 Inka Anglade1*
Inka Anglade1* Thomas H. Dahl1
Thomas H. Dahl1 Bjørn S. B. Kristensen1
Bjørn S. B. Kristensen1 Andreas Hagemann2
Andreas Hagemann2 Arne M. Malzahn2†
Arne M. Malzahn2† Kjell Inge Reitan1
Kjell Inge Reitan1- 1Department of Biology, Norwegian University of Science and Technology, Trondheim, Norway
- 2Department of Fisheries and New Biomarine Industry, SINTEF Ocean, Trondheim, Norway
Introduction: Sustainable growth of the salmon aquaculture industry is highly dependent on the availability of new feed resources as well as improved management and utilization of waste streams from production. Polychaetes are capable of recapturing and recycling valuable biomolecules contained in aquaculture sludge and at the same time constitute a possible resource for aquafeeds.
Material and methods: In this study, the potential of Hediste diversicolor (OF Müller, 1776) to recover nutrients and energy contained aquaculture sludge was evaluated. Hereby, the effect of two different diets, smolt and post-smolt sludge, and different feed levels on polychaete growth, and biochemical and elemental composition was assessed.
Results and discussion: Survival ranged from 80-90%, independent of treatment. Specific growth rates at the highest feed levels were significantly higher than at the lowest feed levels and growth was not affected by the type of diet. Similarly, total lipid and total fatty acid content of polychaetes were positively correlated with feed level while the type of diet had a limited effect. Fatty acid composition of polychaetes reflected feed levels: in both diet groups, polychaetes at lower feed levels showed a higher percentage of saturated fatty acids; monounsaturated fatty acids increased with increased feed supply for smolt sludge-fed group and polychaetes fed post-smolt sludge showed increased polyunsaturated fatty acid content with increasing feed levels. Neither diet nor feed levels affected protein content and amino acid composition of polychaetes. In conclusion, H. diversicolor could be successfully cultivated on both smolt and post-smolt sludge and was rich in proteins and lipids as well as essential amino and fatty acids. The species can be considered a promising candidate for recycling valuable nutrients from different types of side streams salmon aquaculture.
1 Introduction
The global population crossed the threshold of 8 billion in 2022 and is predicted to reach 10 billion by the end of this century (United Nations, 2022). Population growth comes with many challenges, food supply being one of them, and estimates show that there will be a food gap of 56% in 2050 (Searchinger et al., 2019). Aquaculture already plays a vital role in food and protein supply and its importance will increase with agricultural production growing only at a slow rate (Colgrave et al., 2021; Otsuka and Fan, 2021); farmed species as a proportion of global fisheries and aquaculture are projected to grow from 49% in 2020 to 53% in 2030 (FAO, 2022). The Norwegian aquaculture sector has grown continuously since the 1970s and Norway is the world’s largest producer of farmed Atlantic salmon (Salmo salar L.) with over 1.5 million tonnes of salmon produced in 2021 (Shahbandeh, 2020; Fiskeridirektoratet, 2022). In recent years, production has been stagnating due to issues with parasitic infections and environmental concerns. Access to high-quality ingredients for production of aquafeeds could further restrict growth as 94% of assessed marine fish stocks are overexploited or at maximum sustainable capture capacity, hence future supply of fish meal and fish oil will not match demand (Olsen, 2011; FAO, 2022). Next to solving resource supply of feed ingredients, adequate handling of waste streams is imperative for sustainable growth of the Norwegian salmon industry. Both land- and sea-based production of Atlantic salmon generate an immense discharge of dissolved and particulate nutrients (Wang et al., 2012; Meriac, 2019). In 2022, 1.9 million tonnes of salmon feed were used in Norway (Fiskeridirektoratet, 2023). According to Aas and Åsgård (2017), volumes of aquaculture sludge in total salmon production are equivalent to 35-40% of the feed input (WW), giving an aquaculture sludge production of over 700 000 tonnes in 2022. Currently, mostly sludge from land-based smolt and post-smolt production facilities is collected, and land-based sludge makes up roughly 2% of the total sludge production (Aas and Åsgård, 2017). Based on these numbers, we estimate that ca. 15 000 tonnes of aquaculture sludge were collected in 2022. Due to a recent regulation (Meld. St. 16 (2014-2015)) that allows for cultivation of smolt and post-smolt to larger sizes in land-based systems, output of aquaculture sludge from land-based system will increase significantly in the years to come. At the same time, there are incentives to collect aquaculture sludge from sea-based facilities using closed containment or pump systems (Regjeringen, 2015; Olaussen, 2018), meaning that additional volumes of sludge will need to be handled. Previous studies have shown that aquaculture sludge from land-based salmon production is rich in nutrients and can thus constitute an appropriate resource for extractive species. The polychaete Hediste diversicolor (OF Müller, 1776) can successfully grow on aquaculture sludge from different fish species as a sole resource, and moreover, upcycle lipids and fatty acids (FAs) contained in the sludge (Fidalgo e Costa et al., 2000; Jerónimo et al., 2020; Malzahn et al., 2023). Brown et al. (2011) reported that closely related Alitta (Nereis) virens fed waste feed and feces from Atlantic halibut (Hippoglossus hippoglossus L.) was highly effective in utilizing waste streams and had a higher lipid content than the control group. Wang et al. (2019a; 2019b) successfully cultivated H. diversicolor on salmon aquaculture sludge and solid biogas digestate and the polychaetes were efficient in recycling organic nutrients contained in their diets. Overall, both species were found to be rich in marine lipids and FAs, as well as amino acids (AAs) and protein which could make them suitable candidates to be used in aquafeeds (Brown et al., 2011; Wang et al., 2019a; Wang et al., 2019b).
Feeds for salmon smolt and post-smolt differ; typical smolt feed contains 220-240 mg lipids g-1 DW and 425-480 mg protein g-1 DW, while post-smolt feed has a higher lipid content of 315-355 mg g-1 DW and lower protein content of 360-400 mg g-1 DW (BioMar, 2022; EWOS, 2022a; EWOS, 2022b). Accordingly, the lipid and protein content of sludge generated in land-based smolt and post-smolt production can be expected be different in terms of nutritional value and biochemical composition. Further, salt content in the sludges can be different as smolt production uses freshwater while post-smolt are cultivated in brackish or saline water. So far, it has not been investigated how such differences affect the performance of extractive polychaete species. This study aimed to investigate the suitability of using smolt and post-smolt sludge for cultivation of H. diversicolor and evaluate possible effects of the different diets on the polychaetes’ biochemical and elemental composition. Moreover, the goal was to assess the effect of different feed levels on growth, survival, and biochemical and elemental composition of H. diversicolor. Hereby, the hypotheses were that 1) H. diversicolor will perform similarly well on smolt and post-smolt sludge, 2) increased feed availability will result in increased growth, and 3) increased feed availability will affect biochemical composition with regards to lipid and FAs.
2 Material and methods
2.1 Collection and preparation of Hediste diversicolor
Individuals of H. diversicolor used in the experiment were collected from sediment at low tide from the intertidal zone of the Trondheimsfjord in Buvika (63°18’49.2”N 10°10’48.7”E). Collection took place before visible sexual maturation of individuals and prior to the trial which was conducted in November 2019. Thereafter, they were transferred to flow-through holding tanks (52 × 36 × 18 cm L x W x H) with sufficient water flow, containing a 15 cm thick layer of sand. They were acclimated in the tanks for four weeks at 16L:8D and 16°C and fed commercial fish feed (GEMMA DIAMOND 1.0, Skretting AS, Norway) during that period.
2.2 Experimental setup
H. diversicolor was cultivated in two fully automated XR3 cultivation racks operated as flow-through systems (Aquatic habitats, Pentair plc, USA) with 20 cultivation tanks (16-L, 47 x 26 x 21 cm L x W x H) in each rack, of which 16 were used per unit. Tanks were stocked with 15 polychaetes each, yielding a density of 140 ind. m-2. At the start of the trial, unscathed polychaetes were retrieved from the holding tanks, cleaned from sediment, and placed in cups filled with clean seawater to allow for gut evacuation for ≥ 2 h. Subsequently, adherent water was removed and the polychaetes were weighed (balance: Science Education, SE622, VWR Italy), the average initial weight was 250 ± 48 mg (WW). Individuals were then placed into the cultivation tanks which had been filled with a ca. 8 cm thick layer of sandbox sand, leaving a water volume of 8 L. Temperature, salinity, pH and dissolved oxygen levels (% DO) in the tanks were controlled three times per week. Seawater temperature (16.3 ± 0.2°C) and salinity (34.3 ± 0.2 ppt) were set and kept constant by the system. The flow rate was set to 8 L d-1 (100% water exchange daily) to ensure an oxygen saturation ≥ 95%, and the tanks were aerated through air stones placed on top of the sediment. The photoperiod for the room lighting was set to 16L:8D. Dead individuals that were found on the sediment surface throughout the experiment were removed. Polychaetes H. diversicolor were fed two different types of sludge (diets) at four different feeding levels per diet with four replicate tanks each (2x4x4). Polychaetes in system one received smolt sludge (from Lerøy AS, Belsvik), those in system two were fed post-smolt sludge (from LetSea AS, Dønna). Both types of sludge were centrifuged at 3000 rpm for 10 minutes, the supernatant discarded, and the remaining sludge frozen at -20°C to be thawed a day prior to feeding. Feed levels were calculated based on estimated nitrogen content of diets and polychaetes as described by Malzahn et al. (2023). Wang et al. (2019b) established a nitrogen content of H. diversicolor of 95 mg g-1 DW. Accordingly, the different groups in this trial received feed doses equivalent to different percentages of the nitrogen content of polychaetes. Polychaetes fed smolt sludge (S) received 6% of polychaete N (S6), 12% N (S12), 24% N (S24), 45% N (S45), and polychaetes fed post-smolt (PS) sludge, received 6% N (PS6), 13% N (PS13), 25% N (PS25), 47% N (PS47). The polychaetes were fed every second day for 30 days.
