 Rafaela Franco Araújo1,2*
Rafaela Franco Araújo1,2* Marcos Sidney Brito Oliveira2
Marcos Sidney Brito Oliveira2 Abthyllane Amaral Carvalho2João Gabriel Rosário Luz2Marcela Nunes Videira2*Aldi Feiden1
Abthyllane Amaral Carvalho2João Gabriel Rosário Luz2Marcela Nunes Videira2*Aldi Feiden1- 1Aquaculture Management Study Group (GEMAq), State University of Western Paraná, Toledo, Paraná, Brazil
- 2Morphophysiology and Animal Health Laboratory (LABMORSA), State University of Amapá, Macapá, Amapá, Brazil
A systematic review was carried out to document the occurrence of myxozoan parasites in farmed and wild teleost fish in Brazil, considering publications made between 1969 and April 2024. This review followed the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines, and searches were conducted in the Web of Science, Scopus and Pubmed databases. After applying the criteria, 226 articles were selected, recording 286 taxa of myxozoan parasites, of which the genera Henneguya and Myxobolus are the most described in fish in Brazil. Overall, these studies covered ≈ 3% of the more than 4,900 fish species known in Brazilian waters. All reviewed myxozoan genera showed high specificity for infection sites, except Myxobolus, which was a generalist. In the host-parasite interaction network, fish species of importance to Brazilian fishing activity (Pimelodidae, Serrasalmidae, Cichlidae, Bryconidae and Prochilodontidae) obtained the highest number of associations, being the most researched. Regarding the geographic distribution of studies with myxozoans, the majority (79%) occurred with wild fish, originating from all Brazilian river basins, with the exception of the Uruguay River basin. On the other hand, studies that recorded myxozoans in rearing hosts (21% of studies) occurred mainly in the Paraná River basin.
1 Introduction
Among the largest groups that parasitize fish in the Neotropical region, the class Myxozoa Grassé 1970 (Kyger et al., 2021), has approximately 2,596 described species, distributed in 67 genera (Okamura et al., 2018). This class has a cosmopolitan distribution, being found in freshwater and marine environments and some species are pathogenic (Eszterbauer et al., 2020). They are usually microscopic spores measuring approximately 10 to 50 micrometers, with simple morphology and a complex life cycle, which is still little known (Eszterbauer et al., 2020). Generally found in the myxosporean form in vertebrate hosts, they can contain from 2 to 7 valves connected by suture lines and polar capsules that vary from 1 to 7, each capsule containing a polar filament coiled inside in a spiral shape (Lom and Dyková, 1992).
Myxozoans are microparasites with a simplified body organization classified within the Phylum Cnidaria. Closely related to jellyfish, sea anemones and hydras, during evolutionary processes they diverged from the common ancestor to become a parasitic group of vertebrates and invertebrates (Eszterbauer et al., 2020). During their life cycle they have two hosts, one vertebrate (usually fish) and one invertebrate (mainly annelids). In their developmental stages myxozoans are motile during invasion, migration and proliferation due to cellular processes that allow this unique cellular motility (Eszterbauer et al., 2009; Alama-Bermejo et al., 2019). Myxozoans affect the health of fish, and can cause disease and mortality, since there is no effective protection for fish against these parasites (Okamura et al., 2015).
Among the vertebrates infected by parasites, fish have shown greater susceptibility, possibly due to their aquatic environment, in which water facilitates the transmission and dispersion of these organisms (Araujo Neto et al., 2024). Some species of myxozoans are highly pathogenic in both farmed and wild fish and cause serious damage to their hosts and major economic losses to the fishing industry (Naldoni et al., 2019b).
Myxozoan infection in fish can affect different organs and tissues, leading to serious pathological lesions such as reduction of the epithelial area of the gills, myocarditis deformation, displacement, stretching and compression of the capillaries of the gill lamellae, hemorrhage and hypertrophy in the skin, thickening of the external tunic of the swim bladder with granulomatous reaction, perivascular edema in the interlamellar area and stretching of the epithelium of the cornea and urinary bladder (Barassa et al., 2003; Adriano et al., 2005, 2009; Matos et al., 2014; Battazza et al., 2020; Capodifoglio et al., 2020).
Worldwide, studies on the occurrence of Myxozoa parasites causing serious diseases in freshwater and marine host fish have been reviewed by Banu and Rathinam (2023). Among them is proliferative kidney disease caused by Tetracapsuloides bryosalmonae (Canning, Curry, Feist, Longshaw & Okamura, 1999) in salmon species on the European continent (Naldoni et al., 2019a; Oredalen et al., 2022). Furthermore, Kudoa thyrsites (Gilchrist, 1923) and Kudoa paniformis Kabata & Whitaker, 1981 caused muscle myoliquefaction in Scomber scombrus Linnaeus, 1758 (Atlantic mackerel) from Norway (Giulietti et al., 2022). Henneguya ictaluri Kabata & Whitaker, 1981 caused proliferative disease in the gills of catfish and their hybrids, known as “hamburger gill”, which causes severe respiratory distress leading to high mortality and substantial economic losses in the USA aquaculture industry (Stilwell et al., 2023). Kudoa septempunctata Matsukane, Sato, Tanaka, Kamata & Sugita-Konishi, 2010 was detected in the muscle of Paralichthys olivaceus (Temminck & Schlegel, 1846) (flounder) raised in aquaculture systems and has been associated with numerous outbreaks of foodborne illness in Japan, with gastrointestinal symptoms in individuals who ingested this fish (Matsukane et al., 2010; Yahata et al., 2015).
Therefore, following the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines (Moher et al., 2015; Page et al., 2021), a systematic review was conducted to document the occurrence of ichthyoparasites of the Myxozoa Class in farmed teleost fish and wild populations in Brazil, outlining an overview of the geographic distribution, parasite-host interaction, infection sites, in addition to describing the main histopathological changes caused to the fish, aiming to contribute to the knowledge of the health status of the Brazilian ichthyofauna as well as monitoring the status of studies on Myxozoa in Brazil.
2 Materials and methods
2.1 Search strategy
This systematic literature review followed the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines (Moher et al., 2015; Page et al., 2021).
A search was performed in the following databases: Web of Science core collection, Scopus and Pubmed, up to April 2024. Search strings were made for each database, using the search terms “titles” and “abstract” and the Boolean operators search strategy (OR and AND), which were applied to match the following keywords: “fish*” OR “aquaculture” combined with “parasit*” OR “Myxozoa” and “Brazil” (Table 1).

Table 1. Database search strategies.
The lists of records retrieved from each database were merged, and duplicate and inaccessible articles, book chapters, editorial material, errata and conference papers were subsequently excluded. Two independent reviewers evaluated all titles and abstracts, the selection criteria adopted in the review were: studies of parasites of the Myxozoa class; in teleost fish; carried out in Brazil. The bibliography of the selected full texts was reviewed, relevant studies were added to the review and the following selection and exclusion criteria were applied to each article: articles or scientific notes from indexed journals; of any year of publication; written in English, Portuguese or Spanish; which included information on the collection location (municipality or geographic coordinates) and primary research that documented the presence of Myxozoa parasites in fish from natural or farmed environments in Brazil. In short, articles were screened for eligibility using the following exclusion criteria: reviews or checklists that did not present primary research; studies on hosts other than fish; outside the theme of the present study; and unavailable articles.
The lack of uniformity in the designation of study sites, such as geographic coordinates of a fish farm, and in the assessment of the effects of myxozoan infection on fish growth and productivity make it difficult to accurately indicate the location of myxozoans and advances in systematic research on the subject. Furthermore, in some articles analyzed in this review, a lack of data on the infection site by fish species and/or collection site was observed.
2.2 Data collection
Data extracted from the publications included: author, date, title, taxonomy of the parasite (genus and species), site of infection, taxonomy of the host fish (family, genus and species), origin of the host (wild or farmed fish), municipality and state of sample collection, hydrographic basin, geographic coordinates and histopathological alteration. The taxonomic status of the host fish species was checked and validated based on updated data from the FishBase database, ensuring consistency with the real scenario of the Brazilian ichthyofauna (Froese and Pauly, 2024). Each line of the matrix corresponds to a parasite-host-environment interaction, recording the presence of a parasite in a host species in a municipality within the study.
To present the geographic distribution of myxozoan genera infecting wild and/or aquaculture fish in Brazilian River basins (Amazon River, North-Northeast Atlantic, Tocantins River, São Francisco River, Paraná River, Uruguay River, East Atlantic and Southeast Atlantic), we compiled the coordinates of the parasite collection points from the articles in the database. For articles that did not provide geographic coordinates, coordinates were collected using Google Earth software considering the location information that the articles provided (river, municipality, small communities).
2.3 Data analysis
Microsoft Excel® spreadsheets were used to select articles and extract and describe data.
The R software and the “bipartite” package were used to define interaction patterns, with networks drawn using the plot web function (Dormann, 2011; Dormann et al., 2008; R Core Team, 2017) to determine the relationships between parasite and infection site and parasite-host at the genus level. The C-score indices at the network level, number of compartments and genus specificity index of Myxozoa were calculated (SSI) (Dormann, 2011). The C-score index measures the co-occurrence rate of genera in the network and is an indicator of the degree of specificity of these genera, with values ranging from 0 (high co-occurrence) to 1 (low co-occurrence). The SSI assesses the degree of specificity of the parasite genera, ranging from 0 (low specificity) to 1 (high specificity). To classify the specificity levels within this range, we considered values higher than 0.66 to be high, values between 0.33 and 0.66 to be moderate, and values lower than 0.33 to be low. These analyses were performed using R software (R Core Team, 2017).
