 José Carlos García-Gómez1,2*
José Carlos García-Gómez1,2* Amparo Cid-Iturbe1
Amparo Cid-Iturbe1 Enrique Ostalé-Valriberas1*Rocío Espada1
Enrique Ostalé-Valriberas1*Rocío Espada1 José Luis Carballo1
José Luis Carballo1 Darren Andrew Fa2Carlos García-Olaya3
Darren Andrew Fa2Carlos García-Olaya3 Rocio Blanca-Sújar1
Rocio Blanca-Sújar1- 1Laboratorio de Biología Marina de la Universidad de Sevilla (LBM-US)/Área de Investigación I+D+i del Acuario de Sevilla/Estación de Biología Marina del Estrecho (Ceuta), Universidad de Sevilla, Seville, Spain
- 2Natural Sciences and Environment Research Hub, University of Gibraltar, Gibraltar, United Kingdom
- 3Área de Investigación Biológica I+D+i del Acuario de Sevilla, Sevilla, Spain
The limpet (Patella ferruginea), endemic to the western Mediterranean, is the most endangered intertidal mollusk listed in the Habitats Directive (EU). The translocation methodology of this species is a scientific challenge faced by those port infrastructures – where there are established breeding subpopulations - that may be subject to emergency works, restoration, or extension of breakwaters. This study presents the results of an innovative two-phase pilot project (P1A/B and P2), involving the legally authorized translocation of individuals of this species obtained in three relocation experiments in the Marina of Puerto José Banús (Málaga) in July 2021 (phase 1: P1A and B) and May 2022 (phase 2: P2) (study period: July 2021-October 2022). The specimens translocated in both phases were from the same port except for those of P1B, which came from the Marbella Marina as an exceptional case. These translocations were undertaken on concrete breakwater tetrapod units. We show that the somewhat forced, slow and unregulated extraction of specimens to be translocated during the first experiment of the P1A pilot project was the main cause of mortality, as it generated an important exhaustion of the fixation musculature, which resulted in a low survival (66%) after the first month of transfer, whereas during the same period of time. In the subsequent experiments (where the problem was methodologically corrected, a fast extraction technique -less than 5 seconds-, regulated and without injuries), in P1B was 100%, in P2 with cage was 93.3%, and, in P2 without cage it was 83.3%, which reflected an improvement in translocation methodology. The results of this research are compared according to the descriptors of success and failure of each protocol in order to observe the differences and thus enable their replicability.
Introduction
Patella ferruginea Gmelin, 1791 (commonly known as the ferruginean or Mediterranean ribbed limpet), is considered the most endangered endemic marine invertebrate in the Western Mediterranean (Laborel-Deguen and Laborel, 1991a; Ramos, 1998; Espinosa et al., 2014), also by the European Council Directive 92/43/EEC (Council Directive, 1992). It is, therefore, a protected species specifically listed as such in different administrative provisions or international conventions: National Catalogue of Threatened Species (in danger of extinction), Annex IV of the Habitats Directive, Annex II of the Barcelona Convention and Annex II of the Bern Convention. In Spain, this led to the publication of the National Strategy for the conservation of this species (MMAMRM, 2008), under the auspices of the Ministry of the Environment and Rural and Marine Affairs.
Due to the high accessibility of its intertidal habitat, this species has undergone significant regression (Raffaelli and Hawkins, 1996; Haedrich and Barnes, 1997; Rochet and Trenkel, 2003), especially through human exploitation (Aversano, 1986; Guerra-García et al., 2004a; Moreno, 2004), not only for use as fishing bait, due to its muscular foot (Pombo and Escofet, 1996), but also for its ornamental interest. An example of this concerns Lottia gigantea G. B. Sowerby, 1834 (Fenberg and Roy, 2012) or Scutellastra mexicana (Broderip and Sowerby, 1829), the latter being the largest limpet in the world (Dance, 1967; Keen, 1971) and the only known species of patellid in the Eastern Pacific (Carballo et al., 2020). In this sense, P. ferruginea is the largest limpet species in the Mediterranean. Finally, populations of P. ferruginea have also been affected by the reduction of habitat quality and alteration by coastal works, the increase of contaminants (Espinosa et al., 2014; Espinosa and Rivera-Ingraham, 2017) along the coast and changes in sea surface temperature (Freitas et al., 2023). The habitat of this species, the upper midlittoral fringe of the intertidal zone, is highly sensitive and vulnerable to anthropogenic influence. In this zone, limpets have been considered keystone species (Henriques et al., 2017; Marra et al., 2017), and, as Raffaelli and Hawkins (1996) point out, they maintain a mosaic of open spaces that allow the coexistence of many species, thus contributing to increase marine biodiversity. Whilst the keystone species attribute has not been sufficiently argued for P. ferruginea, its potential as a bioindicator taxon of clean and renewed waters has been described and investigated (Espinosa et al., 2007; García-Gómez, 2015), so it can be used as a sentinel species indicator of anthropogenic environmental disturbances or changes. For these reasons, in most ports and marinas of the Alboran Sea (with the particular exception of the ports of Ceuta, Melilla and Gibraltar, whose internal waters are subject to significant renewal), P. ferruginea is common or abundant in the outer docks, being absent -or of accidental presence- in the inner zone (where other limpet species can live).
Distribution and status of populations
The species was widely distributed throughout the western Mediterranean during the Pleistocene (Caton-Thompson, 1946; Laborel-Deguen and Laborel, 1991a) and its presence was abundantly recorded on the coasts of North Africa and Europe until the end of the 19th century. But it was at the beginning of the 20th century, when a clear regression of the species began (Laborel-Deguen and Laborel, 1991a; Templado, 2001). Currently, the species has almost completely disappeared from the western Mediterranean coasts of Europe, from its northern and eastern areas, being relegated to the northern and western coasts of Corsica (Laborel-Deguen and Laborel, 1991a; Sardinia (Porcheddu and Milella, 1991; Doneddu and Manunza, 1992; Cristo et al, 2007; Cristo and Caronni, 2008), Pantellaria and Egadi Islands (Laborel-Deguen and Laborel, 1991a) and Tuscany (Italian peninsula) (Curini–Galletti, 1979; Biagi and Poli, 1986).
The Alboran Sea is the area where the species is most abundant, maintaining high homogeneity and low genetic diversity (Espinosa and Ozawa, 2006; Casu et al., 2011). It is located on the island of Alboran (Paracuellos et al., 2003; Templado et al., 2006), with populations firmly established on Iberian south-Mediterranean coast from Almería to Tarifa (Arroyo et al., 2011), including Gibraltar (Fa et al., 2018) being their western distribution and the strict limit of distribution of the south-Mediterranean coast (García-Gómez and Magariño, 2010), being especially concentrated in external areas of harbors or marina breakwaters (e.g., Motril, Marbella, José Banús, La Duquesa, Sotogrande, La Línea-Alcaidesa, Gibraltar, Algeciras, Tarifa); and North Africa coasts: Ceuta (Guerra-García et al., 2004b; Espinosa, 2006; Espinosa et al., 2009; Rivera–Ingraham et al., 2011a) and Melilla (González-García et al., 2006; González et al., 2015), the Chafarinas Islands (Guallart, 2006), the Algerian islands of Rachgoun (Frenkiel, 1975) and Habibas (Boumaza and Semroud, 2001; Espinosa, 2009), reaching Cape Bon (Espinosa, 2006) and Zembra Island (Tunisia) (Boudouresque and Laborel–Deguen, 1986).
This species of limpet is sequential protandric hermaphrodite with external fertilization (Espinosa et al., 2006; Rivera-Ingraham et al., 2011b), whose reproductive period is between September and December (Frenkiel, 1975). The larvae phase is development with a lecithotrophic larvae that act as limiting factor in the ability of a larva to reach assume the “desperate larva hypothesis” postulates that lecithotrophic larvae become less discriminating in their settlement requirements over time, due to depletion of energy reserves (Botello and Krug, 2006).
Patella ferruginea in coastal infrastructures, unresolved conflict
In addition, inhabiting in natural rocky habitats, P. ferruginea is found on artificial structures, usually on walls, breakwaters and rocks or blocks of breakwaters linked to port infrastructures in the Alboran Sea. In Europe it is the only endangered species that forms important groups of breeding individuals associated with coastal infrastructure breakwaters, even on the smooth, external surfaces of concrete walls linked to such breakwaters (García-Gómez et al., 2011; García-Gómez et al., 2015; Maestre Delgado et al., 2018).
The main conflict in the management of this species in artificial habitats comes from the need to relocate specimens from breakwaters to other places, as a consequence of reconstruction works on breakwaters - either due to severe storms or longer-term deterioration and/or necessary improvements in port infrastructures or their enlargement. In this regard, in Spain, the National Conservation Strategy for this species (MMAMRM, 2008), “discourages the transfer of specimens, except for scientific research or conservation of the species duly justified”. This is because of high mortality when translocating individuals. This position has led to the generation of “an unresolved problem” to relevant Port and Environmental Authorities (the latter competent in the granting of authorizations) by directly affecting future works involving the restoration or modification of the layout of seawalls containing the species and, in the context of natural habitats, by making it impossible to reintroduce adults of the species in geographical areas where it used to exist and has now become locally extinct. However, the Strategy advocates the “establishment of appropriate measures to avoid damage to the usual port activities”, which requires the development of applied research, with concomitant implications, to help to solve the identified problems.
History of translocations of specimens
Previous experiments of translocations by direct extraction of specimens from their source substrata or on plates designed with 3D technology have been carried out: in Corsica (Laborel-Deguen and Laborel, 1991b), Ceuta, Strait of Gibraltar (Espinosa et al., 2008), Islas Hormigas, southern Spain (Guallart et al., 2014), Chafarinas Islands (APM, 2014), Gibraltar (Fa et al., 2018), Zembra Islands, Tunisia (Zarrouk et al., 2018) and between Melilla (North Africa) and La Línea (Algeciras Bay, southern Iberian Peninsula) (LIFE 15/NAT/ES/000987-REMoPaF Project, 2017-2021). In the discussion the results obtained are compared, although except for those from Gibraltar of translocation of specimens on their substrata (without removing from their home scar) that had very satisfactory survival results (Fa et al., 2018), the others have obtained poor or mediocre results. This supports the furtherance of research aimed at improving the methodology of translocations, a workable solution to which is increasingly urgent due to the problems generated by increasingly frequent and more complex coastal engineering.
Objectives
The main objective of this work was to test a new method of translocation of specimens of P. ferruginea in an artificial habitat composed of concrete tetrapods of 6 to 10 tons to assess its suitability for specimen transfer. The method focused on the rapid and manual extraction of specimens (in relaxation phase during the rising tide), transport (in attachment plates and zip bags without fixation) and subsequent relocation of the specimens, in order to obtain higher survival than those obtained in previous experiments and different substrata. This new technique will allow future translocations - when duly justified - to be both viable and sustainable which will also have a direct application for future reintroductions of adult specimens to natural habitats where the species is now absent but where it is known to have previously existed.
Methodology
Study area, physical characteristics of the receptor breakwater and precautions derived from the operational risk
The two ports of Marbella (south of the Iberian Peninsula, Spain) where the pilot project - authorized by the ‘Ministro de Agricultura, Pesca y Desarrollo Rural (CAPyDR) del Gobierno de la Junta de Andalucía - has been carried out are Puerto José Banús (PJB) and Puerto Deportivo de Marbella (PDM). The PJB has been the only receptor of all the specimens translocated from the outer breakwater of the same port (a hostile environment due to its spatial configuration and exposure to waves), as well as the specimens from the PDM (very close to PJB), which has only acted as a donor of a small number of specimens in the pilot project (Figure 1). The PJB breakwater is subject to frequent and intense waves (generated by the wind and amplified by the drastic decrease in depth due to the accumulated sedimentation in the outer area near the breakwater). This presented important health and safety problems in the processes of extraction and relocation of specimens due to the final slope of the breakwater, the size of the tetrapods and the smoothness of their slippery arms, as well as the different current vortices generated when the waves break over them according to their orientation. These reasons sometimes made it necessary to designate team members exclusively to watch the waves (sometimes generated suddenly, next to the breakwater) and, on occasion, the use of safety lines, helmets and vests to access where the limpets were located.
