 M. Virginia Martín1*
M. Virginia Martín1* Ismael Hachero-Cruzado2,3
Ismael Hachero-Cruzado2,3 Juan Carlos Navarro4
Juan Carlos Navarro4 M. Jesús Lago1
M. Jesús Lago1 Eduardo Almansa1
Eduardo Almansa1 Camino Gestal5
Camino Gestal5 Inmaculada Varó4*
Inmaculada Varó4*- 1Centro Oceanográfico de Canarias, Instituto Español de Oceanografía (COC-IEO), CSIC, Santa Cruz de Tenerife, Spain
- 2Centro Oceanográfico de Vigo, Instituto Español de Oceanografía (COV-IEO), CSIC, Vigo, Spain
- 3IFAPA El Toruño, Junta de Andalucía, El Puerto de Santa María, Cádiz, Spain
- 4Instituto de Acuicultura Torre de la Sal (IATS-CSIC), Ribera de Cabanes, Castellón, Spain
- 5Instituto de Investigaciones Mariñas (IIM-CSIC), Vigo, Spain
The aquaculture of Octopus vulgaris is gaining interest due to its high demand, rapid growth, and nutritional value. However, significant challenges remain, particularly in rearing paralarvae to juveniles, mainly due to their specific nutritional needs. This study evaluates the impact of two bioactive compounds— Hytolive® syrup (natural olive fruit extract rich in hydroxytyrosol) and black cumin seed oil (rich in thymoquinone)— using Artemia as a vector, on growth performance, fatty acid profiles, antioxidant responses and lipid peroxidation in O. vulgaris paralarvae from 0 to 30 days post-hatch (dph). Paralarvae were fed one of three diets: control diet (Artemia metanauplii enriched with marine lecithin, CON), control diet supplemented with 10% of Hytolive® syrup (HYT), or control diet supplemented with 30% black cumin seed oil (BCO), with three replicates per treatment. While no significant differences were observed in specific growth rates (SGR) or survival, the HYT and BCO groups exhibited reduced lipid peroxidation at 30 dph, indicating enhanced antioxidant defenses. Additionally, bioactive supplementations altered fatty acid profiles, increasing polyunsaturated fatty acids compared to the control. Although enzymatic antioxidant responses such as catalase (CAT) and superoxide dismutase (SOD) were unaffected, glutathione S-transferase (GST) activity significantly decreased in the BCO group, suggesting a reduced need for detoxification due to lower oxidative stress. This study is the first to demonstrate the transfer of hydroxytyrosol and thymoquinone metabolites from Artemia to O. vulgaris paralarvae and their potential to mitigate oxidative stress. These findings highlight the potential benefits of bioactive-enriched diets in octopus aquaculture and their protective effects during early development. Future research should explore optimal dosages and synergies between bioactive compounds to enhance the immune system and overall health of this species.
1 Introduction
Aquaculture of the common octopus, Octopus vulgaris, has attracted considerable interest in recent years due to the increasing demand for this species in global seafood markets, as well as its rapid growth and high nutritional quality. Despite substantial progress, there are still numerous challenges that need to be addressed for the successful farming of this species. Rearing paralarvae to juvenile stages is one of the most challenging aspects due to the massive mortality during the first life stage, which is the major bottleneck for commercial production, among other reasons, due to their specific nutritional requirements. Several research institutions have worked to improve the breeding, rearing, and feeding techniques of O. vulgaris in captivity, which has allowed significant advances in the field of nutrition and culture technology, resulting in a culture protocol that allows for improved growth and survival of the paralarvae (Tur et al., 2020; Varó et al., 2022). However, there are still numerous aspects that need to be improved, the main one being the reliance on live preys to feed the paralarvae. The impossibility of obtaining these unsustainable preys on a commercial scale, together with the difficulty of rearing them on an experimental scale, has led to the search for other alternatives for feeding paralarvae. The use of Artemia is considered the only viable alternative due to its ease of production and availability. However, its use has often resulted in poor performance, inconsistent growth, and a lack of settlement. Several studies have shown that the Artemia enrichment with long-chain polyunsaturated fatty acids (LC-PUFA) and marine phospholipids can improve the growth, survival and overall wellbeing of paralarvae (Garrido et al., 2018). This enhancement is likely attributable to the increased levels of docosahexaenoic acid (DHA) and polar lipids, both of which play essential roles in the paralarval physiology (Garrido et al., 2018). Despite these improvements, the effectiveness of Artemia as the exclusive diet for paralarvae remains limited, highlighting the need to explore additional enrichment strategies. In this regard, the integration of health-enhancing additives offers a promising opportunity to enhance the nutritional profile of paralarval diets. Consequently, we are considering the enrichment of Artemia with natural bioactive compounds as a promising approach to improve the survival and welfare of paralarvae.
Numerous studies have reported that plant extracts promote diverse biological effects in aquaculture species, including anti-stress activity, growth promotion, appetite stimulation, enhancement of tonicity, immunostimulation, maturation, and anti-pathogen properties in marine organisms, such as fish, shrimp and mollusc (Tadese et al., 2021). The plant extracts that have been tested on finfish and shrimp are numerous. Among them, black cumin seed oil, derived from Nigella sativa, is one of the promising plant-based compounds that has garnered attention in aquaculture in recent years. This oil is recognized for its potential health benefits for cultured organisms, as it may promote the health and growth of aquatic species (Abd El-Hack et al., 2021). It is rich in thymoquinone, thymol, and carvacrol, which are substantial pharmacologically active compounds (Al-Saleh et al., 2006). Thymoquinone accounts for 18.4–24% of the essential oil and is responsible for its antioxidant, anti-inflammatory and antibacterial activities. The strong antioxidant properties of thymoquinone have been shown to reduce oxidative stress and enhance the immunity in fish and shellfish (Latif et al., 2020, 2021; Nur et al., 2020; Mahboub et al., 2022; Niroomand et al., 2024). Some studies also suggest that the inclusion of black cumin oil into the diet may improve feed conversion efficiency, leading to better growth performance (Öz et al., 2018). Overall, research suggests that black cumin oil can be an effective natural supplement in aquaculture to reduce oxidative stress, improve welfare, and enhance growth performance.
On the other hand, olive fruit extract, which contains a high concentration of polyphenols such as hydroxytyrosol and tyrosol, is known for its strong antioxidant properties in fish and crustaceans, helping to neutralize free radicals and reduce oxidative stress and enhance the immune system (Dong et al., 2022a; Hazreen-Nita et al., 2022). Several studies have shown that the inclusion of hydroxytyrosol-rich olive extract can lead to improved growth capacity of the musculoskeletal system and feed utilization (Balbuena-Pecino et al., 2022, 2024; Xia et al., 2024), reduced fat mass by decreasing adipocyte size (Lutfi et al., 2017), reduced hepatic fat accumulation (Dong et al., 2020), attenuated oxidative stress, apoptosis, inflammation and mitochondrial dysfunction (Dong et al., 2022a, b). In addition, olive extract has been shown to be an effective antioxidant during Artemia enrichment, particularly due to its high hydroxytyrosol content (Viciano et al., 2017). Such an antioxidant protective role is reflected in the lipid profile of Artemia, as the addition of hydroxytyrosol preserved the DHA content of enriched metanauplii, potentially improving the nutritional profile, and therefore the health and growth of aquatic species that rely on Artemia as a food source (Viciano et al., 2017).
To the best of our knowledge, these bioactive compounds have not previously been utilized in cephalopod culture. Nevertheless, existing literature suggests that their incorporation into Artemia enrichment protocols may contribute to maintaining the nutritional quality of this live prey and reduce oxidative stress. This, in turn, could potentially improve growth, enhance antioxidant defense mechanisms, and support overall health and survival of octopus paralarvae. Therefore, the aim of this study was to evaluate the effect of black cumin seed oil and Hytolive® syrup (the natural extract obtained from the olive), containing natural bioactive compounds, using Artemia as a vector, on growth performance, fatty acid profiles, antioxidant activities and lipid peroxidation in O. vulgaris paralarvae.
2 Materials and methods
2.1 Ethical statement
All experimental work was performed according to Spanish Law 53/2013 based on the European Union directive on animal welfare (Directive 2010/63/EU) on the protection of animals used for scientific purposes and upon approval by the Bioethics Committee of the Competent Authority (CEIBA 1377-2023).
2.2 Adult broodstock and obtaining paralarvae
Octopus vulgaris broodstock individuals were caught from local fisheries in the Ría de Vigo (NW Spain) using artisanal octopus traps. Individuals were kept in 8000 L tanks of a flow-through seawater system, under natural photoperiod (14L:10D), water temperature of 19-21°C and salinity of 35 PSU. Four octopuses with similar mean wet weight (MWW) were placed in each tank, establishing a sex ratio of three females per male (3:1). Individuals were fed ad libitum on crab and frozen fish. The presence of eggs was verified once a week, and when egg masses were observed in the pot, the ovate female was left alone in the tank by removing the other octopuses.
2.3 Paralarvae rearing and sampling
Octopus hatchlings, with a mean dry weight (DW) of 0.38 ± 0.03 mg, were reared following the guidelines of Iglesias and Fuentes (2014) and Almansa et al. (2025). 100 L black fiberglass cylinder conical tanks with a flow-through seawater system were used at a density of 5 paralarvae L-1 for 30 days (500 paralarvae per tank). A light intensity of 500 lx and a photoperiod of 14L:10D were maintained throughout the experiment. The green water technique was applied to all rearing tanks, daily adding 100,000 cells ml-1 of Isochrysis aff. galbana (T-Iso) and 200,000 cells ml-1 of Nannochloropsis gaditana to each tank, before lights were turned on. The mean water temperature of the rearing tanks was 21°C, salinity was 35 PSU, and dissolved oxygen was near saturation.
Paralarvae were fed one of three experimental diets: a control diet (CON), the control diet supplemented with Hytolive® syrup (Genosa I+D S.I., Málaga, Spain), a natural olive extract rich in hydroxytyrosol (HYT), or the control diet supplemented with black cumin seed oil (Esential’arôms, Spain) (BCO), with three replicates per treatment (9 tanks).
Artemia cysts (Sep-Art AF; INVE Aquaculture, Dendermonde, Belgium) were hatched under standard conditions. Two prey sizes were used throughout the experimental period: nauplii (1 day old) were provide from day 0 to 15, and metanauplii (4 days old) from day 16 to 30.