The coordinates obtained were organized in Microsoft Excel® spreadsheets, saved in csv format and exported to Quantum-Gis (QGIS) for map creation. The shape file layers used (hydrographic grid and federation units) in the preparation of the maps were obtained from the Brazilian Institute of Geography and Statistics (IBGE).
3 Results
The search in the databases located 2,466 works and after applying the selection criteria, 226 articles were included in the data analysis (Figure 1). From these studies, 341 parasite-host-environment interactions were recorded. One hundred and seventy-nine species (286 taxa) of myxozoan parasites of 134 species (136 taxa) of host fish were recorded, and six hybrids. Of the 286 myxozoan taxa found, 245 were in wild fish and only 48 in fish from fish farms.
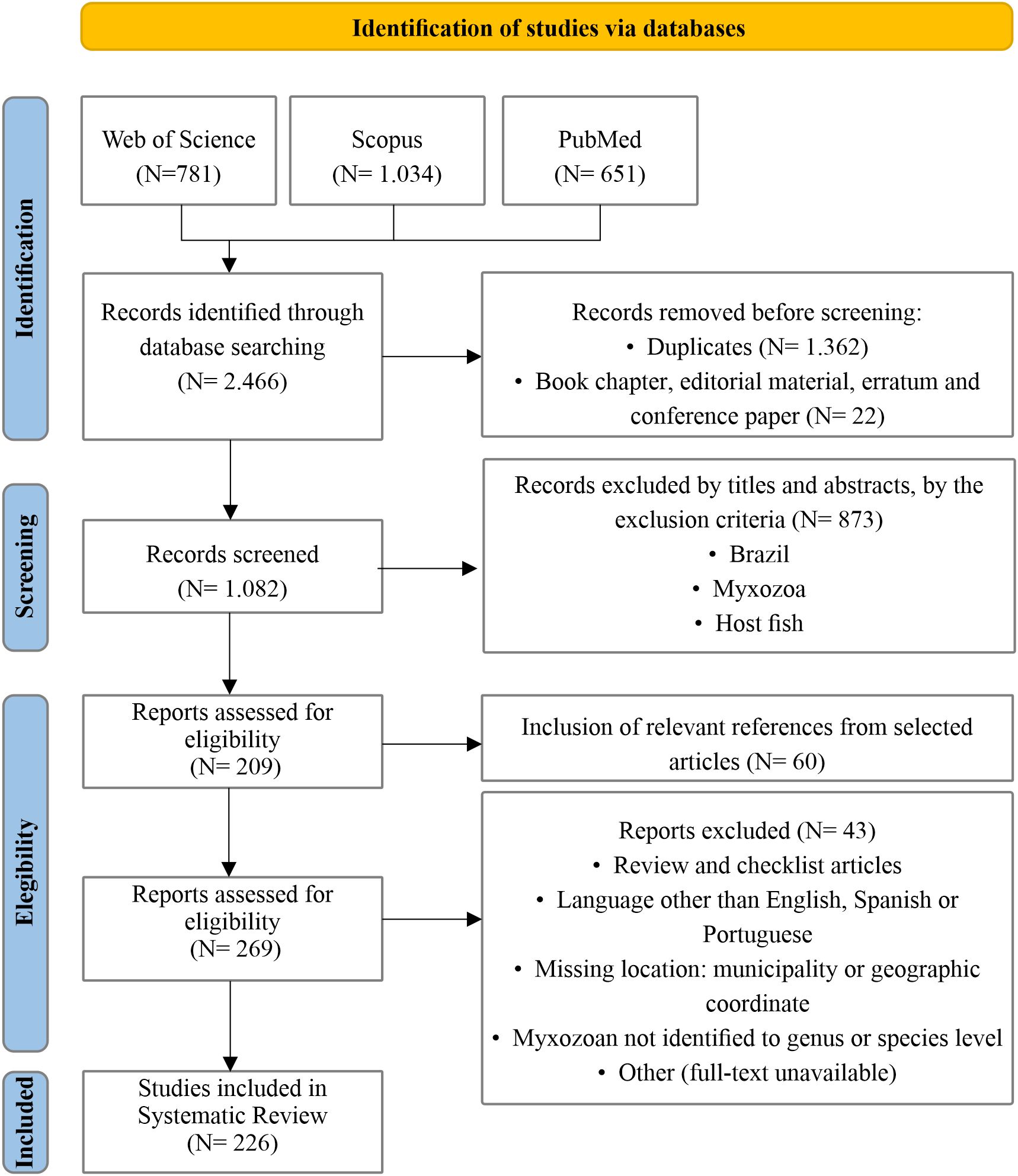
Figure 1. Flowchart of the study search and selection process, as established in the PRISMA 2009 methodology, according to Moher et al. (2015).
A total of 19 genera of Myxozoa occur in Brazil, namely: Agarella (1 species), Ceratomyxa (10), Chloromyxum (1), Coccomyxa (1), Ellipsomyxa (7), Henneguya (68), Hoferellus (2), Kudoa (9), Meglitschia (1), Myxidium (4), Myxobolus (65), Ortholinea (sp.), Sinuolinea (1), Sphaeromyxa (1), Sphaerospora (1), Tetrauronema (1), Thelohanellus (2), Triangulamyxa (2) and Unicauda (2). Henneguya and Myxobolus are the most recurrent in fish in Brazil, where around 80% of studies recorded these genera (Figure 2A).
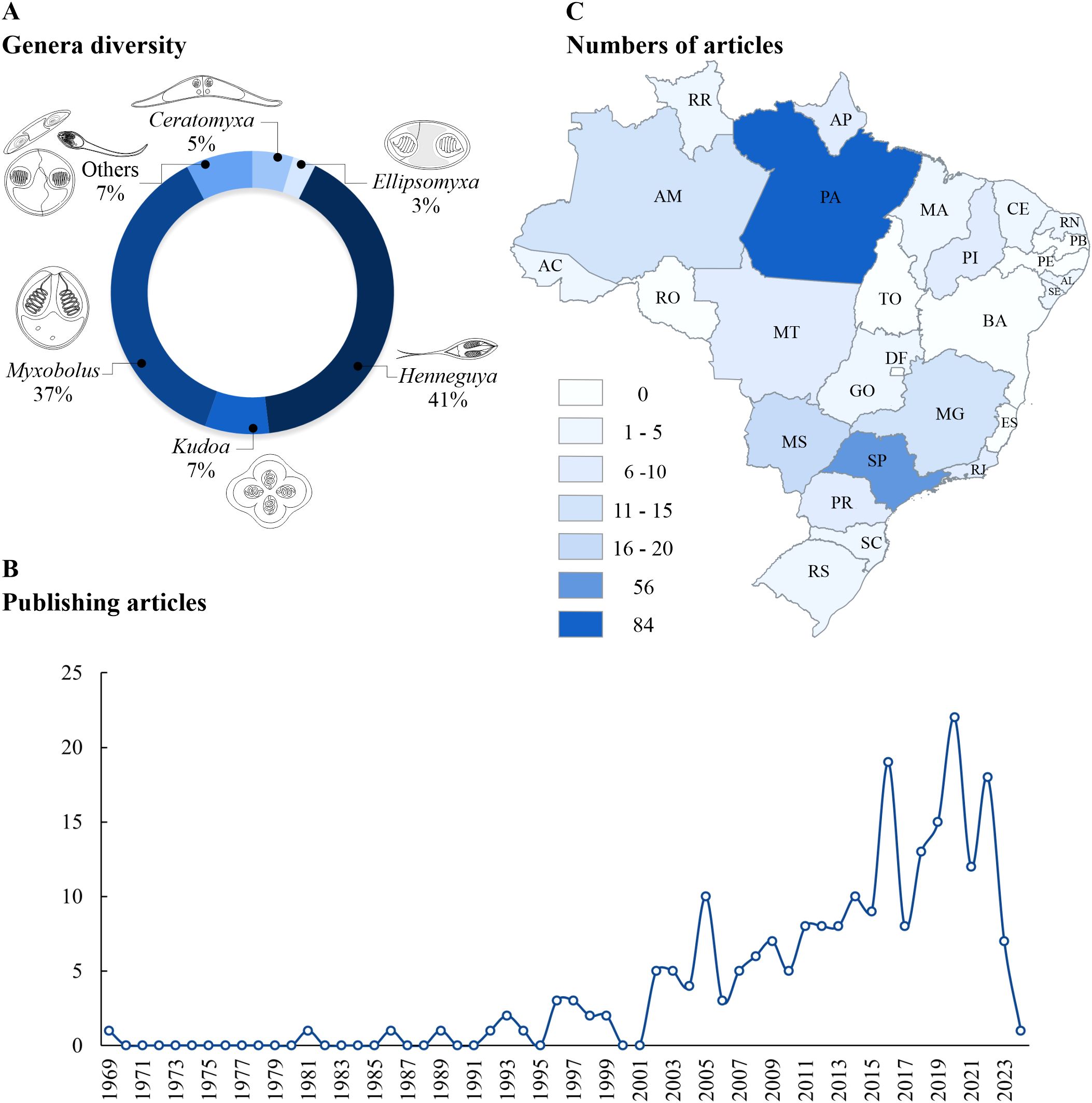
Figure 2. (A) Percentage of Myxozoa genera described in Brazil. “Others” corresponds to Agarella, Chloromyxum, Coccomyxa, Hoferellus, Meglitschia, Myxidium, Ortholinea, Sinuolinea, Sphaeromyxa, Sphaerospora, Tetrauronema, Thelohanellus, Triangulamyxa and Unicauda. (B) Variation in the number of articles published on Myxozoa in fishes in Brazil between 1969 and early 2024. (C) Numbers of jobs performed in Brazilian states. Abbreviations of Brazilian states: AC, Acre; AL, Alagoas; AM, Amazonas; AP, Amapá; BA, Bahia; CE, Ceará; ES, Espírito Santo; GO, Goiás; MA, Maranhão; MG, Minas Gerais; MS, Mato Grosso do Sul; MT, Mato Grosso; PA, Pará; PB, Paraíba; PE, Pernambuco; PI, Piauí; PR, Paraná; RJ, Rio de Janeiro; RN, Rio Grande do Norte; RO, Rondônia; RR, Roraima; RS, Rio Grande do Sul; SC, Santa Catarina; SE, Sergipe; SP, São Paulo; TO, Tocantins.