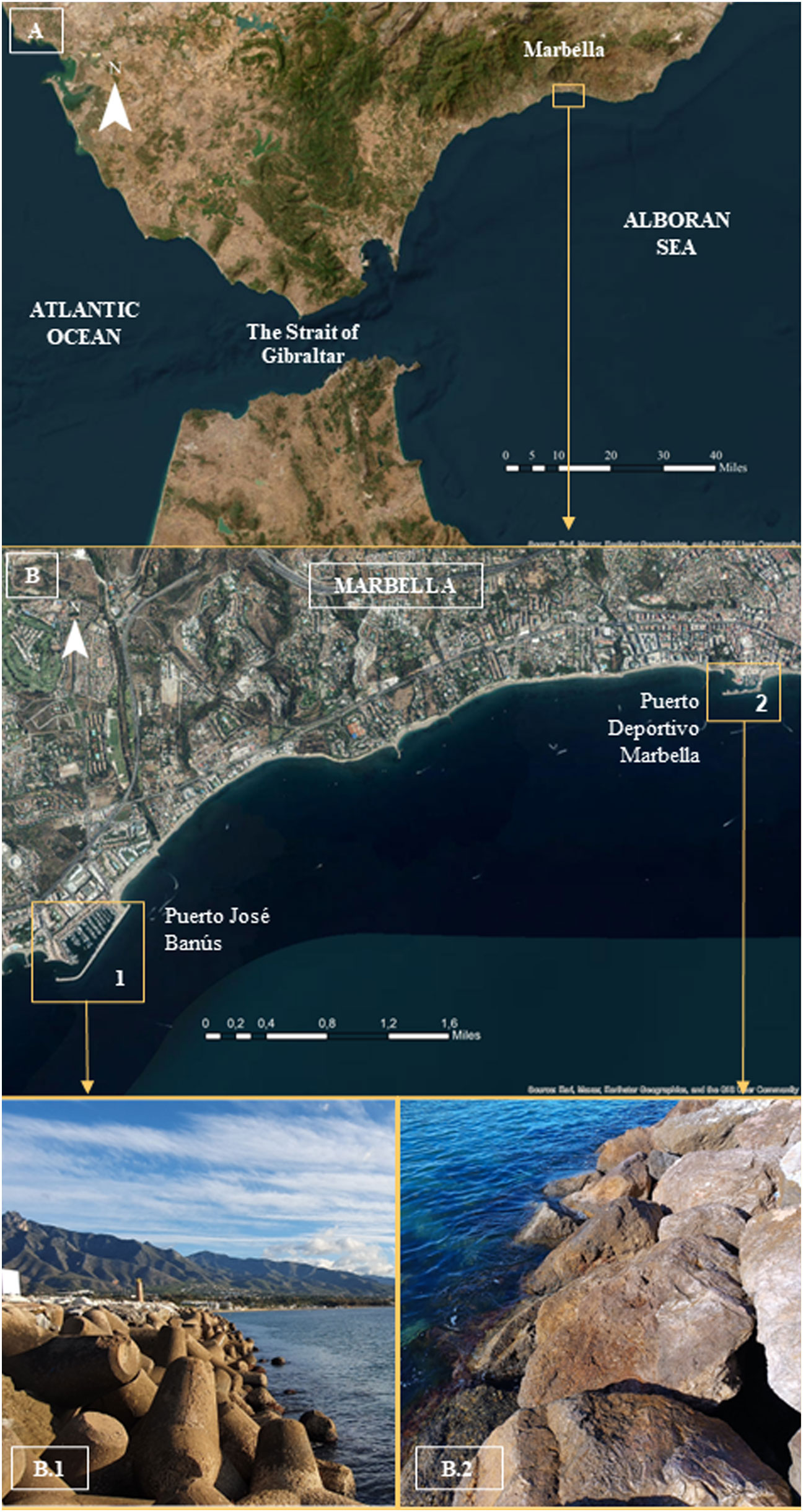
Figure 1 (A) Alboran Sea and location of the study area. (B) Location of PJB (1) and PDM (2), as well as the different substrata of their breakwaters of both phases (P1A and B; P2) of the pilot project: B1, key substratum of concrete tetrapods of the study, used as both donor and receptor zones; B2, riprap substratum of the exclusively donor zone, whose receptor zone was located in the tetrapods of B1.
Authorized specimens, phases of the pilot-project (P1A and B, and P2) and material used
In 2020, the subpopulation of P. ferruginea was censused in PJB, estimated at 1,153 specimens of different sizes (of which 818 were adults, over 30 mm) (CAGPyDS-JA, 2021), along its seawall (1,067 meters long). The pilot project authorized by the Competent Environmental Authority (CAPyDR-JA) for translocation of specimens established on concrete tetrapods was strictly limited to 110 specimens in two phases (P1A, 50 specimens; and P2, 60 specimens). In addition, within the scope of P1 and exceptionally - due to an emergency work situation inside the PDM (14 km from the PJB) - the transfer of 3 adult specimens (P1B) from dolomitic quarry rocks (PDM) to concrete tetrapods in the PJB was authorized, an operation carried out at the end of P1A. This was a special trial because of its significant added value, since the transferred specimens involved transportation between ports, medium and substratum (from natural dolomitic quarry rock to artificial concrete tetrapods), and especially because it allowed modifications to improve P1A before starting P2. This last phase (P2) started in May 2022, with the aim of applying the methodological advances detected in P1A and P1B, as a consequence of the detailed observation of unexpected early mortalities. The translocated specimens (manually extracted from their substrata in the same harbor breakwater of PJB (P1A and P2), have represented 9.5% (110 out of 1,153) of the total contingent found along the exterior arm of the harbor.
The plates used for an express transfer and their subsequent displacement to the receptor area were standardized, made of cement and with a rough, non-slip surface measuring 20 x 20 cm. They were used preliminarily to avoid stressing the limpets (as soon as they were extracted) so that they would use the plates as an intermediate substratum to be installed in a very short time (30 minutes maximum) into pre-established sites, anchored by means of metal bolts previously placed in the receptor area, only requiring to be screwed in. The cages - after previous resistance tests of a previous plastic prototype - were designed in stainless metal mesh (50 X 40 cm), specifically flexible (to mold to the curved arms of the tetrapods and withstand the strong waves), and of low height (to avoid the possible overturning of the transferred limpets and their loss), with flanges secured by flexible aluminum plates on which the cages were screwed to the arms of the tetrapods after drilling them (with a concrete drill bit and a Stanley Fatmax 1250 watt hammer drill) and the use of multifunctional grooved expansion plugs (Fisher type). The cages had an opening to introduce the limpets, which had a metallic fold that acted as a door and allowed its closure with plastic flanges. The cages were sized to contain the limpets for only 3-4 weeks (after which they were disassembled), a critical adaptation period in which, according to pre-existing experiments reviewed in the discussion, the highest mortalities are usually recorded. Rounded-edge spatulas, soft plastic bags (zip type) and plastic coolers were used for the collection, storage and transfer of translocation specimens. To determine mortality and survival within size classes, millimetric precision calipers were used to measure specimens.
Figure 2 shows the zoning of the receptor areas in PJB of the specimens transferred in phases 1 and 2 of the transfer, with their respective control zones - Receptor Control (CR), Donor Control (CD) and Non-Donor Control (CND) for the first phase (P1), and Control (C2) for the second phase (P2). The receptor area for the specimens transferred in the first phase (P1) was established at the east end of the breakwater for TP1A and at the west end of the breakwater for TP1B (3 limpets from PDM). For the second phase (P2), two receptor zones were delimited, one for the translocation of limpets without cages (TP2WCa) and the other for those translocated with cages (TP2Ca), each of them being the donor zone of the other. The limpets translocated in TP1A come from the CD zone and those in TP1B, from PDM, not represented in Figure 2 since it is a separate site to the PJB. The control zones for the P1 phase of translocation are: the CD, the CR (shared with the TP1A zone of translocated limpets), and the non-donor zone CND located at the western end of the PJB breakwater. For the P2 phase, the control zone C2 and adjacent to the CR, which overlaps with the receptor zone TP2WCa, was chosen.
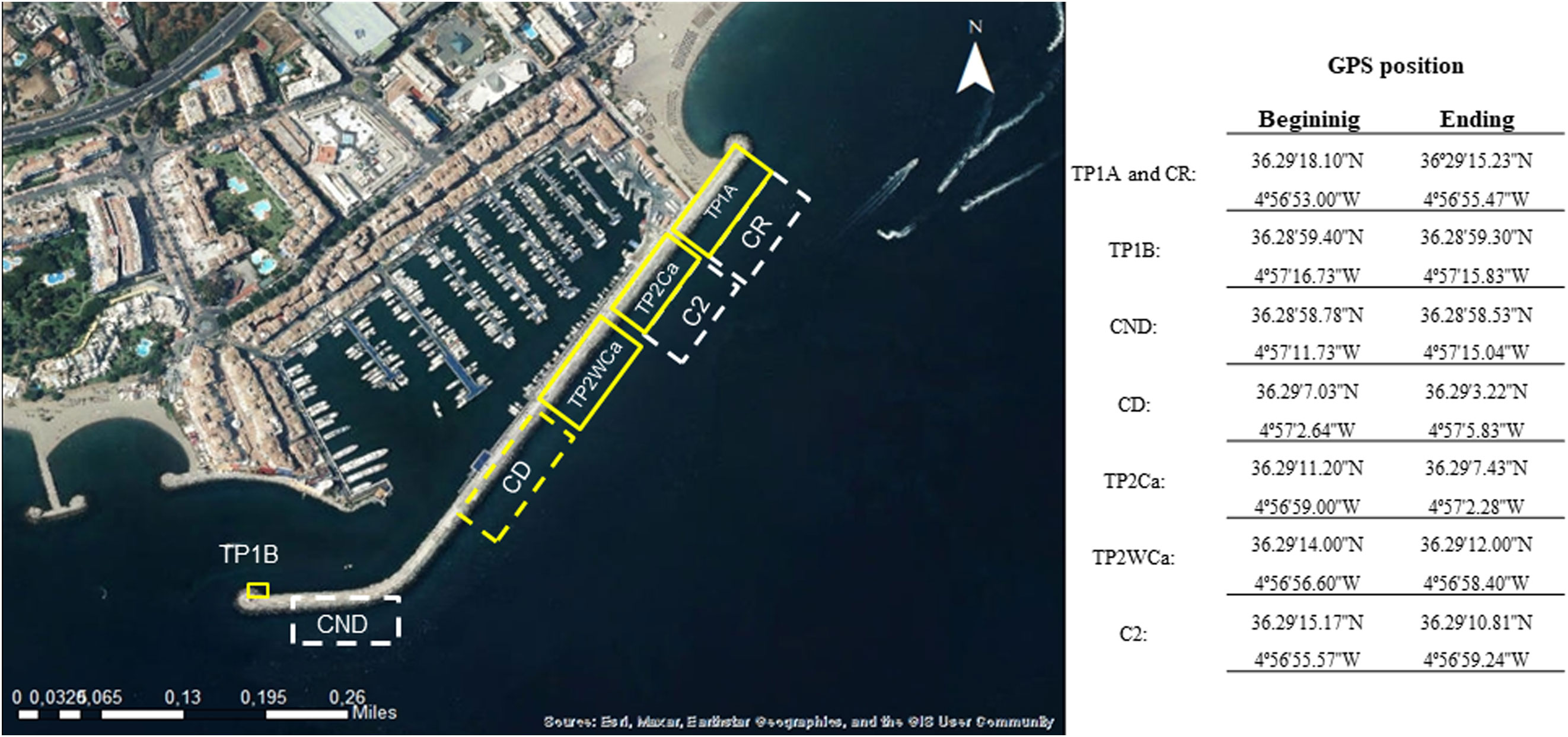
Figure 2 Location of receptor, control, and donor areas in PJB. TP1A and TP1B: receptor areas for limpets translocated in phases P1A and P1B of the pilot project. Receptor areas for limpets translocated in phase P2, TP2Ca: limpets relocated in fixed metal cages; TP2WCa: limpets relocated without cages. The 4 control zones considered are delimited in dashed lines, CR, receptor control; CD, donor control; CND, non-donor control; C2, phase 2 control.
The choice of control zones (untranslocated resident limpets) shared with receptor zones (see Figure 2) was made because it was considered that a comparison of survival percentages between both zones in the same stretch of breakwater and under identical environmental conditions could allow a better deduction of some of the causes that could explain the disappearance of specimens (e.g. extracted by bathers or illegal shell fishermen). One of the difficulties of this type of study lies in the fact that most of the data supporting the mortality are based on missing specimens and not on specimens found dead, whose detailed examination, including necropsy, could help to determine the cause of death or at least rule out other possible causes such as those indicated in the example mentioned above.
Physical and biological parameters
To better understand the oceanographic and atmospheric environmental conditions, sea surface temperature, prevailing wind (at 10 meters height), wave height and chlorophyll a (Chl-a) in the water column were obtained from specialized web databases.
Temperature by annual station was obtained in degrees Celsius (MODIS-Aqua MODISA_L3m_SST_8d_4k vR 2019 dataset) from NASA’s GIOVANNI tool (https://giovanni.gsfc.nasa.gov/giovanni/) at 11 microns and 4 km resolution. This tool allows researchers to visualize and perform small statistical analyses (Acker and Leptoukh, 2007).
The wind data were obtained from the SIMAR point 2025077 https://www.puertos.es/es-es/oceanografia/Paginas/portus.aspx. (official website of Puertos del estado del Ministerio de Transportes, Movilidad y Agenda Urbana de España). HARMONIE-AROME is the meso-scale model used, non-hydrostatic, with high spatial resolution. The data consulted have been obtained at 10 meters above sea level, with a resolution of approximately 2.5 km and a 48 h prediction horizon. Wind speed (extracted by months, for the full year 2021 and up to June 2022, in order to standardize data) is expressed in m/s and wind direction in degrees. Seasonal means and standard deviations were calculated and plotted in bar histograms.
The wave data obtained from the same source as the wind (WAM and WaveWatch models), complemented by the wind fields of the HARMONIE-AROME model. The first two are third-generation spectral models that solve the energy balance equation without making any a priori assumptions about the shape of the wave spectrum. The data have a resolution of 700 m. From the SIMAR point 2025077 we extracted and classified by months, the significant wave height data that approximately represents the mean height of the highest third of waves (significant height, Hs), data extracted from the Puertos del Estado website (complete year 2021 and 2022 until June, using this time scale to make the data uniform). Seasonal means and standard deviation were calculated (winter, spring, summer, autumn 2021 and winter and spring 2022). The data were plotted in a bar histogram with their associated standard errors. The wave height varied from wave to wave, and again the mean height of the highest third of waves (Hs) was plotted.
Chl-a concentration data were extracted from the MODIS-Aqua Level-3 sensor available provided by OBPG (NASA Ocean Biology Processing Group, 2018) at 4 km resolution from the NASA Ocean Color website (http://oceancolor.gsfc.nasa.gov/).
