 Lovrenc Lipej1
Lovrenc Lipej1 Danijel Ivajnšič
Danijel Ivajnšič Petar Kružić
Petar Kružić- 1Marine Biology Station, National Institute of Biology, Piran, Slovenia
- 2Faculty of Natural Sciences and Mathematics, University of Maribor, Maribor, Slovenia
- 3Faculty of Arts, University of Maribor, Maribor, Slovenia
- 4Faculty of Science, University of Zagreb, Zagreb, Croatia
The authors studied the structure of the coastal fish assemblage in a temperate coral reef within a marine protected area in order to provide a baseline information on the occurrence and temporal distribution of fish and to highlight the importance of the coral reef to ichthyofauna. The coastal fish assemblage was investigated at two sites in Veliko jezero (Mljet National Park) in the southern Adriatic Sea with a non-destructive SCUBA visual technique in the period from 2013 to 2021. Altogether, 38 fish taxa were recorded on the right bank (coral reef) and 36 species on the left bank. The presence of the coral reef at a depth range between 9 m to 12 m is the main factor differentiating the fish fauna in these two areas, which are otherwise governed by the same environmental factors. At the coral reef a decrease in fish diversity was discovered with a steady regression from 2013 to 2021.
1 Introduction
Coral reefs are among the most diversified and productive ecosystems and host an outstanding fish fauna (Sale et al., 1984). Scleractinian corals have long been recognized as attracting rich ichthyofauna (Coker et al., 2013) since they provide shelter and microhabitats to a wide array of fishes. The structural and topographical complexity of corals contributes to the diversity and abundance of the fish associated with them and utilizing their resources (Schrandt and Lema, 2011; Graham & Nash, 2013). At the same time, many fish species that use coral reefs as habitat themselves can also have positive effects on the corals, for instance by enhancing the transfer of nutrients or, in the case of herbivore fishes, facilitating coral suppression on the overgrowth of macroalgae (Shaver and Silliman, 2017).
In the Mediterranean Sea Mediterranean stony coral Cladocora caespitosa (Linnaeus, 1758) is a well-known habitat builder, physiologically and morphologically similar to the typical tropical reef-building corals (Zibrowius, 1980; Peirano et al., 1998) known to create large colonies up to 4 m in diameter (Kružić and Požar-Domac, 2002). According to Kružić et al. (2014), C. caespitosa is well adapted to the marked temperature seasonality of the Mediterranean Sea, but extreme seawater temperatures related to global warming have an evident impact on the growth and survival of this endemic coral in the Mediterranean. A growing number of papers have correlated the increase in seawater temperature with increased biological stress and mortality events associated with C. caespitosa in different areas of the Mediterranean (Rodolfo-Metalpa et al., 2011; Kružić et al., 2012; Kersting et al., 2013; Jiménez et al., 2014). Indeed, during the summer of 1997, a prolonged period of high temperatures (93 days with water temperature above 28°C) caused polyp necrosis in 10% of the Mljet bank colonies (Kružić et al., 2014). Mediterranean stony coral with its topographic complexity has been recognized as an important habitat builder for many benthic invertebrates (Koukouras et al., 1998; Pitacco et al., 2017, 2019).
As pointed out in many studies, the coastal ichthyofauna could be considered a good indicator of the state of the environment (Bell, 1983; Harmelin et al., 1995; Lipej et al., 2003; Orlando-Bonaca and Lipej, 2005). Many species of coastal fish are in fact resident and because of their habitat preferences, are almost always expected in a specific microhabitat type. The ichthyofauna of the C. caespitosa bed or banks has attracted little scientific attention. Among the few available sources is the research performed by Pitacco et al. (2019), who studied the fish fauna on beds of C. caespitosa in different locations along the Slovenian coast (Gulf of Trieste). The ichthyofauna of Veliko jezero has also received scarce scientific attention, since previous studies considered fish fauna only partially (Graham et al., 2009; Kraljević and Tutman, 2011).
Since tropical coral reefs are known to harbor remarkably high biodiversity in term of fish fauna (Sale et al., 1984; Hixon, 2011), it would be interesting to establish whether this is also true for the Mediterranean temperate coral reef. Based on these facts, and to gain insight into the structure of fish assemblages in a temperate coral reef, the purpose of this paper is to provide baseline information on the occurrence and distribution of fishes on the temperate coral reef in Veliko jezero and to highlight the importance of coral reefs for coastal ichthyofauna. The present study provides information on the occurrence and distribution of reef fishes in Veliko jezero, part of the Mljet National Park, based on systematic underwater surveys conducted in two selected areas. In addition, two research questions were addressed: did the diurnally active fish fauna on the Cladocora caespitosa coral reef differ from that in the surrounding environment covered with macroalgal vegetation along the depth gradient, and did this change over time?
2 Materials and methods
2.1 Study area
This research was conducted in Veliko jezero (42°46’11.1”N; 17°22’25.3”E), a part of the Mljet National Park in the southern Adriatic Sea (Croatia), which is the oldest marine protected area in the Mediterranean, established on November 11th, 1960 (Kružić, 2002). Veliko jezero was formed during the Mesozoic Era as a karstic depression filled with fresh water (Govorčin et al., 2001). The lake connects with the open sea through a narrow channel (10 m wide and 2.5 m deep), which seems to impact the main ecological characteristics of both the Veliko and Malo jezero seawater lakes (Benović et al., 2000), while the impact of freshwater input is negligible. Veliko jezero covers approximately 24 ha and reaches its greatest depth at 46 m. Salinity is rather high (38.1 - 38.4‰), with small annual oscillations (Kružić, 2002). Near the channel connecting with the sea, a coral reef is located, created by Cladocora caespitosa with a coverage of 650 m2, which makes it one of the biggest coral reefs in the Mediterranean Sea (Kružić and Požar-Domac, 2003). The reef extends at a depth range from 4 to 18 m; however, the main, uninterrupted reef is present between 6 and 12 m. Coral reefs benefit from optimal temperature, strong currents, and the related high organic input (Kružić, 2002).
2.2 Fish assemblage
The sampling of the fish fauna in the Veliko jezero was previously authorized by the authorities of the Mljet National Park. Data on the fish fauna were collected by underwater visual censuses (UVC) performed using SCUBA diving (Harmelin-Vivien et al., 1985; Harmelin-Vivien and Francour, 1992; Patzner and Serrao Santos, 1993; Lipej et al., 2003; Orlando-Bonaca and Lipej, 2005; Lipej et al., 2023). UVC are suitable in marine protected areas since it constitutes a non-destructive method (Harmelin et al., 1995).
The use of UVC is still the most practical and widely used method to assess coral fish fauna (Nagelkerken et al., 2000), especially since its impact on the environment is negligible (Mazzoldi and Girolamo, 1998). Such techniques have been used extensively in MPAs because they are non-destructive and guarantee that the fish community is not affected by sampling (Arcas et al., 2023). In addition, they are particularly suitable in the case of complex substrates, where other techniques cannot be used (Colton and Swearer, 2010), and they are also flexible for implementing different sampling designs in the field (García-Charton et al., 2000) and usually cost-effective (Prato et al., 2017).
For comparing the differences in coastal ichthyofauna between the temperate coral reef and neighboring habitat types, a sampling area characterized by macroalgal coverage was chosen. Fish fauna in Veliko jezero (Mljet National Park) was investigated in the summer period from 2013 to 2021 (2013, 2014, 2015, 2016, 2017 and 2021) in two different areas/environments of Veliko jezero: (a) the temperate coral reef comprising Cladocora caespitosa on the right side of the lake, and (b) the rocky bank on the left side, an area dominated by rocks and massive boulders with a rich algal community (Figure 1), and epifauna in deep layers. In both areas, the rocky bottom changed to a sandy sedimentary bed with a few isolated rocks or coral colonies.
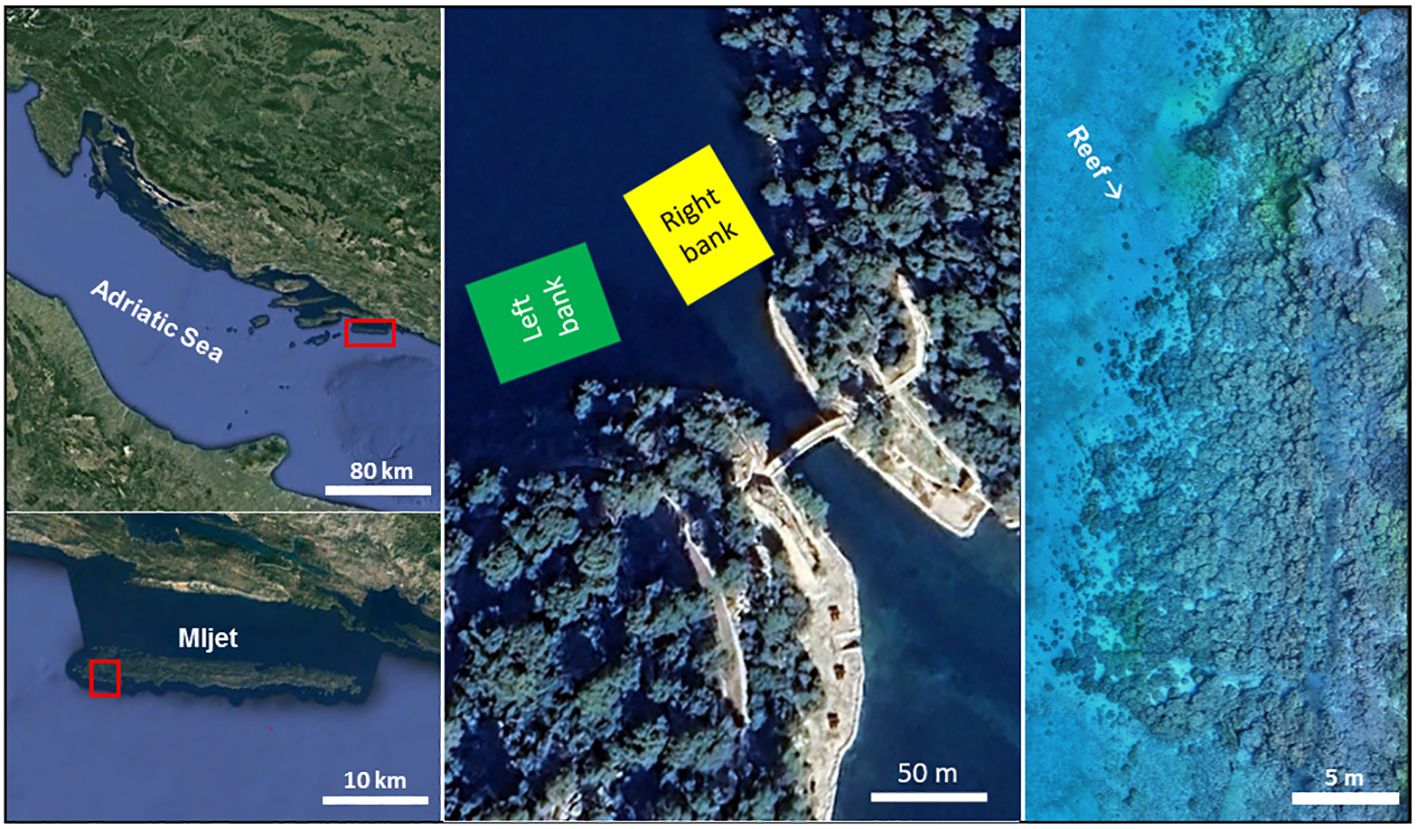
Figure 1 The locations of both study areas inside Veliko jezero in the middle (Google, (2023)), its geographical position in the Adriatic Sea (left; Google, (2023)) and a photogrammetric model of the coral reef (depth range between 9 and 12 m) within the right bank (Rb).