The first recovered article was published in 1969 and only in 1981 was a new article published (Figure 2B). From 2002 onwards, there was a gradual increase in the number of publications on myxozoans infecting fish and between 2011 and 2020. This increase was 175%, that is, it went from 8 to 22 published articles. Over the period from 1969 to 2024, the annual average of published articles was 4.1, ranging from 0 to 22.
The data show a regional centralization in the number of published articles. The state of Pará stands out as a hub for the collection of myxozoans in Brazilian fish, concentrating 84 papers (34% of national production) in the country until the beginning of 2024 (Figure 2C). Pará is followed by São Paulo (56), Mato Grosso do Sul (17), Amazonas (15) and Minas Gerais (11). On the other hand, states that do not have records include Bahia, Espírito Santo, Paraíba, Pernambuco, Rondônia, Tocantins and Distrito Federal.
In the network of interactions between myxozoan genera and the infection sites of host fish, the parasite co-occurrence rate was low at the network level (C-score = 0.691), indicating that most genera have an affinity for only one infection site. Overall, 84% of parasite genera exhibited high specificity for infection sites, 11% exhibited moderate specificity, and 5% exhibited low specificity. Species of the genus Henneguya has a greater affinity for the gills. Ceratomyxa, Ellipsomyxa e Myxidium are specific to the gallbladder, while Kudoa demonstrates specificity for host muscle. Myxobolus species do not show site specificity, recorded at 25 sites on the host (Figure 3; Table 2).
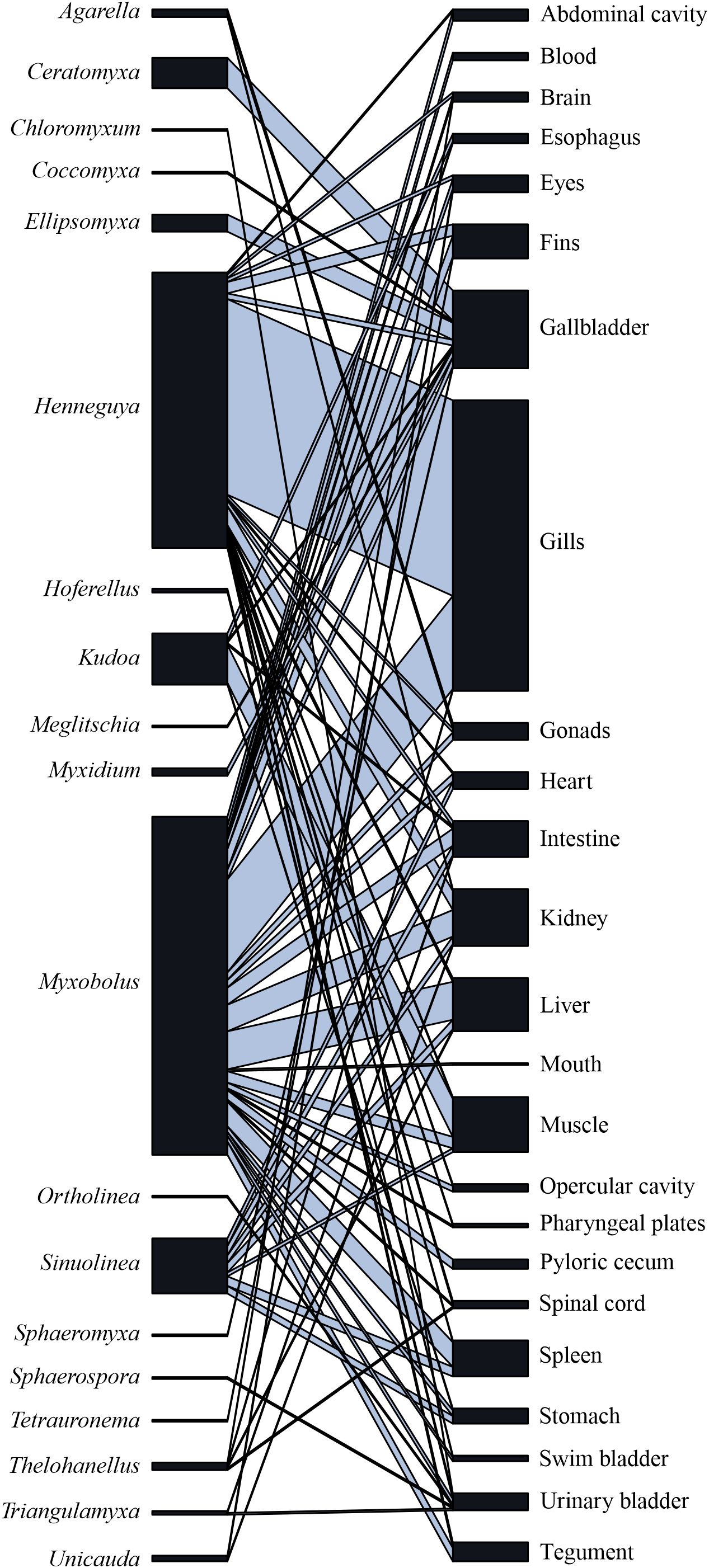
Figure 3. Interactions network between Myxozoa genera and infection site.

Table 2. Specificity indices to assess relationships between myxozoan genera and infection sites in fish from Brazil.
In the interaction network between parasite genera and fish genera, the co-occurrence rate of parasites was moderate at the network level (C = 0,528). Overall, 58% of myxozoan genera showed high specificity for host genera, while 26% showed moderate specificity and 16% exhibited low specificity. The parasites that occurred in the greatest number of host genera were Henneguya (n = 55), Myxobolus (n = 41) and Kudoa (n = 19), being considered generalist parasites (Figure 4; Table 3).
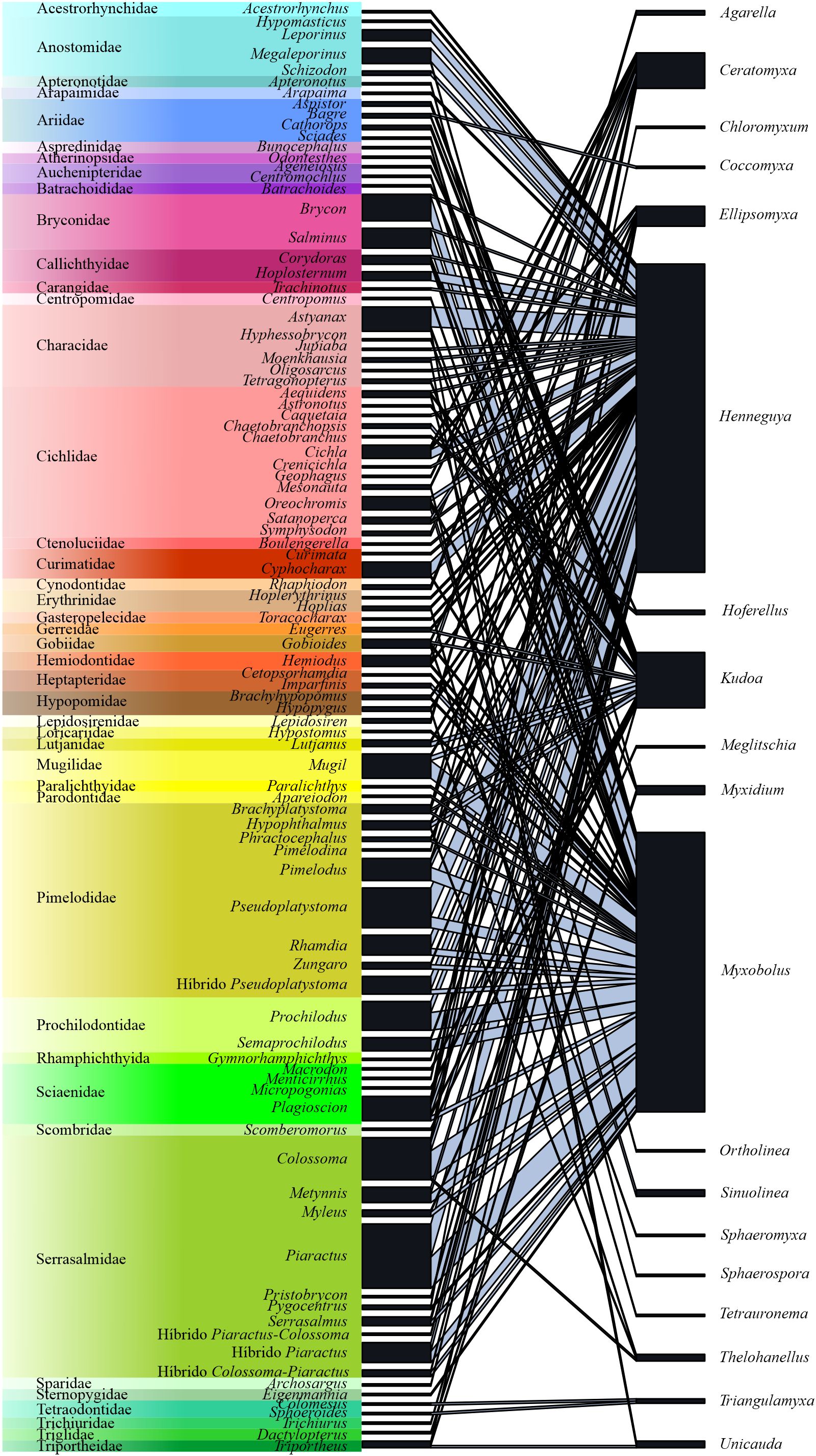
Figure 4. Interactions network between host fish species (n = 341) and Myxozoa genera from Brazil.
