 José Pontón-Cevallos1,2,3*
José Pontón-Cevallos1,2,3* Nathaly Ramírez-Valarezo4
Nathaly Ramírez-Valarezo4 Mireya Pozo-Cajas4
Mireya Pozo-Cajas4 Gabriela Rodríguez-Jácome5
Gabriela Rodríguez-Jácome5 Gabriela Navarrete-Forero6
Gabriela Navarrete-Forero6 Nicolas Moity5
Nicolas Moity5 Gonzalo Villa-Cox7,8
Gonzalo Villa-Cox7,8 Jorge Ramírez-González5
Jorge Ramírez-González5 María José Barragán-Paladines5
María José Barragán-Paladines5 Jorge R. Bermúdez-Monsalve2,9,10
Jorge R. Bermúdez-Monsalve2,9,10 Peter L. M. Goethals1
Peter L. M. Goethals1- 1Department of Animal Sciences and Aquatic Ecology, Faculty of Bioscience Engineering, Ghent University, Ghent, Belgium
- 2Galapagos Marine Research and Exploration Program (GMaRE), Charles Darwin Foundation and Escuela Superior Politécnica del Litoral, Santa Cruz, Ecuador
- 3Facultad de Ciencias de la Vida, Escuela Superior Politécnica del Litoral, Guayaquil, Ecuador
- 4Facultad de Ciencias Naturales, Universidad de Guayaquil, Guayaquil, Ecuador
- 5Charles Darwin Research Station, Charles Darwin Foundation, Santa Cruz, Ecuador
- 6Instituto de Ecología Aplicada, Universidad San Francisco, Quito, Ecuador
- 7Facultad de Ciencias Sociales y Humanísticas, Centro de Investigaciones Rurales, Escuela Superior Politécnica del Litoral, Guayaquil, Ecuador
- 8Department of Agricultural Economics, Faculty of Bioscience Engineering, Ghent University, Ghent, Belgium
- 9Facultad de Ingeniería Marítima y Ciencias del Mar, Escuela Superior Politécnica del Litoral, Guayaquil, Ecuador
- 10Marine Environment Laboratories, International Atomic Energy Agency, Monaco, Monaco
Mangroves provide multiple ecosystem services to coastal communities and their fisheries. In the Galapagos, mangroves have been primarily prized for their role as nursery grounds; though, the importance of cultural benefits, including the development of knowledge systems among fishers, needs to be further explored. Fishers’ local ecological knowledge (LEK) has been employed in the past to answer questions about the marine environment; not yet about mangroves. In such regard, this ethnoecological approach can contribute to unveil mangrove habitat-dependence relationships among fishery taxa across the life cycle, connectivity among habitats, and productivity spatial patterns across the archipelago. To access fishers’ LEK, we conducted 35 semi-structured interviews and 2 facilitated community workshops comprising 3 activities. A photo-elicitation method allowed us to extract values given to mangroves (i.e., ecosystem services), from which indirect use values were considered more relevant for fisheries. As such, the accumulation of LEK might have been crucial for the establishment of these values among fishers. By using a species-habitat association method, we revealed a high perceived connectivity between mangroves and other coastal and marine habitats in the Galapagos. Although this suggests an overall weak dependence to the ecosystem, fishers’ anecdotes allowed us to recognize novel species occurrences and exclusive habitat uses (e.g., spawning in Centropomus viridis). By applying Social Network Analysis to species-habitat association results, we learned that certain migratory commercial species (e.g., Lutjanus argentiventris, Seriola rivoliana) might constitute critical links in marine trophic webs and across seascapes. Finally, through scoring and participatory mapping methods, we revealed the prominence of western islands’ mangroves for fishery productivity, which were linked to biophysical attributes, like forest canopy and freshwater. Yet, other locations in central and southeastern islands might also be important, as they hold similar attributes. Interviews to other mangrove users (e.g., naturalist guides, park rangers, scientists; N = 33), and past scientific studies allowed us to triangulate LEK and to find complementarity among knowledge sources. Despite having an explorative focus, our work already showcases the potential of fishers’ LEK in improving mangrove research in the Galapagos, and the arising opportunities for ecosystem-based fisheries management and mangrove protection.
1 Introduction
Mangroves fringing tropical and subtropical coastlines and estuaries are known to provide manifold benefits to coastal communities and the fisheries they depend on (Nagelkerken et al., 2008; Carrasquilla-Henao and Juanes, 2017). Commonly, these benefits are identified and quantified under an ecosystem services framework, which are defined as any positive benefit that wildlife or ecosystems provide to people (Millennium Ecosystem Assessment, 2005; Barbier et al., 2011). From one side, benefits can result from direct provisioning services, including the supply of food and timber resources (Whitfield, 2017). From another perspective, they derive from indirect supporting and regulating services, mainly the establishment of nursery grounds, but also hazard protection and water sanitation (Igulu et al., 2014). Additionally, tangible (e.g., recreation, ecotourism) and abstract cultural services (e.g., knowledge systems, social relations; Lal, 2003; Nagelkerken et al., 2008) are important for coastal communities depending on small-scaled fisheries and should be accounted when assessing mangrove-fishery linkages from an economic, social, or ecological perspective.
Although the importance of mangroves for fishery production has been globally acknowledged (Alongi, 2002; Lee et al., 2014; Whitfield, 2017; Sheaves et al., 2020), to date, most studies have focused on the mechanisms governing mangrove nursery function and the effects in adjacent fishery production (e.g., penaeid shrimps); with other benefits of the ecosystem receiving less attention (Manson et al., 2005; Carrasquilla-Henao and Juanes, 2017). Although global meta-analyses suggest a positive relationship between mangrove area and fish catch and yield (e.g., see Baran and Hambrey, 1999; Manson et al., 2005; Blaber, 2007; Aburto-Oropeza et al., 2008; Carrasquilla-Henao and Juanes, 2017), there is little evidence of this direct linkage at smaller scales. From this, it is expected that habitat dependence should vary between and within taxa (i.e., across life cycle), as response of regional (e.g., climate) and local environmental conditions (e.g., tide dynamics, turbidity, geomorphology; Castellanos-Galindo and Krumme, 2013). This is particularly relevant if mangroves constitute part of a mosaic of several coastal, estuarine, wetland and freshwaters habitats, which can provide the same opportunities as nursery grounds and for other ecosystem functions (Manson et al., 2005; Blaber, 2007; Benzeev et al., 2017). In such case, mangrove habitat-dependence relationships should be assessed within the context of complex seascape connectivity, particularly for highly-commercial species (e.g., snappers, groupers), which conduct ontogenic migrations from coastal nursery grounds to adult offshore areas (Sheaves, 2009).
Considering the ecological and spatial complexities driving mangrove-fishery linkages, and the weight of cultural ecosystem services in shaping this relationship at the local scale, it is thoughtful to discuss the opportunities that ethnoecological studies, particularly those addressing a Local Ecological Knowledge (LEK) approach, could bring for mangrove research (see Rönnbäck et al., 2007; Carrasquilla-Henao et al., 2019). LEK is defined as contemporary and historical knowledge about the environment based on lived experience that is not restricted by ethnicity or heritage to indigenous people exclusively (which in that case is entitled Traditional Ecological Knowledge - TEK; Tomaselli et al., 2018). As for the marine realm, LEK is provided by direct users of the oceans (e.g., fishers), which accumulate knowledge of adjacent coastal and marine ecosystems through observation, narratives, storytelling, interactions, failure, among others (Berkes et al., 2000; Pilgrim et al., 2008). On the contrary, indirect users (e.g., scientists, government and conservation officers) rely on science and management knowledge (SMK) acquired through formal education, research and conservation practice (Butler et al., 2012).
Ultimately, LEK and SMK provide independent sources of information that can be combined to increase confidence and depth of knowledge (see Drew, 2005; Butler et al., 2012; Madsen et al., 2020). While SMK is considered neutral and objective, LEK is rooted within the history, geography and culture of a particular site without being outdated, static or regressive (Pilgrim et al., 2007; Roux et al., 2019). Several studies in marine science have combined both knowledge sources for applications in ecosystem-based fisheries management, environmental protection, and governance (see Murray et al., 2005; Wilson et al., 2006; Raymond et al., 2010; Figus, 2018). The value of LEK is especially crucial in data-poor regions, where research and monitoring are challenging and limited to smaller spatio-temporal windows to allow the unravelling of causal relationships and trends (Stave et al., 2007; Brook and McLachlan, 2008); this being the case of many tropical oceanic islands (Jupiter et al., 2014).
The Galapagos Islands represent an interesting case study to access and explore fishers’ LEK concerning mangrove-fishery linkages, as other studies have portrayed them as social-ecological laboratory to investigate the early links and interactions between social and natural systems (González et al., 2008; Pontón-Cevallos et al., 2020; Riascos-Flores et al., 2021). The archipelago is surrounded by the Galapagos Marine Reserve (GMR), a multi-use marine protected area in which small-scale fisheries, tourism, research, management and conservation sectors depict competing interests and alliances (Heylings et al., 2002; Jones, 2013). Galapagos mangrove forests and fringing vegetation are in advanced successional stages, given strict environmental protections in place banning logging activity and shrimp farming, which have led to an increase in mangrove cover over the last decade (Moity et al., 2019). Although people in the Galapagos are not indigenous and have only formed bonds with the oceans since the late 18th century (thus, the term TEK cannot be applied), most fishing families hold strong cultural connections with mainland Ecuadorian urban centers (Jones, 2013; Schiller et al., 2013). This long history of natural experiences and interactions with the ocean has resulted in their progressive accumulation of local knowledge about fishing gears and techniques, marine ecosystems, and species’ life histories (Quiroga and Orbes, 1964; González et al., 2008; Schiller et al., 2013; Rodríguez-Jacome, 2020).
The integration of local knowledge in GMR decision-making, as well, as the promotion of mutual respect and collective learning among different knowledge owners, constitute some of the strategies that have been applied to improve governance in the marine reserve (Jones, 2013). However, despite the implementation of participatory monitoring surveys along with the fishing sector (Castrejón and Charles, 2013; Jones, 2013), there are few examples in which fishers’ LEK was integrated with SMK in marine sciences (see Burbano et al., 2014; Rodríguez-Jacome et al., 2019; Cavole et al., 2020). Concerning mangrove ecosystems, although SMK has contributed to highlight the ecological and economic importance of the habitat for several highly commercial species (e.g., Galapagos sailfin grouper - Mycteroperca olfax, snappers, mullets; Tanner et al., 2019; Fierro-Arcos et al., 2021; Plumlee et al., 2022), there are, to date, multiple information gaps that can be potentially tackled by addressing a fishers’ LEK approach. One of these, relates to the elucidation of mechanisms and factors driving habitat-dependence relationships across and within fishery taxa, particularly in the context of connectivity in the Galapagos. Furthermore, there is a need to reveal spatial patterns of fishery productivity across the archipelago at different scales (e.g., whole RMG, bioregions), and the environmental factors driving these patterns. However, cognitive processes around mangrove ecosystems need to be understood first, in order to foresee the applicability of fishers’ LEK in future participative research programs.
In order to explore the contribution of fishers’ LEK in unraveling mangrove-fishery linkages in the Galapagos, we utilized a qualitative approach with fishing communities, comprising two survey instruments: interviews and facilitated community workshops. We specifically explored three research questions: 1) Which and how fishery species associate to mangrove habitats in relation to life cycle and environmental conditions? 2) To what degree mangroves are connected by species movements to other coastal and marine habitats in the Galapagos, and which fishery species constitute critical links for seascape connectivity due to complex trophic interactions in mangroves? 3) Which are the spatial relationships of fishery productivity across the archipelago linked to biophysical attributes of the mangrove ecosystem? To explore these questions, we first dissected mangrove-fishery linkages under an ecosystem services framework to examine how LEK has contributed to shape values towards the ecosystem, and to reason whether it constituted a suitable approach to answer our research questions. We also triangulated and complemented LEK data with interviews to other mangrove users (OMU; e.g., naturalist guides, park rangers, scientists) and published and unpublished ecological data. We finally discussed the opportunities and challenges of addressing a fishers’ LEK approach in future mangrove research in the islands, including its complementarity with SMK and relevance in answering applied research questions.
2 Materials and Methods
2.1 Study Area
The Galapagos Archipelago lies on the Tropical Eastern Pacific about 1000 km off the coast of mainland Ecuador and encompasses 18 main islands and over 200 islets and rocks of volcanic origin (Snell et al., 1996). The coastal and oceanic waters of the archipelago are protected since the establishment of the (GMR) in 1998, which delimited an area of 138,000 km2. Within the marine reserve, tourism and fishing are the main activities contributing to the local economy, each accounting for 52% and 12% (agriculture and livestock included) of the employment in the population, respectively (CGREG, 2021). Fishing activity is only allowed for licensed artisanal fishers, for commercial, subsistence and recreational purposes in designated areas according to the zoning scheme, by which 99% of the GMR is open to fishing (Moity, 2018). Contrarily, industrial fishing has been completely banned since the marine reserve creation. Main fishing ports in the Galapagos are in Santa Cruz (Puerto Arora), San Cristobal (Puerto Baquerizo Moreno), and Isabela (Puerto Villamil), in which fishers are organized in fishing cooperatives and associations (Castrejón, 2011).