The UVC were performed by a skilled diver who swam along a 50 m long, and 1 m wide, horizontal transects at a constant speed (Marconato et al., 1996; Guidetti and Bussotti, 2000). Only diurnally active fish, bigger than 2 cm were counted. All censuses were carried out during the day (between 10:00 h and 16:00 h). In all transects the visibility was good. Sampling was performed by 2 divers. The second diver filmed the transect in order to obtain information on habitat coverage along the transects. Transects were recorded each year in both environments at depths of 3, 6, 9, 12 and 15 m. Thus, altogether 144 transects (rb= reef = 74, lb =left bank = 70) were performed between 2013 and 2021. To test the sampling efficacy, visual transects were performed on the coral reef at the depths of 3, 6 and 9 m from 2012. Fish species were identified mostly visually; in the case of difficulties, the movies taken simultaneously during the transects were accurately analyzed.
2.3 Data analysis
In the first step, all collected data were analyzed and organized into a comma-separated file (CSV) matrix with the following attributes: transect identification number (ID), date (as factor [DMY]), environment (as factor [reef or left bank]), depth (as factor [3, 6, 9, 12, 15 m]), substrate cover in % (Ccae = dense Cladocora caespitosa stands; Cy3= rocks with dense algal vegetation; Cy6 = boulders with dense algal vegetation) and type in % (K1 = bare stones 0.5-10 cm; K2 = bare stones 10-30 cm; P1 = rough sand <0.5 cm), fish species name and abundance. Next, the CSV file was enriched by calculating the following variables: number of fish species per transect (N), fish density per transect (D) and fish diversity (SI) per transect. The latter was determined by applying the Shannon-Wiener Diversity index: SI= −Σ piln(pi), where pi is the relative abundance of each species calculated as the proportion of individuals of a given species to the total number of individuals in the community (Shannon and Weaver, 1949). In the initial step of data analysis, the environment and depth factor variables were used to test potential differences in substrate/habitat type structure along the depth gradient. Mosaic plots were created by using the vcd package (Mayer et al., 2020) and the assoc and mosaic algorithms in the R statistical environment (R Core Team, 2023). To evaluate the interaction between a given fish community and the substrate cover/type (Ccae, Cy3, Cy6, K1, K2, and P1), Canonical Correspondence Analysis (CCA) and the Redundancy analysis (RDA) were performed. The significance of constraints in the CCA models (by = “term”) were additionally tested with the anova function within vegan. It automatically chooses the correct variant for the results of constrained ordination.
To test the assumption of fish species composition and abundance inequality between the coral reef and the algal dominated left bank (concerning the factors year and depth), a multivariate permutation analysis of variance (PERMANOVA; 999 permutations) was performed via the vegan package (Oksanen et al., 2023) in the R environment (R Core Team, 2023). Prior to that, the data were adequately reorganized to meet the basic requirements of this analysis. The weight of abundant fish species was reduced by applying the decostand function. The Euclidean distances between samples in the multidimensional space were thus simultaneously preserved (Legendre and Gallagher, 2001). However, control tests regarding homogeneity of group/factor and permutation dispersion (the betadisper and permutest algorithms) were performed in the following steps. In the case of both dispersion tests yielding insignificance (p>α; α=0.05), the simper function was implemented to identify key contributors of fish community differentiation along the coral reef-left bank environmental contrast. This step was repeated for the start and end of the survey (2013 vs 2021) concerning factor depth (especially for 9 and 12 m) since potential temporal changes in fish community composition and abundance (species turnover) needed to be identified and measured.
In the second analytical part of the study, the focus turned towards fish density and diversity trends in the two Veliko jezero environments (coral reef vs. algal dominated background). Differences in fish density (log transformed) and diversity (SI) by factors environment (reef, left bank), year (2013, 2014, 2015, 2016, 2017 and 2021), and depth (3, 6, 9, 12 and 15 m), were investigated with generalized linear models (GLMs) in the R statistical environment. Additional attention was paid to the 9 and 12 m depth categories where the environmental contrast coral reef-algal background was most pronounced. Differences in fish density (log transformed) and diversity were here tested with simple one-way ANOVA analysis and the corresponding post-hoc test (Tukey Contrasts) considering year as a factor.
3 Results
3.1 Fish species composition and abundance
Analysis of the fish assemblage at these two sites revealed a rather rich diversity of coastal fishes. The 144 visual censuses (74 at the reef and 70 at the left bank) registered a total of 23.486 specimens, expressed in terms of density per 100 m2. The great majority of this abundance comprise specimens of damselfish (Chromis chromis), 65.38% on the right bank and 54.73% on the left bank. Altogether 38 different fish taxa were recorded on the right bank and 36 different species on the left bank (Tables 1A, B). Thirty-four species occurred on both sides, while seven species were present only on the right bank and two species only on the left bank. The number of species per transect on the right bank varied from 5 to 15, on average 8.86 species per 100 m2, while the density per transects varied from 40 to 580 ind./100 m2, on average 189.4 ind./100 m2. If we do not take the damselfish Chromis chromis into consideration the average density becomes 65.57 ind./100 m2.
The number of species per transect on the left bank varied from 3 to 13, on average 8.28 species per 100 m2, while the density per transect varied from 14 to 382 ind./100 m2, on average 135.06 ind./100 m2. If we do not take the damselfish Chromis chromis into consideration, the average density becomes 61.14 ind./100 m2. The highest number of species was represented by sparids (7), followed by labrids, serranids and gobiids (6 each).
In term of frequency of occurrence, the most common species were C. chromis, Coris julis, Serranus scriba, Gobius fallax and Parablennius rouxi (Tables 1A, B). Both species of groupers, Epinephelus marginatus (mean FO% = 49.51) and E. costae (mean FO% = 30.24) were more frequent on the right bank.
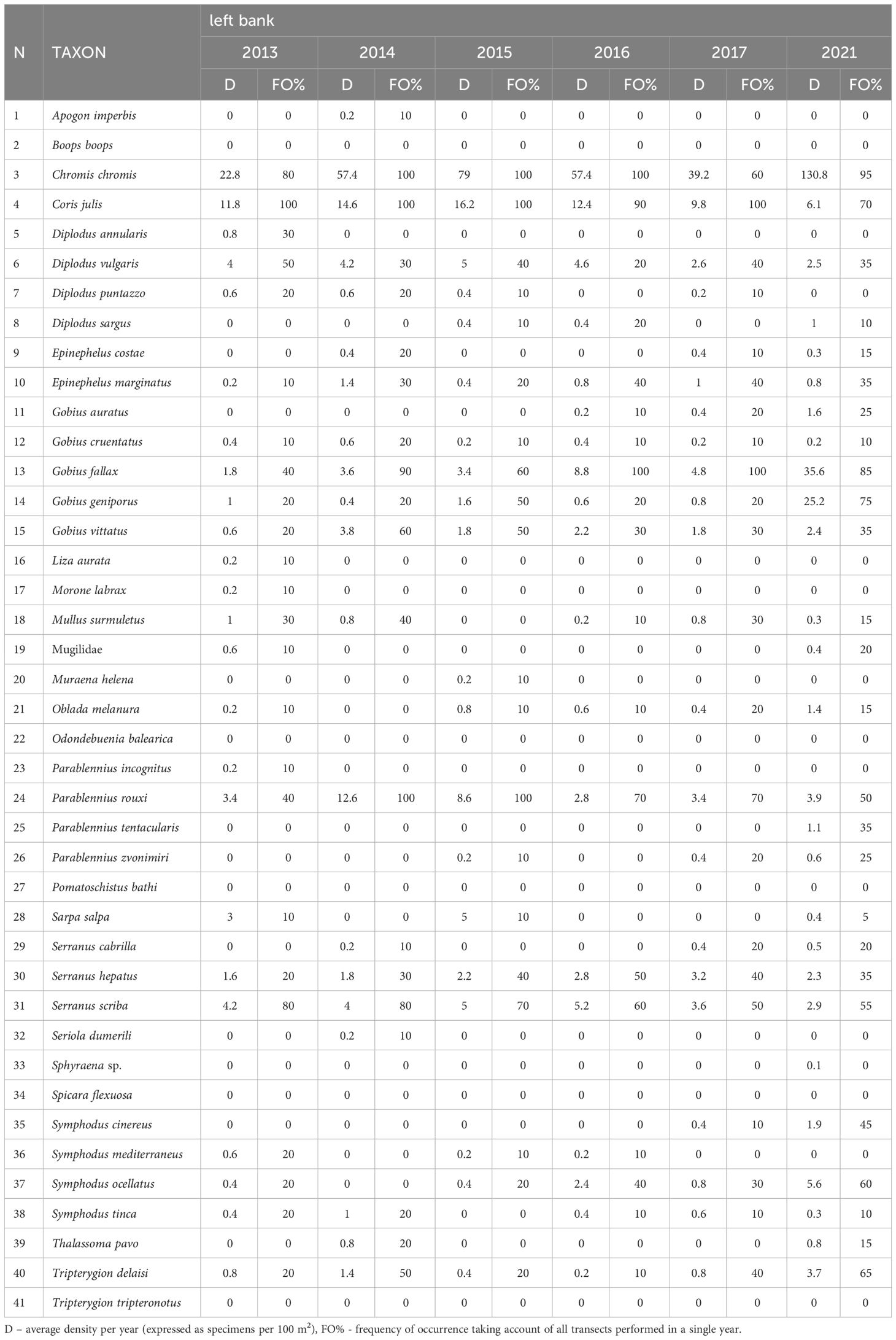
Table 1A The temporal footprint of the diurnally active fish fauna in the algal dominated (left bank).