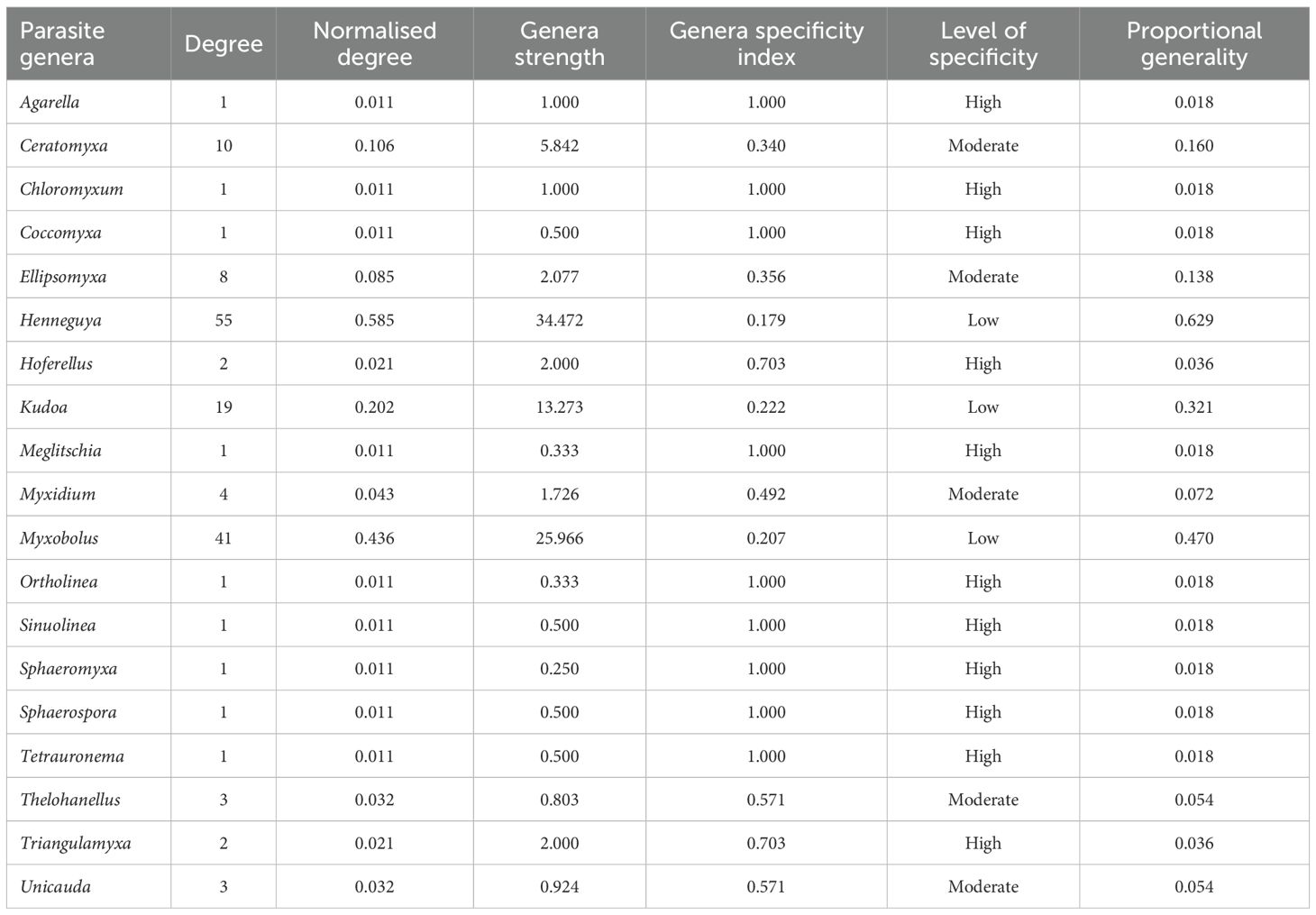
Table 3. Specificity indices to assess the relationships between myxozoan genera and fish genera from Brazil.
In the interaction network between parasite genera and fish families, fish from families Pimelodidae (Rhamdia quelen, Pseudoplatystoma corruscans and its hybrid Pseudoplatystoma reticulatum x Pseudoplatystoma corruscans), Serrasalmidae (Colossoma macropomum, Piaractus mesopotamicus and its hybrid Piaractus mesopotamicus x Piaractus brachypomus), Cichlidae (Cichla monoculus, Oreochromis niloticus), Bryconidae (Salminus brasiliensis and Brycon spp.) and Prochilodontidae (Prochilodus lineatus e Semaprochilodus insignis) harbored a greater number of species of myxozoans, infected mainly by Henneguya and Myxobolus. On the other hand, fish from the families Cichlidae (23 parasite-host-environment interactions) and Sciaenidae (9 interactions) were also parasitized by Ceratomyxa and Ellipsomyxa (Supplementary Table 1).
Overall, 79% of studies on myxozoans are mainly associated with wild fish, with 271 parasite-host-environment interactions, distributed throughout all basins except the Uruguay River basin. On the other hand, only 21% of the studies on parasites are related to rearing hosts, with 70 parasite-host-environment interactions, which are mainly concentrated in the Paraná River basin. The Paraná and Amazon River basins concentrate 38% and 26% of the myxozoan species parasitic on fish in Brazil, respectively (Figure 5A).
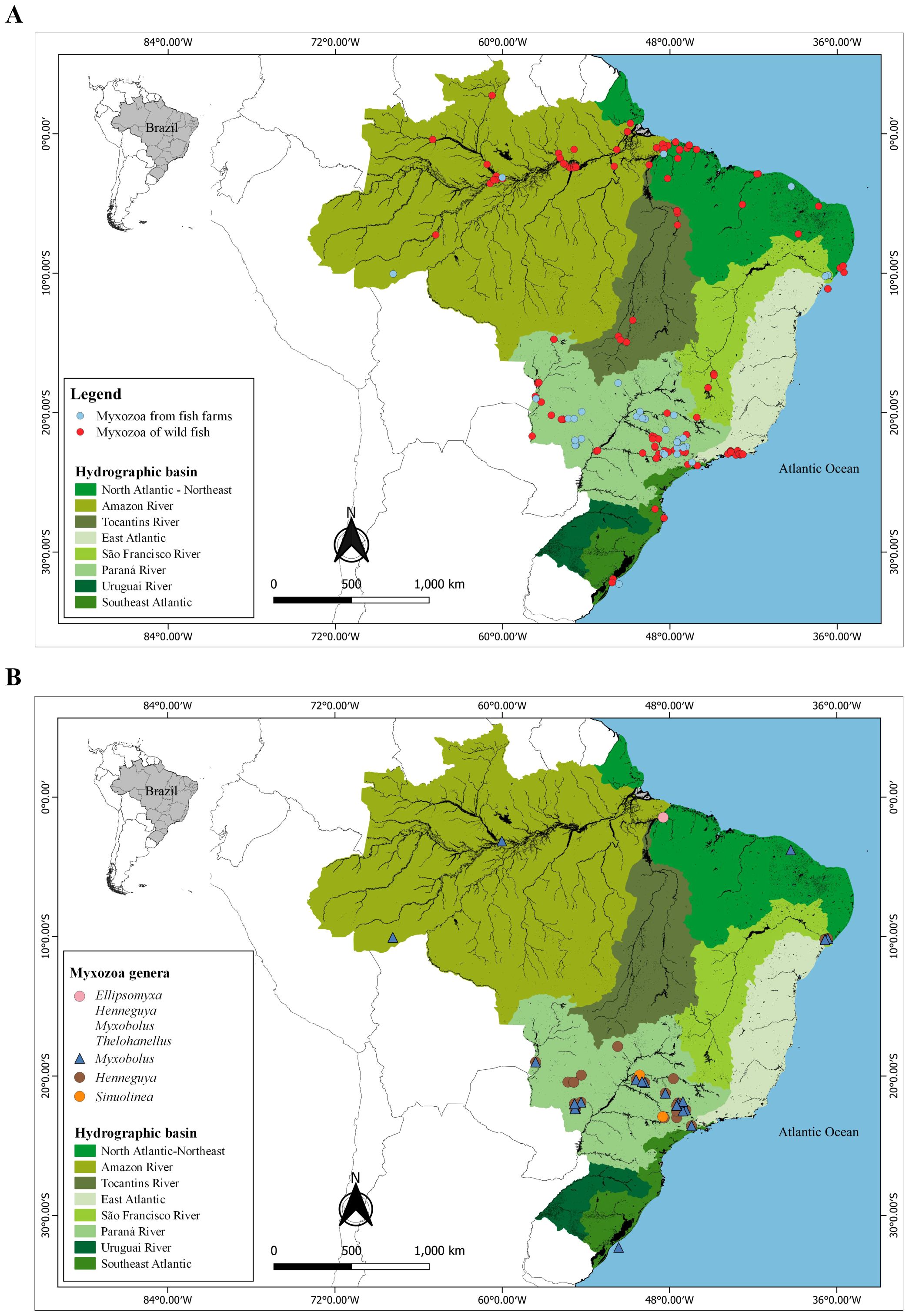
Figure 5. Geographic distribution data of Myxozoa in (A) wild and farmed fish from Brazil. (B) Myxozoa genera from farmed fish from Brazil.
