 Sabit Mohammad Aslam1,2
Sabit Mohammad Aslam1,2 Imre Vass
Imre Vass Milán Szabó
Milán Szabó- 1Institute of Plant Biology, Biological Research Centre, Eötvös Loránd Research Network, Szeged, Hungary
- 2Doctoral School of Biology, University of Szeged, Szeged, Hungary
- 3Climate Change Cluster, University of Technology Sydney, Ultimo, NSW, Australia
Symbiodiniaceae live in endosymbiosis with corals. In the last few decades, mass bleaching events have occurred in the coral reefs, causing damage in the ecosystem and the associated species. Global temperature increase is affecting the algae, disturbing the whole symbiosis and leads to coral bleaching. However, the heat tolerance is strongly determined by the species (formerly genetic clades) harbored by the coral host. We assessed three different strains of Symbiodiniaceae family, i.e., Fugacium kawagutii (CS156), Symbiodinium tridacnidorum (2465), and Symbiodinium microadriaticum (2467), which display different heat tolerance under heat stress conditions. Flash-induced chlorophyll fluorescence relaxation is a useful tool to monitor various components of the photosynthetic electron transport chain and the redox reactions of plastoquinone pool. We observed the appearance of a wave phenomenon in the fluorescence relaxation by heating the strains in combination with microaerobic conditions. The characteristics of this fluorescence wave were found to be strain-specific and possibly related to the transient oxidation and re-reduction of the plastoquinone pool. The appearance of the wave phenomenon appears to be related to cyclic electron flow as well because it is accompanied with enhanced post-illumination chlorophyll fluorescence rise. These results will potentially reveal further details of the role of cyclic electron transport in Symbiodiniaceae and its relevance in heat stress tolerance.
Introduction
The family Symbiodiniaceae comprise of dinoflagellate unicellular microalgae, which are commonly found in endosymbiotic relationship with cnidarians such as corals, sea anemones, and jellyfish. Because Symbiodiniaceae have the ability to perform photosynthesis and synthesize glucose or glycerol, they are responsible for fulfilling the energy requirements of the holobiont, while cnidarian host supplies CO2 and essential inorganic nutrients required for the synthesis of organic compounds (Davy et al., 2012). This relationship is found to be flourishing well under ideal, non-stressed conditions and has a particular significance on coral reefs, which provide shelter to fishes, algae, and hundreds of other associated marine organisms.
The process of reduction of atmospheric CO2 to organic compounds in plants, algae, and certain bacteria using the energy of light is known as photosynthesis. Photosynthesis is divided into two major steps. The first one is known as light reaction or photophosphorylation and occurs in the thylakoid membrane. It includes the absorption of light energy by the photosynthetic antenna system and the transfer of the energy to the reaction centers, where charge separation occurs. Charge separation initiates the series of electron transfer from photolysis of water to ferredoxin-NADP-oxidoreductase (FNR) complex, which reduces NADP+ to NADPH; this process from water to NADPH is denoted as linear electron flow (LEF). The vectorial electron flow is coupled with proton transfer from the stroma to the lumen, which generates proton motive force, which is utilized in adenosine triphosphate (ATP) synthesis. The second reaction, also known as dark reaction or Calvin-Benson-Bassham (CBB) cycle, utilizes the NADPH and ATP generated in the light reaction for the reduction of CO2 into carbohydrates, with the help of the Ribulose-1,5-bisphosphate carboxylase/oxygenase enzyme (RuBisCO) in the stroma.
The ratio of ATP/NADPH generated by LEF alone is not always sufficient to fully support the formation of glyceraldehyde-3-phosphate, the export product of the CBB cycle (Allen, 2002; Kramer and Evans, 2011). Therefore, in photosynthetic organisms, different alternative pathways such as cyclic electron flow (CEF), Mehler reaction, plastoquinone terminal oxidase (PTOX), and flavodiiron (FLV) mediated pathways exist to help overcoming the ATP shortage in normal as well as stress conditions. CEF involves Photosystem I (PSI), where electrons are transferred from the acceptor side of PSI to ferredoxin (Fd) or NAD(P)H, then back to the PQ pool and to the donor side of PSI through the Cyt b6f complex (Cardol et al., 2011). In this way, it generates proton gradient across the thylakoid membrane (Shikanai and Yamamoto, 2017), which further facilitates ATP production, and, thus, maintains the NADPH/ATP ratio under conditions when the acceptor side of PSI is limited. There are two proposed pathways for CEF. One involves Proton gradient regulation5 (PGR5) and PGR5 like photosynthetic phenotype1 (PGRL1) complexes, which mediate the electron transfer from Ferredoxin (Fd) to the PQ pool. Another pathway involves NAD(P)H dehydrogenase (NDH). Two types of NDH complexes exist: type I NDH (NDH-1), which mediates the electron transfer from Fd to the PQ pool in cyanobacteria; whereas, in the case of NDH-2 (or Nda2), which can be found in microalgae, the electron donor to the PQ pool is NADH [reviewed in (Peltier et al., 2016)].
Annual global temperature is increasing year after year. The rate of increase has more than doubled since 1880 from 0.08°C per decade to 0.18°C in 1981 (ncdc-Global Climate Report – Annual, 2020), which is leading to various damages to the ecosystems across the globe. One such example is coral bleaching. Increase in sea water temperature affects adversely the photosynthetic activity of the Symbiodiniaceae, leading to disruption of the symbiotic association (Weis, 2008). This results in the expulsion of symbionts, bleaching of corals which compromises their survival and the persistence of the reef ecosystem (Hughes et al., 2018). However, the extent of destruction is dependent upon the species of Symbiodiniaceae as endosymbiont, as it is classified into nine different genera (formerly clades A–I, (Lajeunesse et al., 2018)), which show different levels of tolerance toward heat stress in different geographical conditions (Abrego et al., 2008; Buxton et al., 2012; Díaz-Almeyda et al., 2017). Various factors such as damage of photosystem proteins (D1) and repair mechanism (Smith et al., 2005), damaged photosynthetic apparatus (Venn et al., 2008), and accumulation of ROS (Downs et al., 2002; Suggett et al., 2008; Lesser, 2011; Hawkins et al., 2015; Rehman et al., 2016; Roberty et al., 2016; Lesser, 2019), thylakoid membrane integrity (Tchernov et al., 2004; Hill et al., 2009; Slavov et al., 2016), antenna proteins (Takahashi et al., 2008) and carbon fixation reactions (Jones et al., 1998; Leggat et al., 2004; Lilley et al., 2010; Hill et al., 2014; Oakley et al., 2014) are found responsible for this large scale bleaching phenotype of corals under heat stress.
Chlorophyll a fluorescence detection is a sensitive tool to monitor the activity of Photosystem II. Illumination of dark-adapted samples leads to the increase in fluorescence from a minimum level (usually denoted as F0) to a maximum (usually denoted as Fm), which decreases again depending on the applied conditions and physiological status of the samples. This variable fluorescence yield is mainly determined by the redox state of the primary electron acceptor of PSII, QA, but is also affected by other processes of the entire electron transport chain. Depending on the application and the selection of the excitation and detection conditions, and measurement protocols, several features of the photosynthetic activity can be revealed (reviewed, e.g., in (Kalaji et al., 2014). A powerful technique is the flash-induced chlorophyll fluorescence relaxation kinetics, which is based on the combined application of weak measuring (or probing) flashes to measure the fluorescence yield resulting from the actual redox state of QA, without being influenced by the probing flashes, at a given time point, and a strong but very short (couple of μs long, quasi single turnover) actinic flash, which causes the transient formation of QA− and, consequently, the increase in the fluorescence yield (Vass et al., 1992; Vass et al., 1999; Cser and Vass, 2007). The fluorescence decreases thereafter, which is monitored over a time range of microseconds to seconds and usually displayed in logarithmic scale due to the wide span of time. Typically, three phases can be discerned due to the reoxidation of QA−: (i) fast phase, which is dominated by the electron transfer to QB, the secondary electron acceptor in PSII, in the timescale of 300–500 µs; (ii) middle phase, which reflects the reoxidation by the plastoquinone that binds to the QB site from the PQ pool after the flash, this occurs in 5–15 ms; and (iii) slow phase, 10–20 s, reflecting the reoxidation of QA−via charge recombination with the oxidized S2 (or S3) states of the donor side of PSII, which occurs from the QAQB- state (this state is in charge equilibrium with QA− QB) (Vass et al., 1999; Deák et al., 2014).
A special case of the fluorescence relaxation profile is the appearance of the so-called wave phenomenon, which, in cyanobacteria, manifests itself in a fluorescence dip at approximately 30–50 ms and a subsequent rise in fluorescence at approximately 1 s; after which, the fluorescence decreases again to its initial level (Deák et al., 2014). This wave phenomenon appeared under microaerobic condition in Synechocystis sp., which causes a strong reduction of the PQ pool due to the depletion of O2, which is the terminal electron acceptor of terminal oxidases (Houille-Vernes et al., 2011; Deák et al., 2014; Ermakova et al., 2016). However, microaerobic conditions alone were not sufficient to induce the wave in eukaryotic microalgae (Volgusheva et al., 2013; Volgusheva et al., 2016; Krishna et al., 2019; Patil et al., 2022). This is because one of the criteria of the appearance of the wave phenomenon is the enhanced reoxidation by PSI relative to the electron influx to the PQ pool from PSII (Deák et al., 2014) and, in certain microalgae, the ratio of PSII : PSI is higher than in cyanobacteria; therefore, the electron flow from PSII toward the PQ pool and the reoxidation of the PQ pool by PSI is well balanced. When the relative activity of PSII is decreased and the PQ pool is highly reduced due to the presence of microaerobic condition, the fluorescence dip and subsequent rise appeared in microalgae (Krishna et al., 2019; Patil et al., 2022). It was also shown that the “bump” phase of the wave is related the re-reduction of the PQ pool via NDH-1 in Synechocystis (Deák et al., 2014) and NDH-2 in Chlamydomonas (Krishna et al., 2019). Therefore, the wave phenomenon potentially gives new information about the rate of electron transfer from stromal components to the PQ pool, electron delivery to various components of the electron transfer chain, and alternative electron transfer pathways, operation of terminal oxidases in regulating the redox level of the PQ pool under various environmental conditions, and balanced regulation of intersystem PSII-PSI electron transport.
