 Xavier Turon1*
Xavier Turon1* Jesús Zarcero1
Jesús Zarcero1 Adrià Antich1
Adrià Antich1 Owen Simon Wangensteen2
Owen Simon Wangensteen2 Enric Ballesteros1
Enric Ballesteros1 Emma Cebrian1
Emma Cebrian1 Candela Marco-Méndez1
Candela Marco-Méndez1 Teresa Alcoverro1
Teresa Alcoverro1- 1Department of Marine Ecology, Centre for Advanced Studies of Blanes (CEAB-CSIC), Catalonia, Spain
- 2Department of Evolutionary Biology, Ecology and Environmental Sciences and Biodiversity Research Institute (IRBio), University of Barcelona, Catalonia, Spain
Against the accelerating pace of worldwide species extinction, reliable biodiversity assessments are critical, both as baselines and to track potential declines. DNA metabarcoding techniques allow for fast and comprehensive assessment of biodiversity in both terrestrial and marine habitats. However, these methods need to be adapted and standardised for each ecosystem in order to be effective. Seagrass meadows are among the most diverse marine habitats and are irreplaceable in terms of the ecosystem services they provide, yet metabarcoding has never been implemented for these systems. In this study, we developed and tested a protocol for metabarcoding the eukaryotic community of meadows of the iconic species, Posidonia oceanica L. (Delile). This seagrass is the main habitat-forming species in Mediterranean coastal waters and is known for its high diversity due to the structural complexity of its canopy and rhizome structures. This habitat is experiencing a range-wide retreat, and there is an urgent need for fast and efficient methods for its biomonitoring and detection of early changes. Our proposed method involves direct sampling of the community, collecting and processing the leaves and rhizome strata separately. To test the utility of the method in distinguishing between different meadow conditions, we sampled two distinct meadows that differ in their prevailing wind and surge conditions, and a nearby rocky reef for comparison. We then adapted a method and pipeline for COI metabarcoding using generalist primers that target the eukaryote diversity present. We detected a high diversity in the two meadows analysed (3,350 molecular operational taxonomic units, dominated by Metazoa and Archaeplastida) and a clear differentiation of the seagrass samples from those of the nearby rocky reefs. The leaves and rhizomes harboured clearly distinct assemblages, and differences were also detected between the two meadows sampled. This new tool has the potential to deliver big biodiversity data for seagrass habitats in a fast and efficient way, which is crucial for the implementation of protection and management measures for this key coastal habitat.
Introduction
The Mediterranean is a Sea under siege by a suite of anthropogenic pressures. Compared to more open seas, this enclosed basin is particularly threatened by stressors related to the current climate crisis (Lejeusne et al., 2010; Garrabou et al., 2022). The habitats dominated by the endemic Mediterranean seagrass Posidonia oceanica L. (Delile) are among the most diverse biologically, and most valuable in terms of ecological services, in the Mediterranean (Traganos et al., 2022). Healthy P. oceanica meadows are responsible for high rates of sediment stabilization and erosion prevention, perform important nursery functions, enhance food webs, and contribute significantly to carbon burial (Campagne et al., 2015). However, P. oceanica meadows are suffering a range-wide decline (Marbà et al., 2014) that is sped up by climate change (Marbà & Duarte, 2010; Jordà et al., 2012), with carry-on consequences for its attendant biodiversity. These effects can be partially balanced by genetic and functional mechanisms (Aurelle et al., 2022; Edgeloe et al., 2022) that can contribute to the seagrass resilience. Besides the ecological and economic consequences that a regression of this habitat would have, centuries of carbon stored by these meadows would be released with meadow decline (Marbà et al., 2014), further aggravating the climate warming trend.
Given the acknowledged precarious state of P. oceanica meadows, several conservation programmes have been established, the effectiveness of which relies upon our ability to provide indicators of ecosystem health that are sensitive to changes in condition, biodiversity and state. These indicators include morphological and structural parameters (e.g., shoot density, cover) and the biodiversity of the associated biota. Biomonitoring of seagrass meadows has become an increasingly key element for assessing ecological impacts, particularly at early stages, to inform the implementation of adequate protection and management measures. Biodiversity assessments are the foundation of any biomonitoring effort. Over the last few decades, conventional morphological techniques have identified several hundreds of species of macro-organisms inhabiting P. oceanica beds (e.g., Borg et al., 2006; Como et al., 2008; Kalogirou et al., 2010), making it one of the most diverse nearshore habitats in temperate seas (reviewed in Piazzi et al., 2016). However, the meiofaunal component of P. oceanica meadows, although a major contributor to their biodiversity (García-Gómez et al., 2022), is often neglected in traditional morphological studies.
Morphology-based census methods are slow to implement and, crucially, are dependent on a worldwide dwindling taxonomic knowledge base (the so-called “taxonomic impediment”, Engel et al., 2021). The scale and rate at which these data can be obtained cannot match the pace at which changes are accruing to these ecosystems. The use of DNA-based methods for taxa identification has been proposed as a new paradigm in biomonitoring (Pawlowski et al., 2021) and has proven useful in marine benthic studies, where it is able to sample a much higher diversity than conventional methods in a fast and efficient way (Steyaert et al., 2020; Duarte et al., 2021). The application of metabarcoding to DNA obtained from the environment (environmental DNA sensu lato, Rodriguez-Ezpeleta et al., 2021), whereby one or a few genetic markers are amplified and used to build amplicon sequencing libraries, is so far the most common approach for DNA-based biodiversity assessment. In the case of the marine benthos, most studies focus on sediment biota (e.g., Fonseca et al., 2014; Brannock et al., 2016; Guardiola et al., 2016; Atienza et al., 2020). Reef communities have received comparatively much less attention, and protocols designed for them include the deployment and recovery of standardized settlement units (Cahill et al., 2018; Obst et al., 2020), or the direct sampling of the community via quadrat scraping or other methods (Wangensteen et al., 2018; De Jode et al., 2019; Shum et al., 2019).
There is as yet no protocol designed for metabarcoding seagrass meadows, and we are in urgent need of developing this tool for these threatened habitats to be able to take prompt action to manage them. One of the main advantages of metabarcoding is its ability to capture, not just the macro-organismal diversity, but also the meio- and micro-organisms present in the habitat (Wangensteen et al., 2018). To date, only Cowart et al. (2015) have applied metabarcoding techniques to sediment cores obtained in Zostera marina meadows to assess their diversity. Reef et al. (2017) also sampled DNA from sediments in seagrass meadows to determine the sources of sediment organic carbon, while Menning et al. (2021) used environmental DNA from water, sediment and leaves of Z. marina to look specifically for seagrass pathogens. Some studies have been performed on DNA from seawater collected in seagrass meadows targeting fish (He et al., 2022; Momota et al., 2022) and dinoflagellates (Jacobs-Palmer et al., 2020). However, it has been shown that, to efficiently capture eukaryotic benthic biodiversity, metabarcoding of water samples is not adequate, and that direct sampling of the benthic assemblage is required (Antich et al., 2021a).
The goal of this paper is to fill this methodological gap by developing and testing a protocol for metabarcoding the eukaryotic community inhabiting P. oceanica meadows, including field methods for direct sampling of the habitat and lab methods and a bioinformatic pipeline customized for the analysis of Cytochrome Oxidase I (COI) amplicons using generalist primers targeting the eukaryote community. This case study is situated in a marine protected area (Spanish National Park), where we sampled two seagrass beds with contrasting levels of wave exposure and nearby rocky shallow water reefs for comparison.
Methods
Sampling
Samples were obtained in September 2020 by scuba diving in the Cabrera Archipelago of the Balearic Islands (Western Mediterranean). This area was declared National Park (the highest protection figure in Spanish law) in 1991, and covers ca. 90,000 Ha of maritime domain. A map of the sampling locations is shown in Figure S1. We collected samples from two seagrass meadows in Cabrera Island, Cala Santa Maria (coordinates: 39.1506, 2.9427) and L’Olló (39.1502, 2.9637). They had different orientations, and are thus subject to different wind and surge conditions, with Cala Santa Maria exposed to northerlies and westerlies, and L’Olló to easterlies. Both meadows were located at 10-12 m depth. A monitoring study in Cala Santa Maria showed the positive effects of the regulations derived from the implementation of the National Park in this seagrass meadow (Marbà et al., 2002). For comparison, we used samples taken at the same dates from a nearby vertical rocky wall located in an islet adjacent to Cabrera (La Imperial: 39.1251, 2.9603), where a metabarcoding monitoring has been going on since 2014 (Wangensteen et al., 2018). We sampled two depths on this wall corresponding to a shallow (-10 m) and a deep habitat (-30 m). These rocky bottom habitats were dominated by algal assemblages with sponges and other invertebrates, and are described in Wangensteen et al. (2018).
Samples of Posidonia oceanica meadows were taken in triplicate at each site with a corer 20 cm in diameter inserted in the meadow down to 10 cm depth. The leaves were cut and placed in plastic bags underwater. The corresponding top 10 cm of the rhizome mat were also placed separately in plastic bags. For the rocky reef habitats, the method developed in Wangensteen et al. (2018) was used: 25x25 quadrats (six at each depth) were scraped to bare rock with hammer and chisel and collected in plastic bags. Samples were fixed within the hour by eliminating water through a sieve of 63 µm mesh size, and replacing it with absolute alcohol. Samples were stored at -20 °C until further processing.