The farmed fish found are parasitized by species from five genera (Ellipsomyxa, Henneguya, Myxobolus, Sinuolinea and Thelohanellus), of which Henneguya (50%) and Myxobolus (44%) are the most frequent, with the remaining genera representing only 6%. Most species of aquaculture fish parasites were recorded in the Paraná River basin (81%), with emphasis on the states of São Paulo and Mato Grosso do Sul. On the other hand, until the present study, no publications had been recorded with myxozoans of fish from the East Atlantic, Southeast Atlantic, Tocantins River basin and Uruguay River basin (Figure 5B).
In terms of parasites of wild fish, species from nineteen different genera were identified, with Henneguya (41%) and Myxobolus (37%) being the most frequent. The Amazon River basin concentrated 30% of the myxozoans, with emphasis on the states of Pará and Amazonas. The Paraná River and North and Northeast Atlantic basins presented 27% and 19% of the studies with myxozoans, respectively (Figures 6A, B)
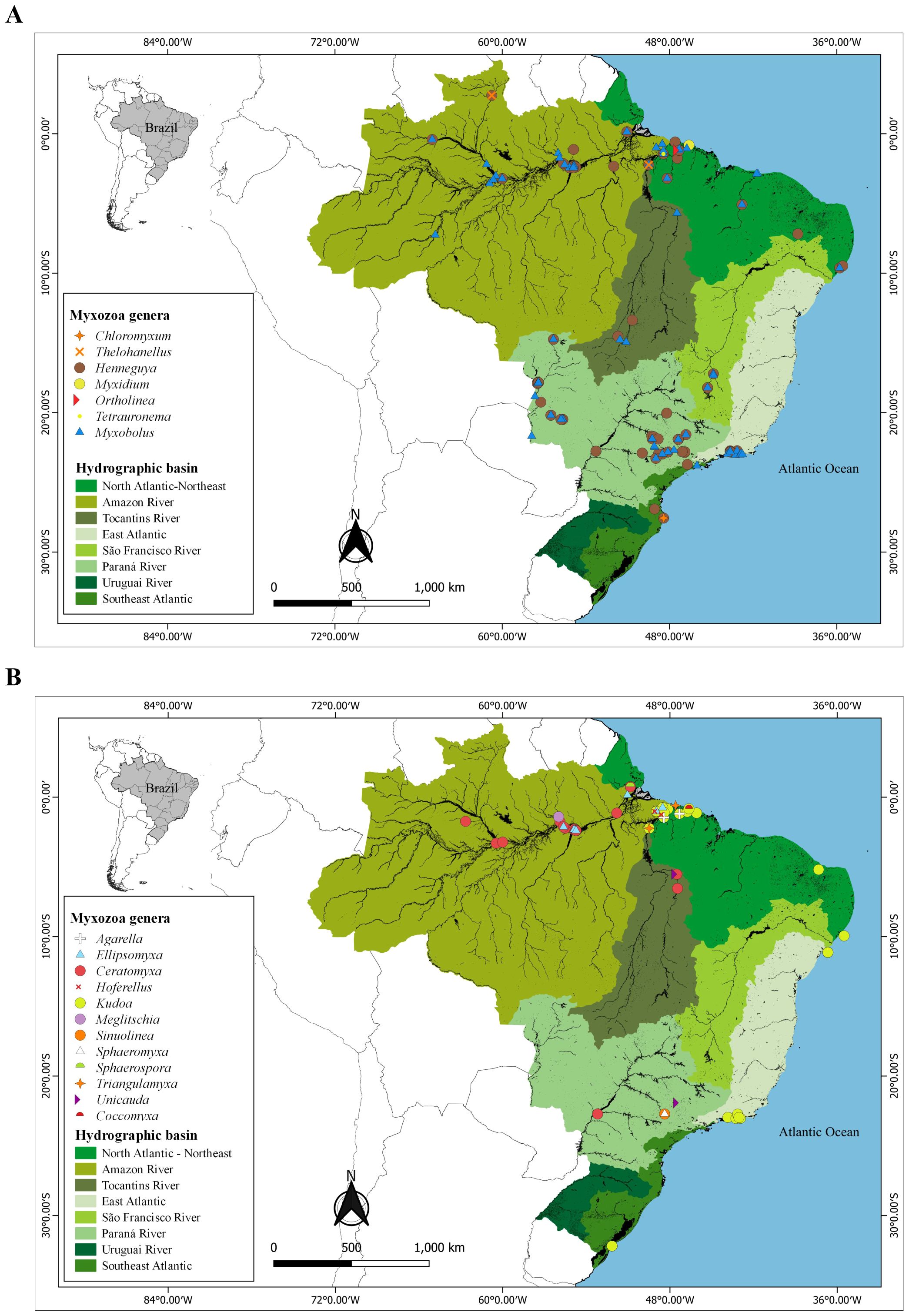
Figure 6. Geographic distribution data of the 19 genera of Myxozoa in wild fishes from Brazil. (A) Chloromyxum, Thelohanellus, Henneguya, Myxobolus, Myxidium, Ortholinea, Tetrauronema e (B) genera Agarella, Ellipsomyxa, Ceratomyxa, Hoferellus, Kudoa, Meglitschia, Sinuolinea, Sphaeromyxa, Sphaerospora, Triangulamyxa, Unicauda and Coccomyxa.
The main histopathological lesions in fish caused by myxozoans are associated with the genera Kudoa, Henneguya and Myxobolus, occurring mainly in the gills and muscle of the host. Common changes are post-mortem liquefactive necrosis of muscle tissue, replacement of muscle fibers, multifocal inflammatory reactions in the gills, lamellar fusion, mucous cell hyperplasia and tissue degeneration (Supplementary Table 2).
4 Discussion
The bibliographic survey recovered 179 species of Myxozoa parasites of fish from Brazil. That shows that this is a highly diverse group in terms of the number of species parasitizing wild and farmed fish in different river basins in Brazil. Overall, Myxozoa species were recorded in 134 species of host fish, representing only 2.7% of the total diversity of approximately 4,900 species of fish currently found in Brazilian waters (Froese and Pauly, 2024). Considering this high diversity of fish in the country, apparently the factor responsible for the low knowledge of the parasitological fauna is the low number of researchers investigating these parasites (Eiras et al., 2011).
Extrapolation patterns of parasite specificity by host are used to estimate parasite diversity from simple equations (Okamura et al., 2018). Using such patterns, Eiras et al. (2011) estimated that each fish species has on average two species of myxozoans, and proposed the existence of about 8,000 species in freshwater fishes from Brazil. However, it is not known whether the curve of known parasites is increasing or in the process of decelerating, having no solid inference of parasite diversity and no adequate method to obtain it (Okamura et al., 2018; Poulin et al., 2020). The currently known diversity of Myxozoa is possibly underestimated, and this may be related to the biased focus on studying parasites of hosts of greater economic value or parasite species of medical, veterinary or zoonotic interest (Poulin et al., 2020).
The first article recovered was published in 1969, where it presented a synopsis of the Myxobolus species of South America and recorded a new species, Myxobolus serrasalmi Walliker, 1969, identified in a freshwater fish in the Amazon region (Walliker, 1969). Previous occurrences were recorded but were not in the languages established in the methodology (Spanish, English or Portuguese), or were not available in indexed journals, so they were excluded from this review according to the eligibility criteria. Over the past 55 years, there has been a gradual increase in the number of publications that reflects the continued interest of specialists in the field. The molecular phylogeny tool has helped in the identification and characterization of myxozoans, as well as in the review of this group (Capodifoglio et al., 2015; Manrique et al., 2016; Rocha et al., 2019; Vieira et al., 2019). Despite the national growth in the number of scientific publications in recent years, from 2021 onwards there has been a drop in the publication of articles referring to myxozoans in fish in Brazil, and the results show that this reduction coincides with the period of the pandemic caused by the SARS-CoV-2 virus, in early 2020, making scientific production unfeasible to some extent.
Since the 1970s, the methodology used to identify and categorize parasites of the Myxozoa class has undergone significant advances. Initially, studies were based exclusively on morphological characteristics observed by optical microscopy, scanning electron microscopy and transmission electron microscopy, focusing on spore shape and size, location of polar capsules, presence of filaments, and preparation of histological slides for measurement and description of the parasites (Rocha et al., 1992). Although these approaches were essential for the initial records of parasitic fauna, they presented limitations regarding the differentiation of cryptic or morphologically similar species, in addition to potential inaccuracies in taxonomy, especially in regions with great ichthyofaunal diversity.