Statistical analysis
To compare the results of survival of translocated limpets and establish if there are statistically significant differences between these results and those obtained for “control” individuals, we used the Log-rank hypothesis test (Mantel, 1966; Peto and Peto, 1972), also known as the Mantel-Cox test, which allows us to compare survival distributions of two samples. This test is based on the χ2 model, in which the events (death) in each group are compared. It provides a p value (probability that the differences are due to chance), in addition to the median or mean survival. To compare the survival of P. ferruginea individuals over time, we used the Kaplan-Meier estimator, also called the limit product estimator, which is a nonparametric statistical method that takes into account each of the times contributed by each individual studied (Jager et al., 2008; Stel et al., 2011). Statistical analyses were performed using SPSS-25 software.
Extraction, transfer, and relocation of specimens
The appropriate transfer period was established between March and May (2021 for P1 and 2022 for P2), since between June and August (both inclusive) there could be days of high temperature (which can cause desiccation at low tide since the specimens have not been able to develop their new scar), and also between September and December, since this is the reproductive period of the species (Frenkiel, 1975). The months from December to February are not recommended due to the frequency of bad weather in winter, especially storms in the area. Prior to the definitive establishment of the methodological transfer protocol, tests were carried out on the mobilization of tetrapods and the cutting of their arms, as well as on a prototype plastic cage with metal fixation, all of which were rejected because of their practical and/or technical infeasibility and, in the case of the cages, because they could not withstand the strong waves (Figure 3: 1, 2 and 6).

Figure 3 1 and 2: Preliminary tests of concrete tetrapod transfers and cutting of tetrapod arm sections with diamond head cutter, which reflected their technical infeasibility; 3: limpet marked with epoxy resin stamped with specific code; 4: limpet placed on a small previously defaunated surface (descaled from barnacle Chthamalus stellatus) to facilitate its new healing and sealing; 5: opening holes in the arm of a tetrapod, with a 1,200 watt hammer drill; 6: discarded prototype plastic cage and aluminum attachment strips, initially tested at P2, to test its wave resistance; 7: all-metal, stainless steel, wave-resistant cage design successfully used at P2; 8 and 9: intermediate attachment plates for extracted limpets, discarded at the beginning of P1A, were replaced with soft plastic “zip-lock” bags (see text); 10: time of extraction of specimens in heavy surf at the PJB breakwater.
Before extraction
Prior to the start of the translocation operation, the donor (extraction) and receptor (relocation) zones were defined. In the donor zone, suitable individuals of P. ferruginea were identified. In the receptor area, relocation points were previously decided for each individual to be relocated, close to the resident limpets, choosing relocation surfaces as similar as possible in terms of structure, slope and roughness, but avoiding those that were very exposed to direct waves, since during the first weeks after relocation limpets are particularly vulnerable to them. In each of the areas, a series of preliminary works were carried out; A) Donor area: each individual limpet to be transferred was identified with a stamped epoxy resin mark with a numerical identification code, photographed and measured (length and width), and the substratum where it was located was characterized. Annotation of its location in a vertical, inclined or horizontal area, rough or smooth, with or without biofouling (e.g. barnacles). B) Receptor area: the choice of suitable relocation sites was based on being as similar as possible to those of its native (donor) area. The upper midlittoral fringe of the species’ intertidal zone was previously identified by the resident limpets-control or, if these are not nearby, by the surface of the P. ferruginea-specific fringe, typically with frequent presence of the lichen Verrucaria amphibia Clemente, 1814 and almost permanent presence of the barnacle Chthamalus stellatus (Poli, 1791), and comprised between the lower edge of the supralittoral Patella rustica L., 1758 belt and by the upper edge of the lower midlittoral shore zone determined by the red alga Ellisolandia elongata (J. Ellis and Solander) Hind and Saunders(2013), (Figure 4).

Figure 4 The dashed parallel lines delimit the strict strip of the upper midlittoral of the intertidal zone where P. ferruginea lives in the Strait of Gibraltar and nearby Mediterranean areas of the Alboran Sea. The upper line is the lower limit of the limpet P. rustica (in the image) and the lower line is the upper limit of the red alga Ellisolandia elongata, both very well defined in the image.
During extraction (donor site)
The extraction of specimens was carried out during the rising tide, high tide, or the beginning of the ebb tide, since it is at this time of emersion that limpets move out of their home scar. During the initial P1A exercise, it was observed that many limpets resisted, resulting in a rather forced extraction, leading to exhaustion of the peripheral muscles of the foot, subsequent weakness in their fixation in the receptor area and ultimately an increased risk of failure to attach or being dislodged by the swell. This was corrected on P1B and P2 by performing a rapid extraction (<5 seconds) that did not allow the animals to react and offer resistance. Extraction was always carried out through the posterior half of the shell and by “lever effect”, pushing the spatula - by the handle - upwards, so that the point of support was the edge of the shell and not soft areas of the body, which could be lethal for the animal, even in the medium term. Once the animals had been extracted by this method, it was checked and recorded whether they had suffered any damage to the foot, since this can also hinder their rapid attachment in the receptor area and, therefore, possible detachment and loss of the animal. If the animals remained too strongly attached for too long during the rising or high tide, since it was impossible to remove them with spatulas, it was planned to do so during low tide together with a small fragment of their substratum to which they were attached, by rapidly fragmenting the concrete or rock with a hammer drill (of at least 1,200 watts) and chisel bit, with additional support from a rotaflex and concrete cutting disc, if necessary. In the receptor area, the fragment of substratum extracted with each animal would be fixed in the receptor area with epoxy marine cement (see discussion).
During transfer
The transfer from the donor to the receptor area was initially carried out (in P1A) on small concrete plates with a granulated surface, with the aim of obtaining a second settlement on these that would minimize the stress of the animal during its transfer to the receptor area (with previously installed bolts), from where the animals could in their own time voluntarily leave the plates and establish their new home scar. However, this was discarded when it was proven in situ that the need for a second transfer to a new substratum actually increased stress to the animal. Instead, soft plastic ziplock bags (flooded with sea water and half-opened to allow water exchange), each containing one animal and transported in the dark in simple plastic coolers, proved to be the best method, avoiding the need for a second settlement of the animals. In most cases, the animals were transported within 30 minutes of extraction, although an interval of up to 3 hours was envisaged. All limpets from P1B and P2 were transferred in this way, which, together with the above-mentioned process of rapid extraction without injury, represented an important advance in the success of the transfers (see results and discussion).
During relocation (receptor zone)
In the receptor zone, the animals were deposited on their new surfaces, also during rising tide, high tide, or the beginning of the ebb tide, taking into consideration that, if they were extracted by surprise, quickly, there would be no muscular exhaustion and they would attach immediately, even if the swell was intense. This would also be an early indication, should the animal not be found during follow-up monitoring, that its absence might be due to other factors not associated with muscle exhaustion. Each transferred specimen was relocated, especially the adults (>3cm), taking into account the proximity of other resident control-adults, since there is evidence that chemical traces possibly reduce the stress of the transferred specimens and maximize the inter-sex reproductive potential (the smaller the distance between males and females, the greater the potential). This tendency for adults to aggregate has been observed previously in littoral stretches where there are few specimens (personal obs.). However, to avoid exceeding the carrying capacity of the system and, for a given area of the intertidal, on rocks or isolated artificial structures, it was ensured that the density of transferred specimens, together with residents, did not exceed the maximum observed, for similar sizes, in other sections of the same receptor area with a higher density of resident limpets.
Temporary monitoring
The specimens transferred in July 2021, from P1A and P1B were monitored for 16 months and those from P2, transferred in May 2022 for 5 months. After the relocation of all specimens, the periodicity of monitoring was as follows: every day for the first two weeks; every 2-3 days for the next two weeks; once a week for the second month; once every two weeks for the third and fourth month; and once a month thereafter, unless significant losses were noticed.
Results
For the study period and experimental area, wind speed and wave height data are shown in Figure 5.

Figure 5 Wind speed (left) and wave height (right) data in the study area during the project timeframe.
A particularly severe storm which took place in April 2022, with correspondingly large wave heights and corresponding material damage caused to the coast, only one of the translocated limpets was found to have disappeared shortly after, with a second loss 15 days later, apparently unrelated to the storm event (Figure 6). This would indicate that, despite the severity of the storm, the species studied showed a great adaptive capacity to the mechanical impact of the waves, and that tenacity and survival in translocated limpets in P1A were more strongly influenced by the condition of the animal (muscular exhaustion and/or physical damage caused by forced physical extraction), than by the strength of wave action experienced.
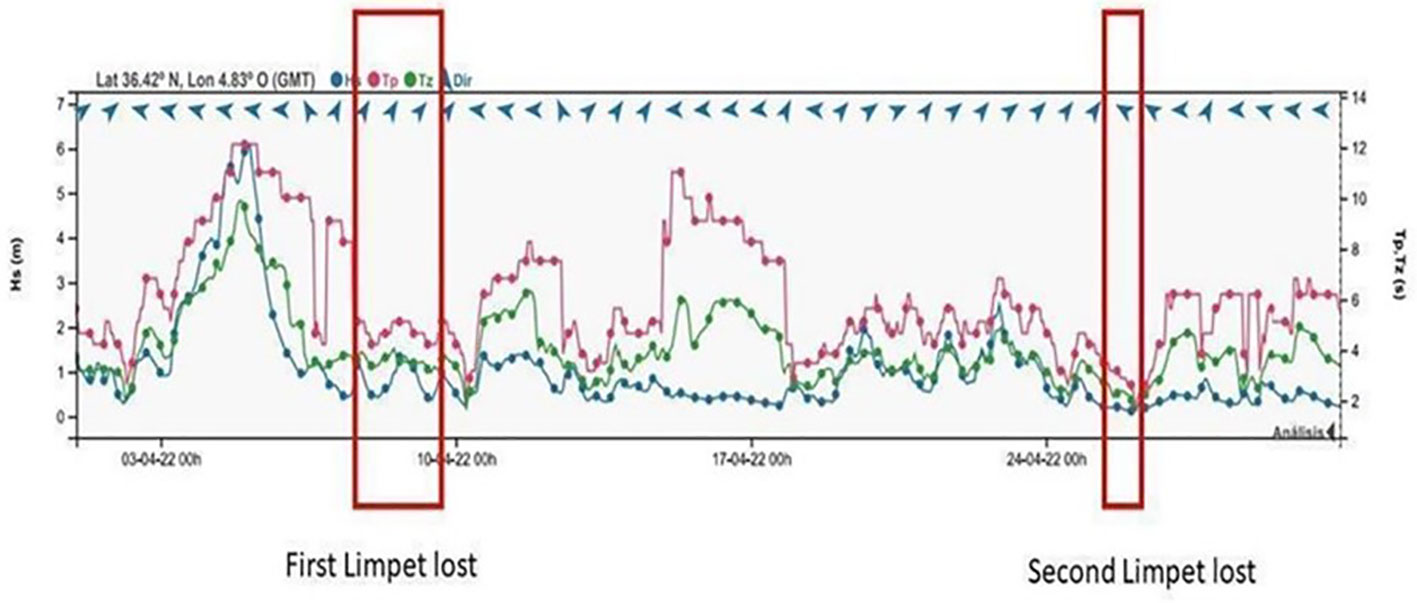
Figure 6 Maximum swell recorded during the study period (first week of April 2022), after which there was only the immediate loss of a single relocated limpet and a subsequent one 15 days later.
Satellite data for temperature and Chlorophyll-a are shown in Figure 7. Chlorophyll-a data for the springs of 2021 and 2022 show appreciable differences, with mean values being appreciably higher in spring 2022. Nonetheless, neither of these parameters were found to influence mortality in both control and receptor areas, given the highly positive outcomes in P1B and P2, once the methodological lessons learnt from P1A had been incorporated to the translocation techniques.
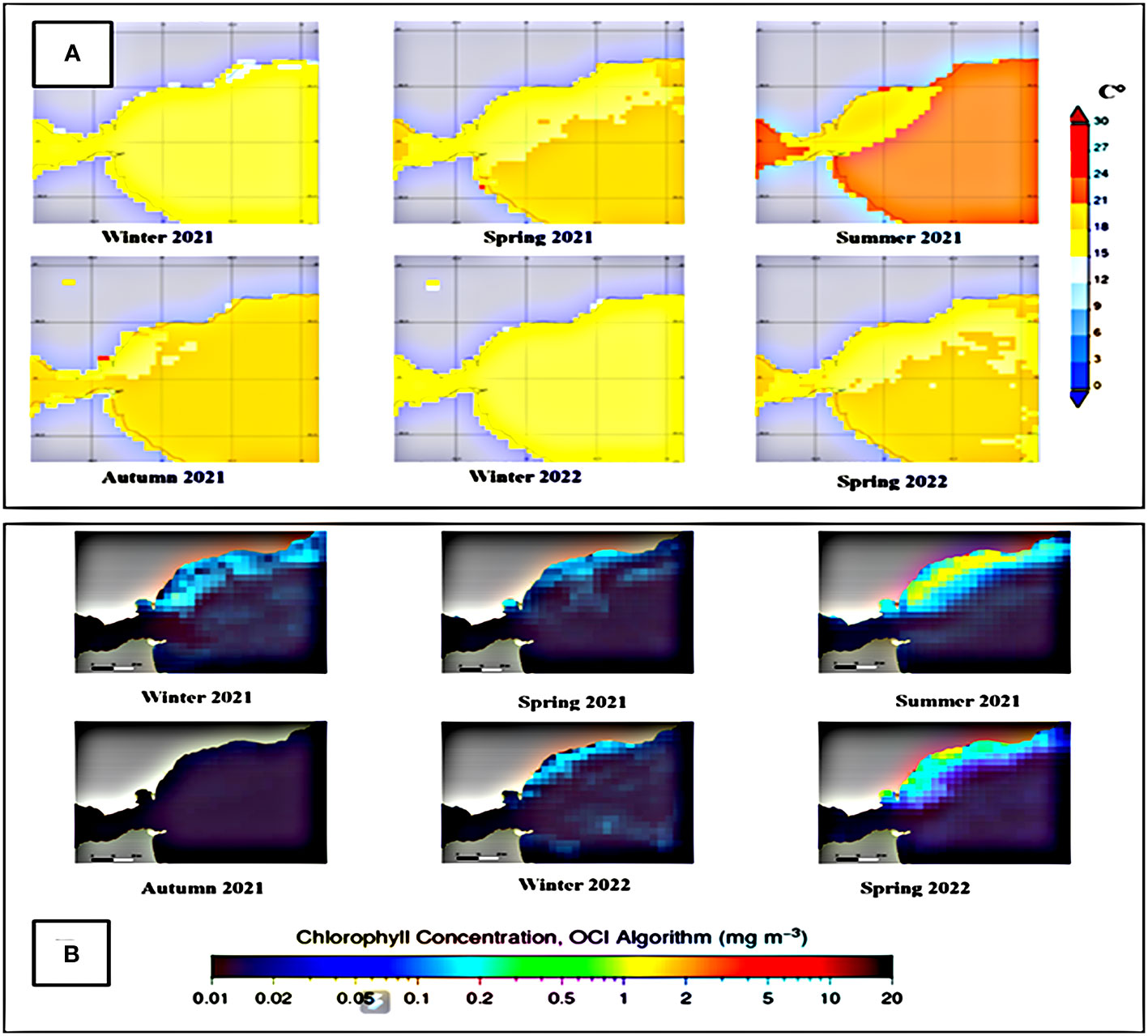
Figure 7 Average temperature (A) and chlorophyll (B) data from autumn 2021 to spring 2022.