Newly hatched nauplii were first fed for 20 hours with T-Iso (10 nauplii· ml-1 and 1·106 cell· ml-1), according to Iglesias and Fuentes (2014), and then enriched for 4 h with Marine Lecithin LC 60® (PhosphoTech Laboratoires, Saint Herblain, France) at a concentration of 0.6 g L-1 and 150 nauplii·ml-1, according to Garrido et al. (2017). Metanauplii were obtained by growing nauplii for four days after hatching with T-Iso as food (5 metanauplii· ml-1 and 4·105 cell·ml-1) and then enriched with marine phospholipids (LC60®) for 4h (120 metanauplii·ml-1 and 0.6 g·L-1). This enrichment strategy was selected based on previous studies which showed that LC60 improve paralarvae growth, probably due to its higher levels of DHA and polar lipids in LC60 enriched Artemia (Garrido et al., 2017; Guinot et al., 2013a, 2013b).
The Control group (CON) was fed with Artemia enriched only with LC60®. The HYT group was fed with LC60-enriched Artemia supplemented with Hytolive® syrup, while the BCO group was fed to LC60-enriched Artemia supplemented with black cumin seed oil. Preliminary trials were conducted to determine the most suitable dietary supplementation levels of these bioactive compounds for paralarvae, given the limited literature available on this subject. For Hytolive®, supplementation levels of 5% and 10% were tested, while for black cumin seed oil, levels of 10%, 20% and 30% were assessed based on previous studies (Guinot et al., 2013a, 2013b). In these preliminary trials, the survival of Artemia at different supplementation levels, as well as lipid peroxidation levels, were assessed. Based on the results obtained, supplementation levels of 10% for Hytolive® and 30% for black cumin oil were selected. These bioactive compounds were added during the LC60 enrichment phase. All Artemia enrichments were carried out in the dark at 4°C with gentle aeration. Prior to their introduction into the culture tanks, Artemia were subject to a disinfection protocol with hydrogen peroxide to minimize bacterial contamination. The treatment consisted of a bath with 8,000 ppm for 5 minutes, with intensive aeration. Afterward, Artemia were thoroughly rinsed with seawater for 10 minutes to remove any residual disinfectant before being transferred to the supply tank.
Paralarvae were fed four times daily (9:00, 13:00, 17:00 and 20:00 h) with enriched Artemia nauplii at a density of 0.5 nauplii·mL-1 from day 0 to 15, and with Artemia metanauplii at a density of 0.15 metanauplii·ml-1 from day 16 to 30.
Samples of octopus paralarvae were collected from each tank at 10-, 20- and 30-days post-hatchling. Specimens were anaesthetized using cold seawater (lower than 2°C), rinsed with distilled water and immediately euthanized by freezing in liquid nitrogen. Samples for biochemical analysis including hydroxytyrosol/tyrosol, quinones/thymoquinones, lipid composition, oxidative damage and antioxidant responses were stored at −80 °C until analysis.
2.4 Growth performance
Growth of paralarvae at 10, 20 and 30 days post hatchling (dph) and survival rate at 30 days from each tank were determined as follows: (a) dry weight (DW; mg) after oven drying for 24 h at 110°C; (b) specific growth rate (SGR; % DW day-1) = (lnW2 − lnW1) x t-1 x 100 where, W1 and W2 are the initial and final mean dry weight of each rearing tank, respectively. Ln is the natural logarithm, t the number of days of the experiment and (c) Survival rate (SR; %) = 100 x Nf x Ni-1, where Nf and Ni are the final and the initial number of paralarvae in the rearing tank of each dietary treatment, respectively.
2.5 Determination of hydroxytyrosol/tyrosol and quinones/thymoquinones
The presence of antioxidant compounds characteristic of olive fruit (hydroxytyrosol/tyrosol) and black cumin oil (quinones/thymoquinones) was analysed in both Artemia and paralarvae samples using mass spectrometry at the Central Service for Experimental Research (SCSIE) at the “Universitat de València”.
Samples (≈ 50 mg w.w.) for phenolic compounds analysis were homogenized with a polytron blender with 0.5 ml of methanol (HPLC grade), sonicated for 2 min, and then incubated at 60°C in a thermoblock at 400 rpm for 60 min. Afterwards, the samples were centrifuged at 2500 g for 10 min and filtered (0.22 μm) before analyses. HPLC analyses were carried out on an Agilent 1260 Infinity instrument (Agilent, Waldbronn, Germany). The chromatography was performed on an analytical column (Waters UPLC C18 column 1.7 μm (2.1 × 50 mm)). The mobile phase consisted of solvent A (water; 0.1% formic acid) and solvent B (methanol; 0.1% formic acid). Separation was carried out in 25 min under the following conditions: 0–5 min, 30% B; 12–18 min, 95% B; 18.5 min, 30% B; 18.5–25 min, 30% B. The injection volume and flow rate were 0.4 ml/min and 5 μL, respectively, and the column temperature was maintained at 35 °C. A Triple TOF™ 5600 system with a DuoSprayTM source operating in the negative ESI mode was used for detection (AB SCIEX, CA, USA). The following parameter settings were used: ion spray voltage, -4500 kV; ion source heater, 450 °C; curtain gas, 25 psi; ion source gas 1, 50 psi; and ion source gas 2, 50 psi; decluttering potential -150 v. For the full MS-IDA (information dependent acquisition) analysis and the MS/MS, experiments were run in negative mode with a scan range from m/z 100 to m/z 1200 and a 250-ms accumulation time for the Full MS and a 100-ms accumulation time for the MS/MS. The CE was 30V and in the MS/MS experiment. The IDA was used to trigger the acquisition of MS/MS spectra for ions matching the IDA criteria. Automated calibration was performed using an external calibrant delivery system (CDS), which infuses calibration solution before sample introduction. Data was qualitatively evaluated using the PeakViewTM software. The quantification of hydroxytyrosol and tyrosol was performed by an external calibration curve using resveratrol (R5010 Sigma-Aldrich) as a standard.
Samples (≈ 50 mg w.w.) for quinones/thymoquinones analysis were homogenized with 1 ml hexane (HPLC grade) using a polytron blender, sonicated for 2 min and vortexed, and then incubated at room temperature in a thermoblock at 150 rpm for 3.5 h. Samples were centrifuged at 1000 g for 15 min, filtered (0.22 μm), and stored at -20°C until analysis. The compounds were analysed by Gas Chromatography- Mass Spectrometry (GC-MS) in an Agilent 5977 A MSD system and an HP-5MS UI (Agilent) (30m x 0.25 mm, 0.25 µm film thickness), using helium as carrier. GC oven temperature was 60°C for 5min, increased to 300 °C at 10°C/min, for 10 min, split flow was adjusted at 1mL/min, and injector temperature was at 250 °C. Mass spectra were recorded at 70 eV. Mass range was from m/z 30 to 650. Computer matching against commercial (Nist, Nist_msms, mainlib, replib, wiley7n) libraries was also used for the identification.
2.6 Lipid composition
Fatty acid composition of paralarvae at 10, 20 and 30 dph were analysed. Total lipids were extracted (Folch et al., 1957), measured gravimetrically (XS105 Mettler Toledo, Switzerland), and stored in chloroform: methanol (2:1; v/v) containing 0.01% butylated hydroxytoluene (BHT) at -20°C until further use. Aliquots of total lipids were subjected to acid catalyzed transmethylation for 16 h at 50°C using 1 mL toluene and 2 mL of 1% (v/v) sulphuric acid in methanol (Christie, 2003). Fatty acid methyl esters (FAME) were extracted with hexane:diethyl ether (1:1; v/v) containing 0.01% BHT and purified by thin-layer chromatography (Silica gel G60, 20 x 20 cm glass plates, Merck, Darmstadt, Germany) using hexane:diethyl ether:acetic acid (85:15:1.5; v/v/v) as a solvent system. Fatty acid methyl esters were then analysed with a Thermo Trace GC Ultra Gas Chromatograph (Thermo Electron Corporation, Waltham, MA, USA), equipped with a fused silica 30 m × 0.25 mm open tubular column (Tracer, TR-WAX, film thickness: 0.25 mm, Teknokroma, Sant-Cugat del Valles, Spain), fitted with an on-column injection system, using helium as a carrier gas, and a flame ionization detector (FID). The analytical temperature was programmed from 50°C to 220°C Peaks were recorded using the Azur software package (version 4.0.2.0. Datalys, France). Individual FAME were identified by comparison with known standards: FAME 37 standard mix (Supelco, Bellefonte, PA, USA) and a well characterized sardine oil (Marinol, Fishing Industry Research Institute, Rosebank, South Africa). The relative amount of each FA was expressed as a percentage of the total amount of FA.
2.7 Antioxidant responses and lipid peroxidation
For the analysis of antioxidant enzymes and lipid peroxidation, pools of 15 paralarvae per tank (three replicates from each treatment at 10-, 20- and 30-days post-hatchling) were collected. Samples were diluted and homogenized in ice-cold 20 mM (w/v) Tris-Cl buffer (pH 7.6) with protease inhibitors (Complete®, Sigma, Madrid, Spain). Homogenates were centrifuged at 15,000 × g for 30 min at 4°C, and supernatants were used for enzymatic assays. All enzymatic assays were performed using a BioTek Synergy HT microplate reader (Winooski, Vermont, USA) and were run in triplicate.
Superoxide dismutase (SOD, EC 1.15.1.1) activity was measured by the Marklund and Marklund (1974) method adapted to a microplate using 30 mM pyrogallol as substrate and measure absorbance at 420 nm (Mesa-Herrera et al., 2019). Catalase (CAT, EC 1.11.1.6) activity was quantified by monitoring the rate of hydrogen peroxide (H2O2) decomposition over time by 240 nm absorbance (Sani et al., 2006). One unit of enzyme activity (U) of CAT and SOD was defined as the amount of enzyme required to transform 1 µmol of substrate per minute (µmol min-1) under the assay conditions. Glutathione S-transferase (GST, EC 2.5.1.18) was determined spectrophotometrically at 340 nm following the protocol of Habdous et al. (2002), and one unit of enzyme activity (U) was defined as the amount of enzyme required to transform 1 nmol of substrate per minute (nmol min-1) under the assay conditions). Lipid peroxidation was quantified by determining the content of malondialdehyde (MDA), evaluated by thiobarbituric acid-reactive substances (TBARS) assay (Ohkawa et al., 1979) and the samples were measured fluorometrically. All enzyme assays were carried out at 25°C. Total soluble protein contents of samples were determined using Bradford Bio-Rad Protein Assay, and the absorbance was read at 595 nm. The results for enzymatic activities were expressed as U mg prot-1, and for lipid peroxidation as nmol MDA mg prot-1.