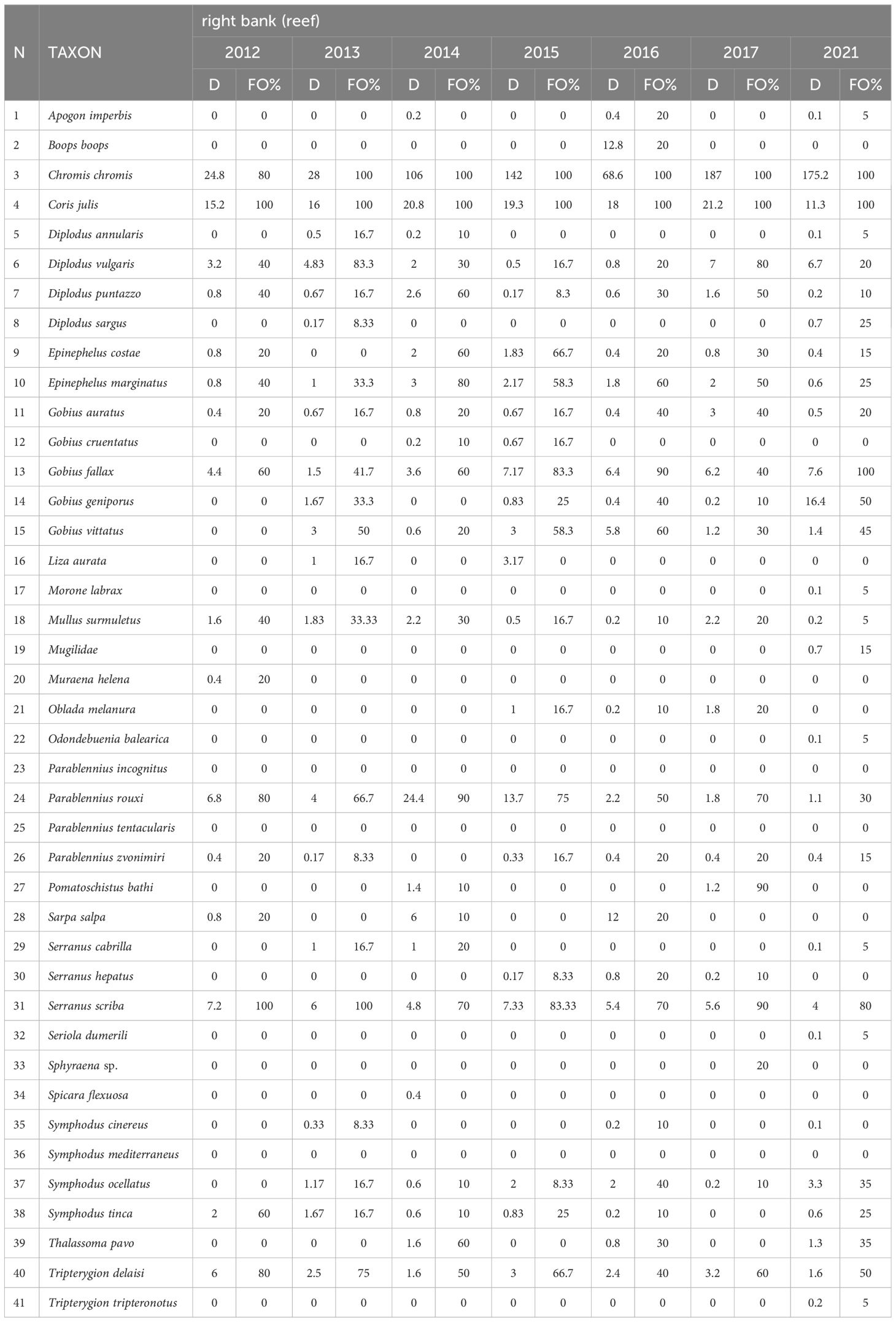
Table 1B The temporal footprint of the diurnally active fish fauna in the reef environment (right bank).
3.2 The fish community-substrate cover/type interaction
The two study areas, which are rather close together, differ significantly (p<α; α=0.05) in substrate/habitat type coverage at the 9 m and 12 m depths (marginally at 15 m, as well), especially concerning rocks and stones with vegetation, stony coral colonies, and stones with turf (Figure 2).
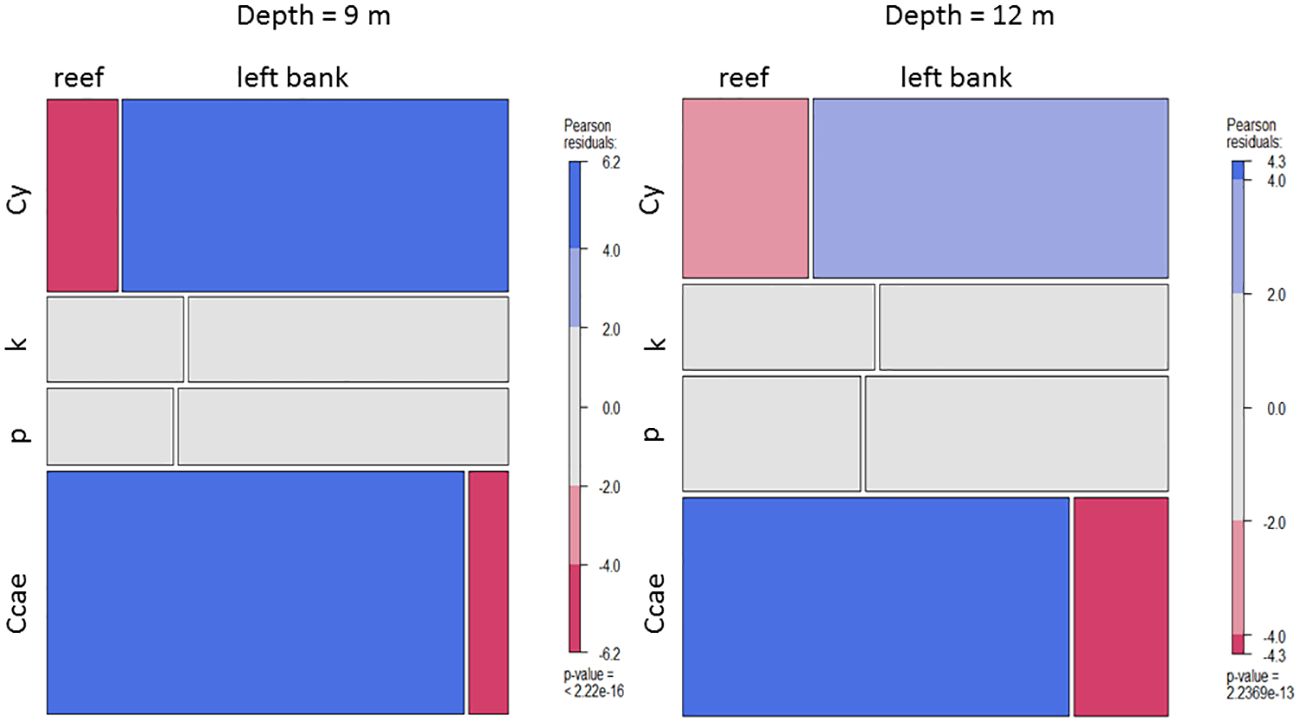
Figure 2 Mosaic plots indicating differences in substrate/habitat type coverage between the reef and the algal dominated left bank environments at 9 m and 12 m depths. Ccae, Cladocora caespitosa; P, sand; K, stones; Cy, large rocks and limestone boulders with dense algal vegetation.
The CCA ordination diagrams in Figure 3 visualize fish species’ preference for habitat types in both environments. The first two axes in the reef (right bank) case explained 55%, and in the algal case (left bank) 53% of the variability among 38 or 36 fish species respectively. However, both CCA models were statistically significant (p<α; α=0.05) concerning constraint ordination.
