 Celeste Sánchez-Noguera
Celeste Sánchez-Noguera Ines D. Lange
Ines D. Lange Jorge Cortés
Jorge Cortés Carlos Jiménez
Carlos Jiménez Christian Wild
Christian Wild Tim Rixen
Tim Rixen- 1Universidad de Costa Rica, Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), San José, Costa Rica
- 2Leibniz Centre for Tropical Marine Research (ZMT), Bremen, Germany
- 3Faculty of Environment, Science and Economy, University of Exeter, Exeter, United Kingdom
- 4Enalia Physis Environmental Research Centre (ENALIA), Nicosia, Cyprus
- 5Marine Ecology Department, University of Bremen, Bremen, Germany
Coral reef development is influenced by a wide variety of factors, including temperature, salinity, nutrient concentrations, and carbonate chemistry. Studies focusing on physicochemical drivers of coral reef distribution and composition in the Eastern Tropical Pacific (ETP) are scarce, and carbonate chemistry and nutrient data for this region are limited. This study measured coral reef composition and physicochemical parameters along the Pacific coast of Costa Rica, over a one-year period at three locations: Santa Elena and Matapalo in the north, and Parque Nacional Marino Ballena in the south. Our results show high seasonal and spatial variability of physicochemical conditions with significant differences mainly explained by inorganic nutrient concentrations, with driving processes also having a strong influence on the variability of carbonate chemistry parameters. Coastal upwelling is the main driver of the seasonal variability in Santa Elena. Comparison of seasonal dissimilarity within locations confirms the presence of a geographical gradient, with stronger influence of the upwelling in Santa Elena relative to Matapalo, where several parameters displayed a lower seasonality and a carbonate system that supports reef development throughout the year. Conversely, in Marino Ballena the river discharges during rainy season exerted a strong control on the seasonal variability. The integrated analysis of coral reef composition and physicochemical parameters suggests that in addition to inorganic nutrients carbonate chemistry also plays a key role in coral distribution. Analyzing the spatial distribution of the main reef builders provides insights into the species-specific tolerance to varying conditions. Pavona clavus is widely distributed in both the northern and southern locations, suggesting that this massive coral is very tolerant to the high variability of physicochemical conditions. The dominant corals in the north (Pavona gigantea and Pocillopora spp.) are highly tolerant to nutrient-enriched cold waters with low aragonite saturation, while one of the main reef-builders in southern locations (Porites cf. lobata) cope better with low salinity, low aragonite saturation and low light intensity caused by river discharges. Understanding the preferences of individual coral species at our study locations can shed light on the environmental factors driving coral reef distribution in other locations of the ETP.
1 Introduction
Eastern Tropical Pacific (ETP) coral reefs extend from Mexico to northern Peru and are characterized by their small size and simple structure with discontinuous distribution. The relatively low diversity of corals in the ETP was increasingly attributed to the prevalence of extreme physical conditions along this coast (Dana, 1975; Cortés, 1997; Glynn et al., 2017a). Eastern boundary upwelling systems limit expansion of coral reefs in the north (California Upwelling system) and towards the south (Humboldt upwelling system). In between, major local wind-driven upwellings occur off Mexico in the Gulf of Tehuantepec, at the border between Nicaragua and Costa Rica in the Gulf of Papagayo (Papagayo upwelling) and in the Gulf of Panama (Panama upwelling). These wind-driven upwelling systems have a strong influence on the local physicochemical conditions in the ETP, particularly by lowering seawater temperature and increasing the availability of chlorophyll and inorganic nutrients (Fiedler and Talley, 2006; Lavín et al., 2006). Although less studied, it is also known that they drive changes in carbonate chemistry parameters. For example, during dry season the Panama upwelling produces a significant increase in total alkalinity (TA) and total dissolved inorganic carbon (DIC) of coastal waters in the Gulf of Panama, with the subsequent significant decrease in pH and aragonite saturation state (Ωa) (Manzello et al., 2008; Manzello, 2010b). Only one study carried out over a 29-hour period in 2009 has measured carbonate chemistry parameters within the Gulf of Papagayo, reporting a sharp decrease in DIC, pH and Ωa during the development of an upwelling event (Rixen et al., 2012). Additionally, the climate anomaly El Niño Southern-Oscillation (ENSO) strongly influences the upwelling conditions in the ETP (Fiedler and Lavín, 2017; Glynn et al., 2017b). During negative ENSO episodes known as El Niño, upwelling weakens, causing seawater temperatures to increase above the average. In the past, ENSO-induced warming events have caused bleaching and significant coral mortality in the Gulf of Papagayo (Jiménez et al., 2001). Conversely, positive ENSO episodes referred to as La Niña intensify local upwelling conditions.
Along the Pacific coast of Costa Rica, coral reef formations are restricted to northern and southern locations (Cortés and Jiménez, 2003; Cortés et al., 2010; Cortés, 2016), while in the middle sections of the littoral scleractinian corals appear as single colonies or in sparse coral communities. Near the coastline, branching corals of the genus Pocillopora built important reef structures in northern locations within the Gulf of Santa Elena and Gulf of Papagayo (Jiménez, 1997, 2001a; Jiménez et al., 2010; Méndez-Venegas et al., 2021), and smaller patch reefs in southern locations near Corcovado and Golfo Dulce (Guzmán and Cortés, 1989; Cortés, 1990; Cortés and Jiménez, 1996). The presence of massive species varies from north to south, with Pavona gigantea, Pavona clavus and Gardineroseris planulata as the main reef builders in the north, while Porites cf. lobata is the main reef-building coral in the south (Cortés, 1990; Jiménez, 2001a; Alvarado et al., 2005).
This observed longitudinal coral distribution (summarized by Cortés and Jiménez, 2003; Glynn et al., 2017a) raises the question of which environmental driver(s) regulate the reef development along this gradient. To address this question, we measured total alkalinity (TA), dissolved inorganic carbon (DIC), temperature, salinity and inorganic nutrient concentrations at three locations along the coast during the dry and rainy season. The selection of these physicochemical parameters responds to their natural variability along this coast due to different processes. For instance, seawater temperature, nutrients and carbonate chemistry are influenced by the Papagayo upwelling, which strongly affects northern locations (Alfaro et al., 2012; Fernández-García et al., 2012; Rixen et al., 2012). In contrast, seasonal changes in river runoff, which impact the southernmost locations, are likely to have a greater effect on nutrient concentrations and salinity (Alvarado and Aguilar, 2009; Alvarado et al., 2009), and likely in the carbonate chemistry during rainy season. In addition to the five physicochemical parameters measured during the study, percentages of benthic community composition were quantified in coral reefs at each location. The main goal of this study was to identify the key physicochemical factors controlling coral reef distribution and species composition at the local scale. Moreover, the specific objective of characterizing seawater carbonate chemistry will enhance baseline information within the ETP. The compiled data allowed the spatial and seasonal comparison of physicochemical conditions and coral reef status, to elucidate which factors can be controlling the differences of the benthic community structure along the Costa Rican Pacific coast.
2 Materials and methods
2.1 Study locations
To answer our research question about the environmental drivers controlling coral reef development and distribution along the Costa Rican Pacific coast, three study locations were chosen based on the presence of coral reefs, which are restricted to northern and southern locations (Cortés and Jiménez, 2003; Cortés et al., 2010; Cortés, 2016): Santa Elena and Matapalo in the north and Parque Nacional Marino Ballena in the south (also referred to as Marino Ballena in this paper) (Figure 1). The number of sampling sites in each location was determined by the presence of coral reef formations. A total of four reefs were selected in Santa Elena (Bajo Rojo: 10.95769, -85.73400; Matapalito: 10.93633, -85.79385; Pochote: 10.93151, -85.80002; Cabros: 10.942267, -85.81363), one in Matapalo (10.53922, -85.76553) and three in Marino Ballena (Tómbolo: 9.14481, -83.75819; Bajo Mauren: 9.11258, -83.74092; Tres Hermanas: 9.10408, -83.70686) (Table 1).
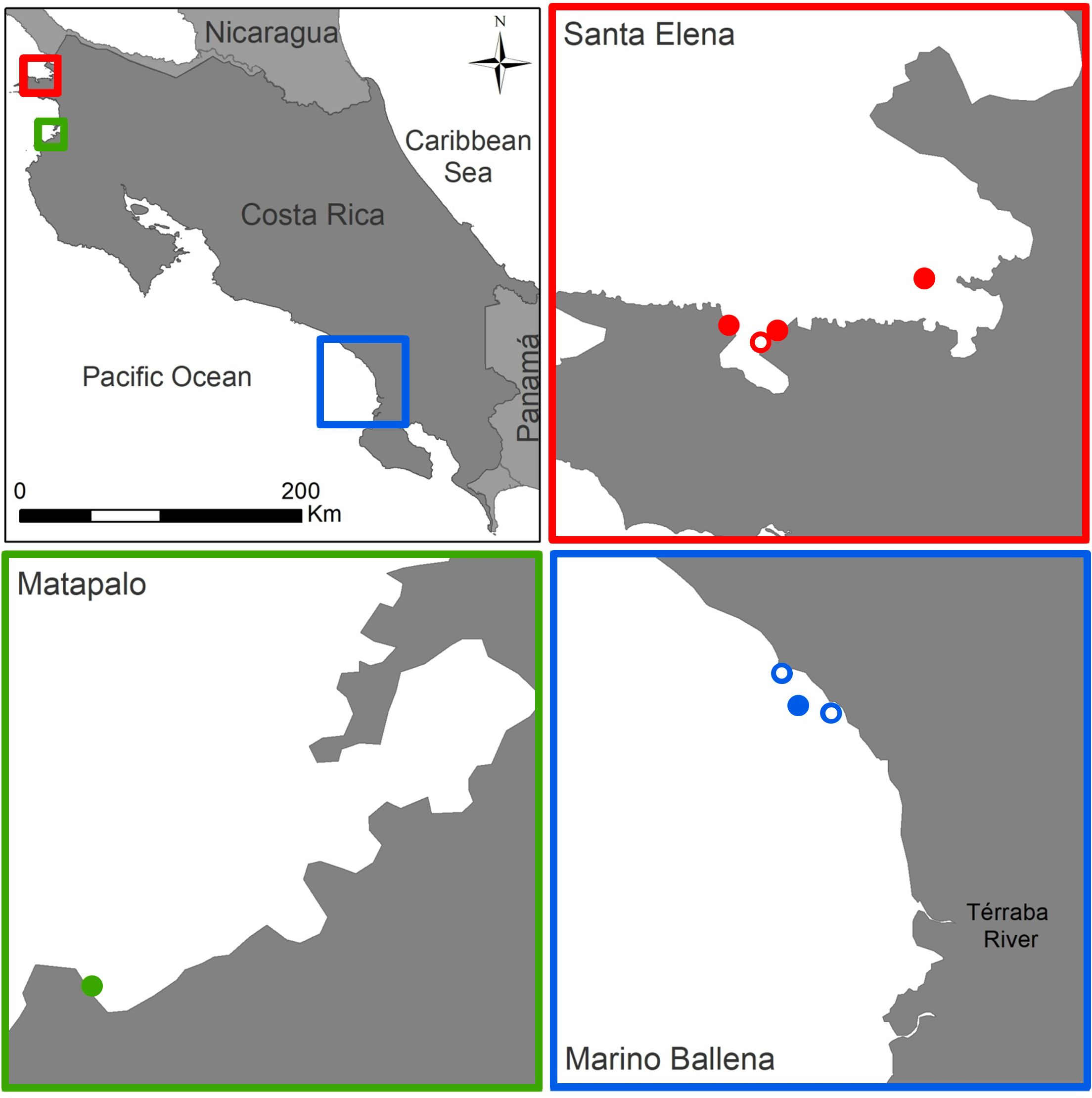
Figure 1. Study areas. Studied locations along the north (Santa Elena, Matapalo) and south (Marino Ballena) Pacific coast of Costa Rica. Circles indicate the sampling reefs in each location (close = water sampling + benthic survey, open = only water sampling).
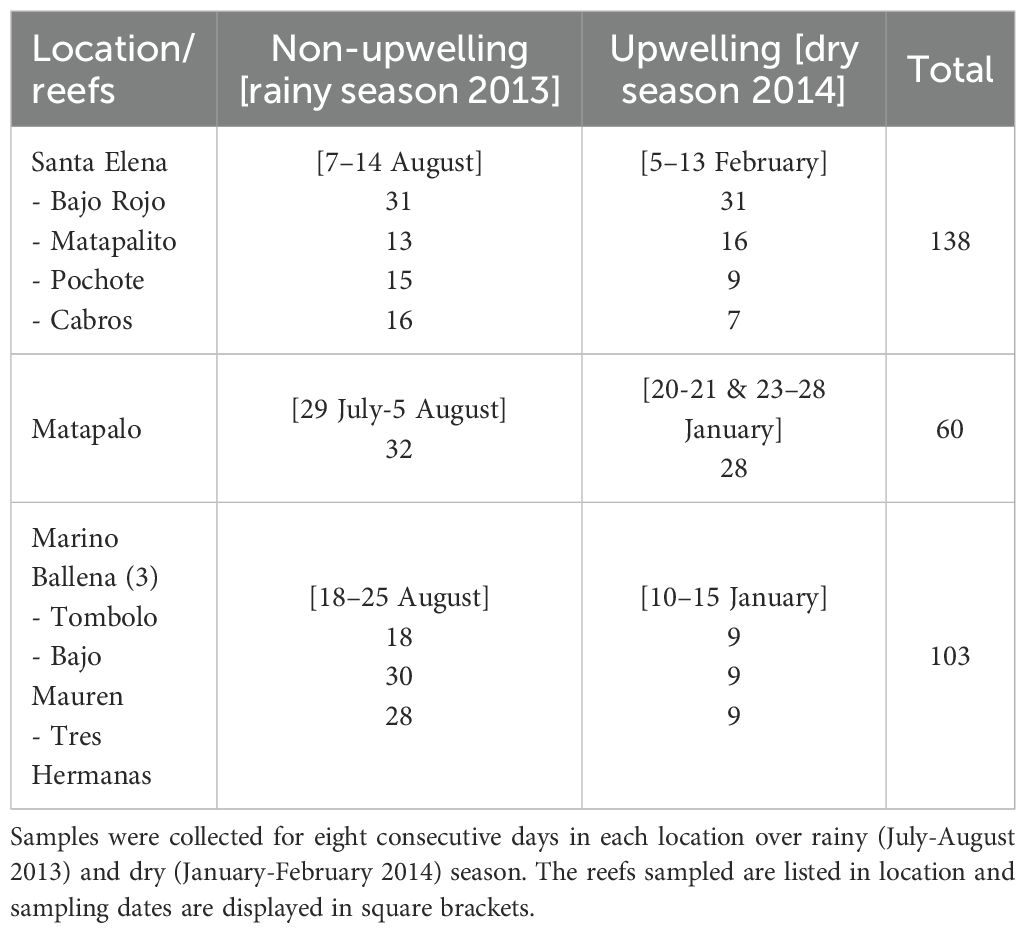
Table 1. Samples were collected for eight consecutive days in each location over rainy (July-August 2013) and dry (January-February 2014) season. The reefs sampled are listed in each location and sampling dates are displayed in square brackets.