Mangroves forests in the Galapagos comprise three species (red – Rhizophora mangle, white – Avicennia germinans, and black – Laguncularia racemosa), in addition to the button mangrove (Conocarpus erectus) which grows as transitional vegetation with the arid coastal zone (Wium-Andersen and Hamann, 1986; Moity and Delgado, 2018). Mangrove forests cover around 35% of the coastline of the archipelago, although most patches exhibit an underdeveloped condition (~85% of mangrove patches have <0.5 ha), due to a rocky coastline mostly exposed to wave activity, a dry climate, and the lack of permanent estuaries and rivers (Moity et al., 2019). The distribution of mangroves is not homogenous across the archipelago. Geologically younger islands (e.g., Isabela, Fernandina) have more mangrove forests than older ones; with Isabela being the island with more mangroves per length of coastline (Moity et al., 2019). Regarding the distribution of mangroves per bioregion (sensu Moity, 2019), 90% of mangrove forests are in the Western and Central-Southeastern (CSE) bioregions (Moity et al., 2019).
2.2 Data Collection
To access knowledge, perceptions and values from fishers we employed a mixed-method qualitative approach comprising two stages: semi-structured qualitative interviews and facilitated community workshops (Newing, 2011). Interviews allowed us, in the first place, to reveal main mangrove-fishery linkages occurring in the Galapagos, and to pinpoint specific knowledge aspects that needed further elaboration and triangulation; these being further explored during workshops. Prior to initiating these activities, we recurred to strategies and techniques (e.g., participant observation, informal conversations, meetings) to prompt multi-stakeholder engagement (e.g., local government, fishing cooperatives, research institutions, conservation organizations) in the conduction of this study, and to identify key informants within fishing communities and other sectors of the society (see Supplementary Material Section 1.1.1.).
During the first stage (November 2019 to February 2020), we conducted semi-structured qualitative interviews to fishers of Santa Cruz, San Cristobal and Isabela, mainly during visits to landing sites. Questionnaires included a mix of open-ended and closed-ended questions aiming to reveal 1) main biophysical attributes, functions and benefits of mangroves (i.e., ecosystem services), 2) key species-habitat associations (of both target and non-target species), including predominant life stages, habitat uses and linkages with environmental conditions, and 3) ‘important’ mangrove locations (e.g., islands, sites within islands) harboring these species. Our universe of participants encompassed both active or retired fishers from any age (or time of experience), but whose primary economic activity is or was recently fishing. We used a non-probabilistic sampling design, in which a chain referral system with snowball sampling was applied (Goodman, 1961). This method allowed us to find potential new respondents and has been proved effective in small islands, like the Galapagos (e.g., Burbano et al., 2014; Cavole et al., 2020). Further details about methods for interviews, including questionnaire questions (Table S1), are presented in Supplementary Material Section 1.1.2.
In order to complement and triangulate knowledge about species-habitat associations and spatial patterns, we conducted interviews to OMU (July to October 2020), comprising local naturalist guides, park rangers and scientists. We selected respondents based on on-ground (rather than theoretical) knowledge, and experience in research, monitoring, tourism or management activities. Interviews were conducted by telephone, and in some cases, finished by e-mail, by employing similar questionnaires to those of fishers (see Supplementary Material Section 1.1.3).
During the second stage, we conducted two facilitated community workshops in Santa Cruz (April 16th, 2021) and Isabela (April 20th, 2021) islands, in which three group activities were performed to, respectively: 1) explore linkages among mangrove ecosystem services and the role of LEK in establishing values towards the ecosystem; 2) to complement and triangulate species-habitat associations in mangroves, and to explore the role of mangrove in the connectivity of these species, and to 3) characterize mangrove sites in regard to biophysical attributes linked to fishery productivity. Group activities were each accompanied by a facilitator and other members of the audience who acted as observers (i.e., scientists, managers and members of the local community comprising 20% of total assistants). Workshops were held in two, rather than three islands, due to a lack of time. Thus, we prioritized to contrast Isabela and Santa Cruz, as fishers from the former have more contact with the prominent ecosystem conditions in the Western and Elizabeth bioregions (see Section 1), while those from the latter, with the Central-Southeastern bioregion (which also comprises San Cristobal).
For group activities, mangrove ecosystem services were further extracted by using a photo-elicitation approach (see Patton, 2002; Richard and Lahman, 2015), in which a set of 20 photograph was handed to each group (Table S2) to build a photo-collage, which was then presented by participants to the rest of the audience (Figure S1). Species-habitat associations to mangroves and connectivity was delved and triangulated (i.e., to untangle potential taxonomic uncertainties), by asking groups to pin illustrations of fishery species to different coastal and marine habitats depicted in a digital representation of a Galapagos seascape (i.e., mangroves, shallow reefs, sandy bays and bottoms, shallow seamounts, epipelagic waters, vertical walls and deep sea; Figure S2). Finally, characterization of mangrove sites was conducted using semi-quantitative scoring during the Santa Cruz workshop, in which groups were asked to score 10 ecosystem variables on 13 sites based on ordinal criteria (i.e., 1, 2 or 3), which represented different states of these variables (‘attributes’) along an environmental continuum (Table S3; Table S4). Some of these sites represented most-salient results from interviews (‘important’ sites; N = 7), while others represented sites near or easily accessible from urbanized areas (‘impacted’ sites; N = 6). Instead, in the Isabela workshop, we employed a participatory mapping approach, in which groups represented and described attributes in these 13 sites and others that they found relevant (Figure S3). We adjusted the methodology from the first to the second workshop, as we found that fishers had problems understanding some variables and scoring criteria employed in the semi-quantitative approach (see Section 4.4). Further details about the group work methods can be found in Supplementary Material Section 1.1.4.
2.3 Data Analysis
First, to explain linkages among biophysical attributes, functions and benefits of mangroves under an ecosystem service framework, we employed Cultural Domain Analysis (CDA). This set of techniques is used to understand how people in a society think about and define their world. CDA is based on the premise that all cultures use a system of categories (‘domains’) to order experience; thus, researchers can determine which items constitute these domains and what values are attached to them (Weller and Romney, 1988; Puri, 2011). For our study, we chose four domains denoting different ecosystem service types (i.e., provisioning, regulating, supporting, and cultural), and then unexclusively assigned all items (keywords) elicited from workshops and interviews to each one of these. Categorization and shared keywords between domains were represented by a Venn diagram. After that, we inspected how LEK might have been critical in establishing perceived linkages among mangrove ecosystem services.
Second, to explore mangrove habitat-dependence relationships across fishery taxa, triangulate knowledge, and assess its complementarity with SMK sources, we consolidated all fishery species (as defined in Molina et al., 2004) reported in mangroves by fishers (i.e., interviews and workshops combined), and OMU, as well as those found in published and unpublished ecological data (Table S7). We highlighted species-habitat associations in which juvenile stages were deemed predominant in mangroves. We also summarized main observations and anecdotes about habitat use in mangroves and movement behavior. Then, we built a dendrogram heatmap to depict patterns of connectivity among coastal and marine habitats in the Galapagos, by pooling target and non-target species. With these data, we also applied Social Network Analysis (SNA) to discern patterns of interaction among species as a result of habitat connectivity. SNA utilizes networks and graph theory to understand social structures; yet has extensively grown across multiple research fields, including biology (Borgatti et al., 2009). A network is composed of a set of nodes (actors) and edges (relationships) that can be represented and analyzed through a variety of mathematical tools (Scott, 2013). For this study, we developed an undirected network representation of species interactions1 (through shared habitats) and grouped them by feeding guild. Based on this representation, we then used estimated hub scores of each species to delimit concentric visual groupings (i.e., using three uniform cutoff intervals), which served us to pinpoint those species that fishers perceive as pivotal in terms of habitat interactions, and thus more relevant for network (ecosystemic) stability. Although other studies have applied a similar approach to analyze species interactions (e.g., Engelhard et al., 2017; Tidbury et al., 2020), they used quantitative rather than qualitative data.
Last, to study spatial patterns in mangrove ecosystems of the Galapagos regarding fishery productivity, we mapped all ‘important’ mangrove locations identified by fishers and OMU during interviews and workshops. Some of these locations were marked on the map by fishers, while others were georeferenced using available databases (CDF, 2018)2. We also mapped biophysical attributes linked to fishery productivity in selected sites (both ‘important’ and ‘impacted’), in which a clear consensus was reached during the semi-quantitative scoring or participatory mapping activities conducted in each workshop (Table S4). These maps were used to investigate spatial and environmental relationships in mangroves. Fishers’ and OMU’s locations were contrasted in terms of spatial distribution, agreement and relative importance given to each location.
R v.4.1.2 (R Core Team, 2021) was used for the implementation of Venn diagram (package ‘ggvenn’; Linlin, 2021), dendrogram heatmap (default package ‘stats’), and Social Network Analysis and representation (package ‘igraph’; Csárdi and Nepusz, 2006). ArcGIS Pro v.2.7 (Esri Inc., 2020) was used to elaborate the maps; whereas Python 3.6. and Adobe Illustrator (Adobe Inc., 2019), to improve visualization.
2.4 Permits and Ethics
Studies and activities involving human participants were conducted under research permits No. PC-41-19, PC-41-20 and PC-41-21 from the Galapagos National Park Directorate, did not not required approval by an ethics committee. Written informed consent for participation was not required for this study in accordance with the national legislation and the institutional requirements. Interviews and workshops were not video- nor voice recorded and conducted only after obtaining informed verbal consent from the participants and assuring them that they could omit questions or end their participation at any stage. Personal identity of participants was kept confidential and anonymous throughout the study.
3 Results
3.1 Sociodemographic Profile of Participants
Qualitative interviews were conducted to 35 artisanal fishers of the Galapagos, from which 18 were held in Santa Cruz, 10 in San Cristobal and 7 in Isabela. 31 of the respondents are currently active fishers (10% of operative fishing population; Vega, 2021), while 4 already retired. None of the respondents were women, as the few found during the visits to landing sites refused to participate. Age of participants ranged from 18 to 81, with a mean of 35 years of experience in either single-species (e.g., lobster, sea cucumber, whitefin) or, more usually, multispecies fisheries (83%). More than half of participants (60%) claimed to have acquired fishing skills locally, while the rest, when living in mainland Ecuador before their arrival to the islands. Furthermore, 43% of participants have additional experience in tourism, agriculture, research, conservation, maritime transport, administrative activities or others. Only 26% mentioned to have at least initiated tertiary education level.
Thirty-three qualitative interviews were conducted to OMU (21 men and 12 women), both by telephone and e-mail, from which 23 comprised naturalist guides, 7 park rangers (active or passive), and 3 scientists. Around 40% of the respondents have participated in environmental research activities, regardless of their main occupation. Finally, 79% mentioned to have completed until tertiary or graduate education level in environmental related fields.
Last, facilitated community workshops from Santa Cruz and Isabela received 14 and 6 local fishers (all men), respectively, with a mean age of 45. Groups in workshops comprised 3-5 fishers with age affinity in the case of Santa Cruz; while in Isabela, they were randomly assigned due to lower assistance.
3.2 Mangrove Ecosystem Services and Local Ecological Knowledge
By using a CDA methodology, we were able to extract 103 keywords depicting biophysical attributes, functions and benefits of mangroves; this allowing us to examine flows and connections between ecosystems services (Figure 1; see Table S5 for a complete list keywords). Other keywords depicted causes and consequences of natural and anthropogenic threats (e.g., marine pollution, habitat loss and disturbance, overfishing, climate-ocean variability), socioeconomic impacts, and responses from society (e.g., coastal clean-ups, reforestation, spatial planning, zoning, tourism regulation); though, these were not further analyzed during this study.