In Brazil, until the first decade of the 21st century, studies remained restricted to this traditional morphological approach, which made the taxonomy of the group even more challenging (Bartošová et al., 2013). From 2010, with the progress of molecular techniques and the consolidation of integrative taxonomy, national studies began to incorporate genetic analyses (such as amplification of the 18S rRNA gene), phylogenetic and morphological analyses in a combined manner (Milanin et al., 2010; Naldoni et al., 2011; Poulin et al., 2020). This methodological modernization provided greater rigor in the discrimination between species, enabled the revision of previously described taxa and boosted the description of new species with greater scientific robustness. Currently, it is recommended that the identification of myxozoans be based on multiple sources of evidence, including biological characteristics of the host, such as specificity, tissue tropism, developmental stages and geographic distribution (Dar et al., 2024).
We centralized our efforts on the collection of Myxozoa in fish in the State of Pará. Research focused on the Amazon generates interest among scientists, as it comprises the largest freshwater discharge on the planet, the most diverse river basin in the world and an ichthyofauna composed of more than 2,700 valid species. (Dagosta & Pinna, 2019; Filizola and Guyot, 2009; Latrubesse et al., 2017; Meade et al., 1985). The Amazon basin encompasses a considerable fraction of all known fish biodiversity in Brazil, concentrating around 55% of the country’s fish species. Additionally, Poulin et al. (2020) documented that latitudinal gradients and their relationship with water temperature influence the species richness of several taxa.
The following pattern of interaction between myxozoan genera and infection sites was detected: tropism of the tissue and/or organ of infection in most myxozoan genera that infect fish, i.e., specific parasites infecting a single site or more than one that are precisely associated. It is suggested that myxozoan species are frequently documented in the gills due to the possibility of myxospore release (Molnár, 2002; Eszterbauer et al., 2013). On the other hand, there were rare reports of parasitic infection in the mouth and pharyngeal plates. The genus Henneguya has a high affinity for the gills of its hosts, while the genus Ceratomyxa has an affinity for the gallbladder. Unlike these aforementioned genera, the genus Myxobolus has low tissue specificity, which corroborates the high diversity of species described within this genus.
Infection sites (histozoic and coelozoic), site specificity and host habitat (freshwater and marine) follow a high pattern of phylogenetic correlation of many myxozoan species (Eszterbauer, 2004; Fiala and Bartoŝová, 2010). Similarly, it was observed that the site of infection of the common ancestor of myxozoans was the Coelozoic renal tubules of freshwater fish and later the first myxozoans infected the gallbladder of marine fish (Fiala and Bartoŝová, 2010). Ceratomyxa, Ellipsomyxa and Myxidium are specific genera of the gallbladder of Brazil, a common characteristic that follows evolution and persists among descendants. However, the genus Myxobolus did not present specificity by site and has been recorded in 25 sites in the hosts, leading to it being considered a generalist parasite. The most numerous myxozoan on the planet in terms of number of species, Myxobolus, invaded various tissues in hosts and subsequently segregated into numerous species during evolution. The genus Henneguya originated from the Myxobolus morphotype, from the emergence of caudal appendages (without movement), demonstrating the importance of the infection site in the evolutionary adaptation of some Myxozoa (Fiala and Bartoŝová, 2010).
In this study, it was noticeable that the most studied fish species were those of importance to Brazilian fishing activity and belonged to the Pimelodidae (30 species of myxozoans), Serrasalmidae (29), Cichlidae (23), Bryconidae (20) and Prochilodontidae (14) families. The results observed reinforce the hypothesis by Eiras et al. (2011) that most studies on fish parasites are directed at species with significant commercial value, whether in the aquaculture market, extractive fishing for human use or in ornamental aquaculture, as they have greater appeal in research funding notices, and can be decisive when evaluating priority research lines. Farmed fish of the genus Piaractus and its hybrid, Pseudoplatystoma, Prochilodus, Rhamdia and Colossoma macropomum Cuvier, 1816 influenced the strong tendency of interaction with Myxozoa parasites. However, few fish have been studied from a parasitological point of view, when compared with their biodiversity.
Parasite species richness is closely associated with ecology and evolution of their host, so that they are correlated with the biodiversity patterns of the sampling area (Poulin et al., 2020). In this study, it was noticeable that research is carried out unevenly across Brazil, with emphasis on the Paraná and Amazon River basins, and it is clear that there are still many areas to be investigated. Spatially biased studies in the northern and southeastern regions also occur due to the work of specialists in sampling areas close to their institutions or the displacement of researchers to the Amazon region, as it presents a great diversity of hosts.
The Paraná River basin concentrates the largest number of studies of myxozoans in farmed fish. This indicator may be related to the fact that the state of São Paulo is notable as the second largest producer of tilapia (Oreochromis niloticus Linnaeus, 1758) in Brazil, added to the occurrence of a significant number of researchers in the area (Godoy et al., 2022). On the other hand, there is a need to research myxozoan parasites in geographic areas of high fish productivity, where there are potentially new species of parasites or possible cases of pathological lesions in fish.
It is possible to observe that the parasitic diversity is related to the diversity of wild fish researched, in which 19 distinct genera were identified, approximately 30% of the Myxosporea genera described in the world (Lom and Dyková, 2006). In contrast, only 8% of parasite genera were found in farmed fish. The genera Henneguya and Myxobolus were the most common parasites in both wild and farmed fish. As observed in parasites of vertebrate hosts worldwide, a weak geographic correspondence between host species richness and parasite discovery effort has been identified, preventing the completion of a complete inventory of parasite biodiversity and making it difficult to predict where new diseases may emerge (Jorge and Poulin, 2018; Poulin et al., 2020).
Myxozoans have been defined as cnidarians evolved into their endoparasitic from free-living ancestors, to a parasitic form that exploited a single host, preceded by pre-adaptation of the parasites that allowed the initial stages of host exploitation in order to achieve greater parasitic fitness through the maintenance of the association (Poulin, 2007; Okamura et al., 2018). The parasites that occurred in the greatest number of host genera were Henneguya, Myxobolus and Kudoa, which are considered generalist parasites.
Parasitism in wild or farmed fish was caused mainly by Henneguya, Myxobolus and Kudoa, often infecting the gills or muscle. Myxozoans are considered potential pathogens for fish, where they generally cause mild lesions and where the hosts reveal signs of discomfort, without presenting an inflammatory reaction at the site of infection or clinical signs of disease (Lom and Dyková, 2006). However, some of these parasites are responsible for serious pathologies associated with outbreaks of infection in various organs of the host, which can negatively affect zootechnical performance and cause the death of fish (Videira et al., 2016; Teixeira et al., 2018; Battazza et al., 2020). Although the reviewed studies focus mainly on the morphological description of the parasites and histopathological changes, quantitative data on economic losses are still scarce, which makes a more accurate assessment of the productive impacts difficult (Tavares-Dias and Martins, 2017).
Treatment of myxozoan diseases in farmed fish remains limited and challenging. Among the few available drugs, fumagillin has been widely used and effective when administered correctly. However, inadequate doses can cause significant side effects, such as loss of appetite, increased mortality, immunosuppression, and severe physiological changes. Therefore, determining the ideal therapeutic dose must carefully consider the fish species, the parasite involved, and the drug’s toxicity profile (Banu and Rathinam, 2023; Kent and Martins, 2017).
In addition to traditional drugs, the use of oregano essential oil, Origanum minutiflorum Schwarz & Davis, 1949, demonstrated antiparasitic effect in asparid fish infected by Myxobolus sp. (Karagouni et al., 2005). In Brazil, there are records of the use of baths with 37% formalin at a concentration of 10 µL/L, which showed a reduction in the parasite load and interruption of mortality in Megaleporinus macrocephalus Garavello & Britski, 1988 infected by Henneguya leporinicola, Martins, Souza, Moraes, Moraes, 1999 being a promising alternative in the national contexto (Martins et al., 1999).
Therefore, preventing infection remains the most effective approach to controlling myxosporidiosis (Noga, 2010). Key measures include good management practices, including reducing stocking density and controlling water quality to reduce fish stress, using pathogen-free water through protected sources, and controlling invertebrate hosts in culture systems (Fontes et al., 2015). Progress in controlling myxozoan infections depends on in-depth studies of the life cycle of these parasites, the validation of safe therapeutic strategies, and the implementation of biosafety measures. Considering the high diversity of species described in Brazil, especially in the genera Henneguya and Myxobolus, it is essential that future research focus on developing therapeutic alternatives suited to the reality of national fish farming.
5 Conclusion
This systematic review study summarized the works related to the occurrence of myxozoans in wild and farmed fish in Brazil. Parasites of the genera Henneguya and Myxobolus were frequently observed in wild and farmed fish distributed throughout Brazilian river basins. Among these fish, those of the families Pimelodidae, Serrasalmidae, Cichlidae, Bryconidae and Prochilodontidae where notable, due to their importance to the for Brazilian fisheries sector. In addition, some pathogenic myxozoans were observed in fish farmed and in fish commonly used in aquariums. Furthermore, myxozoan species were reported in only 2.7% of the more than 4,900 species of fish found in Brazilian waters.
Regarding the sites of infection in the hosts, tropism of the tissue and/or organ of infection was observed in most genera of myxozoans that infect fish. On the other hand, Myxobolus did not show specificity for the sites of infection, being considered a generalist.
There are promising prospects for studies on the life cycle of myxozoans, which may provide indications for interrupting their development in breeding environments. However, as an established activity, actors in the aquaculture production chain must pay attention to new investigations into possible diseases caused by myxozoans that can cause damage to production and preventive management actions. In the context of growing Brazilian aquaculture production, there was a noticeable concentration of publications in the Paraná River basin, in studies of host-parasite interactions in the breeding environment.
