 Rotem Zirler1,2
Rotem Zirler1,2 Lynn Angele Leck1,2
Lynn Angele Leck1,2 Tamar Feldstein Farkash1,2Martina Holzknecht3
Tamar Feldstein Farkash1,2Martina Holzknecht3 Andreas Kroh3
Andreas Kroh3 Vasilis Gerovasileiou4,5Mehmet Fatih Huseyinoglu6,7
Vasilis Gerovasileiou4,5Mehmet Fatih Huseyinoglu6,7 Carlos Jimenez8,9
Carlos Jimenez8,9 Vasilis Resaikos8
Vasilis Resaikos8 Mehmet Baki Yokeş10
Mehmet Baki Yokeş10 Omri Bronstein1,2*
Omri Bronstein1,2*- 1School of Zoology, Faculty of Life Sciences, Tel Aviv University, Tel Aviv, Israel
- 2The Steinhardt Museum of Natural History, Tel Aviv University, Tel Aviv, Israel
- 3Natural History Museum Vienna (NHMW), Vienna, Austria
- 4Department of Environment, Faculty of Environment, Ionian University, Zakinthos, Greece
- 5Hellenic Centre for Marine Research (HCMR), Institute of Marine Biology, Biotechnology and Aquaculture (IMBBC), Thalassocosmos, Heraklion, Crete, Greece
- 6Faculty of Maritime Studies, University of Kyrenia, Kyrenia, Cyprus
- 7Biosphere Research Center, Istanbul, Türkiye
- 8Enalia Physis Environmental Research Centre, Nicosia, Cyprus
- 9Energy, Environment and Water Research Center (EEWRC), The Cyprus Institute, Nicosia, Cyprus
- 10AMBRD Laboratories, Istanbul, Türkiye
The Eastern Mediterranean Sea is an exceptional habitat. Its relative isolation and distinct characteristics create a unique ecosystem recognized as a marine biodiversity hot spot, where one-fifth of the species are endemic. Yet, native Mediterranean biodiversity is under increasing threat, mainly due to massive alien species invasions of Indo-Pacific origin. To date, more than 800 non-indigenous species have been reported in the Eastern Mediterranean Sea, justifying its reputation as one of the most severely affected habitats in the world in terms of marine biological invasions. Here we summarized the Mediterranean invasion dynamics of the long-spined echinoid Diadema setosum (Leske, 1778), one of the most ubiquitous Indo-Pacific sea urchin species. We show an alarming exponential population growth of D. setosum throughout the Eastern Mediterranean since 2018, following more than a decade of ‘invasion lag’ since its first detection in 2006. Molecular analyses illustrate the presence of a single genetic D. setosum clade in the Mediterranean Sea – corresponding to the Arabian Peninsula clade of this species, reinforcing the notion of a Red Sea origin. Our data support the current working hypothesis that the initial introduction of D. setosum occurred in the Northern Levantine Basin from which it gradually expanded in both north-west and south-east trajectories – in contrast to a stepping-stone hypothesis of gradual advancement from the opening of the Suez Canal. Demographic data of D. setosum along the Israeli Mediterranean coastline reveals a well-established population of broad size distributions, from juveniles to adult individuals of remarkably large size. Additionally, we provide evidence of the reproductive capacity of D. setosum in its new environment. Due to the magnitude of Diadema’s ecological footprint, it poses a severe threat to the entire Eastern Mediterranean Sea, including the Levantine Basin and South Aegean Sea, calling for rapid and coordinated action at both national and regional scales.
1 Introduction
The stability of ecosystems depends on their delicate food webs and interactions among the fauna and flora of which they are comprised (Pimm, 1984; McCann, 2000; Hooper et al., 2012; Loreau and de Mazancourt, 2013). The frailty of such interactions is revealed through dramatic changes in the local communities following deviation from equilibrium (Downing et al., 2012; Hooper et al., 2012). One of the many impacts of enhanced anthropogenic activity is the intentional and coincidental transfer of non-indigenous species (NIS) outside of their native distribution range, which under certain circumstances may escalate to species invasion (Galil, 2007). Consequently, invaded regions and their native fauna may be severely affected (Vredenburg, 2004).
A typical invasion largely follows a sequential chain of events (Figure 1) starting with (I) the dispersal of NIS beyond their native range. This facilitates an initial introduction (II) which is normally followed by a (III) lag-phase that may vary in length, during which the new population is maintained at low densities in a restricted area, and often remains undetected. Lag-phase duration may last for decades before NIS become detectible (Crooks, 2005; Azzurro et al., 2016; Zenetos et al., 2019). During establishment (IV), NIS population-growth accelerates, their range expands, and they increasingly interact with the new environment. During this phase, some NIS may turn invasive as they reach exceptionally high abundances and trigger undesired ecological and economic impacts (Mack et al., 2000). Under certain circumstances, NIS populations may grow exponentially and achieve exceptionally high abundances – further expanding their range and changing the ecological equilibriums through competitive exclusion of native species and exhaustion of natural resources. This final stage is largely recognized as population outbreak (V) (Byers et al., 2002; Blackburn et al., 2015).
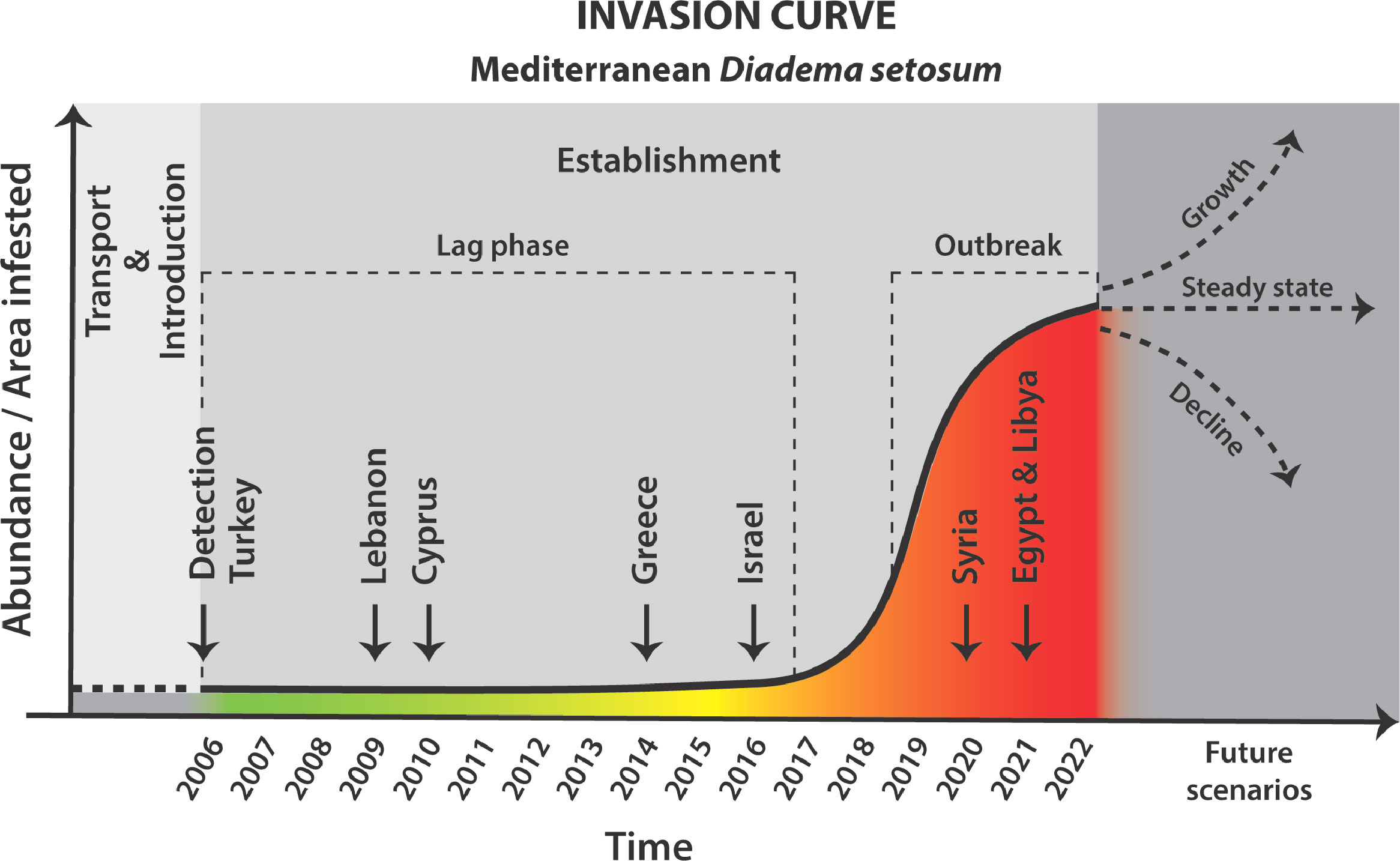
Figure 1 Schematic invasion curve specifying Diadema setosum invasion timeline in the Mediterranean Sea. Following transport and introduction to the new region, founder non-indigenous species (NIS) populations remain restricted in both population size and range. The invasion lag-phase occurs at the onset of NIS establishment and is characterized by restricted spatial distributions and low abundances, which are often undetected. As establishment progresses, population size dramatically increases and includes individuals at varying life stages. Ultimately, population outbreaks may occur as NIS achieve exponential population-growth and range expansion. Vertical dow-facing arrows indicate the year of first D. setosum record for the respective countries.
The Eastern Mediterranean Sea is known as one of the most invaded marine regions in the world (Katsanevakis et al., 2014b; Tsiamis et al., 2020). Though the native biodiversity of the Mediterranean Sea is characterized by a remarkable rate of endemism (accounting for 20% of its species) and was therefore referred as marine biodiversity hot spot (Coll et al., 2010; Gianni et al., 2013; Katsanevakis et al., 2014a), the unique fauna of the Mediterranean Sea is constantly threatened by a massive influx of alien species of Red Sea (RS) origin (Galil, 2007; Coll et al., 2010; Galil et al., 2017). The maritime connection between these two adjacent seas was formed following the opening of the Suez Canal in 1869 (Coll et al., 2010; Soukissian et al., 2017). This manmade route allows marine species to travel from the RS to the Mediterranean (Galil and Goren, 2014) – a process termed Lessepsian Migration (Por, 1971). Originally, the route through the canal was hindered by a series of physical and biological barriers (such as the hyper-saline barrier formed by the Bitter Lakes) that limited, at least temporarily, the intensity of species migration between the RS and Mediterranean (Aron and Smith, 1971; El-Serehy et al., 2014; El-Serehy et al., 2018). However, most of these natural barriers eroded over time, allowing direct (i.e., gamete propagation and active locomotion) and indirect (i.e., human mediated transport) transmission (Carlton, 1987). Consequently, the capacity for successful invasions increased dramatically with time as reflected by the mounting number of reports of new NIS of RS origin in the Eastern Mediterranean (Galil and Goren, 2014; Galil et al., 2015) currently including approximately 800 species (Rotter et al., 2020).
Recently, an ongoing migration is drawing increasing attention as new evidence show alarming rates of population growth of the long-spined sea urchin, Diadema setosum (Leske, 1778) in the Mediterranean Sea. Diadema setosum is one of the most conspicuous Indo-Pacific shallow water echinoids (Bronstein and Loya, 2014; Muthiga and McClanahan, 2020). The two recognized genetic clades of the species are widely distributed throughout the Indo-Pacific, ranging from the RS and Persian Gulf (clade b), through the east coast of Africa in the Indian Ocean, to the west Pacific, off the coast of Japan (clade a) (Lessios et al., 2001; Bronstein et al., 2016; Muthiga and McClanahan, 2020). While diadematoids are largely recognized as omnivores, they mainly feed by grazing on algae scraped from hard substrates (Bronstein and Loya, 2014; Muthiga and McClanahan, 2020). Consequently, Diadema spp., like other members of this family, are potent ‘environmental engineers’, capable of altering the structure and composition of entire benthic communities (Hernández et al., 2008; Ling et al., 2009; Bronstein and Loya, 2014; Goh and Lim, 2015). As algal growth regulators, Diadema grazing restricts algae proliferation, thereby supporting the settlement and development of slower growing benthic organisms – such as corals (Lawrence, 1975; Sammarco, 1982; Bronstein and Loya, 2014; do Hung Dang et al., 2020). Conversely, exceptionally high grazing intensities driven by large Diadema populations, may drive degradation of benthic communities (such as algae, macrophytes, and hard benthic infrastructure; Carreiro-Silva and Mcclanahan, 2001), potentially leading to collapse of entire habitats (Lawrence, 1975; Mokady et al., 1996; Hernández et al., 2008; Qiu et al., 2014).
The capacity of any given species to proliferate and reach high abundances, largely depend on their ability to reproduce. For invasive species in particular, one of the key challenges in concurring new environments, is the ability to reach sexual maturity and reproduce successfully, despite of potentially varying environmental conditions with respect to their native range. While reproduction was extensively studied across the genus Diadema (see Muthiga and McClanahan, 2020 and references therein), and several studies targeted RS populations of D. setosum (Pearse, 1970; Bronstein et al., 2016), none has so far targeted populations from the Eastern Levantine Basin.
The first record of D. setosum in the Mediterranean Sea in 2006, off the Kaş Peninsula, Turkey, (Yokes and Galil, 2006) was a milestone in the successful establishment of large aggregations currently occupying the entire Levantine Basin (Figure 2).
