 Carla Riva-Rossi1
Carla Riva-Rossi1 Javier Ciancio2*
Javier Ciancio2* Daniel Gomez-Uchida3Miguel Pascual4Rodrigo Clarke5Pamela Quiroga1Cecilia Di Prinzio6
Daniel Gomez-Uchida3Miguel Pascual4Rodrigo Clarke5Pamela Quiroga1Cecilia Di Prinzio6 Carolina Lázari7
Carolina Lázari7 John Carlos Garza8
John Carlos Garza8- 1Instituto de Diversidad y Evolución Austral (IDEAus-CONICET), Chubut, Puerto Madryn, Argentina
- 2Centro para el Estudio de Sistemas Marinos (CESIMAR-CONICET), Puerto, Madryn, Chubut, Argentina
- 3Departamento de Zoología, Facultad de Ciencias Naturales y Oceanográficas, Universidad de Concepción & Núcleo Milenio INVASAL, Concepción, Chile
- 4Instituto Patagónico para el Estudio de los Ecosistemas Continentales (IPEEC-CONICET), Puerto Madryn, Chubut, Argentina
- 5Secretaría de Estado de Pesca y Acuicultura, Fundación Anfibia, Santa Cruz, Argentina
- 6Centro de Investigación Esquel de Montaña y Estepa Patagónica (CONICET-FCNyCS- LIESA-UNPSJB), Esquel, Chubut, Argentina
- 7Ocean Sciences Department, University of California Santa Cruz, Santa Cruz, CA, United States
- 8Southwest Fisheries Science Center, National Oceanic and Atmospheric Administration, Santa Cruz, CA, United States
Introduction: Biological invasions are major drivers of biodiversity loss worldwide, and salmonid introductions are among the most transformative events in the Southern Hemisphere. The rapid, large-scale expansion of Chinook salmon (Oncorhynchus tshawytscha) across South America, driven by high migratory capacity, straying rates, and genetic adaptability, constitutes an unprecedented salmonid invasion. In Patagonia, this spread is accelerating, yet the mechanisms enabling long-distance marine dispersal, successful freshwater colonization, and secondary expansion remain poorly understood. A recently established population in the De las Vueltas River (DLVR), an upper tributary of the Santa Cruz River in Argentine Patagonia, was examined as a key node in the invasion network connecting Pacific-origin populations with new Atlantic Ocean basins.
Methods: Two high-resolution SNP panels (96 and 172 loci) were used to investigate the genetic origins and colonization dynamics of the DLVR population. Tissue samples from 70 fish collected in 2014–2015 were genotyped and compared with North American source lineages to infer ancestral origins, and with Pacific and Atlantic naturalized populations to reconstruct recent invasion pathways. Discriminant analysis of principal components (DAPC), genetic mixture modeling, and Bayesian assignment implemented in the R package rubias were performed. Simulations were used to harmonize SNP data across panels and to improve assignment accuracy.
Results: The genetic roots of the DLVR population were traced to stocks from the Lower Columbia River fall and spring runs and the Willamette River spring run. Strong genetic affinities were detected with naturalized populations from Chile’s Aysén Region, particularly the Cobarde and Vargas Rivers, while smaller contributions from Santa Cruz River populations were inferred. These findings indicate colonization through long-distance oceanic or trans-Andean dispersal, followed by secondary expansion within the Santa Cruz basin.
Discussion: The role of ocean connectivity, stepping-stone habitats, and leading-edge dispersal in enabling the rapid eastward spread of Chinook salmon is highlighted by these results. The Santa Cruz River basin is identified as a critical invasion hub, concentrating propagules from multiple lineages and promoting multi-step dispersal into new Atlantic Ocean basins. These insights emphasize the need to incorporate riverine connectivity and invasion hubs into management strategies to mitigate the ecological and evolutionary impacts of Chinook salmon in South America.
Introduction
Biological invasions have accelerated dramatically in the Anthropocene, driven by globalization, trade, and human-induced habitat changes (Hulme, 2009; Pyšek et al., 2008; Seebens et al., 2017). Species with high dispersal capacity are particularly prone to becoming invasive, as their ability to move across heterogeneous landscapes facilitates colonization of new habitats, rapid range expansion, and establishment of self-sustaining populations (Clobert et al., 2012; Nathan and Muller-Landau, 2000). Introductions may be intentional, aiming to provide economic, recreational, or cultural benefits (Gozlan, 2008), or unintentional, occurring as by-products of trade and human activity (Hulme, 2009). Regardless of the vector, highly dispersive species can profoundly alter ecosystems by competing with, preying upon, or transmitting diseases to native species, modifying habitats, and reshaping nutrient and energy flows (Simberloff et al., 2013; Vitule et al., 2009). These ecological effects often translate into socio-economic consequences, including impacts on fisheries, aquaculture, and ecosystem services (Lovell et al., 2006; Pejchar and Mooney, 2009). Monitoring dispersal and establishment of such species is therefore crucial for predicting and managing ecological and economic outcomes (Early et al., 2016).
Successful establishment after dispersal, however, is not guaranteed. It requires compatibility between species’ physiological tolerances and local environmental conditions, access to adequate prey, and the absence of strong biotic resistance (Shea and Chesson, 2002). In diadromous fishes, which alternate between freshwater and marine phases, dispersal outcomes are further constrained by the interplay between ocean currents, prey availability and movements, and the timing of reproductive migrations. These factors directly affect components of fitness such as growth, fecundity, and survival, thereby shaping long-term establishment potential (Pascual and Ciancio, 2007; McDowall, 2002).
Chinook salmon (Oncorhynchus tshawytscha), the largest Pacific salmon, has established self-sustaining populations well beyond its native range in western North America and northeastern Asia, now occurring in the Great Lakes, New Zealand, and multiple Pacific Ocean and Atlantic Ocean basins of southern South America (Busby et al., 1997; Quinn et al., 2001; Correa and Gross, 2008). Their invasion success is linked to complex life-history strategies: anadromous adults spawn in freshwater, juveniles migrate to the ocean for one or several years, and adults return to natal rivers with strong—but not absolute—homing fidelity, allowing occasional straying and colonization of new watersheds (Quinn, 2018; Keefer and Caudill, 2014; Gomez-Uchida et al., 2018). Variation in migration timing and reproductive strategies results in the expression of at least two main ecotypes: “spring-run”, which enter rivers immature and hold in freshwater before spawning, and “fall-run,” which migrate later and spawn soon after river entry. This phenotypic plasticity, combined with high fecundity and broad environmental tolerance, has important repercussions for dispersal dynamics, colonization success, and genetic structure across diverse non-native ecosystems (Healey, 1991; Quinn et al., 2006; Beckman, 2011).
In South America, this invasion represents a large-scale natural experiment combining human-mediated introductions and translocations with subsequent natural dispersal, where human actions altered both the scale and speed of expansion (Correa and Gross, 2008; Wilson et al., 2016). In southern Chile, salmon were introduced for aquaculture and recreational fishing, with sources from the Pacific Northwest and New Zealand (Soto et al., 2007; Correa and Gross, 2008; Riva-Rossi et al., 2012). Stockings in the 1970s–1980s reinforced by net-pen escapes during the 1990s, gave rise to multiple donor-derived populations. Northern basins (Araucanía, Los Lagos) retain higher diversity through admixture, while southern populations (Aysén, Magallanes, Atlantic drainages) are more homogeneous due to secondary dispersal and drift (Riva-Rossi et al., 2012; Gomez-Uchida et al., 2018).
Spread has also been shaped by oceanography. Fjords and archipelagos of southern Chile provided stepping-stone habitats (Antezana, 1999), while large-scale dispersal followed major currents: the Humboldt along the Pacific Ocean and the Brazil–Malvinas confluence on the Atlantic Ocean (Montecino and Lange, 2009; Acha et al., 2004). These systems both constrain and facilitate range expansion. Early models predicted establishment success would hinge on latitude: smolts north of ~40°S would be entrained in the Humboldt Current, whereas those farther south could access the productive Patagonian Shelf (Donaldson and Joyner, 1984). Subsequent observations confirm this, as southern stocks now sustain fast-growing, highly fecund populations often exceeding native performance (Ciancio et al., 2005; Correa and Moran, 2017). This outcome illustrates how oceanography, dispersal, and life-history traits interact to determine invasion trajectories in diadromous fishes.
Over the past four decades, Chinook salmon has colonized rivers between 32° and 52°S along both Pacific and Atlantic slopes. Consequently, their current distribution far exceeds early predictions: the species now occupies at least 48 basins, with reproduction confirmed in more than half, ranging from the Imperial River (39°S) to the Lapataia River (55°S), and extending even to coastal lagoons in Uruguay and Brazil (32°S) (Figueroa-Muñoz et al., 2023; Di Prinzio et al., 2024; Gonçalvez Gowert et al., 2025). While supporting local economies through aquaculture, recreational, and artisanal fisheries (Pascual et al., 2009; García de Leaniz et al., 2010), Chinook salmon also impacts native freshwater fishes and ecosystems by altering nutrient fluxes, trophic interactions, and contaminant dynamics (Vargas et al., 2010; Arismendi et al., 2014; Figueroa-Muñoz et al., 2021, 2022). This rapid expansion reflects the combined effects of propagule pressure, genetic diversity, oceanographic transport, and life-history plasticity. Ocean connectivity thus acts both as a dispersal vector and a structuring force for genetic variation. On the other hand, high fecundity, flexible migration and spawning strategies, broad thermal tolerance, abundant spawning habitats, and low competition or predation have also collectively facilitated rapid niche shifts and successful establishment across Patagonia (Quinn et al., 2001; Pascual et al., 2002; Correa and Gross, 2008; Ciancio et al., 2015; Gomez-Uchida et al., 2018).
The Santa Cruz River, a major Atlantic-draining system, provides a prime setting to explore these processes. In this basin, spawning Chinook salmon were first detected in the early 1980s in the Caterina River, an upper tributary located 500 km upstream from the river mouth (Ciancio et al., 2005; Becker et al., 2007). Subsequent reports in the De las Vueltas River, another upper-basin tributary, suggest ongoing expansion through either local straying or long-distance colonization (Ciancio et al., 2015; Gomez-Uchida et al., 2018). Understanding the genetic origin, diversity, and dispersal dynamics of these populations may provide insights into how ocean currents, fitness consequences, and human-mediated introductions interact to shape the dispersal, colonization, and establishment of diadromous invaders in the Southern Hemisphere.
The specific objectives of this study were to determine the genetic relationships between the De las Vueltas River (DLVR) population and other naturalized and aquaculture stocks across South America, assess the role of multiple source populations in maintaining genetic diversity, and reconstruct colonization pathways. The hypothesis of this study were: 1) the DLVR population originated from contiguous populations within the Santa Cruz basin (e.g., Santa Cruz and Caterina rivers), rather than from an independent colonization event; 2) colonization occurred primarily via short-distance intra-basin dispersal, not through long-distance oceanic dispersal from Pacific sources; and 3) consequently, its genetic diversity is limited by founder effects rather than enhanced by admixture from multiple, genetically diverse sources. These questions were addressed by comparing the genetic composition of DLVR to other naturalized populations using two highly informative SNP panels (Ciancio et al., 2015; Di Prinzio et al., 2015; Gomez-Uchida et al., 2018), enabling both individual- and population-level analyses.