2.8 Statistical analysis
Results are presented as means ± standard deviation (S.D.) after the analysis of triplicate samples. The data were checked for normality and homogeneity of variances by Kolmogorov–Smirnoff and Levene tests, respectively. when necessary, arcsine transformation was performed. A nested two-way ANOVA was applied to DW of the paralarvae, to evaluate inter-tank variability, with treatment, sampling points and tank (as the nested variable) as the factors of variance. The fatty acid profiles of the paralarvae were chemometrically analysed using Principal Component Analysis (PCA) (Nande et al., 2022). A two-way ANOVA was carried out to test for significant differences in oxidative damage and antioxidant responses parameters, with treatment and sampling point as the factors of variance. Also, the effects of three different treatments were analysed by one-way analysis of variance (ANOVA) followed by post hoc Tukey multiple comparison tests. In all statistical tests used, p ≤ 0.05 value was considered statistically significant (95% confidence). The statistical analysis was performed using the IBM SPSS Statistics 25 (SPSS Inc, Chicago, USA).
3 Results
3.1 Growth performance
Data on paralarvae growth performance for each treatment are presented in Table 1. The results showed no significant differences in dry weight (DW) between the CON and HYT groups at 10-, 20-, and 30-days post-hatching (dph) (P > 0.05). However, paralarvae in the BCO group had a lower DW compared to the CON group at 30 dph. The SGR was similar among the three experimental groups, and no significant effects of supplementation were observed (P > 0.05). Similarly, no significant differences in survival (P > 0.05) were detected between the supplemented groups and the control.
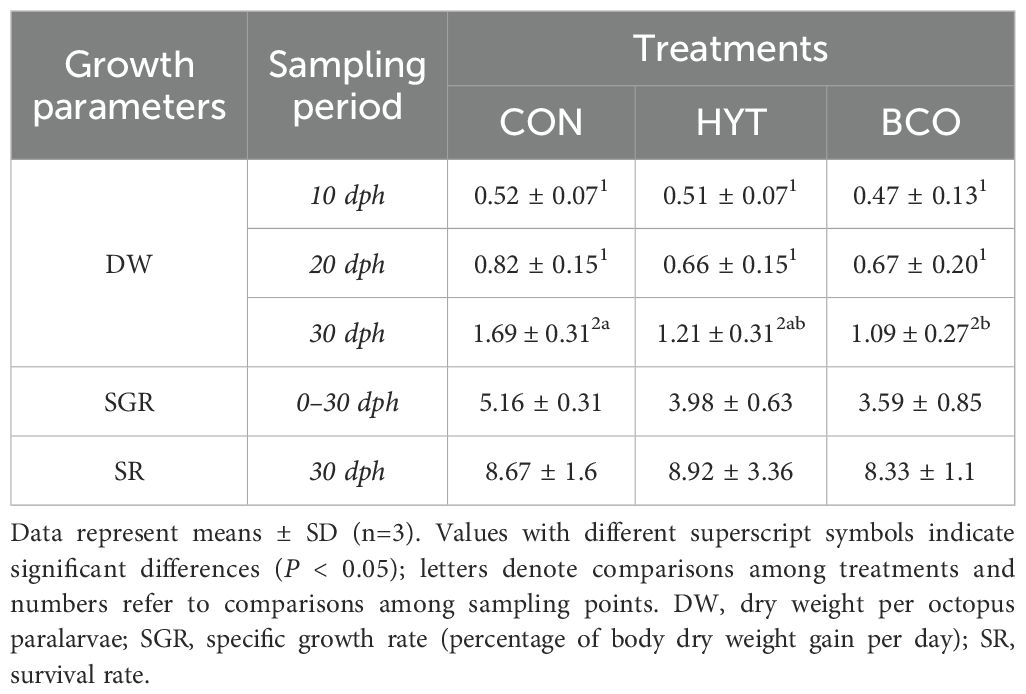
Table 1. Growth performance of Octopus vulgaris paralarvae fed Artemia supplemented with Hytolive (HYT) or black cumin oil (BCO).
3.2 Analysis of hydroxytyrosol/tyrosol and quinones/thymoquinones
The polyphenols detected in paralarvae at 10, 20, and 30 dph were analysed in two groups: the control group (CON), fed LC60-enriched Artemia, and the Hytolive group (HTY), fed LC60-enriched Artemia supplemented with Hytolive. The results are presented in Supplementary Tables S1, S2, respectively. Supplementary Table S3 details the polyphenols found in LC60-enriched Artemia supplemented with Hytolive. Notably, hydroxytyrosol and tyrosol were detected only in LC60-enriched Artemia supplemented with Hytolive (after 4 hours of enrichment) and not in the paralarvae from the HTY group. The mean concentrations of hydroxytyrosol and tyrosol in the Artemia samples supplemented with Hytolive were 92.4 and 61.1 ppb (µg/g), respectively.
Also, chemical compounds detected by GC-MS in paralarvae at 10, 20, and 30 dph were analysed in two groups: the CON group and the black cumin oil group (BCO), the latter fed LC60-enriched Artemia supplemented with black cumin seed oil. The results are summarized in Supplementary Tables S4, S5, respectively. Supplementary Tables S6 and S7 also detail the compounds identified in LC60-enriched Artemia supplemented with black cumin seed oil and the commercial black cumin seed oil itself. Thymoquinone was detected exclusively in the black cumin seed oil samples, indicating that it is completely metabolized in the digestive tract of the Artemia.
3.3 Lipid composition
The total lipids and fatty acids of the paralarvae from the distinct experimental groups are shown in Supplementary Table S8. The fatty acid profiles of the paralarvae from each treatment were statistically analysed using principal component analysis (PCA; Figure 1). The first component accounted for 31.4% of the variance, while the second accounted for 19.4%. The loading plot shows that variables corresponding to the main long-chain polyunsaturated fatty acids (PUFA) n-3 (20:3n-3, 20:4n-3, 20:5n-3, 22:6n-3) were associated with the first component and were inversely related to shorter C14 (14:0, 14:1) and C15 fatty acids, 16:3, and 22:5n-3. The second component is mainly distinguished based on saturated fatty acids like 16:0 and 18:0, as well as the monounsaturated 20:1n-9 loading on the positive side against 20:0 on the negative side. The score plot revealed that the profiles of the paralarvae corresponding to the control group (CON) appeared segregated from the other two dietary groups along the second component. According to age, the profiles of the 10 dph paralarvae tended to segregate from the rest along the first component, showing an age-related pattern, with the profile of the 10 dph paralarvae (more saturated) further separated from those at 20 and 30 dph. The age separation was more evident within the control group, although, in general, scores were dispersed along the axes.
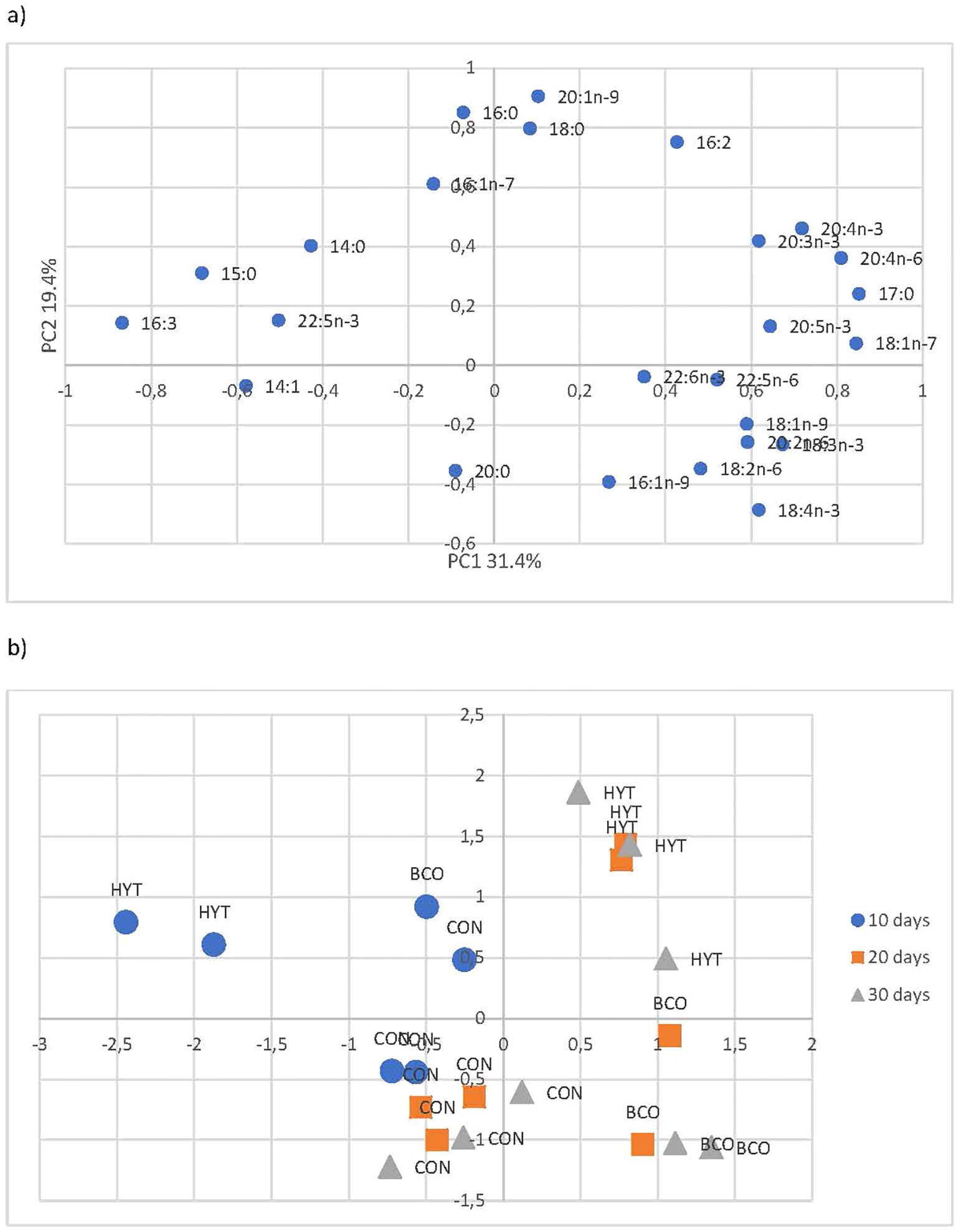
Figure 1. Loading plot (a) and score plot (b) of the principal component analysis (PCA) performed on the fatty acid profiles of the paralarvae of the three experimental groups.