Most coral reefs in northern locations are directly exposed to the seasonal upwelling of Papagayo and are built by branching species (Pocillopora spp.), although there are also reefs that consist of massive corals (Pavona spp.) (Cortés, 1997; Jiménez, 1997, 2001a; Cortés et al., 2010). Two of the study locations selected for this project are located within the Gulf of Papagayo, as both display important reef formations built by different coral species. In southern locations, where the seasonal upwelling is absent, Marino Ballena features several coral communities and coral reefs built by Porites cf. lobata and P. clavus (Alvarado et al., 2005, 2006; Cortés et al., 2010). Golfo Dulce, which is also located in the south and has coral reefs primarily built by Porites cf. lobata (Cortés, 1990), was intentionally excluded from this study. The reason relies on the fact that Golfo Dulce is a tectonic basin with an anoxic layer (Vargas-Zamora et al., 2021), therefore this unique oceanographic condition would have made challenging its comparison with the other locations.
The climate on the Pacific coast of Costa Rica features two well defined seasons, although their timing shows slight variations by location. In the north Pacific, the dry season runs from December to March while the rainy season occurs from May to November (Cambronero-Solano et al., 2021). In contrast, the dry season in the south Pacific is somewhat shorter, lasting from January to March, with the rainy season extending from May to December (Instituto Metereológico Nacional (IMN), 2008). In both regions April serves as a transition period between seasons. In the Gulf of Papagayo, major changes in seawater temperature, nutrient concentration and other physicochemical parameters occur mostly in pulses during the development of the upwelling events over the dry season (Alfaro et al., 2012; Fernández-García et al., 2012; Stuhldreier et al., 2015; Cambronero-Solano et al., 2021). Santa Elena is located in the northernmost section of this gulf and is thought to experience a strongest influence from the seasonal upwelling compared to Matapalo, which is located in the southernmost section of Papagayo. A previous study compared the seasonal changes of physicochemical parameters between these locations, reporting that the upwelling intensity is greater in Santa Elena than in Matapalo, characterized by a decrease in seawater temperature and increase in inorganic nutrient concentrations during the dry season (Stuhldreier et al., 2015). Conversely, during the dry season in Marino Ballena, seawater temperature and salinity increase, while the rainy season leads to higher sedimentation and nutrient levels, along with decreased seawater temperature and salinity (Alvarado and Aguilar, 2009; Alvarado et al., 2009).
Fieldwork for this research was conducted under the research permits No. 028-2013-SINAC and No. SINAC-SE-GASP-PI-R-096-2013, issued by the Sistema Nacional de Áreas de Conservación (SINAC), Ministry of the Environment, Costa Rica.
2.2 Water sampling
The seawater sampling was designed to capture the variability of physicochemical parameters in each location, not within reefs. At each location, water samples were collected during rainy (July-August 2013) and dry (January-February 2014) season (Table 1). Being aware that the Papagayo upwelling is wind-driven and can fluctuate significantly on weekly or daily timescales, the season sampling at each location was conducted during several days in a row. The sampling period per season ranged between 6–8 days in each location, depending on logistics, with the number of sampling days as replicates per season. When possible, samples in each reef were collected at two different tides and depths per day. Sampling depth ranged from 0.5 m below the surface to 11 m, based on the reef depth, tide and sea conditions. Sampling depth was not consistent across reefs, because water column depth varied with the tide range and the sampling hour. Samples were collected at different times of the day, from 05:30 to 16:30, depending on the tide cycles. The sampling was carried out as close as possible to the tide’s peak, but there was a variability in the time of sampling due to the distance between reefs (time required to move from one reef to the next one) and other logistics. In some specific dates, not all reefs were sampled in Santa Elena or Marino Ballena, due to restrictive working conditions such as rough sea or fishing gear deployed near the reef. Additionally, in particular days, samples were collected only at one depth (near the surface or bottom indistinctly) or one tide (low or high indistinctly), due to limited amount of sampling bottles, the availability of boats for sampling (in Matapalo and Marino Ballena most sampling was done taking advantage of touristic trips), the navigation conditions or the distance between reefs.
Sampling was carried out with a 10 L volume Niskin bottle following best practices guidelines for ocean CO2 measurements (Dickson et al., 2007). Discrete water samples for determination of carbonate chemistry parameters were collected in 250 mL air-tight borosilicate bottles, fixed immediately after collection with 200 μL of a 50% saturated HgCl2 solution (35 g L-1 HgCl2) and stored cold until analysis. Nutrient samples were filtered in the field through a 0.45 𝜇m pore size filter and frozen until analysis. Samples for salinity determination were collected in 50 mL plastic bottles and measured in the laboratory with a WTW probe (Cond3310). Seawater temperature (SWT) was measured in situ with HOBO®Pendant Temperature data loggers at Bajo Rojo and Matapalo (10–15 min intervals in dry season between December 2013 and January 2014 and every 30 min during rainy season). The SWT value used was the one recorded the closest to the seawater sampling time. In all the other reefs (Cabros, Matapalito, Pochote, Tómbolo, Tres Hermanas and Bajo Mauren) SWT was also measured in situ, with a WTW probe (Cond3310).
2.3 Reef surveys
To characterize the composition of the benthic community and the main coral reef builders at each location, percentages of benthic coverage were quantified across several reefs. Benthic surveys were carried out at three reefs in Santa Elena (Bajo Rojo, Matapalito, Cabros), one in Matapalo (Matapalo) and one in Marino Ballena (Bajo Mauren) (Figure 1). Due to fieldwork logistics and the fact that we did not intend to study the temporal changes in benthic community composition, reef surveys were conducted at one time point during dry season (Table 2). Data from different reefs were averaged by location. Benthic surveys were conducted following the chain method (Rogers et al., 1994) with five replicates per transect. A 10 m length chain with a known number of links (n = 532) was laid on top of the reef following its contours; nine categories (live coral at species level, dead coral, bleached coral, macroalgae, coralline algae, turf algae, substrate, cyanobacteria and others) were used to quantify the benthic composition under each link. Afterwards, the corresponding number of links for each category was converted to relative benthic cover.
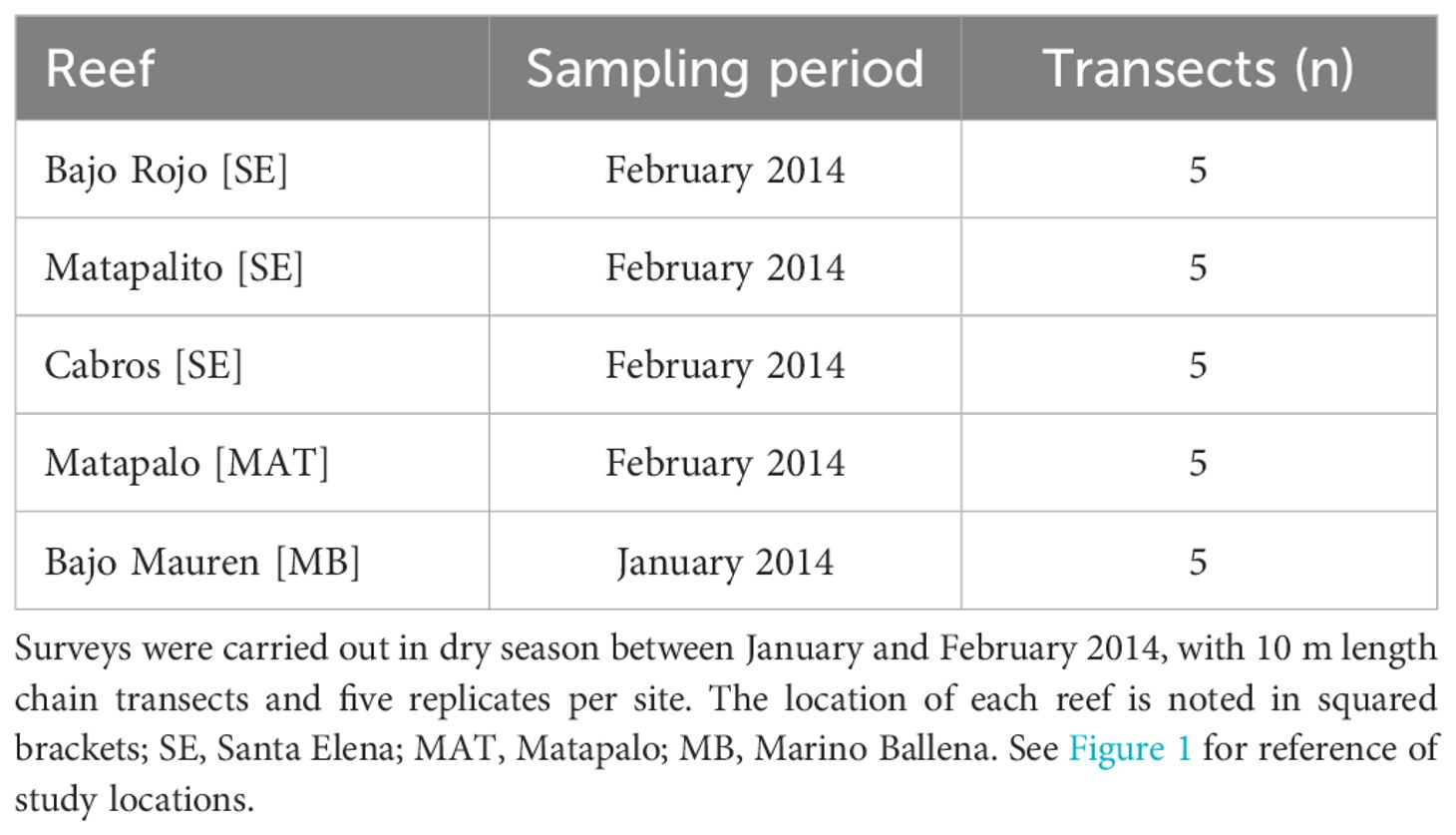
Table 2. Surveys were carried out in dry season between January and February 2014, with 10 m length chain transects and five replicates per site. The location of each reef is noted in squared brackets; SE = Santa Elena; MAT = Matapalo; MB = Marino Ballena. See Figure 1 for reference of study locations.
2.4 Laboratory measurements
Total alkalinity (TA) and dissolved inorganic carbon (DIC) were measured with a VINDTA 3C system (Versatile instrument for the determination of total dissolved inorganic carbon and titration alkalinity), coupled with a UIC CO2 coulometer (model CM5015) and a METROHM©Titrino (model 716 DMS). Instruments were calibrated with Dickson Certified Reference Material (Batch 127) (Dickson et al., 2003). The pH, aragonite saturation state (Ωa) and fCO2 were calculated with CO2SYS as a function of measured parameters (TA, DIC, salinity, nutrients and SWT), with dissociation constants of Mehrbach et al. (1973) for carbonic acid as refit by Dickson and Millero (1987), and Dickson (1990) for boric acid.
Five inorganic nutrients (phosphate, ; silicate, ammonium, ; nitrite, and nitrate, ) were measured through a reagent-based method with a Lachat′sQuikChem© 8500 Series 2 Flow Injection Analysis (FIA) System, following the Standard Operation Procedure from Centro de Investigación en Ciencias del Mar y Limnología (CIMAR) of the University of Costa Rica (PON Nut-08).
2.5 Meteorological data
Meteorological data (rainfall and wind speed) were provided by the Instituto Meteorológico Nacional, from two stations located in the North Pacific (Santa Rosa and Liberia) and two in the South Pacific (Barú and Damas). Wind speed (m/s) and rainfall (mm) data used in this study corresponded to the same dates as the seawater sampling. Wind speed data corresponds to hourly averages and rainfall to daily values measured at each station. The values of wind speed used in this study corresponded to hourly averages closest to the seawater sampling time, and for rainfall values we used the daily value.
2.6 Data analysis
For a general comparison of the physicochemical and meteorological conditions during this study, all values were averaged by location and season (Supplementary Table S1). General statistical differences of physicochemical and meteorological parameters between locations (annual scale) and season were tested by Kruskal-Wallis. For statistical comparisons of carbonate chemistry, we used the salinity-normalized values (nTA, nDIC) (Equation 1):
where S is the measured salinity value of the sample.