Materials and methods
Background on historical introductions and genetic legacy of Chinook salmon in South America
Review works of Basulto, 2003; Becker et al., 2007; Soto et al., 2007; Correa and Gross, 2008; Fernández et al., 2010; Riva-Rossi et al., 2012; Ciancio et al., 2015; Di Prinzio et al., 2015; Correa and Moran, 2017; Gomez-Uchida et al., 2018 were used to obtain Chinook salmon introduction information and its genetic legacy in Chile and Argentina. Historical stocking efforts can be grouped into three main phases. Early attempts between 1886 and 1930 relied on California Central Valley fall-run stocks (McCloud River hatchery), but no successful establishment was detected. The ocean ranching era (late 1970s–1980s) involved large-scale stocking of multiple donor lineages from North America, particularly Lower Columbia River spring (Washington/Oregon), fall-run populations from Puget Sound (Washington), and several coastal Oregon and Washington stocks. These introductions led to the successful establishment of high-diversity artificial populations in southern Chile, especially in the Petrohué (41°S), Cobarde (45°S), Pichicolo (41°S), and Prat (51°S) rivers, with subsequent dispersal into Argentinian rivers such as the Santa Cruz (50°S) (Figure 1). A third phase occurred in the 1990s–2000s, when escapes from net-pen aquaculture operations introduced additional lineages from the Vancouver Island (British Columbia, Canada), Puget Sound, and New Zealand-derived California stocks, further contributing and reinforcing the genetic diversity of established populations, particularly in northern locations (39°-43°S). Genetic studies demonstrate that at least six donor regions contributed to present-day Patagonian Chinook salmon, with varying levels of admixture depending on propagule pressure and secondary dispersal. A synthesis of introduction phases, donor stocks, stocking sites, and their genetic outcomes and present-day distribution is provided in Table 1.
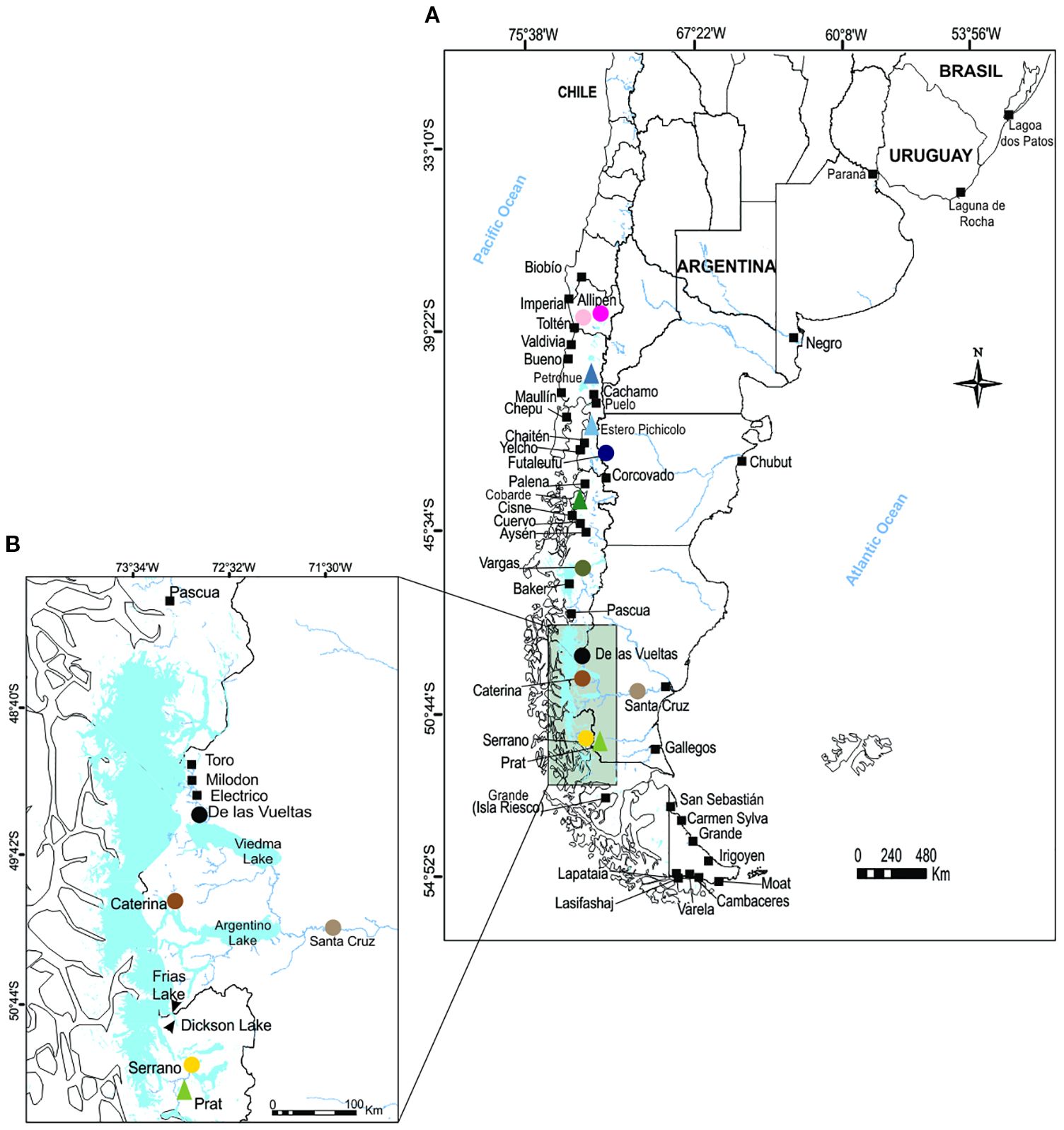
Figure 1. (A) Contemporary distribution of Chinook salmon in South America and sampling locations in Patagonia. Circles indicate naturalized Chinook salmon populations that were genetically sampled, while triangles represent artificial stocks included in the genetic analyses. Squares indicate locations where Chinook salmon presences or spawning populations were reported by Baigún et al., 2022; Figueroa-Muñoz et al. 2023; Espínola et al., 2024). (B) Inset map of the Lake Argentino–upstream Santa Cruz River basin (Argentina) and the Paine–Dickson–Serrano River system (Chile), showing potential dispersal corridors that may facilitate the trans-Andean eastward expansion of Chinook salmon from Pacific Ocean and Atlantic Ocean basins. Colors of the symbols representing genetically analyzed populations are consistent across figures to facilitate their identification in the plots.
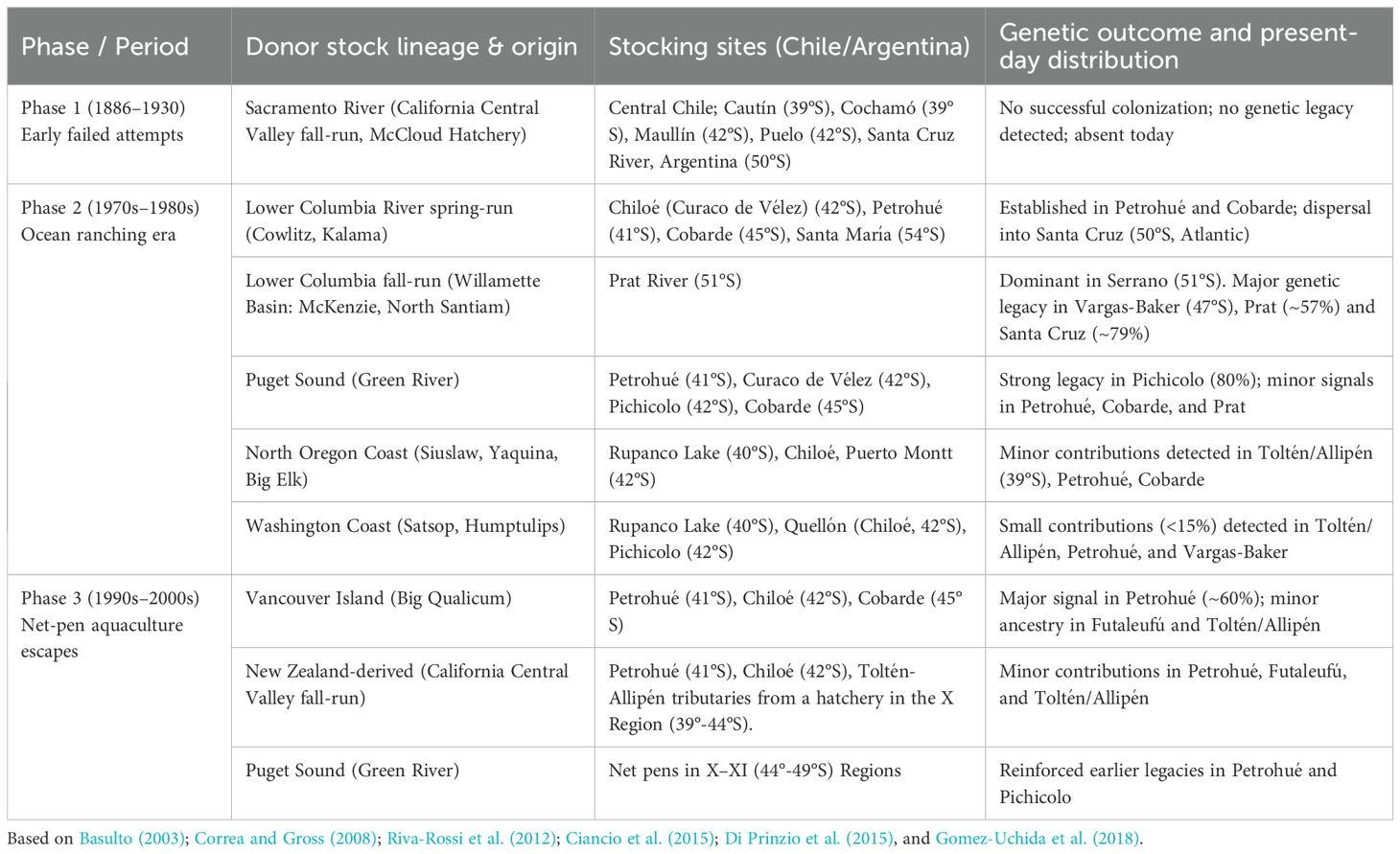
Table 1. Historical stocking efforts of Chinook salmon in South America: phases, donor lineages, stocking sites, genetic outcome and present-day distribution.