Chlorophyll fluorescence has been widely used to determine the photosynthetic efficiency and electron transfer rates in Symbiodiniaceae under various conditions (Gorbunov et al., 2001; Hill et al., 2004; Hill and Ralph, 2008a; Warner et al., 2010; Roth, 2014; Suggett et al., 2015; Warner and Suggett, 2016; Nitschke et al., 2018), and, in combination with other biophysical methods, the alternative electron transport pathways such as CEF (Reynolds et al., 2008; Roberty et al., 2014; Aihara et al., 2016; Dang et al., 2019; Vega De Luna et al., 2020). Active chlorophyll fluorescence kinetics therefore potentially enable bio-optical phenotyping of the coral endosymbiont algae based on an extensive set of parameters of Chl fluorescence induction and relaxation (Hoadley and Warner, 2017; Gorbunov and Falkowski, 2021; Suggett et al., 2022). However, the wave phenomenon, a special but potentially informative signature of the flash-induced Chl fluorescence relaxation in the extended time range of QA− reoxidation (and re-reduction), has not been investigated so far in Symbiodiniaceae.
The aim of the present work was to characterize the flash-induced chlorophyll fluorescence relaxation and the wave phenomenon under the conditions which were previously found to induce wave in cyanobacteria and algae (such as microaerobic treatment and/or diminished PSII activity). Furthermore, strain-specific properties of the photosynthetic electron transport pathways were investigated in three different strains of Symbiodiniaceae belonging to the genera Symbiodinium and Fugacium, under elevated temperatures that were found earlier to induce CEF in Symbiodiniaceae (Aihara et al., 2016; Dang et al., 2019) and to clarify whether these changes are reversible after returning to growth temperature. Heat stress inactivates the donor side of PSII (Tóth et al., 2007a; Hill and Ralph, 2008b); therefore, it increases the PSI : PSII activity ratio, which creates an imbalance in the electron flow from PSII to PQ pool and from PQ pool to PSI. This is an important factor in the appearance of the wave phenomenon. Strain CS156 (Fugacium kawagutii, formerly clade F) and 2465 (Symbiodinium tridacnidorum) and 2467 (Symbiodinium microadriaticum) (formerly clade A3 and A1 respectively) were shown earlier to display different photo-physiological characteristics and functional properties (Suggett et al., 2015). Therefore, they represent contrasting groups in terms of photosynthetic electron transfer processes, which was an important consideration for the species selection for studying flash-induced fluorescence relaxation. It was also investigated whether microaerobic treatment alone, or in combination with acute heat stress, is able to induce the wave phenomenon in different species of Symbiodiniaceae.
Materials and Methods
Symbiodiniaceae Cultures
Symbiodiniaceae cultures CCMP2465 (Symbiodinium tridacnidorum, formerly clade A3), CCMP2467 (Symbiodinium microadriaticum, formerly clade A1), and CS156 (Fugacium kawagutii formerly clade F1) were grown at 24°C under 50 µmol photons m−2 s−1 white light for a week in F/2 media until reaching the mid-log growth phase. Cells were centrifuged at 5,000g and re-suspended in fresh F/2 media in such a way that the final Chlorophyll (Chl) concentration was adjusted to 5 µg/ml. Prepared sample was pre-illuminated under 50 µmol photons m−2 s−1 (growth light condition) for an hour before each measurement.
Measurement of Flash-Induced Chlorophyll Fluorescence Relaxation Kinetics
Flash-induced chlorophyll fluorescence yield (FF) was measured using a double-modulation fluorimeter (FL-3000, Photon Systems Instruments, Brno, Czech Republic) (Trtílek et al., 1997). A 2-ml sample was placed in a cuvette with 1-cm path length and was continuously stirred with a small magnetic stirrer bar in the dark. Four measuring flashes (8 μs, separated with 200-μs intervals, wavelength of 620 nm) were applied to determine minimum fluorescence in the dark (F0), after which, a single turnover saturating actinic flash (30 μs, wavelength of 639 nm) was given to induce the formation of QA−, which resulted in the rise of fluorescence intensity. This is the maximal fluorescence recorded after the single turnover actinic flash, which we specifically denote as Fm(ST). The fluorescence decay resulting from re-oxidation of QA− was measured by applying weak measuring flashes, which have negligible actinic effect and, therefore, do not interfere with the redox state of QA, in the time range from 150 μs to 100 s on a logarithmic time scale.
Measurement of Chlorophyll a Fluorescence Induction Curves
Kautsky induction curve or the polyphasic rise of chlorophyll fluorescence OJIP curve was measured using an FL-3000 fluorimeter (Photon Systems Instruments, Brno, Czech Republic). This method consists of monitoring the kinetics of the chlorophyll fluorescence induction curve in the transition between dark to light adapted state (Kalaji et al., 2017). It measures the transition between the state where all PSII reaction centers are open (referred as O step or F0 at 20 µs) to the state where all reaction centers are closed and the fluorescence intensity reaches its maximum, nominally called as peak (referred as P step or Fm ~ 300 ms) (Strasser and Govindjee, 1992). Two intermediate steps are also observed typically: the J step (at approximately 2 ms) and the I step (at approximately 20 ms). An automated protocol was applied, which provided red actinic light (wavelength at peak, 650 nm; spectral line half-width, 22 nm) with the intensity of 3,500 μmol m−2 s−1 for the induction of OJIP fluorescence transients with a length of 3 s.
Measurement of Post-Illumination Chlorophyll Fluorescence Transient
Post-illumination chlorophyll fluorescence transients (PIFT) were measured using Dual-PAM, Heinz-Walz GmbH, Effeltrich, Germany. Two milliliters of sample was dark adapted for 5 min, then chlorophyll fluorescence was recorded for 6 min. For the first 30 s, the sample was illuminated with a weak measuring light (ML) of approximately 0.1 μmol photons m−2 s−1, then an actinic light (AL, 50 μmol photons m−2 s−1) for 3 min was applied, followed by recording the Chl fluorescence rise in the dark. The same ML intensity was used for the entire measurement sequence.
Activity of Photosystems
The activity of Photosystem II and Photosystem I (Y(II) and Y(I), respectively) were measured using a Dual-PAM, Heinz-Walz GmbH, Effeltrich, Germany. Eight milliliters of the sample (Chl content of 40 μg/ml) with different heat treatments was filtered onto Whatman GF/C filter paper and dark adapted for 5 min. Because the cultures were filtered and spread evenly on the filter paper in thin layer, high chlorophyll content was required to obtain a good signal. Y(II) and Y(I) were determined simultaneously according to (Hoogenboom et al., 2012). Y(II) is calculated as (Fm’-Ft)/Fm’, where Fm’ is the maximum fluorescence upon the saturation pulse and Ft is steady-state value of fluorescence immediately prior to the flash (Maxwell and Johnson, 2000). In dark-adapted state, Y(II) (or Fv/Fm) is calculated as (Fm − F0)/Fm, where Fm is the maximal and F0 is the minimal fluorescence. In analogy to Fm, Pm represents the maximal level of P700 signal obtained by a saturation pulse under far-red illumination, when P700 is fully oxidized (P700+). Y(I) = (Pm′ − P)/Pm, where Pm′ is the maximal level of P700 signal obtained by a saturation pulse at a defined AL illumination and P is the P700 oxidation level at a defined AL illumination (Klughammer and Schreiber, 1994; Klughammer and Schreiber, 2008). The length of the saturation pulse to determine Y(II) and Y(I) was 300 ms (with an intensity of ~10,000 μmol photons m−2 s−1). The intensity of ML for F0 determination was <0.5 μmol photons m−2 s−1.
Experimental Procedure
Differences in the response to acute heat stress within different species of Symbiodiniaceae were observed by recording the changes in photosynthetic parameters including FF, OJIP, PIFT, and photosystem activity at growth condition (24°C), heated (34°C for 2 h), and recovery (for 2 h). Another set of heat stress experiments was also performed at 24°C, 36°C, 38°C, and 40°C for 1 h to monitor the temperature-dependent changes in flash-induced chlorophyll fluorescence relaxation. Microaerobic condition was created by dark, incubating the sample with 7 U ml−1 of glucose oxidase, 60 U ml−1 of catalase, and 10 mM glucose, for 15 min. This microaerobic condition was complemented with the acute heat stress (for 10 min) and the changes in flash-induced chlorophyll fluorescence relaxation were recorded. The same procedure was applied for all Symbiodiniaceae strains.
Statistical Analysis
Statistical analysis was performed using Origin Pro (Origin-Lab Corporation, Northampton, MA, USA). One-way analysis of variance (ANOVA) was performed on independent samples to detect statistically significant differences between treatments for which Tukey’s post-hoc multiple comparison tests (α = 0.05) were applied. Normality tests were performed using the Kolmogorov-Smirnov method and the homogeneity of variance test was performed using Levene’s method.
Results
The Response of the Photosynthetic Activity to Acute Heat Stress in Symbiodiniaceae
In order to reveal the heat-induced changes in the flash-induced chlorophyll fluorescence relaxation, different strains of Symbiodiniaceae were subjected to acute heat stress (Figure 1, upper panels).