Once in the laboratory, the samples of P. oceanica were treated as follows: the leaves were individually scraped with commercial razor blades (replaced between samples) to collect all epiphytes. Whole shoots were processed, including young and older leaves, and both basal and distal portions of them. Rhizomes were similarly scraped with razor blades, detaching the scales and their epiphytes until only the central core remained. All collected matter was then sieved under gentle pressure freshwater through a stainless-steel sieve of 63 µm mesh size and the material retained was kept for further processing. The rocky bottom samples were separated into two fractions using a column of two stainless-steel sieves (1 mm and 63 µm mesh size) under gentle pressure freshwater. This size fractionation was necessary given large size differences of organisms (Wangensteen & Turon, 2017), whereas no size fractionation was required for the seagrass material as it was relatively homogenous in size. The material from the leave and rhizome fractions of P.oceanica, as well as from the two size fractions of the rocky bottom samples was homogenized with a blender (Bosch CleverMix) and constituted the samples for downstream analyses. All equipment was washed and bleached between samples. Three negative controls were added by charring sand samples in a muffle furnace (24 h at 400 °C) and sieving and processing them as the rest of samples.
For DNA extraction, 5 g of each homogenate was processed with the PowerMax Soil DNA Isolation Kit (QIAGEN). DNA concentration was assessed with a Qubit fluorometer (Life Technologies) and was in all cases in excess of 5 ng/µl. Note that in preliminary assays we found that higher DNA yield was obtained using ca. 5 g of material rather than the 10 g that can be processed with this kit.
For the analysis of the COI marker, we amplified the “Leray fragment” of ca. 313 bp (Leray et al., 2013). We used the primers mlCOIintF-XT (forward): 5’-GGWACWRGWTGRACWITITAYCCYCC-3’ (Wangensteen et al., 2018b) and jgHCO2198 (reverse): 5’-TAIACYTCIGGRTGICCRAARAAYCA-3’ (Geller et al., 2013). Three PCR blanks were performed without adding the DNA template. Both primers had 8-base sample-specific tags attached. For a given sample, the same tag was used at the forward and reverse primer (so inter-sample chimeras could be easily spotted). In addition, a variable number (2-4) of fully degenerated positions (Ns) was added to the 5’ end to enhance molecular diversity during sequencing. PCR conditions followed Atienza et al. (2020). PCR products were pooled at equal volume and libraries were prepared with the BIOO NEXTFLEX PCR-Free DNA-Seq Kit (Perkin-Elmer). Sequencing was performed in a partial NovaSeq 2*250 bp run by NovoGene Company.
Bioinformatics pipeline
We processed the sequencing reads following the MJOLNIR pipeline (https://github.com/uit-metabarcoding/MJOLNIR), that combines several software programs, mostly from the OBITools package (Boyer et al., 2016), for the different steps. Illuminapairedend was used to align paired-end reads and keep only those with >40 alignment quality score. Reads were then demultiplexed and primer sequences removed using ngsfilter. Those with mismatched primer tags at any end were discarded. Obigrep and obiuniq were used to perform a length filter (retaining only reads between 299-320 bp) and dereplicate sequences. Singleton sequences (with one read) were eliminated at this step and the Uchime-denovo algorithm from VSEARCH (Rognes et al., 2016) was used to remove chimeric amplicons.
There are two main procedures to reduce metabarcoding datasets: either denoising by merging reads of potentially erroneous sequences with the correct one, or clustering sequences into molecular operational units (MOTUs), pooling the reads and keeping the most abundant sequence as the MOTU representative. It has been shown that, for variable markers, both are complementary (Antich et al., 2021b; Brandt et al., 2021). Denoising is necessary if intra-MOTU variation is the target; in our case, as we were interested in biodiversity at the MOTU level (as a surrogate of species), we proceeded with clustering. We used the SWARM procedure (Mahé et al., 2015), that has the advantage of not having a fixed distance threshold and being independent of input order. It is a fast procedure that relies on a single-linkage method with an initial clustering distance (d), followed by a topological refining of the clusters using abundance structures to divide them into MOTUs. We set parameter d to 13 as recommended for this marker (Antich et al., 2021b).
MOTUs were then submitted to several filters: first, we eliminated all MOTUs for which there were more than 10% of the reads in control samples (negatives or blanks). Second, for each sample, we set to 0 those MOTUs with less than 5 reads (to get rid of potential sample cross-talk). We then removed all MOTUs left with 0 reads across all samples in the previous step. We ran LULU (Frøslev et al., 2017) to remove potentially erroneous MOTUs. With the remaining MOTUs we performed taxonomic assignment using ecotag against a local taxonomy database (downloaded from NCBI and BOLD and curated, current version July 2021), available at https://github.com/uit-metabarcoding/DUFA/ and containing 174,544 COI sequences. Ecotag searches the best hit in the reference database and builds the set of sequences in the database which are at least as similar to the best hit as the query sequence is. Then, the MOTU is assigned to the most recent common ancestor to all these sequences in the NCBI taxonomy tree. With this procedure, the assigned taxonomic rank varies depending on the similarity of the query sequences and the density of the reference database (Boyer et al., 2016). Finally, we filtered the MOTU dataset to eliminate prokaryote (or unassignable) and non-marine sequences. The later were searched manually and deleted whenever the taxonomic assignment was precise enough to determine that they were non-marine.
Community analyses
Most analyses were based on the R package vegan (Oksanen et al., 2019). We obtained MOTU accumulation curves as per read depth with function rarecurve and as per number of samples with specaccum. The rocky reef samples were part of a long-term study (started in 2014, Wangensteen et al., 2018) and, for the purposes of this work, the two size-fractions were combined by averaging the number of reads of each MOTU. We thus had a total of 12 samples of seagrass meadows (2 localities x 2 strata – leaf/rhizome - x 3 replicates) and 12 from rocky bottoms (2 depths x 6 replicates).
To assess the habitat composition, we combined MOTUs into the major eukaryotic super-groups as in Atienza et al. (2020) and, for metazoans, into the different phyla present. MOTU richness and Shannon diversity values were computed from samples rarefied to the minimal number of reads in the samples (function rarefy) and compared across habitat type (two rocky bottom habitats and P. oceanica meadows separated into leaf and rhizome fractions) with ANOVA.
For β-diversity analyses we used the square-root transformed relative read abundance (i.e., the Hellinger transform) of each MOTU in each sample, and computed the Bray-Curtis dissimilarity index (BC). These values were used in a non-metric multidimensional scaling (nmMDS) using the metaMDS function. We also performed permutational analyses of variance (PERMANOVA) on the BC matrix. The PERMANOVA module incorporated in the Primer v6 statistical package (Anderson et al., 2008) was used. We first compared habitat type (rocky-seagrass) as fixed factor. A second test was performed for the seagrass samples with two fixed factors: locality and stratum (leaf or rhizomes). Tests of multivariate dispersions (permdisp) were run when the main factors were significant to determine whether this outcome was a result of different multivariate means or different heterogeneity (spread) of the groups.
Finally, we performed and Indicator Species analysis with the IndVal index (Dufrêne & Legendre, 1997) to assess the MOTUs significantly associated with the different types of habitats. We used the R package indicspecies (De Cáceres and Legendre, 2009) and compared first samples from rocky habitats with those from the P. oceanica meadows. In a second run we compared samples from leaves and rhizomes. With this procedure we identified the MOTUs associated with the rocky habitats first and, within the seagrass meadows, with leaves and rhizomes separately.
Results
The sequencing run of the COI amplifications yielded a total of 53,535,478 reads. After elimination of chimeras and singletons there were 48,618,706 reads remaining, grouped into 1,319,257 unique sequences. These sequences were clustered by SWARM in 10,042 initial MOTUs. After all subsequent filters (particularly the elimination of non-eukaryotes), the final COI dataset consisted of 5,316 MOTUs, 41,557,672 reads and 1,051,377 unique sequences. The average number of reads per sample was 1,135,527 ± 272,513 (mean ± SE), while negatives and blanks had 10,422 ± 2.984 reads. The original sequences have been uploaded to the NCBI SRA archive (Bioproject PRJNA944186). The final MOTU table, with taxonomic assignments and representative sequences, is provided as Supplementary Table S1.
The composition of the samples in terms of proportion of the MOTUs of the different super-groups is presented in Figure 1A (samples from the two P. oceanica meadows combined). Metazoa was the most diverse group in all cases, but more so in the rocky reef samples. The second group was the Archaeplastida, corresponding mostly to rhodophytes. Stramenopiles was the third group in number of MOTUs, and Amebozoa the fourth. In comparatively very low numbers, Alveolata, Fungi, Rhizaria, Hacrobia and Apusozoa followed suit (grouped under “Other” in Figure 1A). The pattern regarding the proportion of reads (rather than of MOTUs) showed a similar trend (Figure 1B), with Metazoa, Archaeplastida, and Stramenopiles as the most abundant groups, while the remaining ones had negligible abundance. The dominance of metazoans in the rocky bottom samples was more marked than when considering MOTU richness, while in number of reads Archaeplastida was clearly more abundant in P. oceanica samples. Of course, the detection of P. oceanica reads would bias abundance estimations. However, our COI primers did not amplify this species due to mismatches in the primer binding region (see in vitro analysis in Supplementary File SF1). This was confirmed by the absence of any plant MOTU (we even searched among the unassigned MOTUs) in our results that matches the expected pattern of high abundance in the seagrass samples and low abundance in the rocky samples.