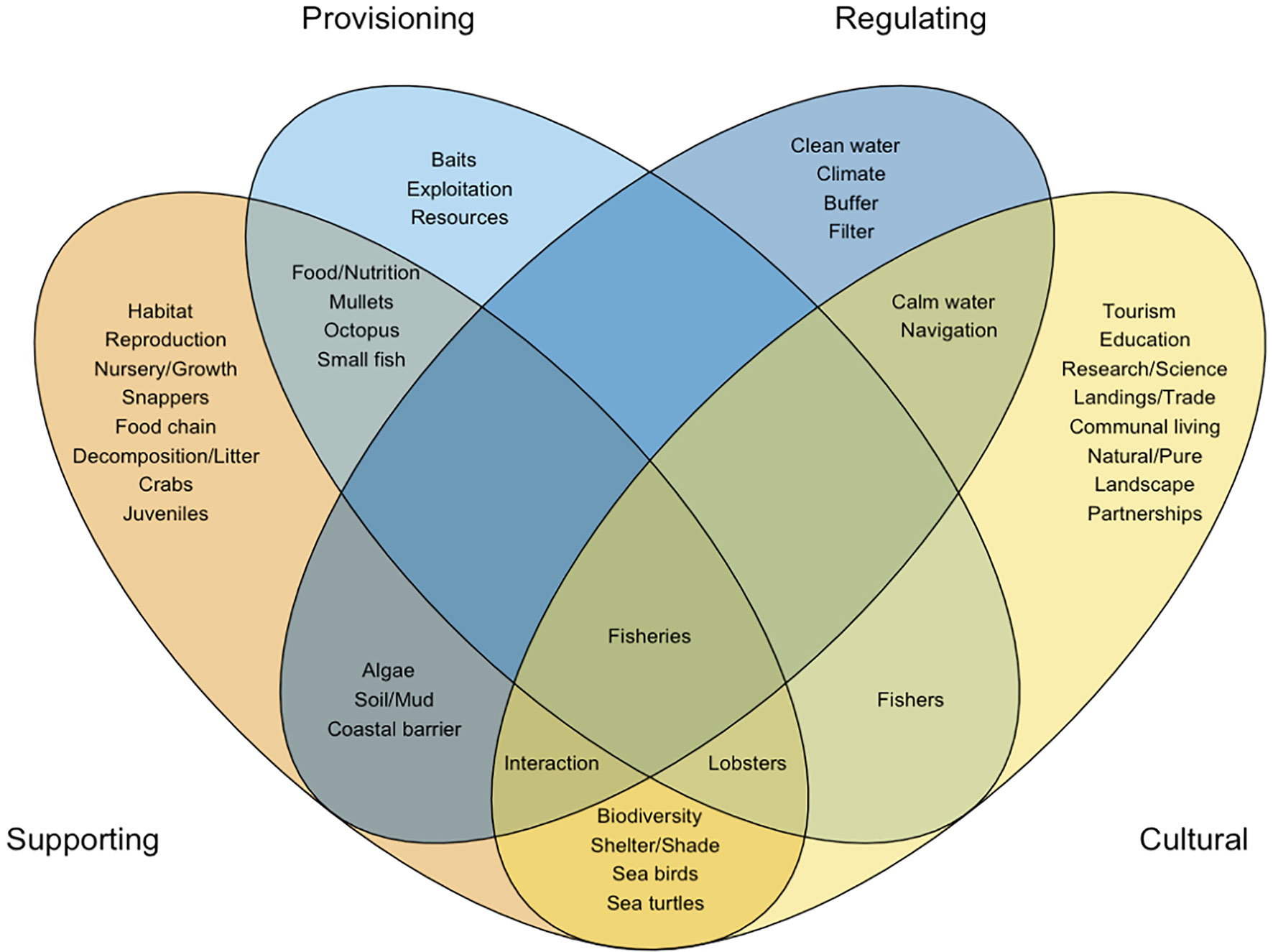
Figure 1 Venn diagram depicting interactions among mangrove ecosystem services represented by keywords extracted from a fishers’ LEK study. Cultural Domain Analysis was used to assign keywords to different ecosystem services categories (‘domains’). Visualization only includes main keywords.
In general, fishers portrayed mangroves as ecosystems that ‘attract marine life’, ‘give life’, or that constitute a ‘source of life’. In relation to value for fisheries, they recognized direct benefits from provisioning services (e.g., supply of food and baits), particularly in regard to subsistence fisheries; though, indirect benefits were more often highlighted and discussed throughout our activities (Figure 1). Participants acknowledged that supporting services relate to habitat functions for fishery species, mainly nursery (i.e., referred to as semilleros in local language, meaning seedbeds), but also reproduction, foraging, and refuge, as well as benefits of nutrient recycling, productivity and trophic webs. Benefits of water sanitation were linked to the facilitation of these supporting services, in addition to those of hazard and erosion control (i.e., regulating services). As such regulating services were deemed relevant for fishery activity, as fishers seek calm, safe and clean waters for vessel navigation, mooring, and landing and commercialization of fish products (Figure 1). Benefits of the provision of shade and shelter were acknowledged in relation to labor and recreation of fishing communities, in addition of being beneficial for wildlife. At the same time, other cultural benefits derived from landscape and opportunities for natural experiences were deemed important for ecotourism, education, research, and management, which constitute sometimes supplementary subsistence activities, but are also essential for social relations (e.g., partnerships) within fishing communities (Figure 1).
As the information and knowledge function of mangroves was valued in the context of fisheries, we examined how LEK might have emerged and adapted within fishing communities and contributed to shape values towards the ecosystem. For one side, recurring presence and high abundance of juvenile stages in mangroves were linked to values towards mangrove nursery function. Additionally, observations and anecdotes of habitat use, migrations and movements among habitats and species interactions among target and non-target species, can explain values towards trophic web and connectivity functions, which have influenced fishers’ perceptions about which species and life stages depend stronger on mangroves, in contrast with other habitats (see Section 3.3 and 3.4). On the other side, knowledge about environmental cycles and conditions on fishery species, and the recognition of certain biophysical attributes in the ecosystem (i.e., mud, crabs, sea birds, forest canopy), were associated to values towards nutrient recycling and productivity functions, which have also influenced fishers’ perceptions of ‘important’ mangrove areas across the archipelago (see Section 3.5).
3.3 Species-Habitat Associations in Mangroves and Nursery Role
Fishers associated numerous fishery species (or taxa) to Galapagos mangrove ecosystems (N = 40), both during interviews and workshops. Species showing the highest relative importance in interviews (i.e., > 38% of respondents) or workshops (i.e., 5 out 5 groups) included snappers (Lutjanus spp.; 6 species pooled), particularly the yellow snapper (L. argentiventris), white snook (Centropomus viridis), mullets (Mugil spp.; 3 species pooled) and green spiny lobster (Panulirus gracilis; Table 1). Other species (or taxa) commonly reported in interviews (i.e., 37-18% of respondents) or workshops (i.e., 4 out of 5 groups) included, Pacific dog snapper (L. novemfasciatus), Galapagos sailfin grouper (referred to as bacalao), Thoburn’s mullet (M. thoburni), salemas (Xenichthys agassizii and Xenocys jessiae), milkfish (Chanos chanos), and herrings (Opisthonema libertate and O. berlangai; Table 1). In addition, fishers also associated several non-target species or taxa to mangroves (N = 25), namely other fishes (e.g., Pacific seahorse - Hippocampus ingens, bullseye puffer - Sphoeroides annulatus), sharks (e.g., whitetip reef shark - Triaenodon obesus), stingrays, sea and aquatic birds, sea lions, and other crustaceans (e.g., shrimps, crabs; Table S6).
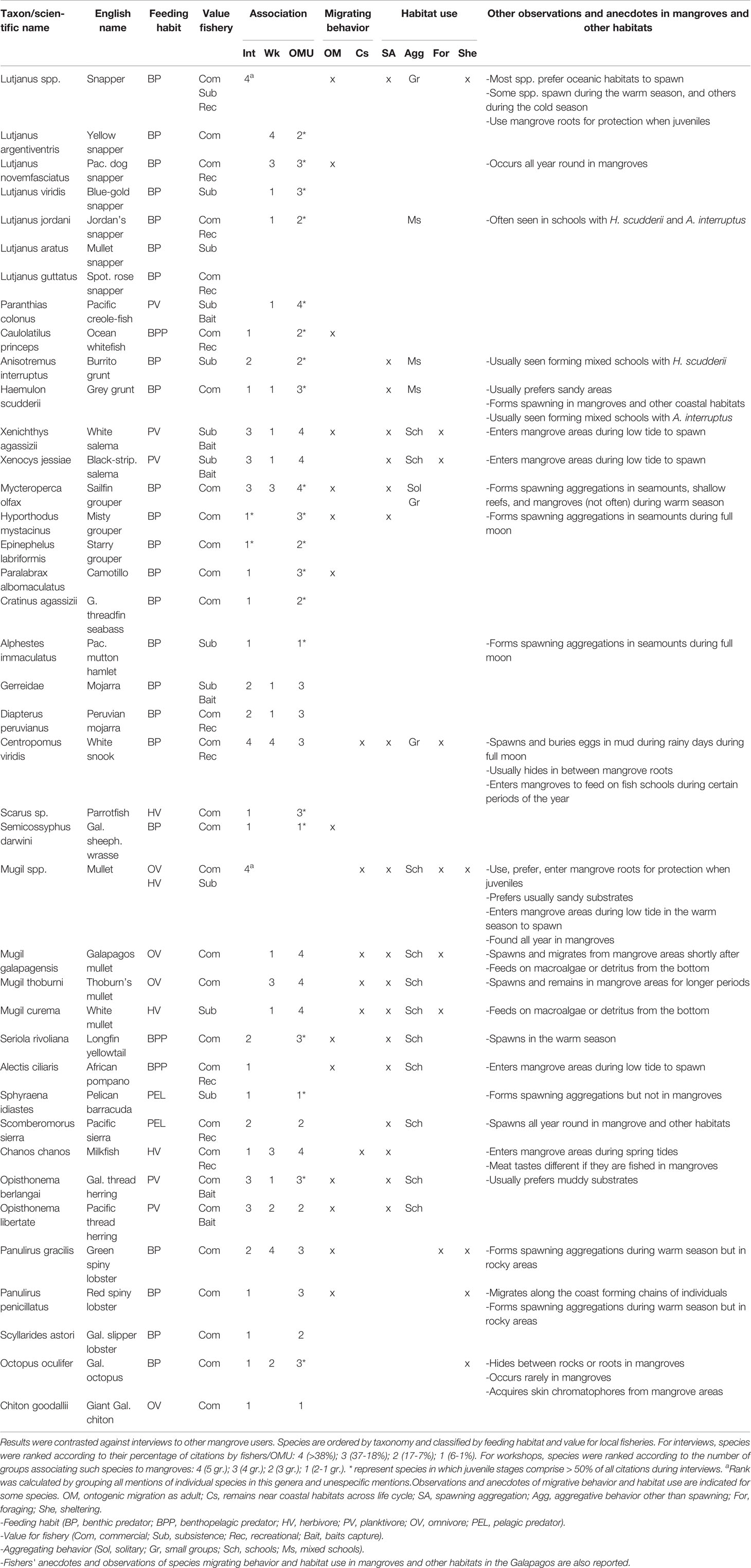
Table 1 Summary of fishery species (or taxa) associated to mangroves by Galapagos fishers during interviews (Int; N = 35), and by groups of fishers during community workshops (Wk; N = 5)- , and by other mangrove users (i.e., naturalist guides, park rangers, scientists; OMU; N = 33).
Although for all fishery species, both adult and juvenile stages were associated to mangroves, the latter stage was predominantly recognized (i.e., > 50% of reports) in species like, snappers, bacalao, starry grouper (Epinephelus labriformis) and misty grouper (Hyporthodus mystacinus; Table 1). On one side, fishers distinguished those fishery species with a marked ontogenic migratory behavior, usually observed solitary or forming small aggregations in mangroves (e.g., bacalao, snappers). On the other side, they distinguished other species remaining near coastal habitats throughout their whole lives (e.g., mullets, salemas, grunts, mojarras), often having shoaling or schooling behaviors; some in association with other species (e.g., grey grunt - Haemulon scudderii and burrito grunt – Anisotremus interruptus; Table 1). In addition, for some non-target species (e.g., sharks, stingrays), juvenile stages were also reported as common in the habitat.
In addition to nursery function, some fishery species in their adult stage were also reported mating or spawning in mangroves (e.g., white snook, mullets, salemas, Pacific thread herring, grunts), foraging on detritus or benthic algae (e.g., green spiny lobster, white mullet - M. curema), or seeking shelter and rest between roots, rocks or tide pools (e.g., Galapagos octopus – Octopus oculifer; Table 1). The effect of environmental cycles (e.g., tides, moon cycle, seasons) and biophysical attributes in the ecosystem (e.g., mud, root systems) were often linked to these habitat uses. In such regard, higher sea temperatures during the warm season and full moon were generally associated with fish mating or spawning behaviors. Species like the white snook, were reported laying and burying their eggs in muddy sea bottoms under these conditions. Tide cycles were linked to periodic movement behavior, as some fishes were reported using spring tides to reach some mangrove areas and then remaining within tide pools (e.g., milkfish, mullets; Table 1).
Interviews conducted to OMU revealed a total of 61 fishery species (or taxa), from which 35 (out of 40) were common with the fishers’ species list (Tables S6, S7). Some highly-commercial species, like the mottled scorpionfish or ‘brujo’ (Pontinus clemensi), yellow fin tuna (Thunnus albacares) and brown sea cucumber (Isostichopus fuscus) were listed in mangroves, yet fishers associated them to other habitats (Table S8). Species like the Pacific creole-fish (Paranthias colonus), grey grunt, salemas, mojarras (Gerreidae), milkfish, and red spiny lobster (P. penicillatus) received higher relative importance than in the case of fishers. Moreover, juvenile stages were predominantly perceived in a higher amount of species; for instance, in grunts, ocean whitefish (Caulolatilus princeps), other serranids, longfin yellowtail (Seriola rivoliana), pelican barracuda (Sphyraena idiastes) and Galapagos octopus (Table 1). Finally, a higher number of non-target species was reported by OMU (Table S6).