Phases P1A and P1B (16 months of monitoring)
The P1A phase revealed the methodological errors of forced manual extraction, causing moderate to severe exhaustion of the foot muscles, resulting in difficulty of adherence in the receptor area and loss of individuals on swell days during the first days or weeks. Figure 8 shows the survival obtained over a time interval of 16 months, showing that the P1A phase obtained a 36% survival percentage while in P1B this was 100%, higher than that of the CR control zone with a survival percentage of 92%.
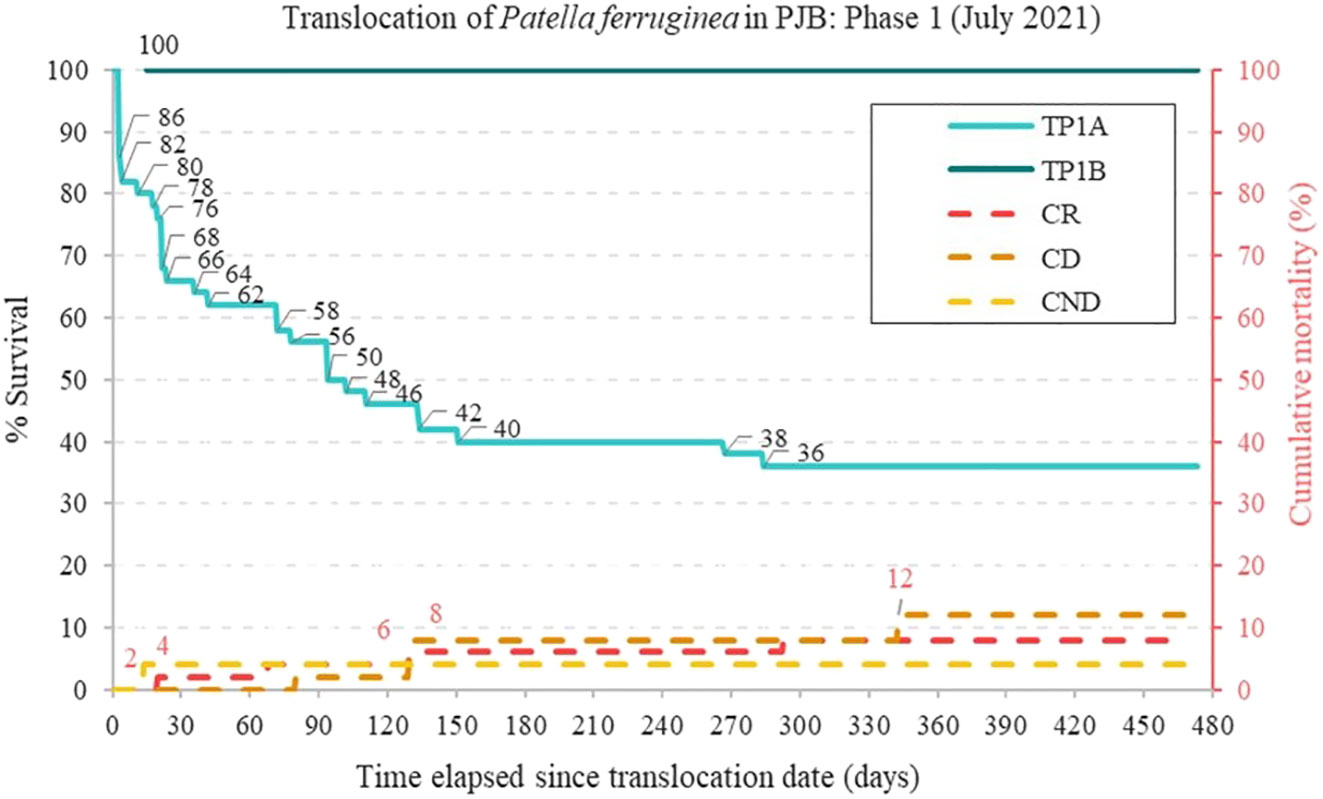
Figure 8 Translocations P1A and P1B with control areas in Puerto José Banús. Change in survival (%) of limpets in the translocation experiments in TP1A (Translocation P1A) and TP1B (Translocation P1B) and cumulative mortality (%) in the secondary axis of the control areas (CR, receptor control; CD, donor control; CND, non-donor control) from 15 July 2021 to 31 October 2022 (16 months). Number of limpets surveyed: NTP1A = 50; NTP1B = 3; NCR = 50; NCD = 50; NCND = 50.
There are significant differences between treatments (χ² = 112.37; p < 0.001). The mean life expectancy of the transferred individuals (TP1A) was 207.66 days and that of those established in the control areas (CD, CND, and CR) was 451.76 days (See Tables 1–3 of data analysis in Supplementary Material). Kaplan-Meier survival function data are shown in Figure 9A. Among the controls set for the P1A phase, there was no significant difference in the survival of individuals after 470 days (χ² = 4.75; p < 0.93).
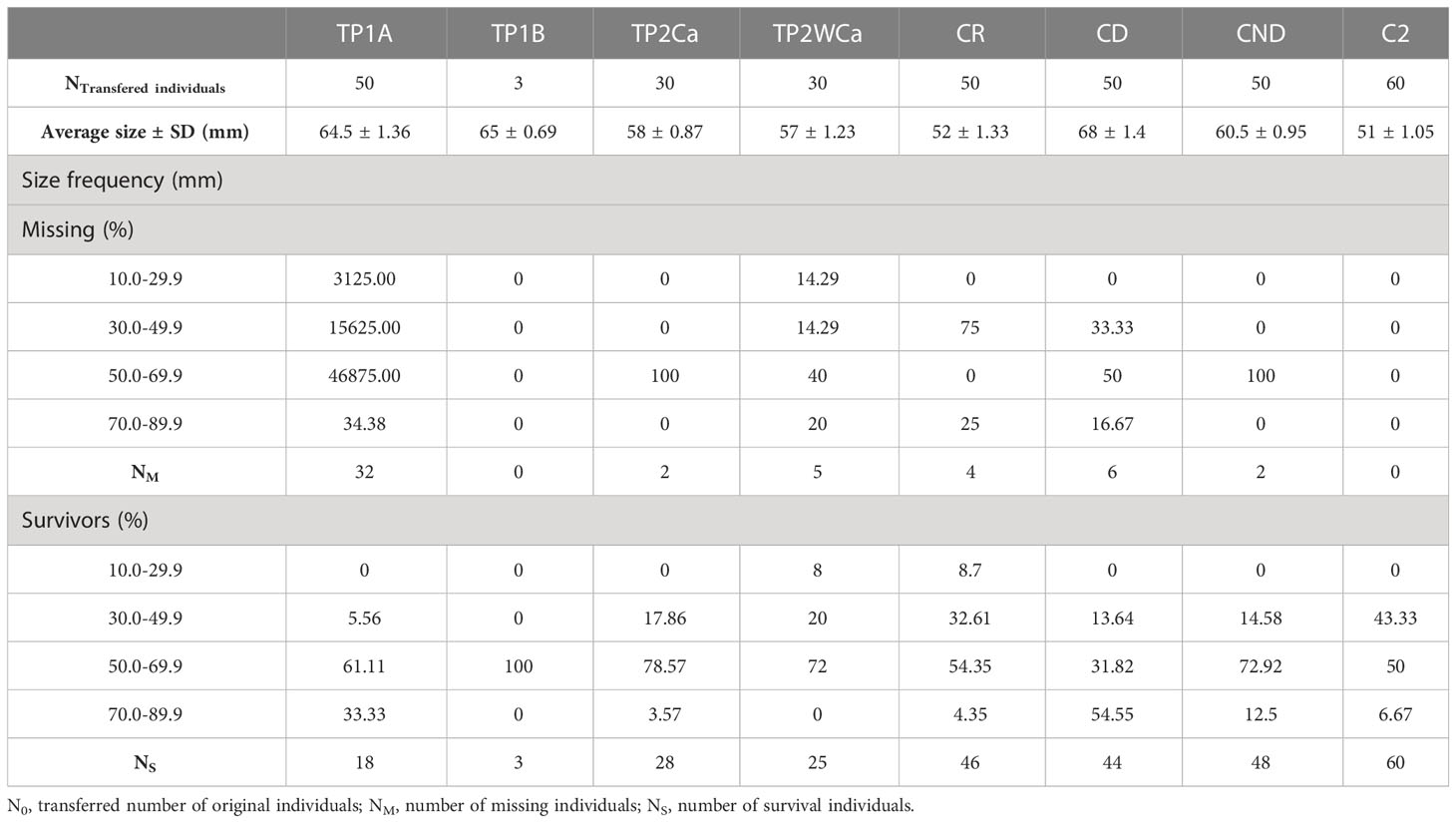
Table 1 Relative size frequency (%) of missing and surviving specimens in PJB phase 1B; TP2Ca, Translocation phase 2 with cage; TP2WCa, Translocation phase 2 without cage) and their controls (CR, receiver control; CD, donor control; CND, non-donor control; C2, phase 2 control).
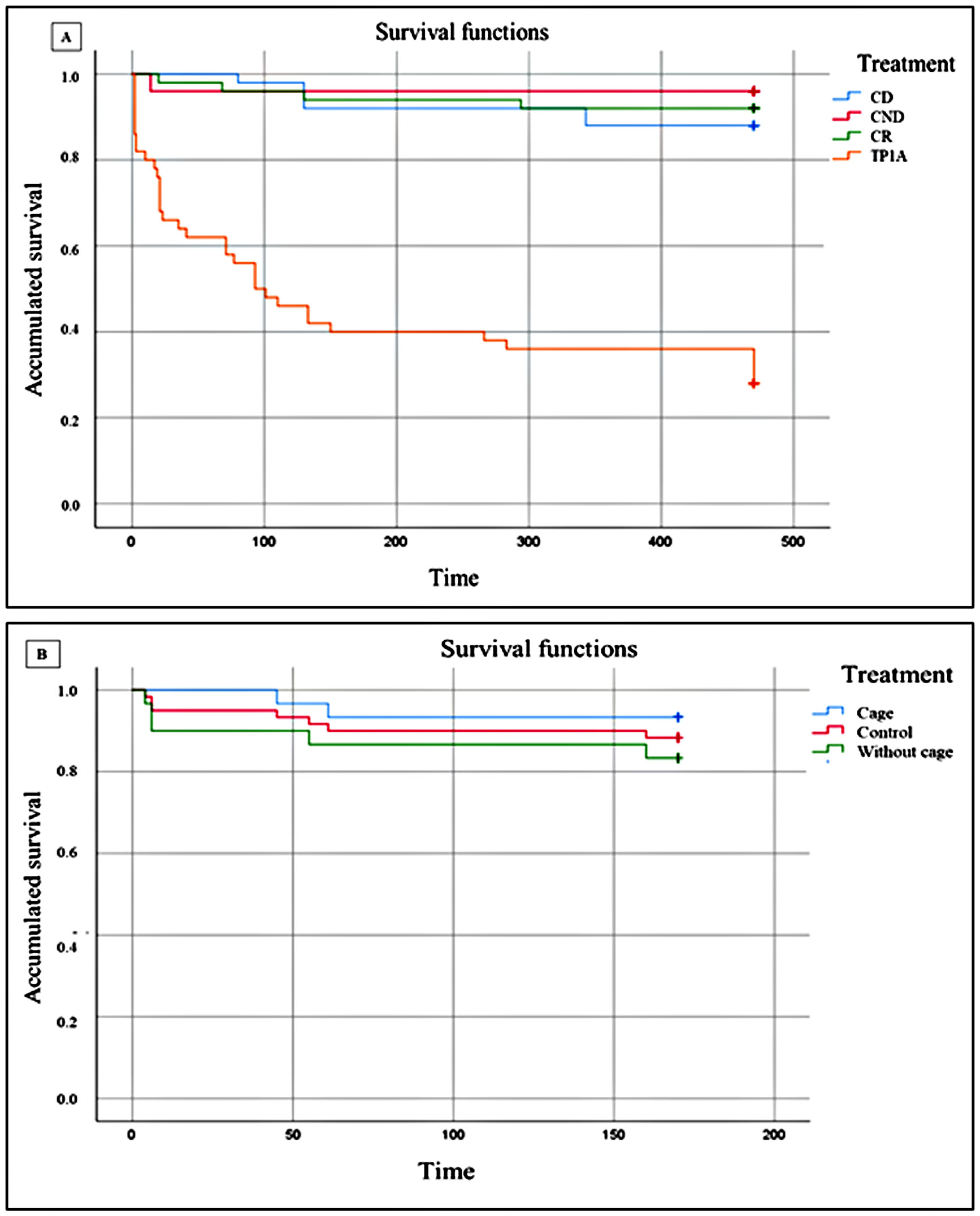
Figure 9 Cumulative survival on a linear scale (Kaplan-Meier function). (A): phase P1 (CD, Control donor; CND, Control non-donor; CR, Control receiver; TP1A, Translocation phase 1A); (B): phase P2 (P2WCa: with cage, P2Ca: without cage; C2: control.