Spatial and temporal variations in physicochemical parameters, as well as benthic community composition, were analyzed using the software programs PRIMER 7, R and PAST. The meteorological data was excluded from these statistical analyses and we expand on this at the end of this section. Moreover, to characterize the variability of the physicochemical conditions of the water column at each location, samples collected at different depths during the same tide were averaged together for each reef, as the sampling depth varied across the reefs. This averaging strategy avoid losing the seasonal replication of measured (SWT, salinity, phosphate, nitrate, nitrite, silicate, ammonium, TA, DIC) and calculated (pH, fCO2, Ωa) physicochemical parameters. Prior to analysis, the physicochemical parameters were log-transformed and standardized, and resemblance matrices were built using Euclidean distances. Differences in physicochemical parameters among locations and seasons were tested with a nested Permutational Multivariate Analysis of Variance (PERMANOVA), including season, location and tide as fixed factors, and reef as a random factor nested within location. Additionally, when the main test indicated significant differences, PERMANOVA pairwise comparisons were conducted. A Similarity Percentage Analysis (SIMPER) was also performed to identify which parameters contributed the most to the observed differences between locations and seasons. To visualize patterns in physicochemical parameters, box-plots and Principal Component Analysis (PCA) were used.
To estimate the general contribution of the main processes driving the carbonate chemistry in our study locations on an annual basis, we did a simple linear regression analysis for each location and used an approach proposed by other authors (Albright et al., 2013); considering that net organic carbon production (photosynthesis-respiration) and net inorganic carbon production (calcification-dissolution) affect TA and DIC differently (Suzuki and Kawahata, 2004). According to the theoretical stoichiometric relationship between these processes and TA and DIC, for every mole of calcium carbonate (CaCO3) produced, DIC decreases by two moles and TA decreases by one mole. In contrast, for every mole of organic carbon produced via photosynthesis, one mole of DIC is consumed while TA remains unchanged (Albright et al., 2013). Therefore, the slope of the simple linear regression (nDIC vs. nTA) indicates the general balance between the net inorganic and organic carbon production at each location. The ratio between these processes (inorganic carbon production:organic carbon production) is given by Equation 2, where m is the slope of the nDIC-nTA relationship (Albright et al., 2013):
In Equation 2, a ratio < 1 points out to a system mainly controlled by organic processes. Conversely, a ratio > 1 indicates that inorganic processes are the main drivers of the system. Further data is needed to identify which specific processes (photosynthesis, respiration, calcification or dissolution) control the overall carbonate system, but this falls outside the objectives of this study.
For analysis of the benthic community composition, differences in percentages of benthic coverage between locations were tested with a nested PERMANOVA, including location as fixed factor and reef as a random factor nested within location. To reduce the influence of dominant taxa, benthic coverage data were fourth-root transformed prior to calculating a Bray-Curtis resemblance matrix. Average coverage values for each benthic category were calculated per location, followed by estimations of the Shannon-Wiener diversity index (H’) and Pielou’s evenness (J’) to evaluate community diversity and distribution.
Finally, it is important to note that meteorological data were obtained from the nearest inland stations to our study areas, which do not measure the exact conditions at our sampling locations. Santa Rosa meteorological station (10.84111, -85.61944) for example, is about 20 km southeast from Santa Elena; Liberia station (10.83889, -85.55222) is about 25 km west from Matapalo; while Barú station (9.27139, -83.88139, rainfall) and Damas station (09.49528, -84.21472, wind speed) are located about 18 km and 70 km northwest from Marino Ballena, respectively. For this reason, we have used wind speed and rainfall data to explain general patterns and tendencies during sampling periods, but not for direct statistical correlations with the physicochemical parameters. Statistical differences of meteorological conditions between locations and season were tested by Kruskal-Wallis.
3 Results
3.1 Environmental parameters
Results of the nested PERMANOVA indicate that physicochemical conditions are significantly different between locations (Pseudo-F = 45.77, P(perm) = 0.003, perms = 8769) and between seasons (Pseudo-F = 38.63, P(perm) < 0.001, perms = 9941). Conversely, tide (Pseudo-F = 0.25, P(perm) = 0.779, perms = 9949) and reef (Pseudo-F = 1.28, P(perm) = 0.201, perms = 9899) did not have a significant effect in these parameters, suggesting that the tide cycle is not relevant for the changes in physicochemical conditions and that the variability within each location is not as important as the variability between locations and seasons. A comparison of the variability of physicochemical conditions on the annual scale indicates that within northern locations, Santa Elena reefs are exposed to a high variation with extreme values either end (Figure 2, Supplementary Table S1); while the variability in Matapalo is lower, particularly for salinity, temperature and carbonate chemistry parameters.
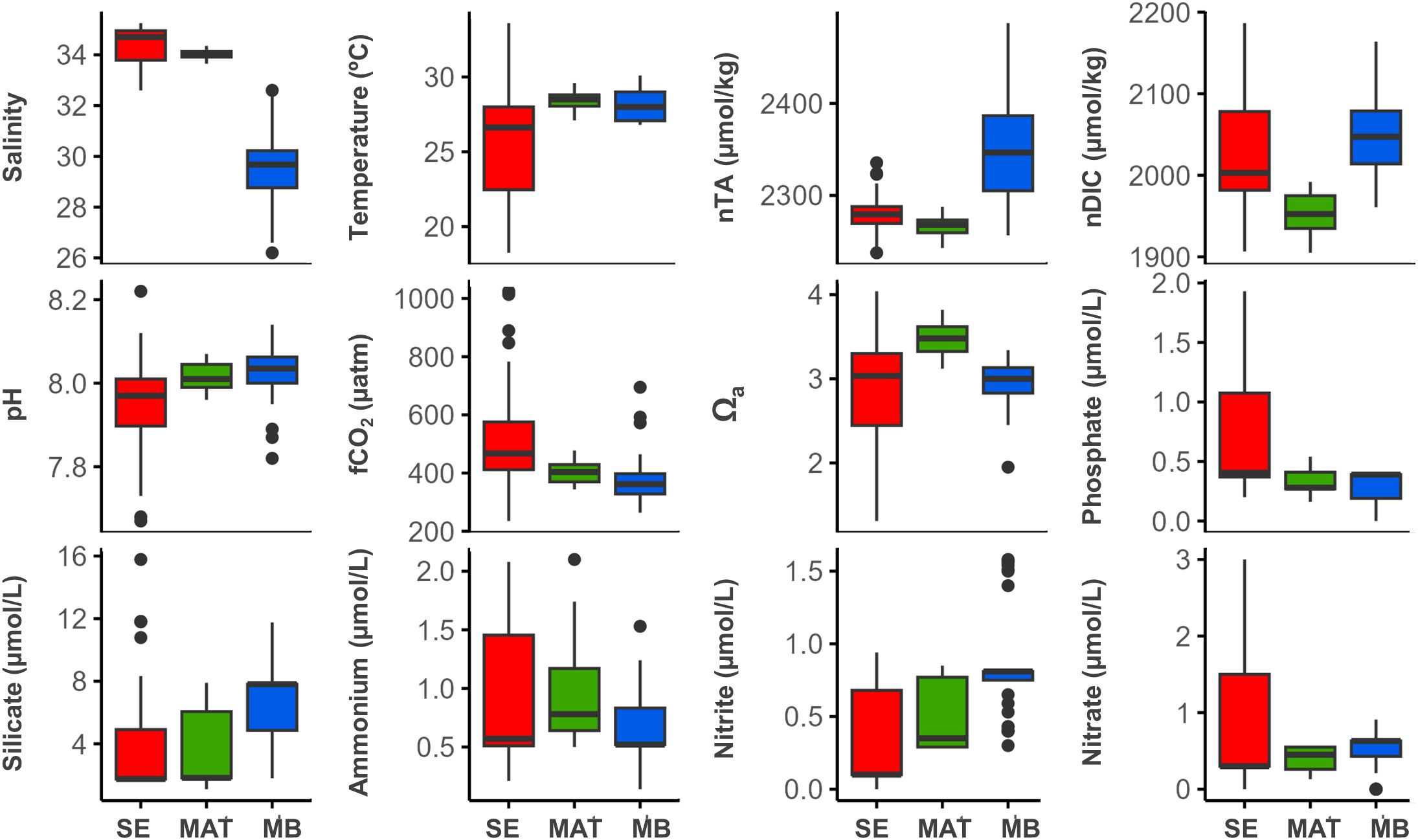
Figure 2. Variability of physicochemical conditions along the Pacific coast of Costa Rica. Annual mean values of physicochemical parameters from the study locations indicate a high spatial variability in seawater conditions, related to the distance from the upwelling of Papagayo. SE = Santa Elena; MAT = Matapalo; MB = Marino Ballena. See Figure 1 for the study locations.
Pairwise tests suggest that the strongest differences in physicochemical conditions occur between the northernmost and the southernmost location (Pseudo-t = 9.53, P(perm) = 0.013, perms = 4322). Santa Elena displays the greatest internal variability, whereas Marino Ballena exhibits the most homogeneous conditions. Seasonal comparisons indicate significant differences between the rainy and the dry season (Pseudo-t = 6.22, p < 0.001, perms = 9940), with physicochemical parameters showing more heterogenous conditions during the dry season.
Average (± standard deviation) of measured parameters by location, independent of seasons, showed that in the northernmost location (Santa Elena) the coastal waters are cooler (25.3 ± 3.3 °C, H = 45.79, p < 0.001) and more saline (34.3 ± 0.8 psu, H = 131.03, p < 0.001), whilst at the southernmost location (Marino Ballena) coastal waters have a lower salinity (29.4 ± 1.7 psu, H = 131.03, p < 0.001) (Supplementary Tables S1, S2). Concentrations of (H = 27.18, p < 0.001) and (H = 10.08, p = 0.006) were highest in Santa Elena, whereas Marino Ballena held the highest average concentrations of (H = 66.06, p < 0.001) and (H = 75.87, p < 0.001) (Supplementary Tables S1, S2). The average nTA values in Santa Elena and Matapalo (< 2300 μmol kg-1) were lower than in Marino Ballena (> 2300 μmol kg-1) (H = 109.31, p < 0.001). On the other hand, in Santa Elena and Marino Ballena annual average nDIC showed concentrations of > 2000 μmol kg-1 whereas nDIC concentrations were lower in Matapalo (< 2000 μmol kg-1) (H = 57.95, p < 0.001) (Supplementary Tables S1, S2).
According to the PCA (Figure 3), five main water parameters explained 82.6% of the variance of the physicochemical conditions. Nitrate (0.663), phosphate (0.583) and ammonium (0.303) contributed 57.2% to the variance (PC1); whilst nitrite (-0.829), phosphate (0.381) and silicate (-0.375) explained a further 25.4% (PC2). These findings were supported by SIMPER analysis, which identified the same five inorganic nutrients as the primary contributors to the observed differences between locations and seasons. According to the comparison between locations, the highest dissimilarity was found between the northernmost (Santa Elena) and the southernmost (Marino Ballena) location (average squared distance = 0.98), primarily driven by phosphate (28.23%), nitrite (27.94%) and nitrate (24.64%), with Santa Elena showing elevated phosphate and nitrate concentrations, whereas Marino Ballena exhibited higher nitrite levels. Meanwhile, there was a moderate dissimilarity (average squared distance = 0.83) between northern locations (Santa Elena and Matapalo), with nitrate (33.02%), phosphate (30.59%) and nitrite (16.17%) contributing nearly to 80% of the observed differences. Nutrient concentrations were generally higher at Santa Elena, except for nitrite, which was slightly greater at Matapalo. In contrast, Matapalo and Marino Ballena showed the lowest dissimilarity (average squared distance = 0.37), mainly due to differences in nitrite (38.32%), silicate (20.14) and ammonium (17.22%), all of which had higher concentrations at Marino Ballena. The SIMPER analysis between seasons indicated that three inorganic nutrients - nitrate (28.19%), phosphate (26.74%) and nitrite (21.56%) – are the primary contributors to the observed seasonal differences. Together, they accounted for over 76% of the dissimilarity between seasons, with notably higher concentrations during the dry season.
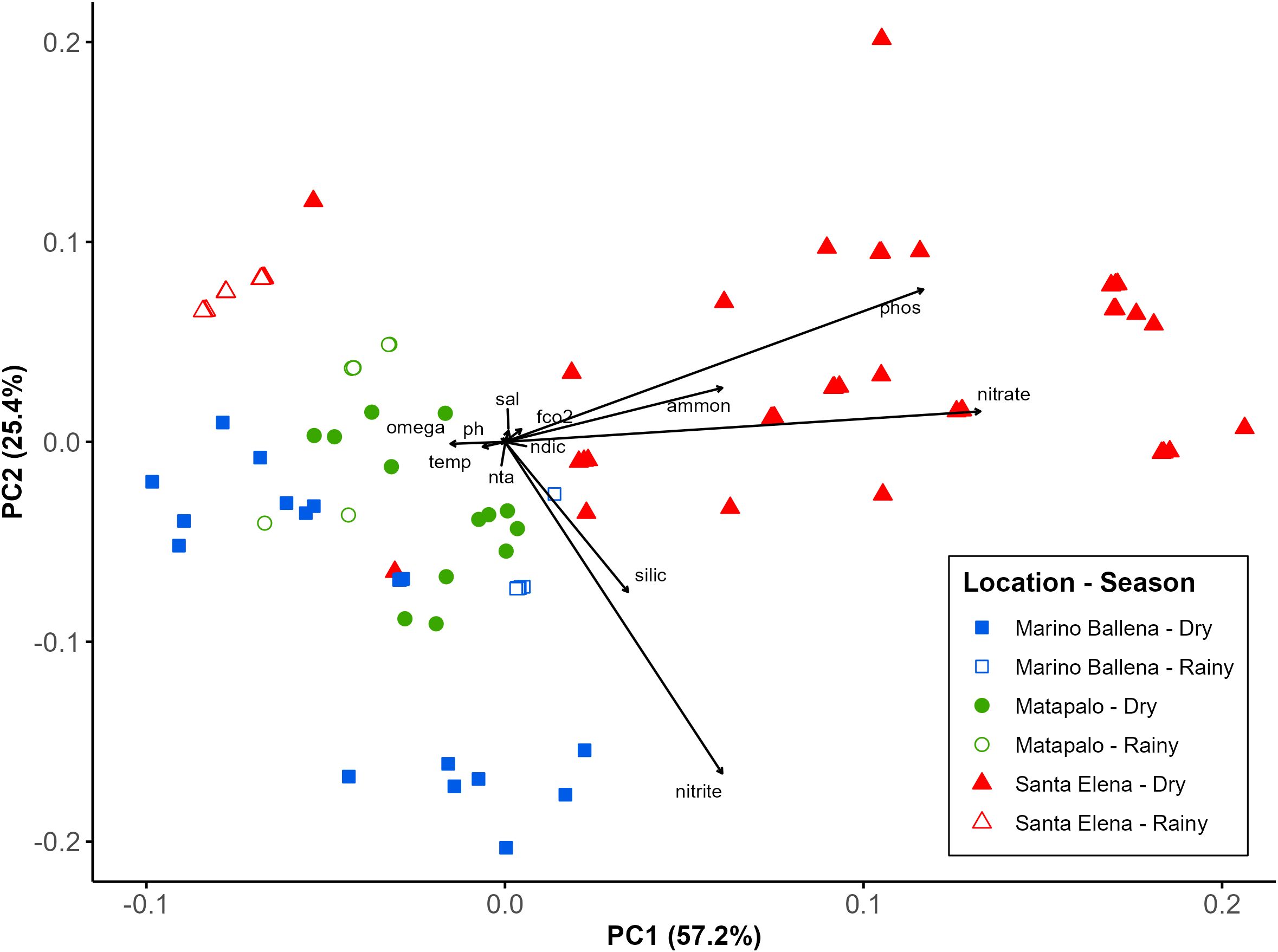
Figure 3. Spatial and temporal variation of physicochemical parameters. Seasonal sampling data from rainy season 2013 (July-August) and dry season 2014 (January-February) were analyzed by location and season based on Principal Component Analysis (PCA). The distance between data points indicates their similarity in seawater conditions. PC1 correlates positively to nitrate (0.663), phosphate (0.583) and ammonium (0.303). PC2 correlates positively to phosphate (0.381) and negatively nitrite (-0.829) and silicate (-0.375).