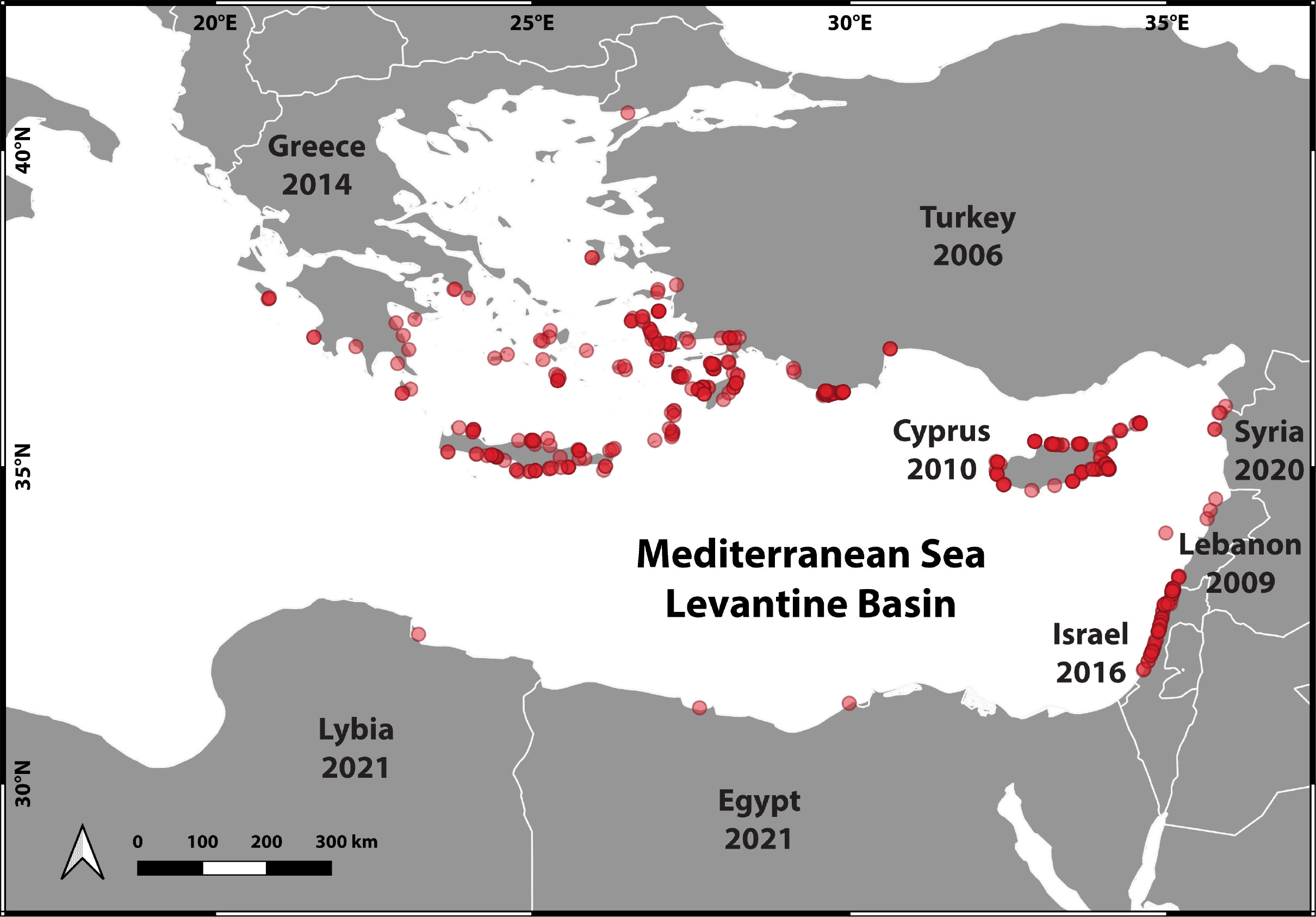
Figure 2 Map of the Eastern Mediterranean Sea. Red dots (transparent red symbols) represent sites of Diadema setosum observations compiled over nearly two decades – from the detection of the first individual in 2006 to the end of 2022. The map was created using QGIS v. 3.28.2 (QGIS Development Team, 2023).
Here we follow the invasion dynamics of D. setosum in the Mediterranean Sea, demonstrating its range expansion and recent accelerated population growth reflecting clear signs of population outbreak. We combine a thorough literature review with extensive survey data and sampling complemented by citizen-science reports, to provide a comprehensive report on the progress of this invasion and depict its true scale. We use molecular data to identify the genetic makeup of the invaders and determine their origin. We show an alarming recent exponential increase in D. setosum abundance in the Eastern Mediterranean and provide evidence of effective reproductive capacity (i.e., the ability of an individual to reproduce) in their new environment. Our results shed new light on both temporal and spatial dynamics of biological invasions in the Mediterranean Sea and facilitate better understanding of the life history, invasion dynamics, genetic makeup, and reproductive biology of the ubiquitous D. setosum.
2 Materials and methods
2.1 Field observations and sample collection
Underwater surveys were conducted between 2016 and 2022 along the Greek, Turkish, Cypriot, and Israeli Mediterranean coastline, and published data were obtained from the literature and complimented by citizen reports (Table S1). Data from a total of 670 reports comprising: 313 reports from scientific literature, 237 underwater survey dives by trained biologists, and 120 citizen science reports, covering a depth range of 0-50 m and spanning over 2,000 km of the Eastern Mediterranean Sea, were compiled. Each identified specimen was noted, indicating the date of observation, depth, substrate type, and precise location.
Data were used to construct two datasets: (1) ‘number of individuals’ – the total number of Diadema setosum individuals encountered on a 45 min dive, and (2) ‘number of observations’ – each independent dive where Diadema setosum are observed was counted as one, regardless of the number of individuals encountered. At all dives, counts were conducted by a single observer, regardless of the number of participants, and dives were regarded as independent reports if conducted at different sites or on different dates.
Considering the potential biases originating from citizen science, strict criteria for evaluating these data were established. Reports missing the exact number of individuals encountered, their depths, and/or precise localities (coordinates) were omitted from downstream analyses. In reports where the number of individuals was ambiguous, values were determined following personal communication with the reporter or else omitted. When depth was given as range, mean values were calculated, and when metadata were missing altogether, the observations were omitted from the analysis. In total, 48 reports were omitted based on these criteria. To avoid instances of misidentification of D. setosum by untrained reporters, such data were included only if accompanied by photographic evidence, allowing unambiguous identification prior to inclusion in our dataset.
When possible, specimens were collected for morphological examinations as well as molecular and histological analyses. In total, 24 D. setosum specimens were collected along the Israeli Mediterranean (specimens are deposited at the Steinhardt Museum of Natural History collection, Tel Aviv University, Israel) (Table S2). Twelve additional tissue samples of adult specimens were collected in Crete, Greece, and five tissue samples were collected from the native Red Sea population (Eilat, Israel), and processed as described below.
2.2 Size distribution
Test diameters (measured at the ambitus) and heights of the 24 D. setosum specimens collected from the Israeli Mediterranean were measured using a Mitutoyo 500-196-30 digital caliper to the nearest 0.01 mm (Table S2). A second distribution representing the native Israeli Red Sea population was obtained from Bronstein’s unpublished data collected between April 2007 and August 2008 in Eilat, Israel, including the sizes of 547 randomly selected D. setosum specimens (Table S3). Both Israeli distributions were then compared with the size distribution of the Greek population, which was modified from Vafidis et al. (2021). The latter providing test diameters of 160 randomly selected individuals, from the vicinity of Dodecanese Islands, between December 2019 and July 2020.
2.3 Molecular analysis
Total genomic DNA was extracted from spine-muscles or gonads using the DNeasy Blood and Tissue Kit (QIAGEN, Hilden, Germany) following the manufacturer’s instructions. PCR amplifications of two mitochondrial and one nuclear markers were performed. Fragments of the mitochondrial (1) cytochrome c oxidase subunit 1 gene (COI) and (2) Lysine-tRNA, ATPase-6 and ATPase-8 region (LYS), as well as the first exon region of the nuclear (3) bindin gene (BIN) were amplified using the Hy-Taq Ready Mix (2x) (Hylabs, Rehovot, Israel). Reaction conditions for the COI fragment using the primers CO1f and CO1a (Lessios et al., 2001) were: 3 min at 95°C followed by 35 cycles of 30 sec at 95°C, 30 sec at 58°C and 1 min at 72°C, ending with a final extension step of 10 min at 72°C (Table 1). Reaction conditions for the LYS fragment using primers LYSa and ATP6b (Lessios et al., 2001) were 3 min at 94°C followed by 35 cycles of 30 sec at 94°C, 30 sec at 55°C and 1 min at 72°C, ending with a final extension step of 10 min at 72°C (Table 1). Reaction conditions for the bindin fragment using the primers DA5A and DAIR (Geyer et al., 2020) were 3 min at 94°C followed by 40 cycles of 30 sec at 94°C, 30 sec at 51°C and 1 min at 72°C, ending with a final extension step of 10 min at 72°C (Table 1). PCR products were visualized on a 1% agarose gel, purified using ExoSAP-IT (Affymetrix) and sequenced in both directions at the TAU sequencing facility. All sequences generated in the present study were deposited in GenBank under accession numbers MT430942-MT430943, MT434142-MT434143, MW387536, MW394192, ON197106-ON197133, ON210773-ON210801 and ON211043-ON211059 (Table S4).
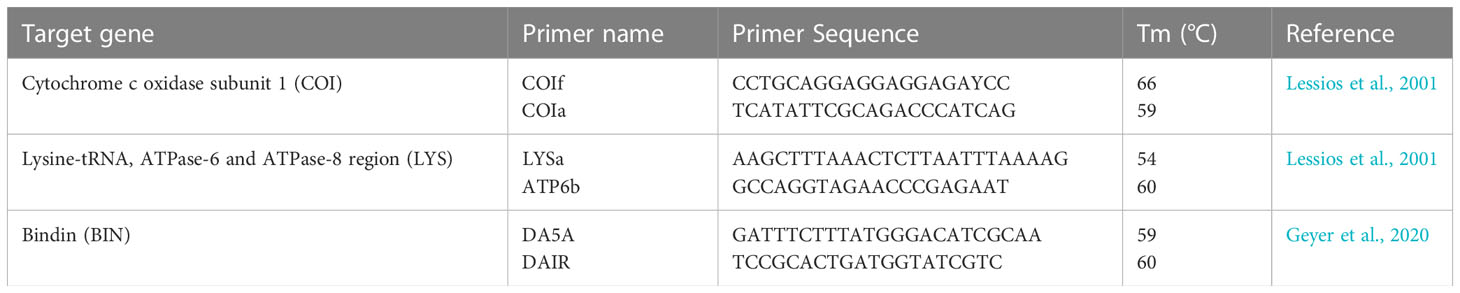
Table 1 List of primers used for the molecular analysis.
Forward and reverse sequences of each locus were assembled, inspected, and edited using SeqTrace (Stucky, 2012). Consensus sequences were edited using AliView v.1.18 (Larsson, 2014) and aligned using MAFFT v.7 alignment server (http://mafft.cbrc.jp/alignment/server/), employing the E-INS-i algorithm. Ambiguous positions were removed using TrimAl v.1.4 (Capella-Gutiérrez et al., 2009) and GUIDANCE2 (Sela et al., 2015), followed by a final manual inspection.
Three datasets were created to facilitate further analyses: (1) COI – comprising 108 COI sequences, 640 bp long, including all publicly available sequences from the family Diadematidae (31 sequences generated in the current study); (2) LYS – comprising 172 Lysin-ATP6 Diadema spp. sequences, 580 bp long, representing all extant species in the genus Diadema (20 sequences generated in the current study); (3) BIN – comprising 157 sequences of the nuclear bindin gene from all extant species of Diadema, 500 bp long (29 sequences generated in the current study; Table S4).
Phylogenetic analyses were conducted using both Maximum Likelihood (ML) and Bayesian Inference (BI) approaches following Bronstein et al. (2017) and Bronstein and Kroh (2018). Briefly, a heuristic search under the Bayesian Information Criterion (BIC) (Schwarz, 1978), as implemented in PartitionFinder2 (Lanfear et al., 2017) was employed to determine the optimal partitioning schemes and models of molecular evolution. ML analyses were performed in parallel with IQtree (Trifinopoulos et al., 2016) and raxmlGUI 2.0 (Edler et al., 2021). IQtree analyses uses ModelFinder (Kalyaanamoorthy et al., 2017) to select the best fit models for each partition, identifying the TN+F+G4 as the best-fit model across all partitions for both COI and LYS datasets, while K2P+G4 was better suited for the BIN datasets. Branch support was evaluated using the ultrafast bootstrap on IQtree (UFBoot, 1000 replicates) as well as standard bootstrap (BS, 1000 replicates). Additional ML analysis were performed with raxmlGUI 2.0 applying the settings ‘ML + thorough bootstrap’, 100 runs, 1000 replicates, using TrN+G4 (for COI and LYS datasets) and K80+G4 (for BIN dataset) as the best-fit models for all partitions as inferred from PartitionFinder2. Bayesian analysis was carried out using MrBayes v. 3.2.2 (Ronquist et al., 2012). We ran two independent runs of three ‘heated’ and one ‘cold’ chain for 10 million generations, sampling parameters and trees every 100th generations. The runs were inspected with Tracer 1.7.1 (Rambaut et al., 2018) to assess convergence. In a conservative approach, the first 25% of trees were discarded as burn-in, and a 50% majority-rule consensus tree was calculated from the remaining trees. Posterior Probabilities (PP) were obtained from the 50% majority-rule consensus of the trees sampled during the stationary phase.
