 Chester J. Sands1,2*
Chester J. Sands1,2* Timothy D. O’Hara3
Timothy D. O’Hara3 Alice Guzzi4,5,6
Alice Guzzi4,5,6 William P. Goodall-Copestake1,7
William P. Goodall-Copestake1,7 Peter Convey1,8,9,10
Peter Convey1,8,9,10 Bhavani E. Narayanaswamy2
Bhavani E. Narayanaswamy2 Rafael Martín-Ledo11Sabine Stöhr12
Rafael Martín-Ledo11Sabine Stöhr12- 1British Antarctic Survey, Natural Environment Research Council, Cambridge, United Kingdom
- 2Scottish Association of Marine Science, Oban, United Kingdom
- 3Museums Victoria, Melbourne, VIC, Australia
- 4Italian National Antarctic Museum (MNA, Section of Genoa), University of Genoa, Genoa, Italy
- 5Department of Earth, Environmental and Life Sciences (DISTAV), University of Genoa, Genoa, Italy
- 6National Biodiversity Future Center (NBFC), Palermo, Italy
- 7Royal Botanic Garden Edinburgh, Edinburgh, United Kingdom
- 8Department of Zoology, University of Johannesburg, Johannesburg, South Africa
- 9Millenium Institute Biodiversity of Antarctic and Sub-Antarctic Ecosystems (BASE), Santiago, Chile
- 10Cape Horn International Center (CHIC), Puerto Williams, Chile
- 11Airea de Zoologia, Facultad de Ciencias, Universidad de Extremadura, Badajoz, Spain
- 12Swedish Museum of Natural History, Stockholm, Sweden
To effectively manage biological assemblages, a sound knowledge of the biodiversity is essential. The Southern Ocean shelf benthic assemblages are typically comprised of species, the names of which are shared across the huge expanse of the region, indicating large population sizes and good connectivity, inferring resilience. This is despite two decades of studies identifying cryptic or unrecognised species in many benthic groups. In this study we examine the common and widespread species Ophioplinthus gelida (Koehler, 1900) and Ophioplinthus martensi (Studer, 1885), both regarded as occurring throughout the Southern Ocean on continental and island shelves. The two species show subtle differences in morphology, despite O. gelida having very plastic character states. Genetic analysis using mitochondrial sequences shows that the two species are part of a radiating clade with ten distinct elements, six of which group as O. gelida and four as O. martensi. Further investigation showed that one of these elements can be attributed to Ophioplinthus carinata (Studer, 1876), not previously identified on the Antarctic shelf, but shown here to be a dominant fraction of the O. “gelida” in our collections. Furthermore, O. “martensi” from its type locality of South Georgia is genetically similar to Ophioplinthus intorta (Lyman, 1878), from Marion Island, but distinct from those collected from the Antarctic shelf, which may then be considered Ophioplinthus inermis (Bell, 1902). A fifth clade of O. martensi collected from waters deeper than 2000 m was not part of this radiation but grouped together with more distantly related Ophioplinthus species. In general, the genus displays a wide range of morphological character states, varying greatly within O. gelida elements, and often shared between species. Several taxa outside of the O. gelida complex may also include unrecognised cryptic species, making reliable field identifications challenging, and greatly increasing the recognised species diversity and regional endemism.
1 Introduction
Species names are, in practical terms, the base unit in conservation biology. This is largely due to conservation management tools such as the IUCN Red List having the requirement that the species included must be formally described, or in the advanced process of being described. This makes sense as items need to be categorised to begin to understand their relevance and their place. However, if the category assigned does not capture the essence of the problem being tackled, it becomes a fallacy that undermines the original purpose: in simple terms, a species name may not be equivalent to a biological species. What makes a biological species remains contentious and species concepts are often adopted by researchers based on where their model organisms best fit (De Queiroz, 2007; Wilkins, 2018). However, when a species name does not represent a biological species, whatever that may be, inaccuracies in ecological estimates, such as species richness and distributions and, ultimately, in conservation status, are likely to be resultant detrimental outcomes. The Convention on Biological Diversity (United Nations, 1992) includes genetic diversity as a base factor of conservation. While the tools exist to measure this diversity, mechanisms to report on it have yet to mature to the stage where they are widely accepted (Hoban et al., 2024).
Notwithstanding the challenges of defining the term, most extant biological species are yet to be described. In the late 20th century, various studies attempted to estimate how many species are present on Earth, with estimates ranging from 3–5 million (May, 1988; Stork, 1993), and with caveats suggesting a far higher figure if those below 1 mm in size are considered. More recently it has been estimated that there may be 8.7 million species (Mora et al., 2011) with only a small proportion of these having been formally described and, specifically, 91% of marine species yet to be described. This presents a huge challenge (Engel et al., 2021), particularly given what is termed the “taxonomic impediment” (Ramsay, 1986; Engel et al., 2021), a shortage, not necessarily of taxonomists (Costello et al., 2013), but in the perceived value of taxonomy coupled with the lack of institutional or political will for funding collections. Given the current imbalance in extinction rate compared to replacement (Turvey and Crees, 2019), it is unlikely we will ever capture contemporary species diversity, with many lost to extinction before they are recognised as existing, let alone threatened. Recent advances in access to and the analyses of DNA sequence data have driven dramatically larger projections of the number of species on Earth (Wiens, 2023), in particular as many of the currently described species appear to be complexes of multiple biological species, a further confounding factor not considered in the 8.7 million species estimate.
Technological advances are contributing to reducing the taxonomic impediment. These include the integration of DNA data that has been transformative through the underpinning of classic taxonomic practice and recent drives to (semi-) automate the generation, collation, analysis and accessibility of all forms of taxonomic data. These are laying the foundations for a vast upscaling of species discovery and monitoring. To take one example, MinION (Oxford Nanopore Technologies) sequencing, introduced a decade ago, has matured to a point where it now provides a highly accessible and affordable sequencing solution that is revolutionising efforts to DNA barcode all species on Earth (referred to as BIOSCAN). This DNA sequencing technology, coupled with high resolution 3D imaging and an AI engine, is being used to rapidly characterise species composition in enormous collections of forest insects (Meier et al., 2024; Vasilita et al., 2024). Such advances hold great promise for tackling the taxonomic impediment through accelerated species discovery, linking to existing voucher specimens, and the production of new vouchers and new species descriptions. However, not all of the world’s biodiversity is as amenable as many insects appear to be to these new automated methods due to both physical constraints and taxonomic complexity (Korshunova et al., 2019; Wattier et al., 2020; Doorenweerd et al., 2024). Many marine benthic species fall into this category, for which further manual groundwork is required to provide robust case studies that, in turn, can be used to inform further advances in taxonomic practice.
Southern Ocean benthic assemblages are recognised to be exceptionally species rich (Clarke and Johnston, 2003). The long isolation of Antarctic shelf benthic fauna from other ocean basins has resulted in highly endemic assemblages, while the ice dynamics of the glacial cycles during the Pliocene and Pleistocene (and earlier eras) are likely to have driven diversification of this fauna as niche availabilities have changed over warming and cooling periods (Clarke and Crame, 1992, 2010; Clarke et al., 2004; Thatje et al., 2005, 2008). The concequences of Pleistocene glacial cycles on diversity have been clearly demonstrated in terrestrial systems, particularly in European and North American fauna and flora (Taberlet et al., 1998; Hewitt, 2000; Paulo et al., 2001; Trewick and Wallis, 2001; Ayoub and Riechert, 2004; Garrick et al., 2004; Schonswetter et al., 2004; Tribsch, 2004; Knowles and Richards, 2005; Gómez and Lunt, 2007; Sommer and Zachos, 2009), and, at the extreme, on Antarctic terrestrial systems (Convey et al., 2008, 2020). The effect of the Pliocene and Pleistocene glacial cycles on the diversity of Southern Ocean benthic assemblages is more difficult to quantify, partially as the elements of the assemblages were first described after the great expeditions of discovery over 100 years ago (Challenger, Belgica, Discovery, Charcot, Scotia etc), while access to fresh material across this huge geographic region is costly and logistically challenging. Where recent samples are available, and where genetic markers have been applied to the collections, the presence of unrecognised cryptic diversity appears to be the norm rather than the exception across multiple groups in these assemblages (Linse et al., 2007; Leese and Held, 2008; Arango et al., 2011; Havermans et al., 2011; Hemery et al., 2012; Dietz et al., 2015; Dömel et al., 2017; Hauquier et al., 2017; González-Wevar et al., 2019, 2022; Jossart et al., 2019; Nirmal et al., 2021; McLaughlin et al., 2023). Rapid divergences leading to high polar speciation rates have been identified in both fish (Rabosky et al., 2018) and brittle stars (O’Hara et al., 2019), probably due to rebound after glaciation-related habitat loss (Convey et al., 2009).
The brittle star genus Ophioplinthus Lyman, 1878 is well represented in Southern Ocean benthic assemblages. First described from material collected off the East Antarctic shelf (Ophioplinthus medusa Lyman, 1878) during the Challenger expedition of 1872–1876, there are currently 36 recognised species (WoRMS, 2025), of which 20 are considered to be exclusive to the Southern Ocean and an additional five are found across the Southern Ocean and lower latitude temperate regions. The remainder are from more northerly regions and not represented in the Southern Ocean. Many of the species were brought into the genus Ophioplinthus from the genera Ophioglypha Lyman, 1860, Ophiurolepis Matsumoto, 1915, Homalophiura Clark, 1915, Theodoria Fell, 1961 and Homophiura Paterson, 1985 (Martynov and Litvinova, 2008). Details of the turbulent taxonomic history of the genus Ophioplinthus and the character states that underly the confusion are eloquently laid out by Martynov and Litvinova (2008) and provide some background to the continuing taxonomic issues discussed in this study. Shortly before Martynov and Litvinova’s (2008) publication, a morphological study demonstrated the poor fit of the generic systematics available at that time (Hunter, 2007). More recently following further availability of molecular and morphological evidence, the genus Ophioplinthus has been placed into the family Ophiopyrgidae Perrier, 1893 (O’Hara et al., 2017, 2018).
Of the 25 species of Ophioplinthus recorded from the Southern Ocean, three stand out due to their abundance: O. martensi (Studer, 1885), O. gelida (Koehler, 1900) and O. brevirima (Mortensen, 1936). Of these O. gelida, originally collected from the Western Antarctic Peninsula continental shelf during the Belgica expedition (1897-1899), is by far the most frequently recorded (based on GBIF search 09.10.2024). The original published description of O. gelida was followed up by subsequent descriptive publications, each highlighting the morphological variation observed in collections of this species (Koehler, 1912, 1922). Koehler’s (1922) descriptions of the Australian Antarctic collection included photographic plates highlighting some of the variation observed and have provided an excellent reference for those subsequently identifying Antarctic ophiuroids, albeit with an underlying assumption that these descriptions are accurate. Mortensen (1936), in his description of O. brevirima, points out that many of the specimens in Koehler’s collections are likely to belong to his newly described species, as they share similarities and both are characterised by a symbiotic relationship with the demosponge Iophon Gray, 1867, a character previously believed to be unique to O. gelida. Indeed, it is likely that many records of O. gelida today are made based on this relationship and may, in fact, represent O. brevirima.
Similarly, Mortensen (1936) discussed specimens of O. martensi from a collection made around South Georgia, the type locality of that species (Studer, 1885). Based on Koehler’s images he synonymised Ophioglypha resistens Koehler, 1911 that was described from Cape Royds in the Ross Sea, as well as Ophiozona inermis Bell, 1902, from the East Antarctic, with O. martensi. Bell’s description of O. inermis is detail-poor and inadequate for comparing with other like species (Bell, 1902), and there is no rationale for this synonymy without examination of the type specimens, something Mortensen was unable to do.
In this study we build on previous work assessing Southern Ocean ophiuroid diversity (Martín-Ledo et al., 2013; Sands et al., 2015, 2021, 2024; Galaska et al., 2017a, 2017b; Jossart et al., 2019; Lau et al., 2021, 2023), in which many cases of hidden diversity were identified and are likely to represent unrecognised, geographically discrete species. Widespread, abundant and well-connected species are typically resilient and of little conservation concern (Weckworth et al., 2013; Lamka and Willoughby, 2024). However, in instances where such species are shown to be broken down into smaller, isolated but formally unrecognised species units, these are likely to be less resilient (Caughley, 1994), and at the same time hidden from conservation management tools such as the IUCN Red List and databases such as GBIF, which, understandably, require a formal species name. As highlighted in a previous study of Amphiura belgicae Koehler, 1901 (Sands et al., 2024), O. gelida and O. martensi have sufficiently complex taxonomic history involving synonymies and poor species descriptions (e.g. Ophiozona inermis as noted above), such that genetic appraisal may find cryptic diversity. Unlike A. belgicae, in which very little morphological variation was found across all specimens examined, O. gelida, in particular, appears to show a large degree of character plasticity (Koehler, 1912, 1922). Due to relatively large egg sizes, both O. martensi and O. gelida are thought to brood their young (Mortensen, 1936), assumed to inhibit gene flow and promote the likelihood of geographically isolated populations. Antarctic species thought to have mobile larval stages, such as the brittle star Ophiuroglypha carinifera (Koehler, 1901) and the Antarctic shelf clade of the snake star Astrotoma agassizii Lyman, 1875, tend to have very little spatial genetic structure across the Antarctic continental shelf (Sands et al., 2015; Jossart et al., 2019). In contrast, the sea star Odontaster validus Koehler, 1906, also a species with a dispersing larval stage, was shown to have genetic diversity coupled with subtle morphological variation, from which new species were described (Janosik and Halanych, 2010; Janosik et al., 2011). Our null hypothesis is that O. gelida is a true species with no clear genetic partitioning, albeit with plastic morphology, while our alternative hypothesis is there is genetic variation on a scale similar to that identified the studies mentioned above, and also in Odontaster validus.
2 Methods
Benthic samples were collected from 502 stations across the Southern Ocean on nine cruises between 2006 and 2017 (Figure 1; Supplementary Table 1) using Agassiz Trawls, mini-Agassiz trawls or Bottom trawls. Material from the trawls was sorted on deck to class, then fine sorted to morphotype in the ships’ laboratories. Sorted specimens were preserved in cold (-20°C) 97% ethanol and kept at -20°C during transit. Preliminary identifications were made at sea, but final determinations were made once material was returned to the United Kingdom with the aid of a Leica M65 stereo microscope. Determinations were primarily undertaken using the literature available, in particular the descriptions of Studer (1876, 1885), Lyman (1878, 1883), Koehler (1901, 1907, 1912, 1922), Mortensen (1925, 1936) and Madsen (1955, 1967). Additional material was gathered from curated collections at the Italian National Antarctic Museum (MNA, Genoa Section) where the biological collection are curated. The specimens were collected from three Antarctic expeditions funded by the Italian National Antarctic Research Program (PNRA), the Alfred Wegener Institute (AWI, Germany) and the National Institute of Water and Atmospheric Research (NIWA, New Zealand). All data collected including metadata, sequences and images are available at dx.doi.org/10.5883/DS-300525.
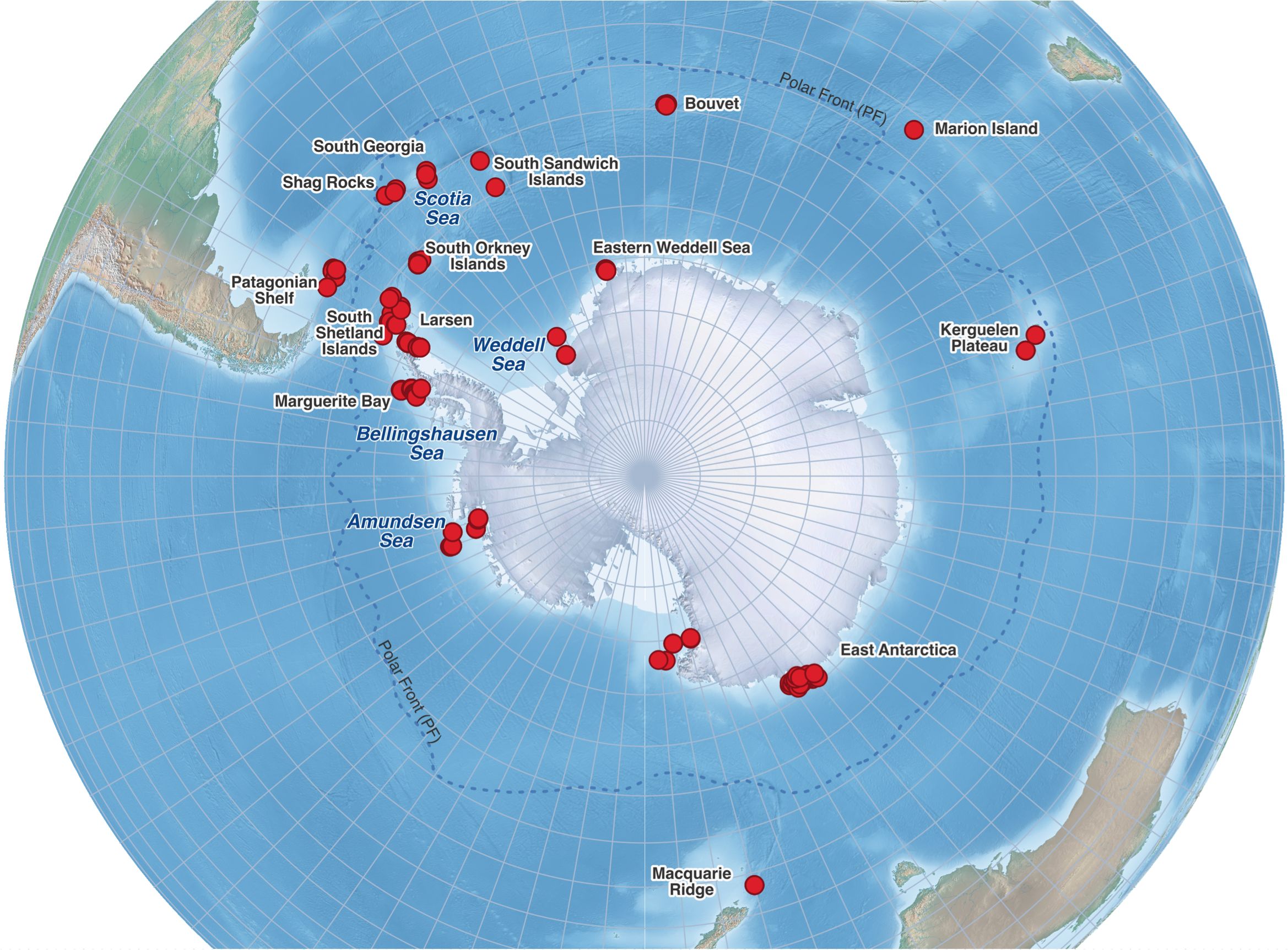
Figure 1. Map of the Southern Ocean showing sampling stations where Ophioplinthus species were collected (red circles).