The PERMANOVA results also revealed a significant interaction between location and season (Pseudo-F = 54.86, P(perm) < 0.001, perms = 9960), indicating that the effect of the season on the physicochemical conditions is not equal in all locations. Pairwise test confirms this result, with the higher variability taking place during dry season in Santa Elena (Pseudo-t = 11.77, P(perm) = 0.026, perms = 7446) and Matapalo (Pseudo-t = 5.91, p < 0.001, perms = 9961). In Marino Ballena, the Pseudo-t value (8.95) also indicates a strong separation in physicochemical conditions between seasons. This difference is statistically significant based on the Monte Carlo p-value (P(MC) < 0.001, perms = 360), although permutation testing (P(perm) = 0.084, perms = 360) offers contradictory results, likely due to the limited permutation count which might affect robustness. SIMPER analysis revealed that seasonal dissimilarity within each location followed a decreasing gradient from north to south, with values of 1.69 for Santa Elena, 0.40 for Matapalo, and 0.31 for Marino Ballena.
During the dry season Santa Elena experienced an increase in salinity (H = 44.38, p < 0.001), nTA (H = 17.83, p < 0.001) and nDIC (H = 48.21, p < 0.001) as well as a sharp drop in SWT (H = 65.34, p < 0.001). At Marino Ballena the increase of nTA (H = 35.781, p < 0.001) and nDIC (H = 33.12, p < 0.001) during the rainy season was accompanied by a slight decrease of salinity (H = 17.31, p < 0.001) and SWT (H = 48.26, p < 0.001). Conversely, salinity (H = 0.17, p = 0.677), nTA (H = 0.76, p = 0.382) and nDIC (H = 0.68, p = 0.409) remained similar during both seasons in Matapalo (Supplementary Tables S1, S3).
The simple linear regression analysis of salinity-normalized values (nDIC vs. nTA) (Figure 4) indicates that Marino Ballena had higher nTA than the other locations, with a steep increase in nTA and nDIC during the rainy season. Santa Elena was the location with the highest nDIC values occurring specifically during the dry season. Measured parameters in Matapalo were the lowest and had similar values during both seasons, overlapping with values measured in Santa Elena during the rainy season. The estimated ratios between inorganic and organic carbon production (Equation 2) were lower in the northern locations, with values of 0.04 in Santa Elena and 0.18 in Matapalo. The estimated inorganic:organic production ratio in Marino Ballena was 0.88.
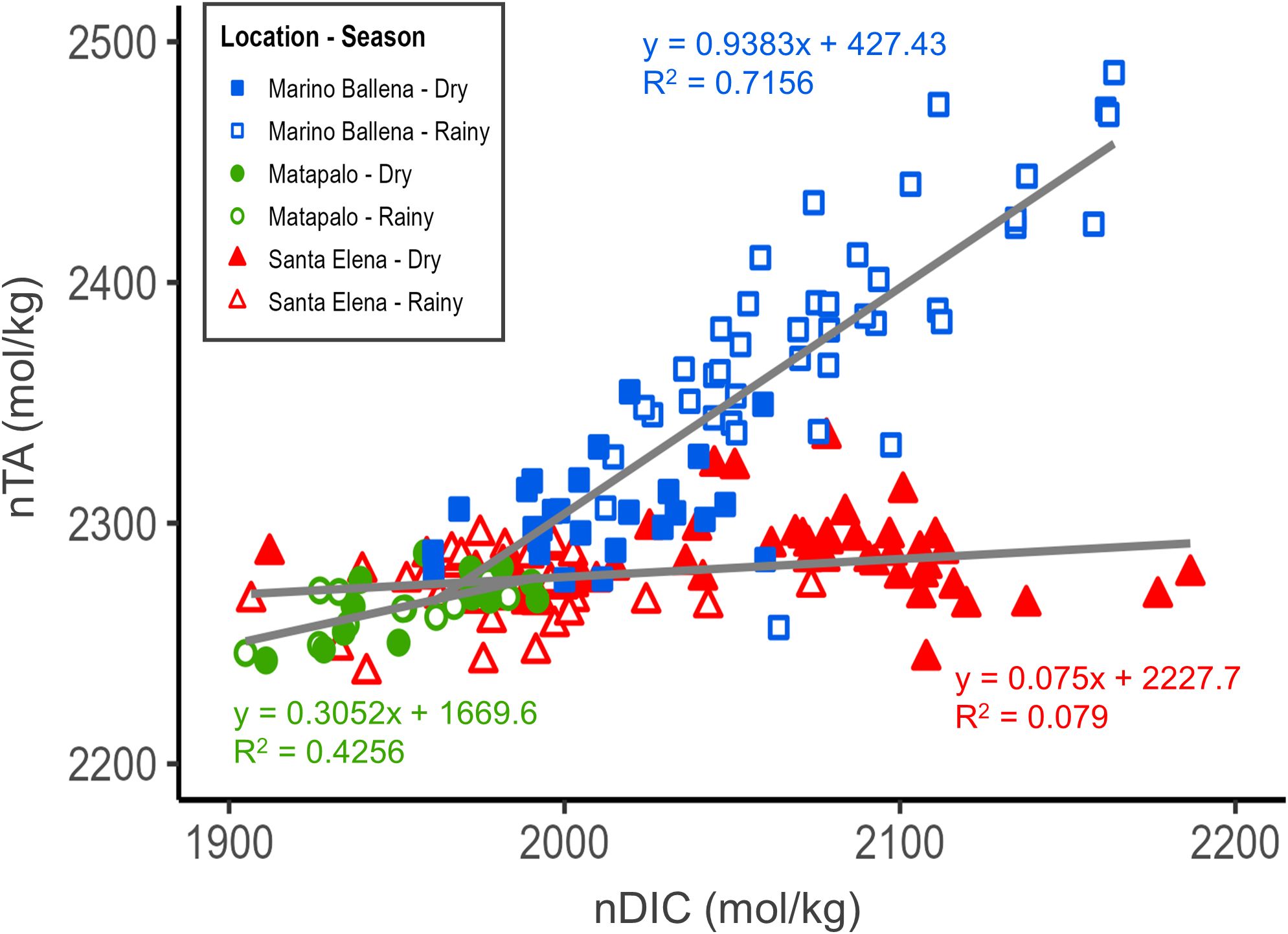
Figure 4. Seasonal variation of carbonate chemistry in three locations at the Pacific coast of Costa Rica. Total alkalinity (TA) and dissolved inorganic carbon (DIC) were normalized to a salinity of 35 (Equation 1). The slope of the nDIC-nTA relationship indicates the balance between the inorganic (calcification-dissolution) and organic processes (photosynthesis-respiration). The ratio between inorganic carbon production:organic carbon production is given by Equation 2.
Meteorological parameters measured near Marino Ballena during the sampling period indicate that this was the location with the lowest wind speeds on the annual scale (1.3 ± 0.7 m s-1, H = 56.33, p < 0.001) and highest rainfall during the rainy season (24.9 ± 12.9 mm day-1, H = 49.28, p < 0.001) (Supplementary Tables S1-S3). The data obtained from the Liberia meteorological station show that Matapalo was the other extreme, with highest wind speeds during the dry season (7.1 ± 3.2 m s-1, H = 6.74, p = 0.009) and lowest rainfall during the rainy season (1.1 ± 1.9 mm day-1, H = 6.77, p = 0.009) (Supplementary Tables S1-S3). Near to Santa Elena, the annual average values of wind speeds and rain were 2.3 ± 1.3 m s-1 and 1.8 ± 3.6 mm day-1, respectively (Supplementary Tables S1, S2).
3.2 Benthic community composition
In all locations, the category “Bleached” accounted for less than 0.2% of benthic coverage, and was excluded from the graphical representation of the percentages of benthic cover (Figure 5). Results from the nested PERMANOVA revealed no significant differences in benthic coverage between locations (Pseudo-F = 0.313, P(perm) = 1.000, perms = 10). In contrast, comparisons within each location indicated significant variation in benthic composition among reefs (Pseudo-F = 19.654, P(perm) = 0.000, perms = 9953). However, findings from within-location comparisons will not be discussed further, as Santa Elena was the only location where more than one reef was surveyed. Regarding benthic composition categories, richness (S) was consistent across all locations. Diversity (H’) showed similar values in northern locations, with slightly higher levels in Marino Ballena. Evenness (J’) was comparable between Matapalo and Marino Ballena, but somewhat lower in Santa Elena (Table 3). Reefs in Santa Elena were mainly composed of live coral (43.37 ± 38.11%), turf (27.67 ± 27.53%), coralline algae (17.24 ± 14.41%) and substrate (11.37 ± 14.96%). Matapalo reef had three dominant categories, live coral (41.39 ± 15.67%), coralline algae (31.50 ± 5.43%) and turf (20.94 ± 6.10%). In the reef evaluated in Marino Ballena the three dominant categories of the benthic composition were coralline algae (44.02 ± 20.19%), live coral (25.71 ± 10.44%) and substrate (13.80 ± 29.29%). Macroalgae were only observed in Santa Elena (0.08 ± 0.21%) and Matapalo (4.92 ± 9.99%). Cyanobacteria were present in all locations but had a very low coverage; with the highest quantified value in Marino Ballena (3.20 ± 2.08%) (Figure 5A). Regarding scleractinian corals, all locations showed low diversity indexes (H’) with values < 1.0, suggesting that these coral reefs are built by few species (Table 3). The reefs in the north were mainly built by P. gigantea and Pocillopora spp., whereas southern reefs are built by P. clavus and P. cf. lobata with minor contribution of Porites panamensis and Psammocora stellata (Figure 5B). Matapalo was the location with lower coral diversity (Table 3), with Pocillopora sp. as the dominant reef builder (Figure 5B).
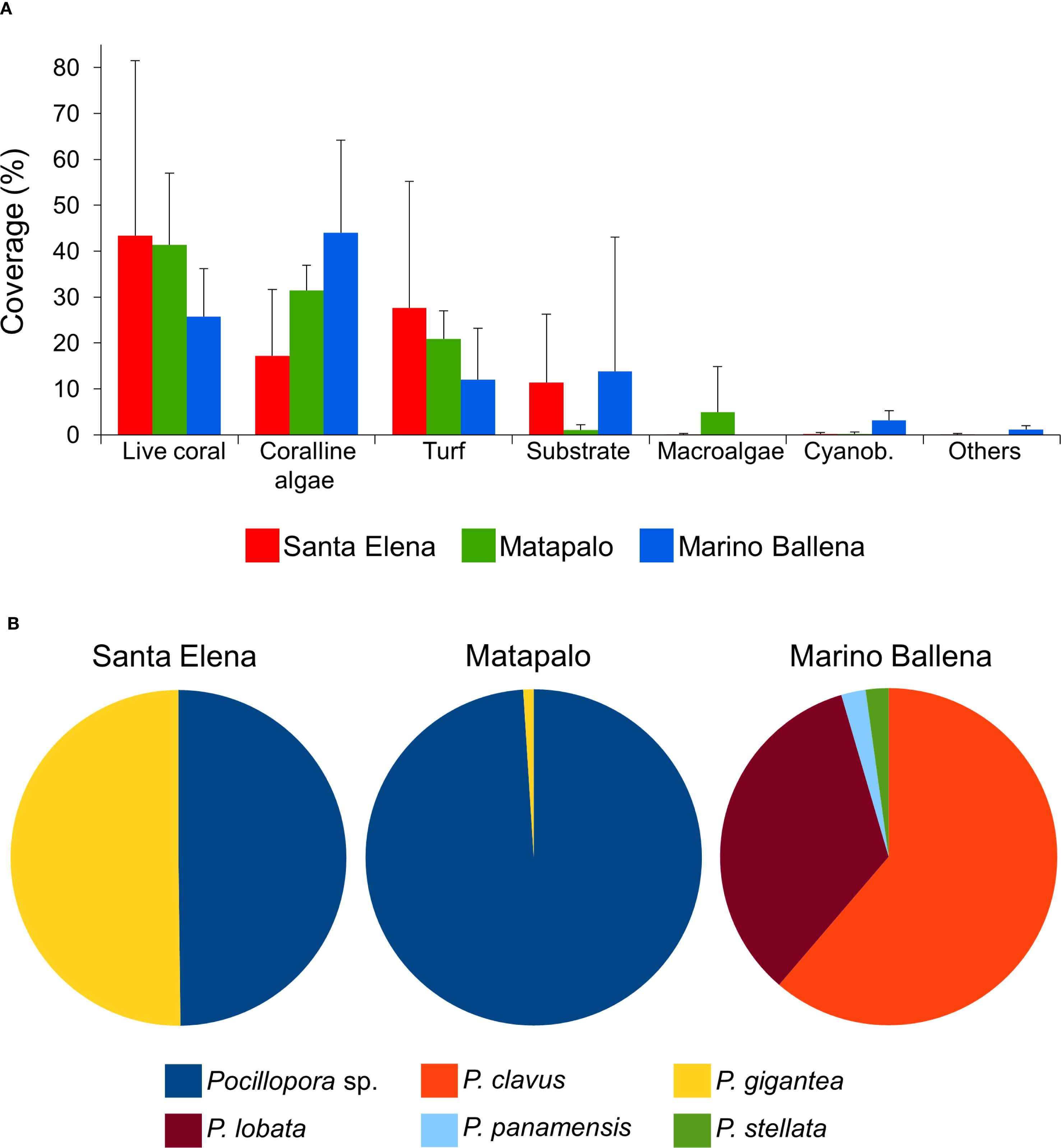
Figure 5. Benthic cover along the Pacific coast of Costa Rica. (A) Percentages of benthic cover reveal the dominant organisms in coral reefs from three locations, Cyanob. = cyanobacteria. (B) Contribution (%) of scleractinian coral species to coral coverage in the study locations.