2.4 Reproductive biology
To evaluate D. setosum’s reproductive capacity in the Mediterranean, 12 individuals were sampled from Plakias (35°09’16.9992”N 24°26’30.0012”E), in southern Crete (Greece), during September 2020, from depths of 10-13 m. Export of material was carried out under a Material Transfer Agreement between HCMR and NHMW dated September 9th 2020. Seven additional samples were collected on different occasions from the Israeli population between December 2019 and September 2021. To facilitate a comparison between the Mediterranean population and the native Red Sea population, five additional D. setosum specimens were sampled from Eilat, Israel (29°30’6.966”N 34°55’3.3924”E) during September 2020, from depth of 5-10 m. Given the rapid decline of Red Sea D. setosum abundance over the past decade (Eviatar and Bronstein, in prep), strict regulations on destructive sampling have been imposed throughout the region, limiting sample availability. Data on localities, sampling dates, moon-phase, sex, and size of individuals used for the reproductive state evaluation are provided in Table S5.
A single gonad from each specimen was extracted, fixed in Bouin’s solution, and prepared for histological analysis following the procedures described in Bronstein and Loya (2015) and Bronstein et al. (2016). Briefly, following fixation, tissue was embedded in paraffin, sectioned to 7 µm using a Shandon M1R microtome, and stained using a standard Hematoxylin and Eosin protocol. Stained slides were examined under a Nikon Eclipse Ni-U light microscope to determine their reproductive stage. We followed the four-stage system of Bronstein et al. (2016) to describe D. setosum’s reproductive cycle: Stage I (spent): Gonads are largely devoid of contents showing ova-free lumen in females and spermatozoan-free lumen in males. A thin layer of nutritive phagocytes (NPs) is present along the ascinal walls in both sexes and may form a pale meshwork across the ascinus. Strongly basophilic previtellogenetic oocytes or primary spermatocytes, staining dark purple with Hematoxylin and eosin, are present along the ascinal wall. Stage II (recovering): NPs proliferate from the gonad ascinal wall, gradually filling the lumen of both ovaries and testis. Limited groups of primary spermatocytes and clusters of previtellogenetic oocytes start appearing in the testicular and ovarian germinal epithelia, respectively, and may occasionally project centrally. Stage III (growing): Both early and late vitellogenetic oocytes may be present along the ovarian wall. All stages of germ cells are evident in the male germinal epithelium and continuously increase in number as new spermatogonia develop basally while spermatocytes migrate to the testicular lumen, where they accumulate as mature spermatozoa, forming visible columns of darkly stained cells. NPs deplete and progressively occupy less space in both males and females. Stage IV (mature): By the end of this stage the NP layer in both ovaries and testes is largely exhausted. Ovaries are packed with mature ova, while oocytes at different maturation stages may still be evident in the germinal epithelium. The testicular lumen is densely packed with spermatozoa. Occasionally some ova and spermatozoa may be evident in the coelom.
2.5 Statistical analyses
Statistical analyses were performed using R (RStudio Team, 2020). As data representing the relationship between years and the number of observations or the number of individuals were counts, and the variance of the count data was greater than the mean, we used a generalized linear model (GLM) with a Poisson distribution and a log-link function (to account for overdispersion) using the stats package (R Core Team, 2022). ‘Year’ was determined as the independent variable, and the number of observations/individuals was determined as the response variable. An a+2 offset was applied to all count data. Pairwise comparisons were conducted between each possible combination of years, for both the number of observations and number of individuals, using the Tukey method (implemented in the R package multcomp; Hothorn et al., 2008). p-values were corrected for multiple comparisons using the Bonferroni correction. Pairwise Kolmogorov-Smirnov tests were performed to check for differences in size frequency distributions between RS, Israeli Mediterranean, and Greek (Crete) Mediterranean populations using the R package dgof (Arnold and Emerson, 2011). Due to multiple testing for the size frequency comparisons, p-value was corrected using the Bonferroni correction. To test for the differences in male-female ratios, Chi-square tests were performed.
3 Results
3.1 Demography
Reports on the presence of D. setosum in the Mediterranean Sea increase with time since the original record in 2006 (Figure 3). By the end of 2017, the number of new observations remained consistently low, comprising no more than 13 annual observations, and an average of 0-2 individuals per report, to a total of 52 observations since the first report in 2006 (Table S1). Between 2018 and 2020 a significant increase in number of observations was noted (Generalized linear model, df=1, p> 0.001; Table S6), summing up to a total of 268 reports (Table S1). Between 2020 through the end of 2022, a significant exponential increase in the number of observations occurred (Generalized linear model, df=1, p<0.001; Table S6 ;Figure 3). In 2018, when the significant increase was first recorded, the sum of documented reports reached 74 (44 of which from the vicinity of Kaş), and by 2021, a total of 205 observations were made throughout the Levantine Basin (Figure 3; Table S1). A non-significant decrease in number of observations was noted in 2022, totaling 145 annual observations (46 of which in Kaş). Prior to 2018, Cyprus held the highest number of annual observations, however, since 2018, the majority of observations shifted to Greece (Figure 3). Overall, Greece holds the highest proportion of observations, accounting for 32% of the total observations (Figure 3).
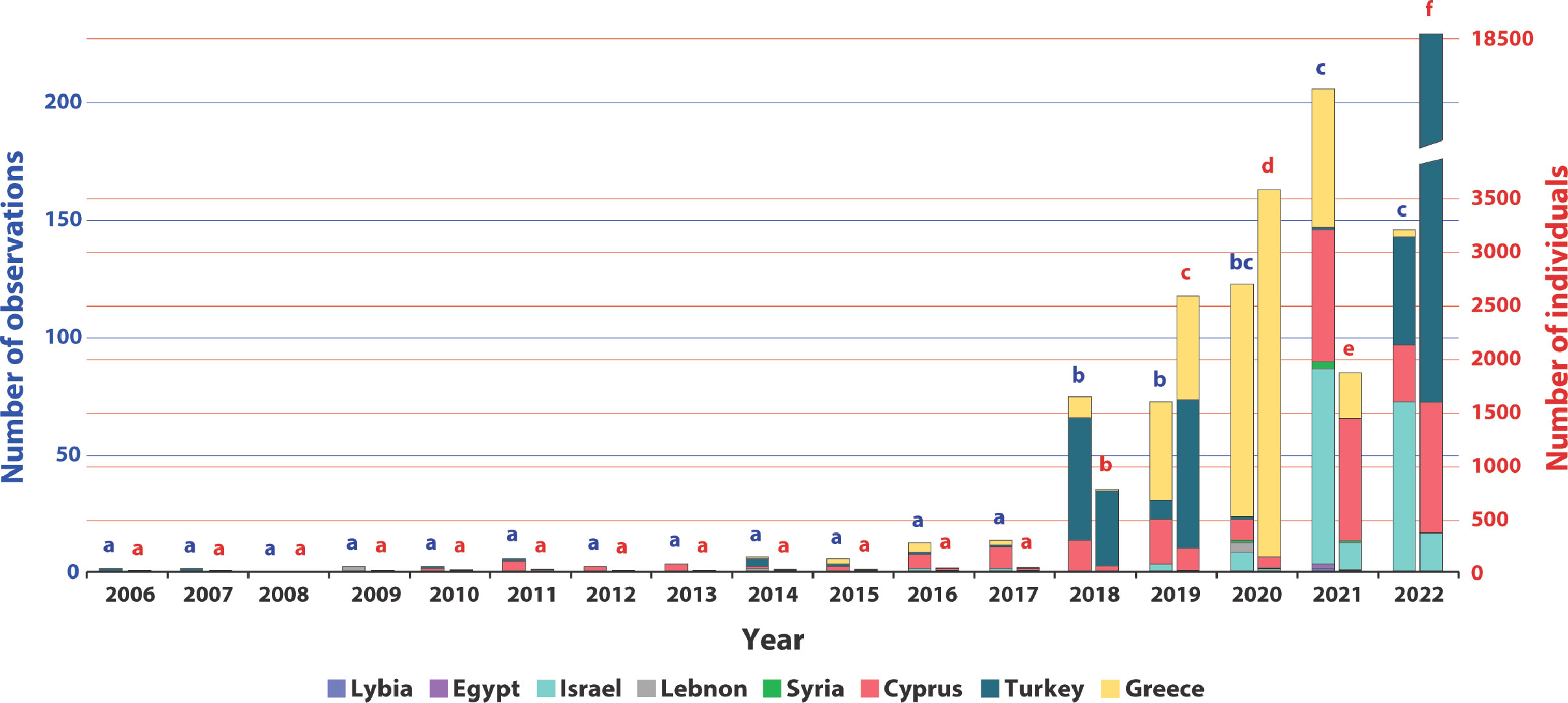
Figure 3 Annual Diadema setosum number of observations and number of individuals in the Mediterranean Sea between 2006 and 2022. Color indicates the country from which the report originates. Column pairs (per year) represent number of observations (left) and number of individuals (right), and correspond to the blue and red scales, respectively.
When considering the annual number of individuals (Figure 3), a similar trend emerges. Between 2006 to 2014, most observations were of single individual, totaling 36 reported individuals during this period. From 2015 to 2017, there was a slight (yet non-significant) increase in the number of reported individuals, with the total number almost doubling within only three years (Figure 3). A significant shift in trend occurred in 2018, during which a total of 757 individuals were observed (Generalized linear model, df=1, p<0.001; Table S6; Figure 3), 428 of which from the vicinity of Kaş. In 2019, the number of individuals further increased (Generalized linear model, df=1, p<0.001; Table S6), resulting in a threefold increase in comparison to 2018 (n=2,563; Figure 3). A further significant increase in the number of individuals occurred in 2020 (Generalized linear model, df=1, p<0.001; Table S6), totaling 3,552 documented individuals, reflecting an exponential growth in D. setosum Mediterranean populations (Figure 3). Despite an increase in the number of reports between 2020 and 2021 (n=122 and n=205, respectively), 2021 was marked by a decrease in the number of observed individuals (n=1847; Figure 3). Still, by the end of 2022, the number of D. setosum individuals reached a record of 18,512 individuals (Generalized linear model, df=1, p<0.001; Table S6). Overall, since the onset of invasion in 2006, the majority of individuals were spotted around the coasts of Turkey (n=19,034) and Greece (n=4,853).
Due to potential biases rising from unequal sampling efforts, a dataset restricted to reports from Kaş, Turkey, was constructed, applying the same analyses for number of observations and number of individuals, as conducted for the total count data. This dataset was further restricted to data obtained through routine scientific surveys. These reports contribute the majority of individuals (63.5%) reported in this study. Furthermore, these reports were collected under a controlled sampling design of routine annual surveys (reporting presence and absences) and facilitated the widest temporal comparison (with reports dating back to the first observation in 2006). Results from these analyses were identical to the results obtained for the pooled data (Generalized linear model, df=1, p<0.001) for both number of observations and number of individuals; Table S7 and S8, validating reported trends.
3.2 Population structure
3.2.1 Size distribution
Body sizes of Diadema setosum varied significantly within and between Mediterranean and RS localities (Figure 4), with all populations compared containing both juvenile and adult individuals. Size-frequency distribution (SFD) of the Israeli population showed specimens with test diameters ranging from a minimum of 11.8 mm to a maximal test diameter of 97.03 mm (Table S2). The majority of specimens (54%) were small-sized (test diameters of 0–40 mm), with medium-sized (40–70 mm) and large-sized (70–100 mm) individuals comprising 25% and 21% of the population, respectively. Similar SFDs were measured in Greece (modified from Vafidis et al., 2021; based on test diameter measurements of n=160 individuals sampled off the Dodecanese Islands) (Kolmogorov-Smirnov; p=0.0964). Similar to the Israeli Mediterranean population, very small (10–20 mm) and very large (80–90 mm) individuals were the least frequent (1% and 4%, respectively). In contrast, SFD of the Israeli population seems to be more biased towards the smaller size groups (10–20 and 20–30 mm) than the Greek population. Nevertheless, both Greek and Israeli populations showed a high proportion of juveniles compared to adults. SFD of D. setosum from its native RS range differed significantly from both Israeli (Kolmogorov-Smirnov; p=0.0211) and Greek (Kolmogorov-Smirnov; p<0.0001) Mediterranean populations. Specimens in the RS had distinctly smaller body size, ranging from 1 to 65 mm. Uniquely, the RS was the only locality where individuals smaller than 10 mm (7.5%) could be observed. The RS population was strongly skewed towards smaller individuals with most individuals ranging between 10 to 30 mm in diameter. The occurrence of individuals larger than 50 mm in the RS was negligible (1.3%).