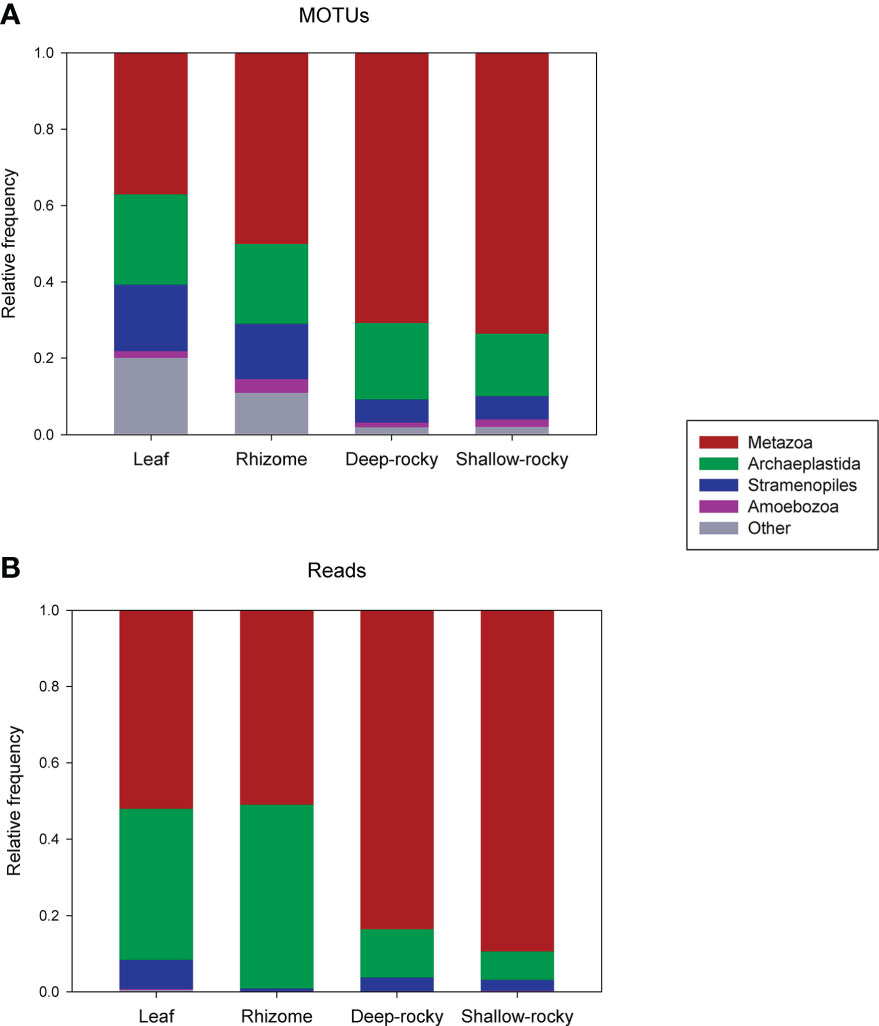
Figure 1 (A), proportion of the assigned MOTUs that belong to the main Super-Groups of Eukarya considered; (B), proportion of reads represented by these Super-Groups.
The distribution of metazoan phyla is shown in Figure 2A as proportion of MOTUs in the samples. 17 phyla were present, of which Arthropoda, Porifera, Annelida and Cnidaria, in this order, were the most diverse overall. A higher diversity of Porifera and Cnidaria was apparent in the rocky reef samples, while more Arthropoda, Annelida and Bryozoa MOTUs were found in the seagrass samples. When considering proportion of reads (Figure 2B), the most abundant phyla were Porifera, Cnidaria, Annelida and Bryozoa. There was a higher abundance of cnidarians in the benthic rocky bottom samples, while there was a high abundance of bryozoans in leaves and of sponges in the rhizomes of P. oceanica samples.
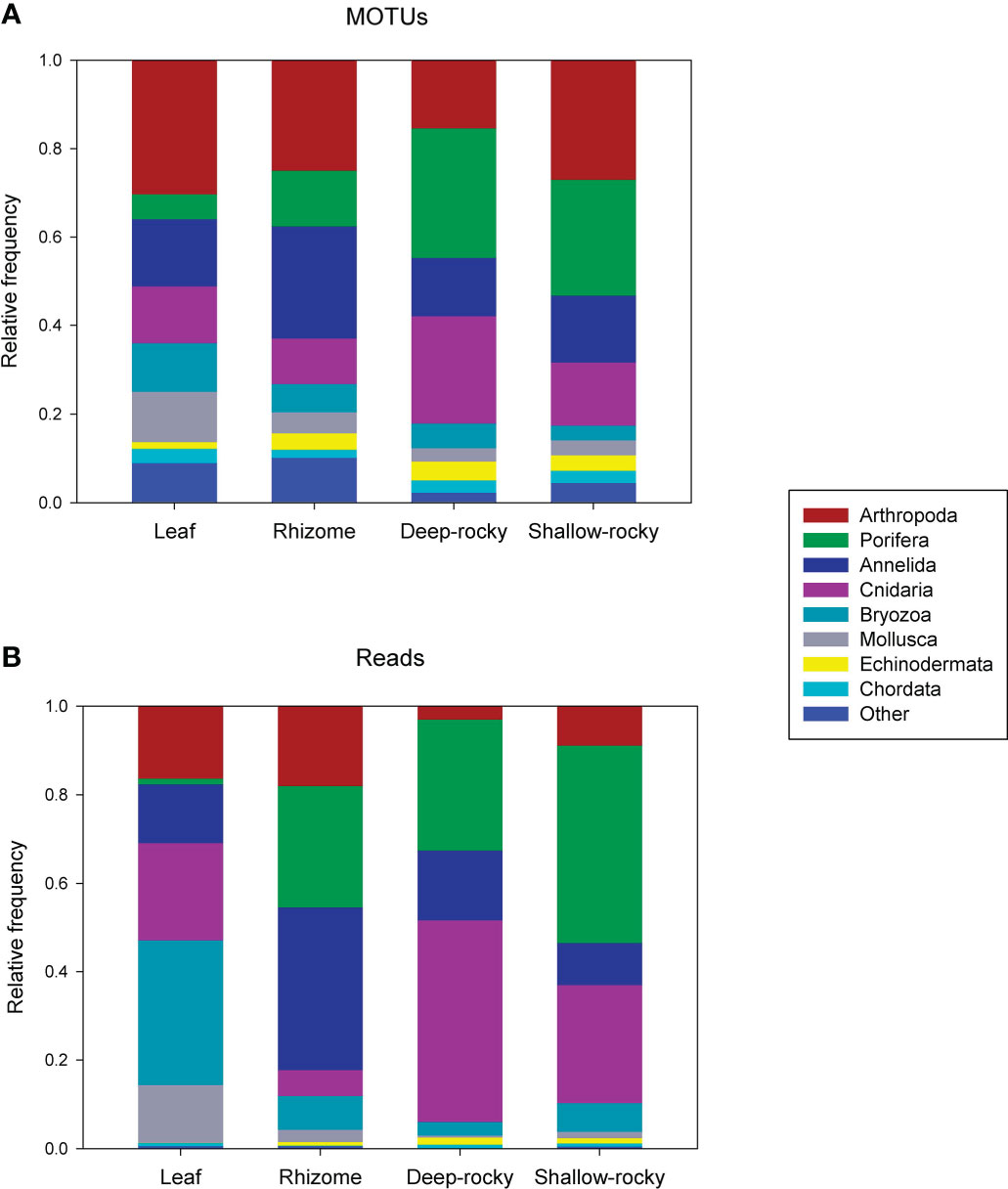
Figure 2 (A), proportion of the assigned MOTUs that belong to the main metazoan phyla found; (B), proportion of reads represented by these phyla.
Overall, the seagrass samples were more diverse (3,350 MOTUs in total) than the rocky reef ones (2,863 MOTUs), and both were quite distinct (only 897 shared MOTUs). Seagrass rhizomes harboured more MOTUs than the leaves (2,403 vs 1,573, with 626 shared MOTUs). The mean MOTU richness per sample of the different habitats varied from 261 to 454 (once samples were rarefied to 39,649 reads, corresponding to the sample with less reads), with lowest values in the rhizome habitat and highest in the leaf and the deep rocky community (Figure 3). However, high variances prevented detection of significant differences (ANOVA, p=0.350). The same pattern was found considering Shannon’s diversity index (Figure 3), with values between ca. 2 to 3.9. For this variable, the diversity values were significantly higher in leaves and the deep rocky reef habitat than in rhizomes (ANOVA p=0.034 followed by Fisher’s LSD post-hoc tests).