At the end of the experiment, polychaetes were retrieved from the tanks and placed in clean seawater for ≥ 2 h to again allow for gut evacuation. They were rinsed and adherent water was removed by placing the worms on paper tissue; afterwards they were weighed, freeze-dried, homogenized, and stored in liquid nitrogen at -80°C for later analyses.
2.3 Survival and growth
Survival was calculated using:
where N0 and Nt is the number of polychaetes (-) at the beginning and the end of the experiment, respectively.
Specific growth rate SGR (d-1) based on weight (WW) of polychaetes H. diversicolor was calculated using the formula:
where Wt is the final mean biomass (mg WW), W0 is the initial mean biomass (mg WW), and t is the duration of the trial (d). The Von Bertalanffy growth function (Von Bertalanffy, 1938) was used to describe the function of specific growth rates:
where SGRmax (d-1) is the mean value of the highest observed SGRs in our trial, k (-) is the growth coefficient, Nfed is nitrogen fed per day, and SGR0 is the value of Nfed for which the SGR is zero.
2.4 Biochemical and elemental analyses
Total lipid content, FA content and FA composition, protein content, total AA content and AA composition, elemental composition, and ash were analyzed for polychaetes and sludge. Ash content was determined by combustion of samples in a muffle furnace at 450°C for 5h. Total lipids were extracted with dichlormethane (CH2Cl2) according to Bligh and Dyer (1959) and subsequently determined gravimetrically. Following Metcalfe et al. (1966), FAs were esterified to fatty acid methyl esters with BF3-methanol and subsequently analyzed using a gas chromatograph (7890B GC, Agilent Technologies, USA) with helium carrier, a WCOT fused-silica capillary column coated with 28 CP-wax 52CB (Holger CP7713). AAs were analyzed following Šližytė et al. (2017) where samples were hydrolyzed for 24h at 110°C using 6 M HCL containing 0.4% mercaptoethanol and then filtered (Whatman glass microfiber filters, grade GF/C, 47 mm). The pH was adjusted to 2.2 before being separated by a high-performance liquid chromatography (HPLC) system (Agilent Infinity 1260, Agilent Technologies, USA) coupled to an online post-column derivatization module (Pinnacle PCX, Pickering laboratories, USA). Protein content was calculated by addition of water-free AAs. Elemental analysis of pooled replicates for each diet and feed level was conducted in accordance with NS-EN ISO 17294-2:2016 by SINTEF Norlab.
2.5 Statistical analysis
All basic statistical analyses were conducted using Sigmaplot for Windows Version 14.0 (Systat Software, Inc., USA). Minitab® 21.1 (Minitab, LLC) was used for principal component analysis (PCA) of FA and AA composition of polychaetes.
Normal distribution of data was tested using Shapiro-Wilk normality tests, homogeneity of variance was evaluated by the Brown-Forsythe test. For data that passed model control, a Welch’s t-test was used when comparing two groups (diets and corresponding feed levels of polychaetes fed different diets) and one-way ANOVA with subsequent Pairwise Multiple Comparison (Holm-Sidak method) was used for comparison of more than two groups. Non-normal distributed data was log-transformed or analyzed using a non-parametric Mann-Whitney Rank Sum Test when comparing two groups and Kruskal-Wallis One Way Analysis of Variance on Ranks for multiple comparison. One-sample t-tests were conducted to determine significant differences in SGRs of polychaetes at the lowest feed levels (S6 and PS6) from zero. Linear regression followed by one-way ANOVA was done to evaluate the effect of diets on composition of polychaetes.
Statistical analysis was performed at the 95% (p < 0.05) confidence level.
3 Results
3.1 Survival and growth
Survival of polychaetes ranged between 80-90% across all feed levels for both diets groups with no significant differences within or between them (Figure 1).
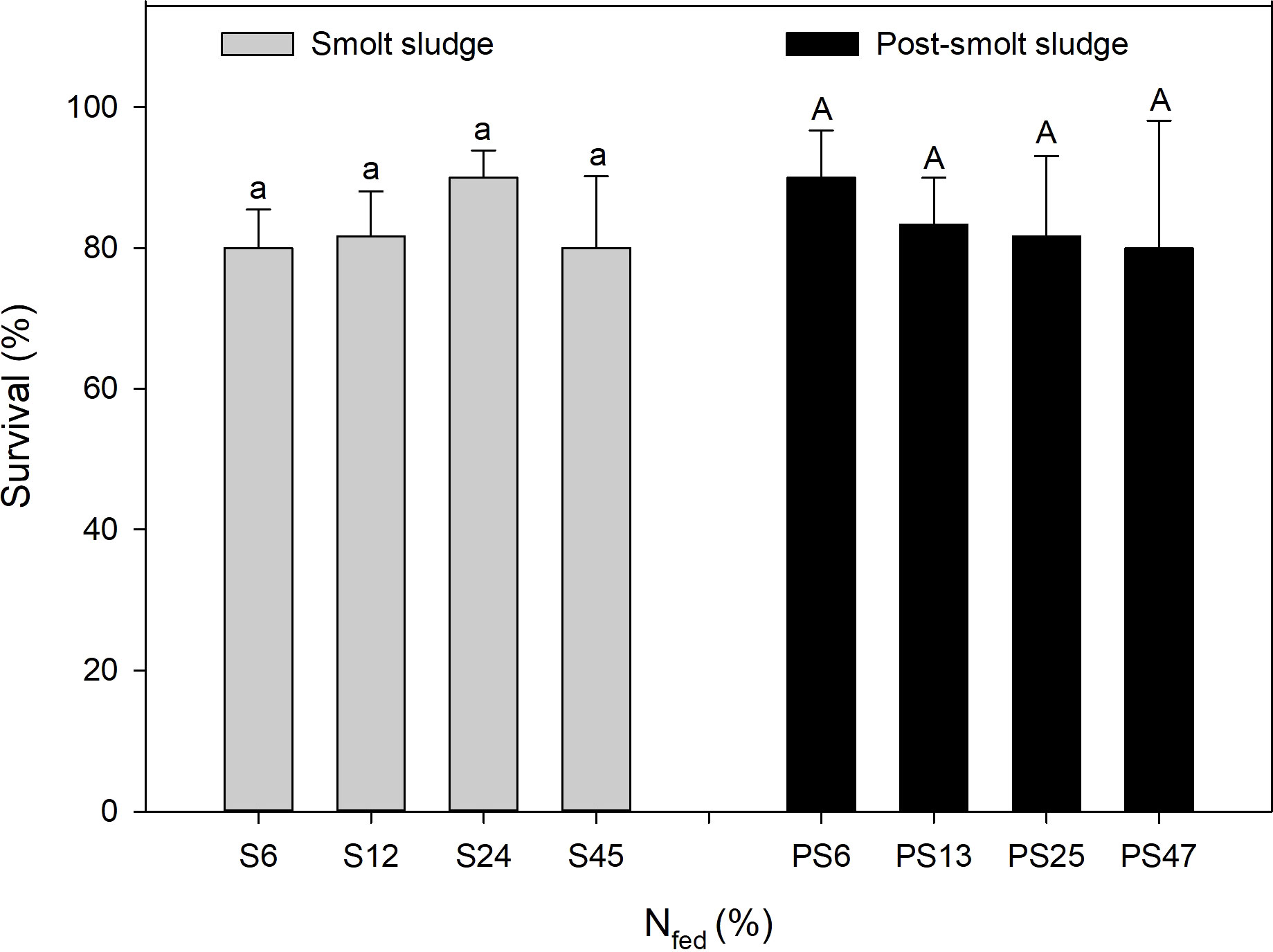
Figure 1 Survival of polychaetes H. diversicolor fed different levels of smolt sludge (S6-S24, n=4; S45, n=3) and post-smolt sludge (PS6-PS47, n=4). Same superscripts indicate non-significant differences (p ≥ 0.05).
Initial mean weight (WW) of individual polychaetes was 250 ± 48 mg across diet groups and feed levels with no significant differences (Figure 2). Polychaetes fed smolt sludge had a significantly higher final weight (WW, mg) in comparison with initial weight (Welch’s t-tests, p < 0.05) at the higher feed levels S24 (408 ± 97 mg) and S45 (382 ± 100 mg). Further, there was a significant difference (Welch’s t-test, p < 0.05) between initial and final mean weight (WW) for polychaetes fed post-smolt sludge at the highest feed level (PS47) with a final mean weight of 432 ± 53 mg. Polychaetes in both diet groups showed increasing SGRs with increasing feed levels (Figure 3). SGRs at the lowest feed levels (S6 and PS6) were not significantly different from zero (one-sample t-test, p ≥ 0.05). For the smolt sludge-fed polychaetes, SGRs ranged from -0.001 d-1 (S6) to 0.017 d-1 (S45), and polychaetes at feed levels S24 and S45 showed significantly higher growth (one-way ANOVA, p < 0.05) than at feed levels S6 and S12. For the polychaetes fed post-smolt sludge, SGRs varied between 0.003 d-1 (PS6) and 0.021 d-1 (PS47). A significantly higher SGR was found for polychaetes at the highest feed level (PS47) compared with the lowest (PS6) (one-way ANOVA, p < 0.05). There were no differences in SGRs of polychaetes fed different diets when comparing corresponding feed levels (S6 vs. PS6, S12 vs. PS13, S24 vs. PS25, and S45 vs. PS47) (Welch’s t-tests, p ≥ 0.05). SGRs of the polychaetes at the different feed levels followed the Von Bertalanffy growth function (Von Bertalanffy, 1938) with a mean maximum growth rate SGRmax of 0.0189 d-1, a growth coefficient k of 0.1067 and an SGR of zero (SGR0) at 5.8% N fed per day. Accordingly, we registered no growth at the lowest feed levels (S6 and PS6) and a steep increase in SGRs until around 15% N fed per day after which the growth function flattened. Maximum growth was approached around 25% N with no significant difference in SGRs between the two highest feed levels in either diet group, S24 and S45, and PS25 and PS47, respectively.