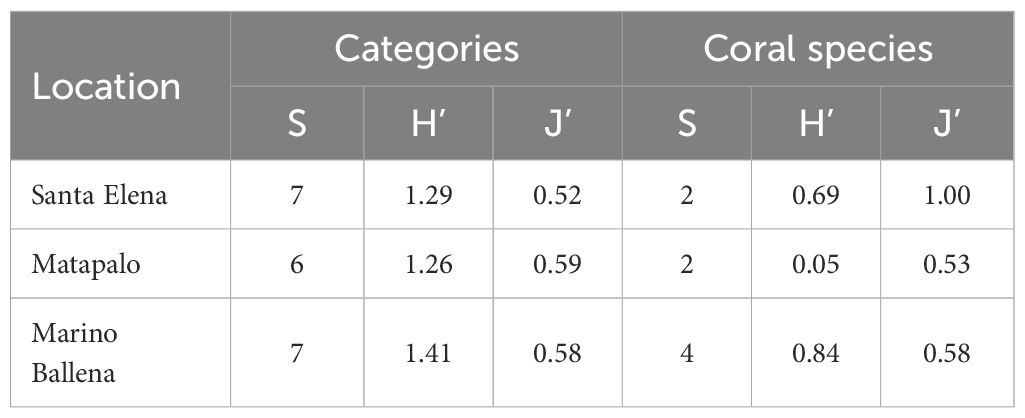
Table 3. Richness (S), diversity (H’) and evenness (J’) of benthic categories and scleractinian coral species in coral reefs at three locations in the Pacific coast of Costa Rica.
4 Discussion
4.1 Evidence of geographical and seasonal variability in the physicochemical conditions along the Pacific coast of Costa Rica
This study shows high spatial variability in the physicochemical parameters on coral reefs along the Costa Rican Pacific coast (Figure 2), and their seasonality seems to be controlled by different processes in each location. The Papagayo upwelling is the main driver of seasonal variability in water parameters in the north, while the rainfall during rainy season controls the variability in the south (Figure 3). Complementing previous studies that described the physical conditions caused by this coastal upwelling (Jiménez, 2001b; Fernández-García et al., 2012), our results show that the changes extend beyond a decrease in SWT and an increase in nutrient concentrations. The Papagayo coastal upwelling is also marked by an increase in nDIC, with the corresponding decline in pH and aragonite saturation state (Ωa) (Figure 4, Supplementary Table S1). Data from this study furthermore confirms the findings of a parallel survey conducted within the same research project which found a geographical gradient in the upwelling intensity between Santa Elena and Matapalo (Stuhldreier et al., 2015), with a greater range in the variability of physicochemical parameters in Santa Elena and a relatively low seasonality in Matapalo (Figures 2, 3, Supplementary Table S1). Therefore, we can affirm that in northern locations the variability in physicochemical parameters depends on the distance from the coastal upwelling. Our results allow comparisons not only within northern locations but also with southern reefs. In the following sections we will discuss the individual environmental parameters and their spatial and seasonal variability in more detail.
4.1.1 Seawater temperature and salinity
The SWT exhibits a reverse seasonality between northern (Santa Elena and Matapalo) and southern (Marino Ballena) locations. Coastal waters in the north are colder from December to April due to the occurrence of the Papagayo upwelling and start to get warmer around May (Alfaro and Cortés, 2012; Alfaro et al., 2012) during the transition to the rainy season. The range of SWT fluctuations in Santa Elena is very high during upwelling season (Cortés et al., 2014; Stuhldreier et al., 2015) and although the lowest value measured during our study was 18 °C, previous studies indicated that SWT in this area can drop to values below 14.0 °C for a few hours during strong upwelling events (Jiménez, 2001b). Conversely, in southern reefs the coastal waters experience a slight cooling during the second half of the year, as the rainy season develops.
This cooling in the south during the rainy season is related to greater freshwater input from numerous rivers near Marino Ballena. The Grande de Térraba River, for example, is located about 10 km southwest of Marino Ballena and drains one of the largest basins in the country (Krishnaswamy et al., 2001). Mean annual precipitation and flow discharge are significantly higher than in any other basin in the north (Waylen and Laporte, 1999). The average water temperature of this river is 25 °C (Umaña-Villalobos and Springer, 2006) and the river discharge is enhanced between the months of May and November (Rojas and Rodríguez, 2008), producing the cooling of the surface waters and a decrease in salinity to values of 26.0 psu (this study and Alvarado and Aguilar, 2009).
4.1.2 Nutrients
This study revealed that inorganic nutrients are the main contributors to the observed differences between locations and seasons in coastal waters along the Costa Rican Pacific. Our results confirm that the high variability of nutrient concentrations in Santa Elena during the dry season is driven by the Papagayo upwelling, with a significant increase of the mean values by 69% in phosphate, 80% in nitrate and 65% in ammonium, as compared to the non-upwelling rainy season. In contrast, most of the inorganic nutrient concentrations in Marino Ballena increased during the rainy season. In these southern reefs, agricultures, land erosion and runoff strongly contribute to the nutrient input carried by the Grande de Térraba River discharge (Alvarado et al., 2009) and other rivers in the area. Consequently, the average concentration of silicate in Marino Ballena during the rainy season was 48% higher as compared to the dry season (Figure 3), and phosphate and nitrate also had a significant increase in their concentrations during the rainy season of up to 125% and 74%, respectively.
4.1.3 Carbonate chemistry parameters
The spatial and temporal analysis of our data supports two major assumptions: 1) the northern Pacific coast of Costa Rica experiences an upwelling-driven geographic gradient in alkalinity, dissolved inorganic carbon and pH, and 2) seasonal variability in seawater carbonate chemistry is mainly driven by the Papagayo upwelling during the dry season in northern locations and by river discharges during the rainy season in the south. Dynamics of Ωa and pH in coastal areas are driven by several processes: i) upwelling of high-CO2 waters (Feely et al., 2008; Rixen et al., 2012; Harris et al., 2013; Sánchez-Noguera et al., 2018b), ii) metabolic processes controlled by planktonic communities and coral reefs (inorganic:organic carbon production ratios) (Gray et al., 2012; Albright et al., 2013, 2015) and iii) inputs from land (Vargas et al., 2016; Dong et al., 2017; Carstensen and Duarte, 2019). In this study, the northernmost location (Santa Elena) and southern location (Marino Ballena) experienced low Ωa values; however, the source of these reduced values and their variability differed between the two locations. In Santa Elena, the lower pH and Ωa was linked to the upwelling of waters with high-CO2 concentrations (Rixen et al., 2012). In contrast, in Marino Ballena the dilution effect by freshwater input was likely responsible for the observed decrease in Ωa values, due to river discharges and increased runoff during the rainy season (Chierici and Fransson, 2009; Harris et al., 2013).
By analyzing the coupled changes of salinity-normalized alkalinity and dissolved inorganic carbon (Figure 4) in combination with the estimated inorganic:organic carbon production ratios, we can shed light on which processes are exerting a major control over the carbonate system in our study locations. In Santa Elena, the observed high variability of nDIC between seasons confirms that the upwelling of CO2-enriched waters is partially responsible for the seasonal fluctuation in seawater carbonate chemistry. This result agrees with other studies (Rixen et al., 2012; Sánchez-Noguera et al., 2018b), showing that in the Papagayo upwelling system deep waters hauled to the surface are enriched in CO2. Additionally, the estimated inorganic:organic carbon production ratio in Santa Elena (< 1) suggests that organic carbon production and respiration also play a role in the seasonal variability of carbonate chemistry. The linear regression analysis of nDIC vs. nTA (Figure 4) suggests that organic carbon production via photosynthesis was enhanced during the rainy season and respiration during the dry season. In line with our findings, other authors have indicated that enhanced primary production can be responsible for decreasing the concentrations of dissolved inorganic carbon, nitrite and nitrate (He et al., 2025), as occurred in Santa Elena during the rainy season. In terms of the enhanced respiration during the dry season, previous studies reported that the abundance of zooplankton in the Gulf of Papagayo is higher during the dry season (Bednarski and Morales-Ramírez, 2004; Rodríguez-Sáenz and Morales-Ramírez, 2012). Therefore, upwelled nutrient-enriched waters can boost the respiration of zooplankton communities, partially contributing to the observed increase in dissolved inorganic carbon during the upwelling season. Similarly, in the California Current System the high concentration of dissolved inorganic carbon is mostly attributed to respiration-derived CO2 (Hauri et al., 2009). On the other hand, the slight enrichment of alkalinity measured in Santa Elena during the dry season could be attributed to the dissolution of CaCO3 in deep waters. Similar to Santa Elena, the estimated inorganic:organic carbon production ratio in Matapalo was also < 1, highlighting the importance of organic production and respiration processes in the carbonate system of northern locations. Moreover, the small seasonal variability in alkalinity and dissolved inorganic carbon measured in Matapalo, in combination with higher pH (> 8.0) and Ωa (> 3.4) values, suggests that CaCO3 is precipitated all year round in this location.
Moving southward to Marino Ballena, the coupled increase of nDIC and nTA (Figure 4) suggest that inorganic processes (i.e. carbonate production or dissolution) could play a key role in the seawater chemistry of this location. However, the estimated inorganic:organic carbon production ratio (0.88) challenges this assumption, indicating that organic processes also play a dominant role in driving the seasonal variability of the carbonate system in this location. Nevertheless, the carbon production ratio in Marino Ballena is considerably higher than in northern locations (Santa Elena = 0.04, Matapalo = 0.18) and closer to a value of 1, implying that the inorganic processes may have a relatively stronger influence on the dynamics of the carbonate system, potentially linked to CaCO3 formation, which is somehow promoted during the dry season. Considering that the Grande de Térraba River drains the largest karstic region in Costa Rica (about 185 km2 of extension) (Ulloa et al., 2011, 2024; Bolz and Calvo, 2018), is very likely that surface and groundwater discharges supply dissolved carbonates to the reefs in this area (Krishnaswamy et al., 2001; Tamše et al., 2014; Kapsenberg et al., 2017; He et al., 2025). This could help explain the slightly elevated nTA observed in Marino Ballena during the dry season, in comparison to Matapalo, a northern location where seasonal upwelling exerts a less pronounced influence on the variability of the carbonate system parameters. A thorough analysis of other carbonate system parameters showed that the fluctuations of nDIC and nTA in Marino Ballena did not correspond with seasonal variations in pH values. The pH levels in Marino Ballena remained consistent across both seasons, with values of about 8.0, which might be caused by the enhanced outwelling of carbonates from nearby mangroves (i.e. Térraba-Sierpe National Wetland) during the rainy season. This is consistent with conclusions from another study, suggesting that mangroves can contribute to buffer coastal acidification (Sippo et al., 2016).
4.2 Environmental conditions shape coral reef composition in the Costa Rican Pacific coast
Even though all study locations displayed a similar benthic richness and diversity; the main reef-building corals are different along the Costa Rican Pacific coast (Figure 5B). Reefs from northern locations are dominated by massive and branching species, whereas reefs from the southern part of this coast are mostly dominated by massive species (this study and Glynn et al., 2017a). A few and small patch reefs of branching-corals have been reported in southern locations (Guzmán and Cortés, 1989; Cortés and Jiménez, 1996), but they are surpassed in number by massive-coral reefs and mostly found in Caño Island, 16 km northwest from the coast. The coral distribution patterns described above (Figure 5B) raise two key questions: 1) Why are the massive reef-building species in the north different from the massive reef-building species in the south? 2) Why are branching corals not among the main reef-builders in the south?
The measured physicochemical parameters revealed spatial and seasonal differences in the conditions along the Costa Rican Pacific coast (Figures 2, 3). However, changes of physicochemical parameters seem to be insufficient to explain why the main reef-building species differ between locations (Figure 5). To find a possible explanation for the observed coral distribution patterns, we also compared our local values with the established global environmental ranges suitable for potential coral reef habitats (Table 4) (Guan et al., 2015). These global values were used to update the ReefHab model (Kleypas, 1995, 1997), originally developed to predict the potential distribution of coral reefs according to the available environmental data. Kleypas’ work was a milestone that stated for the first time the environmental tolerance limits for coral reefs, contributing to understanding and predicting their global distribution. Comparing the physicochemical conditions on the Pacific coast of Costa Rica (this study) with the global thresholds established by the updated ReefHab model (Guan et al., 2015) helps to explain the presence of coral reefs in our study locations, but does not necessarily capture the variability in species composition. Nevertheless, given the limited information available on the environmental tolerance ranges for individual coral genera, we also used these global thresholds as a proxy to understand the differences in the dominant coral species observed at our three study locations. This combined analysis suggests that the composition of the dominant coral species in this study can be mainly explained by phosphate and Ωa, as these parameters divert from the global tolerance thresholds in different ways (Table 4).