This study provides a broad overview of relevant information that can contribute to strategies and actions for the economic development of aquaculture and for the conservation of biodiversity in Brazil. It is suggested that research be expanded in areas with a deficit or absence of studies on myxozoans in fish.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Author contributions
RA: Writing – original draft, Data curation, Formal analysis, Writing – review & editing. MO: Methodology, Writing – review & editing, Formal analysis. AC: Writing – review & editing, Investigation, Data curation. JL: Conceptualization, Investigation, Writing – review & editing. MV: Resources, Supervision, Writing – review & editing. AF: Writing – review & editing, Supervision, Visualization.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Acknowledgments
The authors would like to thank the all members of research group “Sanidade de Organismos Aquáticos da Amazônia” (SOAA) for all support in this study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1630166/full#supplementary-material
References
Adriano E. A., Arana S., Alves A. L., Silva M. R. M., Ceccarelli P. S., Henrique-Silva F., et al. (2009). Myxobolus cordeiroi n. sp., a parasite of Zungaro jahu (Siluriformes: Pimelodiade) from Brazilian Pantanal: Morphology, phylogeny and histopathology. Vet. Parasitol. 162, 221–229. doi: 10.1016/j.vetpar.2009.03.030
Adriano E. A., Arana S., and da Cordeiro N. (2005). An ultrastructural and histopathological study of Henneguya pellucida n. sp. (Myxosporea: Myxobolidae) infecting Piaractus mesopotamicus (Characidae) cultivated in Brazil. Parasite 12, 221–227. doi: 10.1051/parasite/2005123221
Alama-Bermejo G., Holzer A. S., and Bartholomew J. L. (2019). Myxozoan adhesion and virulence: Ceratonova shasta on the move. Microorganisms 7, 1–21. doi: 10.3390/microorganisms7100397
Araujo Neto J. P., Székely C., Molnár K., Pereira C. M. B., Guerreiro S. L., de M., et al. (2024). Morphology and phylogeny of Coccomyxa bragantinensis n. sp. (Cnidaria: Myxozoa) found parasitizing the Coco Sea catfish, Bagre bagre (Siluriformes: Ariidae), captured off the coast of Northern Brazil. Parasitol. Int. 98, 102815. doi: 10.1016/j.parint.2023.102815
Banu H. and Rathinam R. B. (2023). Myxozoan fish diseases: possible treatment and zoonoses. J. Parasitol. Dis. 47, 215–223. doi: 10.1007/s12639-023-01568-9
Barassa B., Cordeiro N. S., and Arana S. (2003). A new species of Henneguya, a gill parasite of Astyanax altiparanae (Pisces: characidae) from Brazil, with comments on histopathology and seasonality. Mem. Inst. Oswaldo Cruz 98, 761–765. doi: 10.1590/S0074-02762003000600009
Bartošová P., Fiala I., Jirků M., Cinková M., Caffara M., Fioravanti M. L., et al. (2013). Sphaerospora sensu stricto: Taxonomy, diversity and evolution of a unique lineage of myxosporeans (Myxozoa). Mol. Phylogenet. Evol. 68, 93–105. doi: 10.1016/j.ympev.2013.02.026
Battazza A., Brasileiro F. C., da S., MaChado E. F., Matos M. G., Santos C. B. T., et al. (2020). Identification and characterization of Sinuolinea niloticus from Nile tilapia (Oreochromis niloticus) farmed in Botucatu, Brazil. Aquac. Int. 28, 1899–1906. doi: 10.1007/s10499-020-00565-6
Capodifoglio K. R. H., Adriano E. A., Naldoni J., Meira C. M., Silva M. R. M., and Maia A. A. M. (2020). Novel myxosporean species parasitizing an economically important fish from the Amazon basin. Parasitol. Res. 119, 1209–1220. doi: 10.1007/s00436-020-06641-3
Capodifoglio K. R. H., Adriano E. A., Silva M. R. M., and Maia A. A. M. (2015). Supplementary data of Henneguya leporinicola (Myxozoa, Myxosporea) a parasite of Leporinus macrocephalus from fish farms in the state of São Paulo, Brazil. Acta Parasitol. 60, 451–458. doi: 10.1515/ap-2015-0062
Dagosta F. C. P. and Pinna M. D.. (2019). The Fishes of the Amazon: distribution and biogeographical patterns, with a comprehensive list of species. Bull. Am. Museum Nat. Hist. 431, 1–163. doi: 10.1206/0003-0090.431.1.1
Dar S. A., Ahmad I., Ahmed I., Kaur H., Khursheed S., Nisar K., et al. (2024). Strategies for describing myxozoan pathogens, dreadful fish diseases in aquaculture. Microb. Pathog. 187, 106512. doi: 10.1016/j.micpath.2023.106512
Dormann C. F. (2011). How to be a specialist? Quantifying specialisation in pollination networks. Netw. Biol. 1, 1–20. doi: 10.0000/issn-2220-8879-networkbiology-2011-v1-0001
Dormann C. F., Gruber B., and Fründ J. (2008). Introducing the bipartite package: analysing ecological networks. R News 8, 8–11.
Eiras J. C., Takemoto R. M., Pavanelli G. C., and Adriano E. A. (2011). About the biodiversity of parasites of freshwater fish from Brazil. Bull. Eur. Assoc. Fish Pathol. 31, 161–168.
Eszterbauer E. (2004). Genetic relationship among gill-infecting Myxobolus species (Myxosporea) of cyprinids: Molecular evidence of importance of tissue-specificity. Dis. Aquat. Organ. 58, 35–40. doi: 10.3354/dao058035
Eszterbauer E., Kallert D. M., Grabner D., and El-Matbouli M. (2009). Differentially expressed parasite genes involved in host recognition and invasion of the triactinomyxon stage of Myxobolus cerebralis (Myxozoa). Parasitology 136, 367–377. doi: 10.1017/S0031182008005398
Eszterbauer E., Sipos D., Forró B., Bartošová P., and Holzer A. S. (2013). Molecular characterization of Sphaerospora molnari (Myxozoa), the agent of gill sphaerosporosis in common carp Cyprinus carpio carpio. Dis. Aquat. Organ. 104, 59–67. doi: 10.3354/dao02584
Eszterbauer E., Sipos D., Kaján G. L., Szegő D., Fiala I., Holzer A. S., et al. (2020). Genetic diversity of serine protease inhibitors in myxozoan (Cnidaria, Myxozoa) fish parasites. Microorganisms 8, 1–14. doi: 10.3390/microorganisms8101502
Fiala I. and Bartoŝová P. (2010). History of myxozoan character evolution on the basis of rDNA and EF-2 data. BMC Evol. Biol. 10, 5–7. doi: 10.1186/1471-2148-10-228
Filizola N. and Guyot J. L. (2009). Suspended sediment yields in the Amazon basin: an assessment using the Brazilian national data set. Hydrol. Process. 23, 3207–3215. doi: 10.1002/hyp.7394
Fontes I., Hallett S. L., and Mo T. A. (2015). ““Comparative epidemiology of myxozoan diseases,”,” in Myxozoan evolution, ecology and development. Eds. Okamura B., Gruhl A., and Bartholomew J. L. (Springer International Publishing), 281–293, Cham). doi: 10.1007/978-3-319-14753-6_15
Froese R. and Pauly D. (2024). FishBase. Available online at: http://www.fishbase.org (Accessed May 21, 2024).
Giulietti L., Karlsbakk E., Cipriani P., Bao M., Storesund J. E., Marathe N. P., et al. (2022). Long-term investigation of the “soft flesh” condition in Northeast Atlantic mackerel induced by the myxosporean parasite Kudoa thyrsites (Cnidaria, Myxozoa): Temporal trends and new molecular epidemiological observations. Fish. Res. 248, 106221. doi: 10.1016/j.fishres.2021.106221
Godoy E. M., David F. S., Fialho N. S., Proença D. C., Camargo T. R., and Bueno G. W. (2022). Environmental sustainability of Nile tilapia production on rural family farms in the tropical Atlantic Forest region. Aquaculture 547, 737481. doi: 10.1016/j.aquaculture.2021.737481
Jorge F. and Poulin R. (2018). Poor geographical match between the distributions of host diversity and parasite discovery effort. Proc. R. Soc B Biol. Sci. 285, 1–9. doi: 10.1098/rspb.2018.0072
Karagouni E., Athanassopoulou F., Lytra A., Komis C., and Dotsika E. (2005). Antiparasitic and immunomodulatory effect of innovative treatments against Myxobolus sp. infection in Diplodus puntazzo. Vet. Parasitol. 134, 215–228. doi: 10.1016/j.vetpar.2005.07.020
Kent M. L. and Martin M. L. (2017). “Farmacologia aplicada à Aquicultura,” in Farmacologia aplicada à Aquicultura., vol. 656 . Eds. Baldisserotto B., Gomes L.C.de, Heinzmann B. M., and da Cunha M. A. (Santa Maria, RS: UFSM).