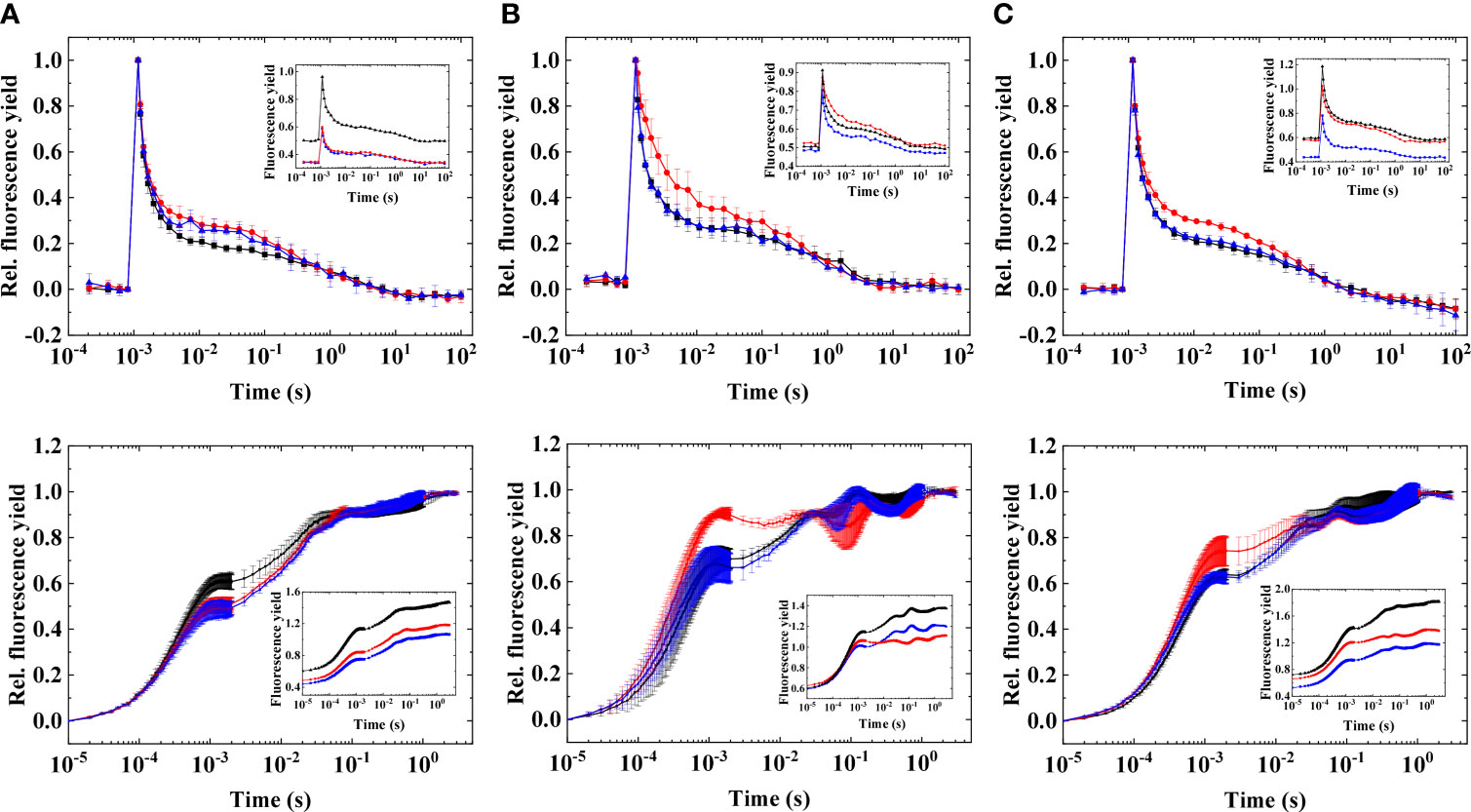
Figure 1 Flash-induced fluorescence relaxation (upper panel) and OJIP induction curves (lower panel) in three different strains of Symbiodiniaceae under elevated temperatures.(A) Fugacium kawagutii (CS156), (B) Symbiodinium tridacnidorum (2465), and (C) Symbiodinium microadriaticum (2467), including control (black), 34°C 2 h (red), and recovery (blue). Main figures: normalized fluorescence traces [mean values of three biological replicates ± standard deviation (S.D.)]. Insets: original data (representative traces). The fluorescence traces were double normalized (F0 to 0 and Fm to 1) to achieve the same initial amplitudes. Insets show original traces without normalization.
Acute heat stress (at 34°C) caused a slight increase in the middle phase (0.003–0.1 s) of the fluorescence relaxation, in all investigated strains, but it was the most expressed in S. tridacnidorum 2465, although these changes were minor at this temperature. Increase in the middle phase corresponds to the elevated reduction of the PQ pool. This was confirmed by fluorescence induction measurements; OJIP traces (Figure 1, lower panels) showed that the largest increase in J phase (at ~1 ms, which indicates the reduction state of the PQ pool (Hill et al., 2004; Tóth et al., 2007b), occurred in S. tridacnidorum 2465, in agreement with the largest increase in the middle phase of the FF transient (Figure 1, upper panels) in S. tridacnidorum 2465. F. kawagutii CS156 displayed the smallest increase in the middle phase of the FF transient, in accordance with the least change in the J phase of the OJIP curve (interestingly, in F. kawagutii CS156, the J phase even decreased at elevated temperatures). The heat-induced changes in the flash-induced fluorescence relaxation traces were nearly completely reversible when the samples were incubated at growth temperature (24°C), in agreement with the decrease of the J phase of the OJIP curves, indicating that the PQ pool becomes more oxidized when the heat-induced changes were relieved.
Elevated heat also caused remarkable changes in the post-illumination Chl fluorescence rise. A transient Chl fluorescence rise occurs after the cessation of light due to the reduction of the PQ pool by stromal reductants; therefore, post-illumination Chl fluorescence rise is indicative of the redox kinetics of the PQ pool (Asada et al., 1993; Reynolds et al., 2008; Gotoh et al., 2010; Deák et al., 2014; Claquin et al., 2021), which showed strain-specific features in Symbiodiniaceae (Figure 2).
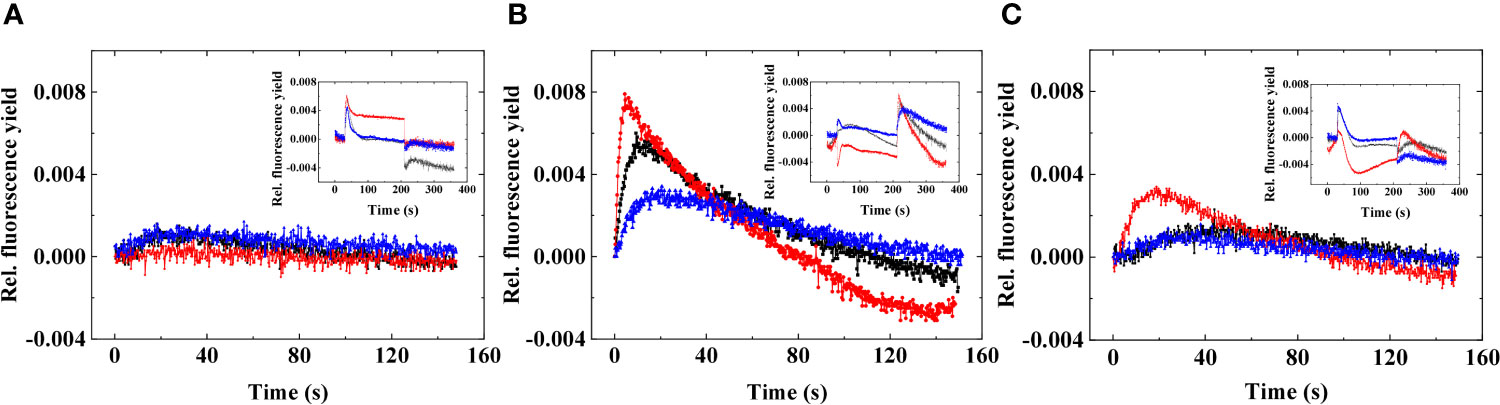
Figure 2 Representative post-illumination Chl fluorescence rise in three different strains of Symbiodiniaceae under elevated temperatures. (A) Fugacium kawagutii (CS156), (B) Symbiodinium tridacnidorum (2465), and (C) Symbiodinium microadriaticum (2467), including control (black), 34°C 2 h (red), and recovery (blue). The fluorescence was adjusted to 0 at the beginning of the post-illumination (dark) phase to display relative changes. Insets show the original traces; here, the minimum fluorescence before the actinic illumination is adjusted to 0 to display relative differences in fluorescence yield.
The smallest response could be observed in F. kawagutii CS156, where acute heat stress did not remarkably alter the post-illumination Chl fluorescence characteristics (Figure 2A). In S. microadriaticum 2467, heat stress accelerated the post-illumination Chl fluorescence rise (Figure 2C), whereas S. tridacnidorum 2465 exhibited the largest response (Figure 2B). In S. tridacnidorum 2465, it was also observed that the post-illumination fluorescence rise was quite large already at 24°C in darkness, indicating a strong dark-induced reduction of the PQ pool (Hill and Ralph, 2008a). At 34°C, the post-illumination fluorescence rise occurred faster than at 24°C and then slowed down when the cells were re-incubated at 24°C.
In order to reveal the changes in the activity of both photosystems under the acute heat stress, PSII and PSI activities were measured (Figure 3).