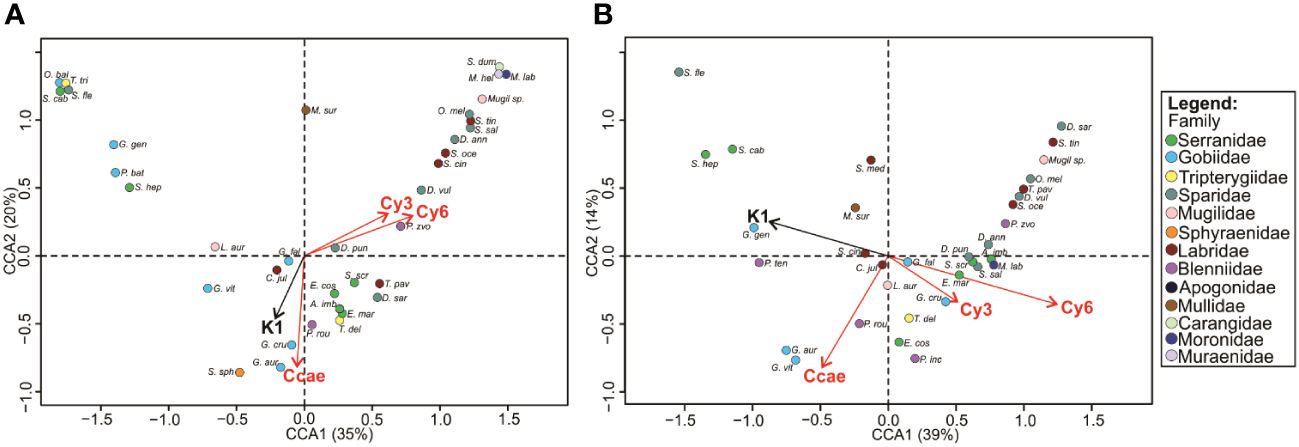
Figure 3 CCA ordination diagrams indicating the link between diurnally active fish fauna and substrate (black arrow)/habitat (red arrows) type on the reef (A); (the first axis loads 35% of variability, and the second axis 20%) and left bank (B); (the first axis loads 39% of variability, and the second axis 14%) environment. Ccae, dense Cladocora caespitosa stands; Cy3, rocks with dense algal vegetation; Cy6, boulder with dense algal vegetation; K1, bare stones 0.5-10 cm; K2, bare stones 10-30 cm; P1, rough sand <0.5 cm.
A clear distinction emerged between coral reef linked fish species (right bank) and those fish that prefer an algal dominated environment (left bank). This is especially true for many labrids (species of the genus Symphodus) and some sparids (Diplodus annularis, D. vulgaris and Oblada melanura). Fish species such as groupers (genus Epinephelus), painted comber (Serranus scriba), rainbow wrasse (Coris julis) and ornate wrasse (Thalassoma pavo) are more closely related to the C. caespitosa dominated bank (right bank). In the substrate type case, a similar fish community-dividing pattern can be observed between bare stone and sand dominated environments. Gobius geniporus, Pomatoschistus bathi, Serranus hepatus, S. cabrilla and some other inhabitants of bare, sandy habitats were found on both banks.
3.3 Differences in fish species composition and abundance – the environment, depth and time perspectives
Comparing the right bank (with the coral reef) fish species community against the algal dominated left bank environment along the depth gradient (3 to 15 m), shows evident differences in the species composition and abundance levels (Figure 4A). However, these statistical significances should be considered with caution since within-group/factor variability and permutation dispersion tests indicated differences, as well. The species contributing most to the detected pattern were Coris julis, Gobius fallax, Serranus hepatus, Mullus surmuletus, Epinephelus costae and Symphodus cinereus. Moreover, this comparison gained in importance and significance as soon as we considered depths where the coral reef versus left bank contrast was magnified. Figure 4B visualizes fish species composition and abundance differences at depths between 9 m and 12 m. Here, the betadisper and permutest results were not significant, and the PERMANOVA analysis could thus be fully trusted. According to the simper function output, these fish species statistically significantly contributed to the detected differences between the two environments: Coris julis (>reef), Serranus scriba (>reef), Gobius geniporus (>left bank), Diplodus vulgaris (>reef), Epinephelus marginatus (>reef), Serranus hepatus (>left bank), Sarpa salpa (only >left bank) and Mullus surmuletus (>left bank).
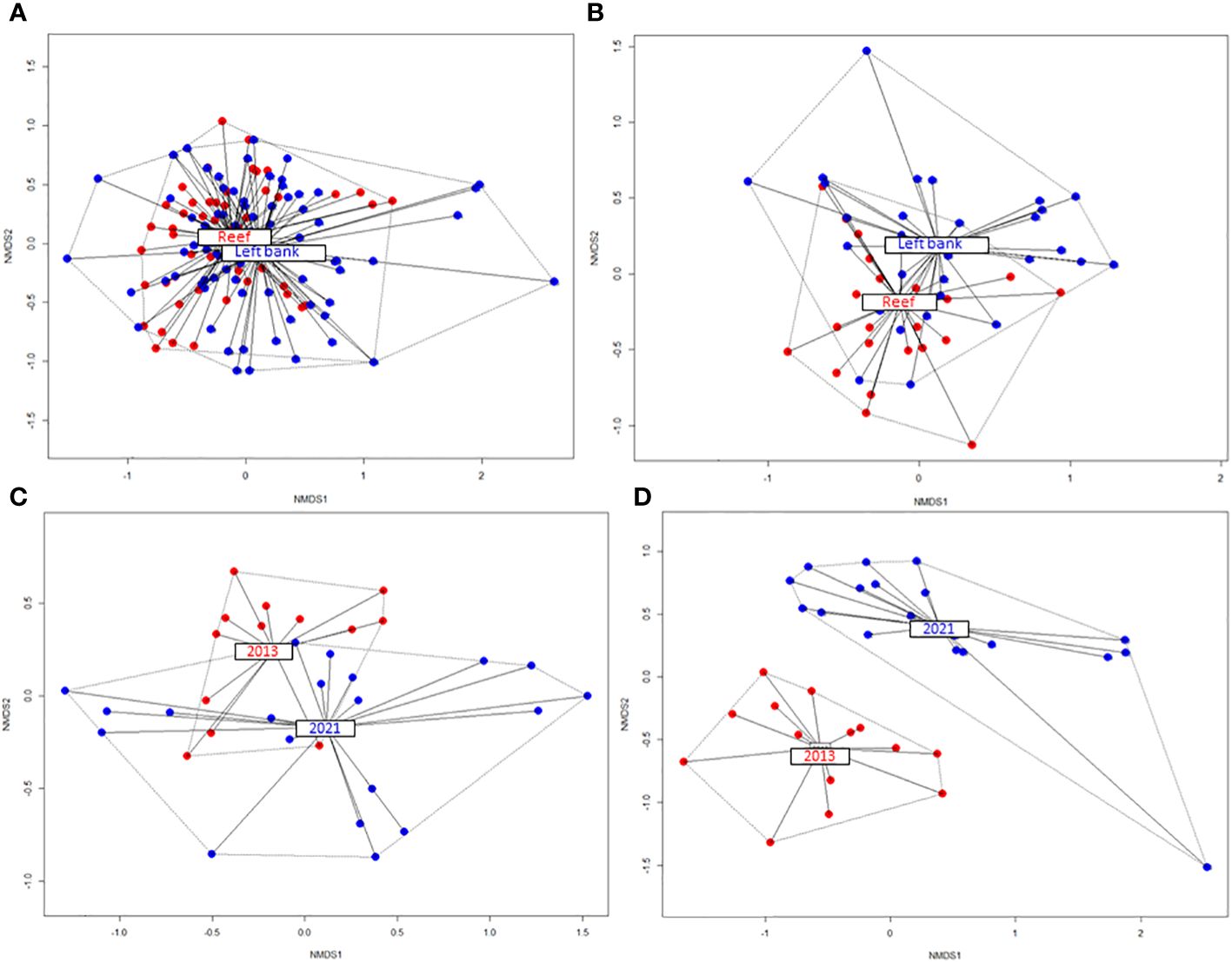
Figure 4 Fish species composition and abundance difference between the right bank (reef) and the algal dominated left bank in the NMDS space (A); the intensified contrast between the two environments in fish species composition and abundance at a 9 m to 12 m depth (B); the species composition shift between 2013 and 2021 on the reef (C) and left bank environment (D).
From the temporal perspective (Figures 4C, D), our results indicated shifts in fish species composition from 2013 to 2021, between depths of 3 m and 15 m, especially in the algal dominated background. While the coral reef area still has some fish faunal overlapping in the NMDS space (with differences in group and permutation dispersion), the left bank experienced instead a clear and significant temporal fish shift.
3.4 Environment-depth-time dynamics in fish density and diversity
The GLM effect plots in Figure 5 summarize the influence of the depth and time factors on the dependent variables of fish density (log transformed) and diversity (SI) on the coral reef and in the algal dominated background environments of Veliko jezero. In both environments, fish density significantly decreased with depth up to a 9 m of depth. Differences emerged between 9 m and 15 m in depth, where on the reef, fish density steadily increased (but never reaching the values found at the 3 m depth), whereas in the algal environment, fish density increased in the 12 m depth category and then decreased to reach its minimum in the deepest parts (15 m). On the reef side, fish density at 9 m of depth differ significantly from other depths. In the algal environment, fish density at depths of 9 m and 15 m differs from other depths. Analysis of the temporal development of fish density showed that, both environmental contrasts yielded a similar sinusoid pattern (one-year turning point difference), with a higher amplitude in the algal background (left bank).

Figure 5 GLM effect plots indicating the influence of depth and time on dependent variables: log-reformed fish density (Dlog) and diversity (SI) on the reef (A) and in the left bank environment (B).