To further facilitate difficult determinations, an Xper3 (Vignes-Lebbe et al., 2016) database of characters and character states was constructed based on those provided in original descriptions for all nominal Ophioplinthus species. Xper3 is a tool to help model morphological character states with given species hypotheses. In total, 37 descriptions were included with 59 characters (descriptors). Given the subjective language used in the descriptions, the numerous different authors and the translation effort, there were many different states for each character. In order to evaluate whether states such as “pear-seed shaped”, “irregular pear-seed shaped”, “oval with point inwards”, and “rounded triangular” (for example) could be rationalised, efforts were made to examine the type specimens where possible, either physically or via images sent by curators (a list of type specimens examined is provided in Supplementary Table 2).
Tissue was taken from an arm tip of specimens and sent to the University of Guelph DNA sequencing service where partial mitochondrial cytochrome c oxidase subunit I sequences were generated and added to the BOLD v4 database along with associated metadata. As part of the BOLD sequencing and databasing process, specimens with sequences of sufficient quality are allocated a Barcode Index Number (BIN), which is determined by an algorithm based on five established species delimitation methods (Ratnasingham and Hebert, 2013). In summary, if a sequence is sufficiently similar to others in the database and sufficiently distinct from all else, it will share their BIN. If a sequence is sufficiently different from all those in the database, it is allocated a new BIN. BINs can be thought of as operational taxonomic units and, as such, could be considered as equivalent to “species”.
The trace files from bidirectional sequences were aligned and checked by eye to form contigs in CodonCode Aligner v9.0.2. Consensus sequences of the contigs were imported to Geneious Prime 2024.0.4. Alignments were conducted in Geneious Prime using MAFFT v7.49 (Katoh and Standley, 2013). Alignments were translated and checked for open reading frame.
To provide a general systematic appreciation of the genus Ophioplinthus, and the relative placement of members currently assigned to O. gelida and O. martensi, all sequenced individuals in our collections of this genus were included, along with a selection of GenBank submissions and outgroups. We acknowledge the limitations of using a single mitochondrial gene, particularly in resolving deeper phylogenetic relationships and that future studies incorporating genomic approaches would enhance phylogenetic resolution and robustness.
Maximum likelihood reconstruction of phylogenetic relationships was conducted in RAxML v8 (Stamatakis, 2014) using a GTR+G model as indicated by JModeltest v2 (Darriba et al., 2012). Bayesian phylogenetic reconstruction was conducted in BEAST 2 v2.7.7 (Bouckaert et al., 2014) using a GTR+G model, four gamma categories and a Yule prior. After optimising prior parameters a long run of 2 x 108 generations was conducted. Tracer v1.7.1 was used to check for mixing and convergence of the parameters. Phylogenetic trees were visualised in FigTree before being further edited in Graphic V3.1.
Summary statistics were calculated using DNAsp v5 (Librado and Rozas, 2009). Within and between group distances (uncorrected P and composite likelihood) were estimated in Mega v10 (Kumar et al., 2018).
In the clades where sample size and geographic spread were sufficient, networks were produced using PopArt (http://popart.otago.ac.nz), using the TCS statistical parsimony method (Clement et al., 2000).
To explore population change over time each clade with sufficient sample size was run through Bayesian Skyline analysis in BEAST 2.7.7. In each case an HKY model was used, a strict clock with a rate of 2.48 × 10–8 applied and run for 1 × 107 generations. This rate, originally estimated from urchin divergence across the isthmus of Panama (Lessios et al., 2001) and previously used for brittle stars (Naughton et al., 2014; Sands et al., 2015, 2024) is used for consistency across our studies. Tracer v1.7.1 was used to check mixing and convergence, and then for the final plot analyses.
Divergence times were estimated using BEAST 2.7.7 using a reduced Ophioplinthus dataset and including the reduced datasets of the Ophiuroglypha lymani Ljungman, 1871 complex with Ophiopyrgidae outgroups (Sands et al., 2015). A GTR+G model was used with a strict clock and rate of 2.48 × 10-8. The analysis was run for 5.4 × 107 generations to ensure sufficient mixing and convergence of parameter estimates. Divergence times of most probable estimates of each of the complexes were visualised using Tracer v1.7.1.
3 Results
3.1 Systematic context
A total of 683 sequences (including outgroups) targeting the 658 bp region of CO1 were used to reconstruct a phylogeny to evaluate the generic systematics of Ophioplinthus, specifically the relationships between O. gelida and O. martensi. Figure 2 presents a simplified version with unsupported branches collapsed. A full unedited phylogeny with all tips labelled with BOLD accession numbers, as well as sequences included from GenBank, is given in Supplementary Figure 1.
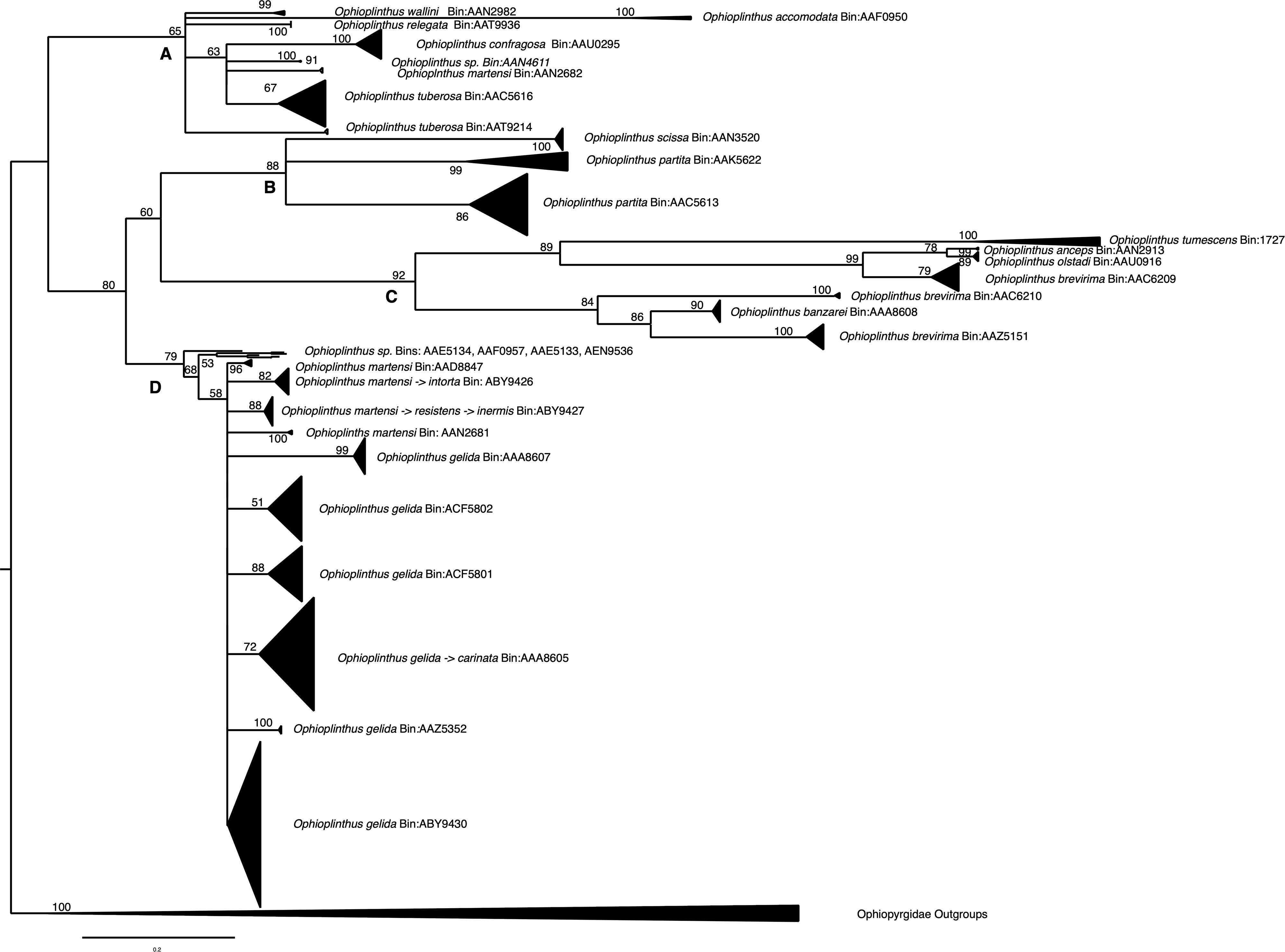
Figure 2. Maximum likelihood phylogeny of 683 Ophioplinthus cytochrome c oxidase subunit 1 partial sequences. Node values are based on bootstrap sampling. Where branch support was below 50% the branches were collapsed. Triangles are collapsed clades where height (vertical side to the right) is proportional to sample size and depth (to the angle joining the phylogeny) is proportional to within clade diversity. The phylogeny is broken into four sub-trees: (A) Temperate and tropical deep-sea species, sub-polar species; (B) “True” Ophioplinthus where included species are morphologically similar to the type species O. medusa; (C) Heavily armoured polar species; (D) Ophioplinthus gelida/martensi polytomy.
The cytochrome c oxidase subunit 1 is a fast-evolving gene and not ideal for phylogenies beyond generic depth, however, it still provides interesting and valuable information that can help inform future systematic and population genetic studies. In this case we recognise four aspects, three of which we touch on briefly here, while the fourth is the primary focus of this study.
First, it appears that given the limits of the samples we had available (Sands Collection = 546, O’Hara Collection = 72, Guzzi (MNA) Collection = 26, GenBank = 45), the genus can be divided into four groups (see Figure 2). These are: A. temperate and tropical deep-sea species [e.g. Ophioplinthus accommodata (Koehler, 1922)], sub-polar [O. confragosa (Lyman, 1878), O. tuberosa (Mortensen, 1936)] and the synonymised genus Theodoria Fell, 1961 [O. relegata (Koehler, 1922), O. wallini (Mortensen, 1925)]; B. “True” Ophioplinthus which includes O. scissa (Koehler, 1908), O. partita (Koehler, 1908), and due to similarity in morphology the type species O. medusa which we were unable to obtain for sequencing; C. the heavily armoured polar species [O. anceps (Koehler, 1908), O. banzarei (Madsen, 1967), O. brevirima (Mortensen, 1936), O. olstadi (Madsen, 1955) and O. tumescens (Koehler, 1922)] and D. a radiating clade of O. gelida and O. martensi.
Second, some currently assigned species are associated with more than one clade/BIN while, in many cases, there are deep divergences between these clades (O. partita BINs: AAK5622 and AAC5613, O. brevirima BINs: AAC6209, AAC6210 and AAZ5151). In the case of O. brevirima some clades/BINs are more closely associated with other discrete species than they are with other clades of O. brevirima, for example, O. brevirima BIN: AAC6209 is more closely related to O. anceps and O. olstadi than it is to the other two clades of O. brevirima.
Third, referring to Supplementary Figure 1 where all sequences are present in an unedited version of the phylogeny, the distribution of the GenBank sequences highlights the difficulties in correctly identifying Ophioplinthus species, particularly distinguishing between O. brevirima and O. gelida. As we note below, this is not a criticism of the identifiers as much as of the inadequate original descriptions and, in some cases, large variation in character states within clades/BINs.
Fourth, this part of our analyses indicates that O. gelida and O. martensi are part of a polytomy or radiation of 10 discrete clades/BINs. This observation provides the main focus of the remainder of the study.
The collection locations of O. gelida and O. martensi are presented in Figure 1, and those of each of the clades of O. gelida and O. martensi are presented in Figures 3 and 4. From these figures the unusually intensive sampling effort available for these taxa can be appreciated, which counterpoints the very limited distributions of some of the clades identified.
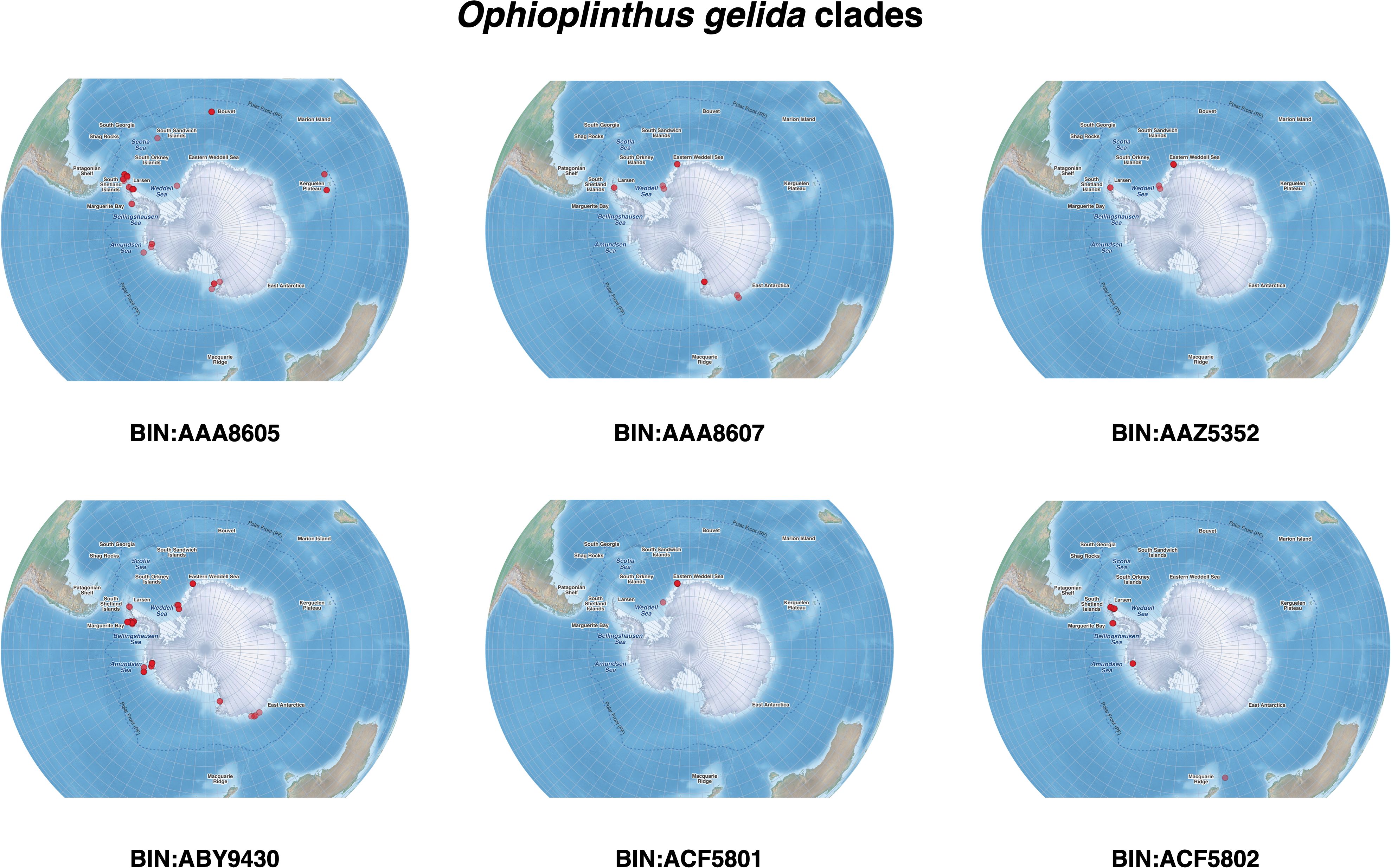
Figure 3. Distribution of Ophioplinthus gelida clades from our sampling efforts.