Fishers’ species list showed also high similarity with the ones reported in past scientific studies, especially from Fierro-Arcos et al. (2021); 23 species in common; (Table S7), who monitored communities using baited underwater cameras (stereo-BRUVS) and underwater visual censuses across two bioregions and multiple sites across the archipelago. In this study, highly-commercial species, like bacalao, snappers and mullets were also predominantly reported as juveniles. Other studies using gill nets, like Llerena-Martillo et al. (2018) and Suárez-Moncada (2012), showed moderate similarities with our results (i.e., 11 and 13 common species, respectively; Table S7), despite only covering few areas in the CSE bioregion. Important food (e.g., bacalao, milkfish) and bait species (e.g., salemas) were absent in the former study. Finally, light-trap studies targeting larval and early juvenile stages, like Verhaegen (2019) and our own data, demonstrate the occurrence of commercial (e.g., mullets, yellow and Pacific dog snappers) and bait species (e.g., herrings) in mangroves, despite also only covering the CSE bioregion (Table S7).
3.4 Connectivity and Trophic Interactions
Fishers reported higher species richness (of target and non-target species) in mangroves in comparison with other habitats, yet many species were shared with sandy bays, shallow reefs, and vertical walls. By contrast, oceanic habitats, like seamounts, epipelagic waters, and the deep ocean shared less species with mangroves (Figure 2; Table S8). Despite prominent differences in species richness, this is considered an artifact in our study (due to methods being habitat-centered), so will not be further discussed in this paper.
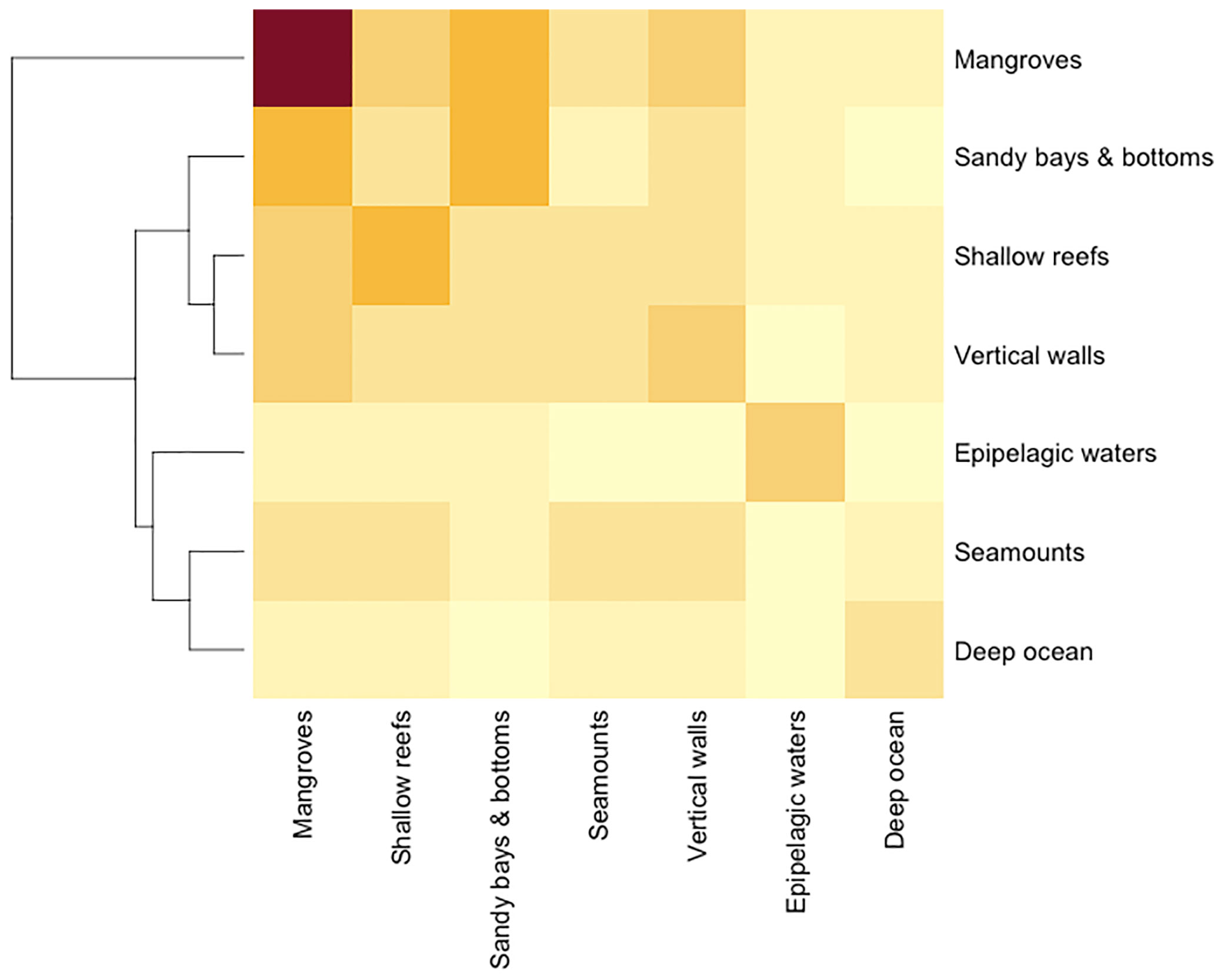
Figure 2 Heatmap depicting similarities among different coastal and marine habitats in the Galapagos in terms of shared species, according to fishers’ LEK. Darker coloration depicts higher species richness (diagonally) or higher similarity between habitats (non-diagonally). Dendrograms represent relative similarity relationships among these habitats.
Based on the analysis of the derived network representation of species interactions, we remarked three clusters in terms on the number of shared habitats (Figure 3). Species in the central cluster shared more habitats with other species, and all were reported in mangroves. Most species in this hub were benthic predators (e.g., snappers, 'bacalao'), omnivores-herbivores (e.g., M. cephalus and M. thoburni) or planktivores (e.g., herrings, salemas), and constitute highly commercial coastal resources in the Galapagos. Contrarily, species in the outer cluster shared the least number of habitats with other species, and were usually pelagic predators (e.g., tunas and marlins), pelagic planktivores (e.g., giant manta ray – Mobula birostris) or deep ocean species (e.g., ‘brujo’; Figure 3). Furthermore, species with highest hub scores (i.e., within the central cluster) included, the longfin yellowtail (1.000), yellow snapper (0.997), Pacific creole-fish (Paranthias colonus; 0.997) and Galapagos octopus (0.997).
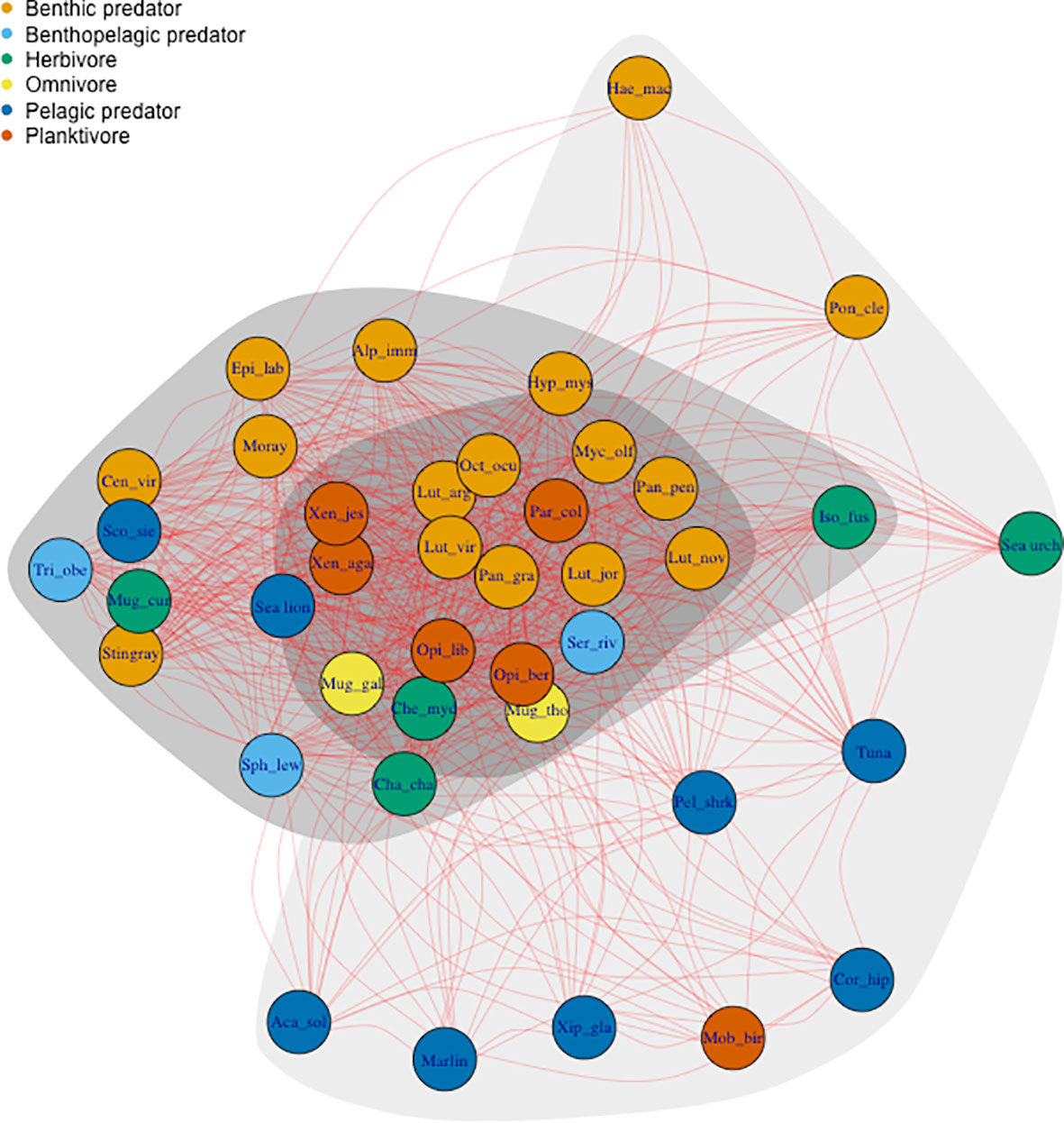
Figure 3 Undirected network representation of species trophic interactions based on shared habitats. Vertices (or nodes) represent species reported by fishers. Edges (or links) represent shared habitats between species. Clusters group species based on ‘hub scores’ estimates during Social Network Analysis and represent cutoff intervals of 1-0.67 (dark grey), 0.66-0.34 (light gray), and 0.33-0 (lightest gray). Species were categorized by feeding type and colored according to legend. Abbreviations for fishery species were coded using three first letters of genera and species in scientific name (see Table 1; Table S8). Other abbreviations: Mob_bir, Mobula birostris; Xip_gla, Xiphius gladius; Aca_sol, Acanthocybium solandri; Cor_hip, Coryphaena hippurus; Pel_shark, other pelagic sharks; Sph_lew, Sphyrna lewini; Iso_fus, Isostichopus fuscus; Moray, moray eels; Sea urch, sea urchins; Tri_obe, Triaenodon obesus.
3.5 Spatial Patterns of Fishery Productivity
Throughout the interviews, fishers identified 7 islands (or parts of islands) across the archipelago containing ‘important’ mangrove locations (Figure 4). Three quarters of the respondents recognized Isabela or sites within the island. Other islands comprised, Santa Cruz, San Cristobal, Fernandina, Santiago, Baltra and Floreana (Figure 4). Almost one quarter of the respondents considered all Galapagos mangrove areas to be equally important.