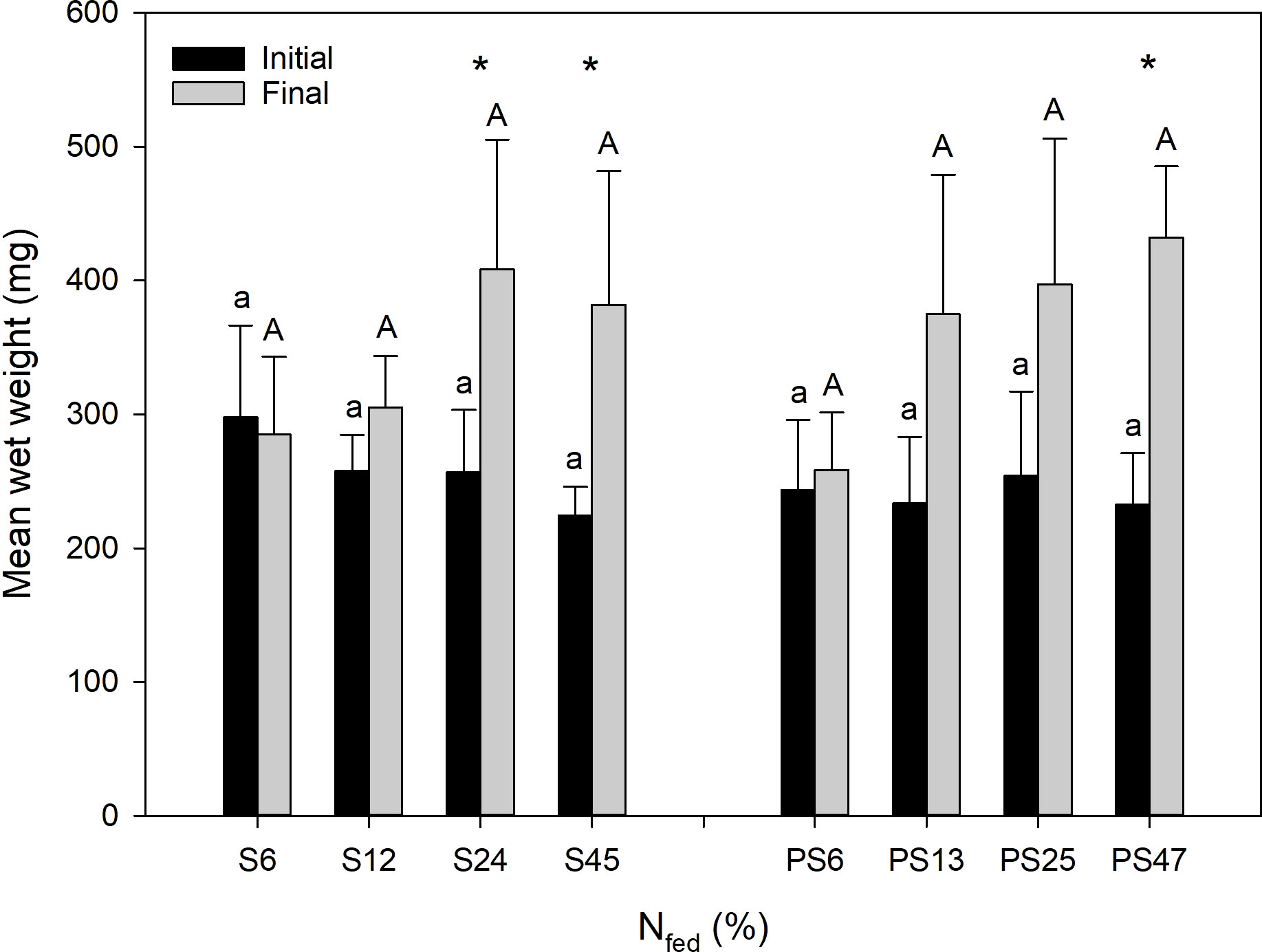
Figure 2 Initial and final mean weights (WW, mg) of polychaetes H. diversicolor fed different levels of smolt sludge (S6-S24, n=4; S45, n=3) and post-smolt sludge (PS6-PS47, n=4). Same superscripts indicate non-significant differences (p ≥ 0.05). Asterisks indicate significant differences between initial and final mean weight (p < 0.05).
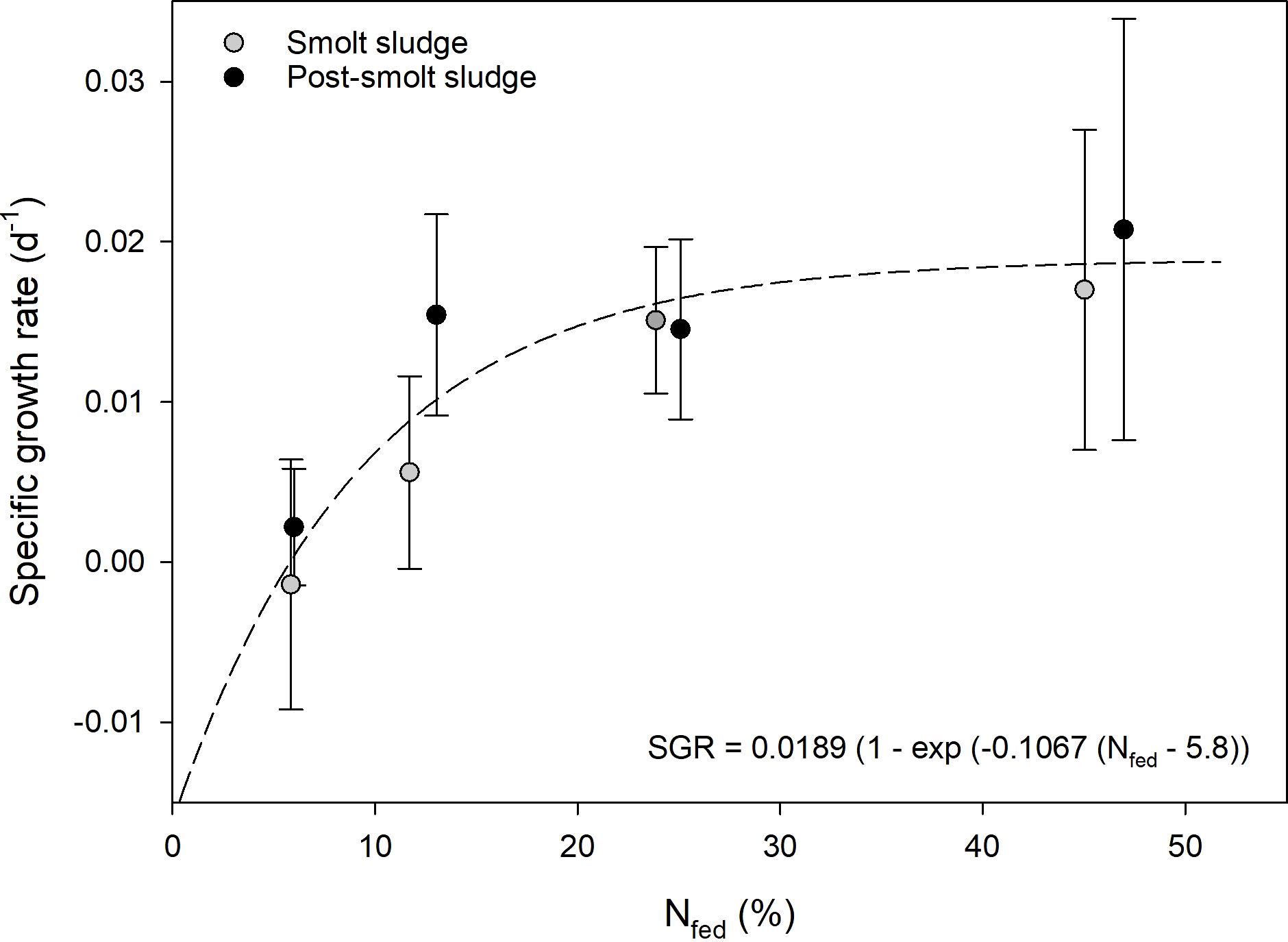
Figure 3 Specific growth rate of polychaetes H. diversicolor fed different levels of smolt sludge (S6-S24, n=4; S45, n=3) and post-smolt sludge (PS6-PS47, n=4).
3.2 Proximate biochemical and elemental composition of diets and polychaetes
3.2.1 Diets
Smolt sludge had a significantly higher proportion of moisture and unidentified material (which includes carbohydrates) than post-smolt sludge (Welch’s t-tests, p < 0.05, Table 1). AA and protein content of the two diets was not significantly different (Welch’s t-tests, p ≥ 0.05). The proportion of ash, lipids and sum of FAs was higher for post-smolt sludge than for smolt sludge (Welch’s t-tests, p < 0.05. Percentages of saturated FAs (SAFAs), polyunsaturated FAs (PUFA) and n-6 FAs were higher for post-smolt sludge compared to smolt sludge while the proportion of monounsaturated FAs (MUFAs) was significantly higher for smolt sludge (Welch’s t-tests, p < 0.05, Table 2). There was no significant difference for arachidonic acid (C20:4n-6; ARA), eicosapentaenoic acid (C20:5n-3; EPA), docosahexaenoic acid (C22:6n-3; DHA) and the sum of n-3 FAs between the two diets (Welch’s t-test, p ≥ 0.05). Content of copper (Cu) and lead (Pb) was higher for smolt sludge than for post-smolt sludge, while post-smolt sludge was higher in calcium (Ca), iron (Fe), zinc (Zn), and cadmium (Cd) (Table 3). Content of arsenic (As) and tin (Sn) was similar in both type of diets.