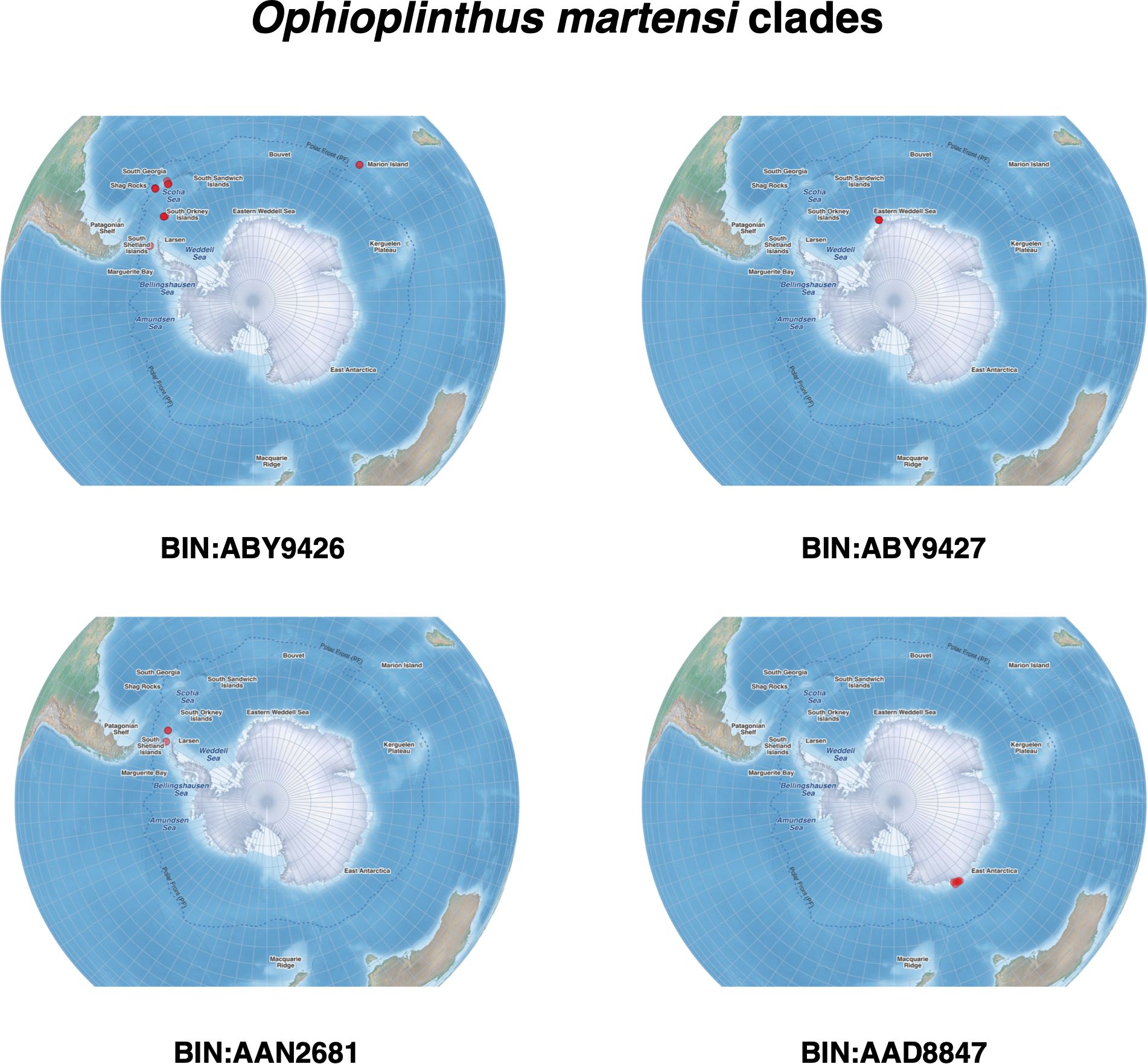
Figure 4. Distribution of Ophioplinthus martensi clades from our sampling efforts.
The summary statistics presented in Table 1 were used to better understand the dynamics of the sequence divergence between clades/BINs. The ratio between CO1 haplotype diversity and nucleotide diversity is a good indicator of potential crypsis in animals (Goodall-Copestake et al., 2012) and, for both O. gelida and O. martensi, the high nucleotide diversity indicates a level of variation beyond that expected within a species. Significant Tajima’s D (DT) and Fu’s S (FS) statistics indicate non-neutral variation in many of the clades that had sufficient sample size to estimate these statistics. As these statistics are significantly negative they indicate either purifying selection, population expansion or both. Fu’s S is particularly sensitive to population expansion and this, along with significant R2 values, strongly suggests a shared historical demographic of population growth within the O. gelida clades.
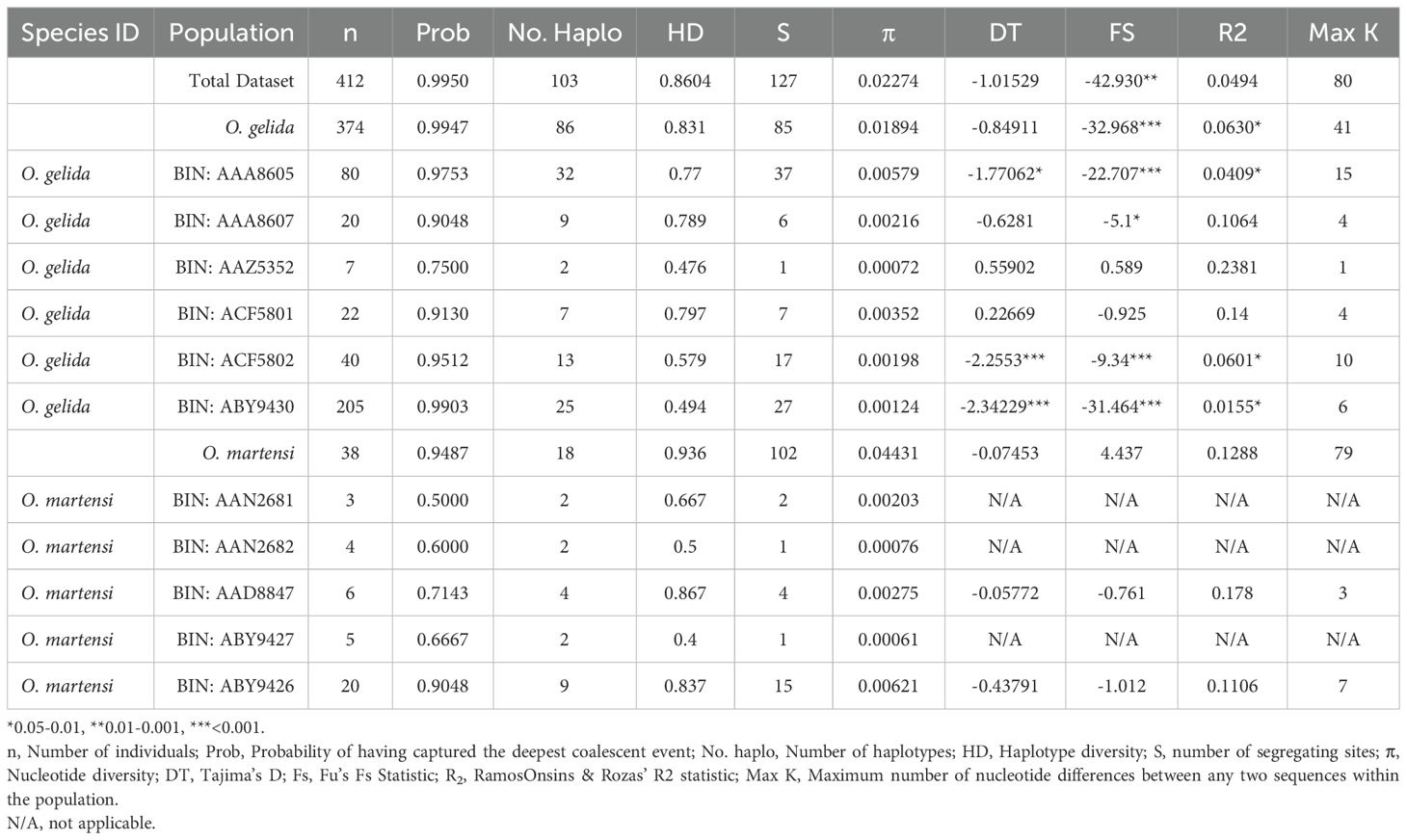
Table 1. Summary statistics of molecular diversity in each of the clades of the Ophioplinthus gelida/martensi radiation where sample size (n) of cytochrome c oxidase subunit 1 sequences was sufficient.
Within and between group distances provided in Table 2 give an indication of the relative variation and divergence of the elements in the polytomy. Uncorrected percentage difference (below the diagonal) and composite likelihood distance (above the diagonal) show typical distances between 3 and 6% between clades, while within-clade variation varied between 0.0007% (BIN: AAZ5352) and 0.005% (BIN: AAA8605).
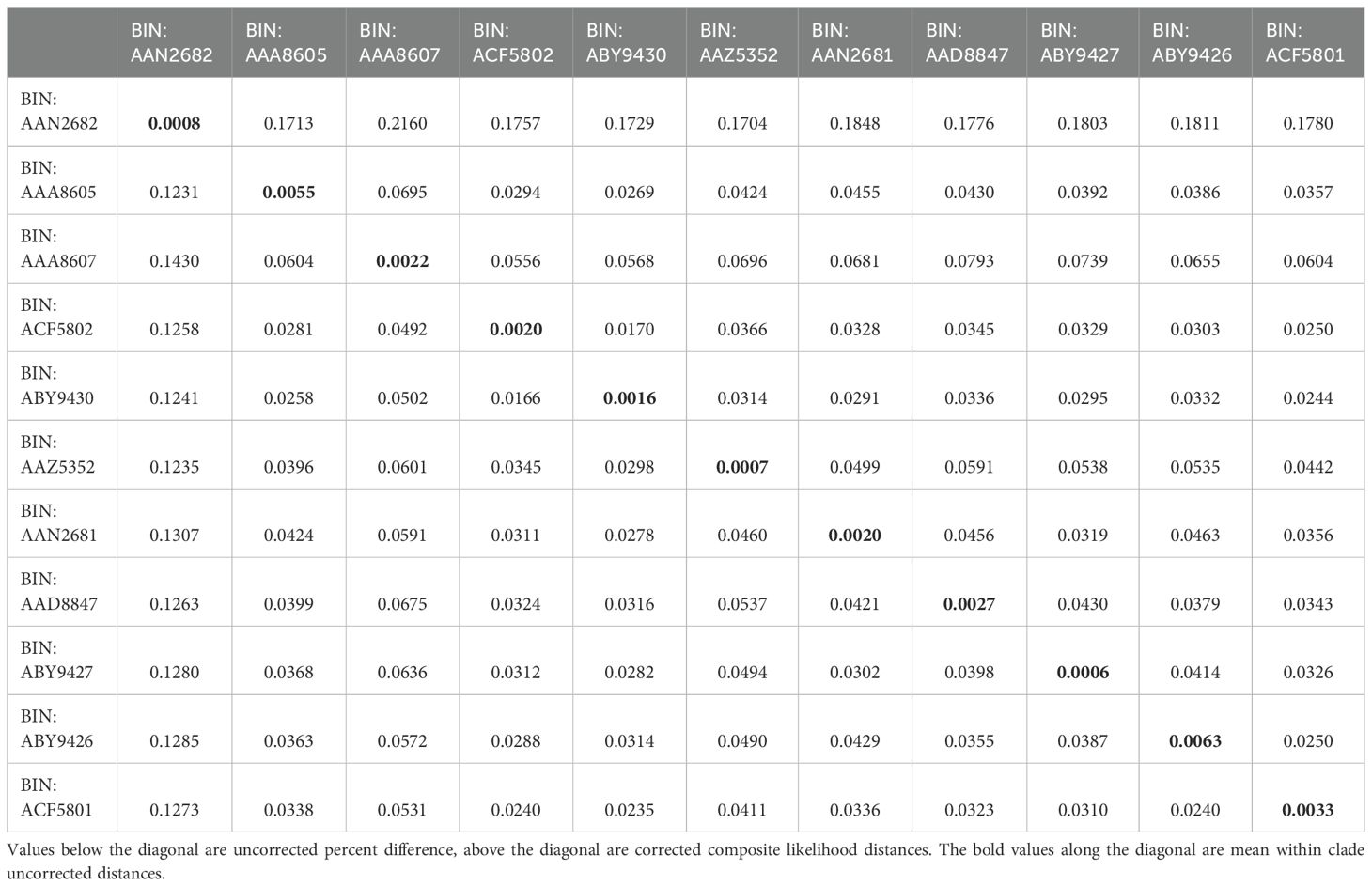
Table 2. Pairwise distances between BINs of the Ophioplinthus gelida/martensi radiation.
3.2 Taxonomic issues
The 10 divergent lineages of O. gelida and O. martensi required taxonomic investigation. The clades may represent intraspecific diversity within the two species, indicating the presence of unidentified cryptic species, or represent described species we failed to identify. Overall, the variation between descriptions and types, and between some syntypes, resulted in a poorly fitting morphological model.
Despite the morphological model’s ineffectiveness, there were two points that stood out after careful evaluation of the characters and re-evaluation of our O. gelida and O. martensi samples and their distributions: First, five of the six clades of O. gelida are restricted to the Antarctic continental shelf and have overlapping ranges. The exception is clade BIN: AA8605, also with a range around the Antarctic continental shelf, but that is represented on the island shelves of the South Sandwich Islands, Bouvet and the Kerguelen Plateau. The species described from Kerguelen are O. carinata (Studer, 1876) and Ophioglypha deshayesi Lyman, 1878, the latter currently considered conspecific with O. carinata. A diagnostic character of O. carinata (given by Lyman, 1878) is an intercalary plate between the adoral shields and the oral plates, usually diamond-shape (circled in Figure 5 image B). This is a character seen in many of our Antarctic shelf O. gelida, and Koehler (1922) describes this character as part of the wide range of variation he noted in the species. Indeed, this character is inconsistently present in all the clades of O. gelida (see Figures 5–9, images B and E). Nevertheless, as the clade is present around Kerguelen, and the only species of Ophioplinthus described from this region is O. carinata, we propose that clade BIN: AAA8605 should be considered O. carinata rather than O. gelida. A single example of clade BIN: AAZ5352 was able to be photographed (Figure 10) and we were unable to judge the morphological variability within this clade. We have considered this part of the O. gelida variation, however, our original taxonomic notes suggest we hesitated with its taxonomic placement due to its similarity with O. martensi. A character matrix to highlight the ineffectiveness of primary morphological characters is provided in Table 3.
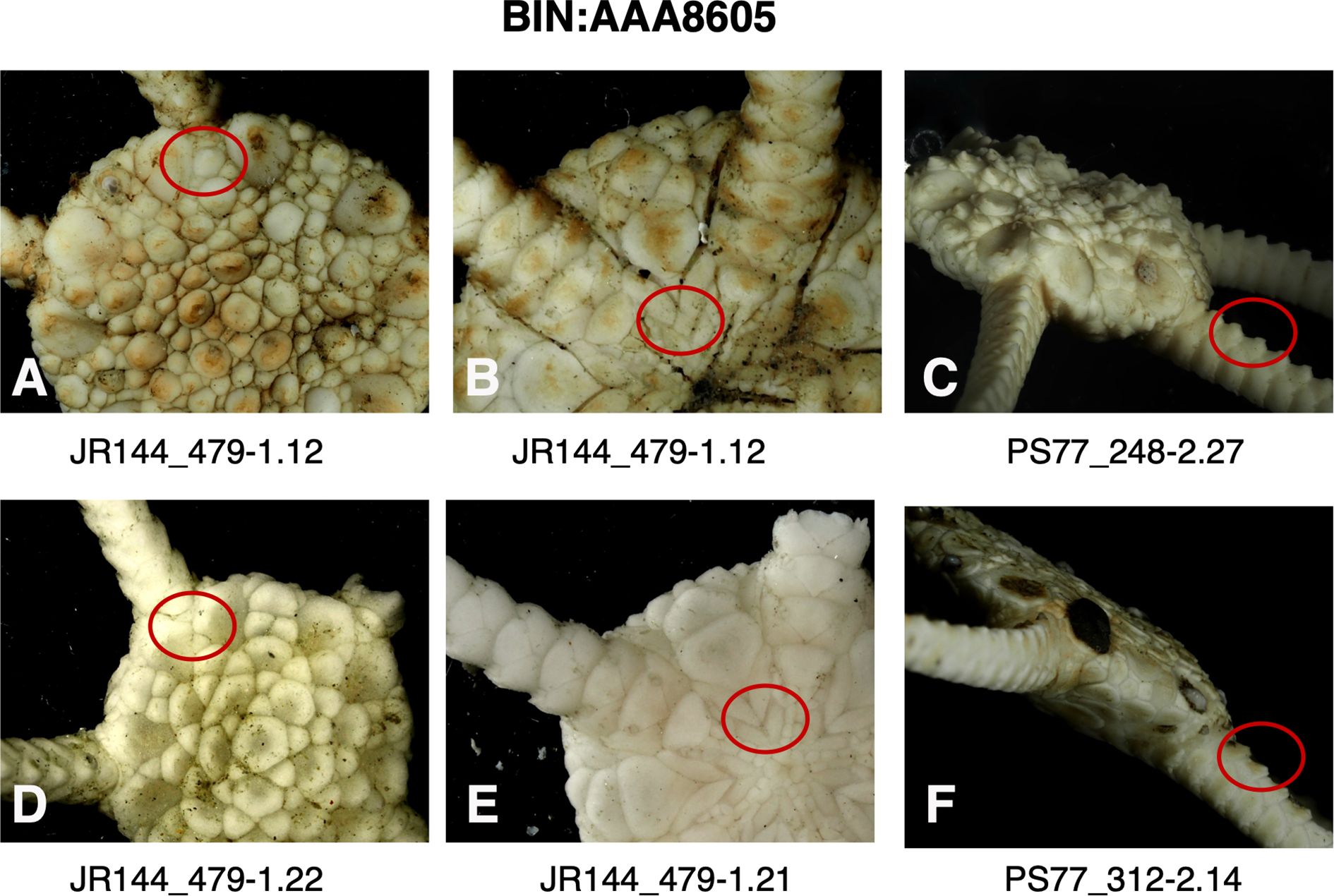
Figure 5. Images of specimens of BIN: AAA8605 (Ophioplinthus gelida->carinata) highlighting (red circle) presence (A) or absence (D) of plates separating radial shields, presence (B) or absence (E) of intercalary plate between adoral shields and oral plates, the relative heights of dorsal arm plate projections (C, F).