In phase P1B, executed at the end of phase P1A and improving the methodology used in the latter, a 100% survival was obtained, despite being the only phase of the pilot project which presented greater risks to success, given that the transfer was executed between two ports (PDM and PJB) and different types and nature of substrata: from dolomitic quarry rocks (PDM) to concrete tetrapods (PJB). The excellent results obtained in this phase following the methodological improvements outlined and after monitoring over the first 3 months led to the approval of phase P2 by the administrative authority.
Phase P2 (5 months of monitoring)
In phase P2, compared to P1A in the first 5 months, a much higher survival was obtained (Figure 10A), confirming the success of the methodological approach applied in phase P1B. There are no significant differences between the different treatments, after 170 days of follow-up from the transfer of P. ferruginea individuals with respect to transferred individuals compared to those in the control area (χ² = 1.49; p = 0.473), showing that, in statistical terms, mortality is similar between transferred individuals and those in the control areas. Regarding the transferred individuals, although some more deaths were detected in translocations without cages (only 3 individuals), there were no significant differences, which suggests - although this should be confirmed with new experiments with a higher N - that in the future it should be possible to dispense with the installation of cages, with the economic and time savings that this would entail (see Tables 4–6, of data analysis in Supplementary Material).
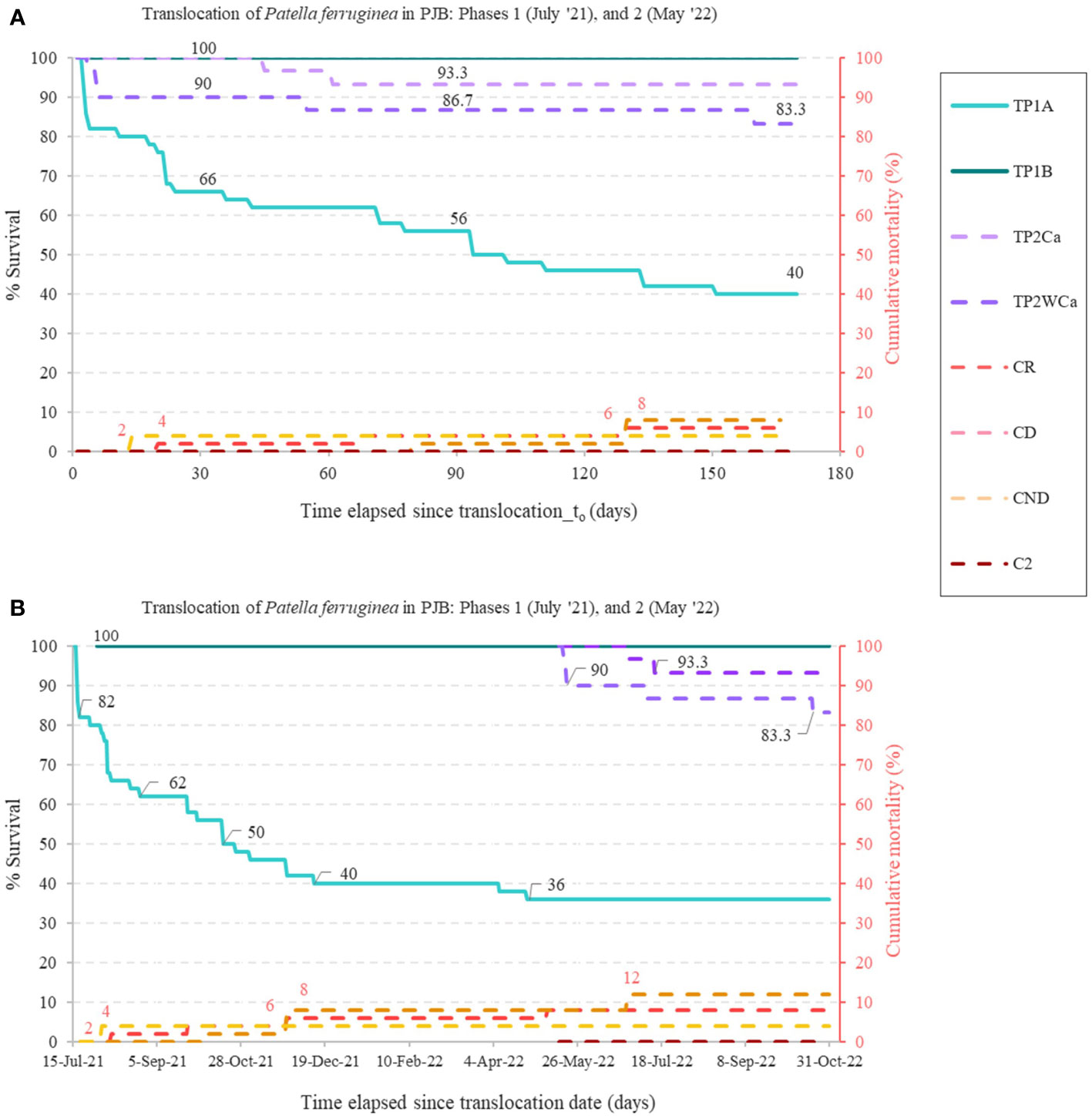
Figure 10 (A), Integrated survival results for P1A (TP1A: Translocation phase 1A), P1B (TP1B: Translocation phase 1B) and P2 in specimens transferred with relocation with cage (TP2Ca) and without cage (TP2WCa), during the first 5.5 months of the translocation, this time being the maximum monitoring time of P2 in this study (from 15 May 2022 to 31 October 2022); and (B), shows the integrated survival results for P1 of 16 months (from 15 July 2021 to 31 October 2022) together with P2 since May 2023. The secondary axis represents the cumulative mortality of the control areas (CR, receiver control; CD, donor control; CND, non-donor control; C2, phase 2 control -0% in this case-). Number of limpets studied: NTP1A = 50; NTP1B = 3; NTP2Ca = 30; NTP2WCa = 30; NCR = 50; NCD = 50; NCND = 50; NC2 = 60.
The survival function (Kaplan-Meier) data for the results illustrated in Figure 10, from P2, are shown in Figure 9B.
Size frequency according to mortality-survival of transferred and control specimens
The results of the relative size frequencies of both missing and surviving specimens in receptor areas (translocated specimens) and in control areas (non-translocated resident specimens) are shown in Table 1.
Considering the mean size of the translocated specimens (TP1A: 64.5 ± 1.36 mm; TP1B: 65 ± 0.69mm; TP2Ca: 58 ± 0.87mm; TP2WCa: 57 ± 1.23mm) and of the control specimens (CR: 52 ± 1.33mm; CD: 68 ± 1.4mm; CND: 60.5 ± 0.95mm; C2: 51 ± 1.05mm), the majority of specimens lost in the TP1A (46.88%, N= 15), TP2Ca (100%, N=2) and TP2WCa (40%, N=2) transfer phases, and in the CD and CND control areas (50%, N=3, and 100%, N=2, respectively) were between 50-69 mm in length, except for 75% (N=3) of the CR control specimens lost that were between 30-49 mm in length (see Table 1).
Regarding the surviving specimens transferred, most of them had a length between 50-69 mm: 61.11% (N=11) of TP1A, 100% (N=3) of TP1B, 78.57% (N=22) of TP2Ca, 72% (N=18) of TP2WCa; and control specimens CR with 54.35% (N=25), CND with 72.92% (N=35) and C2 with 50% (N=30), except control zone CD, where 54.55% (N=24) of the survivors had a length of 70-89 mm.
Summary of survival results
The overall results obtained are summarized in Table 2 and Figure 10B. The highest survivorships were obtained in P1B (TP1B 100%; 16 months of monitoring) and P2 with and without cages (TP2Ca: 93.3% and TP2WCa: 83.3%; 5.5 months of monitoring), due to the methodological improvements applied, derived from the first pilot experiment P1A, with a very poor result (survival at 16 months). Even in the P2 variants (caged and uncaged, TP2Ca and TP2WCa respectively) the results were highly satisfactory, since during the first 5 months of monitoring (established as critical ones as verified in the P1A trial, where survival dropped drastically by about 40%), in the two P2 variants, the survival has at least doubled for the same period. Moreover, translocated limpets from P1B and P2 (with cage) exceeded the survival of resident limpets from both the control-donor CD (88%) and control-receptor CR (92%), which was surprising for mechanically extracted limpets not translocated with their own substrata.
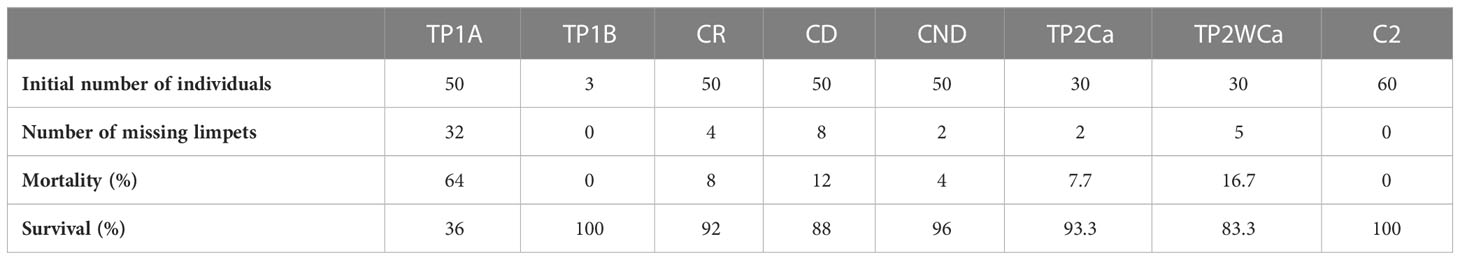
Table 2 Survival of P. ferruginea specimens relocated at the PJB in phase 1A (TP1A, Translocation phase 1A) and phase B (TP1B, Translocation phase 1B) from July 2021 to 31 October 2022, approximately 16 months, including controls (CR, receiver control; CD, donor control; CND, non-donor control), plus translocations in phase 2, with cage (TP2Ca) and without cage (TP2WCa) plus an added control for phase 2 (C2), from 15 May 2022 to October 2022, almost 6 months later.
Synthesis of failure-success methodological descriptors, essential as a basic protocol for further translocation trials
The results of the different methodologies applied in phase P1A and phases P1B and P2 are shown in Table 3, so that they can be replicated in further experiments.
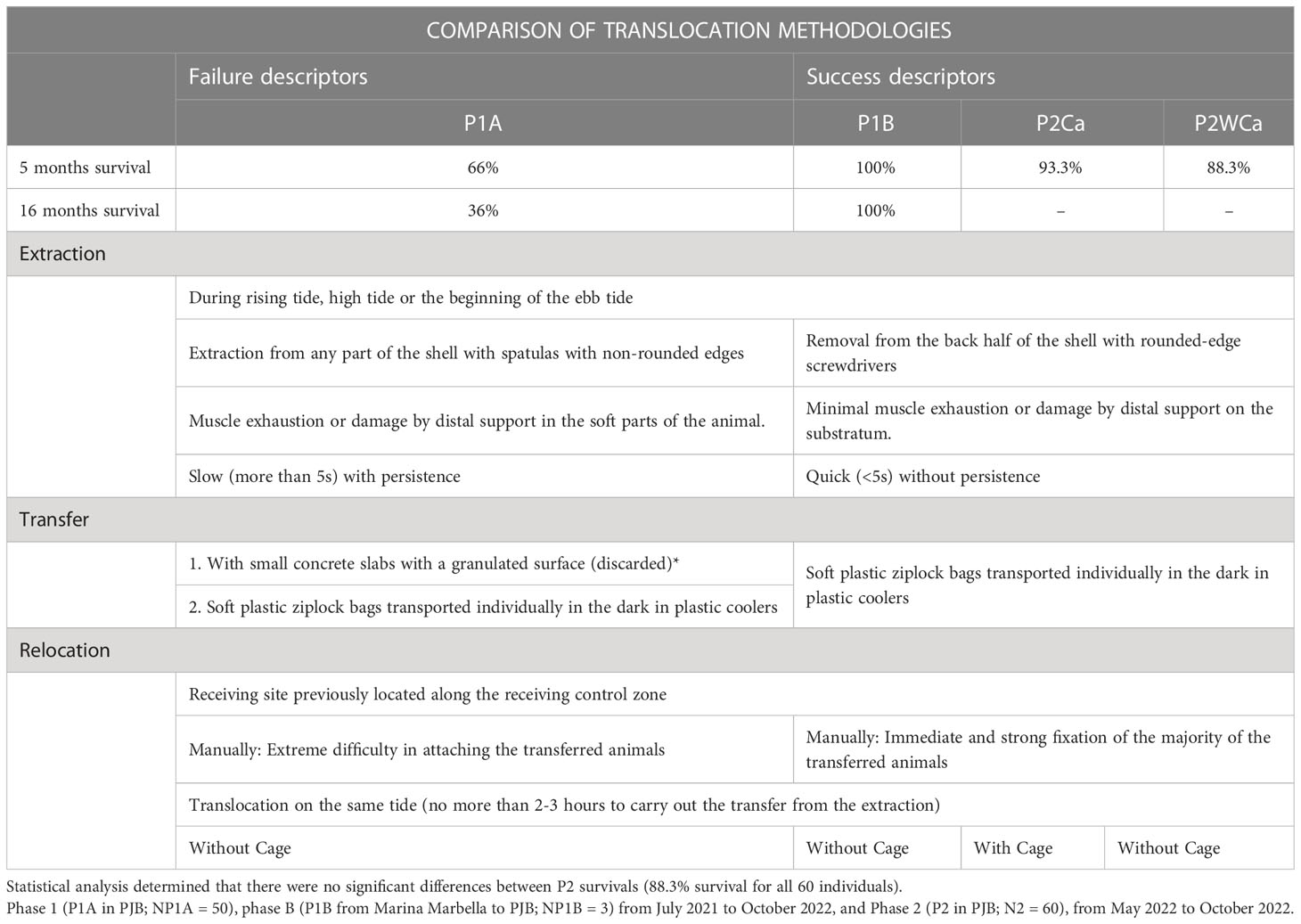
Table 3 Methodological results of failure (P1A) and success (P1B and P2), to be considered in further replication on translocation experiments of P. ferruginea.