Study area and sample collection
For this study, genetic samples were collected exclusively from the De las Vueltas River (DLVR; 49°S), a tributary of Viedma Lake in the upper Santa Cruz River basin. A total of 70 adult individuals were obtained during 2014–2015 through gillnetting, carcass collection, electrofishing, and angling. Fin or muscle tissue was preserved in 95% ethanol for subsequent genetic analyses.
The DLVR drains a 1,283 km² watershed originating in Desierto Lake (506 m a.s.l.) and flowing ~70 km southward into Viedma Lake (250 m a.s.l.). The basin spans a forest–steppe ecotone under a cold-temperate, humid to sub-humid climate with high precipitation (~1,400 mm/year). Hydrological inputs are dominated by snowmelt and glacial runoff, with marked seasonal variation in discharge. Despite its glacial origin, the headwaters lack direct inputs from the Southern Patagonian Icefield, resulting in comparatively clearer and warmer waters. Continuous hydrological connectivity with Andean headwaters provides suitable spawning habitat and a potential dispersal corridor for non-native salmonids, although planned hydropower development in the lower Santa Cruz River may restrict future passage.
Previously published SNP datasets were incorporated to place the DLVR population within the broader context of Chinook salmon invasions in Patagonia. These datasets had been standardized by Gomez-Uchida et al. (2018), who genotyped 9 Patagonian populations using a 172-SNP panel (Supplementary Table S1). Samples originally collected by Riva-Rossi et al. (2012) between 2005 and 2009 included two introduction sites in Chile—Cobarde River (N = 36) and Prat River (N = 30)—and naturally colonized rivers Vargas (N = 24) and Serrano (N = 15) in Chile, and Caterina (N = 45) and Santa Cruz mainstem (N = 18) in Argentina, together with a hatchery broodstock at Estero Pichicolo (N = 25). These samples were obtained from adults by gillnetting, angling, and carcass recovery, and tissues were preserved in ethanol. Additional populations genotyped by Gomez-Uchida et al. (2018) included fish sampled later: adult carcasses from Petrohué River (N = 70; 2013) and juvenile parr from the Allipén (N = 26; 2014) and Toltén rivers (N = 52; 2014). This data were downloaded from Dryad (doi.org/10.5061/dryad.5k45n83).
From the Futaleufú River (43°S), which drains into the Pacific Ocean in southern Chile, the dataset published by Di Prinzio et al. (2015) was incorporated. In this case, 53 adult Chinook salmon were captured by recreational fishers during 2010–2011 in the Argentinean section of the river, and muscle tissue was preserved in 95% ethanol.
Together, the DLVR samples collected in this study and the previously published datasets provided a comprehensive baseline covering both introduction sites and naturalized populations of Chinook salmon across Pacific Ocean and Atlantic Ocean basins in Patagonia (Figure 1).
Laboratory procedures
DNA from the De las Vueltas River samples was extracted using DNeasy 96 Tissue kits on a BioRobot3000 (QIAGEN, Inc., Valencia, CA). Genotypes were collected from 96 SNP loci (Table 1) utilizing 96.96 SNP Genotyping Dynamic Arrays™ on an EP1 system (Fluidigm Corporation, South San Francisco, CA) following the manufacturer’s protocols. Genotyping was conducted using Fluidigm SNP Genotyping Analysis Software (version 2.1.1). The panel comprised 95 loci polymorphic in Chinook salmon and one locus for identifying Coho salmon (O. kisutch) (Clemento et al., 2014) (Supplementary Table S1).
Genotype quality
All reference individuals from Clemento’s (96-SNP, N = 8,031) and Gomez-Uchida’s (172-SNP, N = 341) baselines were retained for downstream analyses (Supplementary Table S1). DLVR samples with genotyping success below 95% or with >10% missing data were excluded, which led to the removal of six loci: Ots_108007-208, Ots_112208-722, Ots_113242-216, Ots_Myc-366, Ots_RAG3 (shared by both panels), and Ots_P50 (172-SNP panel only) and leaving only 66 loci for downstream analysis.
Statistical analysis
Genetic variation in DLVR Chinook salmon was analyzed in GenAlEx 6.5 based on 66–72 SNP loci common to both panels (Clemento et al., 2014; Gomez-Uchida et al., 2018) (Supplementary Table S1). Exact tests for departures from Hardy–Weinberg equilibrium (HWE) and linkage disequilibrium (LD) were conducted, applying a Bonferroni correction (α = 0.05), and GenAlEx 6.5 (Peakall and Smouse, 2012) was used to calculate the number of alleles (Na), observed (HO) and expected heterozygosity (HE), and the percentage of polymorphic loci.
Ancestral and recent origins of DLVR individuals were inferred using the 96-SNP (Clemento et al., 2014) and 172-SNP panels (Warheit et al., 2012; Gómez-Uchida et al., 2018), respectively, with the Bayesian mixture model implemented in the R package rubias (Moran and Anderson, 2019) in R version 4.3.3 (R Core Team, 2023). This framework allowed assignment of individuals to North American reporting units (ancestral origins) and Patagonian baseline populations (recent origins), correcting for unequal representation among source populations. Baseline accuracy was evaluated through self-assignment and simulated mixture tests. Assignment reliability was assessed using posterior probability thresholds ≥ 0.75 and z-scores > -6 for high-confidence assignments (Anderson E., personal communication, March, 2015; Di Prinzio et al., 2015). For recent origins, because DLVR and Futaleufú individuals were successfully genotyped at only 66 and 72 loci, respectively, of the 172-SNP baseline (i.e., after genotype quality analysis), missing genotypes were imputed using reference-based simulations guided by a Discriminant Analysis of Principal Components (DAPC) in the R package adegenet (Jombart et al., 2010) in R version 4.3.3 (R Core Team, 2023). Results are reported only for real individuals. Population structure across Patagonian sites was examined using DAPC (Jombart et al., 2010) to identify reference groups and assess genetic similarity.
Additional methodological details, including genotype quality, MCMC settings, bootstrap replicates, genotype imputation, and assessment of baseline accuracy through simulated mixtures, are provided in the Supplementary Methods.
Results
The genetic composition, population structure, and colonization patterns of Chinook salmon in Patagonia are summarized below, highlighting key findings regarding diversity, admixture, and dispersal, with particular emphasis on the ancestral lineages and recent contributions to the DLVR population.
Genetic origins
Using the 96-SNP panel developed by Clemento et al. (2014), rubias mixture analyses assigned DLVR Chinook salmon primarily to three lineages: Lower Columbia River fall (36 individuals), Lower Columbia River spring (14), and Willamette River spring (19). Assignment probabilities were generally high, with most individuals showing posterior probabilities ≥ 0.75 (Table 2). Z-scores ranged from –5.19 to 1.30 (mean –1.32), with 63 individuals exceeding the –3 threshold. One individual was identified as a Coho salmon and was removed from subsequent analyses.
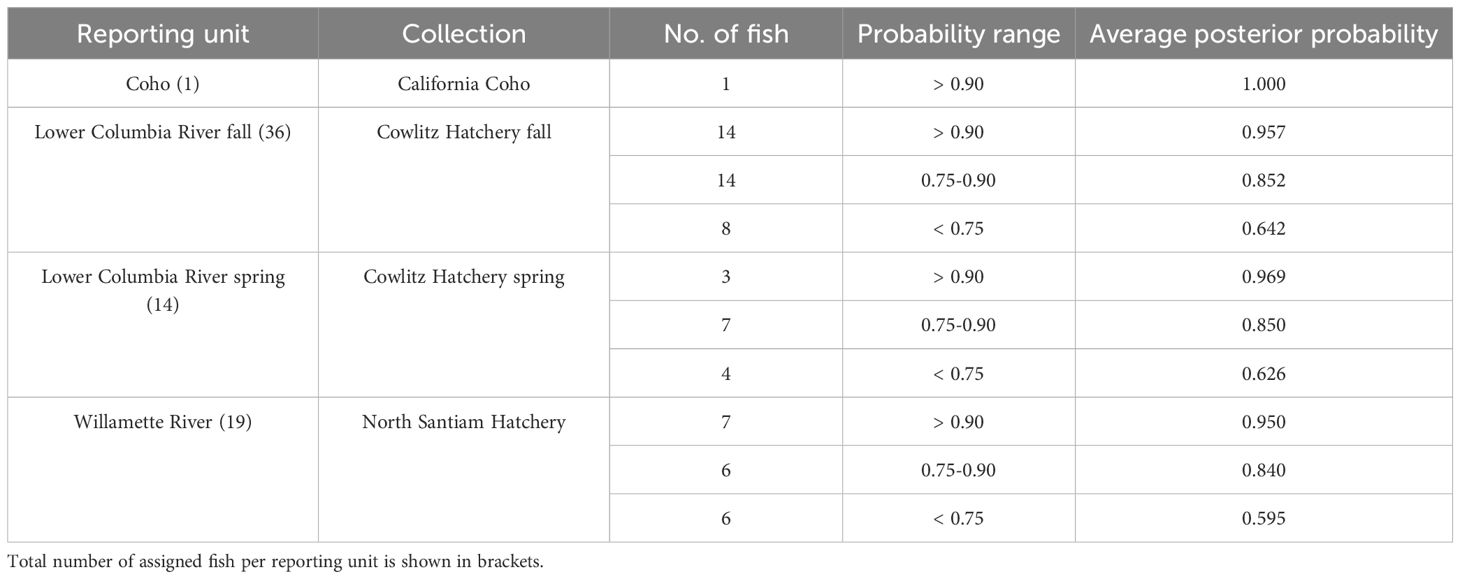
Table 2. Identification of the ancestral roots of DLVR Chinook salmon using rubias.
To investigate the recent origins of DLVR salmon, we compared them with reference populations from Patagonia. DAPC analyses grouped reference populations into four main geographic clusters (Figures 1, 2): Araucanía, Los Lagos, Aysén (including Prat and Serrano), and the Santa Cruz basin in the Atlantic Ocean. Leave-one-out simulations confirmed the resolution of this baseline with high reassignment accuracy (overall 98%, range 95–100%): Araucanía and Los Lagos achieved 100%, Aysén 96%, Santa Cruz 95%, Caterina 93%, while Cobarde showed the lowest value (89%). Among clusters, Araucanía was the most divergent, whereas Los Lagos and Aysén were more closely related, reflecting shared broodstock ancestry. Santa Cruz populations were highly differentiated from all Chilean groups. Within this structure, DLVR clustered almost entirely with the Aysén cluster, with only a few individuals aligning with Santa Cruz (Figure 2), supporting a Pacific rather than local origin.