Fish diversity decreased even more rapidly with the depth gradient. The 9 m depth category represented the turning point in both environments. On the coral reef side, fish species richness increased from this point on, to reach, at a 15 m depth, a similar value as at a 6 m depth. On the left bank, fish diversity rapidly increased from 9 m to 12 m depths, followed then by a decrease towards the 15 m depth, where it reached diversity values comparable to those for 9 m depth. Moreover, on the coral reef side, fish diversity significantly differed for each depth category. On the left bank, the 12 m depth category represented an exception since it had SI values comparable with those for the 3 m depth category.
The temporal pattern in fish diversity on the coral reef side indicated a clear negative and almost linear behavior. Fish diversity decreased each year, whereas on the algal background side, the fish diversity temporal dynamics cannot be described with a linear function. Fluctuations in SI values were evident, but when comparing the start and end of the survey, the difference was significant and the trend clearly negative (fish diversity significantly decreased).
3.5 The critical 9 m to 12 m depths – species turnover and evident biodiversity loss
The temporal trend of fish diversity between 9 m and 12 m in depth showed some differences between the two compared environments. A marginally significant (p=0.061; α=0.05) species composition change was detected in both environments (coral reef and algal dominated background) in only 9 years (the betadisper and permutest algorithm results were insignificant in both cases) (Figures 6A, B). However, this process seems to be less intensive on the coral reef side compared to the algal background.
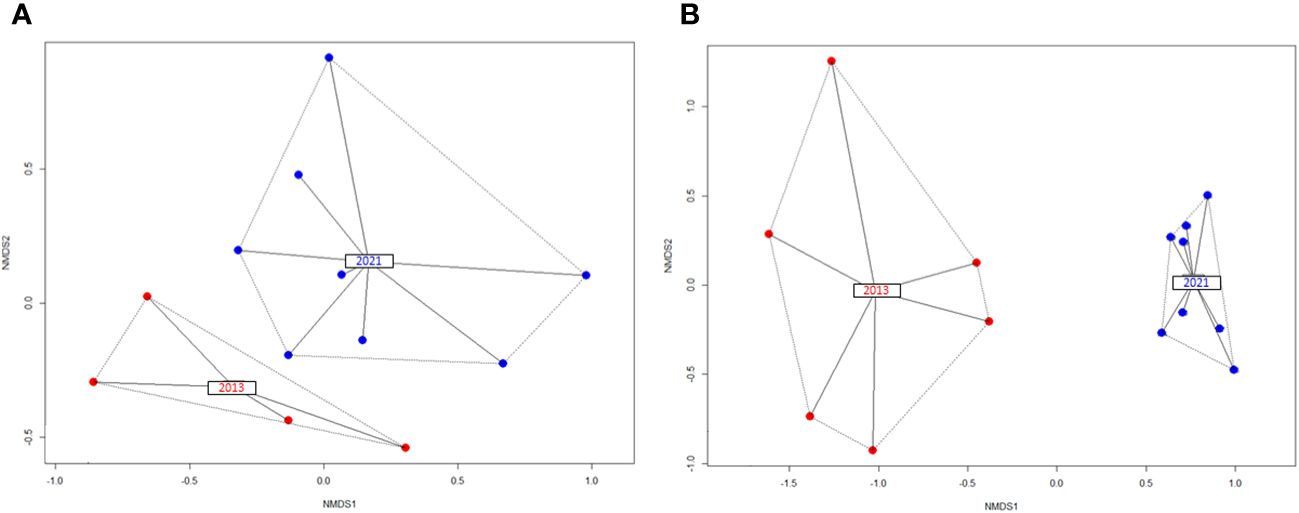
Figure 6 Fish species composition shift in the NMDS space between the years 2013 and 2021 at depths from 9 m to 12 m on the reef (A) and in the left bank environment (B).
What about fish density and diversity in the 9 m to 12 m depth zone? As shown in Figure 7, fish density did fluctuate over time but did not show any clear trend during the given time span (2013 – 2021) in either environment. The temporal trend of fish diversity differed between the coral reef (with a heavy predominance of C. caespitosa) and the left bank. On the reef side, fish density clearly decreased, with a practically steady regression from 2013 to 2021. In the algal dominated background between 9 m and 12 m in depth, fish diversity decreased till 2016, but from then on, it steadily increased; however, differences were not significant. An inverse relation between fish density and diversity was nonetheless evident in this algal dominated environment.
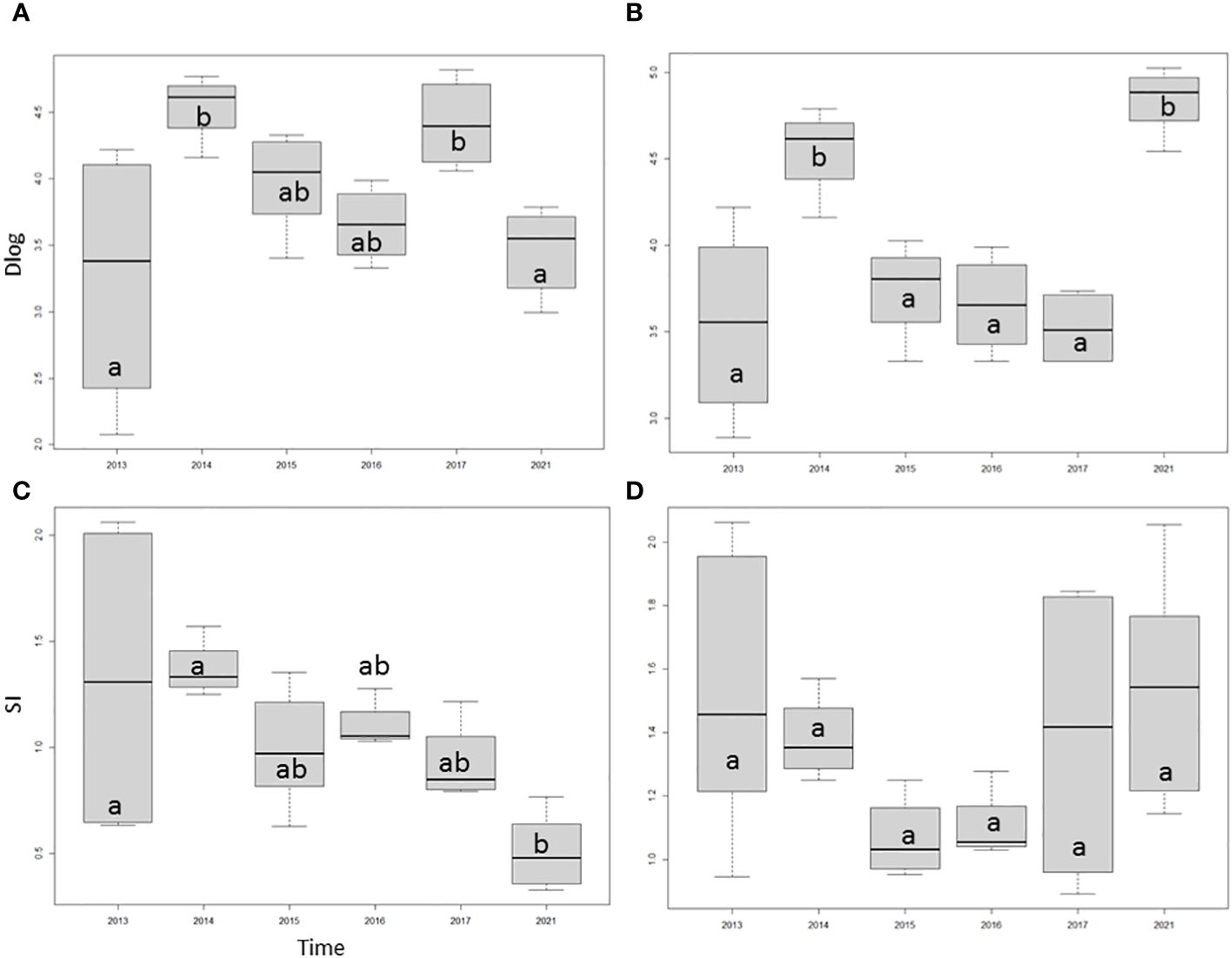
Figure 7 Temporal development of log-transformed fish density (Dlog) and diversity (SI) at the 9 m to 12 m depth belt on the reef (A) and in the algal dominated background (left bank) environment (B). Small letters indicate statistically significant differences concerning Tukey Contrasts (p<α; α=0.05).
4 Discussion
4.1 Habitat diversity
Habitats with high substratum complexity were found to be inhabited by a more diverse fish community than that found on bare gravel or sedimentary bottoms (Macpherson, 1994; Bartholomew et al., 2000; Almany, 2004) since they provide breeding sites, shelter and feeding opportunities. The Mediterranean stony coral is an important habitat builder and provides niches for many tiny invertebrates in the species between corallites (endofauna) or on the surface of the colony (epifauna). The strong sea currents, which occur because of tidal exchange in the channel, appear to favor the growth of the coral reef (see Kružić and Benković, 2008). The bottom sea current in the channel during high tide is stronger than during low tide, and that could be why there are more small colonies in the part of the coral reef towards the inner side of Veliko jezero. Microhabitats within the corallites (inside colonies) are characterized by high diversity and an abundance of many benthic invertebrates (sensu Bettoso et al., 2023). In the study of Pitacco et al. (2019), who investigated the relationship between C. caespitosa, they showed that the number of invertebrate taxa associated with C. caespitosa colonies increases with the colony size, following the SAR equation proposed by Arrhenius (1921). At least 222 invertebrate taxa were found in association with coral colonies in the Gulf of Trieste (Pitacco et al., 2017). The biggest colony in the Gulf of Trieste, measuring about 68 cm in diameter (Zunino et al., 2018), rather smaller, but comparable to the size of most colonies on the Mljet reef, is estimated to host a total of 130 different taxa, predominantly comprising molluscs (39 taxa) and polychaetes (61 taxa). Feeding guilds of coastal fish that specialize in tiny invertebrates probably exploit the feeding potential provided by large colonies of stony coral. In addition, as already discovered for tropical coral reefs, the more complex the corals, the more spaces are available to provide refuge for more fish, and the more niches are provided for various fish species, which decreases interspecies competition (Almany, 2004; Johnson, 2007).