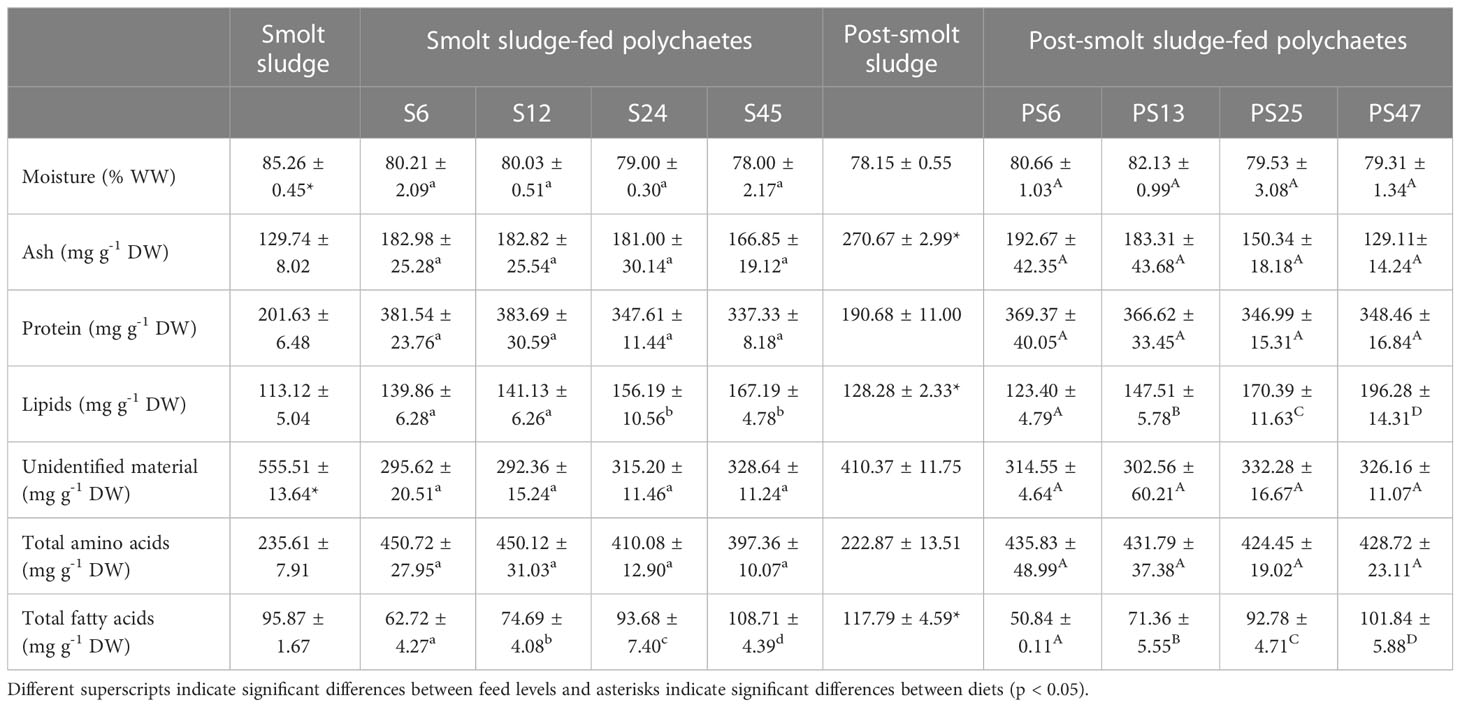
Table 1 Composition of smolt sludge (n=3), post-smolt sludge (n=3) and polychaetes H. diversicolor fed different levels of smolt sludge (S6-S24, n=4; S45, n=3) and post-smolt sludge (PS6-PS47, n=4).

Table 2 Relative fatty acid (FA) composition (% of total FAs) of smolt sludge (n=3), post-smolt sludge (n=3), and polychaetes H. diversicolor fed different levels of smolt sludge (S6-S24, n=4; S45, n=3) and post-smolt sludge (PS6-PS47, n=4).
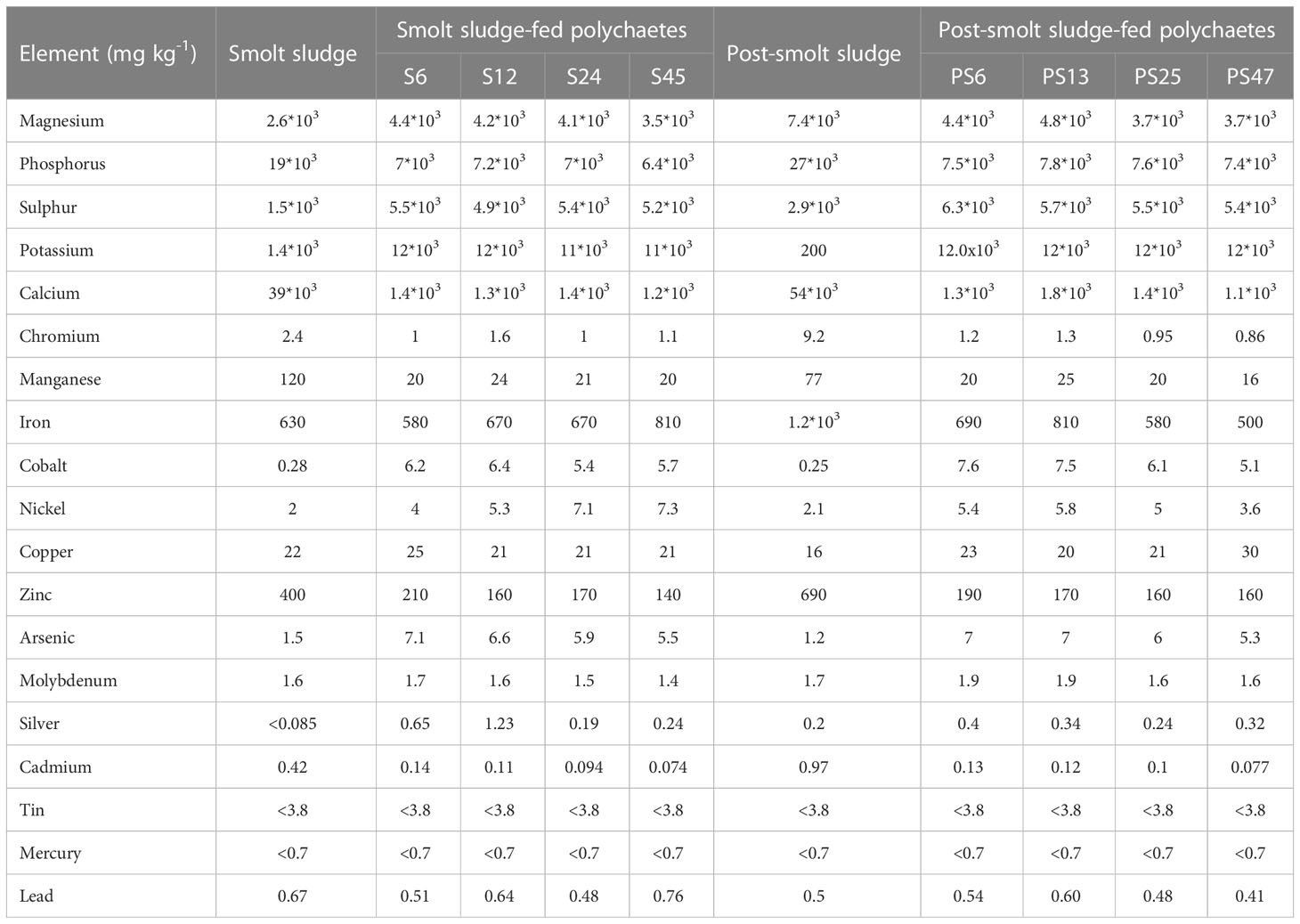
Table 3 Elemental composition of smolt sludge (n=1), post-smolt sludge (n=1), polychaetes H. diversicolor fed different levels of smolt sludge (S6-S45, n=1) and post-smolt sludge (PS6-PS47, n=1).
3.2.2 Polychaetes
Ash content of polychaetes fed smolt sludge and post-smolt sludge was not affected by different feed levels (S6-S45, PS6-47; one-way ANOVA, p ≥ 0.05, Table 1). There was no difference in ash content when comparing the corresponding feed levels of the two diet groups, smolt sludge-fed polychaetes and post-smolt sludge-fed polychaetes, respectively (Welch’s t-test, p ≥ 0.05).
Protein content was also not affected by feed levels of polychaetes fed smolt sludge and post-smolt sludge (one-way ANOVA, p ≥ 0.05, Table 1). Moreover, there was no difference between the corresponding feed levels of the different diet groups (Welch’s t-test, p ≥ 0.05).
For polychaetes fed smolt sludge, lipid content increased significantly from the lower feed levels S6 and S12 to the higher feed levels S24 and S45 (one-way ANOVA, p < 0.05, Table 1). For polychaetes fed post-smolt sludge, lipid content increased significantly with each feed level. When comparing lipid content of polychaetes fed different sludge types at the corresponding feed levels, lipid content of polychaetes at level PS6 was significantly lower than at S6, while at PS47 it was significantly higher than at S45 (Welch’s t-test, p < 0.05). For the other feed levels, no differences were observed.
The proportion of unidentified material, which includes carbohydrates, of polychaetes did not vary significantly between the different feed levels in either of the diet groups (one-way ANOVA, p ≥ 0.05, Table 1). Further, there was no difference between corresponding feed levels (Welch’s t-test, p ≥ 0.05).
Total AA content was similar with no significant difference between feed levels both within and between diet groups (one-way ANOVA, p ≥ 0.05, Table 1). Essential AA (EAA) composition (mg g-1 DW and % of total AAs) was not significantly different when comparing polychaetes fed at different feed levels within diet groups (one-way ANOVA, p ≥ 0.05) and polychaetes from corresponding feed levels (Welch’s t-test, p ≥ 0.05) between diet groups. Further, no difference was detected for non-essential AA (NEAA) composition (mg g-1 DW and % of total AAs) (Supplementary Tables 1, 2).
For both smolt- and post-smolt sludge-fed polychaetes, total FA content increased significantly with each feed level (one-way ANOVA, p < 0.05), with the lowest FA content at the lowest feed levels and the highest FA content at the highest feed levels in both diet groups (Table 1).