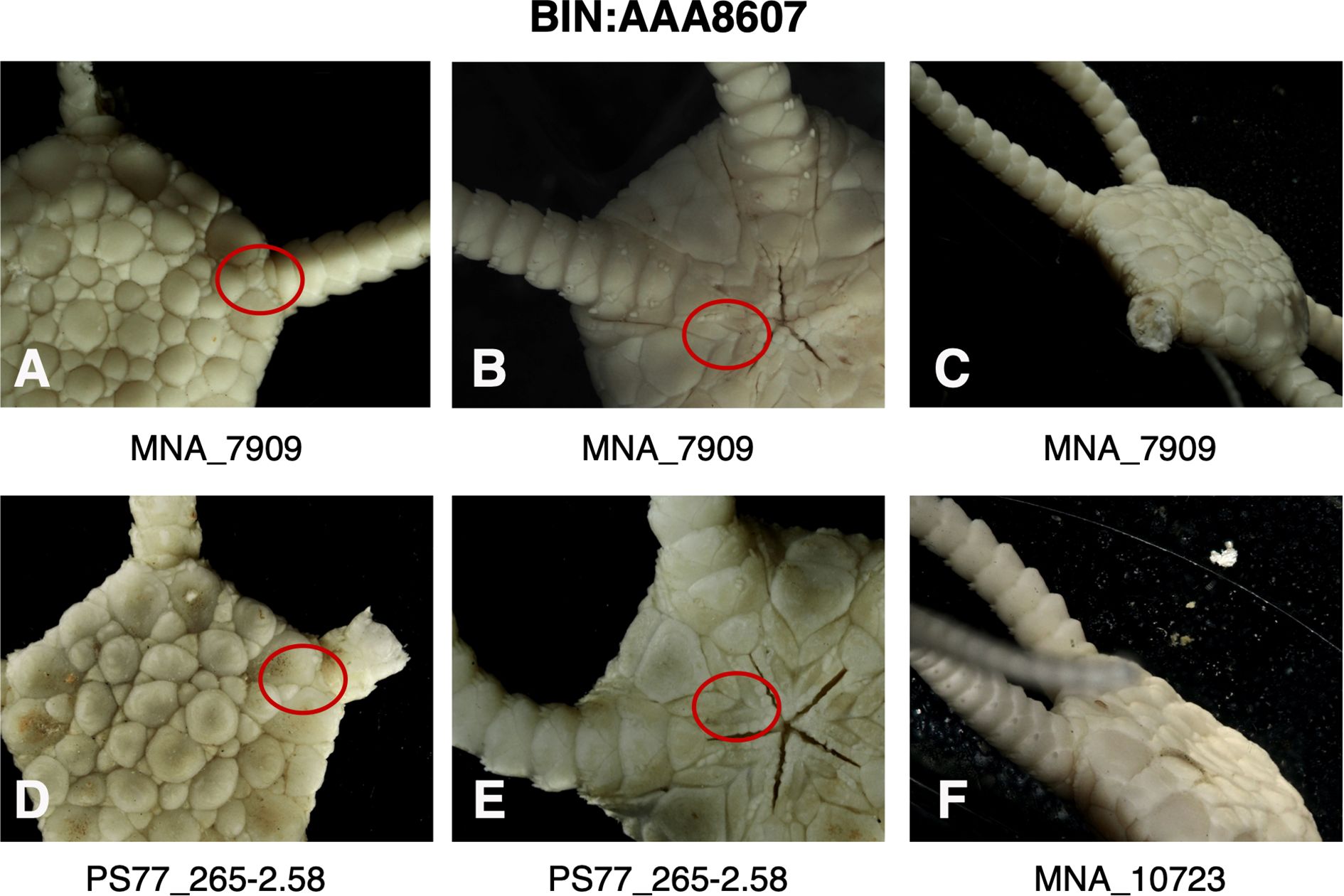
Figure 6. Images of specimens of BIN: AAA8607 (Ophioplinthus gelida) highlighting (red circle) presence (A) or absence (D) of plates separating radial shields, presence (B) or absence (E) of intercalary plate between adoral shields and oral plates, the heights of dorsal arm plate projections (C, F).
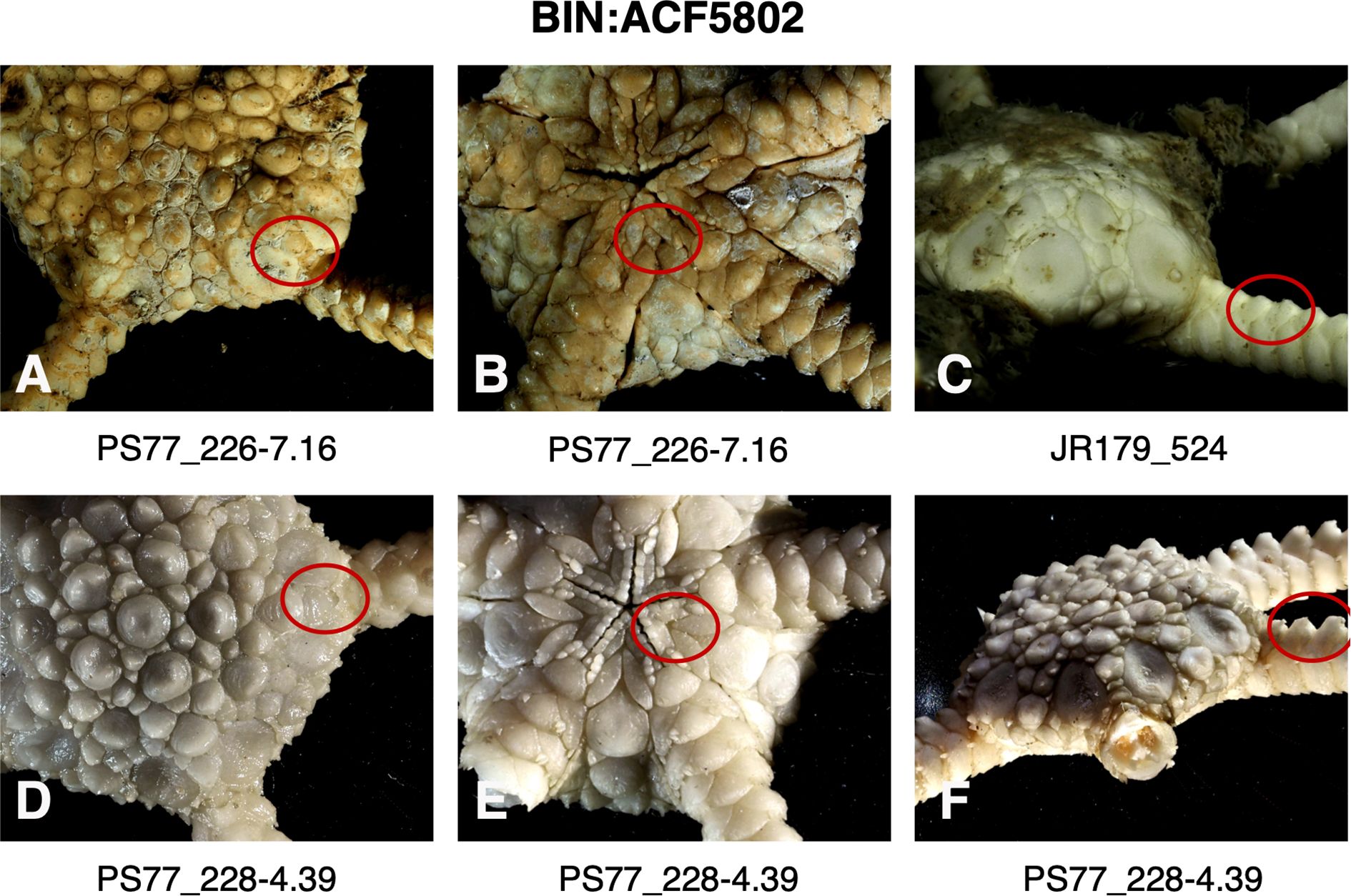
Figure 7. Images of specimens of BIN: ACF5802 (Ophioplinthus gelida) highlighting (red circle) variation of plates separating radial shields (A, D), presence (B) or absence (E) of intercalary plate between adoral shields and oral plates, the relative heights of dorsal arm plate projections (C, F).
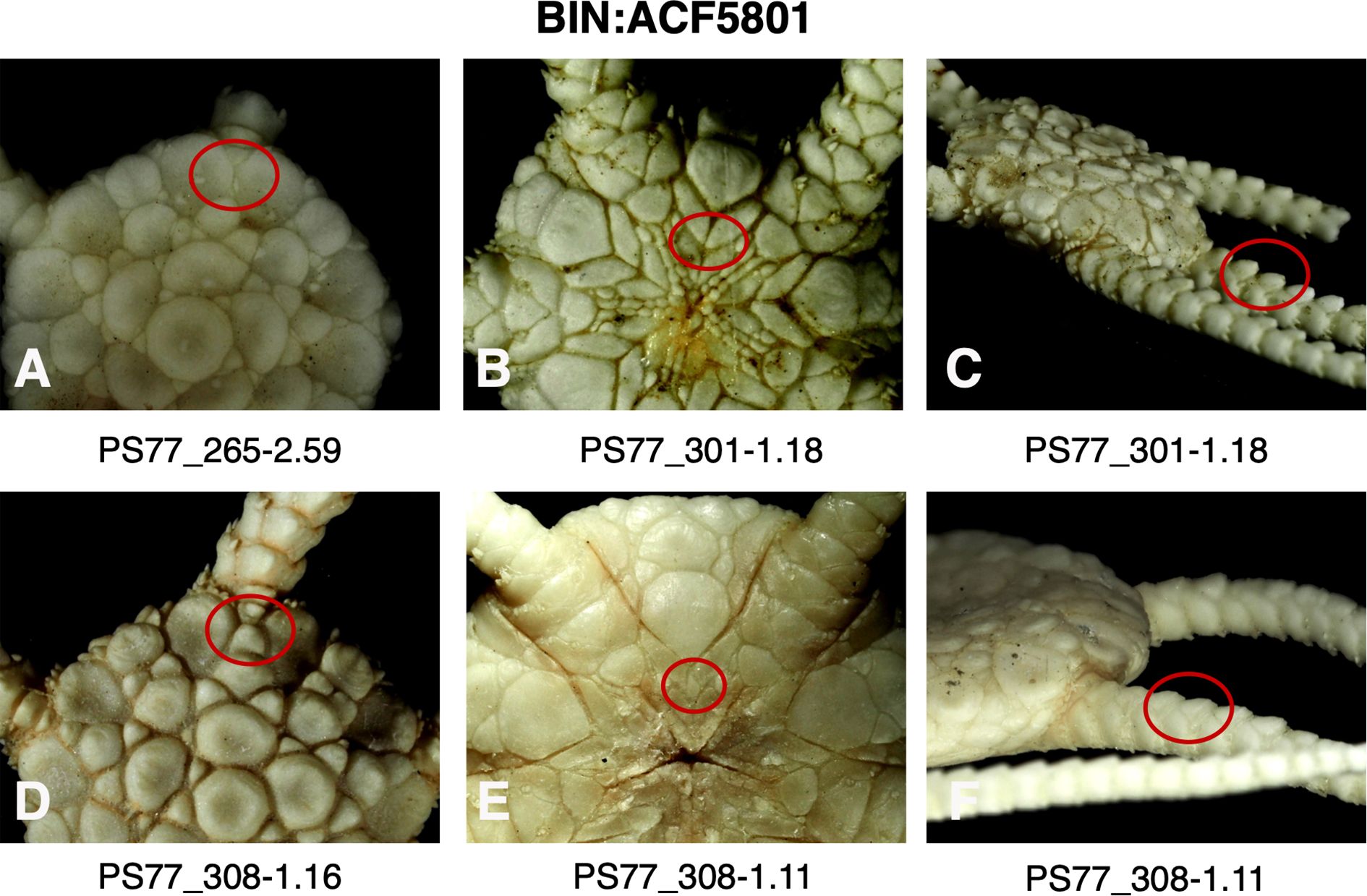
Figure 8. Images of specimens of BIN: ACF5801 (Ophioplinthus gelida) highlighting (red circles) absence (A) and presence (C) of plates separating radial shields, the absence (B) or presence (D) of intercalary plate between oral shields and oral plates, and the relative heights of dorsal arm plate projections (C, F).
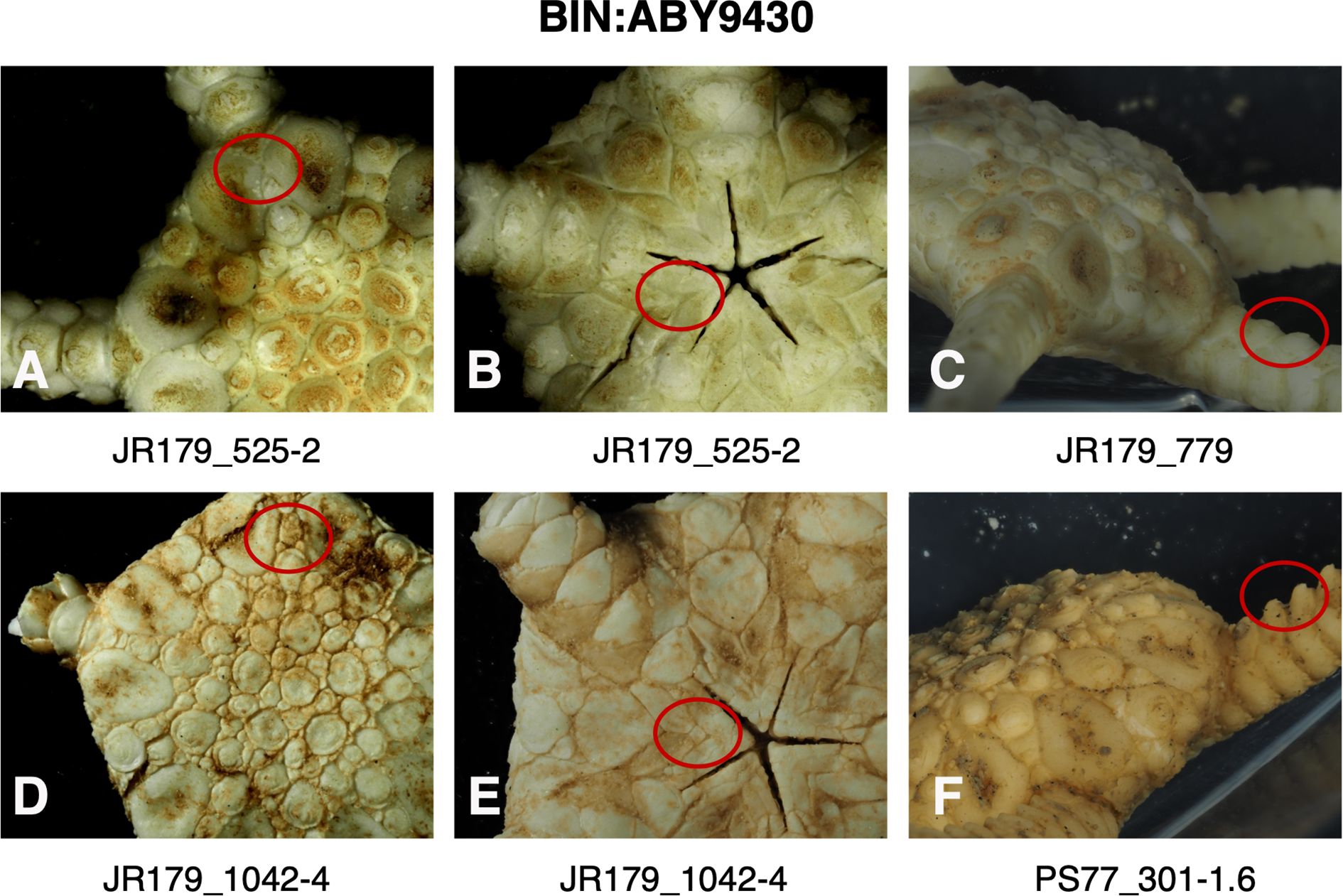
Figure 9. Images of specimens of BIN: ABY9430 (Ophioplinthus gelida) highlighting (red circle) presence (A) or absence (D) of plates separating radial shields, the absence (B) or presence (D) of intercalary plate between oral shields and oral plates, the relative heights of dorsal arm plate projections (C, F).