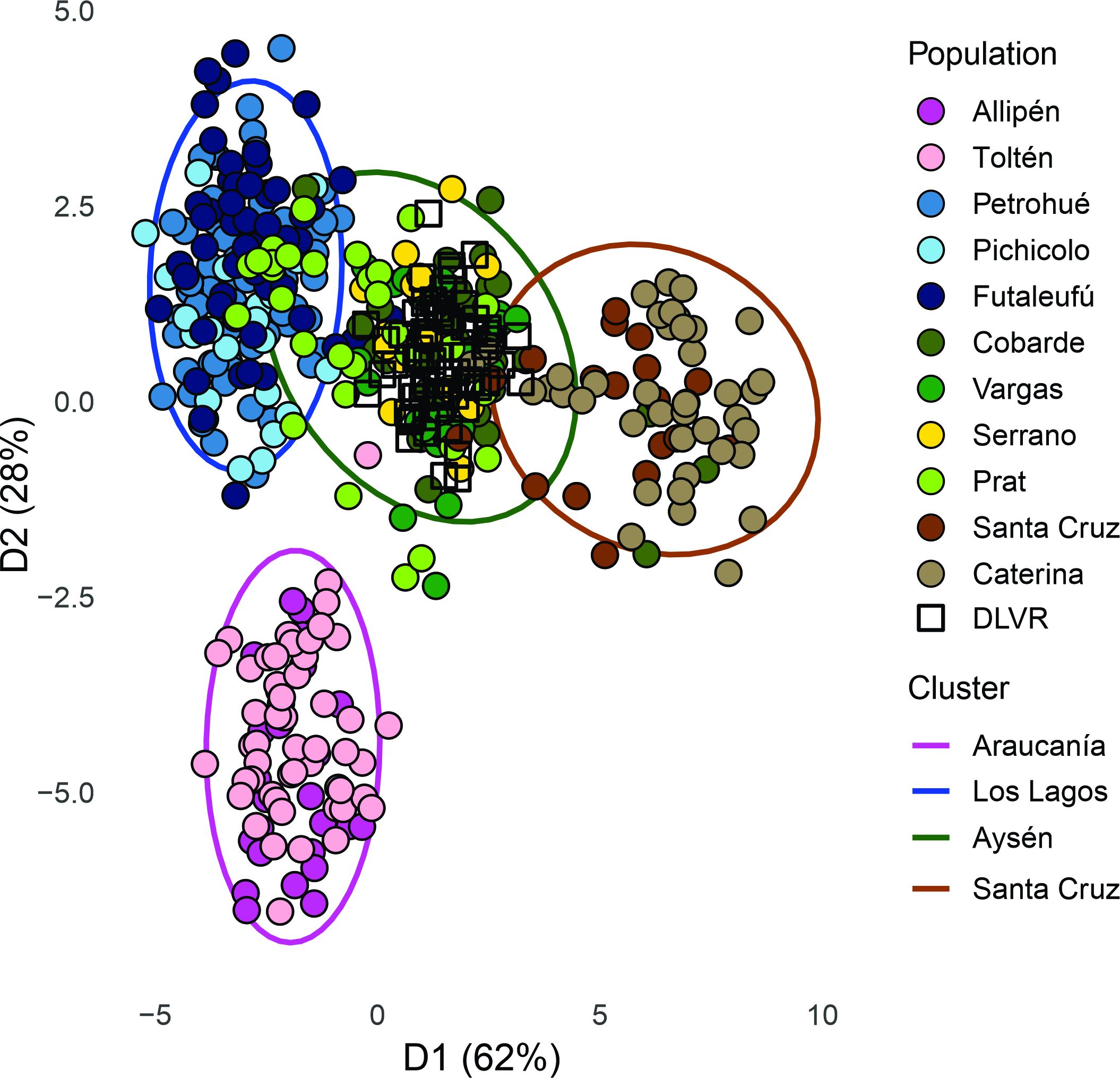
Figure 2. Scatter plot of the discriminant analysis of principal components (DAPC). Each of the four identified clusters is depicted by distinct color inside their 95% inertia ellipses. Colors represent the different clusters and populations within clusters. Dots represent individuals. Squares represent assigned De las Vueltas River (DLVR) individuals. Colors represent the different populations within clusters. The axes represent the first two discriminant functions, respectively.
Simulated mixture analyses in rubias further confirmed the reliability of this four-cluster model, accurately recovering the proportions of each source population without significant bias (Supplementary Figure S1). In contrast, defining a fifth cluster for Prat and Serrano (Magallanes) reduced accuracy: overall assignment dropped to 96%, with only 82% for these rivers, and mixture simulations underestimated contributions from both Aysén and Magallanes (Supplementary Figure S2).
Overall, these results support the robustness of the four-cluster model, showing that it provides a reliable framework for detecting the genetic origins of DLVR individuals. They also highlight that geographic clustering alone may obscure underlying genetic similarities, particularly between neighboring regions such as Aysén and Magallanes.
Genetic diversity
Beyond their origins, the retention of sufficient genetic variation to support the establishment of DLVR salmon was evaluated. No significant departures from Hardy–Weinberg equilibrium (HWE) were detected at any of the 72 loci after Bonferroni correction, and linkage disequilibrium was observed in only one pair of loci. The inbreeding coefficient was slightly negative (FIS = –0.004), indicating a marginal excess of heterozygotes and limited inbreeding. Genetic diversity was moderate to high (Ho = 0.36; He = 0.34), and levels in DLVR were comparable to those of its main source populations, Cobarde and Vargas (Ho = 0.37–0.35; He = 0.37–0.36), slightly higher than some nearby naturalized populations such as Santa Cruz and Caterina, and somewhat lower than in long-established hatchery-influenced populations like Pichicolo and Petrohué (Table 3). When compared with founding lineages from the Pacific Northwest (average Ho = 0.37 and He = 0.36; Ciancio et al., 2015), DLVR genetic diversity fell within the observed range, indicating that the recently established population retains substantial genetic variation sufficient for successful establishment.
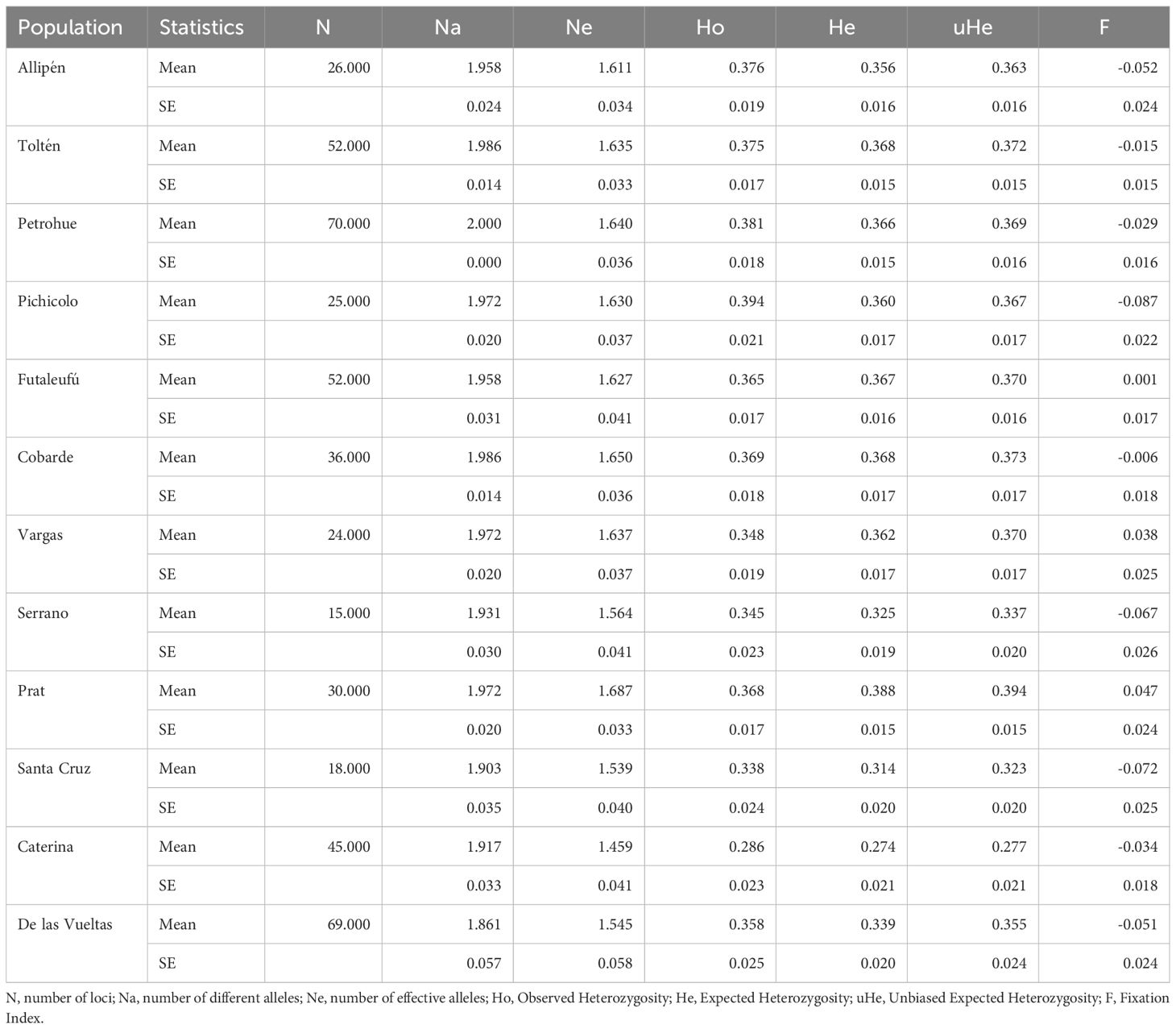
Table 3. Gene diversities per pop across loci for the 72-SNP merged panel.
Colonization and dispersal pathways
Results of DAPC analysis revealed that within clusters, gene flow was evident from the high overlap of population genotypes, particularly among adjacent populations (Figure 2). This was especially noticeable between artificially established populations (Cobarde and Prat) and naturalized populations (Vargas and Serrano) in the Aysén and Magallanes area. Recent common ancestry was also evident between the Prat and Petrohué Rivers, as many individuals from Prat clustered within the Los Lagos group (consistent with artificial propagation programs from Petrohué and Curaco de Vélez stocks). Gene flow was also detected from the Futaleufú River toward both the Aysén region and the Petrohué River, as well as long-distance dispersal between Aysén and the Santa Cruz River basin. In contrast, no evidence of dispersal was observed between the Araucanía populations and those from other regions.
Mixture proportion analysis using rubias assigned most DLVR individuals to the Aysén cluster (56 individuals, 81%; 46% to Cobarde, 35% to Vargas) and the remainder to the Santa Cruz cluster (13 individuals, 19%). Assignment probabilities were generally high, with the majority of individuals in both clusters showing posterior probabilities ≥ 0.75. Z-scores ranged from –2.77 to 3.42 (mean –0.25), with all individuals exceeding the –3 threshold (Table 4). These results highlight the dispersal pattern from Aysén toward the Santa Cruz basin and the contribution of genetic diversity in source populations to the successful establishment at the most distant margin of the watershed.