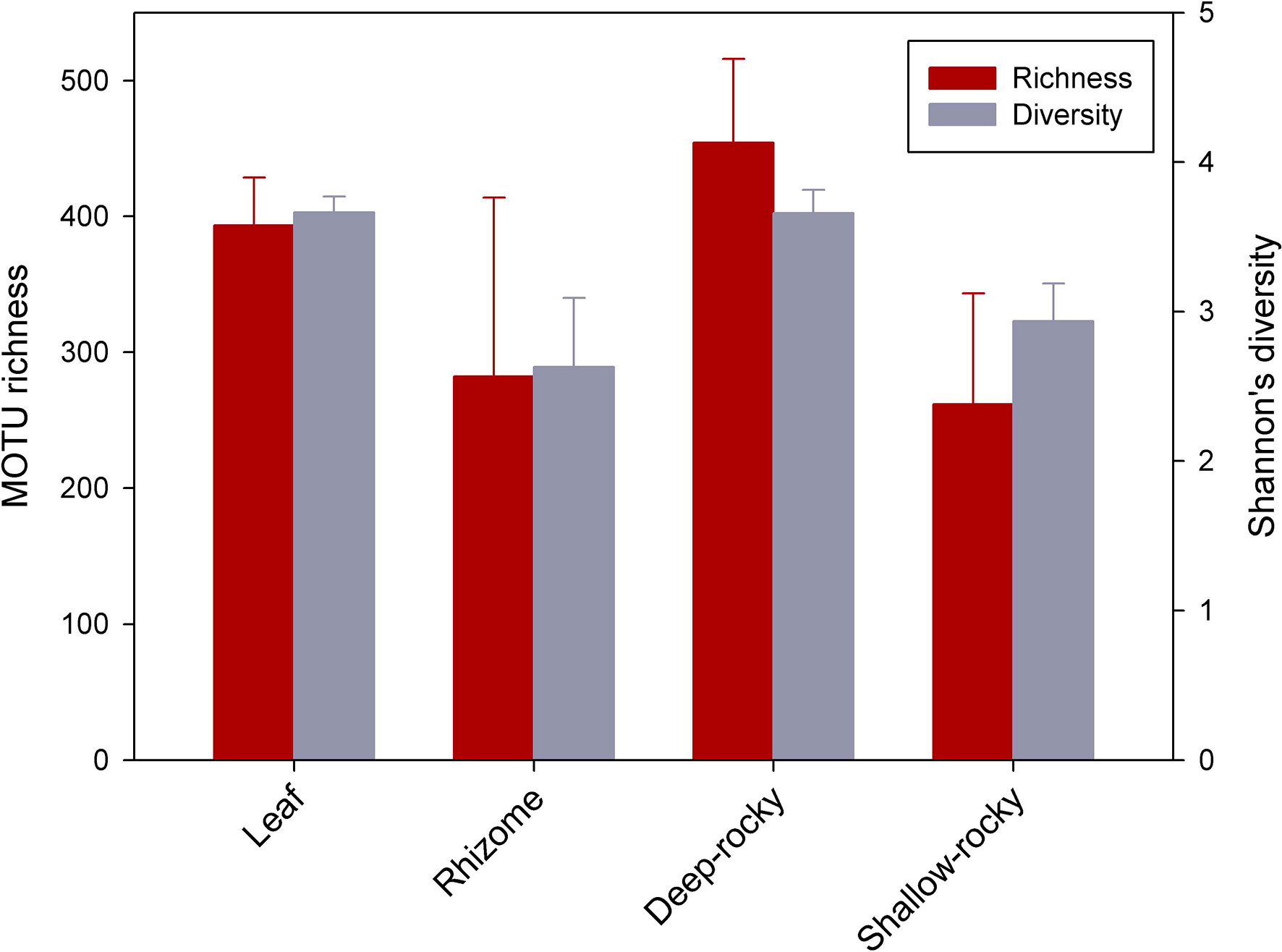
Figure 3 Mean MOTU richness (after rarefaction to a common number of reads), and mean Shannon diversity values of the samples corresponding to the main habitats sampled. Bars are standard errors.
MOTU accumulation curves with increasing read numbers using the rarefied dataset (Figure S2A) showed that, in general, a plateau is reached even at the lowest sequencing depth in our samples. If we exclude the rare MOTUs (i.e., those representing the 5% of the reads), the plateau is more marked (Figure S2B). When we plot MOTU accumulation curves with increasing sample number, the trend is that the number of MOTUs detected increased more or less steadily when combining samples, with no clear levelling-off even when combining the 6 samples available for each habitat type (Figure S3A). The same analysis excluding the rare MOTUs (5% of reads) reveals that accumulation curves tend to plateau (less markedly for the rhizome samples, Figure S3B).
The ordination of the samples in a nmMDS plot (Figure 4) showed a clear separation of the P. oceanica leaves and rhizomes along the first dimension, and the two meadows also formed distinct clusters for each component. The rocky reef samples grouped by depth and were well separated from the P. oceanica samples. The second dimension of the plot separated the shallow rocky reef samples plus the P. oceanica leaf samples from the deep rocky bottom and the rhizome samples.
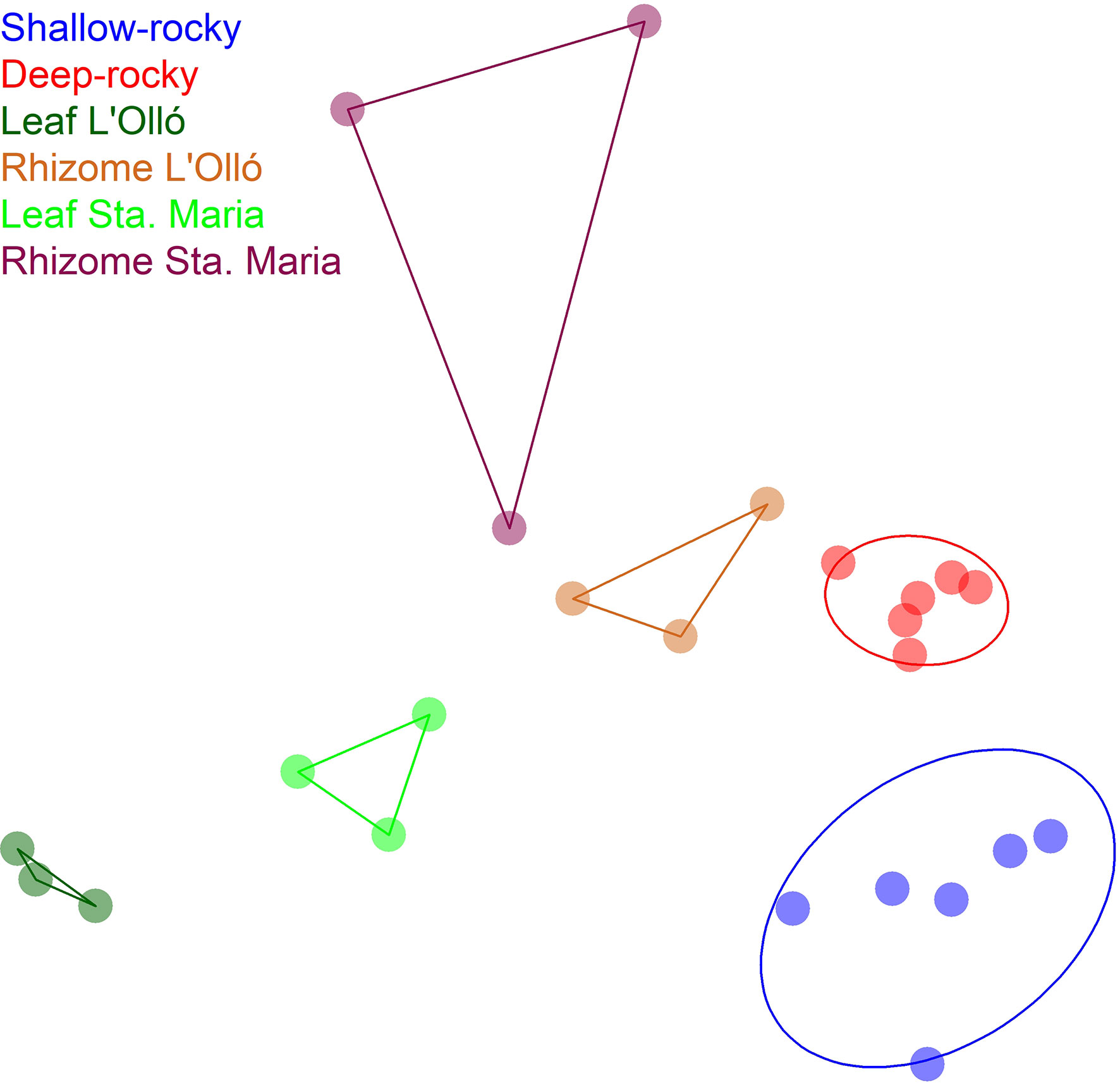
Figure 4 Non-metric multidimensional scaling representation of the samples using the Bray-Curtis distance. Hull lines group the samples of leaves and rhizomes of the two meadows studied. Ellipses are 95% confidence intervals around the centroids of the rocky reef samples. Stress of the configuration: 0.134.