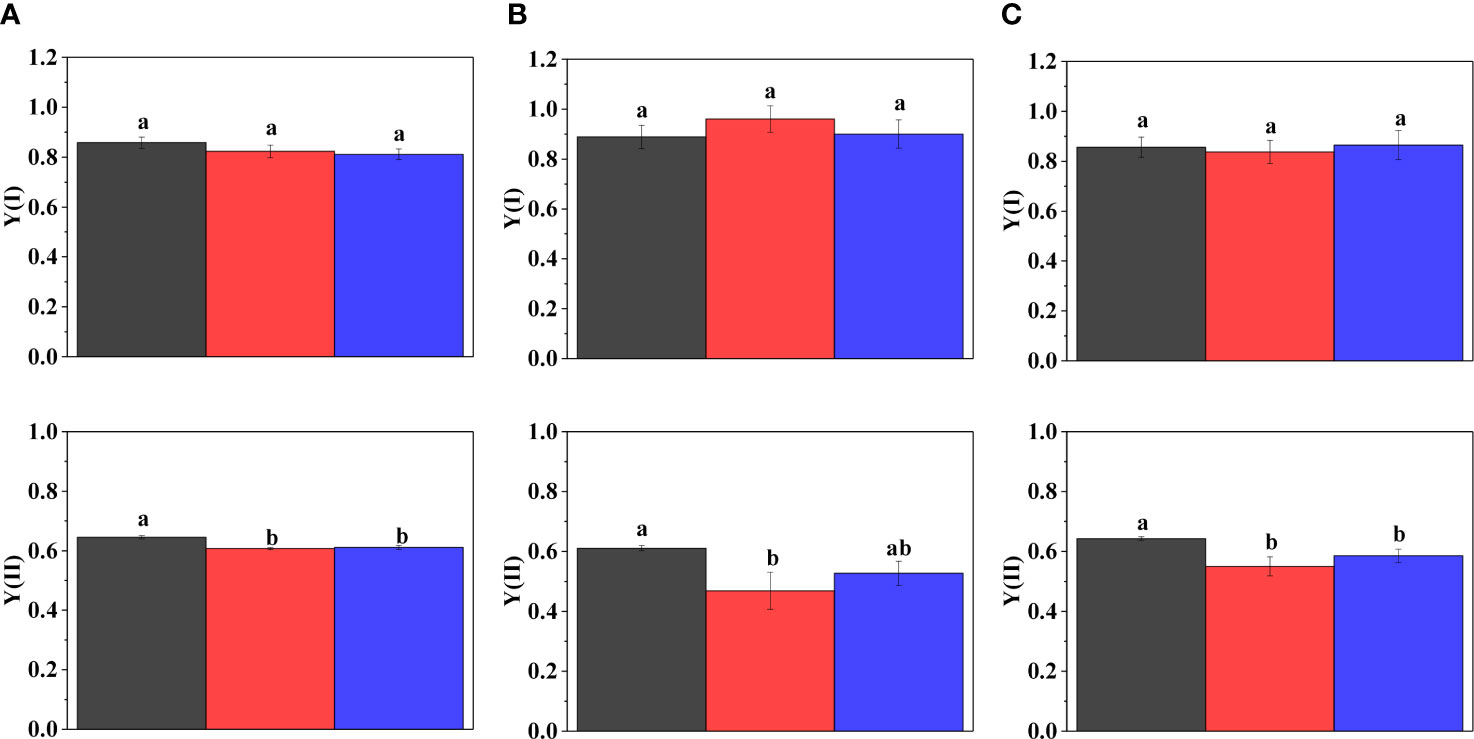
Figure 3 Activity of PSI ((Y(I)), upper panel) and PSII ((Y(II), lower panel) in three different strains of Symbiodiniaceae under elevated temperatures. (A) Fugacium kawagutii (CS156), (B) Symbiodinium tridacnidorum (2465), and (C) Symbiodinium microadriaticum (2467), including control (black), 34°C 2 h (red), and recovery (blue). Different letters above the bars indicate statistically different means (mean values of three biological replicates ± S.D.), and values sharing common letters are not significantly different from one another (p < 0.05).
Acute heat stress caused no change in the activity of Photosystem I (Y(I)) in F. kawagutii CS156 (Figure 3A) and S. microadriaticum 2467 (Figure 3C), but it caused a slight elevation in Y(I) in S. tridacnidorum 2465; however, this was not statistically significant (Figure 3B). The activity of Photosystem II (Y(II)) decreased significantly in all strains, the largest decrease could be observed in S. tridacnidorum 2465. Y(II) recovered only partially under growth temperature in all the three strains.
Temperature Dependence of the Flash-Induced Chlorophyll Fluorescence Relaxation Kinetics in Symbiodiniaceae
To test the inducibility of the wave in fluorescence relaxation in Symbiodiniaceae, cells were exposed to acute heat stress treatment. As shown in Figure 1, acute heat stress at 34°C caused an increase in the middle phase of flash-induced fluorescence relaxation as a result of reduced PQ pool; however, this condition did not induce the fluorescence wave formation. Therefore, it was investigated whether elevated temperatures (above 34°C) caused any further change in the flash-induced fluorescence relaxation (Figure 4).
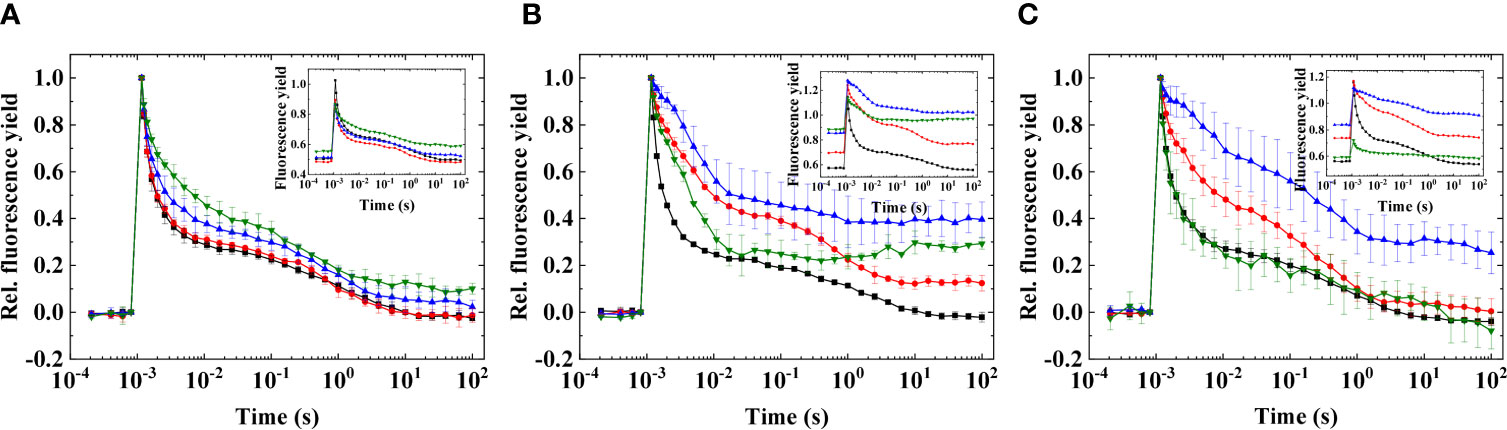
Figure 4 Flash-induced fluorescence relaxation in three different species of Symbiodiniaceae under elevated temperatures. (A) Fugacium kawagutii (CS156), (B) Symbiodinium tridacnidorum (2465), and (C) Symbiodinium microadriaticum (2467), including control (black), 36°C (red), and 38°C (blue), and 40°C (green). The fluorescence traces were double normalized (F0 to 0 and Fm to 1) to achieve the same initial amplitudes (mean values of three biological replicates ± S.D.). Insets show original traces without normalization (representative traces).
Heat treatment with higher temperatures caused marked changes in the flash-induced fluorescence relaxation profiles, and these changes were also strain specific. In F. kawagutii CS156, the flash fluorescence profiles did not change remarkably until 38°C; at 40°C, the middle phase slightly increased indicating the reduced PQ pool (Figure 4A). In S. microadriaticum 2467, the PQ pool became strongly reduced at 36°C and the reduction level increased further at 38°C; however, at 40°C, the fluorescence profile became similar to the profile at the growth temperature (Figure 4C). The reason of this is probably that the signal became very small at 40°C and, therefore, could not be properly measured. S. tridacnidorum 2465 exhibited reduced PQ pool at 36°C because of the elevated middle phase of fluorescence relaxation; however, increasing the temperature further caused a dip and then a slight increase in fluorescence, reminiscent of a wave phenomenon (Figure 4B). These results also indicate the remarkably different heat sensitivity of the different strains, with F. kawagutii CS156 being the least sensitive to elevated temperatures even up to 40°C.
The Induction of the Flash-Induced Wave Phenomenon Is Species Specific in Symbiodiniaceae
It was investigated whether microaerobic conditions alone, which cause a strongly reduced PQ pool due to the depletion of O2 which inhibits the terminal oxidases that keep the PQ pool oxidized (Deák et al., 2014; Ermakova et al., 2016), can induce the wave phenomenon in Symbiodiniaceae or not (Figure 5).
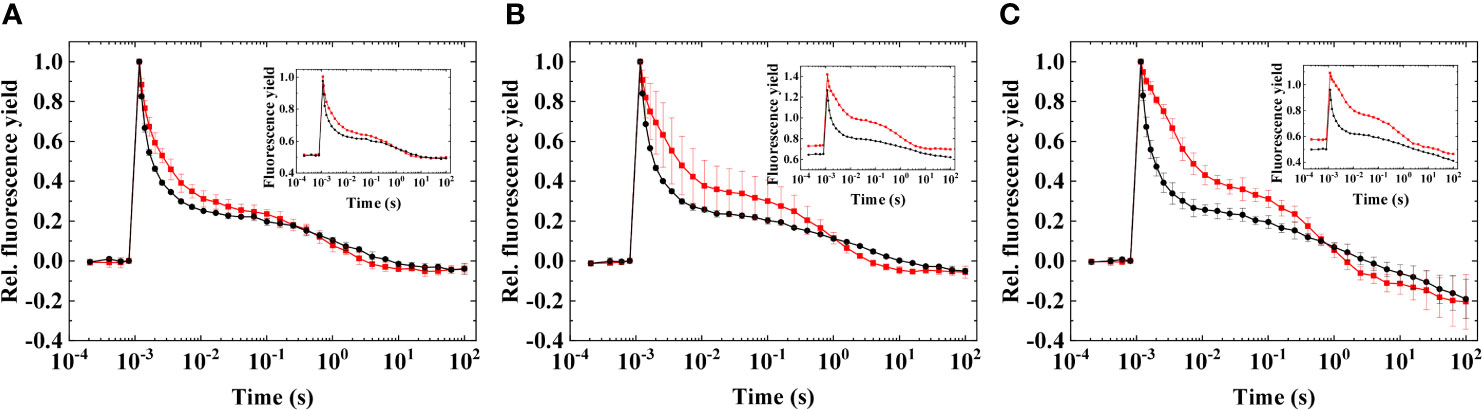
Figure 5 Flash-induced fluorescence relaxation in three different species of Symbiodiniaceae under aerobic and microaerobic conditions, recorded at growth temperature. (A) Fugacium kawagutii (CS156), (B) Symbiodinium tridacnidorum (2465), and (C) Symbiodinium microadriaticum (2467), including control (black) and microaerobic condition (red). Main figures: Normalized fluorescence. Insets: Original data. The fluorescence traces were double normalized (F0 to 0 and Fm to 1) to achieve the same initial amplitudes (mean values of three biological replicates ± S.D.). Insets show original traces without normalization (representative traces).