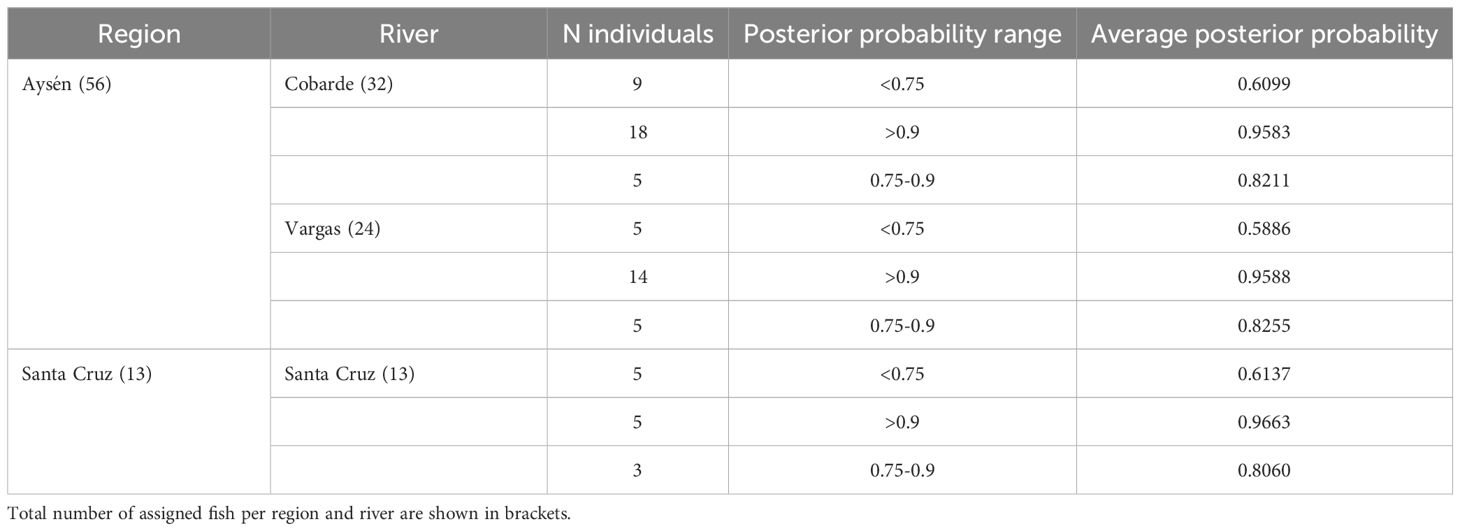
Table 4. Identification of the founding populations of DLVR Chinook salmon using rubias.
Discussion
The invasion of Chinook salmon in South America results from repeated introductions, high propagule pressure, and strong dispersal capacity. Intrinsic traits such as straying, migration, and life-history plasticity promoted admixture and range expansion beyond release sites. Since the end of ocean ranching in 2013, natural dispersal has dominated colonization, supporting establishment in both Pacific- and Atlantic-draining basins while maintaining high genetic diversity (Ciancio et al., 2005; Correa and Gross, 2008; Riva-Rossi et al., 2012; Gomez-Uchida et al., 2018; Figueroa-Muñoz et al., 2023).
Genetic origins
Chinook salmon were introduced into southern South America in three main phases. Early attempts (<1930s), largely using Sacramento River stocks, failed to establish populations in central and southern Chile. Ocean ranching in the 1970s–1980s, based on multiple stocks from the Lower Columbia, Puget Sound, Oregon and Washington, successfully established populations in Los Lagos, Aysén, and Magallanes. Net-pen aquaculture escapes in the 1990s–2000s, mainly from Vancouver Island, Puget Sound, and New Zealand–derived California stocks, reinforced these populations, especially in Los Lagos. In Argentina, early stockings in the Santa Cruz basin (1906–1910) were unsuccessful, and no further intentional releases of Chinook salmon were carried out in this basin. However, by the late 1990s a spawning population was detected in the Caterina River, which originated from Prat River ranching experiments in Chile during the 1980s (Ciancio et al., 2005; Becker et al., 2007).
First recorded in 2006 and now widespread throughout the sub-basin and nearby Andean headwaters, this study revealed that the De las Vueltas River population originated predominantly from long-established Chilean populations in the Cobarde and Vargas Rivers (Aysén cluster), with a smaller contribution from the Santa Cruz cluster. Despite geographic proximity, DLVR does not derive mainly from neighboring Santa Cruz populations, which might have been expected. These results indicate that DLVR arose through gene flow from both distant, long-established Chilean populations and more recent Santa Cruz populations, with Chilean input predominating.
These affinities are embedded within the four-cluster genetic framework corresponding to major geographic and historical introduction zones: Araucanía (Toltén and Allipén Rivers), Los Lagos (Petrohué River and Estero Pichicolo), an “extended Aysén” group (Cobarde, Vargas, Prat, and Serrano Rivers), and Santa Cruz (Santa Cruz and Caterina Rivers) (Gomez-Uchida et al., 2018; Musleh et al., 2020; this study). As expected from their recent origins in Patagonia, DLVR reflects the genetic legacy of founding populations from the Aysén region, while Santa Cruz and Caterina show affinities with sources from Magallanes. Accordingly, DLVR shares the same ancestral background as its founder populations, which ultimately trace back to Lower Columbia River fall and spring lineages, with minor contributions from the Willamette River spring lineage. This shared ancestry among geographically distant regions, such as Aysén (Chile), Santa Cruz (Argentina), and Magallanes (Chile), further explains their observed genetic similarities (Gomez-Uchida et al., 2018). This genetic pattern highlights how common ancestry from original introductions, subsequent admixture, secondary long-distance dispersal and ongoing ocean-mediated connectivity collectively shape the regional population structure across the invasive range.
While genetic analyses outline the broader population structure and ancestry of Chinook salmon, independent evidence from otolith isotope chemistry converges on a similar picture about the mixed origins at DLVR. In particular, Avigliano et al. (2025) showed that only one-third of individuals in the DLVR system were reassigned within its tributaries, with the majority traced to other rivers such as Guanaco, La Leona, and the mainstem Santa Cruz. Genetic evidence likewise places DLVR populations closer to those from Santa Cruz and Aysén, suggesting that De las Vueltas acts less as an isolated breeding unit and more as a demographic sink shaped by multiple dispersal pathways. Importantly, dispersal is captured in complementary but distinct ways by these markers: otolith chemistry provides a short-term environmental snapshot of natal origins, whereas SNPs reveal longer-term, microevolutionary connectivity among populations. Similar integrative approaches have been employed in other systems: for instance, dispersal capacities in Allis shad (Allosa sapidisima) were inferred by Martin et al. (2015) using otolith microchemistry and genetics, while dispersal in anadromous brown trout was tracked by Källo et al. (2023) with the same combination of methods. As in those cases, the congruence between otolith- and SNP-based inferences has been interpreted here as reinforcing the conclusion that DLVR dynamics are shaped by recurrent immigration, source mixing, and the interplay between environmental filtering and evolutionary processes.
The heterogeneous origins of the DLVR population, shaped by contributions from multiple source populations and ongoing dispersal across the Santa Cruz basin, have contributed to its unexpectedly high genetic diversity. Rather than exhibiting the reduced variation typically expected for a recently established population, DLVR shows a slight excess of heterozygotes, indicative of recent admixture or continuing influx of migrants. This pattern underscores the dynamic nature of the invasion front, where secondary dispersal and connectivity among distant and neighboring populations maintain and even enhance genetic variation (Dlugosch and Parker, 2007). Similar processes have been observed in other aquatic invaders, including sea-run brown trout (Salmo trutta) in Argentina, rainbow trout (Oncorhynchus mykiss) in Chile, European green crab (Carcinus maenas) in North America, and topmouth gudgeon (Pseudorasbora parva) in Europe, highlighting the general role of admixture and repeated introductions in promoting establishment, spread, and rapid adaptation (Consuegra et al., 2011; Darling et al., 2008; Gozlan et al., 2010; Giese, 2024).
Overall, these examples reflect a broader trend observed across invasive species. Recent reviews (Kołodziejczyk et al., 2025; Rius et al., 2015) indicate that reductions in genetic diversity are not a general feature of invasions. Among 101 studies of invasive populations, only 19 reported decreased genomic diversity, whereas 43 reported maintenance or even increases in diversity; the majority of the remaining studies indicated high diversity despite lack of direct comparison with native ranges. These findings suggest that invasive populations often retain sufficient genetic variation to sustain evolutionary potential, with losses from founder effects frequently mitigated over time by secondary introductions, admixture, and dispersal.
Colonization and dispersal pathways
The unexpected Pacific origin of the DLVR population highlights the critical role of long-distance ocean dispersal in shaping the distribution of Chinook salmon across Patagonia and South America. Rare dispersal events, combined with local propagule pressure, can drive successful establishment and expansion, as predicted by invasion biology models (Sakai et al., 2001; Lockwood et al., 2013; Narum et al., 2017; Correa and Gross, 2008; Figueroa-Muñoz et al., 2023). Genetic data from this study indicates that Atlantic populations, including DLVR, are not isolated but are continuously influenced by gene flow from Pacific sources, local admixture, and ongoing connectivity. As such, this study supports earlier work emphasizing the role of oceanic dispersal in connecting distant populations, across major geographic barriers (Becker et al., 2007; Riva-Rossi et al., 2012; Gomez-Uchida et al., 2018). Notably, connectivity is maintained even across the formidable Southern Patagonian Ice Field (Antezana, 1999), being facilitated by a network of freshwater and estuarine stepping-stones that connect Pacific and Atlantic drainages. Southward currents, including the West Wind Drift and Cape Horn Current, likely promote the movement of anadromous salmon from southern Chile into Atlantic drainages, whereas the northward Humboldt Current may have limited early ranching success in northern Chile (Glorioso and Flather, 1995, 1997; Sabatini et al., 2004; Ciancio et al., 2005; Montecino and Lange, 2009; Waters, 2008; Zheng et al., 2023). Gene flow from Pacific drainages into Atlantic populations, observed here and by Gomez-Uchida et al. (2018), underscores the ocean as a vector of connectivity across watersheds.
Similar ocean-mediated connectivity has been observed in other migratory marine and estuarine species, including European green crab (Darling et al., 2008), Chilean rockfishes (Eschmeyer and Hureau, 1971; Núñez et al., 2010), and anadromous lampreys (Neira et al., 1988; Potter et al., 1979; Riva-Rossi et al., 2020). These examples demonstrate that ocean currents and stepping-stone habitats can facilitate range expansion and maintain connectivity even across substantial biogeographic barriers. This connectivity is not unique to Chinook salmon. Similar patterns have been observed in other migratory marine and estuarine species of South America: while rockfishes along the Chilean coast show northward and southward dispersal shaped by the Humboldt Current and the West Wind Drift (Eschmeyer and Hureau, 1971; Núñez et al., 2010). Anadromous lampreys of the genus Geotria and the Chilean lamprey (Mordacia lapicida) also exhibit dispersal patterns that closely follow regional oceanographic features (Neira et al., 1988; Potter et al., 1979; Riva-Rossi et al., 2020). These examples reinforce the notion that ocean currents, in combination with stepping-stone habitats, can facilitate rapid range expansion and sustained connectivity even in species facing substantial biogeographic barriers.