Additional information to consider for future translocation trials
Rapid extraction depends on the limpets exhibiting a minimum of activity at rising tide, which is essential to leave sufficient room to introduce the extraction spatula. This does not always happen and, sometimes, a full tide takes place without the limpets leaving their home scars. This poses a serious problem for the viability of this type of action if the number of limpets to be removed is high and the time to carry out the action defined. For this reason, once the pilot project described here was completed, based on subsequent confirmatory tests and methodological extension of other projects currently underway (where this problem has been encountered), the extraction of specimens on fragments of substratum where the limpets were fixed was tested at low tide, using a hammer drill (at least 1250 watts of power) with sharp and flat chisels and/or rotaflex (at least 800 watts of power). Small pieces of substrata (personal observations) were extracted with the animals still attached, and these were translocated to receptor areas where the substratum pieces were fixed with epoxy marine cement. It was found that the ultra-fast vibration of the hammer chisel and rotaflex blade for a very short time (less than 5 minutes for substratum removal) did not affect the limpets and they remained attached once moved, until they voluntarily left their footprint at the recipient site with apparently no apparent sign of stress (personal observations).
Discussion
Advances in translocation methodology: Towards a sustainable resolution of future translocation actions
Given that, as explained above, the breakwaters of port infrastructures provide an advantageous artificial habitat for P. ferruginea to establish in high densities, this becomes a problem when the legitimate owners of such infrastructures must carry out emergency works or plan other works that may be authorized in which a relocation of specimens is required. So far, the history of direct and individual removal of specimens of the protected species has not been satisfactory, given the high mortalities recorded as shown in Table 4. Recent experiments on translocations of specimens on Artificial Inert Mobile Substrata (AIMS), carried out with 3D printing, have been conducted in Melilla (donor area) and La Línea (Algeciras Bay) (188 specimens, 15% survival, at two years; survival was higher in juveniles, <30mm, than in adults, >30mm) (see LIFE projects ReLife and REMoPaF; https://www.liferemopaf.org).
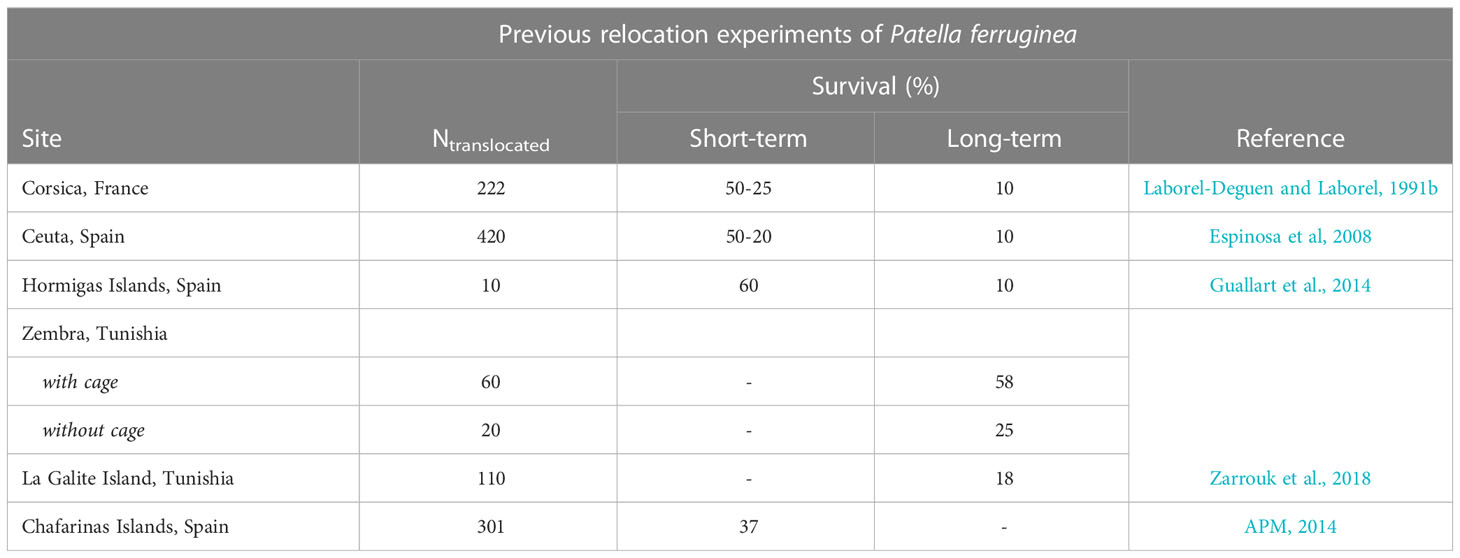
Table 4 Main data recollected from previous direct and individual removals of specimens of Patella ferruginea which proved to be unsuccessful.
In previous experimental trials, only one translocation experiment carried out in Gibraltar moving the limpets together with their home rocks without prior removal of specimens has been successful, (97 specimens, 87% survival at 10 months) (Fa et al., 2018), although a later experiment in the Chafarinas Islands, also using the same method, did not obtain good results (301 specimens, 37% survival, 3 months) (APM, 2014), despite the fact that the transfer of blocks was at a very short distance from the source area of the blocks. However, while the latter may be environmentally sustainable (which is debatable), it is not economically sustainable, given the high costs of the operations due to the logistics and resources required. These difficulties, already complex when dealing with boulders between 100 and 2000 kg, become further compounded when dealing with concrete tetrapods of 6 to 10 Tn- the case we are dealing with - which are almost impossible to reinsert into breakwaters with other tetrapods at the same tidal height and orientation as in their original location, to ensure that the limpets are spatially positioned in the strict upper midlittoral strip that is typical of the species.
The somewhat forced, slow and unregulated extraction of specimens to be translocated during the first experiment of the P1A pilot project presented here, proved to be the main cause of mortality, as it generated an important exhaustion of the foot musculature, which resulted in a relatively low survival (66%) after the first month following translocation. In contrast, in the subsequent experimental trials of this study (where the problem was methodologically corrected, implementing a fast extraction technique - less than 5 seconds - regulated and without injuries), survivorship in P1B and in P2 (both with cages), was 100%, and in P2 (without cages) it was over 90%, reflecting the clear and substantial benefits of the methodological improvements previously outlined. This was confirmed after 5 months, where in P1A a 36% survival was recorded, while for that same period in P2 survivorships of 93.3% (with cages) and 83.3% (without cages) were obtained; and in P1B, a 100% survival was obtained over 16 months of monitoring of this phase, prior to P2. These results could have direct application in future reintroduction projects of adult specimens in natural habitats of the Mediterranean where the species is already extinct or where its populations are at imminent risk of extinction.
Coastal infrastructures, marine biodiversity and P. ferruginea, towards their best integrated management
Bulleri and Chapman (2010) consider it crucial to increase our understanding of the ecological functioning of marine habitats created by urban infrastructure and to incorporate ecological criteria into coastal engineering to preserve biodiversity and Naylor et al. (2017) have proposed the new conservation strategy “integrated greening of grey infrastructure (IGGI)” aimed at enhancing the biodiversity of hard infrastructure that cannot be replaced by green solutions. This is currently reinforced by the emerging “Working with Nature (WwN)” philosophy (Vikolainen et al., 2014; Airoldi et al., 2021; Ruijgrok, 2021; Taljaard et al., 2021) adopted by institutions and companies owning coastal infrastructures. The increase of marinas in recent years to meet the growing demand for water sports (Di Franco et al., 2011), and of commercial harbors - or harbor extensions - due to increasing pressure from shipping (Hanson and Nicholls, 2020), as well as the prospects of global warming, sea level rise and related weather phenomena (such as increasingly frequent and extreme storms) (see Doney et al., 2012; Hanson and Nicholls, 2020), together paint a picture of increasing coastal infrastructures that make it necessary to promote more sustainable designs and materials, as well as greater integration with the local nature and the environment, through ecological research on artificial substrata and eco-engineering, which contribute to generating artificial structures and habitats that enhance biodiversity.
The breakwaters of the ports in the Alboran Sea are artificial habitats of great ecological interest, not only because they provide shelter or allow the growth of protected species, but also because among them there may be dense breeding aggregations of endangered species that must be properly monitored, protected and managed as Dendropoma lebeche or Astroides calycularis among others. (Ostalé-Valriberas et al., 2022). Therefore, P. ferruginea, having dense settlements in artificial habitats of port infrastructures subject to improvement and/or expansion works (in addition to being exposed to other impacts derived from them, related to water quality), requires a delicate integrated management plan to ensure its long-term survival as a species. To this end, it is essential to ensure that this species can be used as a reference to encourage and promote port infrastructures to be increasingly environmentally sustainable, since they often lead to environmental impacts widely identified in the scientific literature, e.g., by subsidiary pollution derived from the isolation of water bodies and the related anthropogenic pressure (Estacio et al., 1997; Seitz et al., 2006; Birch et al., 2008; Masiá et al., 2021), by influencing local change in coastal currents, sediment transport and increased coastal erosion (El-Asmar and White, 2002; Gittman et al., 2015; Critchley and Bishop, 2019; Vona et al., 2020), or by direct or indirect effects on part of the local biota decreasing its biodiversity (Connell and Glasby, 1999; Chapman, 2003; Guerra-García and García-Gómez, 2004; Moschella et al., 2005), in addition to constituting a risk factor for the introduction and dispersal of exotic species with invasive potential (Bulleri and Airoldi, 2005; Glasby et al., 2007).
Protected species established in coastal infrastructures can be very useful in monitoring ecosystem quality in this type of artificial environment. In this respect, P. ferruginea is a bioindicator species of environmental quality in coastal and harbor waters (Espinosa et al., 2006), and, like other indicator species established in harbor breakwaters, it can contribute to the control and valorization of the “ecological potential” contemplated in the European Marine Strategy Framework Directive (MSFD) (Directive 2008/56/EC) for “heavily modified water bodies” (harbor waters). For P. ferruginea this is an additional relevant aspect to ensure its protection and conservation and to revalue its ecological and environmental role as a “sentinel” species for negative environmental changes that may occur in the coastal system.
On the other hand, since entrance to ports is usually restricted, guarded and access to the base of the breakwaters is dangerous, these tend to contain large specimens of protected species, which contracts with the strong extractive pressure the same species will undergo in easily and publicly accessible natural coastal habitats in the surrounding areas. For this reason, and taking P. ferruginea as the main reference, Artificial Marine Micro-Reserves have been proposed in some exhaustively studied ports (García-Gómez et al., 2011; García-Gómez et al., 2015; Firth et al., 2016; Ostalé-Valriberas et al., 2022; Sempere-Valverde et al., 2023).
During the development of this work it has been found that the areas surrounding the PJB and the PDM -as well as those of other nearby ports with important densities of P. ferruginea- benefit of ‘reserve effect’ where the port breakwaters not only recruit larvae from the wild (a phenomenon accentuated by the high spatial heterogeneity of the artificial substrata of the dikes, which benefits the establishment of larvae) but that these recruits then thrive thanks to the restricted access usually in force, leading to the generation of reproductive clusters or subpopulations, with medium and large adults, which in turn enhance genetic connectivity. In this regard, Martins et al. (2010) and Espinosa et al. (2011) indicated that limpet recruitment is influenced by substratum heterogeneity. On the contrary, it is often the case that along the natural rocky areas surrounding the aforementioned ports and those of other ports of the Western Mediterranean coast the species is usually not present, or its presence is accidental and maintained through immigration (except in unfrequented, difficult to access, or protected coastal areas), due to the accessibility of its habitat and anthropogenic pressure. For P. ferruginea, this confirms the importance of the artificial habitats of port infrastructures in maintaining important densities of adult (breeding) specimens of the species, as well as their genetic connectivity.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Ethics statement
Ethical review and approval was not required for the study on animals in accordance with the local legislation and institutional requirements.
Author contributions
JG-G: conceptualization of the work and development the methodology. EO-V, JG-G: application of statistical, mathematical, computational, or other formal techniques to analyze or synthesize study data. EO-V, JG-G, AC-I: data analysis. JG-G, CG-O: participation in the sampled, extraction and relocation of specimens. AC-I, EO-V, RE, JLC: participation in the sampled, relocation of specimens. AC-I, RB-S: monitoring program. CG-O: designs of the two cage prototypes used and, installation and dismantling. JG-G, EO-V, AC-I, RE: writing review & editing. JG-G, EO-V, AC-I, DAF: final revision of the manuscript. JG-G: Management and coordination responsibility for the research activity planning and execution. JG-G: acquisition of the financial support for the project leading to this publication. All authors contributed to the article and approved the submitted version.