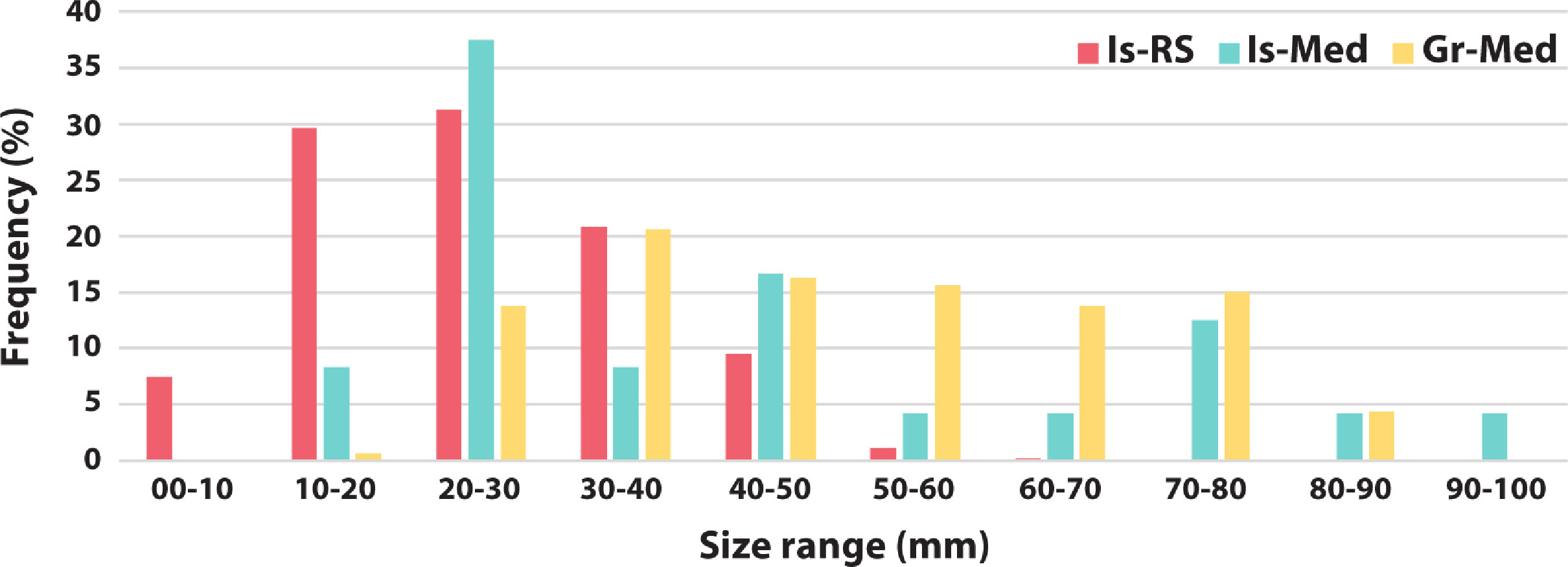
Figure 4 Size-frequency distributions of Diadema setosum at three localities: a native population at the Gulf of Aqaba, Red Sea (Is-RS; n=547), measured between 2007 and 2008; Israeli Mediterranean population (Is-Med; n=24), measured between 2019 and 2021; and Greek population (Gre-Med; n=160), measured between 2019 and 2020 (modified from Vafidis et al., 2021).
3.3 Reproductive biology
Similar male-female ratios were measured in both Crete (Cre-Med; 7 males, 5 females; Chi-square test, X2 = 0.333, df=1, p-value=0.56) and Israeli Mediterranean coastline (Is-Med; four males, three females; Chi-square test, X2 = 0.1428, df=1, p-value=0.7054). The reproductive state of the Cretan specimens (collected in mid-September; Figures 5A–F) appeared to be highly synchronous – showing growing gametes (stage 3), with some individuals having nearly mature oocytes (Figures 5E, F). This sexual synchronization was evenly distributed between males and females (Table S54). All but one of the Israeli Mediterranean specimens (collected during May, and from September to December) were at the recovery stage (stage 2; Figures 5C, D). The only exception was specimen DS21, a male at the growing stage (stage 3), collected in August 2021.
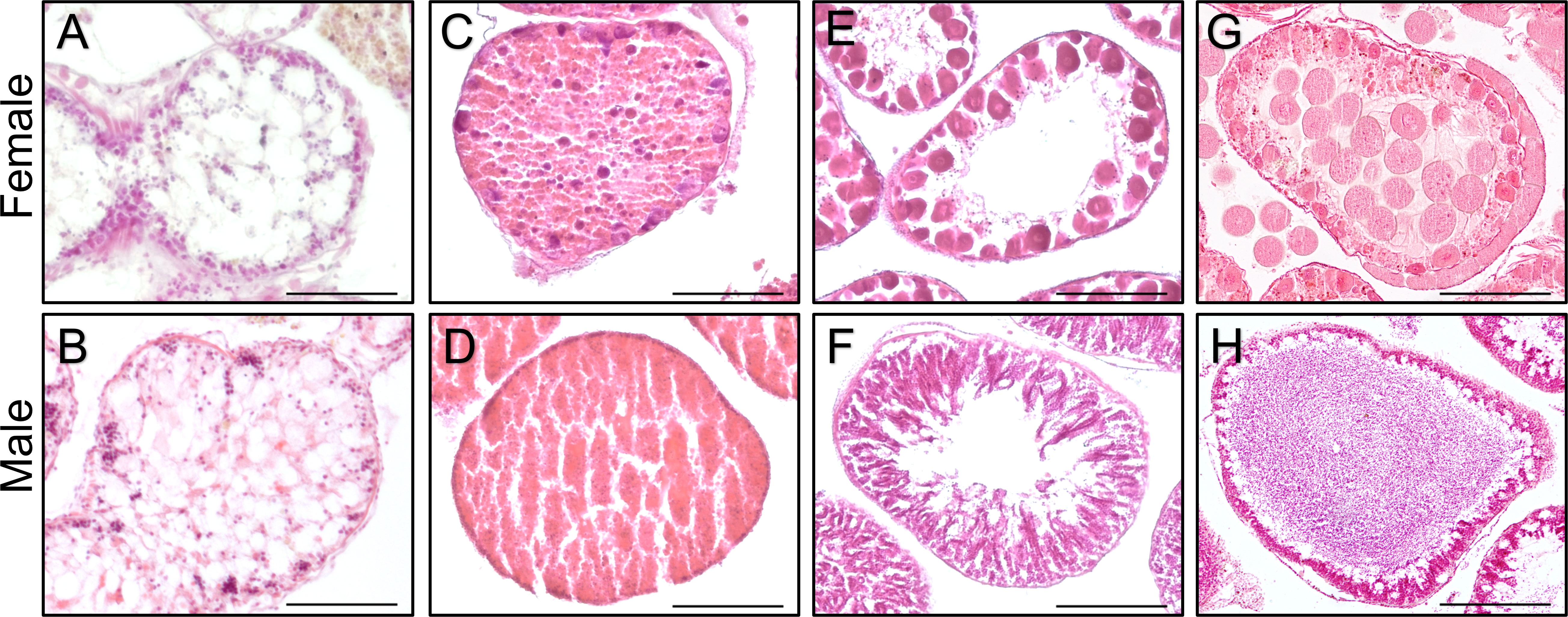
Figure 5 Histological cross-sections of Diadema setosum gonads, representing the four reproductive stages used for classification (following Bronstein et al., 2016). Ovaries (A, C, E, G), and testes (B, D, F, H) are presented. Stage 1 (spent; A, B): the post-spawned gonads are devoid of content, showing mostly free lumen in both sexes. Stage 2 (recovering; C, D): nutritive phagocytes (NPs) are proliferating, filling the acinal lumen. Primary oocytes (C) and spermatocytes (D) might be visible on the germinal epithelia. Stage 3 (growing; E, F) the maturating oocytes (E) migrate toward the lumen and increase in size as vitellogenesis (the process during which the ova store nutrients to support the future larvae) advances. Similarly, in males, spermatocytes migrate towards the lumen as they mature (F), while new spermatogonia keep forming along the germinal epithelium. As gametes proliferate and grow, NPs are continuously depleted and become less abundant. Stage 4 (mature; G, H): the gonads of both sexes are packed with mature oocytes (G) and spermatozoa (H), while NPs are largely exhausted. Photographs (B, G, H) were modified from Bronstein et al. (2016) to illustrate the complete reproductive cycle of D. setosum. Scale bars represent 100 μm.
3.4 Molecular analysis
Sequences of the mitochondrial COI gene of all Diadema species as well as other members of the family Diadematidae were obtained from public databases and included in the current analyses (Figure 6). Complementary to the COI dataset, the mitochondrial LYS gene dataset holds a higher number of Diadema spp. sequences available for comparison with the Mediterranean sequences, although it lacks the representation of D. clarki and other diadematoids (Figure S1). The bindin gene dataset facilitates the phylogenetic reconstruction of all extant Diadema species based on a nuclear gene, thus complementing the two abovementioned mitochondrial genes (Figure S2). As phylogenetic analyses generated congruent topologies, only ML trees are presented – showing both bootstrap support values and posterior probabilities for the respective nodes.

Figure 6 Phylogenetic relationships of the family Diadematidae based on the COI dataset. Maximum likelihood topology is displayed (major clades collapsed), representing all known species of Diadema as well as all publicly available sequences of additional members of the family (Centrostephanus longispinus, Astropyga pulvinate, Astropyga radiata, Echinothrix calamaris, and Echinothrix diadema). All Mediterranean Diadema sequences as well as RS sequences generated in the current study were clustered with D. setosum clade b by both analyses (ML and BI). Bootstrap support values (<65%) and posterior probabilities (<0.65) are shown above nodes, before and after the slash, respectively. Specimens corresponding to GenBank accession numbers LC037355/56/57 (marked with asterisk) were referred to in the literature as Diadema sp. and deposited under D. setosum, however, based on the evidence provided in Chow et al. (2016), these specimens clearly belong to D. clarki. Details on the sequences used to generate the tree are given in Table S4. Scale bar reflects number of changes per site.
All analyses, across methodology and loci, ascribe the Israeli specimens to clade b of D. setosum (Figures 6, S1 and S2). In agreement with previously published results (Lessios et al., 2001; Bronstein et al., 2017; Geyer et al., 2020), D. setosum clade b was resolved as sister to D. setosum clade a in all datasets. However, while both COI and LYS support the early divergence of the former two clades from the other Diadema species, the BIN dataset suggests that the divergence of D. clarki and D. palmeri predated that split. In agreement with the results of Lessios et al. (2001) and Bronstein et al. (2017), both COI and LYS analyses support similar topologies with extant Diadema species retrieved as monophyletic and highly supported. Within the BIN dataset, only three of the eight recognized species of Diadema were resolved as monophyletic clades (D. palmeri, D. clarki and both clades of D. setosum), while the other species remained unresolved (Figure S2).
4 Discussion
4.1 Molecular diagnosis
Two former studies on Mediterranean D. setosum included genetic data in their reports (Bronstein et al., 2017; Bronstein and Kroh, 2018). Still, these studies only utilized two samples – the original specimen collected near Kaş (Turkey) in 2006, and the first confirmed specimen from Israel (2017). The additional data provided in the current study (24 new specimens), as well as the inclusion of a nuclear marker, confirm the identity of Mediterranean Diadema spp. as D. setosum clade b, the clade native to the RS and Persian Gulf. Owing to the geographical proximity of the RS and Mediterranean and given the favorable conditions for clade b in the Eastern Mediterranean (Bronstein et al., 2017), migration of the latter seemed inevitable. Current data supports the notion of a single genetic clade of Diadema (clade b) in the Mediterranean Sea. The Indo-Pacific clade a seems to be absent in the Mediterranean at this time – in agreement with the habitat suitability model of Bronstein et al. (2017). Whether the present Mediterranean population represent the descendants of a single invasion event prior to 2006, or the consequent of multiple, periodic invasions, remains to be answered. Molecular population genetics may help resolve this question once sufficient samples are obtained from across the Eastern Mediterranean.
4.2 Invasion dynamics
Given the conspicuous appearance and non-cryptic nature of D. setosum, early detection of this species in the Mediterranean was possible. Moreover, the hard substrate preference, and depth range preference of Diadema, reaching the shallow depths of the lower intertidal, makes it readily detectible at highly monitored sites along the Mediterranean coastline. Consequently, the first report of this species in 2006 (Yokes and Galil, 2006), likely occurred shortly following the species arrival to the Mediterranean, offering a rare opportunity to study the dynamics of marine invasions, from arrival to population outbreak, in a natural setup.
Since the first detection of D. setosum off Kaş peninsula, Turkey (Yokes and Galil, 2006), evidence for the successful establishment of the species have been accumulating. What was originally anecdotical observations of isolated individuals or small aggregations (Gökoğlu et al., 2007; Crocetta et al., 2015; Gökoğlu et al., 2016; Gerovasileiou et al., 2017; Bronstein and Kroh, 2018; Galanos and Kritikos, 2019; Katsanevakis et al., 2014a; Nader and Indary, 2011; Turan et al., 2011; Tsiamis et al., 2015; Mytilineou et al., 2016; Yapıcı, 2018), has grown to aggregations of dozens (Bilecenoğlu et al., 2019; Katsanevakis et al., 2020; Ammar, 2021) and hundreds (Gül and Aydin, 2021; Vafidis et al., 2021) of individuals.