In addition to oceanic pathways, historical freshwater connections may have also contributed to the colonization of Atlantic Ocean basins. Over the past century, retreat of the Dickson Glacier exposed a lake that may have intermittently drained toward both the Pacific and Atlantic Oceans (Martinic, 2010). This raises the possibility of a link between Dickson Lake (Pacific drainage) and Frías Lagoon (Atlantic drainage), potentially providing a freshwater corridor for Chinook salmon from the Serrano River to reach the Santa Cruz River basin (Figure 1B). Although this connection is no longer active (J. Ciancio, pers. comm.) and remains speculative, it complements the broader picture of multi-pathway dispersal and highlights the need for further investigation into historical and environmental factors shaping invasion dynamics.
Evolutionary consequences of dispersal
The Santa Cruz River basin may act as a central hub in the Chinook salmon invasion network along South America’s Atlantic coast, where connected tributaries and suitable spawning habitats support both local establishment and broader dispersal. Within this system, the De las Vueltas River shows the strongest Pacific ancestry signal, consistent with stepping-stone and leading-edge dynamics in which individuals from established populations colonize peripheral habitats, fostering range expansion and new invasion foci (Hulme et al., 2008; Wilson et al., 2009; Florance et al., 2011). DLVR also receives secondary inputs from nearby tributaries such as Guanaco, La Leona, and upper Santa Cruz (Avigliano et al., 2025). Importantly, these dynamics illustrate how invasive species exploit both natural dispersal pathways and human-facilitated corridors, with human actions reshaping the scale, speed, and ecological consequences of expansion (Blackburn et al., 2011; Wilson et al., 2016). In this context, redistribution hubs like DLVR and Santa Cruz River amplify propagule pressure at the invasion front, accelerating colonization into new basins.
At the invasion front, dispersal ability is often under selection: highly mobile individuals accumulate at low densities, competition is reduced, and spread is accelerated—a pattern also observed in invasive cane toads (Rhinella marina), gypsy moths (Lymantria dispar), and green crabs (Carcinus maenas) (Sharov and Liebhold, 1998; Berthouly-Salazar et al., 2013; Lehnert et al., 2018; Daly et al., 2023; Poloni et al., 2024). In the case of Chinook salmon, such dynamics may favor traits enhancing long-distance migration and marine survival, including rapid growth that shortens ocean residence time and increases survival. Yet dispersal entails trade-offs and risks: individuals investing in migration may contribute less locally, leaving edge populations to function primarily as propagule sources while facing demographic constraints such as reduced genetic diversity, drift, and Allee effects (Nathan and Muller-Landau, 2000; Courchamp et al., 2008; Kanarek and Webb, 2010). For instance, the isolated Caterina population may be limited by small effective size, whereas DLVR sustains continuous immigration from multiple sources, maintaining diversity, mitigating density dependence, and acting as both a local reservoir and a hub for further Atlantic dispersal (Kanarek and Webb, 2010; Letnic et al., 2011).
These dynamics likely drive the colonization of downstream Atlantic basins, including the Paraná River and southern Brazilian lagoons (Espínola et al., 2024; Figueroa-Muñoz et al., 2023; Gonçalves Gowert et al., 2025). The Santa Cruz River thus acts as a redistribution node, amplifying propagule pressure and enabling multi-step dispersal, showing how connectivity, genetic diversity, and adaptive potential transform peripheral populations like DLVR into demographic and evolutionary engines for continental-scale spread (Colautti et al., 2006; Wilson et al., 2009).
Management implications
These dispersal dynamics and hub-mediated expansions highlight how peripheral populations like DLVR can drive multi-step colonization across Patagonian watersheds. By concentrating genetically diverse propagules and facilitating spread into connected and marginal habitats, such nodes underscore the basins that are most at risk of invasion. Despite the end of commercial aquaculture in 2013, Chinook salmon, by virtue of its high dispersal capacity and straying rates, continues to colonize areas far beyond initial introduction sites, posing a continuing risk to basins lacking aquaculture or established feral populations (Pascual et al., 2009; Figueroa-Muñoz et al., 2023).These findings emphasize the importance of proactive monitoring and management strategies that target key dispersal routes rather than solely targeting established populations, as evidence from other invasive species indicates that disrupting movement corridors can be more effective than direct population suppression (Fausch et al., 2009; Britton et al., 2023).
Two large hydroelectric dams currently under construction in the Santa Cruz basin are expected to fragment riverine habitats and obstruct upstream migration of diadromous species—including native lampreys (Geotria macrostoma), exotic anadromous rainbow trout or steelhead (Oncorhynchus mykiss), and Chinook salmon. By restricting access to critical spawning and rearing habitats in the upper basin, the dams threaten the persistence of migratory populations and may halt the expansion of Chinook salmon into headwater systems such as the Río de las Vueltas and other connected basins (Baigún et al., 2022; Quiroga et al., 2015, 2023; Riva-Rossi et al., 2020). Paradoxically, this anthropogenic barrier could act as an inadvertent containment mechanism, illustrating the complex and often contradictory role of large-scale infrastructure in fluvial ecosystems: while imperiling native biodiversity, it may simultaneously help limit the spread of invasive species.
The Santa Cruz case underscores the need for an integrated, forward-looking approach to river governance that blends ecological monitoring, stakeholder engagement, and adaptive planning to reconcile conservation goals with the realities of increasingly modified freshwater systems. Although invasive anadromous salmonids have brought economic benefits through recreational and commercial fishing, the long-term consequences for native biodiversity and ecosystem stability remain poorly understood. Growing tensions among stakeholders—including conservationists, local fishers, and other affected groups—further emphasize the need for inclusive governance frameworks capable of navigating the trade-offs inherent in managing invasive species and infrastructure development (Pascual et al., 2009; García de Leaniz et al., 2010; Narum et al., 2017; Ciancio et al., 2019).
Conclusion
This study demonstrates that the De las Vueltas River (DLVR) Chinook salmon population primarily originates from genetically diverse Pacific stocks, with secondary contributions from established populations in Aysén and, to a lesser extent, Santa Cruz. Genetic evidence and historical records indicate that both long-distance oceanic dispersal and intra-basin connectivity have shaped colonization dynamics. These results underscore the role of invasion hubs and leading-edge populations in facilitating multi-step expansion, where highly mobile individuals establish new populations beyond the core range. From a management perspective, monitoring and disrupting key dispersal routes, particularly at such invasion hubs, is critical to limit further spread into Patagonian basins.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Ethics statement
The animal study was approved by CONICET (Consejo Nacional de Investigaciones Científicas y Tecnológicas). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
CR: Writing – review & editing, Writing – original draft, Investigation, Data curation, Formal analysis, Conceptualization. JC: Investigation, Conceptualization, Funding acquisition, Writing – review & editing. DG-U: Resources, Conceptualization, Writing – review & editing. MP: Writing – review & editing. RC: Writing – review & editing, Investigation. PQ: Writing – review & editing, Visualization. CD: Writing – review & editing. CL: Writing – review & editing. JG: Writing – review & editing, Resources.
Funding
The authors declare financial support was received for the research and/or publication of this article. Funding for this research was provided by the Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) of Argentina to Carla Riva-Rossi, Javier Ciancio, Miguel Pascual, Pamela Quiroga, and Cecilia Di Prinzio and the Norwegian Research Council (grant no. 287438) to Javier Ciancio. Daniel Gómez-Uchida received funding from Chile's government grants FONDECYT Regular 1191256 and Núcleo Milenio INVASAL NCN2021-056.
Acknowledgments
We thank R. Hudson for help with field collections. El Chaltén Municipality, Secretaría de Estado de Pesca y Acuicultura de Santa Cruz, Argentina and Fundación Anfibia provided logistic support. Samples were collected under permit 040/2014 from Subsecretaría de Pesca y Actividades Portuarias de Santa Cruz, Argentina).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1662755/full#supplementary-material
References
Acha E. M., Mianzan H. W., Guerrero R. A., Favero M., and Bava J. (2004). Marine fronts at the continental shelves of austral South America: Physical and ecological processes. J. Mar. Syst. 44, 83–105. doi: 10.1016/j.jmarsys.2003.09.005
Antezana T. (1999). Plankton of southern Chilean fjords: trends and linkages. Sci. Mar. 63, 69–80. doi: 10.3989/scimar.1999.63s169
Arismendi I., Soto D., Penaluna B., Jara C., Leal C., and León-Muñoz J. (2014). Aquaculture, non-native salmonid invasions, and associated declines of native fishes in northern Patagonian lakes. Freshw. Biol. 59, 1–15.
Avigliano E., Niklitschek E., Chung M.-T., Diaz B., Chalde T., Di Prinzio C., et al. (2025). Isotope geochemistry as a natural tag of fish in Patagonian freshwater environments: The invasive Chinook salmon case. Sci. Total Environ. 873, 162395. doi: 10.1016/j.scitotenv.2023.162395
Baigún C., Casalinuovo M., Quiroga P., Riva-Rossi C., Colautti D., Solimano P., et al. (2022). “Fish and Fisheries of the Patagonian Steppe,” in Freshwaters and Wetlands of Patagonia. Eds. Mataloni G. and Quintana R. D. (Springer, Heidelberg, Germany), 351–407.
Basulto S. (2003). El largo viaje de los salmones. Una crónica olvidada. Propagación y cultivo de especies acuáticas en Chile (Santiago: Maval Ltd).
Becker L. A., Pascual M. A., and Basso N. G. (2007). Colonization of the Southern Patagonia ocean by exotic chinook salmon. Conserv. Biol. 21, 1347–1352. doi: 10.1111/j.1523-1739.2007.00761.x
Beckman D. W. (2011). “Introduction to fish reproduction,” in The Physiology of Fishes, 4th edition. Eds. Evans D. H. and Claiborne J. B. (CRC Press, Boca Raton), 295–329.
Berthouly-Salazar C., Hui C., Blackburn T. M., Gaboriaud C., van Rensburg B. J., van Vuuren B. J., et al. (2013). Long-distance dispersal maximizes evolutionary potential during rapid geographic range expansion. Mol. Ecol. 22, 5793–5804. doi: 10.1111/mec.12538
Blackburn T. M., Pyšek P., Bacher S., Carlton J. T., Duncan R. P., Jarošík V., et al. (2011). A proposed unified framework for biological invasions. Trends Ecol. Evol. 26, 333–339. doi: 10.1016/j.tree.2011.03.023
Britton J. R., Lynch A. J., Bardal H., Bradbeer S. J., Coetzee J. A., Coughlan N. E., et al. (2023). Preventing and controlling nonnative species invasions to bend the curve of global freshwater biodiversity loss. Environ. Rev. 31, 310–326. doi: 10.1139/er-2022-0103
Busby P. J., Wainwright T. C., Bryant G. J., Lierheimer L. J., Waples R. S., Waknitz F. W., et al. (1997). Status review of Chinook salmon from Washington, Idaho, Oregon, and California. NOAA Tech. Memo.