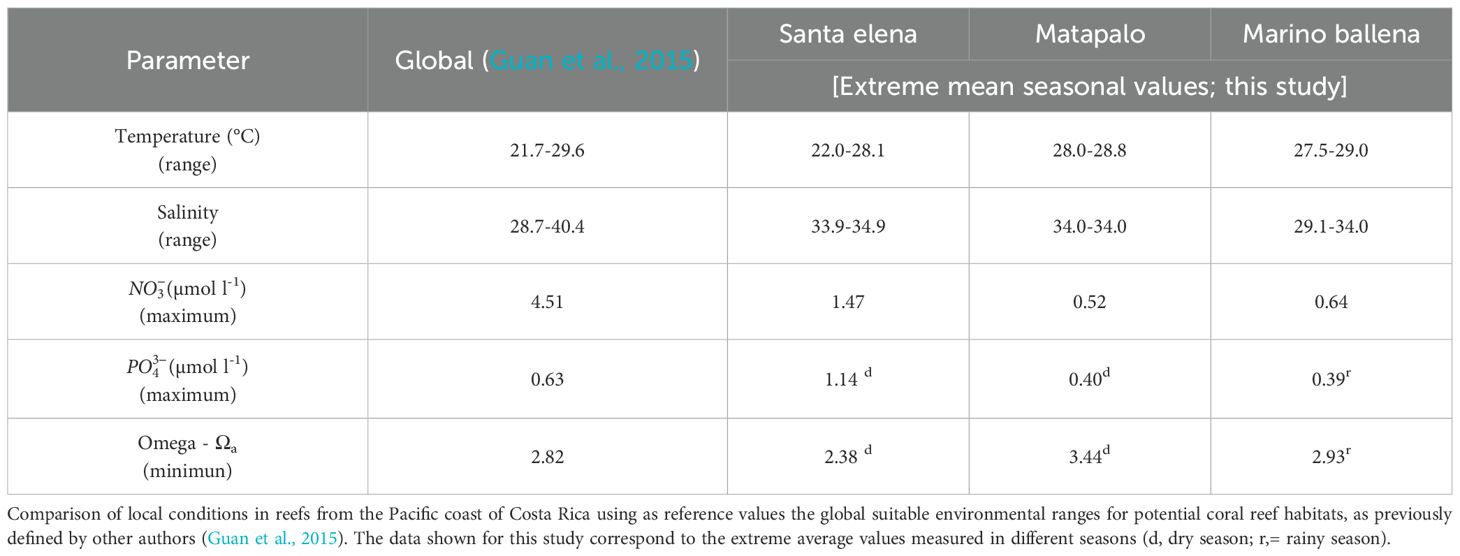
Table 4. Comparison of local conditions in reefs from the Pacific coast of Costa Rica using as reference values the global suitable environmental ranges for potential coral reef habitats, as previously defined by other authors (Guan et al., 2015). The data shown for this study correspond to the extreme average values measured in different seasons (d = dry season, r = rainy season).
In the following paragraphs, we will use the presence of coral reefs as an indicator of the success of those coral species to cope with the combination of the measured physicochemical parameters, therefore a proxy of the suitability of local conditions for the reef development along this coast. To address the first question regarding the observed differences in massive coral species building reefs along this coast (Figure 5B), it is important to mention that even though P. clavus was not present in the reefs surveyed in Santa Elena and Matapalo, this massive coral was reported in the past as the main reef-builder (Jiménez, 1997) or among the most ubiquitous massive species (Palmer et al., 2022) in other northern location. We excluded that P. clavus reef from this study because it experienced a mass mortality several years before, thus the benthic coverage at the time of our study was mainly dominated by turf algae (Sánchez-Noguera et al., 2018a). By considering the historical record of coral reefs in the north Pacific of Costa Rica in addition to our data, we can say that P. clavus succeeds to build reefs in both northern and southern locations. This indicates that this massive coral is highly tolerant to the wide range of physicochemical conditions measured along this coast. Nevertheless, it is likely that they were already living near their physiological tolerance limits in northern locations, which might explain their historical mortality in those areas (Mena et al., 2025). Now we will focus our attention on the massive coral species that are building reefs exclusively in northern or southern locations and are not widely distributed as P. clavus. Our data suggests that P. gigantea, one of the main massive reef-builders in the north Pacific coast of Costa Rica, is highly tolerant to the episodic exposition to low- Ωa cool waters and eutrophic conditions occurring in pulses during upwelling season. This agrees with results from the Pacific coast of Panama, where corals from the genus Pavona exhibited a high tolerance to the combination of thermal stress (lower SWT) and acidification in the coastal upwelling impacted Gulf of Panama (Manzello, 2010a). In contrast, P. cf. lobata building reefs in Marino Ballena indicates that this massive coral thrives in constant conditions of lower Ωa (< 3.0) and lower salinity, and copes very well with low light intensity due to the influx of terrigenous materials occurring predominantly during the rainy season (Alvarado et al., 2009). Despite the mechanisms by which inorganic nutrients affect coral reefs are subject to debate, there is a consensus that unbalanced nutrient enrichment from anthropogenic sources poses negative effects for coral reef functioning (D’Angelo and Wiedenmann, 2014). For example, high nutrient loads have the potential to affect corals indirectly by favoring primary producers, which can outcompete stony corals (Fernández-García et al., 2012), and increasing bioerosion by facilitating shifts to larger and more effective bioeroders (Wizemann et al., 2018). Furthermore, a study from Galapagos concerning reefs affected by different upwelling levels suggested that the skeletal density of P. lobata was diminished due to exposure to elevated phosphate conditions (Manzello et al., 2014). This can contribute to explaining why P. cf. lobata was one of the main reef builders in Marino Ballena. The average seasonal phosphate levels in Marino Ballena were lower than those in Santa Elena, which likely promotes stronger skeletons in Marino Ballena and allows their persistence over time. This suggests that the lower phosphate levels enable P. cf. lobata to build reefs in Marino Ballena, despite the steady Ωa values < 3.0 measured in southern locations. Moreover, other studies involving corals from the genus Porites support our assumption that changes driven by river discharges significantly shape the structure of local coral reefs. For instance, Porites corals in Biscayne Bay, Florida show a high tolerance to changes in salinity (Manzello and Lirman, 2003), while those in Golfo Dulce, Costa Rica, are particularly resilient to high sedimentation (Cortés, 1990). In consequence, this species contributes the most to the reef building under riverine impacted locations.
Now we will move forward to our second question, regarding the minor contribution of branching corals in southern locations. In Costa Rica, most reefs built by Pocillopora spp. have experienced severe mortalities, frequently attributed to warming during ENSO events and to recurrent occurrence of harmful algal blooms (Guzmán et al., 1987, 1990; Glynn et al., 2017a; Sánchez-Noguera et al., 2018a; Palmer et al., 2022). Nevertheless, the current distribution of branching corals and the remaining structures of former reefs can be used as a proxy to identify the most favorable conditions for the success of this genus along the Costa Rican coast. For example, the precipitation of CaCO3 over the whole year at Matapalo, along with its sheltered location, helps explain the presence of a reef built by branching species. This combination of factors contributes to the branching species’ prevalence and relatively high contribution to coral coverage compared to other coral reefs in the same area, which have experienced significant degradation over the last decade (Mena et al., 2025). While branching corals are not entirely absent from southern locations (Guzmán and Cortés, 1989; Cortés and Jiménez, 1996), they contribute much less to the benthic coverage than massive corals in Marino Ballena. This is likely due to the reduced light availability caused by river discharge. Grande de Térraba River, located near Marino Ballena, is the second largest catchment of the country (Granados-Bolaños et al., 2024). Other authors (Alvarado et al., 2005) have previously discussed that the limited presence of Pocillopora spp. in Marino Ballena may be linked to low light penetration in the water column, due to high concentrations of suspended matter derived from river discharge (Alvarado et al., 2009). This hypothesis is confirmed by another study from the Great Barrier Reefs, which showed that the genus Pocillopora is very sensitive to reduced light resulting from high turbidity (Jones et al., 2020). Therefore, we can infer that reduced light availability is one of the main factors limiting reef development by branching corals in southern locations, while high tolerance to salinity fluctuations promotes the prevalence of reefs built by massive corals such as Porites sp.
The results presented in this study provide insights into adaptation limits of individual coral species to local physicochemical conditions. For example, Pavona spp. and Pocillopora spp. reefs from Santa Elena indicate that these species have adapted to the episodic exposition to low- Ωa conditions occurring during upwelling season (2.38, Table 4), when Ωa values are lower than the proposed local (Sánchez-Noguera et al., 2018b) and global thresholds (2.82, Table 4) (Guan et al., 2015) for reef development. Even though the success of the massive and branching species to build coral reefs along the Pacific coast is a good example of adaptation to high local variability, it does not guarantee their survival under climate change scenarios, when the rate of changes will be enhanced (Howes et al., 2015). Costa Rican coral reefs have suffered several ENSO-induced bleaching events over the last decades (Guzmán et al., 1987; Guzmán and Cortés, 1992, 2001; Jiménez and Cortés, 2001, 2003; Jiménez et al., 2001; Cortés et al., 2010; Alvarado et al., 2018), while the current lapse between global bleaching events is shortening the temporal window for reef recovery after associated mass mortality (Hughes et al., 2018). This implies that physicochemical conditions in the ETP are close to the limits that warm water reef ecosystems can tolerate.
4.3 Outlook
The results from our study demonstrate that seawater chemistry properties of coastal waters along the Costa Rican Pacific coast display a high spatial and temporal variability, which is driven by the Papagayo upwelling system during the dry season in the north and river discharge during the rainy season in the south. Matapalo reef, located between the northernmost (Santa Elena) and the southernmost location (Marino Ballena), reveals a relatively low seasonality of physicochemical conditions. This confirms that the seasonal upwelling has a stronger influence further north, in Santa Elena relative to Matapalo. Although carbonate chemistry plays a significant role in determining the composition of coral reefs along the Pacific coast of Costa Rica, it is not the only controlling factor, as inorganic nutrients explain a higher percentage of the observed variability. Our findings indicate that carbonate chemistry is key for promoting favorable physicochemical conditions for reef development in northern locations like Matapalo. However, the sensitivity of coral species to other physicochemical conditions, such as lower salinity and lower light intensity, are responsible for shaping the reef composition in Marino Ballena (Figure 6).
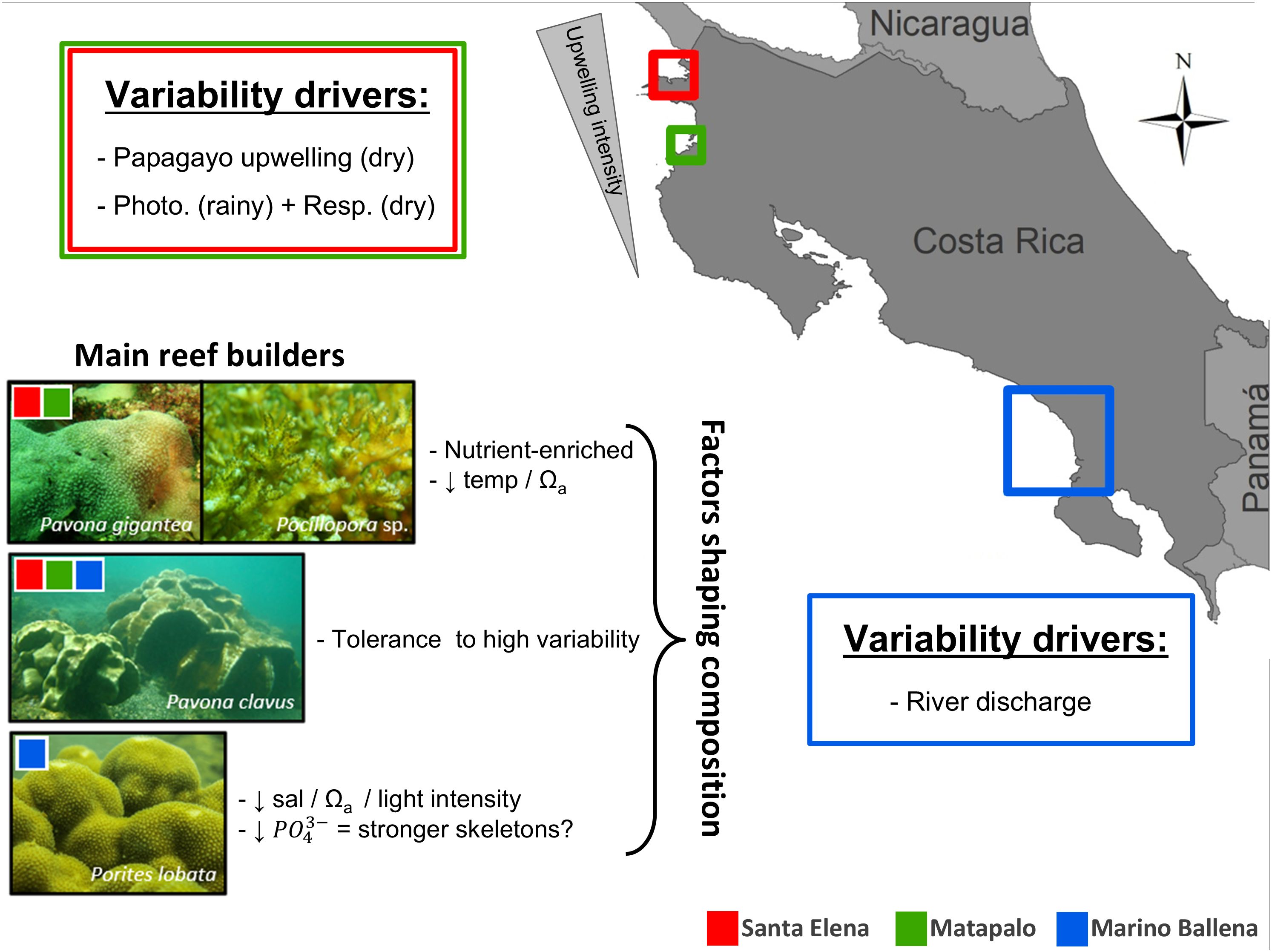
Figure 6. Graphic summary of main findings. Drivers of spatial and seasonal variability in nutrient concentrations and carbonate chemistry, and main factors controlling coral reef composition along the Pacific coast of Costa Rica.