4.2 Fish diversity
The analysis of the fish assemblages on the coral reef and in the adjacent area revealed a rather rich diversity of coastal fish fauna. Previously Graham et al. (2009) published a list of 55 taxa, present in both lakes inside the Mljet National Park. However, data on the occurrence (date, abundance, depth) of each fish species are missing and some doubtful species such as the hammerhead shark (Sphyrna mokkaran) were mentioned. The great majority of other species were also observed in our study, but outside the performed transects. Kraljević and Tutman (2011) mentioned 47 species of juvenile fish in the coves of Mljet National Park. In Veliko jezero they found 12 juvenile species, while at the Soline site, which is the entrance point of the channel connecting Veliko jezero with the open sea, the number of species caught was 37. This number is comparable to the numbers yielded by our study.
In their study of the juvenile fish community in the Mljet National Park Kraljević and Tutman (2011) pointed out that habitat diversity was the main factor in structuring the more diverse fish community. The dominance of juvenile fish highlighted the importance of these two coves inside the Mljet National Park as nursery areas and feeding grounds for many different fish species (Dulčić et al., 1997). Previously, a study of the importance of various Cladocora caespitosa beds for coastal fish fauna was performed in the Slovenian part of the Gulf of Trieste (Pitacco et al., 2017). The sampled areas, characterized by a high density of C. caespitosa coral colonies, hosted moderately rich fish fauna, which represented 1/20 of the 449 species recorded for the Adriatic Sea (Lipej et al., 2022). The fish community’s abundance and diversity in vegetated habitats differ from sandy areas, owing to habitat complexity and protection from predation (sensu Curley et al., 2002). However, as pointed out by Pitacco et al. (2019), fish diversity is likely to respond to habitat heterogeneity, typology of substrate and prevalence of benthic organisms, such as seaweeds and sponges; therefore, the density of coral colonies and their coverage were not appropriate predictors of species richness.
4.3 Differences between the two banks
The difference between the two banks is evident especially at the depth range from 9 m to 12 m (coral reef). At shallower depths the two banks do not differ. The main environmental factor explaining the differences between the banks is spatial heterogeneity, which is expressed in structural complexity. The coral reef (the area from 9 m to 12 m on the right bank) is less structurally differentiated than the huge boulders assessed as the biocoenoses of infralittoral photophilic algae, and at the same time it is not covered with vegetation. Wrasses (family Labridae) are known to be closely associated with rocky bottoms with dense Cystoseira s.l. cover, since they find shelter and build nests inside this vegetation type (Lipej et al., 2009). The rainbow wrasse (Coris julis) prefers less complex habitats (Cuadros et al., 2019), whereas many species of the genus Symphodus prefer more complex, generally densely vegetated habitats with abundant prey such as polychaetes and amphipods (Ruitton et al., 2000).
On the coral reef, a decrease in fish diversity with a steady regression from 2013 to 2021 was discovered, whereas on the left bank the decrease lasted until 2016, after which an increase in species richness was noted. Fish species such as groupers (Epinephelus costae, E. marginatus), painted comber (Serranus scriba), rainbow wrasse (Coris julis) and ornate wrasse (Thalassoma pavo) were more closely related to C. caespitosa. Chittaro (2002) pointed out the importance of microhabitat richness for the increase in coral fish species richness. The painted comber preys on smaller gobies, such as Gobius fallax (Lokovšek et al., 2022), which is the most common and abundant fish prey in the area. Dusky grouper (Epinephelus marginatus) is known to be a mainly piscivorous and resident predator, avoiding large movements to seek their prey (Gracia López and Castelló i Orvay, 2005). The population of the dusky grouper in the study area is composed mainly of smaller (juvenile) specimens (TL<40 cm), which probably prey on smaller fish and crustaceans (Gracia López and Castelló i Orvay, 2005).
4.4 The case of groupers
Groupers (family Epinephelidae) are large, sedentary, slow-growing fish. They are mainly piscivorous and generally reef-resident predators that play a relevant role in regulating fish populations (Howlett et al., 2016). They are economically and ecologically important predators in the rocky coast ecosystems of the Mediterranean Sea. Groupers are generally long-lived (up to 40 years) and take many years to reach sexual maturity, which makes them vulnerable to fishing prior to reproduction (Sadovy de Mitcheson et al., 2012). Owing to increased fishing pressure during recent decades, the dusky grouper (Epinephelus marginatus) has suffered a dramatic decline in the NW Mediterranean and retreats more and more to deeper bottoms (Bruslé, 1985).
Guidetti and Micheli (2011) demonstrated that the analysis of ancient art can be helpful in assessing the status of certain fish species in ancient times. By analyzing more than seventy Etruscan, Greek, and Roman mosaics, they discovered 23 of these depicting groupers, which were much more common in the past than they are today and may have had larger body sizes. A similar phenomenon was observed by Winter et al. (2022), who in the study of three archaeological sites in the Levantine Sea, discovered that groupers bigger than 80 cm were not uncommon in the fish remains obtained from excavation. According to historical evidence, they were more common at very shallow depths of 3 m and less. Nowadays, groupers are generally reported in deep waters, where they find a refuge from fishing in areas without the status of protected area (Winter et al., 2022). The density of E. marginatus seems to be positively correlated to habitat structure, as discovered by Hackradt et al. (2014) for many Spanish Mediterranean protected areas. In our study, juvenile and small size dusky groupers and golden groupers were observed at all studied depths and on both sides, with densities up to 8 ind./100 m2. Howlett et al. (2016) stated that numerous studies have found relationships between coral health and groupers at their early stages when they are more vulnerable.
According to Vacchi et al. (1999) the density of the larger groupers increases with the level of protection, since inside the protected area, they are safe from spear fishing. However, the calculated densities in their study are lower in comparison with our data. As stated by Condini et al. (2018), the creation of MPAs in the Mediterranean Sea was an important step for dusky grouper conservation. Groupers are also known to respond rapidly to the establishment of a protection regime (Claudet et al., 2010). Both studied species, which are known to be residential and territorial species (Chauvet and Francour, 1990; Pastor et al., 2009), have benefited from the protection regime in MPA. This is further evidence of the importance of the NP Mljet as an area that hosts rather high densities of groupers, which are also present in shallow waters and perhaps resemble (although to a minor extent) the situation present in Roman times. Although cases of illegal fishing of groupers seem to occur, we did not observe any decline in density in the period 2013 to 2021 in either of the two study areas.
5 Conclusions
The present study shows that the two banks of Veliko jezero inside the National Park host a rich coastal fish assemblage. The presence of the coral reef mainly at a depth range between 9 m to 12 m is the main factor differentiating the two areas, which are otherwise governed by the same environmental factors. The reef, comprising huge colonies of Cladocora caespitosa, hosts different densities of certain fishes, among which are also two grouper species (E. costae, E. marginatus). The main factors that structure species richness are spatial heterogeneity, vegetation cover and currents. The trend in fish diversity for the period 2013 to 2021 shows an evident decrease in species richness, especially on the right bank. Among many benefits of MPAs they have been shown to protect apex predators that benefit stability to coastal habitats (Rooney et al., 2006; Pettersen et al., 2021) which is evident also in Veliko jezero in the case of groupers. According to the data obtained we may speculate that the marine protected area Veliko jezero maintain high fish diversity and abundance, however, a regular monitoring would ellucidate which specific factors caused the decrease in species richness in recent years.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements. Since in this study we used non-destructive and non-invasive visual census methods for the assessment of fish density, ethical approval was not needed.
Author contributions
LL: Conceptualization, Data curation, Writing – original draft, Writing – review & editing. DI: Conceptualization, Methodology, Writing – original draft, Writing – review & editing, Funding acquisition. VP: Data curation, Writing – original draft, Writing – review & editing. BM: Data curation, Validation, Funding acquisition, Writing – original draft, Writing – review & editing. DT: Data curation, Validation, Writing – original draft, Writing – review & editing. PK: Conceptualization, Data curation, Writing – original draft, Writing – review & editing, Funding acquisition.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was co-financed by the Slovenian Research Agency (ARIS, grant numbers P1-0237 [Coastal Sea Research] and P6-0372 [Slovene Identity and Cultural Awareness in Linguistic and Ethnic Contact Areas in Past and Present]) and by activities included in the ADRICOR project, funded by the Croatian Science Fund (HRZZ) (grant number lP-2O19-04), and the project Development of Research Infrastructure for the International Competitiveness of the Slovenian RRI Space—RI-SI-LifeWatch, co-financed by the Republic of Slovenia, Ministry of Education, Science and Sport and the European Union from the European Regional Development Fund.