Ciancio J. E., Aigo J., Clarke R., and Flikke R. (2019). El polémico salmón invasor. Desde la Patagonia difundiendo saberes 16, 10–19.
Ciancio J. E., Pascual M. A., Lancelotti J., Riva-Rossi C. M., and Botto F. (2005). Chinook Salmon (Oncorhynchus tshawytscha) in the Santa Cruz River, an Atlantic Basin of Patagonia. Environ. Biol. Fishes 74, 217–225. doi: 10.1007/s10641-005-0208-1
Ciancio J. E., Riva-Rossi C., Pascual M. A., Anderson E., and Garza J. C. (2015). The invasion of an Atlantic Ocean river basin in Patagonia by Chinook salmon: new insights from SNPs. Biol. Invasions 17, 2989–2998. doi: 10.1007/s10530-015-0928-x
Clemento A. J., Crandall E. D., Garza J. C., and Anderson E. C. (2014). Evaluation of a single nucleotide polymorphism baseline for genetic stock identification of Chinook salmon (Oncorhynchus tshawytscha) in the California Current large marine ecosystem. Fish. Bull. 112, 112–130. doi: 10.7755/FB.112.2-3.2
Clobert J., Baguette M., Benton T. G., and Bullock J. M. (2012). Dispersal Ecology and Evolution (New York: Oxford University Press).
Colautti R. I., Ricciardi A., Grigorovich I. A., and MacIsaac H. J. (2006). Is invasion success explained by the enemy release hypothesis? Ecol. Lett. 7, 721–733. doi: 10.1111/j.1461-0248.2004.00616.x
Consuegra S., Phillips N., Gajardo G., and García de Leaniz C. (2011). Winning the invasion roulette: escapes from fish farms increase admixture and facilitate establishment of non-native rainbow trout. Evol. Appl. 4, 660–671. doi: 10.1111/j.1752-4571.2011.00189.x
Correa C. and Gross M. R. (2008). Chinook salmon invades southern South America. Biol. Invasions 10, 615–639. doi: 10.1007/s10530-007-9157-2
Correa C. and Moran P. (2017). Polyphyletic ancestry of expanding Patagonian Chinook salmon populations. Sci. Rep. 7, 14338. doi: 10.1038/s41598-017-14531-6
Courchamp F., Berec L., and Gascoigne J. (2008). Allee effects in ecology and conservation (New York: Oxford University Press).
Daly E. Z., Chabrerie O., Massol F., Facon B., Hess M. C. M., Tasiemski A., et al. (2023). A synthesis of biological invasion hypotheses associated with the introduction–naturalisation–invasion continuum. Oikos 2023, e09645. doi: 10.1111/oik.09645
Darling J. A., Bagley M. J., Roman J., Tepolt C. K., and Geller J. B. (2008). Genetic patterns across multiple introductions of the globally invasive crab genus Carcinus. Mol. Ecol. 17, 4992–5007. doi: 10.1111/j.1365-294X.2008.03978.x
Di Prinzio C. Y., Arismendi I. J., and Olivos A. (2024). Revealing a rapid shift in the phenology of the adult spawning migration of an introduced Chinook salmon population in Patagonia. Aquat Ecol. 58, 299–312. doi: 10.1007/s10452-023-10066-2
Di Prinzio C. Y., Riva-Rossi C. M., Ciancio J. E., Garza J. C., and Casaux R. (2015). Disentangling the contributions of ocean ranching and net-pen aquaculture in the successful establishment of Chinook salmon in a Patagonian basin. Environ. Biol. Fishes 98, 1987–1997. doi: 10.1007/s10641-015-0418-0
Dlugosch K. M. and Parker I. M. (2007). Founding events in species invasions: genetic variation, adaptive evolution, and the role of multiple introductions. Mol. Ecol. 17, 431–449. doi: 10.1111/j.1365-294X.2007.03538.x
Donaldson L. R. and Joyner T. (1983). The salmonid fishes as a natural livestock. Sci. Amer. 249, 50–58. doi: 10.1038/scientificamerican0783-50
Early R., Bradley B. A., Dukes J. S., Lawler J. J., Olden J. D., Blumenthal D. M., et al. (2016). Global threats from invasive alien species in the twenty-first century and national response capacities. Nat. Commun. 7, 12485. doi: 10.1038/ncomms12485
Eschmeyer W. N. and Hureau J. C. (1971). Sebastes mouchezi, a senior synonym of Helicolenus tristanensis, with comments on Sebastes capensis and zoogeographical considerations. Copeia 1971, 576–579. doi: 10.2307/1442466
Espínola L. A., Baigún C., Riva-Rossi C., Quiroga P., Abrial E., Rabuffetti A. P., et al. (2024). Non-native anadromous salmonids in the La Plata Basin: long distance colonization? BioInvasions Rec. 13, 251–265. doi: 10.3391/bir.2024.13.1.23
Fausch K. D., Rieman B. E., Dunham J. B., Young M. K., and Peterson D. (2009). Invasion versus isolation: trade-offs in managing native salmonids with barriers to upstream movement. Conserv. Biol. 23, 859–870. doi: 10.1111/j.1523-1739.2008.01159.x
Fernández D. A., Ciancio J. E., Ceballos S. G., Riva-Rossi C. M., and Pascual M. A. (2010). Chinook salmon (Oncorhynchus tshawytscha, Walbaum 1792) in the Beagle Channel, Tierra del Fuego: the onset of an invasion. Biol. Invasions 12, 2991–2997. doi: 10.1007/s10530-010-9731-x
Figueroa-Muñoz G., Arismendi I., Urzúa Á., Soto-Mendoza P., and González J. (2022). Consumption of marine-derived nutrients from invasive Chinook salmon (Oncorhynchus tshawytscha) transfer ω-3 highly unsaturated fatty acids to invasive resident rainbow trout (O. mykiss). Sci. Total Environ. 844, 157077. doi: 10.1016/j.scitotenv.2022.157077
Figueroa-Muñoz G., Gomez-Uchida D., Fierro P., Arriagada P., Górski K., and Habit E. (2021). First record of a synergistic interaction between invasive salmonids in South America. Biol. Invasions 24, 1–8. doi: 10.1007/s10530-021-02629-x
Figueroa-Muñoz G., Olivos J. A., Arismendi I., Fabiano G., Laporta M., Silveira S., et al. (2023). Contemporary distribution of non-native Chinook salmon (Oncorhynchus tshawytscha) in South America. Biol. Invasions 25, 2727–2735. doi: 10.1007/s10530-023-03083-7
Florance D., Webb J. K., Dempster T., Kearney M. R., Worthing A., and Letnic M. (2011). Excluding access to invasion hubs can contain the spread of an invasive vertebrate. Proc. Biol. Sci. 278, 2900–2908. doi: 10.1098/rspb.2011.0032
García de Leaniz C., Gajardo G., and Consuegra S. (2010). From best to pest: changing perspectives on the impact of exotic salmonids in the southern hemisphere. Syst. Biodivers. 8, 447–459. doi: 10.1080/14772000.2010.537706
Giese A. C. (2024). Estructuración geográfica en poblaciones de trucha marrón (Salmo trutta L.) migratoria en ríos de la Patagonia Austral (Bariloche, Argentina: Universidad Nacional del Comahue).
Glorioso P. D. and Flather R. A. (1995). A barotropic model of the currents off SE South America. J. Geophys. Res. 100, 427–440. doi: 10.1029/95JC00942
Glorioso P. D. and Flather R. A. (1997). The patagonian shelf tides. Prog. Oceanogr. 40, 263–283. doi: 10.1016/S0079-6611(98)00004-4
Gomez-Uchida D., Cañas-Rojas D., Riva-Rossi C. M., Ciancio J. E., Pascual M. A., Ernst B., et al. (2018). Genetic signals of artificial and natural dispersal linked to colonization of South America by non-native Chinook salmon (Oncorhynchus tshawytscha). Ecol. Evol. 8, 6192–6209. doi: 10.1002/ece3.4036
Gonçalves Gowert Y. G., Lemos V. M., Corrêa F., Vollrath S., Vieira J. P., Condini M. V., et al. (2025). Multiple tools to investigate the origin of the exotic species Chinook salmon Oncorhynchus tshawytscha (Walbau) (Salmonidae) in the world's largest choked coastal lagoon. J. Fish Biol. 104, 1892–1906. doi: 10.1111/jfb.15728
Gozlan R. E. (2008). Introduction of non-native freshwater fish: Is it all bad? Fish. 9, 106–115. doi: 10.1111/j.1467-2979.2007.00263.x. Pyšek, P., Richardson, D.M.
Gozlan R. E., Andreou D., Asaeda T., Beyer K., Bouhadad R., Burnard D., et al. (2010). Pan-continental invasion of Pseudorasbora parva: towards a better understanding of freshwater fish invasions. Fish. 11, 315–340.
Healey M. C. (1991). “Life history of Chinook salmon,” in Pacific salmon life histories. Eds. Groot C. and Margolis L. (UBC Press, Vancouver), 311–394.
Hulme P. E., Bacher S., Kenis M., Klotz S., Kühn I., Minchin D., et al. (2008). Grasping at the Routes of Biological Invasions: A Framework for Integrating Pathways into Policy. J. Appl. Ecol. 45, 403–414. Available online at: http://www.jstor.org/stable/20143990.
Hulme P. E. (2009). Trade, transport and trouble: managing invasive species pathways in the globalized world. J. Appl. Ecol. 46, 10–18. doi: 10.1111/j.1365-2664.2008.01600.x
Jombart T., Devillard S., and Balloux F. (2010). Discriminant analysis of principal components: a new method for the analysis of genetically structured populations. BMC Genet. 11, 94. doi: 10.1186/1471-2156-11-94
Källo K., Birnie-Gauvin K., Baktoft H., Larsen M. H., Søndergaard M. S., Nielsen A., et al. (2023). Otolith microchemistry combined with genetics reveal patterns of straying and population connectivity in anadromous brown trout (Salmo trutta). Ecol. Freshw. Fish 33, e12760. doi: 10.1111/eff.12760
Kanarek A. R. and Webb C. T. (2010). Allee effects, adaptive evolution, and invasion success. Evol. Appl. 3, 122–135. doi: 10.1111/j.1752-4571.2009.00112.x
Keefer M. L. and Caudill C. C. (2014). Homing and straying by anadromous salmonids: a review of mechanisms and rates. Rev. Fish Biol. Fish. 24, 333–368. doi: 10.1007/s11160-013-9334-6
Kołodziejczyk J., Fijarczyk A., Porth I., Robakowski P., Vella N., Vella A., et al. (2025). Genomic investigations of successful invasions: the picture emerging from recent studies. Biol. Rev. 100, 1396–1418. doi: 10.1111/brv.70005
Lehnert S. J., DiBacco C., Jeffery N. W., Blakeslee A. M. H., Isaksson J., Roman J., et al. (2018). Temporal dynamics of genetic clines of invasive European green crab (Carcinus maenas) in eastern North America. Evol. Appl. 11, 1656–1670. doi: 10.1111/eva.12657
Letnic M., Webb J. K., Jessop T. S., and Dempster T. (2011). Restricting access to invasion hubs enables sustained control of an invasive vertebrate. J. Appl. Ecol. 52, 341–347. doi: 10.1111/1365-2664.12390
Lockwood J. L., Hoopes M. F., and Marchetti M. P. (2013). Invasion Ecology. 2nd edn (Chichester: John Wiley & Sons).