Acknowledgments
To the competent environmental authority, the Consejería de Sostenibilidad, Medio Ambiente y Economía Azul de la Junta de Andalucía, for granting the required authorization to carry out both phases of the pilot project whose results are presented here: and to Puerto José Banús, for having financed this study within the scope of the projects developed by the LBM of the US related to the ferruginean limpet in the Alboran Sea to contribute to its better knowledge, management and conservation. We also thank Juan Núñez Insausti for all the logistic support provided during the course of the pilot project, and Curro Sánchez, Estefanía Martín, Antonio Martín, Altai Pavón, Alfonso (PJB), Curro (PJB), Isabel Cárdenas, Liliana Olaya and their collaboration in the sampling campaigns and/or in some aspects related to the elaboration of the manuscript. Additionally, we express our deep gratitude to the technicians of the Consejería de Agricultura, Pesca y Desarrollo Rural, Eugenio Montes and Manuel Grondona for their valuable collaboration and advice on many different specialized topics related to the subject matter. We also thank MACVALNERA and TECNOAMBIENTE for the technical recommendations and technical tests carried out on the concrete tetrapods.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2023.1166937/full#supplementary-material
References
Acker J., Leptoukh G. (2007). Online analysis enhances use of NASA earth science data. EOS Trans. AGU 88 (2), 14–17. doi: 10.1029/2007EO020003
Airoldi L., Beck M. W., Firth L. B., Bugnot A. B., Steinberg P. D., Dafforn K. A. (2021). Emerging solution to return nature to the urban ocean. Ann. Rev. Mar. Sci. 13 (1), 445–477. doi: 10.1146/annurev-marine-032020-020015
APM (2014). Informe anual del traslado del plan de seguimiento y vigilancia ambiental. rehabilitación del embarcadero del puerto de chafarinas (Puerto de Melilla: Ministerio de Fomento).
Arroyo C., Moreno D. M., Mínguez A. B., de la Linde A., Menéndez J. M. R., de la Rosa J. D. L., et al. (2011). Trabajos de seguimiento de la lapa ferruginosa Patella ferruginea gmelin 1791 en andalucía en el marco de la estrategia nacional de conservación de la especie. Mediterránea: Serie Estudios Biológicos 22), 12–49. doi: 10.14198/MDTRRA2011.ESP.02
Aversano F. R. (1986). Esperimento di insediamento artificiale di Patella ferruginea gmelin 1791 nelle acque del golfo di arzachena (Sardegna settentrionale). Boll. Malacol. 22, 169–170.
Biagi V., Poli D. (1986). Considerazioni su una popolazione di Patella ferruginea gmelin 1971 per le acque del promontorio di piombino. Boll. Malacol. 22, 171–174.
Birch G. F., McCready S., Long E. R., Taylor S. S., Spyrakis G. (2008). Contaminant chemistry and toxicity of sediments in Sydney harbour, Australia: spatial extent and chemistry-toxicity relationships. Mar. Ecol. Prog. Ser. 363, 71–88. doi: 10.3354/meps07445
Botello G., Krug P. J. (2006). ‘Desperate larvae’ revisited: Age, energy and experience affect sensitivity to settlement cues in larvae of the gastropod alderia sp. Mar. Ecol. Progr. Ser. 312, 149–159. doi: 10.3354/meps312149
Boudouresque C. F., Laborel–Deguen F. (1986). Patella ferruginea, in le benthos marin de l’ile de zembra (Parc national, tunisie). Eds. Boudouresque C. F., Harmelin J. G., Grissac A.J. de (Marseille: GIS Posidonie Publishers), 105–110.
Boumaza S., Semroud S. (2001). Inventaire de la population de Patella ferruginea gmelin 1791 des iles habibas (ouest algerien). Rapport du Congres la Commission Internationale pour l’Exploration Scientifique la Mer Mediterranée 36, 361.
Broderip W. J., Sowerby I. G. B. (1829). Observations on new or interesting Mollusca contained, for the most part, in the museum of the zoological society. Zoological J. 4, 359–379.
Bulleri F., Airoldi L. (2005). Artificial marine structures facilitate the spread of a non-indigenous green alga, Codium fragile ssp. tomentosoides, in the north Adriatic Sea. J. Appl. Ecol. 42, 1063–1072. doi: 10.1111/j.1365-2664.2005.01096.x
Bulleri F., Chapman M. G. (2010). The introduction of coastal infrastructures as a driver of change in marine environments. J. Appl. Ecol. 47, 26–35. doi: 10.1111/j.1365-2664.2009.01751.x
CAGPyDS (JA). (2021). Informe sobre: El Proyecto de ampliación del puerto josé banús (Marbella, málaga),” in Programa de gestión sostenible del medio Marino andaluz, 12. Consejería de Agricultura, Ganadería, Pesca y Desarrollo Sostenible. Junta de Andalucía. Sevilla.
Carballo J. L., Yáñez B., Bautista-Guerrero E., García-Gómez J. C., Espinosa F., Tortolero-Langarica J. J., et al. (2020). Decimation of a population of the endangered species Scutellastra mexicana (Broderip and sowerby 1829) (Mollusca, Gastropoda) in the marías island (Eastern ocean pacific) biosphere reserve. Aquat. Conserv.: Mar. Freshw. 30 (1), 20–30. doi: 10.1002/aqc.3239
Casu M., Rivera-Ingraham G. A., Cossu P., Lai T., Sanna D., Dedola G. L., et al. (2011). Patterns of spatial genetic structuring in the endangered limpet Patella ferruginea: implications for the conservation of a Mediterranean endemic. Genetica 139, 1293–1308. doi: 10.1007/s10709-012-9631-3
Caton-Thompson G. (1946). The aterian industry: its place and significance in the paleolithic world. J.R. Anthropol. Inst. G.B. Irel. 76, 87–130. doi: 10.2307/2844512
Chapman M. G. (2003). Paucity of mobile species on constructed seawalls: effects of urbanization on biodiversity. Mar. Ecol. Progr. Ser. 264, 21–29. doi: 10.3354/meps264021
Connell S. D., Glasby T. M. (1999). Do urban structures influence local abundance and diversity of subtidal epibiota? a case study from Sydney harbour, Australia. Mar. Environ. Res. 47, 373–387. doi: 10.1016/S0141-1136(98)00126-3
Council Directive. (1992). On the conservation of natural habitats and of wild fauna and flora. OJ L 206(7), 50. doi: 10.1017/CBO9780511610851.039
Cristo B., Caronni S. (2008). Osservazioni sullo stato di conservazione di Patella ferruginea gmelin 1791, nelle vicinanze di capo ceraso (Golfo di olbia, sardegna nord–orientale). Biol. Mar. Mediterr. 15, 302–303.
Cristo B., Caronni S., Floris A. (2007). Osservazioni su Patella ferruginea gmelin 1791 (Mollusca, Gastropoda) nel golfo di olbia.). Biol. Mar. Mediterr. 14, 344–345.
Critchley L. P., Bishop M. J. (2019). Differences in soft-sediment infaunal communities between shorelines with and without seawalls. Estuaries Coasts 42, 1127–1137. doi: 10.1007/s12237-019-00527-z
Curini–Galletti M. (1979). Ritrovamento di una Patella ferruginea. Notiz. C.I.S.M.A. Roma 1 (1), 53–54.
Di Franco A., Graziano M., Franzitta G., Felline S., Chemello R., Milazzo M. (2011). Do small marinas drive habitat specific impacts? a case study from Mediterranean Sea. Mar. pollut. Bull. 62 (5), 926–933. doi: 10.1016/j.marpolbul.2011.02.053
Doneddu M., Manunza B. (1992). Valutazione dell’impatto antropico relativo alla balneazione estiva su una populazione di Patella ferruginea gmeli Del litorale di aglientu. Boll. Malacol. 28, 161–168.
Doney S. C., Ruckelshaus M., Emmett Duffy J., Barry J. P., Chan F., English C. A., et al. (2012). Climate change impacts on marine ecosystems. Ann. Rev. Mar. Sci. 4, 11–37. doi: 10.1146/annurev-marine-041911-111611
El-Asmar H. M., White K. (2002). Changes in coastal sediment transport processes due to construction of new damietta harbour, Nile delta, Egypt. Coast. Eng. 46 (2), 127–138. doi: 10.1016/S0378-3839(02)00068-6
Espinosa F. (2006). Caracterización biológica del molusco protegido Patella ferruginea gmelin 1791 (Gastropoda: Patellidae): bases para su gestión y conservación (Univ. of Seville).
Espinosa F. (2009). Populational status of the endangered mollusk Patella ferruginea gmelin 1791 (Gastropoda: Patellidae) on Algerian islands (SW Mediterranean). Anim. Biodivers Conserv. 32, 19–28. doi: 10.32800/abc.2009.32.0019
Espinosa F., González A. R., Maestre M. J., Fa D., Guerra-García J. M., García- Gómez J. C. (2008). Responses of the endangered limpet Patella ferruginea to reintroduction under different environmental conditions: survival, growth rates and life-history. Ital. J. Zool. 75 (4), 371–384. doi: 10.1080/11250000801887740
Espinosa F., Guerra-García J. M., FA D., García-Gomez J. C. (2006). Aspects of reproduction and their implications for the conservation of the endangered limpet, Patella ferruginea. Invertebrate Reprod. Dev. 49 (1-2), 85–92. doi: 10.1080/07924259.2006.9652197
Espinosa F., Guerra-García J. M., García-Gómez J. C. (2007). Sewage pollution and extinction risk: an endangered limpet as a bioindicator? Biodiv. Conserv. 16 (2), 377–397. doi: 10.1007/s10531-005-3014-3
Espinosa F., Ozawa T. (2006). Population genetics of the endangered limpet Patella ferruginea (Gastropoda: Patellidae): taxonomic, conservation and evolutionary considerations. J. Zool. Syst. Evol. Res. 44 (1), 8–16. doi: 10.1111/j.1439-0469.2005.00348.x
Espinosa F., Rivera-Ingraham G. A. (2017). Biological conservation of giant limpets: the implications of large size. Adv. Mar. Biol. 76, 105–155. doi: 10.1016/bs.amb.2016.10.002
Espinosa F., Rivera–Ingraham G. A., Fa D. A., García–Gómez J. C. (2009). Effect of human pressure on population size structures of the endangered ferruginean limpet: towards future management measures. J. Coast. Res. 25, 857–863. doi: 10.2112/08-1005.1
Espinosa F., Rivera-Ingraham G., García-Gómez J. C. (2011). Influence of habitat structure and nature of substratum on limpet recruitment: Conservation implications for endangered species. Estuar. Coast. Shelf Sci. 94 (2), 164–171. doi: 10.1016/j.ecss.2011.06.005
Espinosa F., Rivera-Ingraham G. A., Maestre M., González A. R., Bazairi H., García-Gómez J. C. (2014). Updated global distribution of the threatened marine limpet Patella ferruginea (Gastropoda: Patellidae): an example of biodiversity loss in the Mediterranean. Oryx 48 (2), 266–275. doi: 10.1017/S0030605312000580
Estacio F. J., García-Adiego E. M., Fa D. A., García-Gómez J. C., Daza J. L., Hortas F., et al. (1997). Ecological analysis in a polluted area of algeciras bay (Southern spain): external ‘versus’ internal outfalls and environmental implications. Mar. Poll. Bull. 34 (10), 780–793. doi: 10.1016/S0025-326X(97)00046-5
Fa D. A., Finlayson G., Sempere-Valverde J., García-Gómez J. C. (2018). ‘Impossible’re-introduction of the endangered limpet Patella ferruginea gmelin 1791? progress in resolving translocation mortality. Endanger. Species Res. 37, 219–232. doi: 10.3354/esr00921
Fenberg P. B., Roy K. (2012). Anthropogenic harvesting pressure and changes in life history: insights from a rocky intertidal limpet. Am. Nat. 180 (2), 200–210. doi: 10.1086/666613
Firth L. B., Knights A. M., Bridger D., Evans A. J., Mieszkowska N., Moore P. J., et al. (2016). Ocean sprawl: Challenges and opportunities for biodiversity management in a changing world. Oceanogr. Mar. Biol. 54, 193–269. doi: 10.1201/9781315368597-5
Freitas D., Arenas F., Vale C. G., Pinto I. S., Borges D. (2023). Warning of warming limpets: sea temperature effects upon intertidal rocky assemblages. J. Mar. Biol. Assoc. United Kingdom 103, e7. doi: 10.1017/S0025315422001096
Frenkiel L. (1975). Contribution à l’étude des cycles de reproduction des patellidae en algérie. Pubblicacione la Stazzione Zoologica di Napoli 39, 153–189.
García-Gómez J. C. (2015). A guide on environmental monitoring of rocky seabeds in Mediterranean marine protected areas and surrounding zones (Tunis: Marine Biology Laboratory, Department of Zoology, Faculty of Biology, University of Seville. R+D+I Biological Research Area, Seville Aquarium. Ed. RAC/ SPA— MedMPAnet Project). Available at: http://www.rac-spa.org/sites/default/files/doc_medmpanet/final_docs_regional/78.guide_suivi_amp_med_en.pdf.
García-Gómez J. C., Guerra-García J. M., Espinosa F., Maestre M. J., Rivera-Ingraham G., Fa D., et al. (2015). Artificial marine micro-reserves networks (AMMRNs): an innovative approach to conserve marine littoral biodiversity and protect endangered species. Mar. Ecol. 36, 259–277. doi: 10.1111/maec.12167
García-Gómez J. C., López-Fé C. M., Espinosa F., Guerra-García J. M., Rivera-Ingraham G. A. (2011). Marine artificial micro-reserves: a possibility for the conservation of endangered species living on artificial substrata. Mar. Ecol. 32, 6–14. doi: 10.1111/j.1439-0485.2010.00409.x
García-Gómez J. C., Magariño S. (2010). Bucear en el último confín de Europa, la isla de tarifa. instituto de estudios campogibraltareños. Serie Cienc. Naturales 35, 350.