Acknowledgments
This study was conceptualized during the agreed three-month visit (9.1. to 11.4.2023) of the co-author Danijel Ivajnšič to the Department of Biology, Faculty of Science, University of Zagreb. Moreover, the authors would like to express their sincere gratitude to the team of the director, Mr. Sršen and the scientific team of the Mljet National Park for providing us the opportunity to perform a long-term study of the coastal fish fauna of the Veliko jezero and adjacent areas and for their immense support and help. Special thanks also to many researchers for field assistance, including Tihomir Makovec, Leon Lojze Zamuda, Dr. Nicola Bettoso, Dr. Martin Vodopivec, Dr. Roberto Odorico, Dr. Vlado Onofri, Jelena Belamarić, and many others. We would also like to thank Dr. Branko Dragičević from the Institute for Oceanography and Fisheries, who shared with us some remarks about the identification of species from the genus Sphyraena.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Almany G. R. (2004). Differential effects of habitat complexity, predators and competitors on abundance of juvenile and adult coral reef fishes. Oecologia 141, 105–113. doi: 10.1007/s00442-004-1617-0
Arcas E., Valle C., Forcada A. (2023). Evaluation of the economic crisis on the conservation of the ichthyofauna in Marine Protected Areas. Mar. Policy 147, 105347. doi: 10.1016/j.marpol.2022.105347
Bartholomew A., Diaz R. J., Cicchetti G. (2000). New dimensionless indices of structural habitat complexity: predicted and actual effects on a predator’s foraging success. Mar. Ecol. Progr. Ser. 206, 45–58. doi: 10.3354/meps206045
Bell J. D. (1983). Effects of depth and marine reserve fishing restrictions on the structure of a rocky reef fish assemblage in the north-western Mediterranean Sea. J.Appl. Ecol. 20, 357–369. doi: 10.2307/2403513
Benović A., Lučić D., Onofri V., Peharda M., Carić M., Jasprica N., et al. (2000). Ecological characteristics of the Mljet Island seawater lakes (South Adriatic Sea) with special reference to their resident populations of medusae. Scientia marina 64, 197–206. doi: 10.3989/scimar.2000.64s1197
Bettoso N., Faresi L., Pitacco V., Orlando-Bonaca M., Aleffi I. F., Lipej L. (2023). Species richness of benthic macrofauna on rocky outcrops in the Adriatic Sea by using species-area relationship (SAR) tools. Water 15, 318. doi: 10.3390/w15020318
Bruslé J. (1985). Exposé synoptique des données biologiques sur les mérous Epinephelus aeneus (Geoffroy Saint Hilaire 1809) et Epinephelus guaza (Linnaeus 1758) de l’océan Atlantique et de la Méditerranée Vol. 129 (Synopsis sur les pêches: FAO). 64 pp.
Chauvet C., Francour P. (1990). Les mérous du parc national de Port-Cros: aspects socio-démographiques. Bull. Soc Zool. France 114, 5–13.
Chittaro P. M. (2002). Species-area relationships for coral reef fish assemblages of St. Croix, US Virgin Islands. Mar. Ecol. Prog. Ser. 233, 253–261.
Claudet J., Osenberg C. W., Domenici P., Badalamenti F., Milazzo M., Falcón J. M., et al. (2010). Marine reserves: Fish life history and ecological traits matter. Ecol. Appl. 20, 830–839. doi: 10.1890/08-2131.1
Coker D. J., Wilson S. K., Pratchett M. S. (2013). Importance of live coral habitat for reef fishes. Rev. Fish Biol. Fisheries. 24 (1). doi: 10.1007/s11160-013-9319-5
Colton M. A., Swearer S. E. (2010). A comparison of two survey methods: differences between underwater visual census and baited remote underwater video. Mar. Ecol. Prog. Ser. 400, 19–36. doi: 10.3354/meps08377
Condini M. V., Garcıa-Charton J. A., Garcia A. M. (2018). A review of the biology, ecology, behavior and conservation status of the dusky grouper, Epinephelus marginatus (Lowe 1834). Rev. Fish Biol. Fisheries 28, 301–330. doi: 10.1007/s11160-017-9502-1
Cuadros A., Moranta J., Cardona Pascual L., Thiriet P., Francour P., Vidal E., et al. (2019). Juvenile fish in cystoseira forests: influence of habitat complexity and depth on fish behaviour and assemblage composition. Mediterr. Mar. Sci. 20, 380–392. doi: 10.12681/mms.18857
Curley B. G., Kingsford M. J., Gillanders B. M. (2002). Spatial and habitat-related patterns of temperate reef fish assemblages: implications for the design of marine protected areas. Mar. Freshw. Res. 53, 1197–1210. doi: 10.1071/MF01199
Dulčić J., Kraljević M., Grbec B., Pallaoro A. (1997). Composition and temporal fluctuations of inshore juvenile fish populations in the Kornati Archipelago, eastern middle Adriatic. Mar. Biol. 129, 267–277. doi: 10.1007/s002270050167
García-Charton J. A., Pérez-Ruzafa Á., Marcos-Diego C. (2000). “Fish visual census methods for detecting gradients of abundance and biomass across boundaries of MPAs,” in Goñi R., M. Harmelin-Vivien, F. Badalamenti, L. Le Diréach, G. Bernard edit.: Introductory guide to methods for selected ecological studies in marine reserves. (GIS Posidonie publ., Fr.), 1–120.
Guidetti P., Micheli F. (2011). Ancient art serving marine conservation. Front. Ecol. Environ. 9, 374–5. doi: 10.1890/11.WB.019
Google. (2023). Adriatic Sea and Mljet island. Available at: https://www.google.com/maps (Accessed: 10.12.2023).
Gracia López V., Castelló i Orvay F. (2005). Food habits of groupers Epinephelus marginatus (Lowe 1834) and Epinephelus costae (Steindachner 1878) in the Mediterranean coast of Spain. Hidrobiológica 15, 27–34.
Graham W. M., Chiaverano L., d’Ambra I., Mianzan H., Alvarez Colombo G., Acha M., et al. (2009). Fish and jellyfish: using the isolated marine “lakes“ of Mljet Island, Croatia, to explore larger marine ecosystem-based management approaches. Annales Istr. Med. Stud. 19, 39–48.
Graham N. A. J., Nash K. L. (2013). The importance of structural complexity in coral reef ecosystems. Coral Reefs 32), 315–326. doi: 10.1007/s00338-012-0984-y
Guidetti P., Bussotti S. (2000). Nearshore fish assemblages associated with shallow rocky habitats along the southern Croatian coast (Eastern Adriatic Sea). Vie Milieu Life Environ. 50, 171–176.
Guidetti P., Micheli F. (2011). Ancient art serving marine conservation. Front. Ecol. Environ. 9, 374–5. doi: 10.1890/11.WB.019
Hackradt C. W., García-Charton J. A., Harmelin-Vivien M., Pérez-Ruzafa Á., Le Direach L., Bayle-Sempere B., et al. (2014). Response of rocky reef top predators (Serranidae: Epinephelinae) in and around marine protected areas in the western Mediterranean Sea. PloS One 9, e98206. doi: 10.1371/journal.pone.0098206
Harmelin-Vivien M. L., Francour P. (1992). Trawling or visual censuses? Methodological bias in the assessment of fish populations in seagrass beds. Mar. Ecol. 13, 41–51. doi: 10.1111/j.1439-0485.1992.tb00338.x
Harmelin-Vivien M., Harmelin J., Chauvet C., Duval C., Galzin R., Lejeune P., et al. (1985). Evaluation Visuelle Des Peuplements et Populations de Poissons Méthodes et Problèmes. Rev. D’ecologie Terre Vie 40, 467–539. doi: 10.3406/revec.1985.5297
Harmelin J.-G., Bachet F., Garcia F. (1995). Mediterranean Marine Reserves: Fish Indices as Test of Protection Efficiency. P. S. N. Z. I.: Marine Ecology 16 (3), 233–250.
Hixon M. A. (2011). 60 years of coral reef fish ecology: past, present, future. Bull. Mar. Sci. 87, 727–765. doi: 10.5343/bms.2010.1055
Howlett S. J., Stafford R., Waller M., Antha S., Mason-Parker C. (2016). Linking protection with the distribution of grouper and habitat quality in Seychelles. J. Mar. Biol. doi: 10.1155/2016/7851425
Jiménez C., Hadjioannou L., Petrou A., Nikolaidis A., Radoševiću M., Lange M. A., et al. (2014). Mortality of the scleractinian coral Cladocora caespitosa during a warming event in the Levantine Sea (Cyprus). Regional Environ. Change 16, 1963–1973. doi: 10.1007/s10113-014-0729-2
Johnson D. W. (2007). Habitat complexity modifies post-settlement mortality and recruitment dynamics of a marine fish. Ecology 88, 1716–1725. doi: 10.1890/06-0591.1
Kersting D. K., Bensoussan N., Linares C. (2013). Long-term responses of the endemic reef-builder Cladocora caespitosa to Mediterranean warming. PloS One 8, e70820. doi: 10.1371/journal.pone.0070820
Koukouras A., Kühlmann D., Voultsiadou E., Vafidis D., Dounas C., Chintirouglou C., et al. (1998). The macrofaunal assemblage associated with the scleractinian coral Cladocora caespitosa (L.) in the Aegean Sea. Ann. Inst. Océanogr. Paris 74, 97–114.
Kraljević M., Tutman P. (2011). Diel fluctuations of a juvenile fish assemblage in shallow coves of Mljet National Park. Proceedings of the Symposium Branimir Gušić Days. Ecol. Monogr. 9, 229–250.
Kružić P. (2002). Marine fauna of the Mljet National Park (Adriatic Sea, Croatia). 1. Anthozoa. Natura Croatica 11, 265–292.
Kružić P., Benković L. (2008). Bioconstructional features of the coral Cladocora caespitosa (Anthozoa, Scleractinia) in the Adriatic Sea (Croatia). Mar. Ecol. 29, 125–139. doi: 10.1111/j.1439-0485.2008.00220.x
Kružić P., Lipej L., Mavrič B., Rodić P. (2014). Impact of bleaching on the coral Cladocora caespitosa in the eastern Adriatic Sea. Mar. Ecol. Prog. Ser. 509, 193–202. doi: 10.3354/meps10962
Kružić P., Požar-Domac A. (2002). Skeleton growth rates of coral bank of Cladocora caespitosa (Anthozoa, Scleractinia) in lake Veliko jezero (Mljet National Park). Period. Biol. 104, 123–129.