Under microaerobic conditions, a pronounced reduction of the PQ pool could be observed, as indicated by the elevated middle phase of the fluorescence relaxation (Figure 5) and by the increased F0 (Figure 5 inset). Microaerobic conditions caused similar changes across the different strains; however, the induction of the wave phenomenon did not occur in any of them, in agreement with other microalgae, in which microaerobic conditions were not sufficient either to induce the wave phenomenon (Krishna et al., 2019; Patil et al., 2022).
When microaerobic conditions were applied on heat-treated cells, the transient dip and subsequent rise in fluorescence were more expressed, particularly in S. tridacnidorum 2465 (at 38°C; Figure 6B). This temperature resulted in the appearance of the wave phenomenon in F. kawagutii CS156 as well (Figure 6A); however, the largest amplitude could be observed in S. tridacnidorum 2465 in all cases, in which the fluorescence dip falls below the initial dark-adapted F0 level (Supplementary Figures S1–S3, showing the results at 34°C, 36°C, and 40°C heat treatment). In S. microadriaticum 2467, the wave was not obvious; furthermore, a large decrease was observed in the longer timescales (> 10 s) probably due to cell clumping (Figure 6C). This was partially eliminated when 10% ficoll was applied in the cell suspension; however, cell clumping and sedimentation remain an issue in heat-treated samples and microaerobic condition (Supplementary Figure S4). Nevertheless, it is clear that the transient dip and subsequent increase in fluorescence (a wave phenomenon) are the most expressed in S. tridacnidorum 2465.
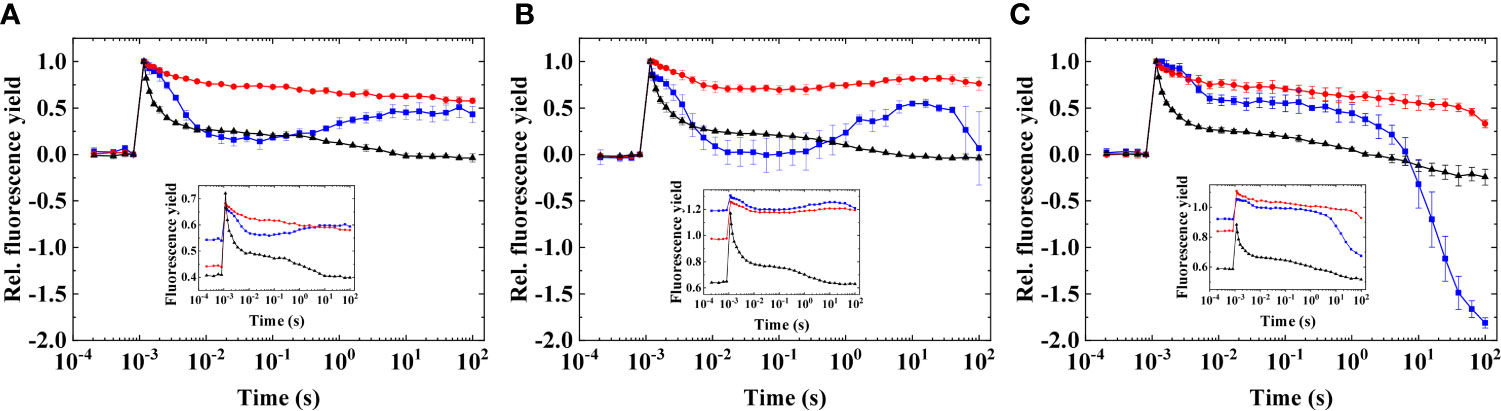
Figure 6 Flash-induced fluorescence relaxation in three different species of Symbiodiniaceae in heat-treated cells in combination with microaerobic conditions. (A) Fugacium kawagutii (CS156), (B) Symbiodinium tridacnidorum (2465), and (C) Symbiodinium microadriaticum (2467), including control (black), heated (at 38°C, red), and heated + microaerobic (blue). Main figures: normalized fluorescence (mean values of three biological replicates ± S.D.). Insets: original data (representative traces). The fluorescence traces were double normalized (F0 to 0 and Fm to 1) to achieve the same initial amplitudes. Insets show original traces without normalization.
Discussion
The Wave Phenomenon in Flash-Induced Chlorophyll Fluorescence Relaxation
Flash-induced chlorophyll fluorescence relaxation kinetics is a highly informative tool to monitor several electron transfer processes in cyanobacteria and microalgae: intersystem PSII-PSI electron flow as well as charge recombination and alternative electron transfer processes such as CEF (Vass et al., 1999; Deák et al., 2014; Volgusheva et al., 2016; Krishna et al., 2019; Havurinne and Tyystjärvi, 2020). Under standard conditions, the fluorescence relaxation displays kinetically well-separated and discernible phases. However, under specific or stress conditions, characteristic waves in the fluorescence relaxation could also be identified, which indicate the reduction-reoxidation sequences of the PQ pool. In cyanobacteria, this wave phenomenon was assigned to the operation of the NDH-1 complex, which mediates the electron transfer from ferredoxin at the acceptor side of PSI to the PQ pool (Deák et al., 2014). In microalgae such as Chlamydomonas reinhardtii, the wave phenomenon is related to the activity of NDH-2 (Krishna et al., 2019; Patil et al., 2022). It is important to note that the nature and the manifestation of the wave phenomenon are quite different in the different taxonomical groups. Whereas, in Synechocystis, microaerobic conditions are sufficient to induce the wave, in microalgae, usually, microaerobic treatment is not sufficient to its manifestation. This is due to the different stoichiometry of photosystems. When the PSI : PSII ratio is high, as in cyanobacteria, strong reduction of the PQ pool is sufficient to induce the wave because the electron withdrawal from the PQ pool by PSI exceeds the electron injection to the PQ pool from PSII. When the PSI : PSII ratio is lower, as in eukaryotic microalgae, the two processes (i.e., electron supply to and withdrawal from the PQ pool) are more balanced, which prevents the induction of the wave. As it was shown, if C. reinhardtii, partial inactivation of PSII, which increases the activity ratio of PSI : PSII, in combination with microaerobic treatment, was required to induce the large dip and subsequent rise of fluorescence after the flash (Patil et al., 2022). Therefore, the ratio of photosystems strongly influences the wave phenomenon, which therefore is an informative indicator of the balance of intersystem electron flow.
Flash-Induced Chlorophyll Fluorescence Relaxation Under Acute Heat Stress in Symbiodiniaceae
As heat stress is a severe problem for coral reefs, it is imperative to establish non-intrusive techniques to monitor heat-induced changes in the physiology and health of coral reefs organisms. Symbiodiniaceae species provide the essential photosynthetic products to the coral host to understand the regulation of photosynthetic electron transport processes of the endosymbiont algae. In our work, treating various Symbiodiniaceae strains with acute heat stress that was shown earlier to damage PSII (Hill and Ralph, 2008b), induce CEF (Aihara et al., 2016; Dang et al., 2019), or affect PSI activity and electron donation between PSII and PSI in intact corals (Szabó et al., 2017) caused some alterations in the fluorescence relaxation profile. This manifested itself in elevated middle phase of the fluorescence relaxation, which is related to enhanced reduction of the PQ pool. These changes were confirmed with fast fluorescence induction and post-illumination Chl fluorescence rise measurements, which showed that the strain S. tridacnidorum 2465 exhibiting the largest change in flash-induced fluorescence relaxation profiles displayed the largest (reversible) increase in the PQ pool reduction (largest post-illumination Chl fluorescence rise and largest increase in J phase of OJIP curves) upon acute heat stress. Conversely, the strain F. kawagutii CS156, which did not exhibit remarkable changes in flash-induced fluorescence relaxation, remained relatively unaltered in terms of the redox state of PQ pool. Therefore, the differences in heat sensitivity of the different species of Symbiodiniaceae as revealed by means of flash-induced fluorescence relaxation and accompanying Chl fluorescence measurements corroborated earlier studies, which identified different functional groups of Symbiodiniaceae (Hennige et al., 2009; Fisher et al., 2012; Suggett et al., 2015; Dang et al., 2019). However, it has to be noted that the acute heat treatment in the 34°C range was insufficient for the manifestation of the wave phenomenon in either Symbiodiniaceae strains investigated here.