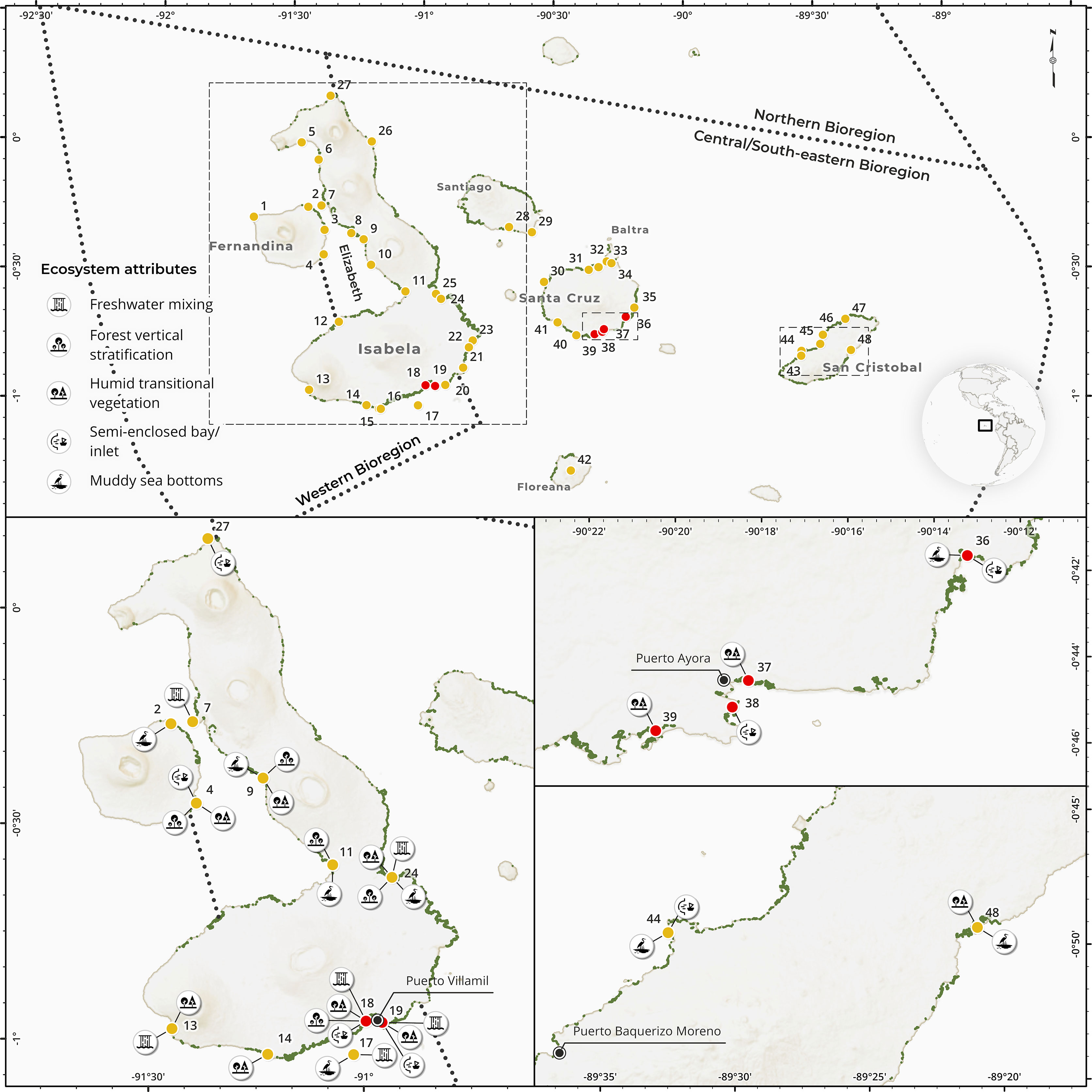
Figure 4 Upper panel. Map of the Galapagos Islands showing ‘important’ mangrove sites (yellow circles) identified by artisanal fishers during interviews. Green polygons along the coast represent mangrove vegetation, as mapped in Moity et al. (2019). Marine bioregions defined by Moity (2019) are overlaid in the map. Site names were number-coded and can be found in Table S9. Lower panels. Subsets maps of Isabela, Santa Cruz and San Cristobal contrasting ‘important’ and ‘impacted’ sites (red circles) regarding ecosystem attributes associated to fishery productivity. Depicted port villages represent locations in which interviews were conducted.
Within these islands, 54 mangrove sites were identified during interviews or workshops, from which 48 could be georeferenced on a map (Figure 4; Table S9). Most salient sites (9-37% of respondents) comprised Cartago Bay, Urbina Bay, Elizabeth Bay, and Punta Albemarle in Isabela; La Tortuga, Puerto Grande, Manglecito, and Rosa Blanca Bay in San Cristobal; and Punta Mangle in Fernandina. Even though most citations corresponded to Isabela or specific sites within the island, we noticed that, in general, fishers tended to recognize locations that were situated closer to their island of residence. For instance, 90% of the respondents from San Cristobal mentioned at least one site in their island.
Other mangrove users interviewed during this study recognized 79% of ‘important’ locations identified by fishers (Table S9), including most-salient sites mentioned above. However, they also remarked unique sites (e.g., Sacacalzon, Islote Venecia in Santa Cruz), some even located in other islands (e.g., Darwin Bay in Genovesa; Pinzon). Higher relative importance was given to Elizabeth Bay and Bolivar Channel in Isabela, Caleta Tortuga Negra and Punta Rocafuerte in Santa Cruz, and Puerto Grande and Manglecito in San Cristobal (Figure S4; Table S9).
We contrasted ecosystem attributes within and between ‘important’ and ‘impacted’ sites during group activities in workshops. Sites concentrating a higher number of attributes in workshops were situated in the Western and Elizabeth bioregions (Figure 4), which were acknowledged by fishers for having the highest mangrove cover. One respondent mentioned that fish in western areas live closer to mangroves throughout their life cycle. OMU linked productivity patterns in the former to the prevalence of nutrient-rich upwellings caused by the cold Cromwell undercurrent. During workshops, sites like Punta Mangle in Fernandina and Urbina Bay, Elizabeth Bay, El Estero, and Cartago Bay in Isabela, were characterized for having the tallest mangrove trees across the archipelago and were equated to mangrove ecosystems in mainland Ecuador (e.g., Gulf of Guayaquil, Esmeraldas). In addition, freshwater mixing was identified in Bolivar Channel (comprising Caleta Tagus and Playa Negra), Caleta Iguana, Roca Union, El Estero, Concha Perla and Cartago Bay in Isabela. On the other side, sites like Garrapatero in Santa Cruz, and Puerto Grande and Rosa Blanca Bay in San Cristobal, were characterized by having mangroves fringing semi-enclosed bays, and the occurrence of muddy sea bottoms; though, forest vertical stratification and freshwater mixing were not reported (Figure 4). In addition, OMU pinpointed sites, like Caleta Tortuga Negra, Punta Rocafuerte and Sacacalzon in Santa Cruz, for their importance as shark nurseries. As such, we were not able to distinguish specific attributes discriminating ‘important’ and ‘impacted’ sites from workshop results, as most prominent spatial patterns resulted from bioregional differences.
4 Discussion
4.1 Mangrove Ecosystem Services and Local Ecological Knowledge
Our study is the first to address a fishers’ LEK approach to study mangrove-fishery linkages in the Galapagos. Although only a reduced percentage of the fisher population participated in the study (from which none were women), it already showcased the potential that fishers’ LEK holds for answering applied research questions about mangrove ecosystems. The application of mixed-methods in our research design (i.e., interviews, different activities in workshops), and the identification of key informants among fishing communities and OMU, allowed us to get access to decades of accumulated ecological knowledge, perceptions and values about mangrove ecosystems, as well as to triangulate and complement information among different knowledge sources. However, we are conscious that these types of ethnoecological studies possess limitations that are absent from the traditional scientific approach. From one side, the sociopolitical context in the Galapagos might have influenced the veracity of some of our results, given recurrent tensions among fishers, scientists and other actors in the archipelago (Quiroga, 2009; Barragán-Paladines, 2015). At the same time, since our methods included several closed-ended questions and semi-qualitative techniques, it might be possible that some of our results do not represent the real vision of fishers themselves, but their understanding of these questions and instructions. The following sections will address separately some of the limitations in each of the research questions investigated in this study.
Before exploring the applicability of fishers’ LEK in answering our research questions, it was necessary in our study to analyze how fishing communities valued mangrove ecosystem in the Galapagos, and how LEK has influenced the establishment of these values. This allowed to identify prominent connections and flows among attributes, functions and benefits of the ecosystem, and to envision ways of integrating LEK with other knowledge sources to complement mangrove research and conservation.
Despite artisanal fishing in the Galapagos mainly occurring in offshore and shallow rocky-reef areas, mangrove ecosystems constitute important grounds for activities, like bait fishing. However, unlike locations, such as Cienaga Grande de Santa Marta in Colombia (Carrasquilla-Henao et al., 2019) and southern Kenya (Rönnbäck et al., 2007), in which direct use values are numerous and prime, in the Galapagos, indirect use (i.e., regulating and supporting services) and non-use values (i.e., cultural services) seemed to play a more important role for local fisheries. Throughout our study, we found that fishers mainly prized mangroves for the facilitation of nursery grounds for a myriad number of marine species, including highly-commercial ones, as well as other habitat uses and ecosystem functions. In addition, non-use values (e.g., shelter, shade, landscape) were also deemed important for the well-being, and social cohesion of fishing communities, which are related to the way they learn from the ecosystem.
Since mangroves are valued for the production of information and knowledge among fishing communities, we deduced that LEK has been determinant in shaping mangrove-fishery linkages in the Galapagos. Although the existence of fishers’ LEK in the archipelago can be rebutted on account of the relatively short history of the local society, the fact that fishers possess fishing traditions inherited or learnt from mainland Ecuador, as well as higher access to remote islands than OMU, constitute two key factors that support the uniqueness and suitability of this knowledge source to answer questions about the marine environment. In this regard, the emergence of LEK has resulted, among other things, in the discernment of the species depending stronger on mangrove ecosystems, and the benefits conferred by the ecosystem in contrast to other coastal habitats in Galapagos. Furthermore, throughout ubiquitous access to multiple areas, fishers have also learned to discriminate key biophysical attributes in the ecosystem and their relationship with ecological functions and environmental conditions, which are valuable to investigate spatial relationships in regard to fishery productivity across the archipelago. Finally, the fact that many fishers have participated in ecotourism, research and conservation activities around mangroves in the past, which sometimes constitute alternative income sources, has likewise played a role in the materialization of this knowledge among fishing communities, and in the strengthening of bonds towards the ecosystem.
4.2 Species-Habitat Associations in Mangroves and Nursery Role
Multiple fishery species in the Galapagos, including many benthic and benthopelagic fishes and invertebrates, were associated to mangrove ecosystems by fishers; some of high commercial importance (e.g., bacalao, snappers) being reported predominantly as juveniles, as other studies confirm (e.g., Fierro-Arcos et al., 2021; Plumlee et al., 2022). In addition, several non-target species, including sharks and aquatic birds, were also associated to mangroves; these establishing critical trophic links in the ecosystem and having a cultural value for local communities. The fact that fishery species listed by fishers highly matched those reported by OMU, as well as those found in past scientific studies (e.g., Fierro-Arcos et al., 2021), allowed us to triangulate our results of species-habitat associations and to discriminate main species associating to the habitat. Triangulation was particularly necessary in this case, due to the high variation in vernacular names among some species (e.g., snappers, grunts), and other issues of reporting during interviews (i.e., species grouping); these constraining the resolution of species taxonomic identities. However, visual aids utilized during the group activity in workshops contributed to partially solve these taxonomic uncertainties, and to pinpoint potential misidentifications in species showing similar morphology; for instance, Jordan’s snapper and grey grunt both being called ‘roncador’, and yellow snapper and Peruvian mojarra (Diapterus peruvianus) both being called ‘chaparra’. As such, it is most likely that fishers’ species list for this habitat did not contain any inaccuracies; thus, can constitute a valid baseline for future studies.
As fishers’ LEK is considered valuable for the understanding of mangrove-fishery linkages in the Galapagos, we underline the opportunities that its integration with SMK could bring in the clarification of habitat-dependence relationships between and within species. We believe that most remarkable insights from fishers derived from their observations and anecdotes about species habitat use, mainly those not yet reported in the scientific literature. Although all fishery species seem to benefit from mangroves during their juvenile stage, it is clear that some are more dependent than others, especially those who also rely on the habitat during their adult stage (e.g., for feeding, mating, spawning). For instance, species like the white snook, salemas, grunts and Pacific thread herring, which have been observed spawning in mangroves, constitute novel reports of species behaviors in the habitat, and might exhibit stronger habitat dependence relationships. Additionally, linkages between habitat use and environmental conditions or biophysical attributes in the ecosystem provided clarification of the mechanisms and factors shaping species-habitat-function relationships. One example of this, is the white snook seeking muddy bottoms to spawn and bury their eggs during the wet season (Table 1).
Furthermore, fishers’ LEK can also complement studies of rare species in Galapagos mangroves, like in other locations (e.g., Barbato et al., 2021) - with species like the misty grouper (Hyporthodus mystacinus) and Galapagos sheephead wrasse (Semicossyphus darwini) constituting novel reports in the habitat (Table S7). On the other hand, species that were given lower relative importance by fishers in contrast to OMU (e.g., Pacific creole-fish grey grunt, milkfish), or that were listed exclusively in mangroves by the latter but to different habitats by the former (e.g., ‘brujo’, brown sea cucumber and yellow-fin tuna), might show weaker dependence to the habitat. These perception discrepancies among mangrove users might result from the fact that fishers are more likely to associate species based on high frequency of sighting, relative abundance or a particular habitat use, while OMU listed species independently of their strength of habitat dependence. Finally, the fact that many species reported as juveniles in mangroves have already been found as larval stages in other studies (e.g., Verhaegen, 2019; own data; Table S7), allowed us to raise the question about the importance of mangroves as larval retention areas in the Galapagos, and to point out the need of more studies targeting these early-life stages, including their taxonomic identification and description.
4.3 Connectivity and Trophic Interactions
Although species-habitat association was probably biased by the fact that a limited number of habitats was presented rather than the real vision of the fishers themselves (i.e., how they see the seascapes of the Galapagos), our results revealed that, in general, coastal seascapes in the Galapagos possess similar macrofaunal assemblages. This suggests low levels of endemism in mangrove ecosystems, though it reveals their role in sustaining species connectivity within coastal seascapes and between coastal and offshore areas (Figure 2). This was expected, as many studies have shown that both vegetated (e.g., seagrass beds, salt marshes) and unvegetated habitats (e.g., coral and shallow rocky reefs), can provide analogous nursery functions to biota as mangroves (Eggleston et al., 2004; Dorenbosch et al., 2005). Despite other studies suggesting high levels of connectivity in fish species between mangroves and shallow reefs in the Galapagos (e.g., Fierro-Arcos et al., 2021), this is the first study to analyze general patterns of connectivity from a fishers’ LEK perspective. Similar qualitative approaches have been employed in Southeastern Puerto Rico to analyze connectivity, while producing relevant results for habitat prioritization and ecosystem-based fisheries management (see García-Quijano, 2007).