For polychaetes fed smolt sludge, a significant increase in SAFAs, MUFAs, PUFAs, ARA, EPA, DHA, and the sum of n-6 FA (mg g-1 DW) was registered from the lowest (S6) to the highest (S45) feed level (one-way ANOVA, p < 0.05, Supplementary Table 3). In terms of relative composition (% of total FAs), the proportion of SAFAs decreased while percentages of MUFAs simultaneously increased with each feed level (one-way ANOVA; p < 0.05, Table 2). For PUFAs, there was no difference for polychaetes between feed levels (one-way ANOVA, p ≥ 0.05, S6-S45). Percentages of essential FAs, ARA, and EPA as well as the sum of n-3 FAs were lower at the highest feed level (S45) compared to the lowest feed level (S6); the sum of n-6 FAs was lowest at the lowest feed level (one-way ANOVA, p < 0.05). There was no clear trend for the polychaetes’ content of DHA, as it was only significantly higher for polychaetes at feed level S24 compared to the other feed levels.
For polychaetes fed post-smolt sludge, absolute values (mg g-1 DW) of MUFAs, PUFAs, ARA, EPA, DHA as well as the sum of n-3 and n-6 FAs were significantly higher at the highest (PS47) than at the lowest (PS6) feed level (one-way ANOVA, p < 0.05, Supplementary Table 3). Considering the relative composition of FAs (% of total FAs), MUFAs showed a decrease with increased feed supply while PUFAs, EPA, DHA, and the sum of n-3 and n-6 FAs were significantly higher at the highest feed level (PS47) than at the lowest (PS6) (one-way ANOVA, p < 0.05, Table 2). The proportion of SAFAs and ARA was not significantly different between feed levels (PS6-PS47)
Arsenic and zinc content of H. diversicolor decreased with increasing feeding level in both diet groups (Table 3). Cadmium content was lowest at the highest feed levels in both diet groups. No trend could be detected for calcium or lead content. Iron content of polychaetes fed smolt sludge increased with increasing feed level, but no trend could be detected for polychaetes fed post-smolt sludge. Copper content decreased with increased feed supply for polychaetes fed smolt sludge but was similar for all polychaetes fed post-smolt sludge. Tin content was below sensitivity limit (3.8 mg kg-1) at all feed levels in both diet groups. Generally, there was no increase with increased feed supply in any of the elements analyzed.
3.3 Dietary effects on biochemical composition of Hediste diversicolor
Linear regression analysis revealed a strong positive correlation (R2 ≥ 0.65; p < 0.05) between supplied lipids, total FAs fed, PUFAs, and n-6 FAs (mg fed g-1 DW) and the respective content of polychaetes (mg g-1 DW) for polychaetes in both diet groups, where lipid, total FA, PUFA and n-6 FA content increased with increased feed supply for polychaetes fed smolt sludge and post-smolt sludge (Table 4).
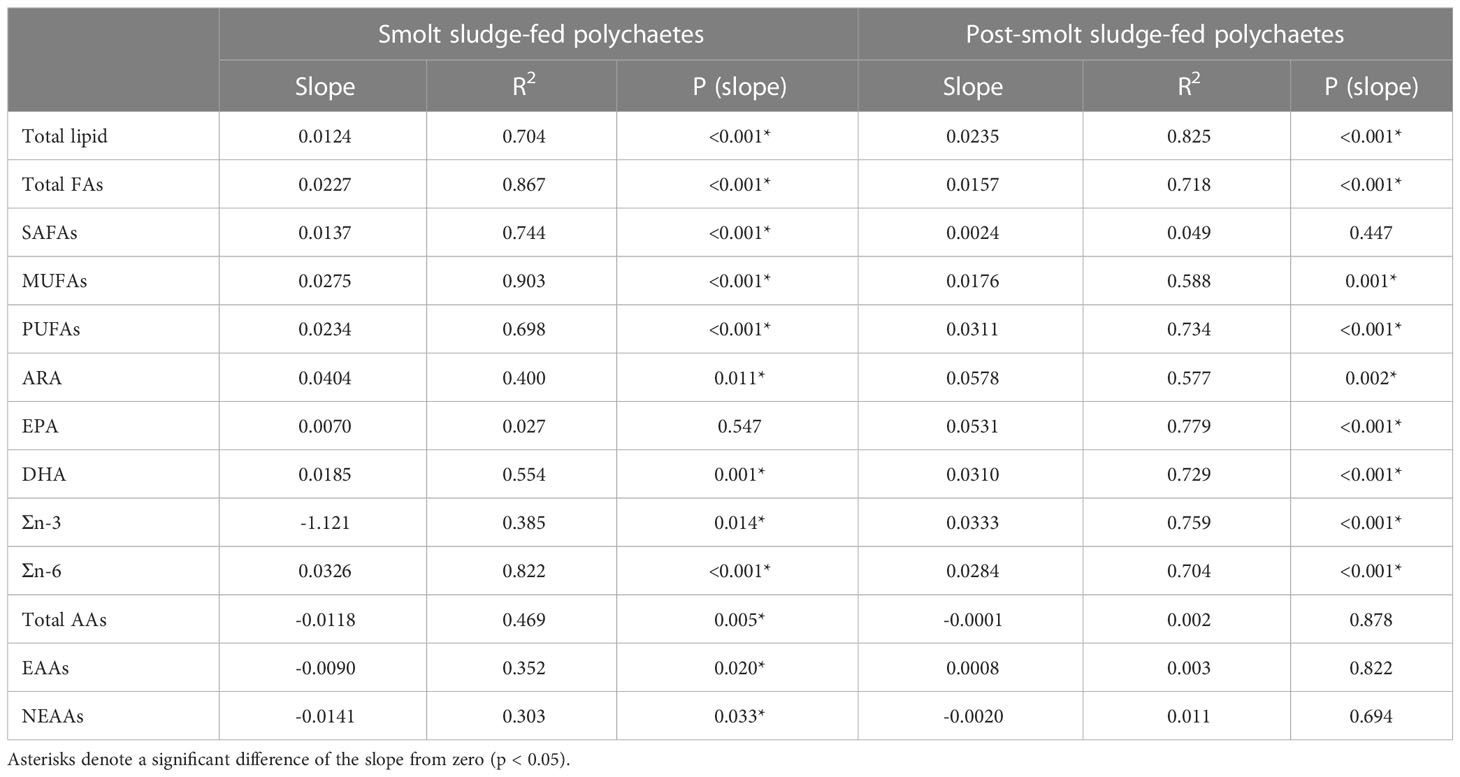
Table 4 Linear regression analysis of total lipid, total fatty acids (FAs), saturated FAs (SAFAs), monounsaturated FAs (MUFAs), polyunsaturated FAs (PUFAs), ARA, EPA, DHA, Σn-3 FAs, Σn-6 FAs, amino acids (AAs), essential AAs (EAAs), and non-essential AAs (NEAAs) for polychaetes H. diversicolor fed increasing rations of corresponding constituent in smolt sludge (S6-S24, n=4; S45, n=3) and post-smolt sludge (PS6-PS47, n=4).
For polychaetes fed smolt sludge, a strong positive correlation (R2 ≥ 0.65; p < 0.05) was additionally found between feed level and corresponding SAFA and MUFA content (mg g-1 DW) (Table 4). Moderate positive correlation (0.4 ≤ R2 < 0.65; p < 0.05) between supplied components and the respective content of polychaetes could be seen for ARA and DHA (mg g-1 DW), while a moderate negative correlation was found for total AAs (mg g-1 DW). There was a weak negative correlation (0.2 ≤ R2 < 0.4; p < 0.05) for n-3 FAs, EAA and NEAA and no correlation for EPA (mg g-1 DW) (R2 < 0.2; p ≥ 0.05).
Further, a strong positive correlation (R2 ≥ 0.65; p < 0.05) between feed level and content of EPA, DHA and n-3 FAs (mg g- 1 DW) was seen for polychaetes fed post-smolt sludge (Table 4). Moderate positive correlation (0.4 ≤ R2 < 0.65; p < 0.05) could be observed for MUFAs and ARA (mg g-1 DW). No correlation was found for the remaining components SAFAs, AAs, EAAs, and NEAAs (mg g-1 DW) (R2 < 0.2; p ≥ 0.05).
PCA showed that relative FA composition (% of total FAs) of polychaetes was affected by both their feed level and moreover, to a certain degree their diet group (Figure 4). PC1 and PC2 combined explained 61% of the variance in the data set. Clear differences could be seen between the polychaetes at different feed levels that were fed the same diet, S6-S45 and PS6-PS47, respectively. Differences in the variance between the two diets were decreasing with higher feed supply (feed level).