Corals from the Pacific coast of Costa Rica have adapted to the local variability in physicochemical parameters. Nevertheless, their tolerance to this variability appears to be species-specific, and does not imply that they can withstand external disturbances that push physicochemical conditions beyond the limits for reef development. Regarding future conservation efforts and reef protection, Matapalo and Marino Ballena seem to be very promising locations. This is mainly to the relatively stable carbonate chemistry conditions in Matapalo and the buffering effect against ocean acidification in Marino Ballena, which is most probably caused by the dissolved carbonates entering from land. However, this calls for integrated coastal management actions to reduce the pressures of local stressors such as siltation, pollution, fishing and tourism.
Data availability statement
The datasets generated and analyzed for this study are publicly available and can be found in PANGAEA (https://www.pangaea.de/), under the following DOI: https://doi.pangaea.de/10.1594/PANGAEA.920884, https://doi.pangaea.de/10.1594/PANGAEA.920876, https://doi.pangaea.de/10.1594/PANGAEA.920881, https://doi.pangaea.de/10.1594/PANGAEA.920883, https://doi.pangaea.de/10.1594/PANGAEA.920882.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
CS-N: Conceptualization, Formal analysis, Methodology, Writing – original draft, Investigation, Data curation. IL: Methodology, Writing – review & editing. JC: Writing – review & editing. CJ: Writing – review & editing. CW: Writing – review & editing, Funding acquisition. TR: Supervision, Writing – review & editing, Funding acquisition, Project administration.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This study was funded by the Leibniz Association as part of COSTACID project from the Leibniz Centre for Tropical Marine Research (ZMT). The publication (APC) of this article was funded by Universidad de Costa Rica through Vicerrectoría de Investgación and Red de Mujeres en Ciencias, Ingenierías y Humanidades (CIHRED-UCR).
Acknowledgments
Special thanks to Minor Lara and family, Fernando Monge and the staff from Scuba Caribe Diving Center for their support in the field. We also thank Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), Hotel RIU Guanacaste (Alberto Leónard) and Scuba Caribe for logistic support. Juan José Alvarado, Gustavo Arias and Jeffrey Sibaja-Cordero provided valuable advice in statistical analysis. The constructive comments of two reviewers improved the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1606253/full#supplementary-material
Supplementary Table 1 | Overall average values (95% confidence intervals) by location and season of measured and calculated (*) physicochemical and meteorological parameters at three locations in the Pacific coast of Costa Rica. Seawater temperature (SWT) was measured in situ and the values for the other parameters were obtained from discrete water samples. For abbreviations of listed parameters please referred to the manuscript. Meteorological data collected at meteorological stations located near the study locations was provided by the Instituto Meteorológico Nacional.
Supplementary Table 2 | Statistical differences of physicochemical and meteorological parameters between locations. Asterisks indicate significant differences (p < 0.05).
Supplementary Table 3 | Statistical differences of physicochemical and meteorological parameters between seasons. Asterisks indicate significant differences (p < 0.05).
References
Albright R., Benthuysen J., Cantin N., Caldeira K., and Anthony K. (2015). Coral reef metabolism and carbon chemistry dynamics of a coral reef flat. Geophys Res. Lett. 42, 3980–3988. doi: 10.1002/2015gl063488
Albright R., Langdon C., and Anthony K. (2013). Dynamics of seawater carbonate chemistry, production, and calcification of a coral reef flat, central Great Barrier Reef. Biogeosciences 10, 6747–6758. doi: 10.5194/bg-10-6747-2013
Alfaro E. J. and Cortés J. (2012). Atmospheric forcing of cool subsurface water events in Bahía Culebra, Gulf of Papagayo, Costa Rica. Rev. Biol. Trop. 60, 173–186. doi: 10.15517/rbt.v60i2.20001
Alfaro E. J., Cortés J., Alvarado J. J., Jiménez C., León A., Sánchez-Noguera C., et al. (2012). Clima y temperatura sub-superficial del mar en Bahía Culebra, Golfo de Papagayo, Costa Rica. Rev. Biol. Trop. 60, 159–171. doi: 10.15517/rbt.v60i2.20000
Alvarado J. J. and Aguilar J. F. (2009). Batimetría, salinidad, temperatura y oxígeno disuelto en aguas del Parque Nacional Marino Ballena, Pacífico, Costa Rica. Rev. Biol. Trop. 57, 19–29. doi: 10.15517/rbt.v57i0.21273
Alvarado J., Beita-Jiménez A., Mena S., Fernández-García C., Cortés J., Sánchez-Noguera C., et al. (2018). Cuando la conservación no puede seguir el ritmo del desarrollo: Estado de salud de los ecosistemas coralinos del Pacífico Norte de Costa Rica. Rev. Biol. Trop. 66, S280–S308. doi: 10.15517/rbt.v66i1.33300
Alvarado J. J., Cortés J., Fernández C., and Nivia-Ruiz J. (2005). Coral communities and coral reefs of Ballena Marine National Park, Pacific coast of Costa Rica. Cienc Mar. 31, 641–651. doi: 10.7773/cm.v31i4.1140
Alvarado J. J., Fernández C., and Cortés J. (2009). Water quality conditions on coral reefs at the Marino Ballena National Park, Pacific Costa Rica. Bull. Mar. Sci. 84, 137–152.
Alvarado J. J., Fernández C., and Nielsen V. (2006). “Arrecifes y Comunidades Coralinas,” in Informe técnico: Ambientes Marino Costeros de Costa Rica. Eds. Nielsen-Muñoz V. and Quesada-Alpízar M. (San José, Costa Rica: The Nature Conservancy), 51–68.
Bednarski M. and Morales-Ramírez A. (2004). Composition, abundance and distribution of macrozooplankton in Culebra Bay, Gulf of Papagayo, Pacific coast of Costa Rica and its value as bioindicators of pollution. Rev. Biol. Trop. 53, 105–118.
Bolz A. and Calvo C. (2018). Carbonate platform development in an intraoceanic arc setting: Costa Rica’s largest limestone sequence - the fila de cal formation (Middle eocene to lower oligocene). Rev. Geológica América Cent. 58, 85–114. doi: 10.15517/rgac.v58i0.32845
Cambronero-Solano S., Tisseaux-Navarro A., Vargas-Hernández J. M., Salazar-Ceciliano J. P., Benavides-Morera R., Quesada-ávila I., et al. (2021). Hydrographic variability in the Gulf of Papagayo, Costa Rica during 2017-2019. Rev. Biol. Trop. 69, S74–S93. doi: 10.15517/rbt.v69is2.48308
Carstensen J. and Duarte C. (2019). Drivers of pH variability in coastal ecosystems. Environ. Sci. Technol. 53, 4020–4029. doi: 10.1021/acs.est.8b03655
Chierici M. and Fransson A. (2009). Calcium carbonate saturation in the surface water of the Arctic Ocean: undersaturation in freshwater influenced shelves. Biogeosciences 6, 2421–2431. doi: 10.5194/bg-6-2421-2009
Cortés J. (1990). The coral reefs of Golfo Dulce, Costa Rica: distribution and community structure. Atoll Res. Bull. 339–346, 1–37. doi: 10.5479/si.00775630.344.1
Cortés J. (1997). Biology and geology of eastern Pacific coral reefs. Coral Reefs 16, S39–S46. doi: 10.1007/s003380050240
Cortés J. (2016). “The Pacific Coastal and Marine Ecosystems,” in Costa Rican Ecosystems. Ed. Kappelle M. (The University of Chicago Press, Chicago), 97–138.
Cortés J. and Jiménez C. (1996). Coastal-marine environments of Parque Nacional Corcovado, Puntarenas, Costa Rica. Rev. Biol. Trop. 44, 35–40.
Cortés J. and Jiménez C. (2003). “Corals and Coral Reefs of the Pacific of Costa Rica: History, Research and Status,” in Latin American Coral Reefs. Ed. Cortés J. (Elsevier Science B.V, Amsterdan, The Netherlands), 361–385.
Cortés J., Jiménez C., Fonseca A., and Alvarado J. (2010). Status and conservation of coral reefs in Costa Rica. Rev. Biol. Trop. 58, 33–50. doi: 10.15517/rbt.v58i1.20022
Cortés J., Samper-Villarreal J., and Bernecker A. (2014). Seasonal phenology of Sargassum liebmannii J. Agardh (Fucales, Heterokontophyta) in an upwelling area of the Eastern Tropical Pacific. Aquat Bot. 119, 105–110. doi: 10.1016/j.aquabot.2014.08.009
D’Angelo C. and Wiedenmann J. (2014). Impacts of nutrient enrichment on coral reefs: new perspectives and implications for coastal management and reef survival. Curr. Opin. Environ. Sustain 7, 82–93. doi: 10.1016/j.cosust.2013.11.029
Dana T. (1975). Development of contemporary Eastern Pacific coral reefs. Mar. Biol. 33, 355–374. doi: 10.1007/bf00390574
Dickson A. (1990). Thermodynamics of the dissociation of boric acid in synthetic seawater from 273.15 to 318.15 K. Deep Sea Res. Part A. Oceanographic Res. Papers 37, 755–766. doi: 10.1016/0198-0149(90)90004-F
Dickson A., Afghan J., and Anderson G. (2003). Reference materials for oceanic CO2 analysis: a method for the certification of total alkalinity. Mar. Chem. 80, 185–197. doi: 10.1016/S0304-4203(02)00133-0
Dickson A. and Millero F. (1987). A comparison of the equilibrium constants for the dissociation of carbonic acid in seawater media. Deep Sea Res. Part A. Oceanographic Res. Papers 34, 1733–1743. doi: 10.1016/0198-0149(87)90021-5
Dickson A., Sabine C., and Christian J. (2007). Guide to best practices for ocean CO2 measurements PICES Special Publication 3. Sidney, BC, Canada: North Pacific Marine Science Organization (PICES).
Dong X., Huang H., Zheng N., Pan A., Wang S., Huo C., et al. (2017). Acidification mediated by a river plume and coastal upwelling on a fringing reef at the east coast of Hainan Island, Northern South China Sea. J. Geophysical Resarch Oceans 122, 7521–7536. doi: 10.1002/2017JC013228
Feely R., Sabine C., Hernández-Ayón J., Ianson D., and Hales B. (2008). Evidence for upwelling of corrosive “Acidified” Water onto the continental shelf. Sci. (1979) 320, 1490–1492. doi: 10.1126/science.1155676
Fernández-García C., Cortés J., Alvarado J., and Nivia-Ruiz J. (2012). Physical factors contributing to the benthic dominance of the alga Caulerpa sertularioides (Caulerpaceae, Chlorophyta) in the upwelling Bahía Culebra, north Pacific of Costa Rica. Rev. Biol. Trop. 60, 93–107. doi: 10.1007/s003380050240
Fiedler P. and Lavín M. (2017). “Oceanographic Conditions of the Eastern Tropical Pacific,” in Coral Reefs of the Eastern Tropical Pacific: Persistence and Loss in a Dynamic Environment. Eds. Glynn P., Manzello D., and Enochs I. (Springer Netherlands, Dordrecht), 59–83.
Fiedler P. and Talley L. (2006). Hydrography of the eastern tropical Pacific: A review. Prog. Oceanogr 69, 143–180. doi: 10.1016/j.pocean.2006.03.008
Glynn P., Alvarado J., Banks S., Cortés J., Feingold J., Jiménez C., et al. (2017a). “Eastern Pacific Coral Reef Provinces, Coral Community Structure and Composition: an Overview,” in Coral Reefs of the Eastern Tropical Pacific: persistence and loss in a dynamic environment. Eds. Glynn P., Manzello D., and Enochs I. (Springer Science+Business Media B.V. Dordrecht, Dordrecht), 107–176. doi: 10.1007/978-94-017-7499-4_5
Glynn P., Mones A., Podestá G., Colbert A., and Colgan M. (2017b). “El Niño-Southern Oscillation: Effects on Eastern Pacific Coral Reefs and Associated Biota,” in Coral reefs of the Eastern tropical Pacific: Persistence and loss in a dynamic environment. Eds. Glynn P., Manzello D., and Enochs I. (Springer Science+Business Media B.V. Dordrecht, Dordrecht), 251–290.
Granados-Bolaños S., Surian N., Birkel C., Alvarado G. E., Quesada-Román A., Galve J. P., et al. (2024). “Fluvial Landscapes of Costa Rica: An Overview of Dynamic Rivers,” in Landscapes and Landforms of Costa Rica. Ed. Quesada-Román A. (Springer International Publishing, Cham), 91–111. doi: 10.1007/978-3-031-64940-0_5
Gray S., DeGrandpre M., Langdon C., and Corredor J. (2012). Short-term and seasonal pH, pCO2 and saturation state variability in a coral-reef ecosystem. Global Biogeochem Cycles 26, 1–13. doi: 10.1029/2011GB004114
Guan Y., Hohn S., and Merico A. (2015). Suitable environmental ranges for potential coral reef habitats in the tropical ocean. PloS One 10, e0128831. doi: 10.1371/journal.pone.0128831
Guzmán H. and Cortés J. (1989). Coral reef community structure at Caño Island, Pacific Costa Rica. Mar. Ecol. 10, 23–41. doi: 10.1111/j.1439-0485.1989.tb00064.x
Guzmán H. and Cortés J. (1992). Cocos Island (Pacific of Costa Rica) coral reefs after the 1982–83 El Niño disturbance. Rev. Biol. Trop. 40, 309–324.