The exponential increase in observations and number of individuals since 2018, reflect a rapid spread and range expansion of D. setosum throughout the Eastern Mediterranean and Southeastern Aegean Sea (Figure 3). During this time, large aggregations of individuals became common, mostly around Turkey, Greece, and Cyprus, marking the shift from establishment to outbreak (Figures 1, 3). The pattern of prolonged periods of low densities upon NIS arrival followed by an exponential increase in abundance, is typical of many biological invasions (Figure 1) (e.g., Aikio et al., 2010; Blackburn et al., 2015; Azzurro et al., 2016). During the initial ‘invasion lag’ the new NIS population is small and represented by only few individuals from the founder population. Routine, intensive marine surveys conducted annually since 2015 by Israel Nature and Parks Authority (INPA) along the Israeli Mediterranean coastline, did not report the presence of D. setosum between 2015 and 2017 (Lazarus et al., 2020). Therefore, the exponential increase in number of observations in the following years, under identical sampling effort, can only be attributed to the rapid proliferation of D. setosum populations. Nevertheless, coordinated, regional scale monitoring is needed to depict the fine scale dynamics of this invasion.
The primary requirement of all living organisms is the ability to feed and experience environmental conditions within the species tolerance range. In this respect, the Eastern Mediterranean, characterized by high water temperatures and solar radiation (in comparison to the western basin) causing high rates of evaporation (Bethoux, 1979; Coll et al., 2010; Schroeder et al., 2016; Soukissian et al., 2017), has recently been shown to be highly suitable for clade b of D. setosum (Bronstein et al., 2017). In turn, these higher temperatures and increased evaporation, drive elevated salinity, forming a longitudinal salinity gradient being highest in the southeast and gradually dissipating to the north and west (Coll et al., 2010; Borghini et al., 2014; Soukissian et al., 2017). Subsequently, the lower salinity near Gibraltar (36.2 psu; Soukissian et al., 2017) resembles the salinity of the northeastern Atlantic, whereas the higher salinity of the southeastern Levant (38.6 psu; Soukissian et al., 2017), resembles the salinity of the northern RS (40.7 psu; Biton and Gildor, 2011). Moreover, the completion of the Aswan dam in 1965, restricted the seasonal flooding of the Nile delta, playing a major role in the overall reduction of Levantine productivity. As the Nile discharge drives large amounts of mud and silt, the dam formation severely limited nutrient drift into the Eastern Mediterranean, decreasing regional productivity by ten folds (Azov, 1991). Consequently, a gradient in habitat suitability for D. setosum now occurs in this region, being most suitable in the Southeastern Levantine Basin, and decreasing to the north and west (Bronstein et al., 2017). As elevated temperatures have been shown to favor both settlement and post-settlement survival of Diadema spp. (Hernández et al., 2010), the significantly higher abundances of D. setosum under the less favorable conditions of the northern Levant (in comparison to the Southeastern Levant), support the assumption of gradual establishment starting in the north and gradually expanding southwards.
The absence of native shallow water (<40m) diadematoids in the Mediterranean suggests little direct niche competition with potentially similar species. The only native Mediterranean diadematoid, Centrostephanus longispinus, is more common at depths of 40 m to over 200 m (Pawson and Miller, 1983; Koukouras et al., 2007; Katsanevakis et al., 2017), with the highest densities recorded at depths of 60–130 m (Furió and Templado, 2012). Still, bathymetric isolation between the native C. longispinus and invasive D. setosum necessitate further investigation, as adult D. setosum specimens were recorded down to depths of 49 m (Katsanevakis et al., 2020) and even 55 m on coralligenous patches off Cyprus (Katsanevakis et al., 2020). At the shallow end, a decade long decline of native shallow-water echinoids in the Eastern Mediterranean, namely Paracentrotus lividus and Arbacia lixula, reached the point of near local extinction at some localities (Yeruham et al., 2015; Rilov, 2016). Under these circumstances, the addition of a strong echinoid competitor to the largely vacant niche is likely to aid the expansion of D. setosum, and further restrict the recovery of native species.
Following successful establishment, the founders of NIS must be capable of reproducing in the new environment in order to maintain sustainable populations over time. Our data based on histological analyses of Mediterranean D. setosum gonads, show evidence for reproductive capacity in populations comprising sexually adult males and females at both Israeli and Greek populations (Figures 5G, H; Table S5), similar to the evidence recently provided from the Dodecanese Island complex (Vafidis et al., 2021). Furthermore, the presence of a wide size distribution among Mediterranean D. setosum, despite being skewed towards juveniles, clearly show individuals of different ages, providing further support for the capacity of D. setosum to successfully reproduce in its new habitat, as reoccurring recruitment from sources outside the Mediterranean at these scales is highly unlikely. As the success of reproduction through broadcast spawning is highly dependent on abundance of spawning individuals (Figures 5G, H; Table S5), growing populations gradually increase the chances of successful fertilizations, leading to the formation of larger larval stocks. Consequently, a higher number of juveniles will potentially be recruited, accelerating the process with time, as these juveniles grow to sexual maturity.
Current evidence suggest population outbreak of D. setosum in the Eastern Mediterranean, being most evident around Turkey and Greece, and likely spreading south along the Eastern coast of the Levant towards Lebanon and Israel (Figure 3). Yet, the true scale and rate of progression from the assumed point of origin in Turkey, is hindered by uneven sampling efforts, reflected, for example, by a single published observation from Lebanon in 2009 (Nader and Indary, 2011). Biased sampling effort is also evident in the drop in reported numbers of individuals between 2020 and 2021 (Figure 3). To the south, D. setosum seems to have recently (2021) expanded to Egypt and Libya (Nour et al., 2022). Interestingly, despite their proximity to the Mediterranean opening of the Suez Canal, Libya and Egypt are the latest countries to report the presence of D. setosum in the Mediterranean. West of the Suez Canal, NIS have to disperse upstream in the prevailing currents, inevitably slowing down their westbound progression (Koukouras et al., 2010). These reports provide further support to the prevailing notion of an initial introduction in Turkey (likely facilitated by man-mediated transport) followed by gradual counter current (Pascual et al., 2017) spread to the south (Bronstein et al., 2017). Considering D. setosum’s delayed southern expansion, further establishment, population growth and potentially outbreak in southeastern Mediterranean countries seems highly probable. To the west, D. setosum may be reaching the edge of its biological range (following the habitat suitability model of Bronstein et al., 2017), potentially providing an explanation to the lack of observation of this species west of Zakynthos, Greece, Ionian Sea (Dimitriadis et al., 2023), despite the close geographic proximity and favoring surface currents from the assumed point of introduction in Kaş.
Maritime traffic plays a major role in transporting NIS to new territories by ballast water transport or as fouling communities on ship hulls (Williams et al., 1988; Lavoie et al., 1999; Costello et al., 2022). Numerous organisms and larvae migrate within ship ballast water, consequently being introduced to new, sometimes distant habitats. The Suez Canal, one of the world’s busiest marine trade routes (Figure 7), provides easy access for alien Red Sea fauna entering the Mediterranean Sea. D. setosum initial detection in the northeastern Levant, 600 km away from Port Said (Egypt), at the Mediterranean side of the Suez Canal, implies that introduction was mediated by ballast water transport, as previously suggested by Yokes and Galil (2006) and Bronstein et al. (2017). Of the three main westbound shipping routes off Port Said, the northernmost one travels to the Aegean Sea, passing tightly south of Rhodes, a short distance from the site of first detection off Kaş (Figure 7). According to this scenario, from this initial point of introduction, the species gradually extended its range, first to the west – being aided by regional currents, and later, at a slower pace, to the south, working its way against the prevailing counter-clockwise surface currents in the Levant Basin (Millot and Taupier-Letage, 2005). The elaborated network of regional shipping routes in the Eastern Mediterranean likely contributed to D. setosum’s local expansion.
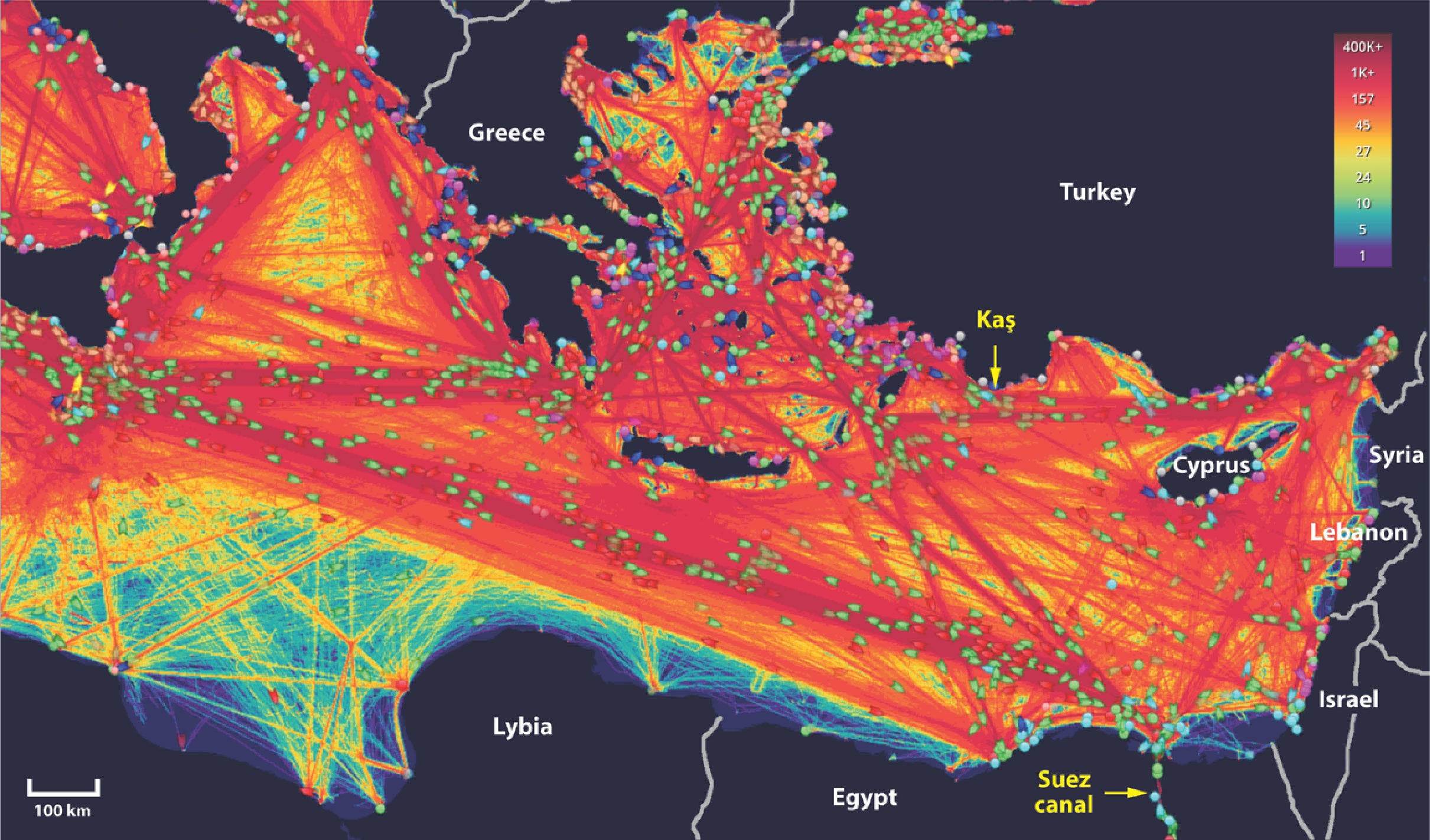
Figure 7 Map of main shipping routes in the Eastern Mediterranean Sea during 2021. Data obtained from https://www.marinetraffic.com/. Color gradient scale represents – routes/2.45km2/year.
4.3 Insights on invasion dynamics from NIS population structure
4.3.1 Size
As demonstrated by the Israeli Mediterranean population, size distribution is multimodal and spans from juveniles to markedly large individuals (Figure 4; Table S2), characteristic of a population with ongoing recruitment and supply of settling larvae (Pecorino et al., 2012). While size-age correlations are highly debated in echinoderm research (Bluhm et al., 1998), it may very well support a qualitative estimation of age. Hence, specimens at a size range of 10–20 mm (e.g., DS4, DS8, and DS18; Table S2) can be estimated as being several months old (Lewis, 1966), while individuals with test diameters of 80–100 mm (such as DS3, DS5, and DS6; Table S2) as being at least several years old. Estimates of the Israeli SFD are based on a limited dataset and clearly more data is needed to capture the true size structure of this population. Nonetheless, the presence of individuals across the species size range, suggests that D. setosum are capable of maintaining stable, sustainable populations in the Mediterranean, demonstrating ongoing recruitment and survival of offspring – most likely originating from sexually reproducing local populations.
Ebert (1982) determined the maximum diameter of D. setosum from Eilat (clade b) to be 83.57 mm – 1.6 times larger than the Indo-pacific clade a (Ebert, 1982; Muthiga and McClanahan, 2020). As such, some of the unusually large Mediterranean specimens (Figure 4, Table S2), reaching an unprecedented large size of 97.03 mm (DS3) may very well be a decade (or more) old. The slow initial growth rate of D. setosum relative to other Diadema species such as D. savignyi (Muthiga and McClanahan, 2020), makes the presence of such large specimens even more surprising. It reflects the high suitability of the new habitat for D. setosum (Bronstein et al., 2017), which encourages body size enhancement. Furthermore, the presence of such large, potentially decade old individuals, suggest that the true date of D. setosum’s arrival to Israel, predates (potentially by several years) current confirmed reports (Bronstein and Kroh, 2018). Similarly, the first specimen collected in Kaş in 2006 was 58 mm, and therefore already several years old. Though the introduction of adults cannot be ruled out at this stage, ballast water transport of larvae is still the most likely means of transport. Under such circumstances, the presence of such large specimens indicates a minimum two-decade long presence of D. setosum in the Mediterranean Sea.