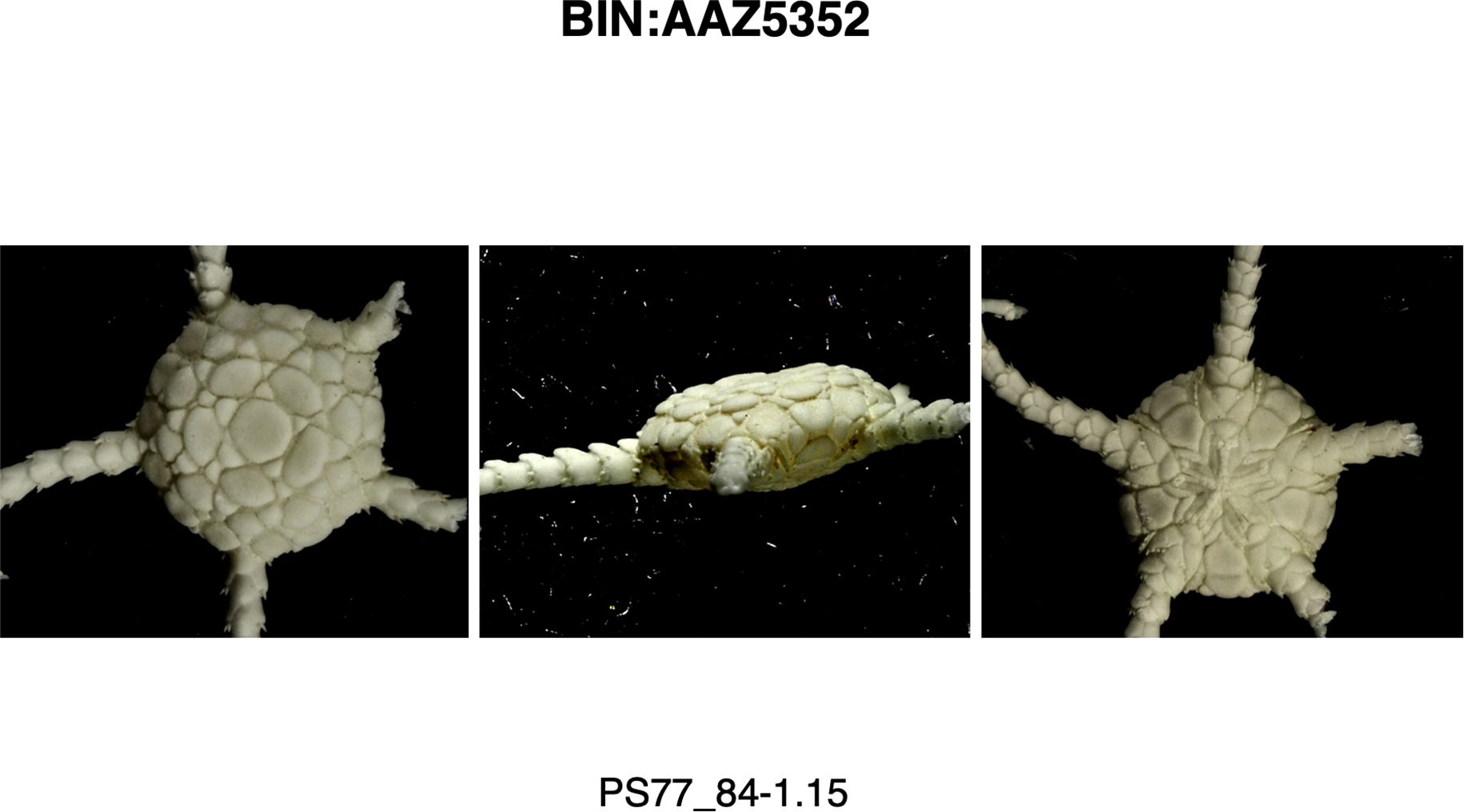
Figure 10. Images of a specimen from BIN: AAZ5352 (Ophioplinthus gelida).
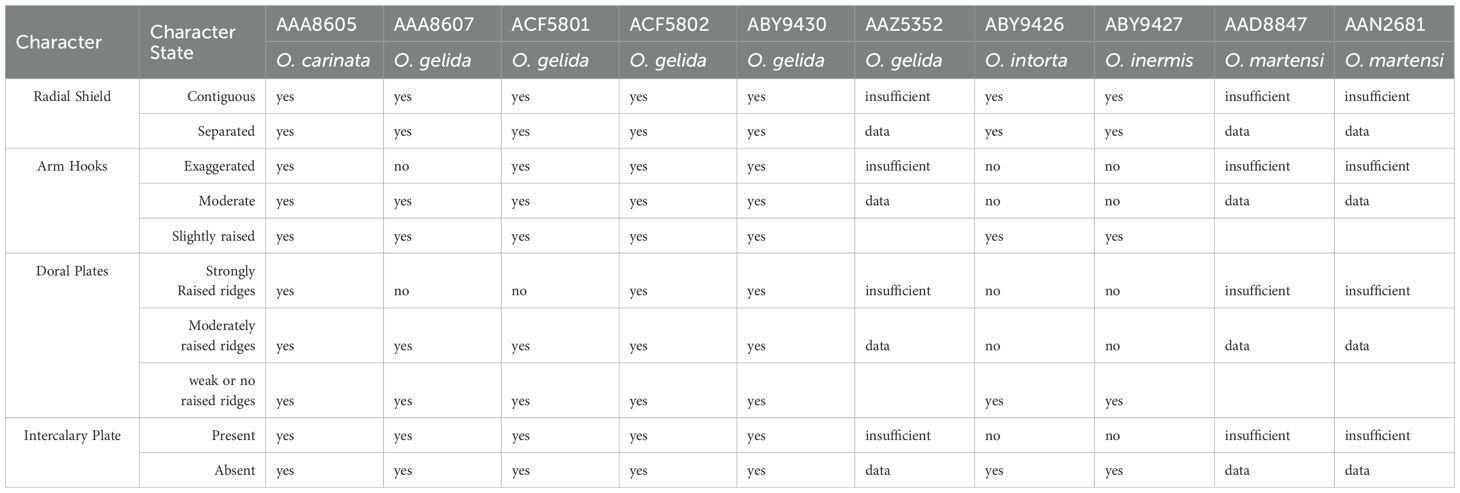
Table 3. A character matrix of exemplar characters and their corresponding character states in the clades of the Ophioplinthus gelida/martensi complex.
Second, O. martensi was described by Studer (1885) from specimens collected around South Georgia. Our South Georgia specimens all fall into clade BIN: ABY9426 (examples in Figure 11). However, specimens from Marion Island had very similar sequences and fall within the same BIN. The Ophioplinthus species identified from around Marion Island is O. intorta (Lyman, 1878). Mortensen (1936) examined Marion Island specimens of Ophioplinthus and referred them to O. martensi. Examination of the type material at the Natural History Museum, London (NHM) and images of the type material of O. martensi from the Museum der Natur Hamburg, along with our own specimens, confirmed that O. intorta is morphologically similar to O. martensi type material but differed subtly from O.”martensi” samples from the Antarctic continental shelf, in that the Antarctic specimens were generally more robust, particularly the arms. This was a very tactile character difference, and not one clearly visible in images (see Figures 11 and 12). We therefore propose that clade BIN: ABY9426 should be considered O. intorta rather than O. martensi.
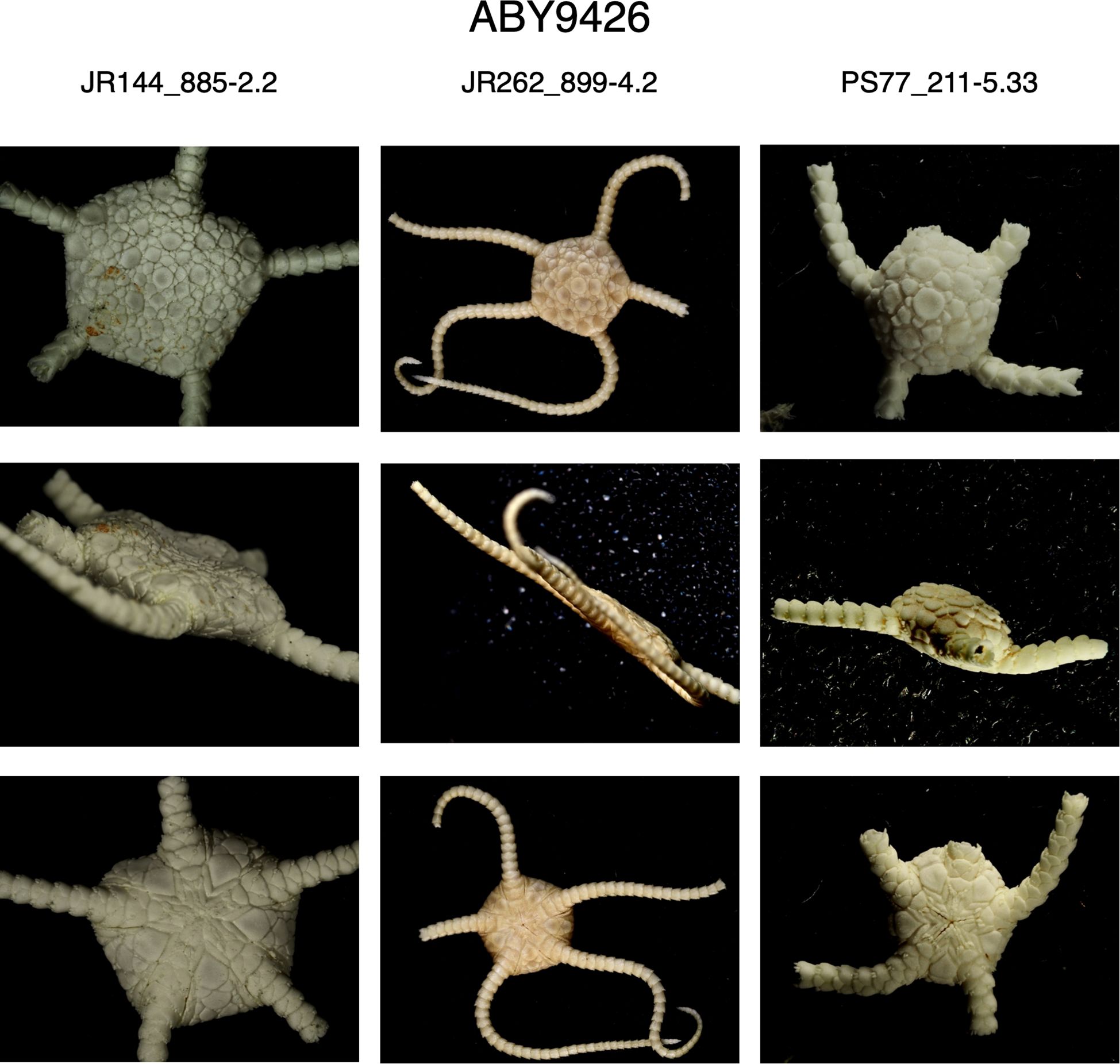
Figure 11. Images of three specimens of BIN: ABY9426 (Ophioplinthus martensi->intorta) from South Georgia shelf region showing aboral (top row), lateral (middle row) and oral perspectives.
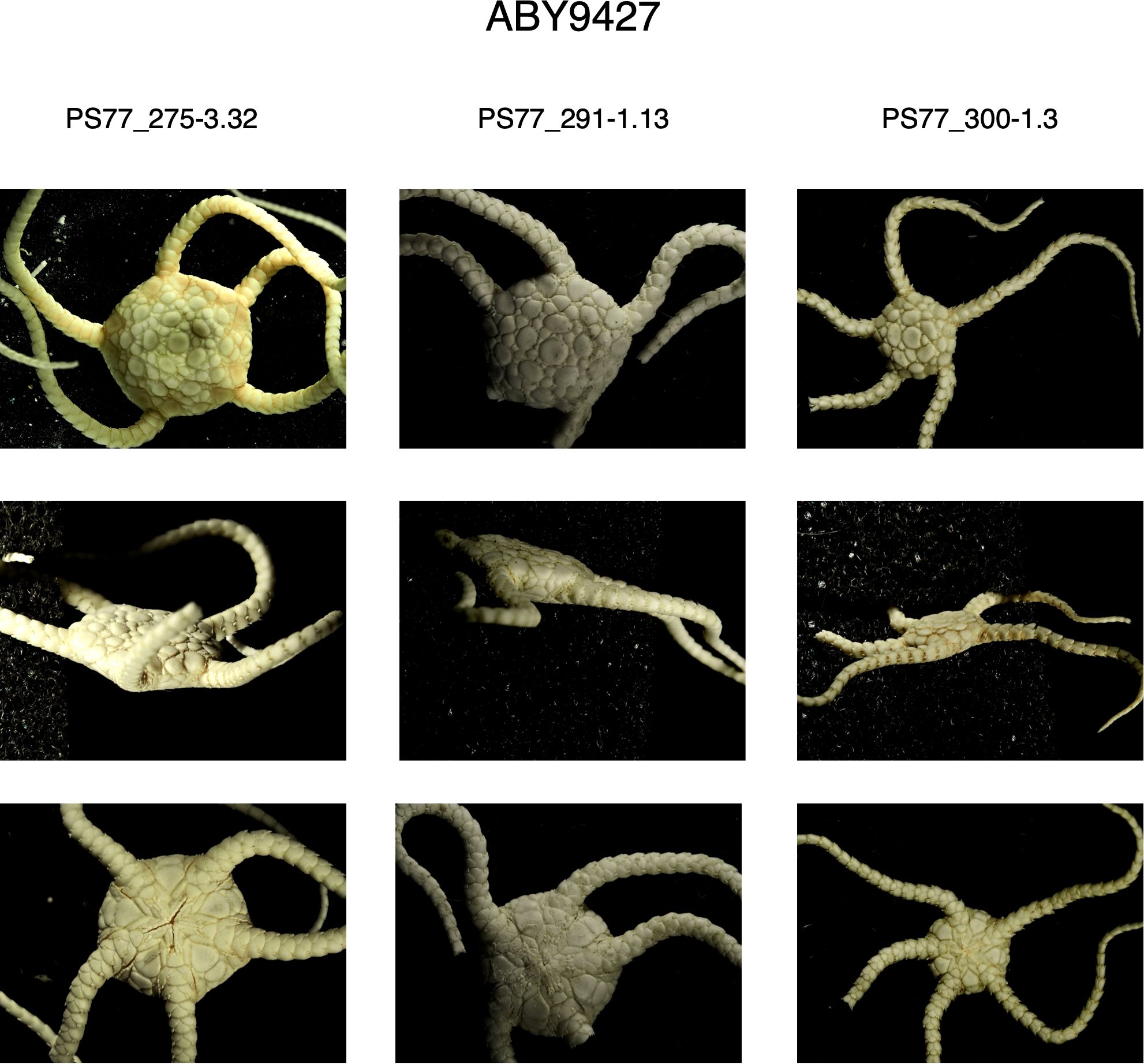
Figure 12. Images of three specimens of BIN: ABY9426 (Ophioplinthus martensi->inermis) from the Weddel Sea, Antarctica, showing aboral (top row), lateral (middle row) and oral perspectives).
Ophioplinthus martensi from the Antarctic Shelf was divided into five separate clades/BINs, each with very limited range within the limit of our sampling. One of these clades (BIN: AAN2682) clustered with the old Theodoria group of species, further highlighting the issues with characters and character states shared between distantly related clades. The four individuals of this clade were collected at depths greater than 2000 m in deep waters off the South Sandwich Islands, while individuals from the other clades were collected at shelf depths (200-800m). Of the other four clades (BIN: AAD8847, BIN: ABY9426, BIN: ABY9427 and BIN: AAN2681), associating them with O. martensi seems illogical as O. martensi was not described from Antarctic waters. Mortensen (1936) synonymised Ophioglypha resistens Koehler, 1911 and Ophiozona inermis Bell, 1902, both collected from the Antarctic continental shelf, with O. martensi, so perhaps the Antarctic clades could represent these species. Samples from BIN: AAD8847 in particular were collected close to both the type localities of O. resistens and O. inermis. Bell’s description of O. inermis is very limited in detail, so the type material of O. inermis at the NHM was examined and compared to images of the type material of O. resistens from Muséum National d’Histoire Naturelle, Paris (MNHN), the type material of O. intorta (also held at the NHM), and our own samples from South Georgia, Marion Island and the Antarctic Shelf. These comparisons show subtle morphological differences in the relative brittleness of the disc and arms, where the Antarctic specimens are more robust, particularly the arms, compared to the sub-Antarctic O. intorta which has more delicate arms, a tactile character better appreciated when handled directly. When Mortensen (1936) synonymised these he only directly assessed specimens from South Georgia, Marion Island and the Patagonian shelf, but no Antarctic specimens, so he was unable to appreciate the variation present, only the similarities given the descriptions. No clear morphological differences between O. inermis, O. resistens and the specimens from our Antarctic clades could be identified.
3.3 Historical demographics
Bayesian skyline plots indicate that, in general, there has been population growth over the past 20,000 years (Figure 13), supporting the inference drawn from the summary statistics, with the most populous clade (O. gelida BIN: ABY9430) showing distinct recent growth over the past 10,000 years. Both O. gelida BIN: ABY9430 and O. carinata BIN: AAA8605 had estimated population sizes an order of magnitude greater than the other clades assessed. This is consistent with our relative sample sizes of the clades. Population networks of these two clades indicate little population structure around the Antarctic shelf, but some structure was evident in the sub-Antarctic island populations of O. carinata (Supplementary Figure 2).