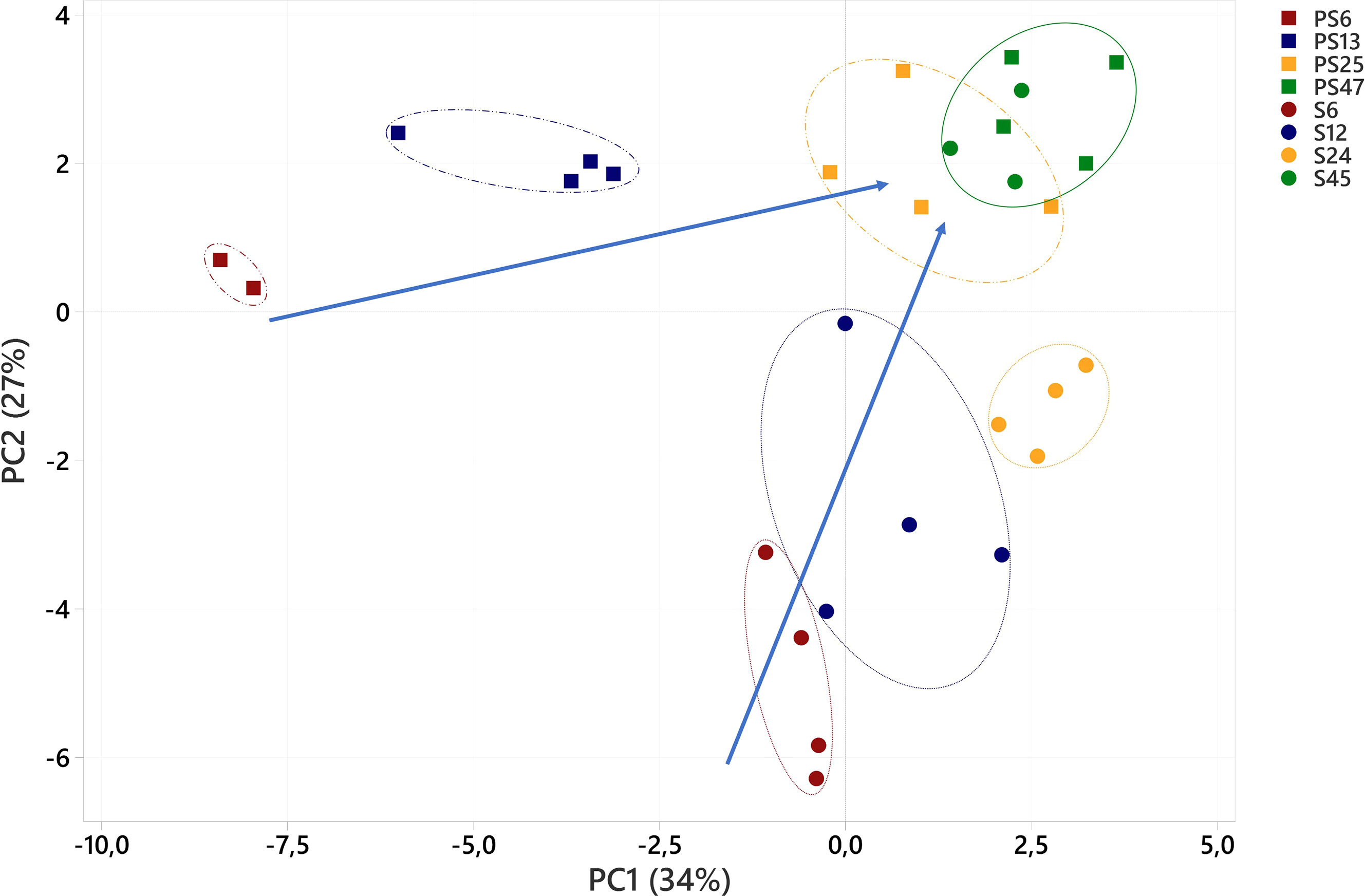
Figure 4 Principal component analysis of fatty acids (% of total fatty acids) in polychaetes H. diversicolor fed different levels of smolt sludge (S6-S24, n=4; S45, n=3) and post-smolt sludge (PS6, n=2, PS13-PS47, n=4).
PCA revealed that AA composition (% of total AAs) was neither impacted by diet nor by feed level (Figure 5). Together, PC1 and PC2 explained 44% of the variance in AA composition.
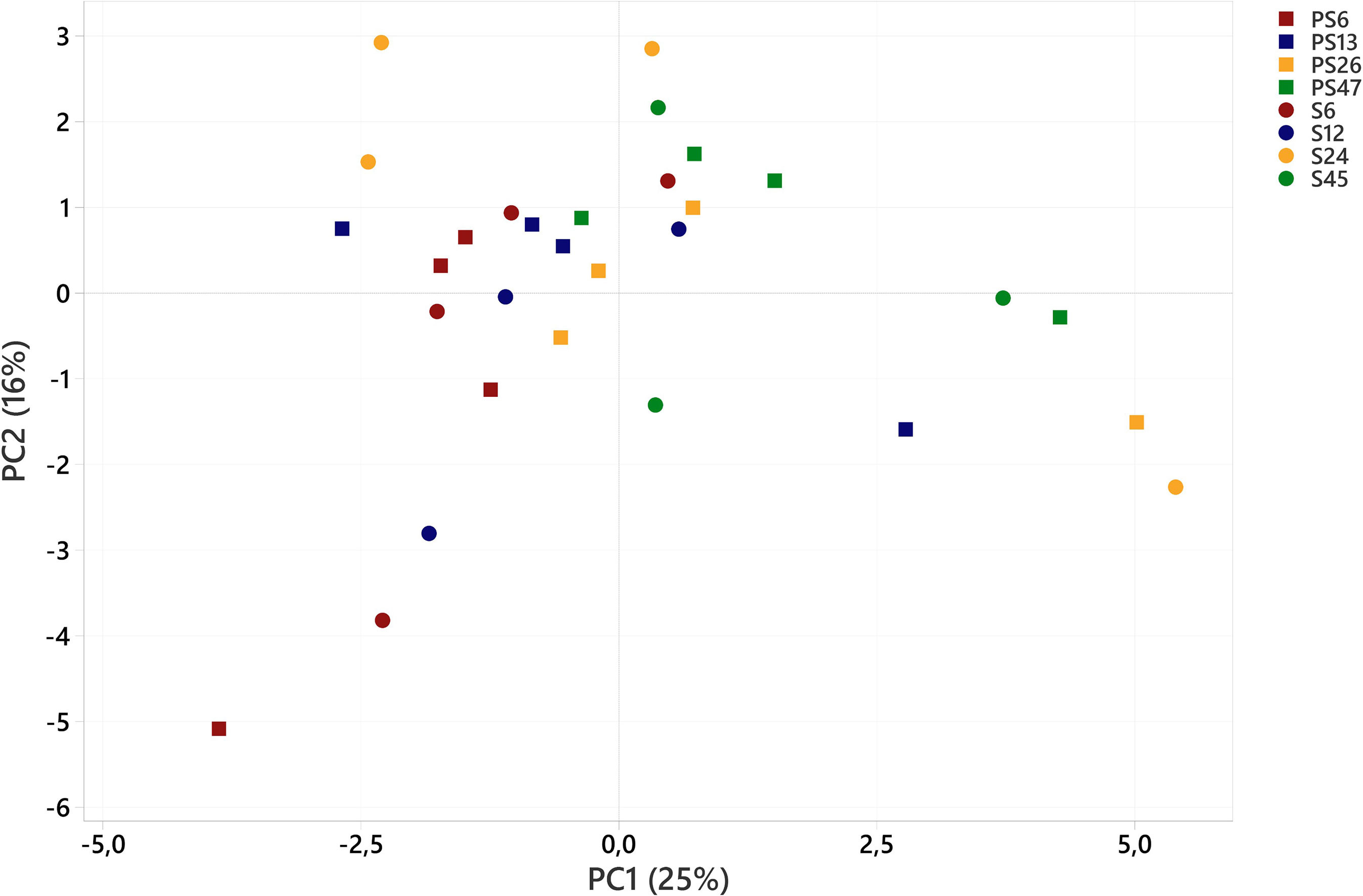
Figure 5 Principal component analysis of amino acids (% of total amino acids) in polychaetes H. diversicolor fed different levels of smolt sludge (S6, S24 n=4, S12, S45, n=3) and post-smolt sludge (PS6, n=2, PS13-PS47, n=4).
4 Discussion
Our study showed that H. diversicolor can successfully be cultivated on waste sludge from both land-based smolt and post-smolt production and specific growth rates at the high feed levels were higher than those in natural populations (Kristensen, 1984). At the lowest feed levels (S6 and PS6), SGRs were not significantly different from zero, suggesting that all energy supplied was invested in basic metabolic processes rather than somatic growth. We found maximum SGRs at the highest feed levels, amounting to 0.017 d-1 for polychaetes fed smolt sludge (S45), and 0.021 d-1 for polychaetes fed post-smolt sludge (PS47). SGRs followed the Von Bertalanffy growth function (Von Bertalanffy, 1938) with levelling off of growth between two highest feed levels as no significant difference was found between these, neither for polychaetes fed smolt sludge (S24 and S45), nor for those fed post-smolt sludge (PS25 and PS47). This implies that maximum growth was found for the used experimental setup. Further increase in feed supply may negatively affect experimental conditions and water quality and would hence not result in increased SGRs. Overall, SGRs in our trial were similar to those reported in other studies with comparable conditions (Bischoff, 2007; Brown et al., 2011). Moreira et al. (2005) established saturation feeding rates for H. diversicolor using one day old Artemia franscicana nauplii at a range of temperatures. They found that 70 mg polychaetes H. diversicolor could consume as much as 80 nauplii per hour, resulting in 1920 nauplii per day if constantly feeding. Following Evjemo and Olsen (1999), one-day old A. franciscana nauplii contain 0.19 µg nitrogen, meaning 1920 nauplii have a nitrogen content equivalent to 31% of nitrogen contained in the 70 mg polychaetes. When taking into account that H. diversicolor does not constantly feed, the numbers reported for feeding saturation by Moreira et al. (2005) are in good agreement with the results we found, even if nitrogen quality in Artemia franscicana is different from that in aquaculture sludge.
Growth of H. diversicolor is affected by different factors. Foremost, it is determined by diet quantity and quality, specifically protein supply and composition (Nesto et al., 2012; Santos et al., 2016). Wang et al. (2019a; 2019b) reported lower specific growth rates of 0.012 d-1 and 0.009 d-1 when providing a lower quantity of smolt sludge to H. diversicolor. In our study, the two diets smolt sludge and post-smolt sludge were found to have a similar protein content, likely explaining the similar growth rates found for the two diets across feed levels. In comparison with our study, Brown et al. (2011) found a higher SGR of 0.033 d-1 when cultivating A. virens on halibut waste which contained 496 mg protein g-1 DW. In addition to a more energy-dense diet and differences between the two species, the small size of individuals in their study may have impacted growth, as the individuals had an initial biomass of 180 mg compared to 250 mg in our study and growth rates of H. diversicolor decline with increasing size (Heip and Herman, 1979).
We aimed to simulate summer conditions by providing long days (16L:8D) and warm temperatures (16°C) to suppress gametogenic development and promote somatic growth (Olive et al., 1997; Last and Olive, 1999). However, given the collection of polychaetes in late fall and a larger size of individuals compared to other studies, it is possible that sexual maturation processes of H. diversicolor had already started. Due to natural changes in day length, temperature, and food availability in fall, energy consumed is invested in sexual maturation instead of somatic growth (Kristensen, 1984). Higher SGR of 0.034 d-1 (Pajand et al., 2017) and 0.06-0.07 d-1 (Nesto et al., 2012) have been reported in studies with smaller, juvenile individuals at higher temperatures.