Further insights might be gained by comparing size frequency distributions between the native population in the RS, the more established Mediterranean population in the Aegean Sea and the proliferating southern Levant population in Israel. Indeed, the Greek population displayed size distributions similar to the Israeli Mediterranean population, showing elevated frequencies of young individuals in the size range of 10 to 30 mm (Figure 4). The abundance of juveniles (compared to adults) at both Greek and Israeli populations, suggest high larval supply for these populations followed by high mortality rates post recruitment. While the drivers of these selective adult mortalities are currently unknown, elevated predation of the less cryptic adults may provide some explanations.
In contrast to the Mediterranean, the native RS population is comprised of significantly smaller individuals (Figure 4). Moreover, new recruits at a size range of 0–10 mm were only identified in the RS, where the preferred recruitment sites, and early-life growth niches of D. setosum, are within crevices and under peddles, rocks and coral debris at shallow depths down to 1 m (Bronstein pers. obs.). While the early recruits’ size-class is inevitably present also in the Mediterranean, its location is currently unknown. The bell-shaped distribution of RS D. setosum suggests strong selective pressure against both very small and very large individuals. Small individuals are more susceptible for predation (McClanahan and Kurtis, 1991; Clemente et al., 2007), whereas large individuals are likely to accumulate fatal illnesses that eventually lead to mortality (Jones, 1985; Buchwald et al., 2015). Thus, the enhanced body size in Mediterranean populations may indicate the current lack of such selective forces in the Levant. The lower abundance of potential Diadema spp. predators (see below), species-specific illnesses, currently present in the new habitat, is probably beneficial for D. setosum populations, supporting enhanced body size and longevity, which in turn increases the species reproductive potential and accelerate the species proliferation.
4.3.2 Depth range
In its native RS range, D. setosum largely occupy shallow subtidal depths down to 20 m, although most populations inhabit much shallower depth of 0 to 10 m (Muthiga and McClanahan, 2020). The bulk of Levantine D. setosum observations were reported from depths of 0–10 m (Table S1), in agreement with previously determined depth range for native D. setosum (Muthiga and McClanahan, 2020). Yet, about 11.5% of the Mediterranean reports seem to exceed this range, occurring at depths of 20–30 m, occasionally in clusters of dozens, and 3.5% of reports come from depths between 35 and 55 m. Several ecological factors may drive this habitat preference pattern. The presence (or absence) of predators may provide a convincing explanation to this deeper-water shift. For instance, the fish species Diplodus sargus and Balistes capriscus, which are known as Diadema spp. predators at depths of 5–20 m in the Canary Islands, are also present the Mediterranean Sea (Clemente et al., 2010; Clemente et al., 2011; Kacem and Neifar, 2014; Exadactylos et al., 2019; Muthiga and McClanahan, 2020). The invasive fish Lagocephalus sceleratus and Torquigener hypselogeneion were also reported to feed on Diadema spp. in the Southeastern Aegean Sea (Ulman et al., 2021; Jimenez pers. Obs.; Huseyinoglu, pers. Obs.), and while the depth range of this species is now documented down to 220 m, it is more abundant at shallow depths of up to 10 m. The presence of such shallow water predators in the Mediterranean may drive D. setosum out of its preferred depth range to waters deeper than 20 m to avoid predation. The capacity of Diadema spp. to occupy greater depths in the Mediterranean may also explain, at least in part, some of the lag phase of the invasion. Dives deeper than 20 m are scarce along the Israeli Mediterranean coastline. In addition, D. setosum has been also recorded in cryptic environments such as the shadowy entrance of marine caves in Greece, Cyprus and Turkey (Digenis et al., 2022; Gerovasileiou et al., 2022; Ragkousis et al., in press). Given the chance that the first individuals spent their initial establishment phase at limited-accessed locations and depths, they might have simply been overlooked. Upon establishment, they proliferated, gradually occupying shallower depths, and became detectible. While the factor driving this ecological release is currently unknown, the hypothesis of detection-avoidance-by-depth gains further support by the most recent report of D. setosum in Libya, reporting the presence of two individuals at a depth of 25 m (Nour et al., 2022).
4.4 Reproductive biology
The reproductive cycle of native Diadema setosum in the RS was intensively studied by Bronstein et al. (2016), illustrating high proportions of sexually mature individuals between July and October (Figure 8). As such, sampling in the current study was conducted during September – at the peak of the expected annual reproductive cycle (Pearse, 1970; Bronstein et al., 2016). In Crete, 92% of the gonads sampled were at the advanced growing stage. The almost uniform synchroneity of the Mediterranean population (for both sexes), are congruent with the findings of Bronstein et al. (2016) and support the ability of D. setosum to reproduce in its new Mediterranean habitat (Figure 8). Like the Crete-Mediterranean population, the RS population seemed synchronized, however, the reproductive cycle at these two locations, varied markedly during fall of 2021. In contrast to the sampling of the Greek population, samples collected along the Israeli Mediterranean were compiled over several months. These specimens demonstrate the presence of both sexes, at the recovering stage (Bronstein et al., 2016), asserting the reproductive capacity of the local population. Further confirmation for D. setosum’s successful reproductive capacity along the Israeli Mediterranean comes from specimen DS21, demonstrating active gametogenesis during the growing stage (following Bronstein et al., 2016). DS21 was sampled in early August, suggesting that the peak of its reproductive cycle leading to spawning likely occur during September to October – in congruence with the RS cycle (Bronstein et al., 2016).

Figure 8 Reproductive cycles of Diadema setosum. Israeli RS (Is-RS, A), Crete, Greece (Cre-Med, B) and Israeli Mediterranean (Is-Med, C) specimens collected between 2019 to 2021. Colors indicate reproductive stages 1–4 (corresponding to stages: Spent (Blue), Recovering (Green), Growing (Purple) and Mature (Red), respectively). Novel data generated during the current study was compared with the 2010 reproductive cycle of RS D. setosum described in Bronstein et al. (2016) (represented as the opaque background illustrations of each radial plot).
4.5 Ecology
Most recently, reports of D. setosum mortality started to accumulate from near Kaş (Turkey), spreading to adjacent coasts off Turkey and Greece. The intensity of these die-offs suggest that the largest, most established Mediterranean population of the species is undergoing mass mortality since July 2022 (Zirler et al. in press). Based on the observed pathology, the characteristic tissue and spine loss suggest a water-born pathogen as the cause of mortalities (currently under investigation), similar to the driver of mortalities in other Diadema species, such as D. antillarum in the Caribbean (Bak et al., 1984; Hughes et al., 1985; Lessios, 1988) and D. africanum in the Canary Islands (Clemente et al., 2014; Hernández et al., 2020; Sangil and Hernández, 2021). Though the spatial scale of these events is, to the best of our knowledge, still confined to the northern Levant, it seems to be rapidly spreading, imposing an immediate threat to both local fauna as well as native RS D. setosum populations, and call for close monitoring of these events.
5 Conclusions
Our data clearly shows that D. setosum is now well established in the Mediterranean Sea. The exponential growth phase currently reached in the Northern Levant marks the shift of D. setosum proliferation to population outbreak. While the full scope and outcomes of this successful invasion are still unclear, the potential risk for the already disrupted environment of the Eastern Mediterranean, increases dramatically. Habitat degradation, depletion of resources, competitive exclusion of native species, hybridization, and the emerging signs of pathogenic infections, are some of the major concerns associated with the current invasion. To preserve the unique ecosystem of the Eastern Mediterranean, coordinated, regional-scale action must be implemented. Regional collaboration will be effective not only for the current D. setosum invasion, but also aid in mitigation of upcoming invasions by NIS that are already making their way to the Mediterranean Sea.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author contributions
Study conception and design: RZ and OB. Data collection: RZ, LL, MHo, VG, MHu, CJ, VR and MY. Analysis and interpretation of results: RZ, OB, LL, TF and AK. Draft manuscript preparation: RZ and OB. All authors reviewed the results and approved the final version of the manuscript.
Funding
This research was supported by the Israel Science Foundation (ISF; grant number 2407/20) and Yad Hanadiv Foundation (grant number 10699) to OB. Field research in Turkey was partially supported by WWF-Turkey.
Acknowledgments
This work was facilitated by the Mediterranean Diadema Response Network (MDRN) – an international collaboration of scientists and stakeholders from Cyprus, Greece, Turkey, Israel, and the United Arab Emirates, aimed at providing real-time, regional scale monitoring of the invasive D. setosum. Institutional support was provided by the Steinhardt Museum of Natural History, Tel Aviv University, Israel. We thank Dr. Nir Stern of the Israel Oceanographic and Limnological Research for facilitating this international collaboration. We extend our gratitude to Dr. Sigal Shefer, Dr. Shevy Rothman, Dr. Boaz Maizel, Dr. Liron Goren, and Alex Geyzner for their contributions to data collection. Field work was supported by the Dive2gether and Mare Mundi Marine Field Station in Plakias, southern Crete, and Enalia Physis Environmental Research Centre and Deep Dive and ScubaCyprus Dive Centers (CJ, MFH, VR).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2023.1152584/full#supplementary-material
References
Aikio S., Duncan R. P., Hulme P. E. (2010). Lag-phases in alien plant invasions: separating the facts from the artefacts. Oikos 119, 370–378. doi: 10.1111/j.1600-0706.2009.17963.x
Ammar I. A. (2021). First record and massive spread of tropical long-spined Sea urchin Diadema antillarum in the Mediterranean Sea (Syrian coast). J. Earth Environ. Sci. Res. 3 (5), 1–4. doi: 10.47363/JEESR/2021(3)158. * Misidentified as Diadema antillarum.
Arnold T. A., Emerson J. W. (2011). Nonparametric goodness-of-fit tests for discrete null distributions. R J. 3, 34–39. doi: 10.32614/RJ-2011-016
Aron W. I., Smith S. H. (1971). Ship canals and aquatic ecosystems: equilibrium has not been achieved since the Erie, welland, and Suez canals were built. Science 174, 13–20. doi: 10.1126/science.174.4004.13
Azov Y. (1991). Eastern Mediterranean–A marine desert? Mar. pollut. Bull. 23, 225–232. doi: 10.1016/0025-326X(91)90679-M
Azzurro E., Maynou F., Belmaker J., Golani D., Crooks J. A. (2016). Lag times in lessepsian fish invasion. Biol. Invasions 18, 2761–2772. doi: 10.1007/s10530-016-1184-4
Bak R. P. M., Carpay M. J. E., de Ruyter van Steveninck E. D. (1984). Densities of the sea urchin Diadema antillarum before and after mass mortalities on the coral reefs on curacao. Mar. Ecol. Prog. series. Oldendorf 17, 105–108. doi: 10.3354/meps017105
Bethoux J. P. (1979). Budgets of the Mediterranean sea. their dependance on the local climate and on the characteristics of the Atlantic waters. Oceanol Acta 2, 157–163.
Bilecenoğlu M., Baki Yokeş M., Draman M. (2019). The invasive sea urchin Diadema setosum provides shelter for coastal fish–first observations from the Mediterranean Sea. Zool Middle East 65, 183–185. doi: 10.1080/09397140.2.019.1571746
Biton E., Gildor H. (2011). Stepwise seasonal restratification and the evolution of salinity minimum in the gulf of aqaba (Gulf of eilat). J. Geophys Res. 116, 1–7. doi: 10.1029/2010JC00686010.1029/2011JC006944
Blackburn T. M., Lockwood J. L., Cassey P. (2015). The influence of numbers on invasion success. Mol. Ecol. 24, 1942–1953. doi: 10.1111/mec.13075
Bluhm B. A., Piepenburg D., von Juterzenka K. (1998). Distribution, standing stock, growth, mortality and production of Strongylocentrotus pallidus (Echinodermata: echinoidea) in the northern barents Sea. Polar Biol. 20, 325–334. doi: 10.1007/s003000050310
Borghini M., Bryden H., Schroeder K., Sparnocchia S., Vetrano A. (2014). The Mediterranean is becoming saltier. Ocean Sci. 10, 693–700. doi: 10.5194/os-10-693-2014
Bronstein O., Georgopoulou E., Kroh A. (2017). On the distribution of the invasive long-spined echinoid Diadema setosum and its expansion in the Mediterranean Sea. Mar. Ecol. Prog. Ser. 583, 163–178. doi: 10.3354/meps12348
Bronstein O., Kroh A. (2018). Needle in a haystack-genetic evidence confirms the expansion of the alien echinoid Diadema setosum (Echinoidea: diadematidae) to the Mediterranean coast of Israel. Zootaxa 4497, 593–599. doi: 10.11646/zootaxa.4497.4.9
Bronstein O., Kroh A., Loya Y. (2016). Reproduction of the long-spined sea urchin Diadema setosum in the gulf of aqaba - implications for the use of gonad-indexes. Sci. Rep. 6, 1–11. doi: 10.1038/srep29569
Bronstein O., Loya Y. (2014). Echinoid community structure and rates of herbivory and bioerosion on exposed and sheltered reefs. J. Exp. Mar. Biol. Ecol. 456, 8–17. doi: 10.1016/j.jembe.2014.03.003
Bronstein O., Loya Y. (2015). Photoperiod, temperature, and food availability as drivers of the annual reproductive cycle of the sea urchin Echinometra sp. from the gulf of aqaba (Red Sea). Coral Reefs 34, 275–289. doi: 10.1007/s00338-014-1209-3
Buchwald R. T., Feehan C. J., Scheibling R. E., Simpson A. G. B. (2015). Low temperature tolerance of a sea urchin pathogen: implications for benthic community dynamics in a warming ocean. J. Exp. Mar. Biol. Ecol. 469, 1–9. doi: 10.1016/j.jembe.2015.04.006
Byers J. E., Reichard S., Randall J. M., Parker I. M., Smith C. S., Lonsdale W. M., et al. (2002). Society for conservation biology directing research to reduce the impacts of nonindigenous species. Cons Biol. 16, 630–640. doi: 10.1046/j.1523-1739.2002.01057.x
Capella-Gutiérrez S., Silla-Martínez J. M., Gabaldón T.. (2009). trimAl: a tool for automated alignment trimming in large-scale phylogenetic analyses. J. Bioinform. 25, 1972–1973. doi: 10.1093/bioinformatics/btp348
Carlton J. T. (1987). Patterns of transoceanic marine biological invasions in the pacific ocean. Bull. Mar. Sci. 41, 452–465.