3.4 Antioxidant status and lipid peroxidation
The effects of dietary supplementation with BCO and HYT on the activities of antioxidant response enzymes in 10, 20 and 30 dph paralarvae are shown in Figures 2-4, as well as in Table 2. No significant effects of supplementation on CAT and SOD activities were observed (P > 0.05), nor were any significant changes detected over time (P > 0.05) (Figures 2, 3; Table 2). However, GST activities were significantly reduced in PL supplemented with BCO at both 20 and 30 dph when compared to the control group (11.2 ± 1.4 vs 14.8 ± 1.0; 12.1 ± 1.9 vs 16.8 ± 2.4 U mg prot-1, P < 0.05) (Figure 4). Additionally, a significant increase in GST activity was detected with the development of paralarvae, with significantly higher values at 30 dph compared to 10 dph for the CON and HYT groups (P < 0.05). However, this increase was not observed in the BCO group.
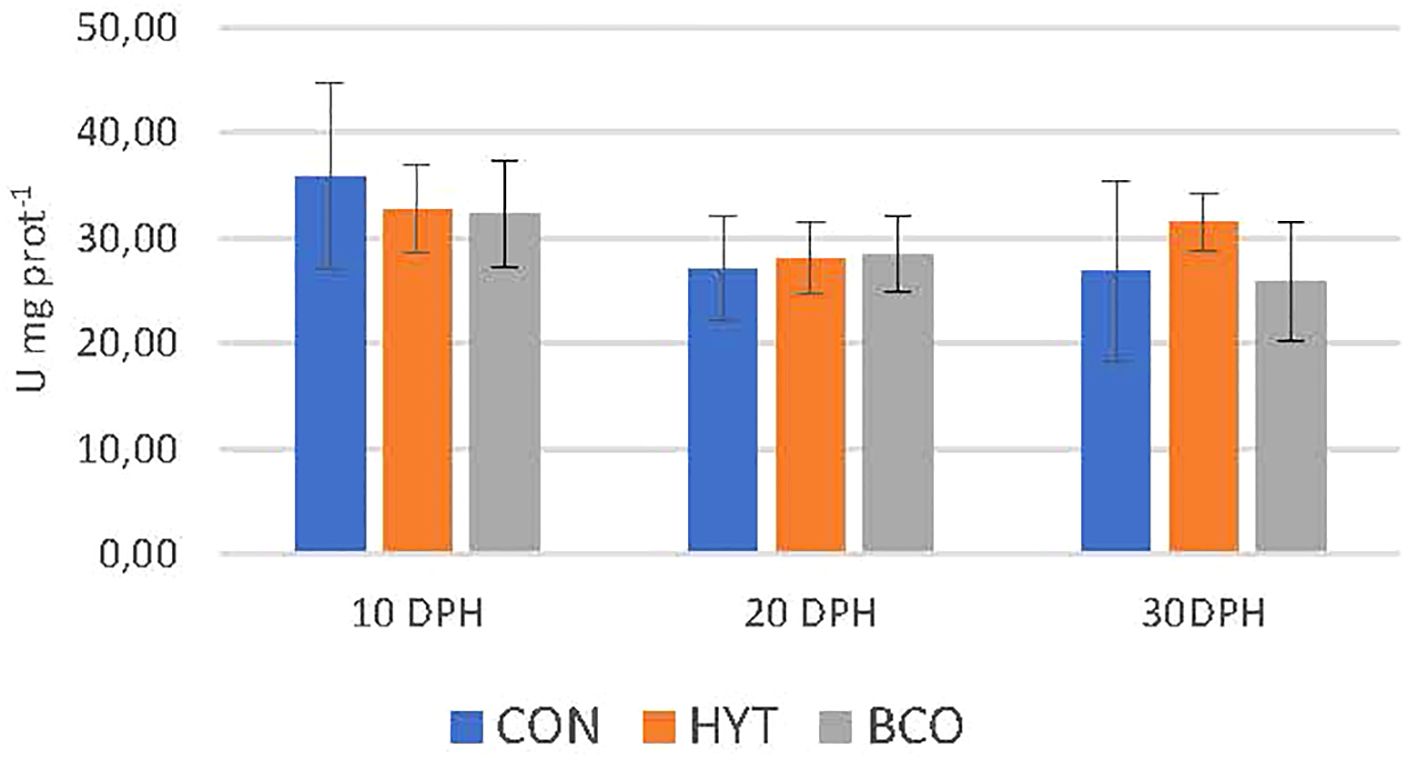
Figure 2. Catalase activity (CAT, U mg prot-1) of 10, 20 and 30 dph paralarvae fed Artemia supplemented with Hytolive (HYT) or black cumin oil (BCO). Values are mean ± SD (n=3).
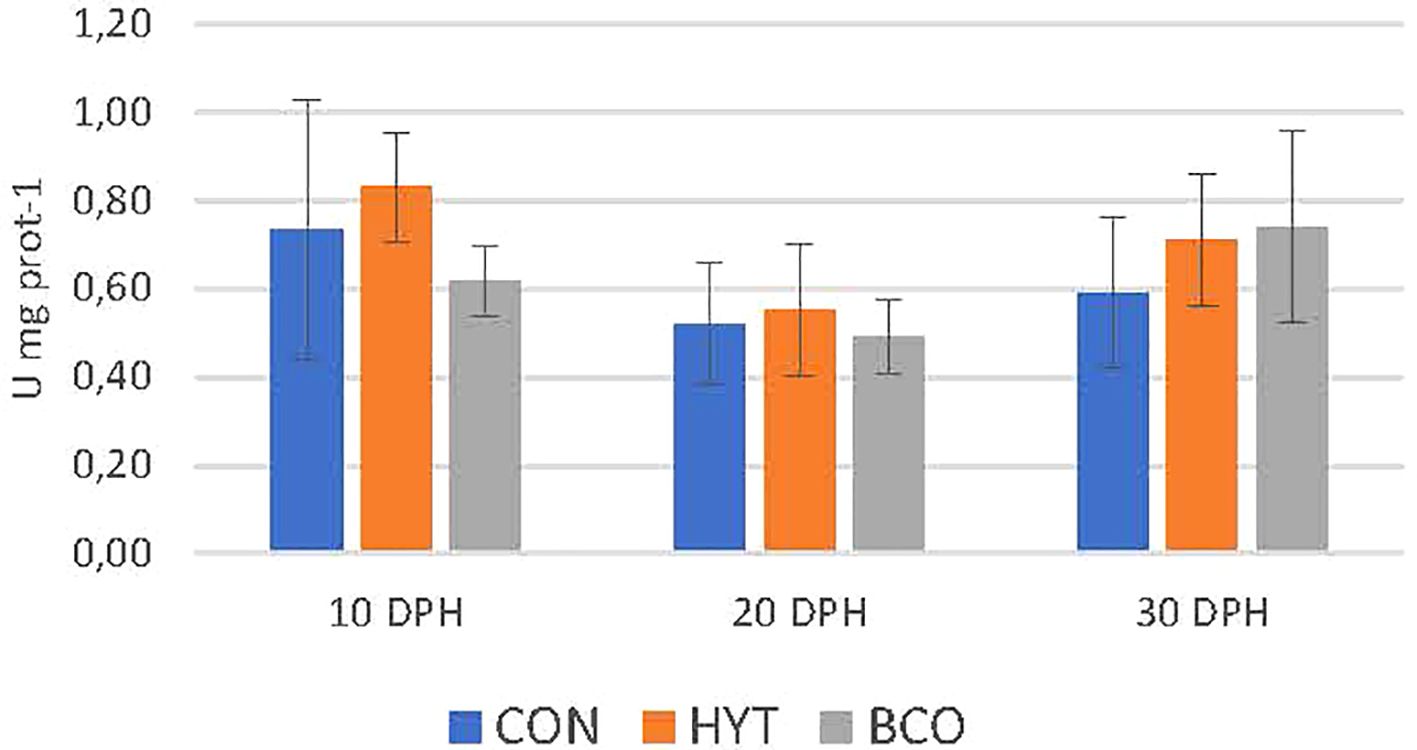
Figure 3. Superoxide dismutase activity (SOD, U mg prot-1) of 10, 20 and 30 dph paralarvae fed Artemia supplemented with Hytolive (HYT) or black cumin oil (BCO). Values are mean ± SD (n=3).