Significantly different ash content between smolt and post-smolt sludge was not reflected in ash content of polychaetes H. diversicolor. Overall, ash content did not differ significantly between feed levels within diet groups or corresponding feed levels of different diet groups, although values at the highest feed levels were lower than those at the lowest. Lower values for ash content of H. diversicolor have been reported in previous studies (Wang et al., 2019a; Wang et al., 2019b).
Polychaetes fed both types of diet had a similar protein content. Since there was no difference between polychaetes fed different diets or feed levels, protein content of H. diversicolor can be considered independent of their diet if feed supply is sufficient. Low feed supply has been presumed to increase protein content since spending more energy on search for feed is correlated with increased muscle stress which leads to formation of muscle tissue that is rich in proteins (Nielsen, 1997; Klinke et al., 2009; Suckow, 2010). Values of protein content in our study were similar to other studies when considering the different methods of protein calculation, suggesting that even though we registered no growth at the lowest feed levels (S6 and PS6), none of the feed levels or diets were so restricting in protein that it would have had a significant effect on protein content of H. diversicolor (Pajand et al., 2017; Wang et al., 2019a; Wang et al., 2019b). Differences in protein quantities supplied to the polychaetes on different feed levels did however affect growth of H. diversicolor as discussed previously.
Total AA content of polychaetes fed smolt sludge and post smolt sludge was within the same range. Both diets had a similar content of AAs but showed somewhat different AA profiles. The differences in AA composition of the supplied diets were not reflected in the composition of H. diversicolor as there were no differences between feed levels or diets and PCA revealed no grouping of diets or feed levels. AA composition is not controlled by diet but rather determined genetically and excess AAs in feed are not used for protein synthesis or stored in tissue but catabolized for energy production (Owen et al., 1979). However, AAs are building blocks for proteins and limited supply of both essential and non-essential AAs will affect animal growth and health (Wu, 2014).
While ash, protein, and AA content as well as AA composition of the two different diets had limited effect on the composition of polychaetes, an effect of lipid content and FA composition could be confirmed in our study. Lipid content increased with increased feed supply for polychaetes fed both diets. As in other studies, linear regression revealed a strong correlation of lipid content in diet and H. diversicolor (Wang et al., 2019a). Accumulation of lipids is linked to excess supply in the provided diets as polychaetes have been shown to store lipids before sexual maturation in times of high food availability (Luis and Passos, 1995; García-Alonso et al., 2008). In our study, post-smolt sludge had a significantly higher lipid content than smolt sludge. This difference resulted in a significant increase in lipid content with each feed level, and additionally an effect when comparing the highest feed levels (S45 vs. PS47) where a significantly higher lipid content was registered for polychaetes fed post-smolt sludge than those fed smolt sludge.
The total FA content of polychaetes in this trial increased with each level in both diet groups. At the highest feed levels, FA content was similar to those found by Pajand et al. (2017) when H. diversicolor was fed beluga sturgeon (Huso huso L.) waste sludge. Generally, the FA composition of diet is described as a determining factor for FA composition of H. diversicolor (Luis and Passos, 1995; Pajand et al., 2017; Malzahn et al., 2023). In our study, we could confirm that differences in FA composition of diets and content of both individual and groups of FAs as well as differences in quantity supplied (feed level) were reflected in the corresponding content of polychaetes. Overall, we could register a strong effect of feed level on the content (mg g-1 DW) of SAFAs, MUFAs, PUFAs, ARA, EPA, DHA as well as the sum of n-3 and n-6 FAs, as levels in almost all groups increased with increased feed supply in both diet groups and linear regression found strong correlation. A similar effect has been reported by Wang et al. (2019a) and Malzahn et al. (2023).
Following relative composition (% of total FAs) of diets, polychaetes reared on both diets had the same major FA. The most abundant SAFA was C16:0 (palmitic acid) while C18:1 n-9 (oleic acid) was the major MUFA. EPA and C18:2 n-6 (linoleic) were the most abundant PUFAs. Differences in proportion of EPA, DHA, and linoleic acid in the two diets were reflected in the polychaetes. Moreover, we saw a relative decrease in SAFAs with increasing feed level in both diet groups which corresponded to a relative increase in MUFAs for polychaetes fed smolt sludge and a relative increase in PUFAs for post-smolt-fed polychaetes, suggesting a selective enrichment and a shift of the FA profile towards a higher proportion of unsaturated FAs by H. diversicolor with increased feed supply as described in previous studies (García-Alonso et al., 2008; Kabeya et al., 2020; Jerónimo et al., 2021).
The polychaetes showed clear differences in their FA composition between feed levels and diets on the lower feed levels. These differences between polychaetes fed different types of sludge are due to differences in lipid content and FA composition of the diets as described by (Luis and Passos, 1995; Pajand et al., 2017). At the higher feed levels, the differences between diets became increasingly smaller, suggesting firstly a convergence of FA profiles with high feed supply and secondly a saturation to the point where the variation in FA composition of polychaetes fed similar diets becomes very small and an equilibrium is reached.
Elemental analysis showed that increased availability of feed did not result in accumulation of the analyzed elements in polychaete tissue. For arsenic, zinc, and cadmium, the lowest content was detected at the highest feed levels. However, even though the content of arsenic decreased in all treatments, it remained higher than the threshold value of EU’s feed ingredients of 2.27 mg arsenic kg-1 feed (DW). All other regulated elements stayed within the thresholds for legislated content (Nærings- og fiskeridepartementet and Landbruks- og matdepartementet, 2015).
5 Conclusion
Our results confirmed the hypotheses that H. diversicolor can grow successfully using smolt sludge and post-smolt sludge as sole feed sources, and that increased feed supply will result in increased specific growth rates up to a certain point. Differences in the composition of diets had no effect on growth. SGRs followed the Von Bertalanffy growth function and showed a levelling off of growth between the two highest feed levels, suggesting that maximum growth for individuals of this size (250 mg) fed smolt and post-smolt sludge with the given abiotic factors was found in this experiment, and growth could not be increased further. Additionally, we concluded a strong effect of lipid and FA content and composition of diets on corresponding content and composition of H. diversicolor, which confirmed our hypothesis that feed supply will affect biochemical composition with regards to lipid and FAs. Ash, protein, and AA content as well as AA composition of polychaetes were not affected by different diets or varying food supply.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
IA: Conceptualization, Formal analysis, Investigation, Writing – original draft, Writing – review & editing. TD: Formal analysis, Investigation, Writing – review. BK: Formal analysis, Investigation, Writing – review. AH: Conceptualization, Investigation, Writing – review & editing, Funding acquisition. AM: Conceptualization, Investigation, Writing – review & editing, Visualization, Project administration, Funding acquisition. KR: Conceptualization, Investigation, Writing – review & editing, Project administration, Funding acquisition. All authors contributed to the article and approved the submitted version.
Funding
We acknowledge funding by the RCN projects POLYCHAETE (#280836), MIND-P (#268338), and the H2020 ERA-NET BlueBio COFUND Project “SIDESTREAM – Secondary bio-production of low trophic organisms utilizing side streams from the Blue and Green sectors to produce novel feed ingredients for European aquaculture” (Grant ID 68), co-funded through national funds provided by the Norwegian Research Council (#311701). The project was carried out in the framework of the national research infrastructure “Norwegian Center for Plankton Technology” (Norwegian Research Council #245937/F50) hosted by SINTEF and NTNU.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2023.1197052/full#supplementary-material
References
Aas T. S., Åsgård T. E. (2017). Estimated content of nutrients and energy in feed spill and faeces in Norwegian salmon culture. Nofima rapportserie 19 (2017), 1–8.
Bischoff A. A. (2007). Solid waste reduction of closed recirculated aquaculture systems by secondary culture of detritivorous organisms (Germany: Ph.D. thesis. Christian-Albrechts-Universität Kiel).
Bligh E. G., Dyer W. J. (1959). A rapid method of total lipid extraction and purification. Can. J. Biochem. 37, 911–917. doi: 10.1139/o59-099
Brown N., Eddy S., Plaud S. (2011). Utilization of waste from a marine recirculating fish culture system as a feed source for the polychaete worm, Nereis virens. Aquaculture 322-323, 177–183. doi: 10.1016/j.aquaculture.2011.09.017
Colgrave M. L., Dominik S., Tobin A. B., Stockmann R., Simon C., Howitt C. A., et al. (2021). Perspectives on future protein production. J. Agric. Food Chem. 69, 15076–15083. doi: 10.1021/acs.jafc.1c05989
Evjemo J. O., Olsen Y. (1999). Effect of food concentration on the growth and production rate of artemia franciscana feeding on algae (T. iso). J. Exp. Mar. Biol. Ecol. 242, 273–296. doi: 10.1016/S0022-0981(99)00104-5
EWOS. (2022a). EWOS CLEAR 4-100 ekstrudert fullfôr til laks og ørret i ferskvann. Bergen, Norway: EWOS AS.
EWOS. (2022b). EWOS CLEAR 200 extruded feed for salmon and trout in fresh water. Bergen, Norway: EWOS AS.
FAO. (2022). The state of world fisheries and aquaculture 2022. towards blue transformation (Rome, Italy:FAO). doi: 10.4060/cc0461en
Fidalgo e Costa P., Narciso L., Cancela da Fonseca L. (2000). Growth, survival and fatty acid profile of Nereis diversicolor (OF müller 1776) fed on six different diets. Bull. Mar. Sci. 67, 337–343. doi: 10.22092/ijfs.2019.118779
Fiskeridirektoratet (2022) Matfiskproduksjon: salg av laks og regnbueørret, og salg per sysselsatt 1994-2021. Available at: https://www.fiskeridir.no/Akvakultur/Tall-og-analyse/Akvakulturstatistikk-tidsserier/Laks-regnbueoerret-og-oerret/Salg%20av%20laks%20og%20regnbue%C3%B8rret (Accessed 17.01. 2023).