PERMANOVA analyses (Table 1) highlighted a significant differentiation between the seagrass meadows and the rocky bottom habitats, and a significantly higher heterogeneity in the former. Among the seagrass samples, there were significant differences linked to the stratum analysed (leaf vs rhizome, with the latter having also more heterogeneity) and the meadow sampled (Table 1), while the interaction was not significant. The stratum factor explained twice the variance than the meadow factor. Accordingly, when the P. oceanica samples were clustered and plotted in a heatmap (Figure 5), there was a clear-cut separation between leaf and rhizome samples. The former clustered separately for the two meadows, while for the rhizomes this separation was not complete.
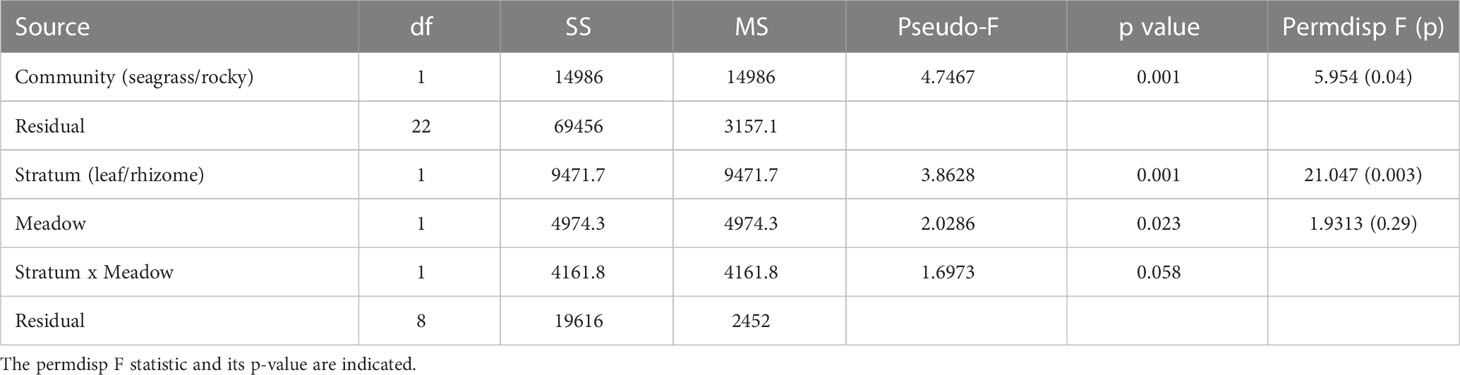
Table 1 Results of the PERMANOVA analyses considering the two main habitat types sampled and, for Posidonia oceanica samples, the effect of the stratum and meadow.
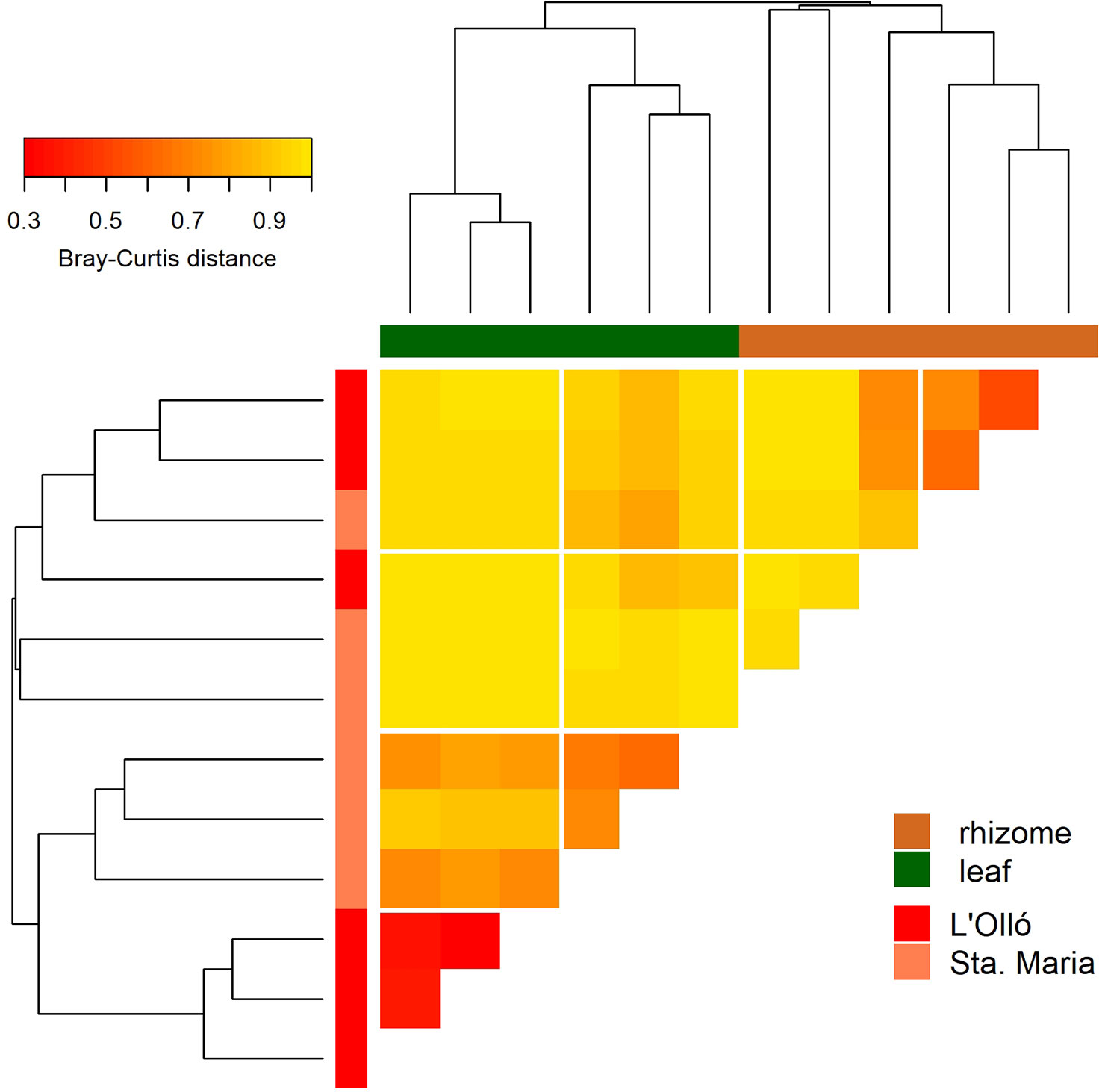
Figure 5 Heatmap and cluster of the Posidonia oceanica samples based on the Bray-Curtis distance. Colour codes indicate stratum (horizontal bars) or meadow (vertical bars).
The results of the Indicator Species analysis are presented in Supplementary Table S2, where the MOTUs significantly associated with the leaf, rhizome, and rocky habitats are listed together with their mean abundance in the respective habitat and the taxonomic assignment. It is noteworthy that leaves and rocky habitat had more MOTUs associated (186 and 220, respectively), than the rhizomes (25 MOTUs). There were also differences in the relative abundance of these indicator MOTUs, that together represented 52.51% and 58.26%, respectively, of the reads in the samples of leaves and rocky habitats, while they comprised only 10.22% of the reads from the rhizome samples.
The composition (at the phylum level) of these MOTUs is summarized in Figure S4 as proportion of MOTUs and reads. The dominant groups among the indicator MOTUs were Rhodophyta, Annelida, and Porifera for leaves, rhizomes, and rocky habitats, respectively. This trend is more marked when considering proportion of reads than that of MOTUs. The number of MOTUs and reads that could not be assigned at phylum level was higher in leaves. We also examined species-level assignments among the indicator MOTUs, but considered only those with a best identity of >95% with the reference database (see Table S2). For the leaf samples, 14 reliable species names could be assigned, and the three most abundant (in relative read abundance) were Laurencia obtusa (Rhodophyta), Mytilus galloprovincialis (Mollusca), and Leptosiphonia brodiei (Rhodophyta). Among the rhizome indicator MOTUs no assignment could be made below the order level. For the rocky samples, 29 indicator MOTUs could be given a species name, and the three more abundant were Ircinia variabilis (Porifera), Paracentrotus lividus (Echinodermata), and Sarcotragus spinulosus (Porifera).
Discussion
The metabarcoding protocol developed here uncovered a high diversity of eukaryotes inhabiting P. oceanica seagrass meadows in the Mediterranean, with 3,350 operational taxonomic units detected in just two meadows. There were clear differences between the leaf and the rhizome stratum, making it necessary to sample and analyse both strata separately in seagrass metabarcoding. Both were markedly different from habitats present in nearby rocky reefs. The stratum (leaf/rhizome) was the main factor explaining differences among seagrass samples but, interestingly, there were also significant differences between the two meadows sampled, which were subject to different regimes of prevailing winds. This proves that the method can detect relatively subtle differences between meadows and, thus, can be a powerful tool to test the effects of diverse stressors on P. oceanica beds, well before these stressors cause visible alterations.
It is surprising that no metabarcoding study has been thus far attempted in P. oceanica meadows, given that this seagrass is the main habitat-forming species of Mediterranean coastal waters, with ca. 20,000 km2 mapped basin-wide in the first 25m of depth (Traganos et al., 2022). So far, the only comprehensive metabarcoding survey of seagrass beds was performed on Zostera marina meadows and used sediment corers instead of sampling the actual habitat created by the plant strata (Cowart et al., 2015). We present here a direct method for sampling P. oceanica beds, which we believe will provide a much more precise information on the biota inhabiting these. Direct sampling has been successfully applied to metabarcoding studies in other highly diverse benthic habitats (Wangensteen et al., 2018; Antich et al., 2023).