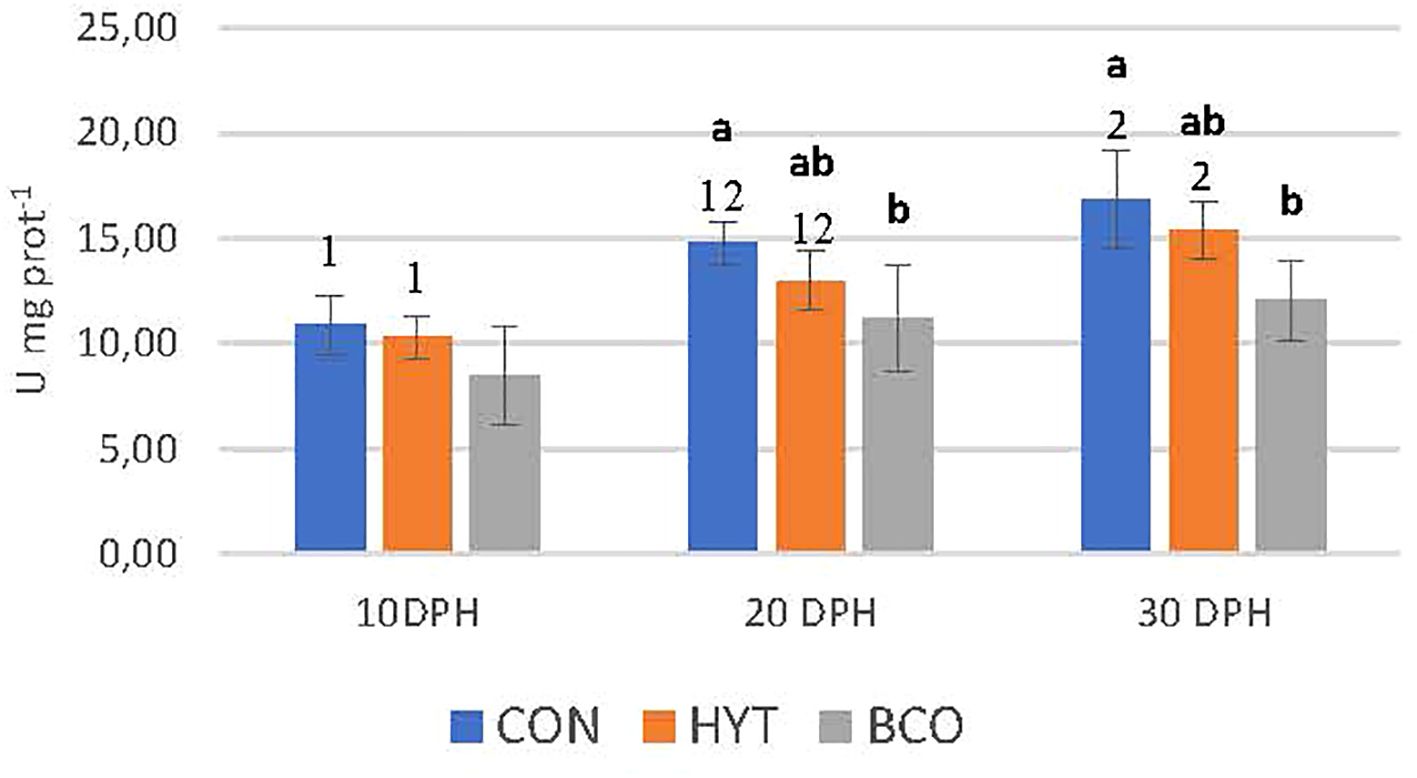
Figure 4. Glutathione S-transferase activity (GST, U mg prot-1) of 10, 20 and 30 dph paralarvae fed Artemia supplemented with Hytolive (HYT) or black cumin oil (BCO). Values are mean ± SD (n=3). Different letters superscripts above the histogram bars indicate statistically significant differences attributed to experimental treatments, whereas different numbers denote significant differences between sampling points (P<0.05).
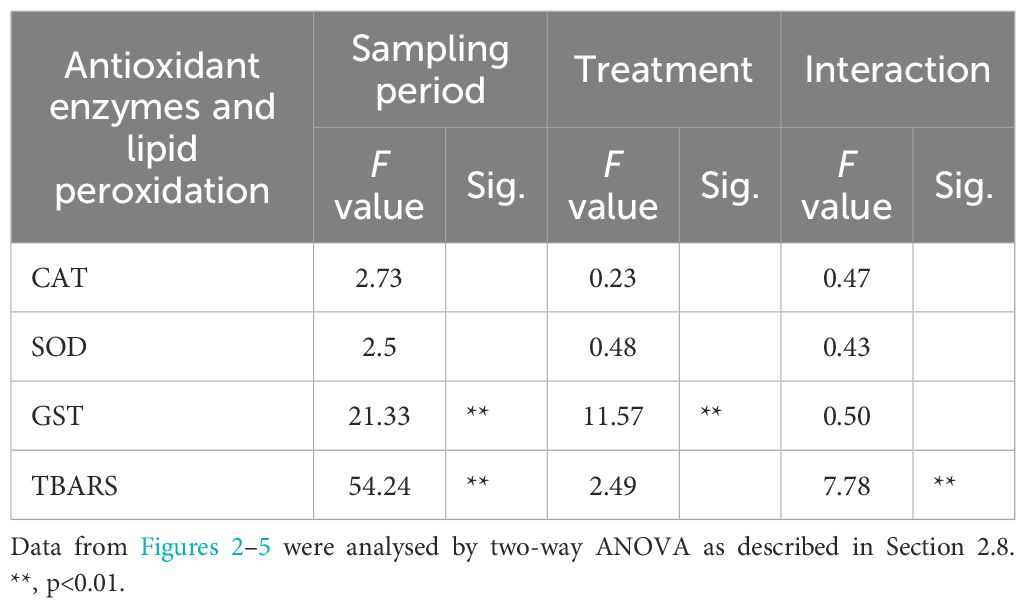
Table 2. Significance of the effects of the sampling period and treatment on oxidative damage and antioxidant responses parameters.
Figure 5 shows the levels of lipid peroxidation products, measured as MDA, in response to the experimental treatments. An increase in MDA levels was detected as the paralarvae developed across all experimental groups, with significantly higher levels recorded at 30 dph compared to 10 dph (Anova P<0.05). However, at 30 dph-paralarvae, the effects of the bioactive compounds became evident, as evidenced by significantly lower TBARS values in paralarvae supplemented with HYT and BCO compared to the control group (0.7 ± 0.2; 0.7 ± 0.1 vs. 1.2 ± 0.2 nmol MDA mg prot-1, P<0.05). Additionally, the two-way ANOVA revealed a highly significant interaction between the sampling period and treatment (P < 0.01) (Table 2).
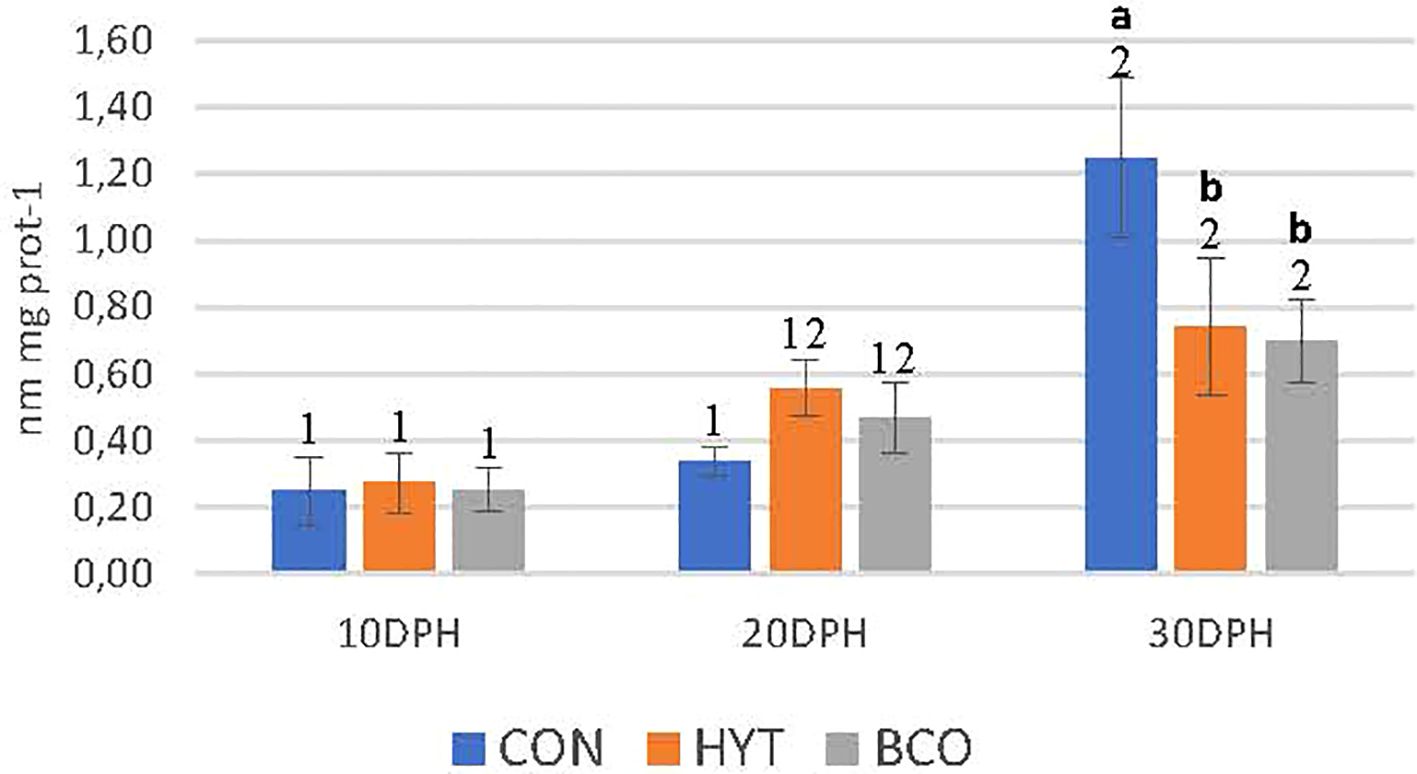
Figure 5. Lipid peroxidation levels (nmol MDA mg-1 protein) in paralarvae at 10, 20, and 30 days post-hatching (dph) that were fed Artemia supplemented with Hytolive (HYT) or black cumin oil (BCO). Values are mean ± SD (n=3). Different letters superscripts above the histogram bars indicate statistically significant differences attributed to experimental treatments, whereas different numbers denote significant differences between sampling points (P<0.05).
4 Discussion
Despite the extensive research efforts in common octopus aquaculture, the culture of the first developmental stages (paralarva) has remained a significant challenge for commercial production. Feeding and nutrition are key factors for understanding its needs under culture conditions. It has been demonstrated that the administration of nutritional supplements, particularly natural bioactive compounds, is an unequivocally effective strategy for enhancing larval survival in aquaculture (Reverter et al., 2014). Medicinal plants have potential as growth promoters, immunomodulators and antioxidants in aquaculture. The beneficial properties of hydroxytyrosol-rich extracts or black seed oil have been extensively studied in mammals and have recently been investigated in fish.
It is well established that hydroxytyrosol and tyrosol do not accumulate in the organism but are rapidly excreted (López de las Hazas et al., 2016). However, the detection of hydroxytyrosol and tyrosol in Artemia supplemented with these compounds suggests that they exhibit some degree of stability during their passage through Artemia’s metanauplii digestive tract. Our results indicate that, after 4 hours of enrichment with LC60 and Hytolive, these compounds are not fully metabolized or excreted. This is the first experimental demonstration that Artemia can serve as a vehicle to deliver these bioactive compounds to paralarvae.