Kružić P., Požar-Domac A. (2003). Banks of the coral Cladocora caespitosa (Anthozoa, Scleractinia) in the Adriatic Sea. Coral Reefs 22, 536. doi: 10.1007/s00338-003-0345-y
Kružić P., Sršen P., Benković L. (2012). The impact of seawater temperature on coral growth parameters of the colonial coral Cladocora caespitosa (Anthozoa, Scleractinia) in the eastern Adriatic Sea. Facies 58, 477–491. doi: 10.1007/s10347-012-0306-4
Legendre P., Gallagher E. D. (2001). Ecologically meaningful transformations for ordination of species data. Oecologia 129, 271–280. doi: 10.1007/s004420100716
Lipej L., Ivajnšič D., Mavrič B., Pitacco V., Trkov D., Orlando Bonaca M. (2023). Coastal fish fauna in the Cystoseira s.l. Algal belts experiences from the northern Adriatic Sea. J. Mar. Sci. Eng. 11, 888. doi: 10.3390/jmse11050888
Lipej L., Kovačić M., Dulčić J. (2022). An analysis of adriatic ichthyofauna—Ecology, zoogeography, and conservation status. Fishes 7, 58. doi: 10.3390/fishes7020058
Lipej L., Orlando-Bonaca M., Ozebek B., Dulčić J. (2009). Nest characteristics of three labrid species in the Gulf of Trieste (northern Adriatic Sea). Acta Adriatica 50, 139–150.
Lipej L., Orlando-Bonaca M., Šiško M. (2003). Coastal fish diversity in three marine protected areas and one unprotected area in the gulf of Trieste (Northern adriatic). P.S.Z.N.: Mar. Ecol. 24, 259–273. doi: 10.1046/j.1439-0485.2003.00843.x
Lokovšek A.,, Orlando-Bonaca M., Trkov D., Lipej L. (2022). An Insight into the Feeding Ecology of Serranus scriba, a Shallow Water Mesopredator in the Northern Adriatic Sea, with a Non-destructive Method. Fishes 7, 210
Macpherson E. (1994). Substrate utilisation in a Mediterranean fish community. Mar. Ecol. Prog. Ser. 114, 211–218. doi: 10.3354/meps114211
Marconato A., Mazzoldi C., De Girolamo M., Stefanni S., Maio G. (1996). L’uso del “Visual census”’Nello studio della fauna ittica costiera. Biol. Mar. Mediterr. 3, 512–513.
Mayer D., Zeileis A., Hornik K. (2020). VCD: Visualizing Categorical Data. R package version 1. 4–8.
Mazzoldi C., Girolamo M. (1998). Littoral fish community of the island lampedusa (Italy): A visual census approach. Ital. J. Zool. 65, 275–280. doi: 10.1080/11250009809386832
Nagelkerken I., Dorenbosch M., Verberk W. C. E. P., Cocheret de la Morinière E., van der Velde G. (2000). Day-night shifts of fishes between shallow-water biotopes of a Caribbean Bay, with emphasis on the nocturnal feeding of Haemulidae and Lutjanidae. Mar. Ecol. Prog. Ser. 194, 55–64. doi: 10.3354/meps194055
Oksanen J., Kindt R., Legendre P., O’Hara R. B., Simpson G. L., Solymos P., et al. (2023). The Vegan Package. Community Ecology Package. Available online at: http://R-Forge.r-Project.Org/Projects/Vegan/ (Accessed 1 February 2023).
Orlando-Bonaca M., Lipej L. (2005). Factors affecting habitat occupancy of fish assemblage in the Gulf of Trieste (Northern Adriatic Sea). Mar. Ecol. 26, 42–53. doi: 10.1111/j.1439-0485.2005.00037.x
Pastor J., Verdoit-Jarraya M., Astruch P., Dalias N., Nelva Pasqual J.-S., Saragoni G., et al. (2009). Acoustic telemetry survey of the dusky grouper (Epinephelus marginatus) in the Marine Reserve of Cerbère-Banyuls: informations on the territoriality of this emblematic species. Comptes Rendus Biologies 332, 732–740. doi: 10.1016/j.crvi.2009.03.010
Patzner R., Serrao Santos R. (1993). Ecology of rocky littoral fishes of the azores. Cour. Forschungsinst. Senckenb. 159, 423–427.
Peirano A., Morri C., Mastronuzzi G., Bianchi C. (1998). The coral Cladocora caespitosa (Anthozoa, Scleractinia) as a bioherm builder in the Mediterranean Sea. Memorie descrittive della Carta geologica d’Italia 52, 59–74.
Pettersen A. K., Marzinelli E. M., Steinberg P. D., Coleman M. A. (2021). Impact of marine protected areas on temporal stability of fish species diversity. Conserv. Biol. 2021, 1–11. doi: 10.1111/cobi.13815
Pitacco V., Crocetta F., Orlando Bonaca M., Mavrič B., Lipej L. (2017). The Mediterranean stony coral Cladocora caespitosa (Linnaeus 1767) as habitat provider for molluscs: colony size effect. J. Sea Res. 129, 1–11. doi: 10.1016/j.seares.2017.08.001
Pitacco V., Mistri M., Lipej L. (2019). Species-Area Relationship (SAR) models as tools for estimating faunal biodiversity associated with habitat builder species in sensitive areas: the case of the Mediterranean stony coral (Cladocora caespitosa). Mar. Environ. Res. 149, 27–39. doi: 10.1016/j.marenvres.2019.05.016
Prato G., Thiriet P., Di Franco A., Francour P. (2017). Enhancing fish Underwater Visual Census to move forward assessment of fish assemblages: An application in three Mediterranean Marine Protected Areas. PloS One 12, e0178511. doi: 10.1371/journal.pone.0178511
R Core Team (2023). R Development Core Team R: A Language and Environment for Statistical Computing 2023 (Vienna, Austria: R Core Team), 2023.
Rodolfo-Metalpa R., Houlbrèque F., Tambutté É., Boisson F., Baggini C., Patti P. P., et al. (2011). Coral and mollusc resistance to ocean acidification adversely affected by warming. Nat. Climate Change 1, 308–312. doi: 10.1038/nclimate1200
Rooney N., McCann K., Gellner G., Moore J. C. (2006). Structural asymmetry and the stability of diverse food webs. Nature 442, 265–269. doi: 10.1038/nature04887
Ruitton S., Francour P., Boudouresque C. F. (2000). Relationships between algae, benthic herbivorous invertebrates and fishes in rocky sublittoral communities of a temperate sea (Mediterranean). Estuar. Coast. Shelf 50, 217–230. doi: 10.1006/ecss.1999.0546
Sadovy de Mitcheson Y., Craig M. T., Bertoncini A. A., Cartpenter K. E., Cheung W. L., Choat J. H., et al. (2012). Fishing groupers towards extinction: a global assessment of threats and extinction risks in a billion dollar fishery. Fish Fisheries. 14, 119–136. doi: 10.1111/j.1467-2979.2011.00455.x
Sale P. F., Douglas W. A., Doherty P. J. (1984). Choice of microhabitats by coral reef fishes at settlement. Coral Reefs 3, 91–99. doi: 10.1007/BF00263759
Schrandt M. N., Lema S. C. (2011). Habitat-associated intraspecific variation in behavior and stress responses in a demersal coral reef fish. Mar. Ecol. Progr. Ser. 143, 153–166. doi: 10.3354/meps09411
Shannon C. E., Weaver W. (1949). The mathematical theory of communication (Urbana, Illinois: University of Illinois Press), 144 pp.
Shaver E. C., Silliman B. R. (2017). Time to cash in on positive interactions for coral restoration. PeerJ 5, e3499. doi: 10.7717/peerj.3499
Vacchi M., La Mesa G., Finoia M. G., Guidetti P., Bussotti S. (1999). Protection measures and juveniles of dusky grouper, Epinephelus marginatus (Lowe 1834) (Pisces, Serranidae), in the marine reserve of Ustica island (Italy, Mediterranean Sea). Mar. Life 9, 63–70.
Winter R., Desidera E. E., Guidetti P., Vermeersch S., Demirel N., Cakırlar C. (2022). Catch of the day: Abundance and size data of groupers (Epinephelidae) and combers (Serranidae) from Middle to Late Holocene Levantine archaeological contexts. J. Island Coast. Archaeology. doi: 10.1080/15564894.2022.2138643
Zibrowius H. (1980). Les Scléractiniaires de la Méditerranée et de l'Atlantique nord-oriental Vol. 11 (Monaco: Mémoires de l'Institut océanographique). 284 Pp.
Zunino S., Pitacco V., Mavrič B., Orlando-Bonaca M., Kružić P., Lipej L. (2018). The ecology of the Mediterranean stony coral Cladocora caespitosa (Linnaeus 1767) in the Gulf of 933 Trieste (northern Adriatic Sea): a 30-year long story. Mar. Biol. Res. 14, 307–320. doi: 10.1080/17451000.2017.1408915
Keywords: fish fauna, spatial heterogeneity, Mediterranean coral reef, Cladocora caespitosa, marine protected area, Veliko jezero
Citation: Lipej L, Ivajnšič D, Pitacco V, Mavrič B, Trkov D and Kružić P (2024) The coastal ichthyofauna of the Mediterranean coral reef: the case of Mljet National Park (Croatia, southern Adriatic Sea). Front. Mar. Sci. 11:1367382. doi: 10.3389/fmars.2024.1367382
Received: 08 January 2024; Accepted: 06 March 2024;
Published: 21 March 2024.
Edited by:
Francesco Tiralongo, University of Catania, ItalyReviewed by:
Ajit Kumar Mohanty, Indira Gandhi Centre for Atomic Research (IGCAR), IndiaDeniz Erguden, Iskenderun Technical University, Türkiye
Copyright © 2024 Lipej, Ivajnšič, Pitacco, Mavrič, Trkov and Kružić. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Danijel Ivajnšič, ZGFuaS5pdmFqbnNpY0B1bS5zaQ==