There were shifts in both diversity and abundance of major groups of eukaryotes between seagrass and rocky bottom habitats, although in all cases metazoans were the dominant group, followed by Archaeplastida. Among metazoans, seagrass leaves had a high abundance (in relative number of reads) of bryozoans and cnidarians, while the rhizome stratum was dominated by sponges and annelids. These results match classical taxonomic approaches that report a high diversity of bryozoans and cnidarians epiphytic on leaves of Posidonia oceanica (Casola et al., 1987; Martínez-Crego et al., 2010) and sponges and annelids in rhizomes (Gambi et al., 2005; Padiglia et al., 2015). In the rocky reef samples, on the other hand, the most abundant groups were cnidarians and sponges, followed by annelids, coherent with previous metabarcoding studies of rocky bottoms (Antich et al., 2023). A higher diversity per sample (both in terms of MOTU richness and Shannon index) was detected in the leaf than in the rhizome samples. A high dispersion of the rhizome samples was also apparent (nmMDS ordination and permdisp analyses), which was coherent with the fact that this habitat harboured more total MOTUs than the leaf habitat, but lower per sample richness. The indicator species analysis detected that leaves and rocky reef habitats had many MOTUs significantly associated with them, while rhizomes were relatively poor in indicator MOTUs. Leaves had a high proportion of significantly associated photosynthetic groups (60 out of 186 indicator MOTUs, comprising 37.26% of the reads), while bryozoans (7.16% of reads) and arthropods (2.96% of reads) were the most abundant indicator metazoan groups. In the rhizomes and the rocky environment, the metazoans were the best represented among the indicator MOTUs, in particular annelids (63.05% of reads) in the rhizomes and sponges and cnidarians in the rocky habitats (38.20% and 19.40% of reads, respectively).
A taxon-by-taxon correspondence between metabarcoding and morphological studies is hardly achievable, due to primer bias and incompleteness of reference databases in metabarcoding, and low coverage of small and cryptic taxa and lack of taxonomic expertise for accurate identification of multiple groups in morphological surveys (Kelly et al., 2017; Steyaert et al., 2020; Keck et al., 2022). When these comparisons have been made, generally a higher diversity was uncovered by metabarcoding and a low overlap in terms of taxa has been found, but in most cases the same ecological patterns have been detected with both types of data (e.g., Cowart et al., 2015; Zaiko et al., 2016; Cahill et al., 2018; Duarte et al., 2023). Thus, morphology and metabarcoding provide complementary windows on taxonomic information, but are consistent in telling the same story in terms of ecological processes. Albeit we haven’t analysed morphologically our samples, our results are in agreement with what is known from morphological studies (see above). Thus, although morphological-based studies remain invaluable, we are confident that general trends of change in seagrass meadows can be reliably detected with the faster method of metabarcoding and can be implemented for biomonitoring this crucial habitat.
Rarefaction curves showed that we could adequately capture the MOTU diversity present in our samples. At the habitat level, our sample size was adequate to detect the MOTUs representing most of the reads, but not sufficient to assess the rare biosphere (those MOTUs comprising less than 5% of reads) present in the communities, and more replicates are advisable in future studies. This higher replication would also better capture the heterogeneity of rhizome samples mentioned above. It was very convenient that our primers did not amplify the seagrass itself, as this would have resulted in a highly reduced number of reads of other organisms, hindering a correct assessment of richness and abundances. It can be noted that ca. 33% of the eukaryote MOTUs could not be assigned to any Super-Group and, within Metazoa, ca. 34% of the MOTUs could not be assigned even at phylum level. These figures point to the need to improve existing gaps in the reference databases (Wangensteen et al., 2018) to reduce the number of unidentified taxa and allow better informed ecological interpretation of the patterns detected.
Taxonomically broad analyses of communities via metabarcoding can be based on several techniques, each with advantages and shortcomings. The use of universal primers is one of them, but other alternatives are the combination of markers, the use of primer cocktails, or PCR-free approaches (Ficetola and Taberlet, 2023). We chose to use the COI marker and a primer set that has been shown to amplify most eukaryote groups (Wangensteen et al., 2018; Antich et al., 2023). However, no primer is truly universal, and we noted above the lack of amplification of the seagrass itself. These primers are also ill-suited for the detection of another important group, the Foraminifera (Macher et al., 2022). Some compromise must be sought depending on the objectives of each study. In our case, for a general assessment of biodiversity, we selected a marker and primer set for which data are being steadily generated on marine communities and will thus allow for comparisons in the future. However, other primer combinations or other markers should be used for studies focused on particular groups.
In this study we used the first 10 cm of the rhizome mat, as we were targeting the living community. However, the well-developed mats of rhizomes and roots can be several metres thick and constitute an archive of past (over millennia) oceanographic conditions and biota. The latter can be analysed by paleoecological methods, including metabarcoding of ancient DNA currently applied to other environments, such as lake sediments (Domaizon et al., 2017). In seagrass beds, Wesselmann et al., 2022 analysed sediments in Mediterranean meadows (Cymodocea nodosa and P. oceanica) to determine decadal changes (over the last century) in metazoan composition. Piñeiro-Juncal et al. (2021), analysed the prokaryote composition of corers (1.5 m long) of soil under P. oceanica meadows at different degradation states. An interesting possibility is that, if corers are obtained including deeper layers of the P. oceanica mat, our protocols can be coupled with ancient DNA techniques applied to the older parts of the sample, thus providing a complete picture of past and present biota in these assemblages. Likewise, the protocol can be adapted for more detailed studies of the leaf epiphytes, for instance considering separately young and old leaves in a shoot, or basal and apical areas of the leaves, depending on the ecological question being addressed.
This study confirms that P. oceanica habitat harbours a rich and diverse ecosystem that ranges across several groups despite being structured by a single species. On the one hand, P. oceanica leaves can serve as habitat for short-lived epifauna and epiflora that are inherently fast-growing, with a turnover rate of less than 100 days (Alcoverro et al., 2001). At the same time, P. oceanica is one of the most stable and long-lived habitats in the Mediterranean (Duarte, 1991), enabling many long-lived species to establish among its complex rhizome structures. The meadow spreads through clonal growth, allowing it to occupy large tracts of the shallow bottoms (Ruggiero et al., 2002), making it an ideal habitat for a vast diversity of species. The P. oceanica meadows can act as “natural samplers” (Mariani et al., 2019; Turon et al., 2020), constituting a natural reservoir of the DNA shed to the environment. The high beta-diversity found between meadows of different orientations is also interesting, as it indicates that species compositions could be highly environmentally determined, with physical forces playing an important role in habitat assembly. Even our limited sampling was sensitive to these compositional differences, highlighting the utility of metabarcoding in tracking potential changes to ecosystem state and function.
In conclusion, this work presents a metabarcoding protocol for seagrass meadows that relies on directly sampling the habitat, collecting and processing the leaf and the rhizome strata separately. We also describe lab and bioinformatics procedures to target the eukaryotic community present in the samples. Our case study in two Mediterranean meadows shows the feasibility and sensitivity of this approach for biomonitoring these important habitats. We trust that this innovative method will spark future research on these threatened and iconic ecosystems.
Data availability statement
The data presented in the study are deposited in the NCBI SRA repository, accession number PRJNA944186.
Author contributions
XT, TA, CM-M and AA designed the research. AA, EB, EC, TA, CM-M and XT performed the samplings. AA, OW and JZ performed laboratory work. AA, JZ, OW and XT did bioinformatics and statistical analyses. XT prepared figures and tables and drafted the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This research was funded by project BIGPARK (2462/2017) of the Spanish Autonomous Organism of National Parks (OAPN), and by projects MARGECH (PID2020-118550RB) and STORM (PID2020-113745RB-I00) of the Spanish Ministry of Science and Innovation (MCIN/AEI/10.13039/501100011033). AA and JZ were supported by grants PRE2018-085664 and PRE2021-097703, respectively of the Spanish Ministry of Science and Innovation and by “ESF Investing in your future”.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2023.1145883/full#supplementary-material
References
Alcoverro T., Cebrian E., Ballesteros E. (2001). The photosynthetic capacity of the seagrass Posidonia oceanica: Influence of nitrogen and light. J. Exp. Mar. Biol. Ecol. 261, 107–120. doi: 10.1016/S0022-0981(01)00267-2
Anderson M. J., Gorley R. N., Clarke K. R. (2008). PERMANOVA+ for PRIMER: Guide to software and statistical methods (Plymouth: Primer-E Ltd).