In paralarvae fed Artemia supplemented with HYT, it was found that hydroxytyrosol and tyrosol were fully metabolized. Nevertheless, secondary metabolites such as homovanillyl alcohol, caffeic acid (both cis- and trans-), p-coumaric acid (both cis- and trans-) and cinnamic acid were detected. It is important to note that these metabolites are not exclusively derived from the metabolism of hydroxytyrosol and tyrosol, as they could also originate from the metabolism of polyphenols present in the microalga I. galbana, which was used for Artemia growth. This observation is supported by the detection of the same metabolites in the control group of paralarvae (CON). However, HYT supplementation appears to enhance polyphenol availability in the paralarvae, which likely contributes to the observed beneficial effects against oxidation (lower levels of lipid peroxidation) in the HYT group.
In contrast, Artemia supplemented with black cumin oil (BCO) did not retain detectable levels of thymoquinone when analysed by GC-MS, suggesting complete metabolism of the compound within Artemia. Nonetheless, potential thymoquinone-derived metabolites, including 3-pentanone, nonane (3,7-dimethyl), heptane (3,4,5-trimethyl), and 1-dodecanol (2-octyl), were identified. These findings suggest that Artemia can transfer thymoquinone metabolites to paralarvae, which may explain the observed antioxidant effects in the BCO group.
Previous studies have shown that incorporating bioactive olive extracts into the diet can enhance growth in some crustaceans and fish species, such as sea bream (Sparus aurata) (Gisbert et al., 2017), Persian sturgeon (Acipenser persicus) (Hosseinnia et al., 2021), Blunt snout bream (Megalobrama amblycephala) (Dong et al., 2022a) and crayfish (Astacus and Astacus leptodactylus) (Parrillo et al., 2017; Sateriale et al., 2023). However, no previous studies have specifically evaluated the effects of olive extracts on cephalopods, including O. vulgaris. The findings from the current study indicate no significant improvement in growth performance in paralarvae of O. vulgaris supplemented with hydroxytyrosol (HYT) after 30 days of dietary inclusion. No effects on specific growth rate (SGR) or survival were observed. Although it is not possible the comparison with other studies on cephalopods, similar findings have been reported in other fish studies. For instance, rainbow trout (Oncorhynchus mykiss) fed diets with varying levels of olive leaf extract for 60 days showed no significant differences in growth performance values, such as weight gain, SGR, or feed utilization (Baba et al., 2018). Likewise, juvenile sea bream supplemented with a hydroxytyrosol-rich extract showed no changes in body weight or other biometric parameters after 4 or 8 weeks of treatment (Balbuena-Pecino et al., 2022).
Regarding paralarvae supplemented with black cumin oil (BCO), no significant differences in SGR and survival rates were also observed compared to the control group. Particularly, the dry weight (DW) of the BCO group was lower. Similar to HYT, no studies have previously investigated the impact of BCO on growth and survival in O. vulgaris paralarvae or other cephalopods. Using other crustacean and fish studies as a reference, some researchers have shown improved growth rates in BCO-supplemented species, including white leg shrimp (Litopenaeus vannamei) (Asadi et al., 2018), common carp (Cyprinus carpio) (Al-Dubakel et al., 2012), rainbow trout (O. mykiss) (Öz et al., 2018; Bektas et al., 2019), and Asian sea bass (Lates calcarifer) (Abdelwahab and El-Bahr, 2012). However, in other species such as tilapia (Oreochromis niloticus), BCO supplementation had no significant effects on growth or feed intake (Ahmed et al., 2018). In all these studies, supplementation levels varied widely, from 0.1% to 30%.
Considering all the data, the growth performance results suggest that the response to bioactive compounds, in terms of growth and survival, is strongly influenced by the species studied and the concentrations of the bioactive extracts used. The paralarvae of O. vulgaris are notable by their unique physiological traits, which are characterized by a high metabolic rate and specialized lipid metabolism, both of which are essential for supporting their rapid growth during early life stages. These paralarvae exhibit a highly aerobic metabolism, that requires substantial energy for processes such as neurosensory development, active pelagic locomotion, and tissue biosynthesis (Morales et al., 2017). Furthermore, the digestive system of paralarvae at this stage exhibits functional limitations, including limited and specific enzymatic activity with a preference for animal proteins and marine phospholipids, and a low capacity to hydrolyse and absorb complex plant lipids. Since these bioactive products are herbal plants, higher inclusion levels of herbal extracts have been reported to negatively impact the growth of other species due to elevated concentrations of anti-nutritional factors (Hazreen-Nita et al., 2022). Therefore, determining the optimum inclusion level of these bioactive compounds in feed formulation is crucially important. In the present study, the tested concentrations of bioactive extracts had no significant effects on SGR or survival of paralarvae. This could indicate that while growth was not enhanced, the supplementation levels used were not harmful, particularly in terms of anti-nutritional factors.
The fatty acid composition of paralarvae from different treatments showed that, although variable, there was a difference in the profiles of the experimental groups by age, with the 10-dph paralarvae having more saturated fatty acid profiles distinct from those of 20 and 30-dph. The paralarvae supplemented with HYT and BCO showed patterns more linked to polyunsaturated fatty acids as compared to the CON group, which can be associated with a protective effect of both dietary treatments against lipid oxidation (Barata et al., 2005).
In order to gain insight into the potential mechanisms through which diverse natural antioxidants influence the antioxidant status of O. vulgaris paralarvae, this study examined a range of biomarkers of oxidative stress. Lipid peroxidation is a well-established biomarker for assessing oxidative stress-induced damage in animal tissues. Oxidative stress occurs when there is an imbalance between the production of reactive oxygen species (ROS) and the antioxidant defenses (Benedetti et al., 2022). When ROS production exceeds an organism’s ability to scavenge them, oxidative damage to macromolecules such as lipids, proteins, and DNA occurs (Valgimigli, 2023). Membrane lipids are particularly susceptible to ROS attack, resulting in increased lipid peroxidation products. In fact, lipid peroxidation has been extensively employed as a biomarker of oxidative stress (Lushchak, 2011). In our study, dietary supplementation with the bioactive compounds HYT and BCO significantly reduced the level of lipid peroxidation in O. vulgaris paralarvae at 30dph, despite an overall increase in lipid peroxidation observed across all groups during development. This reduction suggests an improvement in antioxidant status in paralarvae supplemented with both bioactive compounds. These results are consistent with previous research on the effects of these bioactives in fish (Latif et al., 2020; Dong et al., 2022a). The bioactive compound thymoquinone present in BCO has been shown to inhibit iron-dependent lipid oxidation via the Fenton reaction in a concentration-dependent manner (Nagi and Mansour, 2000). Similarly, hydroxytyrosol and tyrosol from HYT Hydroxytyrosol and tyrosol from Hytolive® have been described as being able to reduce MDA levels in aquatic organisms (Viciano et al., 2017; Dong et al., 2022a, b).
On the other hand, enzymatic and non-enzymatic antioxidant defenses play a key role in preventing oxidative stress by counteracting the effects of reactive oxygen species (ROS). Aquatic organisms use antioxidant enzymes such as SOD and CAT, along with detoxification enzymes like GST, to mitigate the harmful impact of ROS and prevent oxidative damage (Mates, 2000; Martínez-Álvarez et al., 2005). These antioxidants are configured to maintain low levels of ROS within cells, making them essential components of an organism’s defense response (Mates, 2000).
The response to the inclusion of bioactive compounds may vary between individuals and species, depending on factors such as genetics, previous nutritional status, and environmental conditions. In other studies, carried out with different marine vertebrate and invertebrate species, it has been observed that diets enriched with vegetable oils or natural extracts can induce significant changes in the activity of detoxifying enzymes (Xavier et al., 2021).
The results of the present study did not reveal changes in CAT or SOD levels in paralarvae supplemented with HYT or BCO. These enzymes maintained steady activity levels, possibly reflecting transient rather than sustained responses to supplementation (Barata et al., 2005).
However, GST activities were significantly reduced in BCO-supplemented PL after 20 and 30 days of culture compared to the control group. GST is a crucial enzyme involved in the biotransformation process of metabolites. It catalyzes the conjugation of breakdown products of lipid peroxides to glutathione (GSH), facilitating their excretion and decreasing their toxicity (Hellou et al., 2012). Previous studies have shown that GST activity increases in response to external stressors, particularly those that generate ROS (Barata et al., 2005). This increase in activity suggests a detoxification process where GST conjugates with its natural substrates to eliminate toxic metabolites (Townsend and Tew, 2003). Specifically, GST activity is closely linked to the response to oxidative stress in cephalopods, where increased activity has been observed under stress conditions (Semedo et al., 2012). Bioactive compounds in BCO can help restore redox balance by neutralizing free radicals, which in turn preserves antioxidant enzymes like GST (Masella et al., 2004). This mechanism may explain the significant reduction in GST levels in paralarvae supplemented with BCO compared to the control, as well as the maintenance of GST levels throughout development, in contrast to the other two groups. GSTs are upregulated in response to oxidative stress to detoxify byproducts such as lipid peroxides and 4-hydroxynonenal. When oxidative stress is low, the demand for GST-mediated detoxification decreases, resulting in lower GST activity (Awasthi et al., 2017; Sharma et al., 2004). The reduced GST activity in BCO supplemented PL may therefore be attributed to directly scavenging of reactive oxygen species (ROS), which in turn lowers oxidative stress and reduces the demand for GST activity.
5 Conclusions
To the best of our knowledge, this study is the first to demonstrate that the bioactive compounds hydroxytyrosol (HYT) and black cumin oil (BCO) used as supplements in Artemia are transferred to paralarvae, either directly as a parent compound (in the case of HYT) or as metabolites (in the case of BCO). Notably, paralarvae fed these bioactives exhibited an improved profile of long-chain polyunsaturated fatty acids (PUFA), potentially linked to protective effects against oxidative stress.
Furthermore, paralarvae supplemented with HYT and BCO showed lower lipid peroxidation levels (TBARS) compared to the control group after 30 days of supplementation. Additionally, reduced GST activity was observed in the BCO-supplemented group after 20 days, suggesting a potential modulation of oxidative stress responses.