During Social Network Analysis and representation, we found that highly-commercial benthic predators, planktivores and omnivores-herbivores, as well as other culturally-important non-target species (e.g., green sea turtle) obtained higher hub score values in the analysis (i.e., central cluster). This implies that they depend more on connectivity than other predators from epipelagic and deep waters (outer cluster; Figure 3), and on complex trophic interactions (e.g., predation, competition, mutualism) across the habitats they use throughout their life cycle. If such species undergo major population declines or extirpations due to overfishing or other threats, they might produce larger cascading effects in marine trophic webs, as it has been demonstrated in past studies (e.g., Ruiz et al., 2016).
One of the advantages of applying Social Network Analysis with qualitative data, is that it does not require probabilistic sampling or hypothesis-driven approaches, which can be more time-consuming, and still reveal main patterns in multidimensional data (Crossley, 2010). At the same time, this highlights the needs of other data types and knowledge sources in future research (e.g., species densities, biomass, growth and survival rates) to improve our understanding of marine connectivity in the Galapagos. For instance, social network analysis could be applied in the identification of cultural keystone species in mangroves and other habitats (Kilkenny and Nalbarte, 2020), which can then inform species prioritization or be used as focal species in conservation or ecosystem-based fisheries management in the GMR (see King and Beazley, 2005). Although some of our target and non-target species (e.g., sharks, aquatic birds, sea lions) have been identified as keystone species in shallow reefs of the Galapagos (Bustamante et al., 2008; Wolff et al., 2012), this study is the first to report potential cultural keystone species across a matrix of connected habitats. Although smaller animals (e.g., crustaceans, polychaetes) and primary producers might also have a keystone role in mangroves and Galapagos seascapes, large mobile macrofauna species reported during this study might be more suitable as focal species, as they tend to be conspicuous and charismatic and have larger distributions (King and Beazley, 2005).
4.4 Spatial Patterns of Fishery Productivity
Mangrove structure and function are known to vary across bioregional and local levels as response of climatic, geomorphological and ecological forces operating on different spatial scales (e.g., Twilley et al., 1999). Throughout our study, fishers were able to identify ‘important’ locations across the archipelago and to score and recognize biophysical attributes in the ecosystem associated to fishery productivity (Figure 4) - this way allowing us to resolve spatial patterns in the ecosystem. Studies, like Moity et al. (2019), identified differences in mangrove forest development by island and by bioregion, supporting the results of fishers’ LEK that identified mangroves of Isabela as the most outstanding in terms of development. In addition, Fierro-Arcos et al. (2021), already revealed differences in mangrove fish assemblages between the Western and CSE bioregions in Galapagos, with the latter showing higher species richness despite lower mangrove cover. Yet, seasonal sea surface temperature, nutrients and primary productivity might play a role in the discrimination of species assemblages. Our results confirm the existence of a bioregional gradient, mainly as result of prevalent climatic and oceanographic conditions, but also reveal patterns at the local scale, which might derive from differences in geomorphology (open vs. semi-enclosed bay), or freshwater discharges in the ecosystem. As most fishers are familiar with the more productive mangrove systems in mainland Ecuador, they were able to pinpoint sites in which these favorable attributes occur. Moreover, our results highlighted the importance of mangrove areas near or easily accessed from urban areas (‘impacted’ sites) for coastal productivity (e.g., El Estero in Isabela and Garrapatero in Santa Cruz), despite some threats being acknowledged in such locations (e.g., tourism overcapacity, overfishing, contamination). This result is in concordance with the patterns found in our larvae-sampling data, which highlighted higher species richness values in such sites in comparison with others, like La Estacion or Playa Alemanes (Figure 4).
Although differences in perceptions among mangrove users drove discrepancies in the selection and relative importance of islands and sites, our results reveal the relevance of the complementarity of knowledge sources in elucidating spatial patterns in mangroves, which have already been explored in other studies (see Llerena-Martillo et al., 2018; Moity et al., 2019; Fierro-Arcos et al., 2021). This complementarity derives from the fact that fishers can visit remote areas that are hardly visited by OMU (for example, areas outside from mangrove touristic sites), and, most importantly, much frequently and in longer periods of time. This allows fishers to observe ecological processes in different seasons and times of the day, and to remember prominent environmental conditions in the ecosystem and to associate them with the occurrence of certain fishery species. On the other side, as OMU mainly comprised naturalist guides, some locations were selected based on their touristic value and their importance as shark nursery grounds (e.g., Caleta Tortuga Negra) or for sea turtle mating. Dissimilarities in the selection of areas among fishers residing in different islands were also noticeable, since nearby locations tend to be more visited than distant ones. This implies that in Galapagos, as in other tropical oceanic islands, perceptions can be conditioned by local environmental conditions, but also sociopolitical context (Rollero and De Piccoli, 2010; Quintas-Soriano et al., 2018). Nonetheless, we failed to find age differences in the selection of sites, as well as in their characterization; possibly, as we did not include many fishers younger than 30, and older than 60 years old (mean age of 48) – this constraining a proper generational comparison.
Future studies should integrate SMK and LEK to explore more in depth the effects of regional-scale environmental and evolutionary changes in species diversity and composition in mangrove ecosystems (e.g., species interactions, environmental filtering, dispersal, speciation, extinction; Levin, 1992; Chase and Myers, 2011), remarkably in the context of a changing climate (Jennerjahn et al., 2017). From another perspective, this knowledge could be potentially integrated to SMK and applied to inform policy, marine zoning and spatial planning in the islands; for instance, in regard to the definition and selection of Essential Fish Habitats (i.e., those waters and substrate necessary to fish for spawning, breeding, feeding or growth to maturity; Rosenberg et al., 2000) for future zoning schemes in the marine reserve. Other studies exhibiting similar approaches are Bergmann et al. (2004) and Leite and Gasalla (2013). As successful applications always rely on the accuracy of the data, it is important that future research using qualitative approaches engage in designing suitable methods to access knowledge from fishing communities in proficient ways, and in triangulating information. Given recurrent tensions between fishers and scientists in the Galapagos, it might be possible that during the interviews, fishers did not reveal all locations that they consider important. In response, by applying approaches like participatory mapping in the Isabela workshop, we were able to discover additional ‘important’ sites by creating a space to fishers to establish a deeper connection with the researchers, as well as to obtain relevant characteristics about the habitat. Although, the semi-quantitative scoring approach in Santa Cruz workshop was also efficient to pinpoint major spatial patterns, it is likely that researchers’ explanation and understanding of the ecosystem variables and scoring criteria might have affected fishers’ responses. The application of mixed-methods in social research can therefore be valuable for the purpose of triangulation and complementarity and should be considered in this type of studies (Newing, 2011; Murray et al., 2016).
5 Conclusion
Even though Galapagos mangroves do not constitute main fishing grounds, fishing communities in the archipelago have developed deep connections and gained knowledge towards the ecosystem, including species habitat use and environmental linkages, connectivity between mangroves and other habitats, and spatial relationships in relation to fishery productivity. Throughout our research we have designed and implemented a two-stage qualitative approach (i.e., interviews and workshops) to explore fishers’ LEK about mangrove-fishery linkages, discussing in the way, the challenges and opportunities for environmental decision-making. The time dedicated in identifying key informants within the local population was crucial in achieving quality and accuracy in our results. In this regard, our results might only reflect the view of those members of the community that are already ‘environmentally sensitive’, or that have been trained in marine ecology due to the abundance of educational events organized locally; these fishers usually collaborating in this type of studies. In addition, the triangulation of certain aspects of fishers’ LEK against other knowledge owners allowed us to position it as a useful complement to more traditional SMK in the Galapagos, contributing this way, in breaking the social paradigm of ‘fishers as predators of the oceans’ and in highlighting the prospects of engaging fishing communities in mangrove and marine ecosystem research.
Throughout this study, fishers’ LEK allowed us to corroborate and reveal novel fishery species that use mangrove habitats (e.g., Galapagos sheephead wrasse), to underline the major role of mangroves in connectivity against other coastal and marine habitats, and to ratify the prominence of the Western bioregion in fishery productivity as a result of specific biophysical attributes. In such regard, we obtained new insights about mangrove nursery role and other mechanisms associated to habitat dependence, complex species interactions occurring in mangroves in response to trophic interactions, and spatial and biotic-abiotic relationship across mangroves of the archipelago - aspects that have been understudied in the Galapagos. The advantage of relying on a qualitative LEK rather than quantitative SMK approach to investigate these research questions, can derive from the fact that fishing communities are more likely to comprehend complex relationships among species and the environment and to track long-term changes in the ecosystem (Cavole et al., 2020). However, if qualitative data from fishing communities and other marine users shall be used in the future to answer hypothesis-driven questions about the marine ecosystem, the use of probabilistic qualitative methods should be considered (e.g., surveys), as well as other ethnographic methodologies in which the relationship between the researcher and the fishers is much more profound (as opposed to the ephemeral relationship in interviews and workshops). Since qualitative studies including human beings is so complex, and can be influenced by many biases, a multi-methodological approach should be preferred. Another challenge in Galapagos lies in how to position LEK in comparison to traditional SMK, an issue to which this research will contribute. Finally, we suggest that LEK should be implemented in future management instruments in support to the co-management principle of the marine reserve (see Usseglio et al., 2014), principally the five-year fishing calendar (i.e., GMR fisheries management plan). For instance, finfish species suspected to have a high dependence to the habitat (e.g., white snook, yellow snapper) and occurring in several mangrove locations across the archipelago, can justify the creation of species-specific regulations in the fishing calendar, including spatial and other types of management measures (e.g., temporal closures). Ultimately, a LEK approach would even bring other benefits such as raising the sense of belonging and pride among fishing communities, and even improving their relationship with other stakeholders in the archipelago.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Ethics Statement
Ethical review and approval were not required for the study on human participants in accordance with the local legislation and institutional requirements. Written informed consent for participation was not required for this study in accordance with the national legislation and the institutional requirements.
Author Contributions
JP-C, NR-V, and MP-C conceptualized this study and carried our data collection. JP-C, NR, and GV-C conducted data analysis and graphing. JP-C wrote the first draft. GN-F, MB-P, NM, GR-J, JR-G, MP-C, JB-M, and PG provided supervision of the study. All authors contributed significantly to revising the manuscript. All authors contributed to the article and approved the submitted version.
Funding
We thank all funding sources that allowed us to organize and conduct this study in the Galapagos Islands, including the Special Research Fund of Ghent University (Belgium; 01W09317), Global Minds Operational Grant from VLIR-UOS (via Ghent University) and Small Grant of The Rufford Foundation (27919-1). We also thank Ghent University for granting open access publication fees.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We would like to first thank to all the participants of fishing communities of Santa Cruz, San Cristobal and Isabela, including representatives and delegates from fishing cooperatives and associations, for sharing their knowledge and experiences around mangrove ecosystems, as well as providing support in our activities. Also, to local naturalist guides, park rangers and scientists for their participation in interviews. We thank the Galapagos National Park Directorate for granting us consent for the conduction of this study, under research permits No. PC-41-19, PC-41-20 and PC-41-21, and for providing advice on methods and attending workshops. Likewise, we thank local governments (Santa Cruz and Isabela), and research institutions (Charles Darwin Foundation, Universidad de Guayaquil) for supporting our study. This includes the facilitation of visual material for the photo-elicitation activity during workshops, logistics, and diffusion of activities. We thank each individual photographer who granted us authorization for the use of their material during workshops and this manuscript. Finally, we thank María José Rendón, Samuel Cevallos, Javier Farraye and Juan Carlos Cedeño, who helped during interviews, community workshops, and data analysis and visualization. This publication is contribution number 2443 of the Charles Darwin Foundation for the Galapagos Islands.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.911109/full#supplementary-material
Footnotes
- ^ To improve visualization, we excluded species reported only in mangroves (Table S8)
- ^ https://www.darwinfoundation.org/en/datazone. Accessed on 24/11/2021
References
Aburto-Oropeza O., Ezcurra E., Danemann G., Valdez V., Murray J., Sala E. (2008). Mangroves in the Gulf of California Increase Fishery Yields. P. Natl. Acad. Sci. U.S.A. 105 (30), 10456–10459. doi: 10.1073/pnas.0804601105
Adobe Inc. (2019) Adobe Illustrator. Available at: https://adobe.com/products/illustrator.
