz, Y2FybWVuLmpxdWlyb3pAaW5hcGVzY2EuZ29iLm14