Temperature Dependency of the Flash-Induced Chlorophyll Fluorescence Relaxation and the Wave Phenomenon in Symbiodiniaceae
Because the ratio of the activities of PSI : PSII is one of the crucial factors that determine the inducibility of the wave phenomenon, it was an important question to clarify whether the different strains display temperature dependent loss of PSII activity and if the decrease of PSII activity relative to PSI activity results in the wave formation in fluorescence relaxation. Increasing the temperature up to 40°C revealed that, although the wave formation did not occur in any of the investigated strains by heat treatment alone, the dip and the subsequent increase of fluorescence were the most apparent in S. tridacnidorum 2465 (Figure 4B). However, both strong reduction of the PQ pool and partial decrease in PSII activity are typically required in microalgae (C. reinhardtii) for the wave formation (Krishna et al., 2019; Patil et al., 2022). Based on the results of the present work, this is the case for Symbiodiniaceae as well; in heat-treated cells (which caused the loss of PSII activity relative to PSI activity), microaerobic treatment induced the wave formation, which was the most expressed in S. tridacnidorum 2465. The heat-induced changes in flash-induced fluorescence relaxation were also manifested in the relative decrease of variable fluorescence (the difference between maximum fluorescence, Fm(ST), and minimum fluorescence, F0), increase in F0, and changes in the fluorescence relaxation profile (Figure 4). Elevated temperatures were shown to primarily impact oxygen evolving capacity and, thereby, PSII activity in corals, although the oxygen evolving complex was not found to be the primary site of damage of the photochemical apparatus during coral bleaching (Hill and Ralph, 2008b). Furthermore, in Symbiodiniaceae, PSII is more sensitive to heat than PSI (Figure 3) (Hoogenboom et al., 2012); therefore, progressively increasing temperature causes progressively increasing imbalance in the electron flow from PSII that reduces the PQ pool and PQ pool oxidation via the forward electron transfer to PSI, which favors the formation of wave phenomenon. However, heat treatment itself up to 40°C was not sufficient to induce the wave; to observe this phenomenon, microaerobic condition was also crucial, which generates strong redox poise for the PQ pool. The tolerance of reef building corals to various stressors determine the ability of coral reef ecosystems to cope with acute and chronic changes in abiotic environmental conditions (Kühl et al., 1995; Ulstrup et al., 2005; Hughes et al., 2020). Although the temperatures applied in our work are typically well above the normal temperature conditions experienced by coral reefs, extreme environments also exist in which the temperature rises from 36°C to 38°C under which heat tolerant corals can survive. Despite the high temperature tolerance threshold of certain corals, they still remain highly susceptible to bleaching when, for example, peak summer temperature exceeds the upper threshold of their thermal tolerance [reviewed, e.g., in (Camp et al., 2018)]. Therefore, we suggest that, although the temperature treatment scenario in our work could not represent widespread or global environmental conditions, extreme climatic conditions similar to the temperatures applied in the current study may occur and, thus, represent naturally relevant scenarios in some cases. Therefore, the wave phenomenon of flash-induced chlorophyll fluorescence relaxation could be a valuable environmental indicator of heat stress in Symbiodiniaceae, which occurs in combination of hypoxic conditions in coral reef ecosystems. The temperature-dependent features of the formation of the wave phenomenon could decipher strain-specific tolerance to heat stress, although it has to be noted that the capacity for different alternative electron flow processes was not found to be unequivocally correlated with long-term heat tolerance in Symbiodiniaceae (Dang et al., 2019).
Potential Relationship Between the Wave Phenomenon and CEF in Symbiodiniaceae
The formation of the wave was assigned to the operation of the components of CEF, such as the NDH complexes in cyanobacteria (Deák et al., 2014) and some eukaryotic microalgae (Krishna et al., 2019; Patil et al., 2022). Our observations that the wave formation occurred under conditions that was previously found to induce CEF in Symbiodiniaceae (i.e., acute heat stress) (Aihara et al., 2016; Dang et al., 2019) indicate that the wave phenomenon could be potentially assigned to CEF also in this species, although it has to be noted that the wave phenomenon occurred at higher temperatures as compared to the temperature that induced CEF in the earlier studies. The genome of certain strains of Symbiodiniaceae contains copies of both the PGR5/PGRL1 and NDH-2 (Aihara et al., 2016); therefore, it is possible that these pathways play a potential role in the wave formation in this genera; however, at this stage, no experimental evidence can be provided for the involvement of specific components or CEF pathways in the induction of the wave. Other alternative electron transfer pathways in Symbiodiniaceae such as Mehler reaction, PTOX, or FLV proteins (Roberty et al., 2014) could possibly contribute to the regulation of the PQ pool reduction state and the dip-rise kinetics of fluorescence relaxation. However, the wave phenomenon was observed to largest extent in microaerobic condition of heat-treated cells, under which photoreduction of oxygen by these alternative electron transport pathways is not functional. This argues for the involvement of CEF, which also accelerates under microaerobic conditions in Symbiodiniaceae (Aihara et al., 2016). However, similarly to C. reinhardtii, it is quite unlikely that the wave reflects an actual flash-induced cycling of electrons from the acceptor side of PSI to the PQ pool (Patil et al., 2022) because of the rise of the fluorescence after the dip phase occurs in several seconds and the re-reduction of P700+via CEF occurs in ms timescale [e.g., (Alric, 2010)]. Furthermore, it is unlikely that the components like PGR5/PGRL1 mediate fast CEF pathways in Symbiodiniaceae, similar to recent findings in Chlamydomonas as it has been shown that the maximal rate of CEF was similar in the pgrl1 mutant and in wild-type Chlamydomonas (Nawrocki et al., 2019). However, the extent of CEF can be modulated from very low values to a maximal value by modulating the redox state of the donor and acceptor side of PSI (Nawrocki et al., 2019); therefore, it is plausible to investigate the relationship between the different kinetic phases in flash-induced fluorescence relaxation profiles and the kinetics of CEF and its regulatory components, which requires further studies in Symbiodiniaceae.
In conclusion, flash-induced fluorescence relaxation is a sensitive indicator of heat stress in Symbiodiniaceae. These algae displayed the so-called wave phenomenon when the PSII activity is partially decreased and the PQ pool becomes strongly reduced in microaerobic conditions. The wave phenomenon and heat sensitivity of the flash-induced fluorescence relaxation displayed species-specific features; therefore, this method is highly informative about the species-specific characteristics of the redox reactions of the PQ pool. The appearance of the wave phenomenon could possibly be related to CEF processes in Symbiodiniaceae; however, this remains to be investigated further. The flash-induced fluorescence relaxation is therefore a valuable non-intrusive marker of the stress-induced changes in photosynthetic electron transport and therefore could be used as a sensitive detection tool for coral bleaching.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Author Contributions
Conceptualization, IV and MS; methodology, IV, SM, PP, and MS; validation, IV and MS; formal analysis, IV, MS, and SM; investigation, SM, PP, and MS; resources, IV and MS; data curation, IV and MS; writing—original draft preparation, IV, SM, and MS; writing—review and editing, IV and MS; visualization, SM and MS; supervision, IV and MS; project administration, IV and MS; funding acquisition, IV and MS. All authors contributed to the article and approved the submitted version.
Funding
This research was funded by the National Research, Development and Innovation Office, grant number FK128977 and the Hungarian Academy of Sciences, MTA Premium Postdoctoral Research Program, grant number PREMIUM-2017–38.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors thank Prof. Michael Kühl, University of Copenhagen, for providing some of the strains.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.932355/full#supplementary-material
References
Abrego D., Ulstrup K. E., Willis B. L., Van Oppen M. J. (2008). Species-Specific Interactions Between Algal Endosymbionts and Coral Hosts Define their Bleaching Response to Heat and Light Stress. Proc. Biol. Sci. 275, 2273–2282. doi: 10.1098/rspb.2008.0180
Aihara Y., Takahashi S., Minagawa J. (2016). Heat Induction of Cyclic Electron Flow Around Photosystem I In the Symbiotic Dinoflagellate Symbiodinium. Plant Physiol. 171, 522–529. doi: 10.1104/pp.15.01886
Allen J. F. (2002). Photosynthesis of ATP—Electrons, Proton Pumps, Rotors, and Poise. Cell 110, 273–276. doi: 10.1016/S0092-8674(02)00870-X