Antich A., Palacín C., Cebrian E., Golo R., Wangensteen O. S., Turon X. (2021b). Marine biomonitoring with eDNA: Can metabarcoding of water samples cut it as a tool for surveying benthic communities? Mol. Ecol. 30, 3175–3188. doi: 10.1111/mec.15641
Antich A., Palacín C., Wangensteen O. S., Turon X. (2021a). To denoise or to cluster, that is not the question: Optimizing pipelines for COI metabarcoding and metaphylogeography. BMC Bioinf. 22 (1), 177. doi: 10.1186/s12859-021-04115-6
Antich A., Palacín C., Zarcero J., Wangensteen O. S., Turon X. (2023). Metabarcoding reveals high-resolution biogeographical and metaphylogeographical patterns through marine barriers. J. Biogeogr. 50, 515–527. doi: 10.1111/jbi.14548
Atienza S., Guardiola M., Praebel K., Antich A., Turon X., Wangensteen O. S. (2020). DNA Metabarcoding of deep-sea sediment communities using COI: Community assessment, spatio-temporal patterns and comparison with 18S rDNA. Diversity 12, 123. doi: 10.3390/d12040123
Aurelle D., Thomas S., Albert C., Bally M., Bondeau A., Boudouresque C. F., et al. (2022). Biodiversity, climate change, and adaptation in the Mediterranean. Ecosphere 13, e3915. doi: 10.1002/ecs2.3915
Borg J. A., Rowden A. A., Attrill M. J., Schembri P. J., Jones M. B. (2006). Wanted dead or alive: High diversity of macroinvertebrates associated with living and ‘dead’ Posidonia oceanica matte. Mar. Biol. 149, 667–677. doi: 10.1007/s00227-006-0250-3
Boyer F., Mercier C., Bonin A., Le Bras Y., Taberlet P., Coissac E. (2016). Obitools: A unix-inspired software package for DNA metabarcoding. Mol. Ecol. Res. 16, 176–182. doi: 10.1111/1755-0998.12428
Brandt M. I., Trouche B., Quintric L., Günther B., Wincker P., Poulain J., et al. (2021). Bioinformatic pipelines combining denoising and clustering tools allow for more comprehensive prokaryotic and eukaryotic metabarcoding. Mol. Ecol. Res. 21, 1904–1921. doi: 10.1111/1755-0998.13398
Brannock P. M., Wang L., Ortmann A., Waits D. S., Halanych K. M. (2016). Genetic assessment of meiobenthic community composition and spatial distribution in coastal sediments along northern gulf of Mexico. Mar. Environ. Res. 119, 166–175. doi: 10.1016/j.marenvres.2016.05.011
Cahill A. E., Pearman J. K., Borja A., Carugati L., Carvalho S., Danovaro R., et al. (2018). A comparative analysis of metabarcoding and morphology-based identification of benthic communities across different regional seas. Ecol. Evol. 8, 8908–8920. doi: 10.1002/ece3.4283
Campagne C. S., Salles J. M., Boissery P., Deter J. (2015). The seagrass Posidonia oceanica: Ecosystem services identification and economic evaluation of goods and benefits. Mar. Poll. Bull. 97, 391–400. doi: 10.1016/j.marpolbul.2015.05.061
Casola E., Scardi M., Mazzella L., Fresi E. (1987). Structure of the epiphytic community of Posidonia oceanica leaves in a shallow meadow. Mar. Ecol. 8, 285–296. doi: 10.1111/j.1439-0485.1987.tb00189.x
Como S., Magni P., Baroli M., Casu D., De Falco G., Floris A. (2008). Comparative analysis of macrofaunal species richness and composition in Posidonia oceanica, Cymodocea nodosa and leaf litter beds. Mar. Biol. 153, 1087–1101. doi: 10.1007/s00227-007-0881-z
Cowart D. A., Pinheiro M., Mouchel O., Maguer M., Grall J., Miné J., et al. (2015). Metabarcoding is powerful yet still blind: A comparative analysis of morphological and molecular surveys of seagrass communities. PloS One 10, e0117562. doi: 10.1371/journal.pone.0117562
De Cáceres M., Legendre P. (2009). Associations between species and groups of sites: indices and statistical inference. Ecology 90, 3566–3574. doi: 10.1890/08-1823.1
De Jode A., David R., Dubar. J., Rostan J., Guillemain D., Sartoretto S., et al. (2019). Community ecology of coralligenous assemblages using a metabarcoding approach. 3ème Symposium Méditerranéen sur la Conserv. du Coralligène at autres Bio-Concrétions, 41–45.
Domaizon I., Windegardner A., Capo E., Gauthier J., Gregory-Evaes I. (2017). DNA-Based methods in paleolimnology: New opportunities for investigating long-term dynamics of lacustrine biodiversity. J. Paleolimnol 58 (1), 21. doi: 10.1007/s10933-017-9958-y
Duarte C. M. (1991). Allometric scaling of seagrass form and productivity. Mar. Ecol. Prog. Ser. 77, 289–300. doi: 10.3354/meps077289
Duarte S., Leite B. R., Feio M. J., Costa F. O., Filipe A. F. (2021). Integration of DNA-based approaches in aquatic ecological assessment using benthic macroinvertebrates. Water 13, 331. doi: 10.3390/w13030331
Duarte S., Vieira P. E., Leite B. R., Teixeira M. A. L., Neto J. M., Costa F. O. (2023). Macrozoobenthos monitoring in Portuguese transitional waters in the scope of the water framework directive using morphology and DNA metabarcoding. Estuar. Coast. Shelf Sci. 281, 108207. doi: 10.1016/j.ecss.2022.108207
Dufrêne M., Legendre P. (1997). Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecol. Monogr. 67, 345–366. doi: 10.2307/2963459
Edgeloe J. M., Severn-Ellis A. A., Bayer P. E., Mehravi S., Breed M. F., Krauss S. L., et al. (2022). Extensive polyploid clonality was a successful strategy for seagrass to expand into a newly submerged environment. Proc. R. Soc B, 20220538. doi: 10.1098/rspb.2022.0538
Engel M. S., Ceríaco L. M. P., Daniel G. M., Dellapé P. M., Lobl I., Marinov M., et al. (2021). The taxonomic impediment: A shortage of taxonomists, not the lack of technical approaches. Zool. J. Linn. Soc 193, 381–387. doi: 10.1093/zoolinnean/zlab072
Ficetola G. F., Taberlet P. (2023). Towards exhaustive community ecology via DNA metabarcoding. Mol. Ecol. doi: 10.1111/mec.16881
Fonseca V. G., Carvalho G. R., Nichols B., Quince C., Johnson H. F., Neill S. P., et al. (2014). Metagenetic analysis of patterns of distribution and diversity of marine meiobenthic eukaryotes. Global Ecol. Biogeogr. 23, 1293–1302. doi: 10.1111/geb.12223
Frøslev T. G., Kjøller R., Bruun H. H., Ejrnæs R., Brunbjerg A. K., Pietroni C., et al. (2017). Algorithm for post-clustering curation of DNA amplicon data yields reliable biodiversity estimates. Nat. Commun. 8, 1188. doi: 10.1038/s41467-017-01312-x
Gambi M. C., Trani B., Buia M. C. (2005). Taxonomic diversity and distribution of polychaete and isopod borers on the sheaths of the seagrass posidonia oceanica: analysis at regional scale along the coast off Sardinia (Italy). Ital. J. Zool. 72, 141–151. doi: 10.1080/11250000509356664
García-Gómez G., García-Herrero A., Sánchez N., Pardos F., Izquierdo-Muñoz A., Fontaneto D., et al. (2022). Meiofauna is an important, yet often overlooked, component of biodiversity in the ecosystem formed by Posidonia oceanica. Invert. Biol. 141, e12377. doi: 10.1111/ivb.12377
Garrabou J., Gómez-Gras D., Medrano A., Ponti M., Schlegel R., Bensoussan N., et al. (2022). Marine heatwaves drive recurrent mass mortalities in the Mediterranean Sea. Glob. Change Biol. 28, 5708–5725. doi: 10.1111/gcb.16301
Geller J., Meyer C., Parker M., Hawk A. (2013). Redesign of PCR primers for mitochondrial cytochrome c oxidase subunit I for marine invertebrates and application in all-taxa biotic surveys. Mol. Ecol. Res. 13, 851–861. doi: 10.1111/1755-0998.12138
Guardiola M., Wangensteen O. S., Taberlet P., Coissac E., Uriz M. J., Turon X. (2016). Spatio-temporal monitoring of deep-sea communities using metabarcoding of sediment DNA and RNA. PeerJ 4, e2807. doi: 10.7717/peerj.2807
He X., Stanley R. R. E., Rubidge E. M., Jeffery N. W., Hamilton L. C., Westfall K. M., et al. (2022). Fish community surveys in eelgrass beds using both eDNA metabarcoding and seining: Implications for biodiversity monitoring in the coastal zone. Can. J. Fish. Aquat. Sci. 79, 1335–1346. doi: 10.1139/cjfas-2021-021
Jacobs-Palmer E., Gallego R., Ramon-Laca A., Kunselman E., Cribari. K., Horwith M., et al. (2020). A halo of reduced dinoflagellate abundances in and around eelgrass beds. PeerJ 8, e8869. doi: 10.7717/peerj.8869
Jordà G., Marbà N., Duarte C. M. (2012). Mediterranean Seagrass vulnerable to regional climate warming. Nat. Clim. Change 2, 821–824. doi: 10.1038/nclimate1533