Guzmán H. and Cortés J. (2001). Changes in reef community structure after fifteen years of natural disturbances in the Eastern Pacific (Costa Rica). Bull. Mar. Sci. 69, 133–149.
Guzmán H., Cortés J., Glynn P., and Richmond R. (1990). Coral mortality associated with dino-flagellate blooms in the eastern Pacific (Costa Rica and Panama). Mar. Ecol. Prog. Ser. 60, 299–303. doi: 10.3354/meps060299
Guzmán H., Cortés J., Richmond R., and Glynn P. (1987). Efectos del fenómeno de “El Niño Oscilacion Sureña” 1982/83 en los arrecifes coralinos de la Isla del Caño, Costa Rica. Rev. Biol. Trop. 35, 325–332.
Harris K., DeGrandpre M., and Hales B. (2013). Aragonite saturation state dynamics in a coastal upwelling zone. Geophys Res. Lett. 40, 2720–2725. doi: 10.1002/grl.50460
Hauri C., Gruber N., Plattner G., Alin S., Feely R., Hales B., et al. (2009). Ocean acidification in the california current system. Oceanography 22, 60–71. doi: 10.5670/oceanog.2009.97
He S., Gordon S., and Maiti K. (2025). Carbonate and nutrient dynamics in a mississippi river influenced eutrophic estuary. Estuaries Coasts 48, 63. doi: 10.1007/s12237-025-01494-4
Howes E., Joos F., Eakin M., and Gattuso J. (2015). An updated synthesis of the observed and projected impacts of climate change on the chemical, physical and biological processes in the oceans. Front. Mar. Sci. 2. doi: 10.3389/fmars.2015.00036
Hughes T., Anderson K., Connolly S., Heron S., Kerry J., Lough J., et al. (2018). Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Sci. (1979) 359, 80–83. doi: 10.1126/science.aan8048
Instituto Metereológico Nacional (IMN) (2008). El clima, su variabilidad y cambio climático en Costa Rica (San José: Comité Regional de Recursos Hidráulicos (CRRH)).
Jiménez C. (1997). Corals and coral reefs of Culebra Bay, Pacific coast of Costa Rica: Anarchy in the reef. Proc. 8th Int. Coral Reef Symposium 1, 329–334.
Jiménez C. (2001a). Arrecifes y ambientes coralinos de Bahía Culebra, Pacífico de Costa Rica: Aspectos biológicos, económico-recreativos y de manejo. Rev. Biol. Trop. 49, 215–231.
Jiménez C. (2001b). Seawater temperature measured at the surface and at two depths (7 and 12 m) in one coral reef at Culebra Bay, Gulf of Papagayo, Costa Rica. Rev. Biol. Trop. 49, 153–161.
Jiménez C., Bassey G., Segura A., and Cortés J. (2010). Characterization of the coral communities and reefs of two previously undescribed locations in the upwelling region of Gulf of Papagayo (Costa Rica). Rev. Mar. Cost. 2, 95–108. doi: 10.15359/revmar.2.8
Jiménez C. and Cortés J. (2001). Effects of the 1991–92 El Niño on scleractinian corals of the Costa Rican central Pacific coast. Rev. Biol. Trop. 49, 239–250.
Jiménez C. and Cortés J. (2003). Coral cover change associated to El Niño, Eastern Pacific, Costa Rica 1992-2001. Mar. Ecol. 24, 179–192. doi: 10.1046/j.1439-0485.2003.03814.x
Jiménez C., Cortés J., León A., and Ruíz E. (2001). Coral bleaching and mortality associated with the 1997–98 El Niño in an upwelling environment in the Eastern Pacific (Gulf of Papagayo, Costa Rica). Bull. Mar. Sci. 69, 151–169.
Jones R., Giofre N., Luter H. M., Neoh T. L., Fisher R., and Duckworth A. (2020). Responses of corals to chronic turbidity. Sci. Rep. 10, 4762. doi: 10.1038/s41598-020-61712-w
Kapsenberg L., Alliouane S., Gazeau F., Mousseau L., and Gattuso J.-P. (2017). Coastal ocean acidification and increasing total alkalinity in the northwestern Mediterranean Sea. Ocean Sci. 13, 411–426. doi: 10.5194/os-13-411-2017
Kleypas J. (1995). “A Diagnostic Model for Predicting Global Coral Reef Distribution,” in Recent Advances in Marine Science and Technology. Eds. Bellwood O., Choat H., and Saxena N. (Honolulu, HI, USA: PACON International and James Cook University), 211–220.
Kleypas J. (1997). Modeled estimates of global reef habitat and carbonate production since the last glacial maximum. Paleoceanography 12, 533–545. doi: 10.1029/97PA01134
Krishnaswamy J., Halpin P., and Richter D. (2001). Dynamics of sediment discharge in relation to land-use and hydro-climatology in a humid tropical watershed in Costa Rica. J. Hydrol (Amst) 253, 91–109. doi: 10.1016/S0022-1694(01)00474-7
Lavín M., Fiedler P., Amador J., Ballance L., Färber-Lorda J., and Mestas-Nuñez A. (2006). A review of eastern tropical Pacific oceanography: Summary. Prog. Oceanogr 69, 391–398. doi: 10.1016/j.pocean.2006.03.005
Manzello D. (2010a). Coral growth with thermal stress and ocean acidification: lessons from the eastern tropical Pacific. Coral Reefs 29, 749–758. doi: 10.1007/s00338-010-0623-4
Manzello D. (2010b). Ocean acidification hotspots: Spatiotemporal dynamics of the seawater CO2 system of eastern Pacific coral reefs. Limnol Oceanogr. 55, 239–248. doi: 10.4319/lo.2010.55.1.0239
Manzello D., Enochs I., Bruckner A., Renaud P., Kolodziej G., Budd D., et al. (2014). Galaípagos coral reef persistence after ENSO warming across an acidification gradient. Geophys Res. Lett. 41, 9001–9008. doi: 10.1002/2014GL062501
Manzello D., Kleypas J., Budd D., Eakin C., Glynn P., and Langdon C. (2008). Poorly cemented coral reefs of the eastern tropical Pacific: possible insights into reef development in a high-CO2 world. Proc. Natl. Acad. Sci. U.S.A. 105, 10450–10455. doi: 10.1073/pnas.0712167105
Manzello D. and Lirman D. (2003). The photosynthetic resilience of Porites furcata to salinity disturbance. Coral Reefs 22, 537–540. doi: 10.1007/s00338-003-0327-0
Mehrbach C., Culberson C., Hawley J., and Pytkowicz R. (1973). Measurement of the apparent dissociation constants of carbonic acid in seawater at atmospheric pressure. Limnol Oceanogr 18, 897–907. doi: 10.4319/lo.1973.18.6.0897
Mena S., Quesada-Perez F., Sánchez-Noguera C., Salas-Moya C., Alvarado J. J., López-Garro A., et al. (2025). Estructura comunitaria de ecosistemas coralinos en sitos de importancia para la conservación de la biodiversidad marina del Pacífico Norte de Costa Rica. Rev. Biol. Trop. 73, e63715. doi: 10.15517/rev.biol.trop.v73iS1.63715
Méndez-Venegas M., Jiménez C., Bassey-Fallas G., and Cortés J. (2021). Condición del arrecife coralino de Playa Blanca, Punta Gorda, uno de los arrecifes más extensos de la costa Pacífica de Costa Rica. Rev. Biol. Trop. 69, 194–207. doi: 10.15517/rbt
Palmer C., Jimenez C., Bassey G., Ruiz E., Cubero T. V., Diaz M. M. C., et al. (2022). Cold water and harmful algal blooms linked to coral reef collapse in the Eastern Tropical Pacific. PeerJ 10, e14081. doi: 10.7717/peerj.14081
Rixen T., Jiménez C., and Cortés J. (2012). Impact of upwelling events on the sea water carbonate chemistry and dissolved oxygen concentration in the Gulf of Papagayo (Culebra Bay), Costa Rica: Implications for coral reefs. Rev. Biol. Trop. 60, 187–195. doi: 10.15517/rbt.v60i2.20004
Rodríguez-Sáenz K. and Morales-Ramírez A. (2012). Composición y distribución del mesozooplanDICon en una zona de afloramiento costero (Bahía Culebra, Costa Rica) durante La Niña 1999 y el 2000. Rev. Biol. Trop. 60, 143–157. doi: 10.15517/rbt.v60i2.19998
Rogers C., Garrison G., Grober R., Hillis Z.-M., and Franke M. (1994). Coral Reef Monitoring Manual for the Caribbean and Western Atlantic. St. John, U.S. Virgin Islands: National Park Service, Virgin Islands National Park. p. 114.
Rojas J. and Rodríguez O. (2008). Diversidad y abundancia ictiofaunística del río Grande de Térraba, sur de Costa Rica. Rev. Biol. Trop. 56, 1429–1447. doi: 10.15517/rbt.v56i3.5720
Sánchez-Noguera C., Jiménez C., and Cortés J. (2018a). Desarrollo costero y ambientes marino-costeros en Bahía Culebra, Guanacaste, Costa Rica. Rev. Biol. Trop. 66, S309–S327. doi: 10.15517/rbt.v66i1.33301
Sánchez-Noguera C., Stuhldreier I., Cortés J., Jiménez C., Morales A., Wild C., et al. (2018b). Natural ocean acidification at Papagayo upwelling system (north Pacific Costa Rica): Implications for reef development. Biogeosciences 15, 2349–2360. doi: 10.5194/bg-15-2349-2018
Sippo J. Z., Maher D. T., Tait D. R., Holloway C., and Santos I. R. (2016). Are mangroves drivers or buffers of coastal acidification? Insights from alkalinity and dissolved inorganic carbon export estimates across a latitudinal transect. Global Biogeochem Cycles 30, 753–766. doi: 10.1002/2015GB005324
Stuhldreier I., Sánchez-Noguera C., Rixen T., Cortés J., Morales A., and Wild C. (2015). Effects of seasonal upwelling on inorganic and organic matter dynamics in the water column of eastern Pacific coral reefs. PloS One 10, e0142681. doi: 10.1371/journal.pone.0142681
Suzuki A. and Kawahata H. (2004). “Reef water CO2 system and carbon production of coral reefs: Topographic control of system-level performance,” in Global Environmental Change in the Ocean and on Land. Eds. Shiyomi M., Kawahata H., Koizumi H., Tsuda A., and Awaya Y. (TERRAPUB, Tokyo), 229–248. Available online at: http://faculty.petra.ac.id/dwikris/docs/cvitae/docroot/html/www.terrapub.co.jp/e-library/kawahata/pdf/229.pdf (Accessed March 7, 2017).
Tamše S., Ogrinc N., Walter L., Turk D., and Faganeli J. (2014). River Sources of dissolved inorganic carbon in the gulf of trieste (N adriatic): stable carbon isotope evidence. Estuaries Coasts 38, 151–164. doi: 10.1007/s12237-014-9812-7
Ulloa A., Aguilar T., Goicoechea C., and Ramirez R. (2011). Descripción, clasificación y aspectos geológicos de las zonas kársticas de Costa Rica. Rev. Geológica America Cent. 45, 53–74. doi: 10.15517/rgac.v0i45.1910
Ulloa A., Vargas M., Argüello A., Álvarez Y., Hidalgo A., and Deleva S. (2024). “Main karstic Landscapes and Caves of COSTA RICA,” in Landscapes and Landforms of Costa Rica. Ed. Quesada-Román A. (Springer International Publishing, Cham), 133–149. doi: 10.1007/978-3-031-64940-0_7
Umaña-Villalobos G. and Springer M. (2006). Variación ambiental en el río Grande de Térraba y algunos de sus afluentes, Pacífico sur de Costa Rica. Rev. Biol. Trop. 54, 265–272. doi: 10.15517/rbt.v54i1.26850
Vargas C., Contreras P., Pérez C., Sobarzo M., Saldías G., and Salisbury J. (2016). Influences of riverine and upwelling waters on the coastal carbonate system off Central Chile and their ocean acidification implications. J. Geophys Res. Biogeosci 121, 1468–1483. doi: 10.1002/2015JG003213
Vargas-Zamora J. A., Gómez-Ramírez E., and Morales-Ramírez Á. (2021). A fjord-like tropical ecosystem, Pacific coast of Costa Rica : overview of research in Golfo Dulce. Rev. Biol. Trop. 69, 773–796. doi: 10.15517/rbt.v69i3.46406
Waylen P. and Laporte M. (1999). Flooding and the El Niño-Southern Oscillation phenomenon along the Pacific coast of Costa Rica. Hydrol Process 13, 2623–2638. doi: 10.1002/(SICI)1099-1085(199911)13:16<2623::AID-HYP941>3.0.CO;2-H
Keywords: coastal variability, coral reefs, drivers, Papagayo upwelling, Eastern Tropical Pacific
Citation: Sánchez-Noguera C, Lange ID, Cortés J, Jiménez C, Wild C and Rixen T (2025) Environmental conditions and carbonate chemistry variability influencing coral reef composition along the Pacific coast of Costa Rica. Front. Mar. Sci. 12:1606253. doi: 10.3389/fmars.2025.1606253
Received: 04 April 2025; Accepted: 08 September 2025;
Published: 06 October 2025.
Edited by:
Cecilia Chapa-Balcorta, Universidad del Mar, MexicoReviewed by:
Fiorella Prada, Rutgers, The State University of New Jersey, United StatesFrancisco Medellín Maldonado, National Autonomous University of Mexico, Mexico
Copyright © 2025 Sánchez-Noguera, Lange, Cortés, Jiménez, Wild and Rixen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Celeste Sánchez-Noguera, Y2VsZXN0ZS5zYW5jaGV6bm9ndWVyYUB1Y3IuYWMuY3I=
‡ORCID: Celeste Sánchez-Noguera, orcid.org/0000-0002-9541-4788