It is reasonable to assume that bioactives may offer protective benefits during the early stages of development. This fact is of high relevance for future studies on the diet of O. vulgaris due to the high economic interest in this species and the recent advances in its culture. Future studies should focus on establishing the optimal dose of the selected bioactive compounds, testing higher concentrations without compromising the viability of the live prey. Furthermore, the combination of both bioactives should be investigated to decipher if any synergistic effects could boost the immune system of O. vulgaris. Finally, extending the evaluation to subsequent developmental stages would facilitate a more comprehensive understanding of their long-term impact.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Ethics statement
The animal studies were approved by Bioethics Committee of the CSIC (CEIBA 1377-2023). The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent was obtained from the owners for the participation of their animals in this study.
Author contributions
MM: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Supervision, Writing – original draft, Writing – review & editing, Funding acquisition, Project administration, Validation, Visualization. IH-C: Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – review & editing, Resources, Validation. JN: Data curation, Formal analysis, Investigation, Methodology, Validation, Visualization, Writing – review & editing. ML: Data curation, Formal analysis, Methodology, Resources, Visualization, Writing – review & editing. EA: Conceptualization, Funding acquisition, Project administration, Resources, Supervision, Writing – review & editing. CG: Conceptualization, Funding acquisition, Project administration, Supervision, Validation, Writing – review & editing. IV: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This study was supported by OCTOMICS Project (AGL-2017-89475-C2) funded by the Spanish Ministerio de Ciencia e Innovación, and by “ERDF A way of making Europe”. This study was also funded by the programme THINKINAZUL, supported by the Spanish Ministerio de Ciencia e Innovación with funding from European Union NextGenerationEU (PRTR-C17.I1) and Gobierno de Canarias (SD2218/6897). We acknowledge support of the publication fee by the CSIC Open Access Publication Support Initiative (PROA) through its Unit of Information Resources for Research (URICI).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1629293/full#supplementary-material
References
Abd El-Hack M. E., Abdelnour S. A., Khafaga A. F., Taha A. E., and Abdel-Latif H. M. R. (2021). “Nigella sativa seeds and its derivatives in fish feed,” in Black cumin (Nigella sativa) seeds: Chemistry, Technology, Functionality, and Applications. Food Bioactive Ingredients. Ed. Fawzy Ramadan M. (Springer, Cham). doi: 10.1007/978-3-030-48798-0_19
Abdelwahab A. M. and El-Bahr S. M. (2012). Influence of black cumin seeds (Nigella sativa) and turmeric (Curcuma longa Linn.) mixture on performance and serum biochemistry of Asian Sea Bass, Lates calcarifer. World J. Fish Mar. Sci. 4, 496–503. doi: 10.5829/idosi.wjfms.2012.04.05.6478
Ahmed H. A., Abusina G., Elnady A., Abdelrahman H., and Abedo A. E. (2018). Evaluation of substituting black seed meal (Nigella sativa L.) as protein source in Nile tilapia diets. Bioscience Res. 15, 1191–1198.
Al-Dubakel A., Al-Mhawe B., Majeed M., and Shaeyal L. (2012). Preliminary study on the effect of dietary black seed (Nigella sativa) on growth and blood glucose of common carp (Cyprinus carpio) fingerlings. Ira. J. Thi. Qar. Univ. Agricult. Res. 1, 41–50.
Al-Saleh I. A., Billedo G., and El-Doush I. I. (2006). Levels of selenium, dl-α-tocopherol, dl-γ- tocopherol, all-trans-retinol, thymoquinone and thymol in different brands of Nigella sativa seeds. J. Food Composition Anal. 19, 167–175. doi: 10.1016/j.jfca.2005.04.011
Almansa E., Márquez L., Rosas C., Martín M. V., Navarro J. C., Uriarte I., et al. (2025). “Octopods aquaculture: Reproduction, rearing technology, nutritional physiology, welfare and health status,” in “Aquaculture and living resource management, vol. 2. Ed. Pereira L. (CRC Press (Taylor & Francis Group), ISBN: 978-1-032-34632-8.
Asadi M. S., Gharaei A., Mirdar Harijani J., and Arshadi A. (2018). A Comparison between dietary effects of Cuminum cyminum essential oil and Cuminum cyminum essential oil, loaded with iron nanoparticles, on growth performance, immunity and antioxidant indicators of white leg shrimp (Litopenaeus vannamei). Aquacult Nutr. 24, 1466–1473. doi: 10.1111/anu.12683
Awasthi Y., Ramana K., Chaudhary P., Srivastava S., and Awasthi S. (2017). Regulatory roles of glutathione-S-transferases and 4-hydroxynonenal in stress-mediated signalling and toxicity. Free Radical Biol. Med. 111, 235–243. doi: 10.1016/j.freeradbiomed.2016.10.493
Baba E., Acar Ü., Yılmaz S., Zemheri F., and Ergün S. (2018). Dietary olive leaf (Olea europea L.) extract alters some immune gene expression levels and disease resistance to Yersinia ruckeri infection in rainbow trout Oncorhynchus mykiss. Fish shellfish Immunol. 79, 28–33. doi: 10.1016/j.fsi.2018.04.063
Balbuena-Pecino S., Montblanch M., García-Meilán I., Fontanillas R., Gallardo Á., Gutiérrez J., et al. (2022). Hydroxytyrosol-rich extract from olive juice as an additive in gilthead sea bream juveniles fed a high-fat diet: Regulation of somatic growth. Front. Physiol. 13, 966175. doi: 10.3389/fphys.2022.966175
Balbuena-Pecino S., Montblanch M., Rosell-Moll E., González-Fernández V., García-Meilán I., Fontanillas R., et al. (2024). Impact of hydroxytyrosol-rich extract supplementation in a high-fat diet on gilthead sea bream (Sparus aurata) lipid metabolism. Antioxidants 13, 403. doi: 10.3390/antiox13040403
Barata C., Navarro J. C., Varó I., Riva M. C., Arun S., and Porte C. (2005). Changes in antioxidant enzyme activities, fatty acid composition and lipid peroxidation in Daphnia magna during the aging process. Comp. Biochem. Physiology Part B 140, 81–90. doi: 10.1016/j.cbpc.2004.09.025
Bektaş Z. H., Savaşer S., Akçimen U., Ceylan M., Yener O., and Bulut C. (2019). Using of black cumin seed powder (Nigella sativa) as immunostimulant and growth promoter in rainbow trout, Oncorhynchus mykiss (Walbaum). Turkish J. Fisheries Aquat. Sci. 19, 987–999.
Benedetti M., Giuliani M. E., Mezzelani M., Nardi A., Pittura L., Gorbi S., et al. (2022). Emerging environmental stressors and oxidative pathways in marine organisms: Current knowledge on regulation mechanisms and functional effects. Biocell 46, 37. doi: 10.32604/biocell.2022.017507
Dong Y., Li L., Espe M., Lu K., and Rahimnejad S. (2020). Hydroxytyrosol Attenuates Hepatic Fat Accumulation via Activating Mitochondrial Biogenesis and Autophagy through the AMPK Pathway. J. Agric. Food Chem. 68, 9377–9386. doi: 10.1021/acs.jafc.0c03310
Dong Y., Xia T., Yu M., Wang L., Song K., Zhang C., et al. (2022a). Hydroxytyrosol attenuates high-fat-diet-induced oxidative stress, apoptosis and inflammation of blunt snout bream (Megalobrama amblycephala) through its regulation of mitochondrial homeostasis. Fishes 7, 78. doi: 10.3390/fishes7020078
Dong Y., Yu M., Wu Y., Xia T., Wang L., Song K., et al. (2022b). Hydroxytyrosol promotes the mitochondrial function through activating mitophagy. Antioxidants 11, 893. doi: 10.3390/antiox11050893
Folch J., Lees M., and Stanley G. S. (1957). A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 226, 497–509. doi: 10.1016/S0021-9258(18)64849-5
Garrido D., Martín M. V., Rodríguez C., Iglesias J., Navarro J. C., Estévez A., et al. (2018). Meta-analysis approach to the effects of live prey on the growth of Octopus vulgaris paralarvae under culture conditions. Rev. Aquac. 10, 3–14. doi: 10.1111/raq.12142
Garrido D., Varó I., Morales A. E., Hidalgo M. D. C., Navarro J. C., Hontoria F., et al. (2017). Assessment of stress and nutritional biomarkers in cultured Octopus vulgaris paralarvae: effects of geographical origin and dietary regime. Aquaculture 468, 558–568. doi: 10.1016/j.aquaculture.2016.11.023
Gisbert E., Andree K. B., Quintela J. C., Calduch-Giner J. A., Ipharraguerre I. R., and Pérez-Sánchez J. (2017). Olive oil bioactive compounds increase body weight and improve gut health and integrity in gilthead sea bream (Sparus aurata). Br. J. Nutr. 117, 351–363. doi: 10.1017/S0007114517000228
Guinot D., Monroig Ó., Hontoria F., Amat F., Varó I., and Navarro J. C. (2013b). Enriched on-grown Artemia metanauplii actively metabolise highly unsaturated fatty acid-rich phospholipids. Aquaculture 412–413, 173–178. doi: 10.1016/j.aquaculture.2013.07.030
Guinot D., Monroig Ó., Navarro J. C., Varó I., Amat F., and Hontoria F. (2013a). Enrichment of Artemia metanauplii in phospholipids and essential fatty acids as a diet for common octopus (Octopus vulgaris) paralarvae. Aquaculture Nutr. 19, 837–844. doi: 10.1111/anu.12048
Habdous M., Vincent-Viry M., Visvikis S., and Siest G. (2002). Rapid spectrophotometric method for serum glutatione S-transferases activity. Clinica Chimica Acta 326, 131–142. doi: 10.1016/S0009-8981(02)00329-7
Hazreen-Nita M. K., Kari Z. A., Mat K., Rusli N. D., Sukri S. A. M., Harun H. C., et al. (2022). Olive oil by-products in aquafeeds: Opportunities and challenges. Aquaculture Rep. 22, 100998. doi: 10.1016/j.aqrep.2021.100998
Hellou J., Ross N. W., and Moon T. W. (2012). Glutathione, glutathione S-transferase, and glutathione conjugates, complementary markers of oxidative stress in aquatic biota. Environ. Sci. pollut. Res. 19, 2007–2023. doi: 10.1007/s11356-012-0909-x
Hosseinnia E., Khara H., Farokhroz M., Yousefi Jourdehi A., and Kazemi R. (2021). Effects of dietary olive oil and butylated hydroxytoluene (BHT) on growth, blood, and immunity indices in juvenile Persian sturgeon (Acipenser persicus). Iranian J. Fisheries Sci. 20, 810–827.