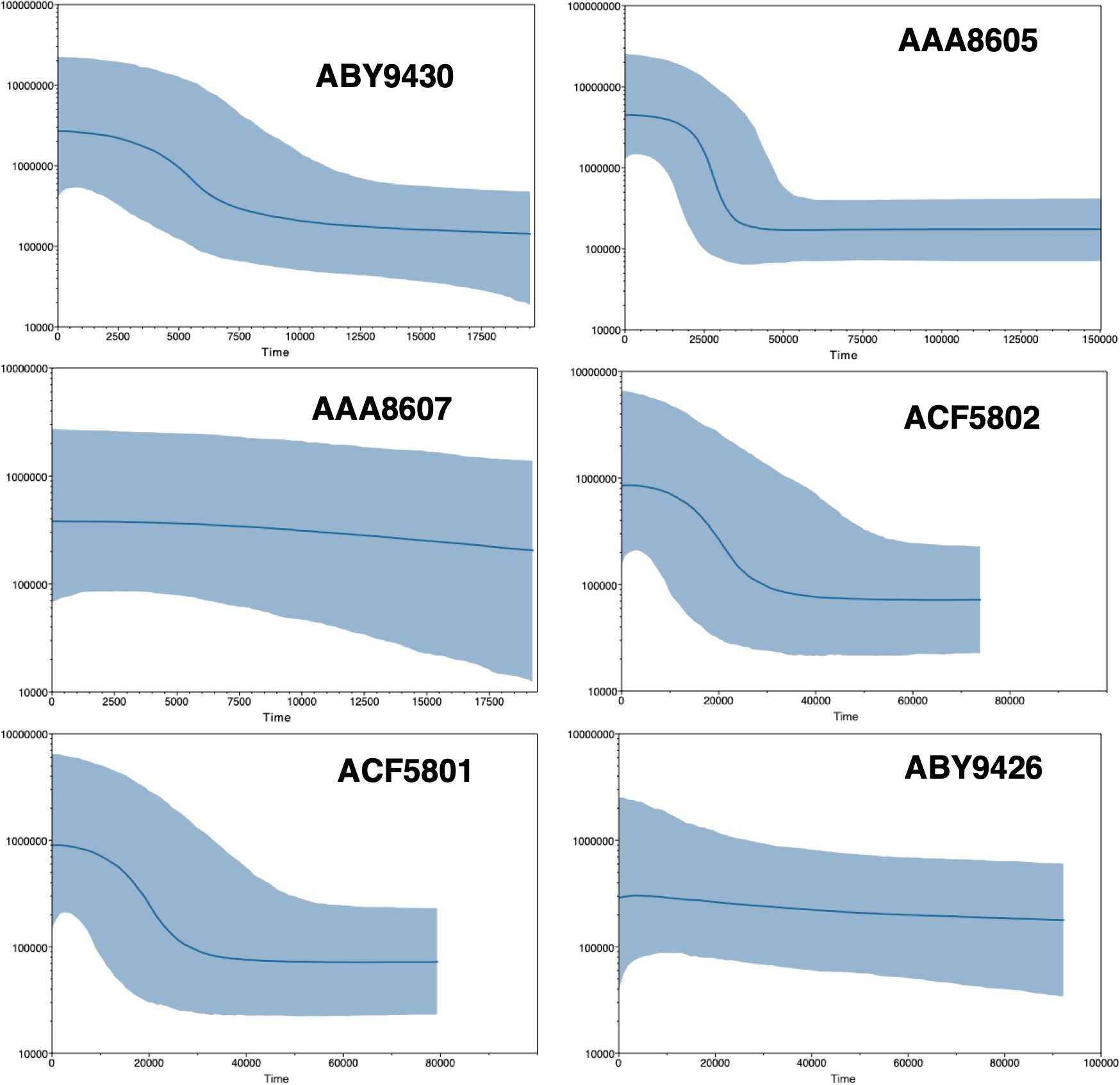
Figure 13. Bayesian skyline plots of sufficiently sampled clades of the Ophioplinthus gelida complex. The X axis is time in years; the Y axis is samples size. The blue lines are most probable estimates and lighter blue shading the 95% highest probable density.
Most probable divergence time estimates of the Ophioplinthus gelida and Ophiuroglypha lymani complexes were very closely aligned (mean O. gelida = 0.422 my, SE 2.5 × 10-3, mean O. lymani = 0.46 my, SE 2.69 × 10-3). The Amphiura belgicae complex has a division that is older (mean = 7.3 my, SE 4.27 × 10-3) but, when the products of this division, the A. belgicae complex from the Antarctic shelf (mean = 0.46 my, SE 2.73 × 10-3) and A. belgicae/A. eugeniae from the Patagonian shelf (mean = 0.44 my, SE 2.62 × 10-3), are taken into account, again the divergence times do not substantially differ from the O. gelida or O. lymani complexes.
4 Discussion
Our study has identified that there has been a recent radiation of Ophioplinthus species into a clade of 10 divergence lineages that morphologically contains O. gelida and O. martensi. Further investigation indicates that the species O. carinata has been misidentified as O. gelida on the Antarctic continental shelf, and that O. intorta and O. inermis are part of what has been termed O. martensi. Most importantly, our study does not delimit species, although it appears that some, such as O. carinata and O. intorta, naturally fall into clades in the radiation. However, it does identify genetic diversity equivalent to species level where morphological characters and taxonomic efforts have failed. We highlight here that genetic diversity needs to be taken into account by conservation managers and policy makers as species names alone fail to capture true diversity present in the Southern Ocean. We strongly advocate the continuing use of CO1, despite its limitations, to identify regions of diversity due to it is relatively low cost (particularly in relation to admittedly more thorough genomic methods), relatively fast application (particularly in comparison to morpho–taxonomy, especially where microstructural analyses need to be applied, which for ophiuroids should be standard practice), and amenability to direct comparisons across all species in the assemblage.
The results of this study align with many other investigations of Southern Ocean benthic biodiversity, indicating that genetic diversity is substantially greater than currently recognised taxonomic diversity. This pattern is apparent not only with Southern Ocean Ophiuroidea (Hunter and Halanych, 2008; Sands et al., 2015, 2021, 2024; Galaska et al., 2017a; Jossart et al., 2019), but also Asteroidea (Janosik and Halanych, 2010; Jossart et al., 2021; Moreau et al., 2021), Echinoidea (Díaz et al., 2011; David et al., 2016), Crinoidea (Wilson et al., 2007; Hemery et al., 2012) and Holothuroidea (O’Loughlin et al., 2011). Indeed, it seems to be a consistent theme among many benthic groups (Raupach and Wägele, 2006; Linse et al., 2007; Leese and Held, 2008; Leese et al., 2008; Krabbe et al., 2010; Schüller, 2011; Dietz et al., 2015; Brasier et al., 2016; Dömel et al., 2017; Hauquier et al., 2017; González-Wevar et al., 2019, 2022). These few examples of the general trend highlight the current extreme underestimation of benthic diversity in the Southern Ocean. At present this situation is being perpetuated because in very few cases have identified cryptic or unrecognised species been subsequently formally described (Janosik and Halanych, 2010; McLaughlin et. al., 2023).
Ophioplinthus species are commonly collected from the Antarctic and sub-Antarctic shelf regions in trawls or dredges, and most often identified as O. gelida. Given the broad range of character states provided by Koehler, particularly in his 1922 monograph, and highlighted in this study, this is understandable (refer to Supplementary Figure 1 for many examples of misidentifications present in GenBank and BOLD). We have found that characters often used in species descriptions, for example, separation of radial shields, presence of an intercalary plate between adoral shields and oral shields, and the form and extent of the keel on the dorsal arm plates (see Figures 5-8) vary both within and between clades (Table 3). Other character states, such as pronounced protuberances on the dorsal arm plates Figures 5-9 panels C and F), and the arrangement of the ventral interradial plates (Figures 5-9B, E) can also vary substantially within and between clades, and yet share character states with other clades (e.g. Figure 5B and Figure 6E share similar ventral interradial plate arrangements despite being different clades). How, then, are collectors, specifically those studying biodiversity, to correctly separate O. gelida from O. carinata or interpret the six clades within O. gelida? Without further investigations, preferably with nuclear markers and morphological microstructure, the outcomes of the study of O. gelida is that it is a radiation consisting of 10 divergent lineages at some point along the speciation trajectory. We now consider some possible scenarios.
First, each clade, or BIN, may represent a unique biological species. If so, one is O. carinata, and one represents O. gelida sensu stricto, with the other four clades being yet-to-be described species. Generating DNA sequence from the syntypes held in the Institute of Natural Sciences in Brussels would help clarify where O. gelida sensu stricto belongs under this scenario (if, indeed, they all belong to the same clade). Although our collections include BIN: AAA8605 (O. carinata), BIN: ABY9430 and BIN: ACF5802 from locations close to the type locality of the O. gelida syntypes, the syntypes are not distinguishable morphologically from some individuals of each of these clades and may belong to any of them.
Second, there is the possibility that the clades of O. gelida may be lineages from a single genetically diverse species. Further population genomic work will be required to explore this scenario. In this case the species would be referred to as O. carinata while O. gelida would become a synonym. However, the six O. gelida clades are genetically equivalent to the four clades of O. martensi in the radiation, although O. martensi is morphologically different from O. gelida. This observation suggests that each clade may be a species.
Third, both explanations may apply, with some clades representing biological species and some biological species being a combination of clades. That the radiation falls into two morphogroups (O. gelida and O. martensi) may suggest this scenario. Our collections confirm that O. carinata is distributed across the Antarctic shelf as well as Antarctic and sub-Antarctic islands, unlike other O. gelida-like clades. This suggests that O. carinata, at least, is likely to be a cohesive species (Templeton, 2001) distinct from the other O. gelida-like clades. Likewise, O. intorta is restricted to the sub-Antarctic implying that it is distinct from other O. martensi-like clades.
As with other cryptic radiations we have studied, the O. gelida/martensi radiation is relatively recent. Although studies have provided a range of clock-rates for ophiuroids, the rates vary and are derived from tropical species not closely related to the Southern Ocean species we have been studying (Naughton et al., 2014; Lessios and Hendler, 2022). Even so, it appears that the radiations occur in the Pleistocene, and our results here suggest that the timing is congruent with radiations in Ophiuroglypha lymani and Amphiura belgicae. Given the recent divergence, there is a likelihood that neither micromorphology nor molecular ecological approaches will delineate clear species boundaries. In all three complexes we see some species-level variation within each, for example, Ophiuroglypha carinifera within the O. lymani complex and A. eugeniae within the A. belgicae complex (Sands et al., 2015, 2024). Even in well-studied animals such as Australian lizards, where sampling is comparatively straightforward, sample sizes high, and good genomic data are available, the short time since divergence coupled with ongoing but uneven hybridisation and introgression among genetic populations cause fuzzy boundaries or low confidence in defining groups (Prates et al., 2024).
Of note is the comparatively high sampling effort achieved in the current study in terms of numbers of samples taken and the spatial scale covered. A total of 502 trawls were conducted over more than a year of total sea time with teams enabling 24 hour working days (Table 1). However, spread across the enormity of the Southern Ocean, it appears that the sampling was patchy (Figure 1), and this is further reflected in the limited spatial coverage of most of our clades of interest (e.g. BIN: AAZ5352, BIN: ACF5801 and BIN: ACF5802, Figure 3, and all O. martensi clades, Figure 4). It may be that our collections reflect the distribution of the clades but, given the patchy nature of the assemblages on the Antarctic shelf sea floor due in part to a mosaic of successional stages (Gutt et al., 1996; Gutt, 2000), it is likely that our sampling is not sufficient to capture the full diversity present at each station, and even in each region. Furthermore, there are some species of Southern Ocean Ophioplinthus that, despite our sampling effort, we have not managed to record in our collections (e.g. O. medusa and O. frigida). This indicates there are discoveries yet to be made. Prior to the identification of potential species complexes in Southern Ocean Ophiuroidea, rarefaction curves of ophiuroid species sampled from regions across the Western Antarctic seas were steep (Sands et al., 2013), indicating that more effort is required to be able to accurately appreciate the diversity and distribution of ophiuroids in the Southern Ocean.
It is, however, clear that multiple genetic lineages exist, indicating a substantially higher diversity than is currently recognised in terms of species names. As genetic diversity is recognised as a critical component of the CBD, some means of taking this observation into account for conservation management purposes is necessary. Suggestions such as evolutionarily significant units (ESUs, Ryder, 1986), management units (MUs, Moritz, 1994) and molecular operational taxonomic units (MOTUs, Floyd et al., 2002) have been posited as surrogates to species. However, although still discussed in the literature, there has been little uptake of these suggestions in regard to implementation into policy or practice likely due to the intensive, expensive and time-consuming work required to collect, define and identify such units, particularly in non-model organisms, precisely what this study demonstrates. In this context, work conducted on forest invertebrates, where large numbers of samples can rapidly be sequenced, photographed and categorised in databases for use for AI identification is an encouraging development (Meier et al., 2024; Vasilita et al., 2024), but unlikely to be adopted by those researching deep sea benthos.
An alternative would be a biogeographic approach using available genetic distribution data to identify regions of diversity. This has long been suggested as a conservation management strategy (Avise, 1989; Bermingham and Moritz, 1998; Moritz and Faith, 1998; Arbogast and Kenagy, 2001). Rather than a species-by-species approach, regions that hold genetically distinct populations of a few representative species could be considered as conservation targets (Faith, 1992; Moritz and Faith, 1998; Laity et al., 2015). Practical examples of such research are abundant in the literature both on land (Smith et al., 2000; Shaffer et al., 2022) and in the sea (DeBoer et al., 2014; von der Heyden et al., 2014). However, examples of such studies directly implemented into policy are difficult to find, with that of Shaffer et al. (2022) being a notable exception.
There have been several studies that highlight the need for more active conservation management in the Southern Ocean, with a specific emphasis on benthic assemblages (Douglass et al., 2014; Brooks et al., 2020; Brasier et al., 2021). Douglass et al. (2014) point out that many of the ecotypes they identified are not under any active protection. In addition, they acknowledge that the assemblages they recognise do not consider genetic variation and thus do not capture this important criterion of the CBD. Our findings here, together with previous studies (Sands et al., 2015, 2024; Jossart et al., 2019), strongly indicate that there are unique lineages, likely to be biological species, often with very limited distributions, across the Southern Ocean. It is interesting that the estimated times of radiations of the O. gelida complex do not appear to differ from those of the Ophiuroglypha lymani complex or the two radiating clades of the Amphiura belgicae/eugeniae complex. The seemingly congruent timing of these events suggests the divergences may have been initiated by a shared historical event, allowing us to speculate that other cryptic radiations identified in previous studies may also be the result of the same event.
In conclusion, the plastic morphology of the O. carinata/O. gelida/O. martensi/O. intorta/O. inermis complex will continue to inhibit identification of collections. Ophioplinthus carinata make up a substantial proportion of the collections currently identified as O. gelida from the Antarctic shelf, a fact not recognised until now. We suggest that specimens assigned as O. martensi in collections from around the sub-Antarctic island shelves be considered as Ophioplinthus intorta (Lyman, 1878), while those collected around the Antarctic shelf may be considered Ophioplinthus inermis (Bell, 1902) comb. nov., resurrected from its synonymy with O. martensi. To better understand the species status of the clades in this complex, detailed micro-morphological examination of skeletal plates should be undertaken, along with population genomic studies to more robustly identify species boundaries. Despite relatively intensive sampling, we still do not have sufficient representation of each clade of this complex to confidently describe their distributions. The entire genus Ophioplinthus requires further systematic investigation, with several species outside of the O. gelida complex showing deep divergences that are also likely to represent unrecognised and undescribed species, greatly increasing the recognised diversity of Southern Ocean brittle stars and endemism of regional assemblages.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
CS: Resources, Project administration, Funding acquisition, Formal Analysis, Validation, Data curation, Conceptualization, Methodology, Writing – original draft, Writing – review & editing, Investigation. TO’H: Data curation, Investigation, Writing – review & editing, Resources. AG: Data curation, Resources, Writing – review & editing, Investigation. WG-C: Writing – review & editing, Investigation, Supervision. PC: Project administration, Supervision, Writing – review & editing. BN: Project administration, Writing – review & editing, Supervision. RM-L: Resources, Writing – review & editing, Data curation. SS: Writing – review & editing, Methodology, Supervision.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. Funding for sequencing was largely provided by the Census of Antarctic Marine Life. Curation and taxonomic determinations were funded by SYNTAX grant and an Antarctic Science Bursary to CS. CS and PC are supported by NERC core funding to the ‘Biodiversity, Evolution and Adaptation’ team.
Acknowledgments
We would like to thank the captains and crews of the RRS James Clark Ross and FS Polarstern for assistance in collecting samples. Samples were collected during cruises JR144, JR179, JR230, JR262, JR275, PS77 and PS82. We thank Rachel Downey and Camille Moreau for assistance sorting samples. A special thanks to Elena Zaikina who assisted RM-L with taxonomic determinations and documentation. We thank Prof. Andreas Schmit-Rhaesa of Museum de Natur Hamburg, Dr Yves Samyn of the Institute of Natural Sciences, Brussels, Shirley Sorokin of South Australian Museum, and Prof. Torsten Struck and Åse Wilhelmsen of University of Oslo Natural History Museum for provision of type material imagery. Thanks to Dr Hugh Carter of London Natural History Museum and Dr Stefano Schiaparelli of the Italian National Antarctic Museum for access to the collections.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1615695/full#supplementary-material
Supplementary Figure 1 | Unedited RAxML phylogeny of all sequences, with accession numbers (BOLD and GenBank), allocated BINs and putative (initial) identifications.
Supplementary Figure 2 | Population networks of the two most populous clades of the Ophioplinthus gelida complex. Circles are distinct haplotypes (unique sequences), circle diameter is proportional to number of sequences represented, hash lines are unsampled haplotypes. Each line represents a step change.
Supplementary Table 1 | A list of cruises and stations included in this study.
Supplementary Table 2 | A list of all relevant type material examined for this study.