Alongi D. M. (2002). Present State and Future of the World's Mangrove Forests. Environ. Conserv. 29 (3), 331–349. doi: 10.1017/S0376892902000231
Baran E., Hambrey J. (1999). Mangrove Conservation and Coastal Management in Southeast Asia: What Impact on Fishery Resources? Mar. Pollut. Bull. 37 (8-12), 431–440. doi: 10.1016/S0025-326X(99)00076-4
Barbato M., Barría C., Bellodi A., Bonanomi S., Borme D., Ćetković I., et al. (2021). The Use of Fishers’ Local Ecological Knowledge to Reconstruct Fish Behavioral Traits and Fishers’ Perception of Conservation Relevance of Elasmobranchs in the Mediterranean Sea. Mediterr. Mar. Sci. 22 (3), 603–622. doi: 10.12681/mms.25306
Barbier E. B., Hacker S. D., Kennedy C., Koch E. W., Stier A. C., Silliman B. R. (2011). The Value of Estuarine and Coastal Ecosystem Services. Ecol. Monogr. 81, 169–193. doi: 10.1890/10-1510.1
Barragán-Paladines M. J. (2015). Exploring Governance in Galapagos Marine Reserve (St. John’s: Memorial University of Newfoundland).
Benzeev R., Hutchinson N., Friess D. A. (2017). Quantifying Fisheries Ecosystem Services of Mangroves and Tropical Artificial Urban Shorelines. Hydrobiologia 803 (1), 225–237. doi: 10.1007/s10750-017-3299-8
Bergmann M., Hinz H., Blyth R. E., Kaiser M. J., Rogers S. I., Armstrong M. (2004). Using Knowledge From Fishers and Fisheries Scientists to Identify Possible Groundfish ‘Essential Fish Habitats’. Fish. Res. 66 (2-3), 373–379. doi: 10.1016/j.fishres.2003.07.007
Berkes F., Colding J., Folke C. (2000). Rediscovery of Traditional Ecological Knowledge as Adaptive Management. Ecol. Appl. 10 (5), 1251–1262. doi: 10.1890/1051-0761(2000)010[1251:ROTEKA]2.0.CO;2
Blaber S. J. (2007). Mangroves and Fishes: Issues of Diversity, Dependence, and Dogma. B. Mar. Sci. 80 (3), 457–472.
Borgatti S. P., Mehra A., Brass D. J., Labianca G. (2009). Network Analysis in the Social Sciences. Science 323(5916), 892–895. doi: 10.1126/science.1165821
Brook R. K., McLachlan S. M. (2008). Trends and Prospects for Local Knowledge in Ecological and Conservation Research and Monitoring. Biodiver. Conserv. 17 (14), 3501–3512. doi: 10.1007/s10531-008-9445-x
Burbano D. V., Mena C. F., Guarderas P., Vinueza L., Reck G. (2014). “Shifting Baselines in the Galapagos White Fin Fishery, Using Fisher’s Anecdotes to Reassess Fisheries Management: The Case of the Galapagos Grouper,” in The Galapagos Marine Reserve: A Dynamic Social-Ecological System. Eds. Denkinger J., Vinueza L. (New York, NY: Springer International Publishing), 159–185.
Bustamante R. H., Okey T. A., Banks S. (2008). “Biodiversity and Food-Web Structure of a Galapagos Shallow Rocky-Reef Ecosystem,” in Food Webs and the Dynamics of Marine Reefs. Eds. McClanahan T. R., Branch G. M. (New York, NY: Oxford University Press, Inc), 135–161.
Butler J. R., Tawake A., Skewes T., Tawake L., McGrath V. (2012). Integrating Traditional Ecological Knowledge and Fisheries Management in the Torres Strait, Australia: The Catalytic Role of Turtles and Dugong as Cultural Keystone Species. Ecol. Soc 17 (4), 34. doi: 10.5751/ES-05165-170434
Carrasquilla-Henao M., Ban N., Rueda M., Juanes F. (2019). The Mangrove-Fishery Relationship: A Local Ecological Knowledge Perspective. Mar. Policy 108, 103656. doi: 10.1016/j.marpol.2019.103656
Carrasquilla-Henao M., Juanes F. (2017). Mangroves Enhance Local Fisheries Catches: A Global Meta-Analysis. Fish. Fish. 18 (1), 79–93. doi: 10.1111/faf.12168
Castellanos-Galindo G. A., Krumme U. (2013). Tidal, Diel and Seasonal Effects on Intertidal Mangrove Fish in a High-Rainfall Area of the Tropical Eastern Pacific. Mar. Ecol. Prog. Ser. 494, 249–265. doi: 10.3354/meps10512
Castrejón M. (2011). Co-Manejo Pesquero En La Reserva Marina De Galápagos Tendencias, Retos Y Perspectivas De Cambio. (México, D.F: Fundación Charles Darwin, Kanankil, Plaza y Valdés).
Castrejón M., Charles A. (2013). Improving Fisheries Co-Management Through Ecosystem-Based Spatial Management: The Galapagos Marine Reserve. Mar. Policy 38, 235–245. doi: 10.1016/j.marpol.2012.05.040
Cavole L. M., Andrade-Vera S., Marin Jarrin J. R., Dias D. F., Aburto-Oropeza O., Barrágan-Paladines M. J. (2020). Using Local Ecological Knowledge of Fishers to Infer the Impact of Climate Variability in Galápagos’ Small-Scale Fisheries. Mar. Policy 121, 104195. doi: 10.1016/j.marpol.2020.104195
Charles Darwin Foundation (2018) Datazone. Available at: https://www.darwinfoundation.org/en/datazone (Accessed November 24, 2021).
Chase J. M., Myers J. A. (2011). Disentangling the Importance of Ecological Niches From Stochastic Processes Across Scales. Philos. T. R. Soc B. 366(1576), 2351–2363. doi: 10.1098/rstb.2011.0063
Consejo de Gobierno del Régimen Especial de Galápagos (CGREG) (2021). Plan De Desarrollo Sustentable Y Ordenamiento Territorial Del Régimen Especial De Galápagos, Plan Galápagos 2030 (Puerto Baquerizo Moreno, Galápagos: CGREG).
Crossley N. (2010). The Social World of the Network: Combining Qualitative and Quantitative Elements in Social Network Analysis (Sociologica 1). Sociologica 1, 1—34. doi: 10.2383/32049
Csárdi G., Nepusz T. (2006).The Igraph Software Package for Complex Network Research. InterJournal. Complex Syst. 1695 (5), 1—19.
Dorenbosch M., Grol M. G. G., Christianen M. J. A., Nagelkerken I., van der Velde G. (2005). Indo-Pacific Seagrass Beds and Mangroves Contribute to Fish Density and Diversity on Adjacent Coral Reefs. Mar. Ecol. Prog. Ser. 302, 63–76. doi: 10.3354/meps302063
Drew J. A. (2005). Use of Traditional Ecological Knowledge in Marine Conservation. Conserv. Biol. 19 (4), 1286–1293. doi: 10.1111/j.1523-1739.2005.00158.x
Eggleston D. B., Dahlgren C. P., Johnson E. G. (2004). Fish Density, Diversity, and Size-Structure Within Multiple Back Reef Habitats of Key West National Wildlife Refuge. B. Mar. Sci. 75 (2), 175–204.
Engelhard S. L., Huijbers C. M., Stewart-Koster B., Olds A. D., Schlacher T. A., Connolly R. M. (2017). Prioritizing Seascape Connectivity in Conservation Using Network Analysis. J. Appl. Ecol. 54 (4), 1130–1141. doi: 10.1111/1365-2664.12824
Esri Inc. (2020). ArcGIS Pro (Version 2.5) (Redlands, CA: Esri Inc). Available at: https://www.esri.com/en-us/arcgis/products/arcgis-pro/overview.
Fierro-Arcos D., Marín Jarrín J. R., Aburto-Oropeza O., Harvey E. S., Rastoin-Laplane E., Salinas-de-León P. (2021). Mangrove Fish Assemblages Reflect the Environmental Diversity of the Galapagos Islands. Mar. Ecol. Prog. Ser. 664, 183–205. doi: 10.3354/meps13628
Figus E. C. (2018). Using Local Knowledge to Inform Commercial Fisheries Science and Management in Poland and Alaska (Fairbanks, AK: University of Alaska Fairbanks).
García-Quijano C. G. (2007). Fishers' Knowledge of Marine Species Assemblages: Bridging Between Scientific and Local Ecological Knowledge in Southeastern Puerto Rico. Am. Anthropol. 109 (3), 529–536. doi: 10.1525/aa.2007.109.3.529
González J. A., Montes C., Rodríguez J., Tapia W. (2008). Rethinking the Galapagos Islands as a Complex Social-Ecological System: Implications for Conservation and Management. Ecol. Soc 13 (2), 13. doi: 10.5751/ES-02557-130213
Goodman L. A. (1961). Snowball Sampling. Ann. Math. Stat. 32 (1), 148–170. doi: 10.1214/aoms/1177705148
Heylings P., Bensted-Smith R., Altamirano M. (2002). “Zonificación E Historia De La Reserva Marina De Galápagos,” in Reserva Marina De Galápagos: Línea Base De La Biodiversidad. Eds. Danulat E., Edgar G. J. (Santa Cruz, CA: Fundación Charles Darwin y Servicio Parque Nacional de Galápagos), 10–22.
Igulu M. M., Nagelkerken I., Dorenbosch M., Grol M. G., Harborne A. R., Kimirei I. A., et al. (2014). Mangrove Habitat Use by Juvenile Reef Fish: Meta-Analysis Reveals That Tidal Regime Matters More Than Biogeographic Region. PloS One 9, e114715. doi: 10.1371/journal.pone.0114715
Jennerjahn T. C., Gilman E., Krauss K. W., Lacerda L. D., Nordhaus I., Wolanski E. (2017). “Mangrove Ecosystems Under Climate Change”,” in Mangrove Ecosystems: A Global Biogeographic Perspective”. Eds. Rivera-Monroy V. H., Lee S. H., Kristensen E., Twilley R. R. (Cham: Springer International Publishing), 211–244.
Jones P. J. (2013). A Governance Analysis of the Galápagos Marine Reserve. Mar. Policy 41, 65–71. doi: 10.1016/j.marpol.2012.12.019
Jupiter S., Mangubhai S., Kingsford R. T. (2014). Conservation of Biodiversity in the Pacific Islands of Oceania: Challenges and Opportunities. Pacific Conserv. Biol. 20 (2), 206–220. doi: 10.1071/PC140206
Kilkenny M., Nalbarte L. (2020). “Keystone Sector Identification: A Graph Theory-Social Network Analysis Approach,” in The Web Book of Regional Science. Ed. Loveridge S. (Morgantown, WV: West Virginia University Regional Research Institute).
King M. C., Beazley K. F. (2005). Selecting Focal Species for Marine Protected Area Network Planning in the Scotia–Fundy Region of Atlantic Canada. Aquat. Conserv. 15 (4), 367–385. doi: 10.1002/aqc.682
Lal P. (2003). Economic Valuation of Mangroves and Decision-Making in the Pacific. Ocean Coast. Manage. 46 (9-10), 823–844. doi: 10.1016/S0964-5691(03)00062-0
Lee S. Y., Primavera J. H., Dahdouh-Guebas F., McKee K., Bosire J. O., Cannicci S., et al. (2014). Ecological Role and Services of Tropical Mangrove Ecosystems: A Reassessment. Global Ecol. Biogeogr. 23 (7), 726–743. doi: 10.1111/geb.12155
Leite M. C., Gasalla M. A. (2013). A Method for Assessing Fishers’ Ecological Knowledge as a Practical Tool for Ecosystem-Based Fisheries Management: Seeking Consensus in Southeastern Brazil. Fish. Res. 145, 43–53. doi: 10.1016/j.fishres.2013.02.013
Levin S. A. (1992). The Problem of Pattern and Scale in Ecology: The Robert H. MacArthur Award Lecture. Ecology 73 (6), 1943–1967. doi: 10.2307/1941447
Llerena-Martillo Y., Peñaherrera-Palma C., Espinoza E. R. (2018). Fish Assemblages in Three Fringed Mangrove Bays of Santa Cruz Island, Galapagos Marine Reserve. Rev. Biol. Trop. 66 (2), 674–687. doi: 10.15517/rbt.v66i2.33400
Madsen E. K., Elliot N. B., Mjingo E. E., Masenga E. H., Jackson C. R., May R. F., et al. (2020). Evaluating the Use of Local Ecological Knowledge (LEK) in Determining Habitat Preference and Occurrence of Multiple Large Carnivores. Ecol. Indic. 118, 106737. doi: 10.1016/j.ecolind.2020.106737
Manson F. J., Loneragan N. R., Skilleter G. A., Phinn S. R. (2005). An Evaluation of the Evidence for Linkages Between Mangroves and Fisheries: A Synthesis of the Literature and Identification of Research Directions. Oceanogr. Mar. Biol. 43, 493–524.
Millennium Ecosystem Assessment (2005). Ecosystems and Human Well-Being: Wetlands and Water Synthesis (Washington, DC: World Resources Institute).
Moity N. (2018). Evaluation of No-Take Zones in the Galapagos Marine Reserve, Zoning Plan 2000. Front. Mar. Sci. 5. doi: 10.3389/fmars.2018.00244
Moity N. (2019). Marine Bioregions for Coastal Subtidal Shallow Rocky Reef of the Galapagos Archipelago, GIS Layer for Edgar Et Al. 2004 (Pangaea). doi: 10.1594/PANGAEA.907105
Moity N., Delgado B. (2018). “Manglares,” in Atlas De Galápagos, Ecuador: Especies Nativas E Invasoras (Quito: Fundación Charles Darwin (FCD) y WWF-Ecuador), 61–63.