Carreiro-Silva M., Mcclanahan T. R. (2001). Echinoid bioerosion and herbivory on Kenyan coral reefs: the role of protection from fishing. J. Exp. Mar. Biol. Ecol. 262, 133–153. doi: 10.1016/s0022-0981(01)00288-x
Chow S., Konishi K., Mekuchi M., Tamaki Y., Nohara K., Takagi M., et al. (2016). DNA Barcoding and morphological analyses revealed validity of Diadema clarki ikeda 1939 (Echinodermata, echinoidea, diadematidae). Zookeys 2016, 1–16. doi: 10.3897/zookeys.585.8161
Clemente S., Hernández J. C., Brito A. (2011). Context-dependent effects of marine protected areas on predatory interactions. Mar. Ecol. Prog. Ser. 437, 119–133. doi: 10.3354/meps09243
Clemente S., Hernández J. C., Rodríguez A., Brito A. (2010). Identifying keystone predators and the importance of preserving functional diversity in sublittoral rocky-bottom areas. Mar. Ecol. Prog. Ser. 413, 55–67. doi: 10.3354/meps08700
Clemente S., Hernández J. C., Toledo K., Brito A. (2007). Predation upon Diadema aff. antillarum in barren grounds in the canary islands. Sci. Mar. 71, 745–754. doi: 10.3989/scimar
Clemente S., Lorenzo-Morales J., Mendoza J. C., Lopez C., Sangil C., Alves F., et al. (2014). Sea Urchin Diadema africanum mass mortality in the subtropical eastern Atlantic: role of waterborne bacteria in a warming ocean. Mar. Ecol. Prog. Ser. 506, 1–14. doi: 10.3354/meps10829
Coll M., Piroddi C., Steenbeek J., Kaschner K., Lasram F. B. R., Aguzzi J., et al. (2010). The biodiversity of the Mediterranean Sea: estimates, patterns, and threats. PloS One 5. doi: 10.1371/journal.pone.0011842
Costello K. E., Lynch S. A., McAllen R., O'Riordan R. M., Culloty S. C. (2022). Assessing the potential for invasive species introductions and secondary spread using vessel movements in maritime ports. Mar. pollut. Bull. 177, 113496. doi: 10.1016/j.marpolbul.2022.113496
Crocetta F., Agius D., Balistreri P., Bariche M., Bayhan Y. K., Çakir M., et al. (2015). New Mediterranean biodiversity records (October 2015). Mediterr Mar. Sci. 16 (3), 682–702. doi: 10.12681/mms.1477
Crooks J. A. (2005). Lag times and exotic species: the ecology and management of biological invasions in slow-motion. Ecoscience 12, 316–329. doi: 10.2980/i1195-6860-12-3-316.1
Digenis M., Arvanitidis C., Dailianis T., Gerovasileiou V. (2022). Comparative study of marine cave communities in a protected area of the south-Eastern Aegean Sea, Greece. J. Mar. Sci. Eng. 10, 660. doi: 10.3390/jmse10050660
Dimitriadis C., Neave E. F., Shum P., Mariani S., D’Amen M., Azzurro E. (2023). First records of Sphyraena chrysotaenia (Klunzinger 1884) and Diadema setosum (Leske 1778) in the marine protected area of zakynthos island (Ionian Sea – greece): a result of a joint monitoring strategy. Acta Adriatica 64, 1–4. doi: 10.32582/aa.64.1.3
do Hung Dang V., Fong C. L., Shiu J. H., Nozawa Y. (2020). Grazing effects of sea urchin Diadema savignyi on algal abundance and coral recruitment processes. Sci. Rep. 10, 1–9. doi: 10.1038/s41598-020-77494-0
Downing A. S., Van Nes E. H., Janse J. H., Witte F., Cornelissen I. J. M., Scheffer M., et al. (2012). Collapse and reorganization of a food web of mwanza gulf, lake Victoria. Ecol. Appl. 22, 229–239. doi: 10.1890/11-0941.1
Ebert T. A. (1982). Longevity, life history, and relative body wall size in sea urchins. Ecol. Monogr. 52, 353–394. doi: 10.2307/2937351
Edler D., Klein J., Antonelli A., Silvestro D. (2021). raxmlGUI 2.0: a graphical interface and toolkit for phylogenetic analyses using RAxML. Methods Ecol. Evol. 12, 373–377. doi: 10.1111/2041-210X.13512
El-Serehy H. A., Abdallah H. S., Al-Misned F. A., Irshad R., Al-Farraj S. A., Almalki E. S. (2018). Aquatic ecosystem health and trophic status classification of the bitter lakes along the main connecting link between the red Sea and the Mediterranean. Saudi J. Biol. Sci. 25, 204–212. doi: 10.1016/j.sjbs.2017.12.004
El-Serehy H. A., Al-Misned F. A., Shafik H. M., Al-Rasheid K. A., Bahgat M. M. (2014). Surface microplankton composition at a hyper saline oligotrophic environment of bitter lake on the Suez canal, Egypt. Turk J. Fish Aquat Sci. 14, 439–448. doi: 10.4194/1303-2712-v14_2_14
Exadactylos A., Vafidis D., Tsigenopoulos C. S., Gkafas G. A. (2019). High connectivity of the white seabream (Diplodus sargus, l. 1758) in the Aegean Sea, Eastern Mediterranean basin. Animals 9. doi: 10.3390/ani9110979
Furió J. G., Templado J. (2012). Centrostephanus longispinus,” in VV.AA., bases ecológicas preliminares para la conservación de las especies de interés comunitario en españa: invertebrados (Madrid: Ministerio de Agricultura, Alimentación y Medio Ambiente), 1–58.
Galanos C. J., Kritikos S. (2019). Diadema setosum (Leske 1778) (Echinodermata, echinoidea, diadematidae), first record for simi island, hellas, eastern Mediterranean. Parnassiana Arc 7, 15–19.
Galil B. (2007). Loss or gain? invasive aliens and biodiversity in the Mediterranean Sea. Mar. pollut. Bull. 55, 314–322. doi: 10.1016/j.marpolbul.2006.11.008
Galil B., Boero F., Campbell M. L., Carlton J. T., Cook E., Fraschetti S., et al. (2015). ‘Double trouble’: the expansion of the Suez canal and marine bioinvasions in the Mediterranean Sea. Biol. Invasions 17, 973–976. doi: 10.1007/s10530-014-0778-y
Galil B., Goren M. (2014). “Metamorphoses: bioinvasions in the Mediterranean Sea,” in The Mediterranean Sea: its history and present challenges (Netherlands: Springer), 463–478. doi: 10.1007/978-94-007-6704-1_27
Galil B., Marchini A., Occhipinti-Ambrogi A., Ojaveer H. (2017). The enlargement of the Suez canal-erythraean introductions and management challenges. Manage. Biol. Invasions 8, 141–152. doi: 10.3391/mbi.2017.8.2.02
Gerovasileiou V., Akel E. H., Akyol O., Alongi G., Azevedo F., Babali N., et al. (2017). ). new Mediterranean biodiversity records (July 2017). Mediterr Mar. Sci. 18 (2), 355–384. doi: 10.12681/mms.13771
Gerovasileiou V., Bancila R. I., Katsanevakis S., Zenetos A. (2022). Introduced species in Mediterranean marine caves: an increasing but neglected threat. Mediterr Mar. Sci. 23, 995–1005. doi: 10.12681/mms.31284
Geyer L. B., Zigler K. S., Tiozzo S., Lessios H. A. (2020). Slow evolution under purifying selection in the gamete recognition protein bindin of the sea urchin Diadema. Sci. Rep. 10. doi: 10.1038/s41598-020-66390-2
Gianni F., Bartolini F., Airoldi L., Ballesteros E., Francour P., Guidetti P., et al. (2013). Conservation and restoration of marine forests in the Mediterranean Sea and the potential role of marine protected areas. Adv. Oceanogr Limnol 4, 83–101. doi: 10.1080/19475721.2013.845604
Goh B. P. L., Lim D. Y. F. (2015). Distribution and abundance of sea urchins in Singapore reefs and their potential ecological impacts on macroalgae and coral communities. Ocean Sci. J. 50, 211–219. doi: 10.1007/s12601-015-0018-0
Gökoğlu M., Balcı B. A., Çolak H., Banbul Acar B. A. (2007). Akdeniz’de uzun dikenli deniz kestanesi’nin (Diadema setosum; leske 1778) İkinci kaydı. in XI. sualtı bilim ve teknolojisi toplantısı (İstanbul: Koç Üniversitesi), 115.
Gökoğlu K., Kuşat M. (2022). Bir deniz akvaryumunun su temin Ünitesi civarında gözlemlenen makro faunadan seçilmiş türler. Acta Aquatica Turcica 1). doi: 10.22392/actaquatr.990402
Gökoğlu M., Teker S., Julian D. (2016). Antik Kent phaselis in bazı bentik indo-pasifik türleri. J. Interdiscip. Mediterr. Stud. 2, 225–233. doi: 10.18367/pha.16015
Gül M. R., Aydin M. (2021). “Density and habitat preference of an invasive species (Diadema setosum, Leske 1778) in the Mediterranean Sea,” in In book of proceedings XII international scientific agriculture symposium, Jahorina, Bosnia Herzegovina. 990–995.
Hernández J. C., Clemente S., Girard D., Pérez-Ruzafa Á., Brito A. (2010). Effect of temperature on settlement and postsettlement survival in a barrens-forming sea urchin. Mar. Ecol. Prog. Ser. 413, 69–80. doi: 10.3354/meps08684
Hernández J. C., Clemente S., Sangil C., Brito A. (2008). The key role of the sea urchin Diadema aff. antillarum in controlling macroalgae assemblages throughout the canary islands (eastern subtropical atlantic): an spatio-temporal approach. Mar. Environ. Res. 66 (2), 259–270. doi: 10.1016/j.marenvres.2008.03.002
Hernández J. C., Sangil C., Lorenzo-Morales J. (2020). Uncommon southwest swells trigger sea urchin disease outbreaks in Eastern Atlantic archipelagos. Ecol. Evol. 10 (15), 7963–7970. doi: 10.1002/ece3.6260
Hooper D. U., Adair E. C., Cardinale B. J., Byrnes J. E. K., Hungate B. A., Matulich K. L., et al. (2012). A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature 486, 105–108. doi: 10.1038/nature11118
Hothorn T., Bretz F., Westfall P. (2008). Simultaneous inference in general parametric models. Biometrical J. 50 (3), 346–363. doi: 10.1002/bimj.200810425
Hughes T. P., Keller B., Jackson J., Boyle M. (1985). Mass mortality of the echinoid Diadema antillarum philippi in Jamaica. Bull. Mar. Sci. 36, 377–384.
Jones G. M. (1985). Paramoeba invadens n. sp. (Amoebida, paramoebidae), a pathogenic amoeba from the sea urchin, Strongylocentrotus droebachiensis, in Eastern Canada. J. Protozool 32, 564–569. doi: 10.1111/j.1550-7408.1985.tb03075.x
Kacem H., Neifar L. (2014). The reproductive biology of the grey triggerfish Balistes capriscus (Pisces: balistidae) in the gulf of gabès (south-eastern Mediterranean Sea). J. Mar. Biol. Assoc. United Kingdom 94, 1531–1537. doi: 10.1017/S0025315414000824
Kalyaanamoorthy S., Minh B. Q., Wong T. K. F., von Haeseler A., Jermiin L. S. (2017). ModelFinder: fast model selection for accurate phylogenetic estimates. Nat. Methods 14, 587–589. doi: 10.1038/nmeth.4285
Katsanevakis S., Acar Ü., Ammar I., Balci B. A., Bekas P., Belmonte M., et al. (2014a). New Mediterranean biodiversity records (October 2014). Mediterr Mar. Sci. 15, 675–695. doi: 10.12681/mms.1123
Katsanevakis S., Coll M., Piroddi C., Steenbeek J., Lasram F. B. R., Zenetos A., et al. (2014b). Invading the Mediterranean Sea: biodiversity patterns shaped by human activities. Mediterr Mar. Sci. 1. doi: 10.3389/fmars.2014.00032
Katsanevakis S., Poursanidis D., Hoffman R., Rizgalla J., Rothman S. B. S., Levitt-Barmats Y., et al. (2020). Unpublished Mediterranean records of marine alien and cryptogenic species. Bioinvasions Rec 9, 165–182. doi: 10.3391/bir.2020.9.2.01
Katsanevakis S., Sini M., Dailianis T., Gerovasileiou V., Koukourouvli N., Topouzelis K., et al. (2017). Identifying where vulnerable species occur in a data poor context: combining satellite imaging and underwater occupancy surveys. Mar. Ecol. Prog. Ser. 577, 17–32. doi: 10.3354/meps12232
Koukouras A., Kitsos M. S., Tzomos T., Tselepides A. (2010). Evolution of the entrance rate and of the spatio-temporal distribution of lessepsian Crustacea decapoda in the Mediterranean Sea. Crustaceana 83 (12), 1409. doi: 10.1163/001121610X539498
Koukouras A., Sinis A. I., Bobori D., Kazantzidis S., Kitsos M.-S. (2007). The echinoderm (Deuterostomia) fauna of the Aegean Sea, and comparison with those of the neighbouring seas. J. Biol. Res. 7, 67–92.