References
Arango C. P., Soler-Membrives A., and Miller K. J. (2011). Genetic differentiation in the circum—Antarctic sea spider Nymphon australe (Pycnogonida; Nymphonidae). Deep Sea Res. Part II Top. Stud. Oceanogr. 58, 212–219. doi: 10.1016/j.dsr2.2010.05.019
Arbogast B. S. and Kenagy G. J. (2001). Comparative phylogeography as an integrative approach to historical biogeography. J. Biogeogr. 28, 819–825. doi: 10.1046/j.1365-2699.2001.00594.x
Avise J. C. (1989). A role for molecular genetics in the recognition and conservation of endangered species. Trends Ecol. Evol. 4, 279–281. doi: 10.1016/0169-5347(89)90203-6
Ayoub N. A. and Riechert S. E. (2004). Molecular evidence for Pleistocene glacial cycles driving diversification of a North American desert spider, Agelenopsis aperta. Mol. Ecol. 13, 3453–3465. doi: 10.1111/j.1365-294X.2004.02335.x
Bell F. J. (1902). Reports on the collections of Natural History made in the Antarctic regions during the voyage of the “Southern Cross”. VIII. Echinoderma. 214–220, 2 pl.
Bermingham E. and Moritz C. (1998). Comparative phylogeography - concepts and applications. Mol. Ecol. 7, 367–369. doi: 10.1046/j.1365-294x.1998.00424.x
Bouckaert R., Heled J., Kühnert D., Vaughan T., Wu C.-H., Xie D., et al. (2014). BEAST 2: A software platform for bayesian evolutionary analysis. PloS Comput. Biol. 10, e1003537. doi: 10.1371/journal.pcbi.1003537
Brasier M. J., Barnes D., Bax N., Brandt A., Christianson A. B., Constable A. J., et al. (2021). Responses of southern ocean seafloor habitats and communities to global and local drivers of change. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.622721
Brasier M. J., Wiklund H., Neal L., Jeffreys R., Linse K., Ruhl H., et al. (2016). DNA barcoding uncovers cryptic diversity in 50% of deep-sea Antarctic polychaetes. Open Sci. 3, 160432. doi: 10.1098/rsos.160432
Brooks C. M., Chown S. L., Douglass L. L., Raymond B. P., Shaw J. D., Sylvester Z. T., et al. (2020). Progress towards a representative network of Southern Ocean protected areas. PloS One 15, e0231361. doi: 10.1371/journal.pone.0231361
Caughley G. (1994). Directions in conservation biology. J. Anim. Ecol. 63, 215–244. doi: 10.2307/5542
Clark H. L. (1915). Catalogue of recent ophiurans: based on the collection of the Museum of Comparative Zoölogy. Memoirs Museum Comp. Zoology 25(4), 165–376.
Clarke A., Aronson R., Crame J., Gili J., and Blake D. (2004). Evolution and diversity of the benthic fauna of the Southern Ocean continental shelf. Antarct. Sci. 16, 559–568. doi: 10.1017/S0954102004002329
Clarke A. and Crame J. A. (1992). The Southern Ocean benthic fauna and climate change: a historical perspective. Philos. Trans. R. Soc Lond. B. Biol. Sci. 338, 299–309. doi: 10.1098/rstb.1992.0150
Clarke A. and Crame J. A. (2010). Evolutionary dynamics at high latitudes: speciation and extinction in polar marine faunas. Philos. Trans. R. Soc B Biol. Sci. 365, 3655–3666. doi: 10.1098/rstb.2010.0270
Clarke A. and Johnston N. M. (2003). Antarctic marine benthic diversity. Oceanogr. Mar. Biol. Annu. Rev. 41, 47–114.
Clement M., Posada D., and Crandall K. A. (2000). TCS: a computer program to estimate gene genealogies. Mol. Ecol. 9, 1657–1659. doi: 10.1046/j.1365-294x.2000.01020.x
Convey P., Biersma E. M., Casanova-Katny A., and Maturana C. S. (2020). “Refuges of antarctic diversity,” in Past Antarctica. Eds. Oliva M. and Ruiz-Fernández J. (Academic Press), 181–200. doi: 10.1016/B978-0-12-817925-3.00010-0
Convey P., Gibson J. A. E., Hillenbrand C.-D., Hodgson D. A., Pugh P. J. A., Smellie J. L., et al. (2008). Antarctic terrestrial life - challenging the history of the frozen continent? Biol. Rev. 83, 103–117. doi: 10.1111/j.1469-185X.2008.00034.x
Convey P., Stevens M. I., Hodgson D. A., Smellie J. L., Hillenbrand C.-D., Barnes D. K. A., et al. (2009). Exploring biological constraints on the glacial history of Antarctica. Quat. Sci. Rev. 28, 3035–3048. doi: 10.1016/j.quascirev.2009.08.015
Costello M. J., Wilson S., and Houlding B. (2013). More taxonomists describing significantly fewer species per unit effort may indicate that most species have been discovered. Syst. Biol. 62, 616–624. doi: 10.1093/sysbio/syt024
Darriba D., Taboada G. L., Doallo R., and Posada D. (2012). jModelTest 2: more models, new heuristics and parallel computing. Nat. Methods 9, 772–772. doi: 10.1038/nmeth.2109
David B., Saucède T., Chenuil A., Steimetz E., and De Ridder C. (2016). The taxonomic challenge posed by the Antarctic echinoids Abatus bidens and Abatus cavernosus (Schizasteridae, Echinoidea). Polar Biol. 39, 897–912. doi: 10.1007/s00300-015-1842-5
DeBoer T. S., Naguit M. R. A., Erdmann M. V., Ablan-Lagman M. C. A., Carpenter K. E., Toha A. H. A., et al. (2014). Concordance between phylogeographic and biogeographic boundaries in the Coral Triangle: conservation implications based on comparative analyses of multiple giant clam species. Bull. Mar. Sci. 90, 277–300. doi: 10.5343/bms.2013.1003
De Queiroz K. (2007). Species concepts and species delimitation. Syst. Biol. 56, 879–886. doi: 10.1080/10635150701701083
Díaz A., Féral J.-P., David B., Saucède T., and Poulin E. (2011). Evolutionary pathways among shallow and deep-sea echinoids of the genus Sterechinus in the Southern Ocean. Deep Sea Res. Part II Top. Stud. Oceanogr. 58, 205–211. doi: 10.1016/j.dsr2.2010.10.012
Dietz L., Arango C. P., Dömel J. S., Halanych K. M., Harder A. M., Held C., et al. (2015). Regional differentiation and extensive hybridization between mitochondrial clades of the Southern Ocean giant sea spider Colossendeis megalonyx. Open Sci. 2, 140424. doi: 10.1098/rsos.140424
Dömel J. S., Melzer R. R., Harder A. M., Mahon A. R., and Leese F. (2017). Nuclear and mitochondrial gene data support recent radiation within the sea spider species complex Pallenopsis patagonica. Front. Ecol. Evol. 4. doi: 10.3389/fevo.2016.00139
Doorenweerd C., San Jose M., Geib S., Barr N., and Rubinoff D. (2024). Genomic data reveal new species and the limits of mtDNA barcode diagnostics to contain a global pest species complex (Diptera: Tephritidae: Dacinae). Syst. Entomol. 49, 279–293. doi: 10.1111/syen.12616
Douglass L. L., Turner J., Grantham H. S., Kaiser S., Constable A., Nicoll R., et al. (2014). A hierarchical classification of benthic biodiversity and assessment of protected areas in the Southern Ocean. PloS One 9, e100551. doi: 10.1371/journal.pone.0100551
Engel M. S., Ceríaco L. M. P., Daniel G. M., Dellapé P. M., Löbl I., Marinov M., et al. (2021). The taxonomic impediment: a shortage of taxonomists, not the lack of technical approaches. Zool. J. Linn. Soc 193, 381–387. doi: 10.1093/zoolinnean/zlab072
Faith D. P. (1992). Conservation evaluation and phylogenetic diversity. Biol. Conserv. 61, 1–10. doi: 10.1016/0006-3207(92)91201-3
Fell H. B. (1961). The fauna of the Ross Sea: Ophiuroidea. New Z. Department Sci. Ind. Res. Bull. 142, 1–79.
Floyd R., Abebe E., Papert A., and Blaxter M. L. (2002). Molecular barcodes for soil nematode identification. Mol. Ecol. 11, 839–850.
Galaska M. P., Sands C. J., Santos S. R., Mahon A. R., and Halanych K. M. (2017a). Crossing the divide: admixture across the antarctic polar front revealed by the brittle star Astrotoma agassizii. Biol. Bull. 232, 198–211. doi: 10.1086/693460
Galaska M. P., Sands C. J., Santos S. R., Mahon A. R., and Halanych K. M. (2017b). Geographic structure in the Southern Ocean circumpolar brittle star Ophionotus victoriae (Ophiuridae) revealed from mtDNA and single-nucleotide polymorphism data. Ecol. Evol. 7(3), 475–485. doi: 10.1002/ece3.2617
Garrick R. C., Sands C. J., Rowell D. M., Tait N. N., Greenslade P., and Sunnucks P. (2004). Phylogeography recapitulates topography: very fine-scale local endemism of a saproxylic “giant” springtail at Tallaganda in the Great Dividing Range of south-east Australia. Mol. Ecol. 13, 3329–3344. doi: 10.1111/j.1365-294X.2004.02340.x
Gómez A. and Lunt D. H. (2007). “Refugia within refugia: patterns of phylogeographic concordance in the Iberian Peninsula,” in Phylogeography of Southern European Refugia: Evolutionary perspectives on the origins and conservation of European biodiversity. Eds. Weiss S. and Ferrand N. (Springer Netherlands, Dordrecht), 155–188. doi: 10.1007/1-4020-4904-8_5
González-Wevar C. A., Gérard K., Rosenfeld S., Saucède T., Naretto J., Díaz A., et al. (2019). Cryptic speciation in Southern Ocean Aequiyoldia eightsii (Jay 1839): Mio-Pliocene trans-Drake Passage separation and diversification. Prog. Oceanogr. 174, 44–54. doi: 10.1016/j.pocean.2018.09.004
González-Wevar C. A., Segovia N. I., Rosenfeld S., Maturana C. S., Jeldres V., Pinochet R., et al. (2022). Seven snail species hidden in one: Biogeographic diversity in an apparently widespread periwinkle in the Southern Ocean. J. Biogeogr. 49, 1521–1534. doi: 10.1111/jbi.14453
Goodall-Copestake W. P., Tarling G. A., and Murphy E. J. (2012). On the comparison of population-level estimates of haplotype and nucleotide diversity: a case study using the gene cox1 in animals. Heredity 109, 50–56. doi: 10.1038/hdy.2012.12
Gray J. (1867). Notes on the Arrangement of Sponges, with the Descriptions of some New Genera. Proc. Zoological Soc. London. 1867(2), 492–558.
Gutt J. (2000). Some “driving forces” structuring communities of the sublittoral Antarctic macrobenthos. Antarct. Sci. 12, 297–313. doi: 10.1017/S0954102000000365
Gutt J., Starmans A., and Dieckmann G. (1996). Impact of iceberg scouring on polar benthic habitats. Mar. Ecol. Prog. Ser. 137, 311–316. doi: 10.3354/meps137311
Hauquier F., Leliaert F., Rigaux A., Derycke S., and Vanreusel A. (2017). Distinct genetic differentiation and species diversification within two marine nematodes with different habitat preference in Antarctic sediments. BMC Evol. Biol. 17, 120. doi: 10.1186/s12862-017-0968-1
Havermans C., Nagy Z. T., Sonet G., De Broyer C., and Martin P. (2011). DNA barcoding reveals new insights into the diversity of Antarctic species of Orchomene sensu lato (Crustacea: Amphipoda: Lysianassoidea). Deep Sea Res. Part II Top. Stud. Oceanogr. 58, 230–241. doi: 10.1016/j.dsr2.2010.09.028
Hemery L. G., Eléaume M., Roussel V., Améziane N., Gallut C., Steinke D., et al. (2012). Comprehensive sampling reveals circumpolarity and sympatry in seven mitochondrial lineages of the Southern Ocean crinoid species Promachocrinus kerguelensis (Echinodermata). Mol. Ecol. 21, 2502–2518. doi: 10.1111/j.1365-294X.2012.05512.x
Hewitt G. M. (2000). The genetic legacy of the Quaternary ice ages. Nature 405, 907–913. doi: 10.1038/35016000
Hoban S., da Silva J. M., Hughes A., Hunter M. E., Kalamujić Stroil B., Laikre L., et al. (2024). Too simple, too complex, or just right? Advantages, challenges, and guidance for indicators of genetic diversity. BioScience 74, 269–280. doi: 10.1093/biosci/biae006
Hunter R. L. (2007). Morphological cladistic analysis of Ophiurolepis Matsumoto 1915 (Ophiurida: ophiuridae) from the Southern ocean. Zootaxa 1401, 33–51. doi: 10.11646/zootaxa.1401.2
Hunter R. L. and Halanych K. M. (2008). Evaluating connectivity in the brooding brittle star Astrotoma agassizii across the Drake Passage in the Southern Ocean. J. Hered. 99, 137–148. doi: 10.1093/jhered/esm119
Janosik A. M. and Halanych K. M. (2010). Unrecognized Antarctic biodiversity: A case study of the genus Odontaster (Odontasteridae; Asteroidea). Integr. Comp. Biol. 50, 981–992. doi: 10.1093/icb/icq119
Janosik A. M., Mahon A. R., and Halanych K. M. (2011). Evolutionary history of Southern Ocean Odontaster sea star species (Odontasteridae; Asteroidea). Polar Biol. 34, 575–586. doi: 10.1007/s00300-010-0916-7
Jossart Q., Kochzius M., Danis B., Saucède T., and Moreau C. V. E. (2021). Diversity of the Pterasteridae (Asteroidea) in the Southern Ocean: a molecular and morphological approach. Zool. J. Linn. Soc 192, 105–116. doi: 10.1093/zoolinnean/zlaa097
Jossart Q., Sands C. J., and Sewell M. A. (2019). Dwarf brooder versus giant broadcaster: combining genetic and reproductive data to unravel cryptic diversity in an Antarctic brittle star. Heredity 123, 622–633. doi: 10.1038/s41437-019-0228-9
Katoh K. and Standley D. M. (2013). MAFFT Multiple Sequence Alignment Software Version 7: Improvements in performance and usability. Mol. Biol. Evol. 30, 772–780. doi: 10.1093/molbev/mst010
Knowles L. L. and Richards C. L. (2005). Importance of genetic drift during Pleistocene divergence as revealed by analyses of genomic variation. Mol. Ecol. 14, 4023–4032. doi: 10.1111/j.1365-294X.2005.02711.x
Koehler R. (1900). Note preliminaire sur les Echinides et les Ophiures de l'Expédition Antarctique Belge. Bull. Academie Royale Belgique. 11(3), 814–820.
Koehler R. (1901). Expédition Antarctique Belge. Résultats du voyage de S. Y. “Belgica” en 1897-98-99. Echinides Ophiures. Zoologie 1–42.
Koehler R. (1906). Note préliminaire sur les Échinodermes recueillis par l’expédition, antarctique franYaise du Dr. Charcot (Échinides, Astéries et Ophiures). Bull. du Museum Paris 1905, 464–470.
Koehler R. (1907). Revision de la collection des Ophiures du Museum d’histoire Naturelle Paris. Bull. Sci. Fr. Belg. 41, 279–351.
Koehler R. (1908). Astéries, Ophiures et Échinides de l’Expédition Antarctique Nationale Écossaise. Trans. R. Soc Edinb. 46, 529–649.
Koehler R. (1911). Astéries, Ophiures, et Échinides de l’Expédition Antarctique Anglaise de 1907-1909. British Antarctic Expedition 1907-1909. Rep. Sci. Investig. 2 Biol. 4, 25–66.
Koehler R. (1912). “Echinodermes (Astéries, Ophiures et Échinides),” in Deuxième Expédition Antarctique Française, (1908–10) (Paris: Ministère de l'instruction publique), 272.