Moity N., Delgado B., Salinas-de-León P. (2019). Mangroves in the Galapagos Islands: Distribution and Dynamics. PloS One 14 (1), e0209313. doi: 10.1371/journal.pone.0209313
Molina L., Danulat E., Oviedo M., González J. A. (2004). Guı́a De Especies De Interés Pesquero En La Reserva Marina De Galápagos. Puerto Ayora, Galapagos: Fundación Charles Darwin, Agencia Española de Cooperación Internacional and Dirección Parque Nacional Galápagos.
Murray G., Bavington D., Neis B. (2005). “Local Ecological Knowledge, Science, Participation and Fisheries Governance in Newfoundland and Labrador: A Complex, Contested and Changing Relationship,” in Participation in Fisheries Governance. Ed. Gray T. S. (Dordrecht: Springer), 269–290.
Murray G., D’Anna L., MacDonald P. (2016). Measuring What We Value: The Utility of Mixed Methods Approaches for Incorporating Values Into Marine Social-Ecological System Management. Mar. Policy 73, 61–68. doi: 10.1016/j.marpol.2016.07.008
Nagelkerken I., Blaber S. J. M., Bouillon S., Green P., Haywood M., Kirton L. G., et al. (2008). The Habitat Function of Mangroves for Terrestrial and Marine Fauna: A Review. Aquat. Bot. 89 (2), 155–185. doi: 10.1016/j.aquabot.2007.12.007
Newing H. (Ed.) (2011). “Developing the Methodology,” in Conducting Research in Conservation: Social Science Methods and Practice (London and New York, NY: Routledge), 43–65.
Patton M. Q. (2002). Qualitative Evaluation and Research Methods. 2nd ed (Thousand Oaks, CA: Sage Publications, Inc).
Pilgrim S., Cullen L., Smith D., Pretty J. (2007). Hidden Harvest or Hidden Revenue-A Local Resource Use in a Remote Region of Southeast Sulawesi, Indonesia. Indian J. Tradit. Know. 6, 150–159.
Pilgrim S. E., Cullen L. C., Smith D. J., Pretty J. (2008). Ecological Knowledge is Lost in Wealthier Communities and Countries. Environ. Sci. Technol. 42 (4), 1004–1009. doi: 10.1021/es070837v
Plumlee J. D., Kenworthy M. D., Gittman R. K., Keller D. A., Garnett E. E., Vaca-Pita L., et al. (2022). Remarkable Euryhalinity of a Marine Fish Lutjanus Novemfasciatus in Mangrove Nurseries. Ecology 103 (2), e03582. doi: 10.1002/ecy.3582
Pontón-Cevallos J. F., Bruneel S., Marín Jarrín J. R., Ramírez-González J., Bermúdez-Monsalve J. R., Goethals P. L. (2020). Vulnerability and Decision-Making in Multispecies Fisheries: A Risk Assessment of Bacalao (Mycteroperca Olfax) and Related Species in the Galapagos’ Handline Fishery. Sustainability 12 (17), 6931. doi: 10.3390/su12176931
Puri R. K. (2011). “Documenting Local Environmental Knowledge and Change,” in Conducting Research in Conservation. Ed. Newing H. (London and New York, NY: Routledge), 168–191.
Quintas-Soriano C., Brandt J. S., Running K., Baxter C. V., Gibson D. M., Narducci J., et al. (2018). Social-Ecological Systems Influence Ecosystem Service Perception. Ecol. Soc 23 (3), 3. doi: 10.5751/ES-10226-230303
Quiroga D. (2009). Crafting Nature: The Galapagos and the Making and Unmaking of a "Natural Laboratory". J. Polit. Ecol. 16 (1), 123–140. doi: 10.2458/v16i1.21695
Quiroga D., Orbes A. (1964). Apuntes E Informaciones Sobre Las Pesquerías En El Archipiélago De Colón (Islas Galápagos). Inst. Nacional Pesca Boletín Informativo 1 (5), 4–18.
Raymond C. M., Fazey I., Reed M. S., Stringer L. C., Robinson G. M., Evely A. C. (2010). Integrating Local and Scientific Knowledge for Environmental Management. J. Environ. Manage. 91 (8), 1766–1777. doi: 10.1016/j.jenvman.2010.03.023
R Core Team (2021). R: A Language and Environment for Statistical Computing (Vienna: R Foundation for Statistical Computing). Available at: https://www.R-project.org/.
Riascos-Flores L., Bruneel S., van der Heyden C., Deknock A., Van Echelpoel W., Forio M. A., et al. (2021). Polluted Paradise: Occurrence of Pesticide Residues Within the Urban Coastal Zones of Santa Cruz and Isabela (Galapagos, Ecuador). Sci. Total Environ. 763, 142956. doi: 10.1016/j.scitotenv.2020.142956
Richard V. M., Lahman M. K. (2015). Photo-Elicitation: Reflexivity on Method, Analysis, and Graphic Portraits. Int. J. Res. Method. Eda. 38 (1), 3–22. doi: 10.1080/1743727X.2013.843073
Rodríguez-Jacome G. (2020). Cultural Heritage, Tourism and Local Development in The Case of Santa Cruz-Galápagos, Ecuador (Barcelona: Universitat Autònoma de Barcelona).
Rodríguez-Jacome G., Moity N., Ramírez-González J., Andrade-Vera S., Barragán-Paladines M. J. (2019). “Climate Change Impacts in the Artisanal Fishery of Galapagos: Fishers’ Knowledge,” in Technical Report No.6 (Puerto Arora, Galapagos: Charles Darwin Foundation).
Rollero C., De Piccoli N. (2010). Place Attachment, Identification and Environment Perception: An Empirical Study. J. Environ. Psychol. 30 (2), 198–205. doi: 10.1016/j.jenvp.2009.12.003
Rönnbäck P., Crona B., Ingwall L. (2007). The Return of Ecosystem Goods and Services in Replanted Mangrove Forests: Perspectives From Local Communities in Kenya. Environ. Conserv. 34 (4), 313–324. doi: 10.1017/S0376892907004225
Rosenberg A., Bigford T. E., Leathery S., Hill R. L., Bickers K. (2000). Ecosystem Approaches to Fishery Management Through Essential Fish Habitat. B. Mar. Sci. 66 (3), 535–542.
Roux M. J., Tallman R. F., Martin Z. A. (2019). Small-Scale Fisheries in Canada's Arctic: Combining Science and Fishers’ Knowledge Towards Sustainable Management. Mar. Policy 101, 177–186. doi: 10.1016/j.marpol.2018.01.016
Ruiz D. J., Banks S., Wolff M. (2016). Elucidating Fishing Effects in a Large-Predator Dominated System: The Case of Darwin and Wolf Islands (Galápagos). J. Sea Res. 107, 1–11. doi: 10.1016/j.seares.2015.11.001
Schiller L., Alava J. J., Grove J., Reck G., Pauly D. (2013). “A Reconstruction of Fisheries Catches for the Galapagos Islands 1950-2010,” in Working Paper Series #2013-11 (Fisheries Centre: The University of British Columbia).
Sheaves M. (2009). Consequences of Ecological Connectivity: The Coastal Ecosystem Mosaic. Mar. Ecol. Prog. Ser. 391, 107–115. doi: 10.3354/meps08121
Sheaves M., Abrantes K., Barnett A., Benham C., Dale P., Mattone C., et al. (2020). The Consequences of Paradigm Change and Poorly Validated Science: The Example of the Value of Mangroves to Fisheries. Fish. Fish. 21 (5), 1067–1075. doi: 10.1111/faf.12479
Snell H. M., Stone P. A., Snell H. L. (1996). A Summary of Geographical Characteristics of the Galapagos Islands. J. Biogeogr. 23 (5), 619–624. doi: 10.1111/j.1365-2699.1996.tb00022.x
Stave J., Oba G., Nordal I., Stenseth N. C. (2007). Traditional Ecological Knowledge of a Riverine Forest in Turkana, Kenya: Implications for Research and Management. Biodiver. Conserv. 16 (5), 1471–1489. doi: 10.1007/s10531-006-9016-y
Suárez-Moncada J. M. (2012). Comunidades Biológicas En Los Manglares De Isabela, Santa Cruz Y San Cristóbal (Guayaquil: Universidad de Guayaquil).
Tanner M. K., Moity N., Costa M. T., Marin Jarrin J. R., Aburto-Oropeza O., Salinas-de-León P. (2019). Mangroves in the Galapagos: Ecosystem Services and Their Valuation. Ecol. Econ. 160, 12–24. doi: 10.1016/j.ecolecon.2019.01.024
Tidbury H., Taylor N., van der Molen J., Garcia L., Posen P., Gill A., et al. (2020). Social Network Analysis as a Tool for Marine Spatial Planning: Impacts of Decommissioning on Connectivity in the North Sea. J. Appl. Ecol. 57 (3), 566–577. doi: 10.1111/1365-2664.13551
Tomaselli M., Kutz S., Gerlach C., Checkley S. (2018). Local Knowledge to Enhance Wildlife Population Health Surveillance: Conserving Muskoxen and Caribou in the Canadian Arctic. Biol. Conserv. 217, 337–348. doi: 10.1016/j.biocon.2017.11.010
Twilley R. R., Rivera-Monroy V. H., Chen R., Botero L. (1999). Adapting an Ecological Mangrove Model to Simulate Trajectories in Restoration Ecology. Mar. Pollut. Bull. 37, 404–419. doi: 10.1016/S0025-326X(99)00137-X
Usseglio P., Schuhbauer A., Friedlander A. (2014). “Collaborative Approach to Fisheries Management as a Way to Increase the Effectiveness of Future Regulations in the Galapagos Archipelago,” in The Galapagos Marine Reserve: A Dynamic Social-Ecological System. Eds. Denkinger J., Vinueza L. (New York, NY: Springer International Publishing), 159–185.
Vega F. (2021)“En Galápagos Existen 420 'Pescadores' Con Licencia Que No Pescan Ni Un Resfrío”. In: Bitácora Ambiental. Available at: https://www.bitacoraec.com/post/en-galápagos-existen-420-pescadores-con-licencia-que-no-pescan-ni-un-resfr%C3%ADo?fbclid=IwAR0uLJ891Fc5zfstvHrlcVKhxweJJ8vRPZOZy6dlXOpJsi0o08uSpPOcevA (Accessed January 13, 2022).
Verhaegen C. (2019). DNA Barcoding of Larvae of Commercially Important Fish Species in the Galapagos Islands (Brussels: Universiteit Antwerpen, Universiteit Gent and Vrije Universiteit Brussel).
Weller S. C., Romney K. A. (1988). Systematic Data Collection (Newbury Park, CA: Sage Publications, Inc).
Whitfield A. K. (2017). The Role of Seagrass Meadows, Mangrove Forests, Salt Marshes and Reed Beds as Nursery Areas and Food Sources for Fishes in Estuaries. Rev. Fish. Biol. Fisher. 27 (1), 75–110. doi: 10.1007/s11160-016-9454-x
Wilson D. C., Raakjær J., Degnbol P. (2006). Local Ecological Knowledge and Practical Fisheries Management in the Tropics: A Policy Brief. Mar. Policy 30 (6), 794–801. doi: 10.1016/j.marpol.2006.02.004
Wium-Andersen S., Hamann O. (1986). Manglares De Las Islas Galápagos. Inst. Geográfico Mil. Rev. Geográfica 23, 101–122.
Keywords: mangroves, fishers, fisheries, local ecological knowledge, Galapagos Islands, ecosystem services, research, ethnoecology
Citation: Pontón-Cevallos J, Ramírez-Valarezo N, Pozo-Cajas M, Rodríguez-Jácome G, Navarrete-Forero G, Moity N, Villa-Cox G, Ramírez-González J, Barragán-Paladines MJ, Bermúdez-Monsalve JR and Goethals PLM (2022) Fishers’ Local Ecological Knowledge to Support Mangrove Research in the Galapagos. Front. Mar. Sci. 9:911109. doi: 10.3389/fmars.2022.911109
Received: 02 April 2022; Accepted: 05 May 2022;
Published: 02 June 2022.
Edited by:
Manuel J. Zetina-Rejón, Instituto Politécnico Nacional (IPN), MexicoReviewed by:
Ilana Rosental Zalmon, State University of the North Fluminense Darcy Ribeiro, BrazilSalvatore Siciliano, Fundação Oswaldo Cruz (Fiocruz), Brazil
Copyright © 2022 Pontón-Cevallos, Ramírez-Valarezo, Pozo-Cajas, Rodríguez-Jácome, Navarrete-Forero, Moity, Villa-Cox, Ramírez-González, Barragán-Paladines, Bermúdez-Monsalve and Goethals. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: José Pontón-Cevallos, am9zZWZlcm5hbmRvLnBvbnRvbmNldmFsbG9zQHVnZW50LmJl