Gittman R. K., Joel F. F., Popowich A. M., Keller D. A., Bruno J. F., Currin C. A., et al. (2015). Engineering away our natural defenses: An analysis of shoreline hardening in the US. Front. Ecol. Environ. 13, 301–307. doi: 10.1890/150065
Glasby T. M., Connell S., Holloway M. G., Hewitt C. L. (2007). Nonindigenous biota on artificial structures: Could habitat creation facilitate biological invasions? Mar. Biol. 151, 887–895. doi: 10.1007/s00227-006-0552-5
González J. A., Paredes P., Enrique C., Calzado P., Bueno I. (2015). Patella ferruginea, patrimonio del litoral melillense: biología, ecología y conservación. GEEPP Ediciones, 157.
González-García J. A., Bueno del Campo I., García Peña H., Bazairi H. (2006). “Las poblaciones de Patella ferruginea gmeli (Mollusca, Gastropoda, patellidae) en los acantilados de melilla y tres forcas,” in XIV simposio ibérico de estudios de biología Marina. Eds. Ballesteros M., Palacín C., Turon X.(Barcelona), 152–153.
Guallart J. (2006). Estado de patella ferruginea en las islas chafarinas y estudios previos para la traslocación de ejemplares (Madrid: Technical scientific report (unpublished) for Organismo Autónomo de Parque Nacionales, Ministerio de Medio Ambiente).
Guallart J., Templado J., Acevedo I., Calvo M., Machordom. A., Luque Á.A., et al. (2014). Informe de la reintroducción de ejemplares de la lapa Patella ferruginea (catalogada “en peligro de extinción”) en las islas hormigas (Murcia). Unpublished, 26.
Guerra-García J. M., Corzo J., Espinosa F., Fa D. A., García-Gómez J. C. (2004a). Extinction risk and harbours as marine reserves. J. Moll. Stud. 70, 96–98. doi: 10.1093/mollus/70.1.96
Guerra-García J. M., Corzo J., Espinosa F., García-Gómez J. C. (2004b). Assessing habitat use of the endangered marine mollusc Patella ferruginea (Gastropoda, patellidae) in northern Africa: preliminary results and implications for conservation. Biol. Conserv. 116, 319–326. doi: 10.1016/S0006-3207(03)00201-5
Guerra-García J. M., García-Gómez J. C. (2004). Crustacean assemblages and sediment pollution in an exceptional case study: a harbour with two opposing entrances. Crustaceana 77, 353–370. doi: 10.1163/1568540041181538
Haedrich R. L., Barnes S. M. (1997). Changes over time of the size structure in an exploited shelf fish community. Fish. Res. 31 (3), 229–239. doi: 10.1016/S0165-7836(97)00023-4
Hanson S. E., Nicholls R. J. (2020). Demand for ports to 2050: Climate policy, growing trade and the impacts of sea-level rise. Earth's Future 8, e2020EF001543. doi: 10.1029/2020EF001543
Henriques P., Delgado J., Sousa R., Ray S. (2017). Patellid limpets: an overview of the biology and conservation of keystone species of the rocky shores. Organismal Mol. malacology, 71–95. doi: 10.5772/67862
Hind K. R., Saunders G. W. (2013). A molecular phylogenetic study of the tribe Corallineae (Corallinales, Rhodophyta) with an assessment of genus-level taxonomic features and descriptions of novel genera. J. Phycol. 49 (1), 103–114.
Jager K. J., Van Dijk P. C., Zoccali C., Dekker F. W. (2008). The analysis of survival data: the Kaplan–Meier method. Kidney Int. 74 (5), 560–565. doi: 10.1038/ki.2008.217
Keen A. M. (1971). Sea Shells of tropical West america. marine mollusks from Baja California to Peru. 2nd ed (Stanford, California: Stanford University Press).
Laborel-Deguen F., Laborel J. (1991a). ““Statut de Patella ferruginea gmelin en méditerranée,”,” in Les Espèces marines à protéger en méditerranée. Eds. Boudouresque. C. F., Avon. M., Gravez V. (Marseille: GIS Posidonie Publishers), 91–103.
Laborel-Deguen F., Laborel J. (1991b). “Une tentative de réintroduction de Patella ferruginea gmel. (Gasteropoda) dans le parc national de port-cros (Var, France),” in Les Espèces marines à protéger en méditerranée, boudouresque. Eds. Boudouresque C. F., Avon. M., Gravez V. (Marseille, France: GIS Posidonie publ.), 129–132.
LIFE REMoPaF Project (2017-2021). LIFE-15/NAT/ES/000987-REMoPaF (UE). Available at: https://www.liferemopaf.org.
Maestre Delgado M. J., Espinosa F., Fa D., Pavón A., García-Gómez J. C. (2018). Estado de la población de Patella ferruginea en el área del estrecho de Gibraltar. Almoraima. Rev. Estudios Campogibraltareños 49, 115–130).
Mantel N. (1966). Evaluation of survival data and two new rank order statistics arising in its consideration. Cancer Chemother. Rep. 50, 163–170.
Marra S., Coppa S., Camedda A., Massaro G., de Lucia G. A. (2017). The exploitation of limpets in a Mediterranean marine protected area: assessing the effectiveness of protection in the intertidal zone. Mediterr. Mar. Sci. 18 (3), 406–423. doi: 10.12681/mms.2087
Martins G. M., Thompson R. C., Neto A. I., Hawkins S. J., Jenkins S. R. (2010). Enhancing stocks of the exploited limpet patella candei d’Orbigny via modifications in coastal engineering. Biol. Conserv. 143 (1), 203–211. doi: 10.1016/j.biocon.2009.10.004
Masiá P., Ardura A., Gaitan M., Gerber S., Rayon-Vina F., Garcia-Vazquez E. (2021). Maritime ports and beach management as sources of coastal macro-, meso-, and microplastic pollution. Environ. Sci. Poll. Res. 28, 30722–30731. doi: 10.1007/s11356-021-12821-0.org/10.3354/meps326011
MMAMRM (2008). “Estrategia para la conservación de la lapa ferrugínea (Patella ferruginea) en españa,” in Ministerio de medio ambiente y medio rural y Marino (MMAMRM), 49, Madrid.
Moreno D. (2004). “Prosobranquios y heterobranquios. fauna andaluza, in proyecto andalucía. naturaleza XIV, zoología II, los animales esquizocelomados,” in Moluscos, anélidos, sipuncúlidos y equiúridos. Eds. Tinaut J. A., Pascual F. (Sevilla: Publicaciones Comunitarias, Grupo Hércules), 79–114.
Moschella P. S., Abbiati M., Aberg P., Airoldi L., Anderson J. M., Bacchiocchi F., et al. (2005). Low-crested coastal defence structures as artificial habitats for marine life: Using ecological criteria in design. Coast. Eng. 52 (10–11), 1053–1071. doi: 10.1016/j.coastaleng.2005.09.014
NASA Goddard Space Flight Center, Ocean Ecology Laboratory, Ocean Biology Processing Group (2018). Moderate-resolution imaging spectroradiometer (MODIS) aqua chlorophyll data (Greenbelt, MD, USA: NASA OB. DAAC). doi: 10.5067/AQUA/MODIS/L3M/CHL/2018.07/20/2022
Naylor L. A., MacArthur M., Hampshire S., Bostock K., Coombes M. A., Hansom J. D., et al. (2017). Rock armour for birds and their prey: ecological enhancement of coastal engineering. Proc. Institution Civil Engineers-Maritime Eng. 170 (2), 67–82. doi: 10.1680/jmaen.2016.28
Ostalé-Valriberas E., Sempere-Valverde J., Pavón-Paneque A., Coppa S., Espinosa F., García-Gómez J. C. (2022). Artificial marine micro-reserves as a new ecosystem-based management tool for marine conservation: The case of Patella ferruginea (Gastropoda, patellidae), one of the most endangered marine invertebrates of the Mediterranean. Mar. Policy 136, 104917. doi: 10.1016/j.marpol.2021.104917
Paracuellos M., Nevado J. C., Moreno D., Giménez A., Alesina J. J. (2003). Conservational status and demographic characteristics of Patella ferrguinea gmelin 1791 (Mollusca, Gastropoda) on the alboran island (Western Mediterranean). Anim. Biodiv. Conserv. 26, 29–37.
Peto R., Peto J. (1972). Asymptotically efficient rank invariant test procedures. J. R. Stat. Soc Ser. A 135 (2), 185–198. doi: 10.2307/2344317
Pombo O. A., Escofet A. (1996). Effect of exploitation on the limpet Lottia gigantea: a field study in Baja California (USA). Pac. Sci. 50, 393–403. Available at: http://hdl.handle.net/10125/2914.
Porcheddu A., Milella I. (1991). “Aperçu sur l’ecologie et sur la distribution de Patella ferruginea (L.) gmelin 1791 en mers italiennes,” in Les Espèces marines à protéger en méditerranée. Eds. Boudouresque C. F., Avon M., Gravez V. (Marseille: GIS Posidonie Publishers), 119–128.
Ramos M. A. (1998). Implementing the habitats directive for mollusc species in Spain. J. Conchol. Special Publication. 2, 125–132.
Rivera–Ingraham G. A., Espinosa F., García–Gómez J. C. (2011a). Conservation status and updated census of Patella ferruginea (Gastropoda, patellidae) in ceuta: distribution patterns and new evidence of the effects of environmental parameters on population structure. Anim. Biodiv. Conserv. 34 (1), 83–99.
Rivera-Ingraham G. A., Espinosa F., García-Gómez J. C. (2011b). Environmentally mediated sex change in the endangered limpet Patella ferruginea (Gastropoda: Patellidae). J. Moll. Stud. 77 (3), 226–231. doi: 10.1093/mollus/eyr007
Rochet M. J., Trenkel V. M. (2003). Which community indicators can measure the impact of fishing? a review and proposals. Can. J. Fish. Aquat. Sci. 60, 86–99. doi: 10.1139/f02-164
Ruijgrok E. (2021). Natural solutions versus technical solutions: How ecosystem benefits can make a difference in public decisions. Res. Urban. Ser. 7, 261–282. doi: 10.47982/rius.7.137
Seitz R. D., Lipcius R. N., Olmstead N. H., Seebo M. S., Lambert D. M. (2006). Influence of shallow-water habitats and shoreline development on abundance, biomass, and diversity of benthic prey and predators in Chesapeake bay. Mar. Ecol. Progr. Ser. 326, 11–27. doi: 10.3354/meps326011
Sempere-Valverde J., Guerra-García J. M., García-Gómez J. C., Espinosa F. (2023). “Coastal urbanization, an issue for marine conservation,” in Coastal habitat conservation. Ed. Espinosa F. (Laboratorio biología Marina, Universidad de Sevilla, Sevilla, Spain: Elsevier-Academic Press), 41–79.
Stel V. S., Dekker F. W., Tripepi G., Zoccali C., Jager K. J. (2011). Survival analysis I: the Kaplan-Meier method. Nephron Clin. Pract. 119 (1), c83–c88. doi: 10.1159/000324758
Taljaard S., Slinger J. H., Arabi S., Weerts S. P., Vreugdenhil H. (2021). The natural environment in port development: A ‘green handbrake’ or an equal partner? Ocean Coast. Manage. 199, 105390. doi: 10.1016/j.ocecoaman.2020.105390
Templado J. (2001). “Patella ferruginea (Gmelin 1791),” in Los Invertebrados no insectos de la directiva hábitats en españa. Eds. Ramos M. A., Bragado D., Fernández J. (Madrid: Ediciones Serie Técnica, Organismo Autónomo Parques Nacionales, Dirección General de Conservación de la Naturaleza, Ministerio de Medio Ambiente), 41–49.
Templado J., Calvo M., Moreno D., Flores A., Conde F., Abad R., et al. (2006). Flora y fauna de la reserva Marina de pesca de la isla de alborán (Madrid, Spain: Secretaría General de Pesca Martítima, MAPA).
Vikolainen V., Bressers H., Lulofs K. (2014). A shift toward building with nature in the dredging and port development industries: Managerial implications for projects in or near natura 2000 areas. Environ. Manage. 54 (1), 3–13. doi: 10.1016/10.1007/s00267-014-0285-z
Vona I., Gray M. W., Nardin W. (2020). The impact of submerged breakwaters on sediment distribution along marsh boundaries. Water 12 (4), 1016. doi: 10.3390/w12041016
Keywords: translocation adults, survival, advances methodology, conservation, endangered Patellogastropoda
Citation: García-Gómez JC, Cid-Iturbe A, Ostalé-Valriberas E, Espada R, Carballo JL, Fa DA, García-Olaya C and Blanca-Sújar R (2023) Advances in the management and translocation methodology of the endangered mollusk Patella ferruginea in artificial habitats of port infrastructures: implications for its conservation. Front. Mar. Sci. 10:1166937. doi: 10.3389/fmars.2023.1166937
Received: 15 February 2023; Accepted: 28 March 2023;
Published: 20 April 2023.
Edited by:
Alan Hodgson, Rhodes University, South AfricaReviewed by:
Phillip Fenberg, University of Southampton, United KingdomRafael Sarda, Spanish National Research Council (CSIC), Spain
Copyright © 2023 García-Gómez, Cid-Iturbe, Ostalé-Valriberas, Espada, Carballo, Fa, García-Olaya and Blanca-Sújar. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Enrique Ostalé-Valriberas, ZW5yaXF1ZW9zdGFsZXZhbHJpYmVyYXNAZ21haWwuY29t; José Carlos García-Gómez, amNnYXJjaWFAdXMuZXM=