Korshunova T., Picton B., Furfaro G., Mariottini P., Pontes M., Prkić J., et al. (2019). Multilevel fine-scale diversity challenges the ‘cryptic species’ concept. Sci. Rep. 9, 6732. doi: 10.1038/s41598-019-42297-5
Krabbe K., Leese F., Mayer C., Tollrian R., and Held C. (2010). Cryptic mitochondrial lineages in the widespread pycnogonid Colossendeis megalonyx Hoek 1881 from Antarctic and Subantarctic waters. Polar Biol. 33, 281–292. doi: 10.1007/s00300-009-0703-5
Kumar S., Stecher G., Li M., Knyaz C., and Tamura K. (2018). MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 35, 1547–1549. doi: 10.1093/molbev/msy096
Laity T., Laffan S. W., Gonzalez-Orozco C. E., Faith D. P., Rosauer D. F., Byrne M., et al. (2015). Phylodiversity to inform conservation policy: An Australian example. Sci. Total Environ. 534, 131–143. doi: 10.1016/j.scitotenv.2015.04.113
Lamka G. F. and Willoughby J. R. (2024). Habitat remediation followed by managed connectivity reduces unwanted changes in evolutionary trajectory of high extirpation risk populations. PloS One 19, e0304276. doi: 10.1371/journal.pone.0304276
Lau S. C. Y., Strugnell J. M., Sands C. J., Silva C. N. S., and Wilson N. G. (2021). Evolutionary innovations in Antarctic brittle stars linked to glacial refugia. Ecol. Evol. 11 (23), 17428–17446. doi: 10.1002/ece3.8376
Lau S. C. Y., Strugnell J. M., Sands C. J., Silva C. N. S., and Wilson N. G. (2023). Genomic insights of evolutionary divergence and life history innovations in Antarctic brittle stars. Mol. Ecol. 32, 3382–3402. doi: 10.1111/mec.16951
Leese F. and Held C. (2008). Identification and characterization of microsatellites from the Antarctic isopod Ceratoserolis trilobitoides: nuclear evidence for cryptic species. Conserv. Genet. 9, 1369–1372. doi: 10.1007/s10592-007-9491-z
Leese F., Kop A., Wagele J.-W., and Held C. (2008). Cryptic speciation in a benthic isopod from Patagonian and Falkland Island waters and the impact of glaciations on its population structure. Front. Zool. 5, 19. doi: 10.1186/1742-9994-5-19
Lessios H. A. and Hendler G. (2022). Mitochondrial phylogeny of the brittle star genus Ophioderma. Sci. Rep. 12, 5304. doi: 10.1038/s41598-022-08944-0
Lessios H. A., Kessing B. D., and Pearse J. S. (2001). Population structure and speciation in tropical seas: global phylogeography of the sea urchin Diadema. Evolution 55, 955–975. doi: 10.1111/j.0014-3820.2001.tb00613.x
Librado P. and Rozas J. (2009). DnaSP v5: a software for comprehensive analysis of DNA polymorphism data. Bioinformatics 25, 1451–1452. doi: 10.1093/bioinformatics/btp187
Linse K., Cope T., Lörz A.-N., and Sands C. J. (2007). Is the Scotia Sea a center of Antarctic marine diversification? Some evidence of cryptic speciation in the circum-Antarctic bivalve Lissarca notorcadensis (Arcoidea: Philobryidae). Polar Biol. 30, 1059–1068. doi: 10.1007/s00300-007-0265-3
Lyman T. (1860). Descriptions of new Ophiuridae, belonging to the Smithsonian Institution and to the Museum of Comparative Zoology at Cambridge. Proc. Boston Soc. Natural History 1859-61. 7, 193–204.
Lyman T. (1875). Ophiuridae and Astrophytidae: including those dredged by the late Dr. William Stimpson (University Press).
Lyman T. (1878). Ophiuridae and Astrophytidae of the”Challenge” expedition. Part I. Bull. Mus. Comp. Zool. Harv. Coll. 5, 65–168.
Lyman T. (1883). Reports on the results of dredging, under the supervision of Alexander Agassiz, in the Caribbean Sea, (1878-79), and on the east coast of the United States, during the summer of 1880, by the U.S. coast survey steamer “Blake”, commander J.R. Bartlett, U.S.N., commanding. XX. - Report on the Ophiuroidea. Bull. Mus. Comp. Zool. Harv. Coll. 10, 227–287.
Madsen F. J. (1955). Echinoderms other than holothurians collected in sub-Antarctic and Antarctic seas, mainly by the Norvegia-expeditions 1928-30. Sci. Res. Norw Antarct Exped 1927–1928 37, 1–17.
Madsen F. J. (1967). Ophiuroidea. Br. Aust. New Z. Antarctic Res. Expedition 1929–1931 Rep. – Ser. B (Zool. Botany) 9, 120–145.
Martín-Ledo R., Sands C. J., and López-González P. J. (2013). A new brooding species of brittle star (Echinodermata: Ophiuroidea) from Antarctic waters. Polar Biol. 36, 115–126. doi: 10.1007/s00300-012-1242-z
Martynov A. V. and Litvinova N. M. (2008). Deep-water Ophiuroidea of the northern Atlantic with descriptions of three new species and taxonomic remarks on certain genera and species. Mar. Biol. Res. 4, 76–111. doi: 10.1080/17451000701840066
Matsumoto H. (1915). A new classification of the Ophiuroidea. Proc. Acad. Natural Sci. Philadelphia 67, 43–92.
May R. M. (1988). How many species are there on Earth? Science 241, 1441–1449. doi: 10.1126/science.241.4872.1441
McLaughlin E. L., Wilson N. G., Rouse G. W., McLaughlin E. L., Wilson N. G., and Rouse G. W. (2023). Resolving the taxonomy of the Antarctic feather star species complex Promachocrinus ‘kerguelensis’ (Echinodermata: Crinoidea). Invertebr. Syst. 37, 498–527. doi: 10.1071/IS22057
Meier R., Srivathsan A., Oliveira S. S., Balbi M. I. P. A., Ang Y., Yeo D., et al. (2024). “Dark taxonomy”: a new protocol for overcoming the taxonomic impediments for dark taxa and broadening the taxon base for biodiversity assessment. Cladistics. 41, 223–238. doi: 10.1111/cla.12609
Mora C., Tittensor D. P., Adl S., Simpson A. G. B., and Worm B. (2011). How many species are there on earth and in the ocean? PloS Biol. 9, e1001127. doi: 10.1371/journal.pbio.1001127
Moreau C., Jossart Q., Danis B., Eléaume M., Christiansen H., Guillaumot C., et al. (2021). The high diversity of Southern Ocean sea stars (Asteroidea) reveals original evolutionary pathways. Prog. Oceanogr. 190, 102472. doi: 10.1016/j.pocean.2020.102472
Moritz C. (1994). Defining ‘Evolutionarily Significant Units’ for conservation. Trends Ecol. Evol. 9, 373–375. doi: 10.1016/0169-5347(94)90057-4
Moritz C. and Faith D. P. (1998). Comparative phylogeography and the identification of genetically divergent areas for conservation. Mol. Ecol. 7, 419–429. doi: 10.1046/j.1365-294x.1998.00317.x
Mortensen T. (1925). On a small collection of echinoderms from the Antarctic Sea. Arkiv för Zoologi 17A, 1–12.
Mortensen T. (1936). “Echinoidea and Ophiuroidea,” in Discovery Reports (Cambridge: Cambridge University Press), 199–348.
Naughton K. M., O’Hara T. D., Appleton B., and Cisternas P. A. (2014). Antitropical distributions and species delimitation in a group of ophiocomid brittle stars (Echinodermata: Ophiuroidea: Ophiocomidae). Mol. Phylogenet. Evol. 78, 232–244. doi: 10.1016/j.ympev.2014.05.020
Nirmal B., Mohan K., Prakasam M., Tripati A., Mortyn P. G., and Rodríguez-Sanz L. (2021). Pleistocene surface-ocean changes across the Southern subtropical front recorded by cryptic species of Orbulina universa. Mar. Micropaleontol. 168, 102056. doi: 10.1016/j.marmicro.2021.102056
O’Hara T. D., Hugall A. F., Thuy B., Stöhr S., and Martynov A. V. (2017). Restructuring higher taxonomy using broad-scale phylogenomics: The living Ophiuroidea. Mol. Phylogenet. Evol. 107, 415–430. doi: 10.1016/j.ympev.2016.12.006
O’Hara T. D., Hugall A. F., Woolley S. N. C., Bribiesca-Contreras G., and Bax N. J. (2019). Contrasting processes drive ophiuroid phylodiversity across shallow and deep seafloors. Nature 565, 636–639. doi: 10.1038/s41586-019-0886-z
O’Hara T. D., Stöhr S., Hugall A. F., Thuy B., and Martynov A. (2018). Morphological diagnoses of higher taxa in Ophiuroidea (Echinodermata) in support of a new classification. Eur. J. Taxon. 0, 1–35. doi: 10.5852/ejt.2018.416
O’Loughlin M. P., Paulay G., Davey N., and Michonneau F. (2011). The Antarctic region as a marine biodiversity hotspot for echinoderms: Diversity and diversification of sea cucumbers. Deep Sea Res. Part II Top. Stud. Oceanogr. 58, 264–275. doi: 10.1016/j.dsr2.2010.10.011
Paterson G. L. (1985). The deep-sea Ophiuroidea of the north Atlantic Ocean. Bull. Br. Museum (Natural History) 49, 1–162.
Paulo O. S., Dias C., Bruford M. W., Jordan W. C., and Nichols R. A. (2001). The persistence of Pliocene populations through the Pleistocene climatic cycles: evidence from the phylogeography of an Iberian lizard. Proc. R. Soc Lond. - Ser. B Biol. Sci. 268, 1625–1630. doi: 10.1098/rspb.2001.1706
Prates I., Hutchinson M. N., Singhal S., Moritz C., and Rabosky D. L. (2024). Notes from the taxonomic disaster zone: Evolutionary drivers of intractable species boundaries in an Australian lizard clade (Scincidae: Ctenotus). Mol. Ecol. 33, e17074. doi: 10.1111/mec.17074
Rabosky D. L., Chang J., Title P. O., Cowman P. F., Sallan L., Friedman M., et al. (2018). An inverse latitudinal gradient in speciation rate for marine fishes. Nature 559, 392–395. doi: 10.1038/s41586-018-0273-1
Ratnasingham S. and Hebert P. D. N. (2013). A DNA-based registry for all animal species: the barcode index number (BIN) system. PloS One 8, e66213. doi: 10.1371/journal.pone.0066213
Raupach M. J. and Wägele J. W. (2006). Distinguishing cryptic species in Antarctic Asellota (Crustacea: Isopoda)-a preliminary study of mitochondrial DNA in Acanthaspidia drygalskii. Antarct. Sci. 18, 191–198. doi: 10.1017/S0954102006000228
Ryder O. A. (1986). Species conservation and systematics: the dilemma of subspecies. Trends Ecol. Evol. doi: 10.1016/0169-5347(86)90059-5
Sands C. J., Goodall-Copestake W., Stöhr S., Narayanaswamy B. E., Convey P., O’Hara T., et al. (2024). The diverse and widespread Southern Ocean ophiuroid Amphiura belgicae should be considered a species complex. Front. Ecol. Evol. 12. doi: 10.3389/fevo.2024.1416418
Sands C. J., Griffiths H. J., Downey R. V., Barnes D. K. A., Linse K., and Martín-Ledo R. (2013). Observations of the ophiuroids from the West Antarctic sector of the Southern Ocean. Antarct. Sci. 25, 3–10. doi: 10.1017/S0954102012000612
Sands C. J., O’Hara T., Barnes D. K. A., and Martín-Ledo R. (2015). Against the flow: evidence of multiple recent invasions of warmer continental shelf waters by a Southern Ocean brittle star. Evol. Popul. Genet. 3. doi: 10.3389/fevo.2015.00063
Sands C. J., O’Hara T. D., and Martín-Ledo R. (2021). Pragmatic assignment of species groups based on primary species hypotheses: the case of a dominant component of the Southern Ocean benthic fauna. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.723328
Schonswetter P., Tribsch A., Stehlik I., and Niklfeld H. (2004). Glacial history of high alpine Ranunculus glacialis (Ranunculaceae) in the European Alps in a comparative phylogeographical context. Biol. J. Linn. Soc 81, 183–195. doi: 10.1111/j.1095-8312.2003.00289.x
Schüller M. (2011). Evidence for a role of bathymetry and emergence in speciation in the genus Glycera (Glyceridae, Polychaeta) from the deep Eastern Weddell Sea. Polar Biol. 34, 549–564. doi: 10.1007/s00300-010-0913-x
Shaffer H. B., Toffelmier E., Corbett-Detig R. B., Escalona M., Erickson B., Fiedler P., et al. (2022). Landscape genomics to enable conservation actions: the California conservation genomics project. J. Hered. 113, 577–588. doi: 10.1093/jhered/esac020
Smith T. B., Holder K., Girman D., O’Keefe K., Larison B., and Chan Y. (2000). Comparative avian phylogeography of Cameroon and Equatorial Guinea mountains: implications for conservation. Mol. Ecol. 9, 1505–1516. doi: 10.1046/j.1365-294x.2000.01032.x
Sommer R. S. and Zachos F. E. (2009). Fossil evidence and phylogeography of temperate species: “glacial refugia” and post-glacial recolonization. J. Biogeogr. 36, 2013–2020. doi: 10.1111/j.1365-2699.2009.02187.x
Stamatakis A. (2014). RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30, 1312–1313. doi: 10.1093/bioinformatics/btu033
Stork N. E. (1993). How many species are there? Biodivers. Conserv. 2, 215–232. doi: 10.1007/BF00056669
Studer T. (1876). Echinodermen aus dem antarktischen Meere und zwei neue Seeigel von den Papua-Inseln, gesammelt auf de Reise S.M.S. Gazelle um die Erde. Monatsbericht K. Preuss. Akad. Wiss. Ze Berl., 452–465.
Studer T. (1885). Die Seesterne Süd-Georgiens nach der Ausbeute der deutschen Polarstation in 1882 und 1993. Jahrbuch der Hamburgischen Wissenchaftlichen Anstalten 2, 143–166.
Taberlet P., Fumagalli L., Wustsaucy A. G., and Cosson J. F. (1998). Comparative phylogeography and postglacial colonization routes in Europe. Mol. Ecol. 7, 453–464. doi: 10.1046/j.1365-294x.1998.00289.x
Templeton A. R. (2001). Using phylogeographic analyses of gene trees to test species status and processes. Mol. Ecol. 10, 779–791. doi: 10.1111/j.1365-294X.2001.01199.x
Thatje S., Hillenbrand C.-D., and Larter R. (2005). On the origin of Antarctic marine benthic community structure. Trends Ecol. Evol. 20, 534–540. doi: 10.1016/j.tree.2005.07.010
Thatje S., Hillenbrand C.-D., Mackensen A., and Larter R. (2008). Life hung by a thread: endurance of Antarctic fauna in glacial periods. Ecology 89, 682–692. doi: 10.1890/07-0498.1
Trewick S. A. and Wallis G. P. (2001). Bridging the “beech-gap”: New Zealand invertebrate phylogeography implicates pleistocene glaciation and pliocene isolation. Evolution 55, 2170–2180. doi: 10.1111/j.0014-3820.2001.tb00733.x
Tribsch A. (2004). Areas of endemism of vascular plants in the Eastern Alps in relation to Pleistocene glaciation. J. Biogeogr. 31, 747–760. doi: 10.1111/j.1365-2699.2004.01065.x
Turvey S. T. and Crees J. J. (2019). Extinction in the anthropocene. Curr. Biol. 29, R982–R986. doi: 10.1016/j.cub.2019.07.040
United Nations (1992). Convention on biological diversity (Rio de Janeiro: United Nations Environment Program). Available online at: https://www.cbd.int/doc/legal/cbd-en.pdf (Accessed May 4, 2025).
Vasilita C., Feng V., Hansen A. K., Hartop E., Srivathsan A., Struijk R., et al. (2024). Express barcoding with NextGenPCR and MinION for species-level sorting of ecological samples. Mol. Ecol. Resour. 24, e13922. doi: 10.1111/1755-0998.13922
Vignes-Lebbe R., Chesselet P., Diep Thi M. H., Rakotoarisoa N. R., Blackmore S., and Riera B. (2016). “Xper3: new tools for collaborating, training and transmitting knowledge on botanical phenotypes,” in Botanists of the Twenty-First Century: Roles Challenges and Opportunities (UNESCO, Paris), 228–239.
von der Heyden S., Beger M., Toonen R. J., van Herwerden L., Juinio-Meñez M. A., Ravago-Gotanco R., et al. (2014). The application of genetics to marine management and conservation: examples from the Indo-Pacific. Bull. Mar. Sci. 90, 123–158. doi: 10.5343/bms.2012.1079
Wattier R., Mamos T., Copilaş-Ciocianu D., Jelić M., Ollivier A., Chaumot A., et al. (2020). Continental-scale patterns of hyper-cryptic diversity within the freshwater model taxon Gammarus fossarum (Crustacea, Amphipoda). Sci. Rep. 10, 16536. doi: 10.1038/s41598-020-73739-0
Weckworth B. V., Musiani M., DeCesare N. J., McDevitt A. D., Hebblewhite M., and Mariani S. (2013). Preferred habitat and effective population size drive landscape genetic patterns in an endangered species. Proc. R. Soc B Biol. Sci. 280, 20131756. doi: 10.1098/rspb.2013.1756
Wiens J. J. (2023). How many species are there on Earth? Prog. problems. PloS Biol. 21, e3002388. doi: 10.1371/journal.pbio.3002388
Wilson N., Hunter R., Lockhart S., and Halanych K. (2007). Multiple lineages and absence of panmixia in the “circumpolar” crinoid Promachocrinus kerguelensis from the Atlantic sector of Antarctica. Mar. Biol. 152, 895–904. doi: 10.1007/s00227-007-0742-9
WoRMS Editorial Board. (2025). World register of marine species (WoRMS). Available online at: https://www.marinespecies.org (Accessed April 16, 2025).
Keywords: diversity, taxonomy, conservation biology, speciation, benthos, Ophiuroidea, Southern Ocean
Citation: Sands CJ, O’Hara TD, Guzzi A, Goodall-Copestake WP, Convey P, Narayanaswamy BE, Martín-Ledo R and Stöhr S (2025) And then there were many: insights from the tangled taxonomy of the Antarctic brittle star Ophioplinthus gelida (Echinodermata: Ophiuroidea). Front. Mar. Sci. 12:1615695. doi: 10.3389/fmars.2025.1615695
Received: 21 April 2025; Accepted: 23 June 2025;
Published: 29 July 2025.
Edited by:
Didier Alain Jollivet, Centre National de la Recherche Scientifique (CNRS), FranceReviewed by:
Saskia Brix, Senckenberg Museum, GermanyCarlos Leiva, Spanish National Research Council (CSIC), Spain
Copyright © 2025 Sands, O’Hara, Guzzi, Goodall-Copestake, Convey, Narayanaswamy, Martín-Ledo and Stöhr. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Chester J. Sands, Y2pzYW5AYmFzLmFjLnVr