Iglesias J. and Fuentes L. (2014). Octopus vulgaris. Paralarval culture. Cephalopod culture, 427–450. doi: 10.1007/978-94-017-8648-5
Latif M., Faheem M., Hoseinifar S. H., and Van Doan H. (2020). Dietary black seed effects on growth performance, proximate composition, antioxidant and histo-biochemical parameters of a culturable fish, rohu (Labeo rohita). Animals 11, 48. doi: 10.3390/ani11010048
Latif M., Faheem M., Hoseinifar S. H., and Van Doan H. (2021). Protective efficacy of Nigella sativa seeds against diethyl phthalate induced growth retardation, oxidative stress and histo-biochemical damages in Labeo rohita. Aquaculture 533, 736065. doi: 10.1016/j.aquaculture.2020.736065
López de las Hazas M. C. L., Piñol C., Macià A., Romero M. P., Pedret A., Solà R., et al. (2016). Differential absorption and metabolism of hydroxytyrosol and its precursors oleuropein and secoiridoids. J. Funct. Foods 22, 52–63. doi: 10.1016/j.jff.2016.01.030
Lushchak V. I. (2011). Environmentally induced oxidative stress in aquatic animals. Aquat. Toxicol. 101, 13–30. doi: 10.1016/j.aquatox.2010.10.006
Lutfi E., Babin P. J., Gutierrez J., Capilla E., and Navarro I. (2017). Caffeic acid and hydroxytyrosol have anti-obesogenic properties in zebrafish and rainbow trout models. PloS One 12, e0178833. doi: 10.1371/journal.pone.0178833
Mahboub H. H., Elsheshtawy H. M., Sheraiba N. I., Fahmy E. M., Mohamed E. A., Abdelnaeim N. S., et al. (2022). Dietary black cumin (Nigella sativa) improved hemato-biochemical, oxidative stress, gene expression, and immunological response of Nile tilapia (Oreochromis niloticus) infected by Burkholderia cepacia. Aquaculture Rep. 22, 100943. doi: 10.1016/j.aqrep.2021.100943
Marklund S. and Marklund G. (1974). Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 47, 469–474. doi: 10.1111/j.1432-1033.1974.tb03714.x
Martínez-Álvarez R. M., Morales A. E., and Sanz A. (2005). Antioxidant defenses in fish: biotic and abiotic factors. Rev. Fish Biol. fisheries 15, 75–88. doi: 10.1007/s11160-005-7846-4
Masella R., Varì R., D’Archivio M., Di Benedetto R., Scazzocchio B., Giovannini C., et al. (2004). Extra virgin olive oil biophenols inhibit cell-mediated oxidation of LDL by increasing the mRNA transcription of glutathione-related enzymes. J. Nutr. 134, 785–791. doi: 10.1093/jn/134.4.785
Mates J. M. (2000). Effects of antioxidant enzymes in the molecular control of reactive oxygen species toxicology. Toxicology 153, 83–104. doi: 10.1016/S0300-483X(00)00306-1
Mesa-Herrera F., Quinto-Alemany D., and Diaz M. (2019). A sensitive, accurate, and versatile method for the quantification of superoxide dismutase activities in biological preparations. Reactive oxygen species 7 , 10–20. doi: 10.20455/ros.2019.809
Morales A. E., Cardenete G., Hidalgo M. C., Garrido D., Martín M. V., and Almansa E. (2017). Time course of metabolic capacities in paralarvae of the common octopus, Octopus vulgaris, in the first stages of life. Searching biomarkers of nutritional imbalance. Front. Physiol. 8. doi: 10.3389/fphys.2017.00427
Nagi M. N. and Mansour M. A. (2000). Protective effect of thymoquinone against doxorubicin–induced cardiotoxicity in rats: A possible mechanism of protection. Pharmacol. Res. 41, 283–289. doi: 10.1006/phrs.1999.0585
Nande M., Monroig Ó., MaChado A. M., Castro L. F. C., Lopes-Marques M., Capitão A., et al. (2022). Dietary lipids as modulators of fatty acid profile and gene expression patterns on body compartments of Octopus vulgaris paralarvae. Aquaculture 556, 738293. doi: 10.1016/j.aquaculture.2022.738293
Niroomand M., Akbarzadeh A., Ebrahimi-Dorche E., Sobhani S. A., Sheikhahmadi A., and Jalali S. A. H. (2024). The effects of dietary black cumin (Nigella sativa) seed meal on immune response of Pacific white shrimp (Penaeus vannamei). J. Appl. Aquaculture 36, 666–685. doi: 10.1080/10454438.2023.2294099
Nur I., Munaeni W., and Abidin L. O. B. (2020). Assessment of antibacterial and immunostimulating activity of black cumin (Nigella sativa) extract against vibriosis in white shrimp (Litopenaeus vannamei). Thai J. Veterinary Med. 50, 549–557. doi: 10.56808/2985-1130.3061
Ohkawa H., Ohishi N., and Yagi K. (1979). Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Analytical Biochem. 95, 351–358. doi: 10.1016/0003-2697(79)90738-3
Öz M., Dikel S., and Durmus M. (2018). Effect of black cumin oil (Nigella sativa) on the growth performance, body composition and fatty acid profile of rainbow trout (Oncorhynchus mykiss). Iranian J. Fisheries Sci. 17, 713–724. doi: 10.22092/ijfs.2018.116826
Parrillo L., Coccia E., Volpe M. G., Siano F., Pagliarulo C., Scioscia E., et al. (2017). Olive mill wastewater-enriched diet positively affects growth, oxidative and immune status and intestinal microbiota in the crayfish, Astacus leptodactylus. Aquaculture 473, 161–168. doi: 10.1016/j.aquaculture.2017.02.013
Reverter M., Bontemps N., Lecchini D., Banaigs B., and Sasal P. (2014). Use of plant extracts in fish aquaculture as an alternative to chemotherapy: current status and future perspectives. Aquaculture 433, 50–61. doi: 10.1016/j.aquaculture.2014.05.048
Sani M., Sebaï H., Gadacha W., Boughattas N. A., Reinberg A., and Mossadok B. A. (2006). Catalase activity and rhythmic patterns in mouse brain, kidney and liver. Comp. Biochem. Physiol. Part B 145, 331–337. doi: 10.1016/j.cbpb.2006.08.005
Sateriale D., Facchiano S., Kaldre K., Forgione G., De Cristofaro G. A., Pagliarulo C., et al. (2023). Benefits of polyphenol-based synbiotics in crustacean diet. Fishes 8, 255. doi: 10.3390/fishes8050255
Semedo M., Reis-Henriques M. A., Rey-Salgueiro L., Oliveira M., Delerue-Matos C., Morais S., et al. (2012). Metal accumulation and oxidative stress biomarkers in octopus (Octopus vulgaris) from Northwest Atlantic. Sci. Total Environ. 433, 230–237. doi: 10.1016/j.scitotenv.2012.06.058
Sharma R., Yang Y., Sharma A., Awasthi S., and Awasthi Y. (2004). Antioxidant role of glutathione S-transferases: protection against oxidant toxicity and regulation of stress-mediated apoptosis. Antioxidants Redox signalling 6 2, 289–300. doi: 10.1089/152308604322899350
Tadese D., Song C., Sun C. X., Bo L., Zhou Q., Xu P., et al. (2021). The role of currently used medicinal plants in aquaculture and their action mechanisms: A review. Rev. Aquaculture 14, 816–847. doi: 10.1111/raq.12626
Townsend D. M. and Tew K. D. (2003). The role of glutathione-S-transferase in anti-cancer drug resistance. Oncogene 22, 7369–7375. doi: 10.1038/sj.onc.1206940
Tur R., Domingues P., Almansa E., García-Fernández P., Lago M. J., and Pérez E. (2020). Procedimiento para el cultivo de paralarvas del pulpo común, Octopus vulgaris. – Patente IEON° 201731369.
Valgimigli L. (2023). Lipid peroxidation and antioxidant protection. Biomolecules 13, 1291. doi: 10.3390/biom13091291
Varó I., Prado-Álvarez M., Ortea I., Morales A. E., García-Fernández P., Domingues P., et al. (2022). Proteogenomic study of the effect of an improved mixed diet of live preys on the aquaculture of Octopus vulgaris paralarvae. Front. Mar. Sci. 8, 817701. doi: 10.3389/fmars.2021.817701
Viciano E., Monroig Ó., Barata C., Peña C., and Navarro J. C. (2017). Antioxidant activity and lipid peroxidation in Artemia nauplii enriched with DHA-rich oil emulsion and the effect of adding an external antioxidant based on hydroxytyrosol. Aquaculture Res. 48, 1006–1019. doi: 10.1111/are.12943
Xavier M. J., Conceição L. E. C., Valente L. M. P., Colen R., Rodrigues A. C. M., Rocha R. J. M., et al. (2021). Dietary natural plant extracts can promote growth and modulate oxidative status of Senegalese sole postlarvae under standard/challenge conditions. Anim. (Basel) 11, 1398. doi: 10.3390/ani11051398
Keywords: O. vulgaris, paralarvae, bioactive compounds, stress biomarkers, lipids
Citation: Martín MV, Hachero-Cruzado I, Navarro JC, Lago MJ, Almansa E, Gestal C and Varó I (2025) Effect of natural bioactive compounds on growth and welfare in Octopus vulgaris paralarvae. Front. Mar. Sci. 12:1629293. doi: 10.3389/fmars.2025.1629293
Received: 15 May 2025; Accepted: 05 August 2025;
Published: 01 September 2025.
Edited by:
Luca Parma, University of Bologna, ItalyReviewed by:
Khaled Mohammed Geba, Menoufia University, EgyptErcüment Genç, Ankara University, Türkiye
Copyright © 2025 Martín, Hachero-Cruzado, Navarro, Lago, Almansa, Gestal and Varó. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: M. Virginia Martín, dmlyZ2luaWEubWFydGluQGllby5jc2ljLmVz; Inmaculada Varó, aW5tYUBpYXRzLmNzaWMuZXM=