Kalogirou S., Corsini-Foka M., Sioulas A., Wennhage H., Pihl L. (2010). Diversity, structure and function of fish assemblages associated with posidonia oceanica beds in an area of the eastern Mediterranean Sea and the role of non-indigenous species. J. Fish Biol. 77, 2338–2357. doi: 10.1111/j.1095-8649.2010.02817.x
Keck F., Blackman R. C., Bossart R., Brantschen J., Couton M., Hürlemann S., et al. (2022). Meta-analysis shows both congruence and complementarity of DNA and eDNA metabarcoding to traditional methods for biological community assessment. Mol. Ecol. 31, 1820–1835. doi: 10.1111/mec.16364
Kelly R. P., Closek C. J., O’Donnell J. L., Kralj J. E., Shelton A. O., Samhouri J. F. (2017). Genetic and manual survey methods yield different and complementary views of an ecosystem. Front. Mar. Sci. 3. doi: 10.3389/fmars.2016.00283
Lejeusne C., Chevaldonné P., Pegent-Martini C., Boudouresque C. F., Pérez T. (2010). Climate change effects on a miniature ocean: the highly diverse, highly impacted Mediterranean Sea. Trends Ecol. Evol. 25, 250–260. doi: 10.1016/j.tree.2009.10.009
Leray M., Yang J. Y., Meyer C. P., Mills S. C., Agudelo N., Ranwez V., et al. (2013). A new versatile primer set targeting a short fragment of the mitochondrial COI region for metabarcoding metazoan diversity: Application for characterizing coral reef fish gut contents. Front. Zool. 10, 34. doi: 10.1186/1742-9994-10-34
Macher J. N., Bloska D. M., Holzmann M., Girard E. B., Pawlowski J., Renema W. (2022). Mitochondrial cytochrome c oxidase subunit I (COI) metabarcoding of foraminifera communities using taxon-specific primers. Peer J. 10, e13952. doi: 10.7717/peerj.13952
Mahé F., Rognes T., Quince C., de Vargas C., Dunthorn M. (2015). Swarm v2: highly-scalable and high-resolution amplicon clustering. PeerJ 3, e1420. doi: 10.7717/peerj.1420
Marbà N., Díaz-Almela E., Duarte C. M. (2014). Mediterranean Seagrass (Posidonia oceanica) loss between 1842 and 2009. Biol. Conserv. 176, 183–190. doi: 10.1016/j.biocon.2014.05.024
Marbà N., Duarte C. M. (2010). Mediterranean Warming triggers seagrass (Posidonia oceanica) shoot mortality. Glob. Change Biol. 16, 2366–2375. doi: 10.1111/j.1365-2486.2009.02130.x
Marbà N., Duarte C. M., Holmer M., Martínez R., Basterretxea G., Orfila A., et al. (2002). Effectiveness of protection of seagrass (Posidonia oceanica) populations in cabrera national park (Spain). Environ. Conserv. 29, 509–518. doi: 10.1017/S037689290200036X
Mariani S., Baillie C., Colosimo G., Riesgo A. (2019). Sponges as natural environmental DNA samplers. Curr. Biol. 29, R395–R402. doi: 10.1016/j.cub.2019.04.031
Martínez-Crego B., Prado P., Alcoverro T., Romero J. (2010). Composition of epiphytic leaf community of Posidonia oceanica as a tool for environmental biomonitoring.” estuar. Coast. Shelf Sci. 88, 199–208. doi: 10.1016/j.ecss.2010.03.026
Menning D. M., Gravley H. A., Cady M. N., Pepin D., Wyllie-Echeverria S., Ward D. H., et al. (2021). Metabarcoding of environmental samples suggest wide distribution of eelgrass (Zostera marina) pathogens in the north pacific. Metabarcoding Metagenom. 5, 35–42. doi: 10.3897/mbmg.5.62823
Momota K., Hosokawa S., Komuro T. (2022). Small-scale heterogeneity of fish diversity evaluated by environmental DNA analysis in eelgrass beds. Mar. Ecol. Prog. Ser. 688, 99–112. doi: 10.3354/meps13994
Obst M., Exter K., Allcock A. L., Arvanitidis C., Axberg A., Bustamante M., et al. (2020). A marine biodiversity observatory network for genetic monitoring of hard-bottom communities (ARMS-MBON). Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.572680
Oksanen J., Blanchet F. G., Friendly M., Kindt R., Legendre P., McGlinn D., et al. (2019). Vegan: Community ecology package (R package version 2.5-7). Available at: https://CRAN.R-project.org/package=vegan.
Padiglia A., Cadeddu B., Demurtas D., Bertolino M., Manconi R., Pronzato R. (2015). Sponges of Posidonia oceanica meadows (Sardinia, W-Mediterranean Sea). PeerJ PrePrints 3, e1044v1. doi: 10.7287/peerj.preprints.1044v1
Pawlowski J., Bonin A., Boyer F., Cordier T., Taberlet P. (2021). Environmental DNA for biomonitoring. Mol. Ecol. 30, 2931–2936. doi: 10.1111/mec.16023
Piazzi L., Balata D., Ceccherelli J. (2016). Epiphyte assemblages of the Mediterranean seagrass Posidonia oceanica: A review. Mar. Ecol. 37, 3–41. doi: 10.1111/maec.12331
Piñeiro-Juncal N., Kaal J., Moreira J. C. F., Martínez Cortizas A., Lambais M. R., Otero X. L., et al. (2021). Cover loss in a seagrass Posidonia oceanica meadow accelerates soil organic matter turnover and alters soil prokaryotic communities. Org. Geochem. 151, 104140. doi: 10.1016/j.orggeochem.2020.104140
Reef R., Atwood T. B., Samper-Villarreal J., Adame M. F., Sampayo E. M., Lovelock C. E. (2017). Using eDNA to determine the source of organic carbon in seagrass meadows. Limnol. Oceanogr. 62, 1254–1265. doi: 10.1002/lno.10499
Rodríguez-Ezpeleta N., Morisette O., Bean C. W., Manu S., Banerjee P., Lacoursière-Rousel A., et al. (2021). Trade-offs between reducing complex terminology and producing accurate interpretations from environmental DNA: Comment on “Environmental DNA: What’s behind the term?” Mol. Ecol. 30, 4601–4605. doi: 10.1111/mec.15942. by Pawlowski et al.
Rognes T., Flouri T., Nichols B., Quince C., Mahé F. (2016). VSEARCH: a versatile open source tool for metagenomics. PeerJ 4, e2548. doi: 10.7717/peerj.2584
Ruggiero M. V., Turk R., Procaccini G. (2002). Genetic identity and homozygosity in north-Adriatic populations of Posidonia oceanica: An ancient, post-glacial clone? Conserv. Genet. 3, 71–74.
Shum P., Barney B. T., O’Leary J. K., Palumbi S. R. (2019). Cobble community DNA as a tool to monitor patterns of biodiversity within kelp forest ecosystems. Mol. Ecol. Res. 19, 1470–1485. doi: 10.1111/1755-0998.13067
Steyaert M., Priestley V., Osborne O., Herraiz A., Arnold R., Savolainen O. (2020). Advances in metabarcoding techniques bring us closer to reliable monitoring of the marine benthos. J. App. Ecol. 57, 2234–2245. doi: 10.1111/1365-2664.13729
Traganos D., Lee C. B., Blume A., Poursanidis D., Cizmek H., Deter J., et al. (2022). Spatially explicit seagrass extent mapping across the entire Mediterranean. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.871799
Turon M., Angulo-Preckler C., Antich A., Praebel K., Wangensteen O. S. (2020). More than expected from old sponge samples: A natural sampler DNA metabarcoding assessment of marine fish diversity in nha trang bay (Vietnam). Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.605148
Wangensteen O. S., Palacín C., Guardiola M., Turon X. (2018). DNA Metabarcoding of littoral hard-bottom communities: High diversity and database gaps revealed by two molecular markers. PeerJ 6, e4705. doi: 10.7717/peerj.4705
Wangensteen O. S., Turon X. (2017). “Metabarcoding techniques for assessing biodiversity of marine animal forests,” in Marine animal forests Rossi. Eds. Bramanti L. S., Gori A., Orejas C. (Berlin, Germany: Springer International Publishing), 445–473. doi: 10.1007/978-3-319-21012-4_53
Wesselmann M., Geraldi N. R., Marbà N., Hendricks I. E., Díaz-Rúa R., Duarte C. M. (2022). eDNA reveals the associated metazoan diversity of Mediterranean seagrass sediments. Diversity 14, 549. doi: 10.3390/d14070549
Keywords: benthos, seagrass, metabarcoding, Posidonia oceanica, cytochrome oxidase I
Citation: Turon X, Zarcero J, Antich A, Wangensteen OS, Ballesteros E, Cebrian E, Marco-Méndez C and Alcoverro T (2023) Metabarcoding the eukaryotic community of a threatened, iconic Mediterranean habitat: Posidonia oceanica seagrass meadows. Front. Mar. Sci. 10:1145883. doi: 10.3389/fmars.2023.1145883
Received: 16 January 2023; Accepted: 22 March 2023;
Published: 06 April 2023.
Edited by:
Ana Riesgo, Museo Nacional de Ciencias Naturales CSIC, SpainReviewed by:
Cole G. Easson, Middle Tennessee State University, United StatesPere Ferriol, University of the Balearic Islands, Spain
Copyright © 2023 Turon, Zarcero, Antich, Wangensteen, Ballesteros, Cebrian, Marco-Méndez and Alcoverro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xavier Turon, eHR1cm9uQGNlYWIuY3NpYy5lcw==