Lanfear R., Frandsen P. B., Wright A. M., Senfeld T., Calcott B. (2017). Partitionfinder 2: new methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 34, 772–773. doi: 10.1093/molbev/msw260
Larsson A. (2014). AliView: A fast and lightweight alignment viewer and editor for large datasets. J. Bioinform. 30, 3276–3278. doi: 10.1093/bioinformatics/btu531
Lavoie D. M., Smith L. D., Ruiz G. M. (1999). The potential for intracoastal transfer of non-indigenous species in the ballast water of ships. Estuar. Coast. Shelf Sci. 48, 551–564. doi: 10.1006/ecss.1999.0467
Lawrence J. M. (1975). On the relationships between marine plants and sea urchins. Oceanography Mar. Biology: Annu. Rev. 13, 213–286.
Lazarus M., Frid O., Yahel R. (2020). Israeli Mediterranean Marine protected areas surveys (Marine bioblitz) - final report for 2015, 2017 and 2019. Israel Nature and Parks Authority 1, 1–57.
Lessios H. A. (1988). Mass mortality of Diadema antillarum in the Caribbean: what have we learned? Annu. Rev. Ecol. Syst. 55 (5), 371–393. doi: 10.1146/annurev.es.19.110188.002103
Lessios H. A., Kessing B. D., Pearse J. S. (2001). Population structure and speciation in tropical seas: global phylogeography of the sea urchin Diadema. Evol. (N Y) 55, 955–975.
Lewis J. B. (1966). Growth and breeding in the tropical echinoid Diadema antillarum philippi. Bull. Mar. Sci. 16, 151–158.
Ling S. D., Johnson C. R., Frusher S. D., Ridgway. K. R. (2009). Overfishing reduces resilience of kelp beds to climate-driven catastrophic phase shift. Proc. Natl. Acad. Sci. U.S.A. 106 (52), 22341–22345. doi: 10.1073/pnas.0907529106
Loreau M., de Mazancourt C. (2013). Biodiversity and ecosystem stability: a synthesis of underlying mechanisms. Ecol. Lett. 16, 106–115. doi: 10.1111/ele.12073
Mack R. N., Simberloff D., Lonsdale W. M., Evans H., Clout M., Bazzaz F. A. (2000). Biotic invasions: causes, epidemiology, global consequences, and control. Ecol. Appl. 10, 689–710. doi: 10.1890/1051-0761(2000)010[0689:BICEGC]2.0.CO;2
McClanahan T. R., Kurtis J. D. (1991). Population regulation of the rock-boring sea urchin Echinometra mathaei (de blainville). J. Exp. Mar. Biol. Ecol. 147, 121–146. doi: 10.1016/0022-0981(91)90041-t
Millot C., Taupier-Letage I. (2005). “Circulation in the Mediterranean Sea” in The Mediterranean Sea. Ed. Saliot A. (Berlin, Heidelberg: Springer Berlin Heidelberg), 29–66. doi: 10.1007/b107143
Mokady O., Lazar B., Loya Y. (1996). Echinoid bioerosion as a major structuring force of red Sea coral reefs. Biol. Bull. 190, 367–372. doi: 10.2307/1543029
Muthiga N. A., McClanahan T. R. (2020). “Diadema,” in Sea Urchins: biology and ecology developments in aquaculture and fisheries science. Ed. Lawrence J. M. (Elsevier), 397–418. doi: 10.1016/B978-0-12-819570-3.00023-8
Mytilineou C., Akel E. H. K., Babali N., Balistreri P., Bariche M., Boyaci Y., et al. (2016). New mediterranean biodiversity records (November 2016). Mediterr Mar. Sci. 17, 794–821. doi: 10.12681/mms.1976
Nader M. R., Indary S. (2011). First record of Diadema setosum (Leske 1778) (Echinodermata, echinoidea, diadematidae) from Lebanon, Eastern Mediterranean. Aquat Invasions 6, 23–25. doi: 10.3391/ai.2011.6.S1.005
Nour O. M., al Mabruk S. A. A., Adel M., Corsini-Foka M., Zava B., Deidun A., et al. (2022). First occurrence of the needle-spined urchin Diadema setosum (Leske 1778) (Echinodermata, diadematidae) in the southern Mediterranean Sea. Bioinvasions Rec 11, 199–205. doi: 10.3391/bir.2022.11.1.20
Pascual M., Rives B., Schunter C., MaCpherson E. (2017). Impact of life history traits on gene flow: a multispecies systematic review across oceanographic barriers in the Mediterranean Sea. PloS One 12. doi: 10.1371/journal.pone.0176419
Pawson D. L., Miller J. E. (1983). Systematics and ecology of the sea-urchin genus Centrostephanus (Echinodermata: echinoidea) from the Atlantic and Eastern pacific oceans. Smithson Contrib Mar. Sci. 1–15. doi: 10.5479/si.01960768.20.1
Pearse J. S. (1970). Reproductive periodicities of indo-pacific invertebrates in the gulf of suez. III. the echinoid Diadema setosum (Leske). Bull. Mar. Sci. 20, 697–720.
Pecorino D., Lamare M. D., Barker M. F. (2012). Growth, morphometrics and size structure of the diadematidae sea urchin Centrostephanus rodgersii in northern new Zealand. Mar. Freshw. Res. 63 (7), 624–634. doi: 10.1071/mf12040
Pimm S. L. (1984). The complexity and stability of ecosystems. Nature 307, 321–326. doi: 10.1038/307321a0
Por F. D. (1971). One hundred years of Suez canal–a century of lessepsian migration: retrospect and viewpoints. Syst. Zool. 20, 138–152.
QGIS Development Team (2023). QGIS geographic information system (Open Source Geospatial Foundation Project). Available at: http://qgis.osgeo.org.
Qiu J. W., Lau D. C. C., Cheang C.c., Chow W.k. (2014). Community-level destruction of hard corals by the sea urchin Diadema setosum. Mar. pollut. Bull. 85, 783–788. doi: 10.1016/j.marpolbul.2013.12.012
Rambaut A., Drummond A. J., Xie D., Baele G., Suchard M. A. (2018). Posterior summarization in Bayesian phylogenetics using tracer 1.7. Syst. Biol. 67, 901–904. doi: 10.1093/sysbio/syy032
R Core Team (2022). R: a language and environment of statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.R-project.org/.
Rilov G. (2016). Multi-species collapses at the warm edge of a warming sea. Sci. Rep. 6. doi: 10.1038/srep36897
Ronquist F., Teslenko M., van der Mark P., Ayres D. L., Darling A., Höhna S., et al. (2012). Mrbayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61, 539–542. doi: 10.1093/sysbio/sys029
Rotter A., Klun K., Francé J., Mozetič P., Orlando-Bonaca M. (2020). Non-indigenous species in the Mediterranean Sea: turning from pest to source by developing the 8Rs model, a new paradigm in pollution mitigation. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00178
Sammarco P. W. (1982). Effects of grazing by Diadema antillarum philippi (Echinodermata: echinoidea) on algal diversity and community structure. J. Exp. Mar. Biol. Ecol. 65, 83–105. doi: 10.1016/0022-0981(82)90177-0
Sangil C., Hernández J. C. (2021). Recurrent large-scale sea urchin mass mortality and the establishment of a long-lasting alternative macroalgae-dominated community state. Limnol Oceanogr 67 (S1), 1–14. doi: 10.1002/lno.11966
Schroeder K., Chiggiato J., Bryden H. L., Borghini M., Ben Ismail S. (2016). Abrupt climate shift in the Western Mediterranean Sea. Sci. Rep. 6, 1–7. doi: 10.1038/srep23009
Schwarz G. (1978). Estimating the dimension of a model. Ann. Stat. 6 (2), 461–464. doi: 10.1214/aos/1176344136
Sela I., Ashkenazy H., Katoh K., Pupko T.. (2015). GUIDANCE2: accurate detection of unreliable alignment regions accounting for the uncertainty of multiple parameters. Nucleic Acids Res 43, 7–14. doi: 10.1093/nar/gkv318
Soukissian T. H., Denaxa D., Karathanasi F., Prospathopoulos A., Sarantakos K., Iona A., et al. (2017). Marine renewable energy in the Mediterranean Sea: status and perspectives. Energies (Basel) 10. doi: 10.3390/en10101512
Stucky B. J. (2012). Seqtrace: a graphical tool for rapidly processing DNA sequencing chromatograms. J. Biomolecular Techniques 23, 90–93. doi: 10.7171/jbt.12-2303-004
Trifinopoulos J., Nguyen L.-T., von Haeseler A., Minh B. Q. (2016). W-IQ-TREE: a fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 44, W232–W235. doi: 10.1093/nar/gkw256
Tsiamis K., Aydogan O., Bailly N., Balistreri P., Bariche M., Carden-Noad S., et al. (2015). New Mediterranean biodiversity records (July 2015). Mediterr Mar. Sci. 16 (2), 472–488. doi: 10.12681/mms.1440
Tsiamis K., Azzurro E., Bariche M., Çinar M. E., Crocetta F., de Clerck O., et al. (2020). Prioritizing marine invasive alien species in the European union through horizon scanning. Aquat Conserv. 30, 794–845. doi: 10.1002/aqc.3267
Turan C., Erguden D., Uygur N. (2011). On the occurrence of Diadema setosum (Leske 1778) in antakya bay, Eastern Mediterranean Sea. J. Black Sea/Mediterranean Environ. 17, 4–8.
Ulman A., Yildiz T., Demirel N., Canak O., Yemişken E., Pauly D. (2021). The biology and ecology of the invasive silver-cheeked toadfish (Lagocephalus sceleratus), with emphasis on the Eastern Mediterranean. NeoBiota 68, 145–175. doi: 10.3897/neobiota.68.71767
Vafidis D., Antoniadou C., Voulgaris K., Varkoulis A., Apostologamvrou C. (2021). Abundance and population characteristics of the invasive sea urchin Diadema setosum (Leske 1778) in the south Aegean Sea (eastern Mediterranean). J. Biol. Res. (Greece) 28, 1–14. doi: 10.1186/s40709-021-00142-9
Vredenburg V. T. (2004). Reversing introduced species effects: experimental removal of introduced fish leads to rapid recovery of a declining frog. Proc. Natl. Acad. Sci. 101, 7646–7650. doi: 10.1073/pnas.0402321101
Williams R. J., Griffithsb B. F., van der Wal E. J., Kellyb J. (1988). Cargo vessel ballast water as a vector for the transport of non-indigenous marine species. Estuar. Coast. Shelf Sci. 26, 409–420. doi: 10.1016/0272-7714(88)90021-2
Yapıcı S. (2018). Unusual observation of the alien sea urchin Diadema setosum (Leske 1778) in the Aegean Sea: recent and recorded occurrences. Thalassas: Int. J. Mar. Sci. 34, 267–269. doi: 10.1007/s41208-017-0060-z/investigated
Yeruham E., Rilov G., Shpigel M., Abelson A. (2015). Collapse of the echinoid Paracentrotus lividus populations in the Eastern Mediterranean - result of climate change? Sci. Rep. 5. doi: 10.1038/srep13479
Yokes B., Galil B. S. (2006). The first record of the needle-spined urchin Diadema setosum (Leske 1778) (Echinodermata: echinoidea: diadematidae) from the Mediterranean Sea. Aquat Invasions 1, 188–190. doi: 10.3391/ai.2006.1.3.15
Keywords: Diadema, Levantine Basin, Mediterranean, non-indigenes species, population outbreak, alien species, invasion dynamics
Citation: Zirler R, Leck LA, Farkash TF, Holzknecht M, Kroh A, Gerovasileiou V, Huseyinoglu MF, Jimenez C, Resaikos V, Yokeş MB and Bronstein O (2023) Gaining a (tube) foothold – trends and status following two decades of the long-spined echinoid Diadema setosum (Leske, 1778) invasion to the Mediterranean Sea. Front. Mar. Sci. 10:1152584. doi: 10.3389/fmars.2023.1152584
Received: 27 January 2023; Accepted: 15 May 2023;
Published: 31 May 2023.
Edited by:
Pedro Morais, Florida International University, United StatesReviewed by:
José Carlos Hernández, University of La Laguna, SpainSimone Farina, Anton Dohrn Zoological Station Naples, Italy
Copyright © 2023 Zirler, Leck, Farkash, Holzknecht, Kroh, Gerovasileiou, Huseyinoglu, Jimenez, Resaikos, Yokeş and Bronstein. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Omri Bronstein, YnJvbnN0ZWluQHRhdWV4LnRhdS5hYy5pbA==