Alric J. (2010). Cyclic Electron Flow Around Photosystem I in Unicellular Green Algae. Photosynth. Res. 106, 47–56. doi: 10.1007/s11120-010-9566-4
Asada K., Heber U., Schreiber U. (1993). Electron Flow to the Intersystem Chain from Stromal Components and Cyclic Electron Flow in Maize Chloroplasts, as Detected in Intact Leaves by Monitoring Redox Change. Plant Cell Physiol. 34, 39–50. doi: 10.1093/oxfordjournals.pcp.a078398
Buxton L., Takahashi S., Hill R., Ralph P. J. (2012). Variability in the Primary Site of Photosynthetic Damage in Symbiodinium SO. (Dinophyceae) Exposed to Thermal Stress. J. Phycol. 48, 117–126. doi: 10.1111/j.1529-8817.2011.01099.x
Camp E. F., Schoepf V., Mumby P. J., Hardtke L. A., Rodolfo-Metalpa R., Smith D. J., et al. (2018). The Future of Coral Reefs Subject to Rapid Climate Change: Lessons from Natural Extreme Environments. Front. Mar. Sci. 5. doi: 10.3389/fmars.2018.00004
Cardol P., Forti G., Finazzi G. (2011). Regulation of Electron Transport in Microalgae. Biochim. Biophys. Acta (BBA)-Bioenerg. 1807, 912–918. doi: 10.1016/j.bbabio.2010.12.004
Claquin P., Rene-Trouillefou M., Lopez P. J., Japaud A., Bouchon-Navaro Y., Cordonnier S., et al. (2021). Singular Physiological Behavior of the Scleractinian Coral Porites Astreoides in the Dark Phase. Coral reefs 40, 139–150. doi: 10.1007/s00338-020-02023-4
Cser K., Vass I. (2007). Radiative and Non-Radiative Charge Recombination Pathways in Photosystem II Studied by Thermoluminescence and Chlorophyll Fluorescence in the Cyanobacterium Synechocystis 6803. Biochim. Biophys. Acta 1767, 233–243. doi: 10.1016/j.bbabio.2007.01.022
Dang K. V., Pierangelini M., Roberty S., Cardol P. (2019). Alternative Photosynthetic Electron Transfers and Bleaching Phenotypes Upon Acute Heat Stress in Symbiodinium and Breviolum spp. (Symbiodiniaceae) in Culture. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00656
Davy S. K., Allemand D., Weis V. M. (2012). Cell Biology of Cnidarian-Dinoflagellate Symbiosis. Microbiol. Mol. Biol. Rev. 76, 229–261. doi: 10.1128/MMBR.05014-11
Deák Z., Sass L., Kiss É., Vass I. (2014). Characterization of Wave Phenomena in the Relaxation of Flash-Induced Chlorophyll Fluorescence Yield in Cyanobacteria. Biochim. Biophys. Acta (BBA)-Bioenerg. 1837, 1522–1532. doi: 10.1016/j.bbabio.2014.01.003
Díaz-Almeyda E. M., Prada C., Ohdera A. H., Moran H., Civitello D. J., Iglesias-Prieto R., et al. (2017). Intraspecific and Interspecific Variation in Thermotolerance and Photoacclimation in Symbiodinium Dinoflagellates. Proc. R. Soc B: Biol. Sci. 284, 20171767. doi: 10.1098/rspb.2017.1767
Downs C., Fauth J. E., Halas J. C., Dustan P., Bemiss J., Woodley C. M. (2002). Oxidative Stress and Seasonal Coral Bleaching. Free Rad. Biol. Med. 33, 533–543. doi: 10.1016/S0891-5849(02)00907-3
Ermakova M., Huokko T., Richaud P., Bersanini L., Howe C. J., Lea-Smith D. J., et al. (2016). Distinguishing the Roles of Thylakoid Respiratory Terminal Oxidases in the Cyanobacterium Synechocystis Sp. PCC 6803. Plant Physiol. 171, 1307–1319. doi: 10.1104/pp.16.00479
Fisher P. L., Malme M. K., Dove S. (2012). The Effect of Temperature Stress on Coral-Symbiodinium Associations Containing Distinct Symbiont Types. Coral Reefs 31, 473–485. doi: 10.1007/s00338-011-0853-0
Gorbunov M. Y., Falkowski P. G. (2021). Using Chlorophyll Fluorescence Kinetics to Determine Photosynthesis in Aquatic Ecosystems. Limnol. Oceanogr. 66, 1–13. doi: 10.1002/lno.11581
Gorbunov M. Y., Kolber Z. S., Lesser M. P., Falkowski P. G. (2001). Photosynthesis and Photoprotection in Symbiotic Corals. Limnol. Oceanogr. 46, 75–85. doi: 10.4319/lo.2001.46.1.0075
Gotoh E., Matsumoto M., Ogawa K. I., Kobayashi Y., Tsuyama M. (2010). A Qualitative Analysis of the Regulation of Cyclic Electron Flow Around Photosystem I From the Post-Illumination Chlorophyll Fluorescence Transient in Arabidopsis: A New Platform for the In Vivo Investigation of the Chloroplast Redox State. Photosynth. Res. 103, 111–123. doi: 10.1007/s11120-009-9525-0
Havurinne V., Tyystjärvi E. (2020). Photosynthetic Sea Slugs Induce Protective Changes to the Light Reactions of the Chloroplasts They Steal From Algae. Elife 9, 1–29 doi: 10.7554/eLife.57389.sa2
Hawkins T. D., Krueger T., Wilkinson S. P., Fisher P. L., Davy S. K. (2015). Antioxidant Responses to Heat and Light Stress Differ With Habitat in a Common Reef Coral. Coral Reefs 34, 1229–1241. doi: 10.1007/s00338-015-1345-4
Hennige S. J., Suggett D. J., Warner M. E., Mcdougall K. E., Smith D. J. (2009). Photobiology of Symbiodinium Revisited: Bio-Physical and Bio-Optical Signatures. Coral Reefs 28, 179–195. doi: 10.1007/s00338-008-0444-x
Hill R., Larkum A. W., Frankart C., Kühl M., Ralph P. J. (2004). Loss of Functional Photosystem II Reaction Centres in Zooxanthellae of Corals Exposed to Bleaching Conditions: Using Fluorescence Rise Kinetics. Photosynth. Res. 82, 59–72. doi: 10.1023/B:PRES.0000040444.41179.09
Hill R., Ralph P. J. (2008a). Dark-Induced Reduction of the Plastoquinone Pool in Zooxanthellae of Scleractinian Corals and Implications for Measurements of Chlorophyll a Fluorescence. Symbiosis 46, 45–56.
Hill R., Ralph P. J. (2008b). Impact of Bleaching Stress on the Function of the Oxygen Evolving Complex of Zooxanthellae From Scleractinian Corals. J. Phycol. 44, 299–310. doi: 10.1111/j.1529-8817.2008.00468.x
Hill R., Szabó M., Ur Rehman A., Vass I., Ralph P. J., Larkum A. W. (2014). Inhibition of Photosynthetic CO2 Fixation in the Coral Pocillopora Damicornis and its Relationship to Thermal Bleaching. J. Exp. Biol. 217, 2150–2162. doi: 10.1242/jeb.100578
Hill R., Ulstrup K. E., Ralph P. J. (2009). Temperature Induced Changes in Thylakoid Membrane Thermostability of Cultured, Freshly Isolated, and Expelled Zooxanthellae From Scleractinian Corals. Bull. Mar. Sci. 85, 223–244.
Hoadley K. D., Warner M. E. (2017). Use of Open Source Hardware and Software Platforms to Quantify Spectrally Dependent Differences in Photochemical Efficiency and Functional Absorption Cross Section Within the Dinoflagellate Symbiodinium Spp. Front. Mar. Sci. 4. doi: 10.3389/fmars.2017.00365
Hoogenboom M. O., Campbell D. A., Beraud E., Dezeeuw K., Ferrier-Pages C. (2012). Effects of Light, Food Availability and Temperature Stress on the Function of Photosystem II and Photosystem I of Coral Symbionts. PloS One 7, e30167. doi: 10.1371/journal.pone.0030167
Houille-Vernes L., Rappaport F., Wollman F.-A., Alric J., Johnson X. (2011). Plastid Terminal Oxidase 2 (PTOX2) is the Major Oxidase Involved in Chlororespiration in Chlamydomonas. Proc. Natl. Acad. Sci. U.S.A 108, 20820–20825. doi: 10.1073/pnas.1110518109
Hughes D. J., Alderdice R., Cooney C., Kühl M., Pernice M., Voolstra C. R., et al. (2020). Coral Reef Survival Under Accelerating Ocean Deoxygenation. Nat. Climate Change 10, 296–307. doi: 10.1038/s41558-020-0737-9
Hughes T. P., Kerry J. T., Baird A. H., Connolly S. R., Dietzel A., Eakin C. M., et al. (2018). Global Warming Transforms Coral Reef Assemblages. Nature 556, 492–496. doi: 10.1038/s41586-018-0041-2
Jones R. J., Hoegh-Guldberg O., Larkum A. W. D., Schreiber U. (1998). Temperature-Induced Bleaching of Corals Begins With Impairment of the CO2 Fixation Mechanism in Zooxanthellae. Plant Cell Environ. 21, 1219–1230. doi: 10.1046/j.1365-3040.1998.00345.x
Kalaji H. M., Schansker G., Brestic M., Bussotti F., Calatayud A., Ferroni L., et al. (2017). Frequently Asked Questions About Chlorophyll Fluorescence, the Sequel. Photosynth. Res. 132, 13–66. doi: 10.1007/s11120-016-0318-y
Kalaji H. M., Schansker G., Ladle R. J., Goltsev V., Bosa K., Allakhverdiev S. I., et al. (2014). Frequently Asked Questions About In Vivo Chlorophyll Fluorescence: Practical Issues. Photosynt. Res. 122, 121–158. doi: 10.1007/s11120-014-0024-6
Klughammer C., Schreiber U. (1994). An Improved Method, Using Saturating Light-Pulses, for the Determination of Photosystem-I Quantum Yield via P700+-Absorbency Changes at 830 Nm. Planta 192, 261–268. doi: 10.1007/BF01089043
Klughammer C., Schreiber U. (2008). Saturation Pulse Method for Assessment of Energy Conversion in PSI. PAM Appl. Notes 1, 11–14.
Kramer D. M., Evans J. R. (2011). The Importance of Energy Balance in Improving Photosynthetic Productivity. Plant Physiol. 155, 70–78. doi: 10.1104/pp.110.166652
Krishna P. S., Morello G., Mamedov F. (2019). Characterization of the Transient Fluorescence Wave Phenomenon That Occurs During H2 Production in Chlamydomonas Reinhardtii. J. Exp. Bot. 70, 6321–6336. doi: 10.1093/jxb/erz380
Kühl M., Cohen Y., Dalsgaard T., Jørgensen B. B., Revsbech N. P. (1995). Microenvironment and Photosynthesis of Zooxanthellae in Scleractinian Corals Studied With Microsensors for O2;, pH and Light. Mar. Ecol. Prog. Ser. 117, 159–172. doi: 10.3354/meps117159
Lajeunesse T. C., Parkinson J. E., Gabrielson P. W., Jeong H. J., Reimer J. D., Voolstra C. R., et al. (2018). Systematic Revision of Symbiodiniaceae Highlights the Antiquity and Diversity of Coral Endosymbionts. Curr. Biol. 28, 2570–2580.e2576. doi: 10.1016/j.cub.2018.07.008
Leggat W., Whitney S., Yellowlees D. (2004). Is Coral Bleaching Due to the Instability of the Zooxanthellae Dark Reactions? Symbiosis 37, 137–153.
Lesser M. P. (2011). ““Coral Bleaching: Causes and Mechanisms,”,” in Coral Reefs: An Ecosystem in Transition. Eds. Dubinsky Z., Stambler N. ( Springer) Dordrecht, 405–419.