Kyger R., Luzuriaga-Neira A., Layman T., Sandberg T. O. M., Singh D., Huchon D., et al. (2021). Myxosporea (Myxozoa, Cnidaria) lack DNA cytosine methylation. Mol. Biol. Evol. 38, 393–404. doi: 10.1093/molbev/msaa214
Latrubesse E. M., Arima E. Y., Dunne T., Park E., Baker V. R., D’Horta F. M., et al. (2017). Damming the rivers of the Amazon basin. Nature 546, 363–369. doi: 10.1038/nature22333
Lom J. and Dyková I. (1992). ““Myxosporidia (Phylum Myxozoa),”,” in Protozoan parasites of fishes. Developments in Aquaculture and Fisheries Science. Eds. Lom J. and Dyková I. (Elsevier, Amsterdam), 159–235.
Lom J. and Dyková I. (2006). Myxozoan genera: Definition and notes on taxonomy, life-cycle terminology and pathogenic species. Folia Parasitol. (Praha). 53, 1–36. doi: 10.14411/fp.2006.001
Manrique W. G., Figueiredo M. A. P., de Andrade Belo M. A., Martins M. L., and Azevedo C. (2016). Ultrastructural description of Myxobolus cuneus (Myxosporea) in the skeletal muscle and kidney of tropical farmed fish Piaractus mesopotamicus (Characiformes: Characidae). Parasitol. Res. 115, 2505–2510. doi: 10.1007/s00436-016-5026-1
Martins M. L., de Souza V. N., de Moraes J. R., and de Moraes F. R. (1999a). Gill infection of Leporinus macrocephalus Garavello & Britski 1988 (Osteichthyes: Anostomidae) by Henneguya leporinicola n. sp. (Myxozoa: Myxobolidae). Description, histopathology and treatment. Rev. Bras. Biol. 59, 527–534. doi: 10.1590/s0034-71081999000300018
Matos E., Videira M., Velasco M., Sanches O., São Clemente S. C., and Matos P. (2014). Infection of the heart of Pimelodus ornatus (Teleostei, Pimelodidae), by Myxobolus sp. (Myxozoa, Myxobolidae). Rev. Bras. Parasitol. Vet. 23, 543–546. doi: 10.1590/S1984-29612014083
Matsukane Y., Sato H., Tanaka S., Kamata Y., and Sugita-Konishi Y. (2010). Kudoa septempunctata n. sp. (Myxosporea: Multivalvulida) from an aquacultured olive flounder (Paralichthys olivaceus) imported from Korea. Parasitol. Res. 107, 865–872. doi: 10.1007/s00436-010-1941-8
Meade R. H., Dunne T., Richey J. E., Santos U. D. M., and Salati E. (1985). Storage and remobilization of suspended sediment in the lower amazon river of Brazil. Sci. (80-.). 228, 488–490. doi: 10.1126/science.228.4698.488
Milanin T., Eiras J. C., Arana S., Maia A. A. M., Alves A. L., Silva M. R. M., et al. (2010). Phylogeny, ultrastructure, histopathology and prevalence of Myxobolus oliveirai sp. nov., a parasite of Brycon hilarii (Characidae) in the Pantanal wetland, Brazil. Mem. Inst. Oswaldo Cruz 105, 762–769. doi: 10.1590/S0074-02762010000600006
Moher D., Shamseer L., Clarke M., Ghersi D., Liberati A., Petticrew M., et al. (2015). Preferred reporting items for systematic review and meta-analysis protocols (PRISMA-P) 2015 statement. Syst. Rev. 4, 1–9. doi: 10.1186/2046-4053-4-1
Molnár K. (2002). Site preference of fish myxosporeans in the gill. Dis. Aquat. Organ. 48, 197–207. doi: 10.3354/dao048197
Naldoni J., Adriano E. A., Hartigan A., Sayer C., and Okamura B. (2019a). Malacosporean myxozoans exploit a diversity of fish hosts. Parasitology 146, 968–978. doi: 10.1017/S0031182019000246
Naldoni J., Arana S., Maia A. A. M., Silva M. R. M., Carriero M. M., Ceccarelli P. S., et al. (2011). Host-parasite-environment relationship, morphology and molecular analyses of Henneguya eirasi n. sp. parasite of two wild Pseudoplatystoma spp. in Pantanal Wetland, Brazil. Vet. Parasitol. 177, 247–255. doi: 10.1016/j.vetpar.2010.12.008
Naldoni J., Zatti S. A., da Silva M. R. M., Maia A. A. M., and Adriano E. A. (2019b). Morphological, ultrastructural, and phylogenetic analysis of two novel Myxobolus species (Cnidaria: Myxosporea) parasitizing bryconid fish from São Francisco River, Brazil. Parasitol. Int. 71, 27–36. doi: 10.1016/j.parint.2019.03.009
Okamura B., Gruhl A., and Bartholomew J. L. eds. (2015). Myxozoan evolution, ecology and development., Springer. Cham: Springer International Publishing. doi: 10.1007/978-3-319-14753-6
Okamura B., Hartigan A., and Naldoni J. (2018). Extensive uncharted biodiversity: the parasite dimension. Integr. Comp. Biol. 58, 1132–1145. doi: 10.1093/icb/icy039
Oredalen T. J., Sæbø M., and Mo T. A. (2022). Patterns of Tetracapsuloides bryosalmonae infection of three salmonid species in large, deep Norwegian lakes. J. Fish Dis. 45, 185–202. doi: 10.1111/jfd.13548
Page M. J., McKenzie J. E., Bossuyt P. M., Boutron I., Hoffmann T. C., Mulrow C. D., et al. (2021). The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. BMJ 372, n71. doi: 10.1136/bmj.n71
Poulin R. (2007). Evolutionary ecology of parasites. 2nd (Princeton: Princeton University Press). doi: 10.1515/9781400840809.8
Poulin R., Presswell B., and Jorge F. (2020). The state of fish parasite discovery and taxonomy: a critical assessment and a look forward. Int. J. Parasitol. 50, 733–742. doi: 10.1016/j.ijpara.2019.12.009
R Development Core Team (2017). R: a language and environment for statistical computing (Vienna: R Foundation for statistical computing). Available online at: http://www.r-project.org/.
Rocha E., Matos E., and Azevedo C. (1992). Henneguya amazonica n.sp. (Myxozoa, Myxobolidae), parasitizing the gills of Crenicichla lepidota Heckel 1840 (Teleostei, cichlidae) from amazon river. Eur. J. Protistol. 28, 273–278. doi: 10.1016/S0932-4739(11)80233-6
Rocha S., Rocha F., Casal G., Mendonça I., Oliveira E., Al-Quraishy S., et al. (2019). Supplemental diagnosis and phylogeny of Myxobolus absonus (Cnidaria, Myxozoa) from the eye of the freshwater fish Pimelodus maculatus (Siluriformes, Pimelodidae). Acta Trop. 191, 87–97. doi: 10.1016/j.actatropica.2018.12.032
Stilwell J. M., Camus A. C., Ware C., Walker C. M., Stanton J. B., Leary J. H., et al. (2023). Influence of channel catfish and channel × Blue catfish hybrids on myxozoan community composition in catfish aquaculture ponds. N. Am. J. Aquac. 85, 241–251. doi: 10.1002/naaq.10293
Tavares-Dias M. and Martins M. L. (2017). An overall estimation of losses caused by diseases in the Brazilian fish farms. J. Parasitol. Dis. 41, 913–918. doi: 10.1007/s12639-017-0938-y
Teixeira R. J., Eiras J. C., Spadacci-Morena D. D., Xavier J. G., and Lallo M. A. (2018). Infecção das brânquias de tilápia do Nilo (Oreochromis niloticus) por Myxosporea. Pesqui. Veterinária Bras. 38, 1085–1090. doi: 10.1590/1678-5150-PVB-4859
Videira M., Velasco M., Malcher C. S., Santos P., Matos P., and Matos E. (2016). An outbreak of myxozoan parasites in farmed freshwater fish Colossoma macropomum (Cuvier 1818) (Characidae, serrasalminae) in the amazon region, Brazil. Aquac. Rep. 3, 31–34. doi: 10.1016/j.aqrep.2015.11.004
Vieira D. H. M. D., Pelegrini L. S., Abdallah V. D., Kozlowiski, and De Azevedo R. K. (2019). Parasitology International Supplementary studies on Henneguya guanduensis (Cnidaria: Myxosporea) infecting gills and intestine of Hoplosternum littorale in Brazil: Ultrastructural and molecular data. Parasitol. Int. 70, 27–32. doi: 10.1016/j.parint.2019.01.009
Walliker D. (1969). Myxosporidea of some Brazilian freshwater fishes. J. Parasitol. 55, 942–948. doi: 10.2307/3277155
Keywords: aquaculture, Myxozoa, parasites, pathogenicity, river basins
Citation: Araújo RF, Oliveira MSB, Carvalho AA, Luz JGR, Videira MN and Feiden A (2025) Systematic review of myxozoan occurrence in Brazilian fish. Front. Mar. Sci. 12:1630166. doi: 10.3389/fmars.2025.1630166
Received: 06 June 2025; Accepted: 22 July 2025;
Published: 26 August 2025.
Edited by:
Daniel Cerqueda-García, Instituto de Ecología (INECOL), MexicoReviewed by:
Archana Sinha, Central Inland Fisheries Research Institute (ICAR), IndiaAmelia Paredes-Trujillo, Universidad de Ciencias y Artes de Chiapas Centro Universitario de Informacion y Documentacion, Mexico
Copyright © 2025 Araújo, Oliveira, Carvalho, Luz, Videira and Feiden. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rafaela Franco Araújo, YXJhdWpvcmZkZUBnbWFpbC5jb20=; Marcela Nunes Videira, bWFyY2VsYS52aWRlaXJhQHVlYXAuZWR1LmJy