Fiskeridirektoratet (2023) Forbruk av fôr fordelt på art 2017-2022 (produksjonsområde). Available at: https://www.fiskeridir.no/Akvakultur/Tall-og-analyse/Biomassestatistikk/Biomassestatistikk-etter-produksjonsomraade (Accessed 15.01. 2023).
García-Alonso J., Müller C. T., Hardege J. (2008). Influence of food regimes and seasonality on fatty acid composition in the ragworm. Aquat. Biol. 4, 7–13. doi: 10.3354/ab00090
Heip C., Herman R. (1979). Production of Nereis diversicolor o. f. müller (Polychaeta) in a shallow brackish-water pond. Estuar. Coast. Shelf Sci. 8, 297–305. doi: 10.1016/0302-3524(79)90047-1
Jerónimo D., Lillebø A. I., Rey F., Ii H. K., Domingues M. R. M., Calado R. (2021). Optimizing the timeframe to produce polychaetes (Hediste diversicolor) enriched with essential fatty acids under different combinations of temperature and salinity. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.671545
Jerónimo D., Lillebø A. I., Santos A., Cremades J., Calado R. (2020). Performance of polychaete assisted sand filters under contrasting nutrient loads in an integrated multi-trophic aquaculture (IMTA) system. Sci. Rep. 10, 1–10. doi: 10.1038/s41598-020-77764-x
Kabeya N., Gür İ., Oboh A., Evjemo J. O., Malzahn A. M., Hontoria F., et al. (2020). Unique fatty acid desaturase capacities uncovered in Hediste diversicolor illustrate the roles of aquatic invertebrates in trophic upgrading. Philos. Trans. R. Soc Lond. B Biol. Sci. 375, 20190654. doi: 10.1098/rstb.2019.0654
Klinke R., Pape H.-C., Kurtz A., Silbernagl S. (2009). Physiologie (Stuttgart, Germany: Georg Thieme Verlag).
Kristensen E. (1984). Life cycle, growth and production in estuarine populations of the polychaetes Nereis virens and N. diversicolor. Ecography 7, 249–250. doi: 10.1111/j.1600-0587.1984.tb01128.x
Last K. S., Olive P. J. W. (1999). Photoperiodic control of growth and segment proliferation by Nereis (Neanthes) virens in relation to state of maturity and season. Mar. Biol. 134, 191–199. doi: 10.1007/s002270050537
Luis O. J., Passos A. M. (1995). Seasonal changes in lipid content and composition of the polychaete nereis (Hediste) diversicolor. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 111, 579–586. doi: 10.1016/0305-0491(95)00029-8
Malzahn A. M., Villena-Rodríguez A., Monroig Ó., Johansen Å., Castro L. F. C., Navarro J. C., et al. (2023). Diet rather than temperature determines the biochemical composition of the ragworm Hediste diversicolor (OF müller 1776) (Annelida: nereidae). Aquaculture 569, 739368. doi: 10.1016/j.aquaculture.2023.739368
Meriac A. (2019). Smolt production and the potential for solid waste collection in Norway. Nofima rapportserie 25 (2019), 1–12.
Metcalfe L. D., Schmitz A. A., Pelka J. R. (1966). Rapid preparation of fatty acid esters from lipids for gas chromatographic analysis. Anal. Chem. 38, 514–515. doi: 10.1021/ac60235a044
Moreira S. M., Moreira-Santos M., Guilhermino L., Ribeiro R. (2005). A short-term sublethal in situ toxicity assay with Hediste diversicolor (Polychaeta) for estuarine sediments based on postexposure feeding. Environ. Toxicol. Chem. 24, 2010. doi: 10.1897/04-473r1.1
Nærings- og fiskeridepartementet and Landbruks- og matdepartementet (2015). Forskrift om fôrvarer: grenseverdier for uønskede stoffer i fôrvarer som nevnt i direktiv 2002/32/EF artikkel 3 annet ledd. Oslo, Norway: Lovdata.
Nesto N., Simonini R., Prevedelli D., Da Ros L. (2012). Effects of diet and density on growth, survival and gametogenesis of Hediste diversicolor (OF müller 1776) (Nereididae, polychaeta). Aquaculture 362, 1–9. doi: 10.1016/j.aquaculture.2012.07.025
Nielsen K. S. (1997). Animal physiology: adaptation and environment (Cambridge, UK: Cambridge University Press).
Olaussen J. O. (2018). Environmental problems and regulation in the aquaculture industry. insights from Norway. Mar. Policy 98, 158–163. doi: 10.1016/j.marpol.2018.08.005
Olive P., Fletcher J., Rees S., Desrosiers G. (1997). Interactions of environmental temperature with photoperiod in determining age at maturity in a semelparous polychaete Nereis (Neanthes) virens sars. J. Therm. Biol. 22, 489–497. doi: 10.1016/S0306-4565(97)00067-3
Olsen Y. (2011). Resources for fish feed in future mariculture. Aquac. Environ. Interact. 1, 187–200. doi: 10.3354/aei00019
Otsuka K., Fan S. (2021). Agricultural development: new perspectives in a changing world (Washington, D.C., USA: International Food Policy Research Institute (IFPRI). doi: 10.2499/9780896293830
Owen O., Reichard G., Patel M., Boden G. (1979). Energy metabolism in feasting and fasting. Horm. Energy Metab. 169–188. doi: 10.1007/978-1-4757-0734-2_8
Pajand Z. O., Soltani M., Bahmani M., Kamali A. (2017). The role of polychaete Nereis diversicolor in bioremediation of wastewater and its growth performance and fatty acid composition in an integrated culture system with Huso huso (Linnaeus 1758). Aquac. Res. 48, 5271–5279. doi: 10.1111/are.13340
Regjeringen (2015). Forutsigbar og miljømessig bærekraftig vekst i norsk lakse- og ørretoppdrett, meld. st. 16 (2014-2015) melding til stortinget, tilråding fra nærings- og fiskeridepartementet 20 (Regjeringen Solberg: godkjent i statsråd samme dag).
Santos A., Granada L., Baptista T., Anjos C., Simões T., Tecelão C., et al. (2016). Effect of three diets on the growth and fatty acid profile of the common ragworm Hediste diversicolor (O.F. müller 1776). Aquaculture 465, 37–42. doi: 10.1016/j.aquaculture.2016.08.022
Searchinger T., Waite R., Hanson C., Ranganathan J., Dumas P., Matthews E., et al. (2019). Creating a sustainable food future: a menu of solutions to feed nearly 10 billion people by 2050 (Washington, D.C., USA: WRI).
Shahbandeh M. (2020) Distribution of salmon production worldwide in 2018, by leading country. Available at: https://www.statista.com/statistics/1182142/leading-salmon-producers-worldwide/ (Accessed 17.01. 2023).
Šližytė R., Opheim M., Storrø I., Sterten H. (2017). Simple technologies for converting rest raw materials of Atlantic salmon (Salmo salar) into high-quality, valuable, and tasty feed ingredients. J. Aquat. Food Prod. Technol. 26, 604–619. doi: 10.1080/10498850.2016.1247124
Suckow B. (2010). Untersuchungen zur integrierten aquakultur mittels detritivorer organismen als verwerter der feststoffe in der marinen aquakultur (Germany: Diplomarbeit. Carl von Ossietzky Universität Oldenburg).
United Nations (2022). World population prospects 2022: summary of results. UN DESA/POP/2022/TR/NO. 3 (New York City, USA: Department of Economic and Social Affairs, Population Division).
Von Bertalanffy L. (1938). A quantitative theory of organic growth (inquiries on growth laws. II). Hum. Biol. 10, 181–213.
Wang H., Hagemann A., Reitan K., Ejlertsson J., Wollan H., Handå A., et al. (2019a). Potential of the polychaete Hediste diversicolor fed on aquaculture and biogas side streams as an aquaculture food source. Aquac. Environ. Interact. 11, 551–562. doi: 10.3354/aei00331
Wang X., Olsen L. M., Reitan K. I., Olsen Y. (2012). Discharge of nutrient wastes from salmon farms: environmental effects, and potential for integrated multi-trophic aquaculture. Aquac. Environ. Interact. 2, 267–283. doi: 10.3354/aei00044
Wang H., Seekamp I., Malzahn A. M., Hagemann A., Carvajal A. K., Slizyte R., et al. (2019b). Growth and nutritional composition of the polychaete Hediste diversicolor (OF müller 1776) cultivated on waste from land-based salmon smolt aquaculture. Aquaculture 502, 232–241. doi: 10.1016/j.aquaculture.2018.12.047
Keywords: Hediste diversicolor, nutrient recycling, polychaetes, salmon aquaculture, aquaculture sludge, total lipid, fatty acids
Citation: Anglade I, Dahl TH, Kristensen BSB, Hagemann A, Malzahn AM and Reitan KI (2023) Biochemical composition of Hediste diversicolor (OF Müller, 1776) (Annelida: Nereidae) reared on different types of aquaculture sludge. Front. Mar. Sci. 10:1197052. doi: 10.3389/fmars.2023.1197052
Received: 30 March 2023; Accepted: 04 May 2023;
Published: 22 May 2023.
Edited by:
Paola Gabriela Scodelaro Bilbao, CONICET Centro de Recursos Naturales Renovables de la Zona Semiárida (CERZOS), ArgentinaReviewed by:
Ingrid Lupatsch, AB Agri Ltd, United KingdomAna Pombo, Center for Marine and Environmental Sciences (MARE- IPLeiria), Portugal
María E. Bravo, CONICET Institute for Basic, Environmental and Applied Geosciences (IGEBA), Argentina
Copyright © 2023 Anglade, Dahl, Kristensen, Hagemann, Malzahn and Reitan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Inka Anglade, aW5rYS5zZWVrYW1wQG50bnUubm8=
†Present address: Arne M. Malzahn, Institute of Marine Ecosystem and Fishery Science, University of Hamburg, Hamburg, Germany