Lesser M. P. (2019). Phylogenetic Signature of Light and Thermal Stress for the Endosymbiotic Dinoflagellates of Corals (Family Symbiodiniaceae). Limnol. Oceanogr. 64, 1852–1863. doi: 10.1002/lno.11155
Lilley R. M., Ralph P. J., Larkum A. W. (2010). The Determination of Activity of the Enzyme Rubisco in Cell Extracts of the Dinoflagellate Alga Symbiodinium Sp. By Manganese Chemiluminescence and its Response to Short-Term Thermal Stress of the Alga. Plant Cell Environ. 33, 995–1004. doi: 10.1111/j.1365-3040.2010.02121.x
Maxwell K., Johnson G. N. (2000). Chlorophyll Fluorescence—a Practical Guide. J. Exp. Bot. 51, 659–668. doi: 10.1093/jexbot/51.345.659
Nawrocki W. J., Bailleul B., Cardol P., Rappaport F., Wollman F. A., Joliot P. (2019). Maximal Cyclic Electron Flow Rate is Independent of PGRL1 in Chlamydomonas. Biochim. Biophys. Acta (BBA)-Bioenerg. 1860, 425–432. doi: 10.1016/j.bbabio.2019.01.004
Nitschke M. R., Gardner S. G., Goyen S., Fujise L., Camp E. F., Ralph P. J., et al. (2018). Utility of Photochemical Traits as Diagnostics of Thermal Tolerance Amongst Great Barrier Reef Corals. Front. Mar. Sci. 5. doi: 10.3389/fmars.2018.00045
NOAA National Centers for Environmental Information State of the Climate:Monthly Global Climate Report for 2020, published online January 2021, https://www.ncei.noaa.gov/access/monitoring/monthly-report/global/202013/supplemental/page-1
Oakley C. A., Schmidt G. W., Hopkinson B. M. (2014). Thermal responses of Symbiodinium photosynthetic carbon assimilation. Coral Reefs 33, 501–512. doi: 10.1007/s00338-014-1130-9
Patil P. P., Mohammad Aslam S., Vass I., Szabó M. (2022). Characterization of the Wave Phenomenon of Flash-Induced Chlorophyll Fluorescence in Chlamydomonas Reinhardtii. Photosynth. Res. doi: 10.1007/s11120-022-00900-3
Peltier G., Aro E.-M., Shikanai T. (2016). NDH-1 and NDH-2 Plastoquinone Reductases in Oxygenic Photosynthesis. Annu. Rev. Plant Biol. 67, 55–80. doi: 10.1146/annurev-arplant-043014-114752
Rehman A. U., Szabó M., Deák Z., Sass L., Larkum A., Ralph P., et al. (2016). Symbiodinium sp. Cells Produce Light-Induced Intra-and Extracellular Singlet Oxygen, which Mediates Photodamage of the Photosynthetic Apparatus and has the Potential to Interact with the Animal Host in Coral Symbiosis. New Phytol. 212, 472–484. doi: 10.1111/nph.14056
Reynolds J. M., Bruns B. U., Fitt W. K., Schmidt G. W. (2008). Enhanced Photoprotection Pathways in Symbiotic Dinoflagellates of Shallow-Water Corals and Other Cnidarians. Proc. Natl. Acad. Sci. U.S.A 105, 13674–13678. doi: 10.1073/pnas.0805187105
Roberty S., Bailleul B., Berne N., Franck F., Cardol P. (2014). PSI Mehler Reaction is the Main Alternative Photosynthetic Electron Pathway in Symbiodinium sp., Symbiotic Dinoflagellates of Cnidarians. New Phytol. 204, 81–91. doi: 10.1111/nph.12903
Roberty S., Furla P., Plumier J. C. (2016). Differential Antioxidant Response Between two Symbiodinium Species from Contrasting Environments. Plant Cell Environ. 39, 2713–2724. doi: 10.1111/pce.12825
Roth M. S. (2014). The engine of the reef: Photobiology of the Coral–Algal Symbiosis. Front.Microbiol. 5. doi: 10.3389/fmicb.2014.00422
Shikanai T., Yamamoto H. (2017). Contribution of Cyclic and Pseudo-Cyclic Electron Transport to the Formation of Proton Motive Force in Chloroplasts. Mol. Plant 10, 20–29. doi: 10.1016/j.molp.2016.08.004
Slavov C., Schrameyer V., Reus M., Ralph P. J., Hill R., Büchel C., et al. (2016). “Super-Quenching” State Protects Symbiodinium from Thermal Stress—Implications for Coral Bleaching. Biochim. Biophys. Acta (BBA)-Bioenerg. 1857, 840–847. doi: 10.1016/j.bbabio.2016.02.002
Smith D. J., Suggett D. J., Baker N. R. (2005). Is Photoinhibition of Zooxanthellae Photosynthesis the Primary Cause of Thermal Bleaching in Corals? Global Change Biol. 11, 1–11. doi: 10.1111/j.1529-8817.2003.00895.x
Strasser R. J., Govindjee, (1992). “The Fo and the O-J-I-P Fluorescence Rise in Higher-Plants and Algae” in Regulation of Chloroplast Biogenesis. Ed. Argyroudi-Akoyunoglou J. H., 423–426. Nato ASI Series. Springer New York, NY
Suggett D. J., Goyen S., Evenhuis C., Szabó M., Pettay D. T., Warner M. E., et al. (2015). Functional Diversity of Photobiological Traits Within the Genus Symbiodinium Appears to be Governed by the Interaction of Cell Size With Cladal Designation. New Phytol. 208, 370–381. doi: 10.1111/nph.13483
Suggett D. J., Nitschke M. R., Hughes D. J., Bartels N., Camp E. F., Dilernia N., et al. (2022). Toward Bio-Optical Phenotyping of Reef-Forming Corals Using Light-Induced Fluorescence Transient-Fast Repetition Rate Fluorometry. Limnol. Oceanogr. Methods 20, 172–191. doi: 10.1002/lom3.10479
Suggett D. J., Warner M. E., Smith D. J., Davey P., Hennige S., Baker N. R. (2008). Photosynthesis and production of hydrogen peroxide by symbiodinium (pyrrhophyta) phylotypes with different thermal tolerances. J. Phycol. 44, 948–956. doi: 10.1111/j.1529-8817.2008.00537.x
Szabó M., Larkum A. W., Suggett D. J., Vass I., Sass L., Osmond B., et al. (2017). Non-intrusive assessment of photosystem ii and photosystem i in whole coral tissues. Front. Mar. Sci. 4, 269. doi: 10.3389/fmars.2017.00269
Takahashi S., Whitney S., Itoh S., Maruyama T., Badger M. (2008). Heat stress causes inhibition of the de novo synthesis of antenna proteins and photobleaching in cultured symbiodinium. Proc. Natl. Acad. Sci. U.S.A 105, 4203–4208. doi: 10.1073/pnas.0708554105
Tchernov D., Gorbunov M. Y., De Vargas C., Yadav S. N., Milligan A. J., Haggblom M., et al. (2004). Membrane Lipids of Symbiotic Algae are Diagnostic of Sensitivity to Thermal Bleaching in Corals. Proc. Natl. Acad. Sci. U.S.A 101, 13531–13535. doi: 10.1073/pnas.0402907101
Tóth S. Z., Schansker G., Garab G., Strasser R. J. (2007a). Photosynthetic Electron Transport Activity in Heat-Treated Barley Leaves: The Role of Internal Alternative Electron Donors to Photosystem II. Biochim. Biophys. Acta-Bioenerg. 1767, 295–305. doi: 10.1016/j.bbabio.2007.02.019
Tóth S. Z., Schansker G., Strasser R. J. (2007b). A Non-Invasive Assay of the Plastoquinone Pool Redox State Based on the Ojip-Transient. Photosynth. Res. 93, 193–203. doi: 10.1007/s11120-007-9179-8
Trtílek M., Kramer D. M., Koblížek M., Nedbal L. (1997). Dual-Modulation Led kinetic Fluorometer. J. Lumin. 72-74, 597–599. doi: 10.1016/S0022-2313(97)00066-5
Ulstrup K. E., Hill R., Ralph P. J. (2005). Photosynthetic Impact of Hypoxia on in Hospite Zooxanthellae in the Scleractinian Coral Pocillopora Damicornis. Mar. Ecol. Prog. Ser. 286, 125–132. doi: 10.3354/meps286125
Vass I., Kirilovsky D., Etienne A.-L. (1999). UV-B Radiation-Induced Donor- and Acceptor-Side Modifications of Photosystem II In the Cyanobacterium Synechocystis sp. pcc 6803. Biochemistry 38, 12786–12794. doi: 10.1021/bi991094w
Vass I., Styring S., Hundal T., Koivuniemi A., Aro E., Andersson B. (1992). Reversible and Irreversible Intermediates During Photoinhibition of Photosystem II: Stable Reduced QA Species Promote Chlorophyll Triplet Formation. Proc. Natl. Acad. Sci. U.S.A 89, 1408–1412. doi: 10.1073/pnas.89.4.1408
Vega De Luna F., Córdoba-Granados J. J., Dang K.-V., Roberty S., Cardol P. (2020). In vivo Assessment of Mitochondrial Respiratory Alternative Oxidase Activity and Cyclic Electron Flow Around Photosystem I: On Small Coral Fragments. Sci. Rep. 10, 17514. doi: 10.1038/s41598-020-74557-0
Venn A. A., Loram J. E., Douglas A. E. (2008). Photosynthetic Symbioses in Animals. J. Exp. Bot. 59, 1069–1080. doi: 10.1093/jxb/erm328
Volgusheva A., Kruse O., Styring S., Mamedov F. (2016). Changes in the Photosystem II: Complex Associated with Hydrogen Formation in Sulfur Deprived Chlamydomonas Reinhardtii. Algal Res. 18, 296–304. doi: 10.1016/j.algal.2016.06.025
Volgusheva A., Styring S., Mamedov F. (2013). Increased Photosystem II: Stability Promotes H2 Production in Sulfur-Deprived Chlamydomonas Reinhardtii. Proc. Natl. Acad. Sci. U.S.A 110, 7223–7228. doi: 10.1073/pnas.1220645110
Warner M. E., Lesser M. P., Ralph P. J. (2010). “Chlorophyll Fluorescence in Reef Building Corals,” in Chlorophyll a Fluorescence in Aquatic Sciences: Methods and Applications. Eds. Suggett D. J., Prášil O., Borowitzka M. A. ( Springer) Dordrecht, 209–222.
Warner M. E., Suggett D. J. (2016). “The Photobiology of Symbiodinium spp.: Linking Physiological Diversity to the Implications of Stress and Resilience”, in The Cnidaria, Past, Present and Future. Eds. Goffredo S., Dubinsky Z. ( Springer) Cham, 489–509.
Keywords: photosynthesis, chlorophyll fluorescence, heat stress, cyclic electron flow, Symbiodiniaceae
Citation: Mohammad Aslam S, Patil PP, Vass I and Szabó M (2022) Heat-Induced Photosynthetic Responses of Symbiodiniaceae Revealed by Flash-Induced Fluorescence Relaxation Kinetics. Front. Mar. Sci. 9:932355. doi: 10.3389/fmars.2022.932355
Received: 29 April 2022; Accepted: 22 June 2022;
Published: 22 July 2022.
Edited by:
Christine Ferrier-Pagès, Centre Scientifique de Monaco, MonacoReviewed by:
Stephane Roberty, University of Liège, BelgiumKenneth Hoadley, University of Alabama, United States
Copyright © 2022 Mohammad Aslam, Patil, Vass and Szabó. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Imre Vass, dmFzcy5pbXJlQGJyYy5odQ==; Milán Szabó, c3phYm8ubWlsYW5AYnJjLmh1