Lovell S. J., Stone S. F., and Fernandez L. (2006). The economic impacts of aquatic invasive species: a review of the literature. Agric. Resour. Econ. Rev. 35, 195–208. doi: 10.1017/S1068280500010157
Martin J., Rougemont Q., Drouineau H., Launey S., Jatteau P., Bareille G., et al. (2015). Dispersal capacities of anadromous Allis shad population inferred from a coupled genetic and otolith approach. Can. J. Fish. Aquat. Sci. 72, 991–1003. doi: 10.1139/cjfas-2014-0510
Martinic M. B. (2010). ¿Un enigma histórico-geográfico resuelto? La intercomunicación de las cuencas hídricas del Paine y Lago Argentino. Magallania 38, 27–40.
McDowall R. M. (2002). The origin of the salmonid fishes: marine, freshwater … or neither? Rev. Fish Biol. Fish. 11, 171–179. doi: 10.1023/A:1020370328194
Montecino V. and Lange C. B. (2009). The Humboldt Current System: Ecosystem components and processes, fisheries, and sediment studies. Prog. Oceanogr. 83, 65–79. doi: 10.1016/j.pocean.2009.07.041
Moran B. M. and Anderson E. C. (2019). Bayesian inference from the conditional genetic stock identification model. Can. J. Fish. Aquat. Sci. 76, 551–560. doi: 10.1139/cjfas-2018-0143
Musleh S. S., Seeb L. W., Seeb J. E., Ernst B., Neira S., Harrod C., et al. (2020). Mixed-stock analyses of migratory, non-native Chinook salmon at sea and assignment to natal sites in fresh water at their introduced range in South America. Biol. Invasions 22, 3175–3182. doi: 10.1007/s10530-020-02319-0
Narum S. R., Gallardo P., Correa C., Matala A., Hasselman D., Sutherland B. J. G., et al. (2017). Genomic patterns of diversity and divergence of two introduced salmonid species in Patagonia, South America. Evol. Appl. 10, 402–416. doi: 10.1111/eva.12464
Nathan R. and Muller-Landau H. C. (2000). Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends Ecol. Evol. 15, 278–285. doi: 10.1016/S0169-5347(00)01874-7
Neira F. J., Bradley J. S., Potter I. C., and Hilliard R. W. (1988). Morphological variation among widely dispersed larval populations of anadromous Southern Hemisphere lampreys (Geotriidae and Mordaciidae). Zool. J. Linn. Soc 92, 383–408. doi: 10.1111/j.1096-3642.1988.tb01730.x
Núñez J. J., González M. T., and Pérez-Losada M. (2010). Testing patterns of adaptive radiation in a rockfish species complex (Sebastes spp.) from the Southern Hemisphere. Mol. Phylogenet. Evol. 56, 791–801. doi: 10.4067/S0718-19572010000400005
Pascual M., Macchi P., Urbanski J., Marcos F., Riva-Rossi C., Novara M., et al. (2002). Evaluating Potential Effects of Exotic Freshwater Fish From Incomplete Species Presence–absence Data. Biol. Invasions 4, 101–113. doi: 10.1023/A:1020513525528
Pascual M. A. and Ciancio J. E. (2007). “Introduced anadromous salmon and trout in Patagonia: risks, uses, and a conservation paradox,” in Ecological and Genetic Implications of Aquaculture Activities. Ed. Bert T. M. (Springer, Dordrecht), 255–284. doi: 10.1007/978-1-4020-6150-6_12
Pascual M. A., Lancelotti J. L., Ernst B., Ciancio J. E., Aedo E., and García-Asorey M. (2009). Scale, connectivity, and incentives in the introduction and management of non-native species: the case of exotic salmonids in Patagonia. Front. Ecol. Environ. 7, 533–540. doi: 10.1890/070127
Peakall R. and Smouse P. E. (2012). GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research—an update. Bioinformatics 28, 2537–2539. doi: 10.1093/bioinformatics/bts460
Pejchar L. and Mooney H. A. (2009). Invasive species, ecosystem services and human well-being. Trends Ecol. Evol. 24, 497–504. doi: 10.1016/j.tree.2009.03.016
Poloni S., Lutscher F., and Lewis M. A. (2024). Evolutionary dynamics at the leading edge of biological invasions. bioRxiv 2024.12.13.628423. doi: 10.1101/2024.12.13.628423
Potter I. C., Prince P. A., and Croxall J. P. (1979). Data on the adult marine and migratory phases in the life cycle of the southern hemisphere lamprey, Geotria australis Gray. Environ. Biol. Fishes. 4, 65–69.
Pyšek P., Richardson D. M., Pergl J., Jarošík V., Sixtová Z., and Weber E. (2008). Geographical and taxonomic biases in invasion ecology. Trends Ecol. Evol. 23, 237–244. doi: 10.1016/j.tree.2008.02.002
Quinn T. P. (2018). The behavior and ecology of Pacific salmon and trout. 2nd ed. (Seattle: University of Washington Press).
Quinn T. P., Kinnison M. T., and Unwin M. J. (2001). Evolution of Chinook salmon (Oncorhynchus tshawytscha) populations in New Zealand: pattern, rate and process. Genetica 112–113, 493–513. doi: 10.1023/A:1013348024063
Quinn T. P., McGinnity P., and Cross T. F. (2006). Long-term declines in body size and shifts in run timing of Atlantic salmon in Ireland. J. Fish Biol. 88, 81–99. doi: 10.1111/j.0022-1112.2006.01017.x
Quiroga P., Baker C. F., Baigún C., Stewart M., Lancelotti J. L., and Riva-Rossi C. (2023). Distribution and abundance of the Patagonian lamprey (Geotria macrostoma) in the largest unfragmented river of southern Patagonia, Argentina. Environ. Biol. Fishes. 106, 1933 –1951. doi: 10.1007/s10641-023-01472-4
Quiroga A. P., Lancelotti J., Riva-Rossi C., Tagliaferro M., García-Asorey M., and Pascual M. (2015). Dams versus habitat: predicting the effects of dams on habitat supply and juvenile rainbow trout along the Santa Cruz River, Patagonia. Hydrobiologia. doi: 10.1007/s10750-015-2217-1
R Core Team (2023). R: A Language and Environment for Statistical Computing (Vienna: R Foundation for Statistical Computing). Available online at: https://www.R-project.org/.
Rius M., Turon X., Bernardi G., Volckaert F. A. M., and Viard F. (2015). Marine invasion genetics: from spatio-temporal patterns to evolutionary outcomes. Biol. Invasions 17, 869–885. doi: 10.1007/s10530-014-0792-0
Riva-Rossi C., Barrasso D. A., Baker C. F., Quiroga A. P., Baigún C., and Basso N. G. (2020). Revalidation of the Argentinian pouched lamprey Geotria macrostoma (Burmeister 1868) with molecular and morphological evidence. PloS One 15, e0233792. doi: 10.1371/journal.pone.0233792
Riva-Rossi C. M., Pascual M., Aedo-Marchandt E., Mezga B., Basso N., Ciancio J., et al. (2012). The invasion of Patagonia by Chinook salmon (Oncorhynchus tshawytscha): inferences from mitochondrial DNA patterns. Genetica 140, 439–453. doi: 10.1007/s10709-012-9692-3
Sabatini M. E., Reta R., and Matano R. P. (2004). Circulation and zooplankton biomass distribution over the southern Patagonian shelf during late summer. Cont. Shelf Res. 24, 1359–1373. doi: 10.1016/j.csr.2004.04.003
Sakai A. K., Allendorf F. W., Holt J. S., Lodge D. M., Molofsky J., With K. A., et al. (2001). The population biology of invasive species. Annu. Rev. Ecol. Evol. Syst. 32, 305–332. doi: 10.1146/annurev.ecolsys.32.081501.114037
Seebens H., Blackburn T. M., Dyer E. E., Genovesi P., Hulme P. E., Jeschke J. M., et al. (2017). No saturation in the accumulation of alien species worldwide. Nat. Commun. 8, 14435. doi: 10.1038/ncomms14435
Sharov A. A. and Liebhold A. M. (1998). Bioeconomics of Managing the Spread of Exotic Pest Species with Barrier Zones. Ec. App. 8, 833–845. doi: 10.2307/2641270
Shea K. and Chesson P. (2002). Community ecology theory as a framework for biological invasions. Trends Ecol. Evol. 17, 170–176. doi: 10.1016/S0169-5347(02)02495-3
Simberloff D., Martin J. L., Genovesi P., Maris V., Wardle D. A., Aronson J., et al. (2013). Impacts of biological invasions: what's what and the way forward. Trends Ecol. Evol. 28, 58–66. doi: 10.1016/j.tree.2012.07.013
Soto D., Arismendi I., Di Prinzio C. D., and Jara F. (2007). Establishment of Chinook salmon (Oncorhynchus tshawytscha) in Pacific basins of southern South America and its potential ecosystem implications. Rev. Chil. Hist. Nat. 80, 81–98. doi: 10.4067/S0716-078X2007000100007
Vargas P. V., Arismendi I., and Lara G. (2010). Evidencia de solapamiento de micro-hábitat entre juveniles del salmón introducido Oncorhynchus tshawytscha y el pez nativo Trichomycterus areolatus en el río Allipén, Chile. Rev. Biol. Mar. Oceanogr. 45, 285–292. doi: 10.4067/S0718-19572010000200010
Vitule J. R. S., Freire C. A., and Simberloff D. (2009). Introduction of non-native freshwater fish can certainly be bad. Fish. 10, 98–108. doi: 10.1111/j.1467-2979.2008.00312.x
Warheit K. I., Duda J. J., and Pess G. R. (2012). Population genetic structure of Chinook salmon from western Washington. Trans. Am. Fish. Soc 141, 442–453. doi: 10.1080/00028487.2012.668573
Waters J. M. (2008). Driven by the West Wind Drift? A synthesis of southern temperate marine biogeography, with new directions for dispersalism. J. Biogeogr. 35, 417–427. doi: 10.1111/j.1365-2699.2007.01724.x
Wilson J. R. U., Dormontt E. E., Prentis P. J., Lowe A. J., and Richardson D. M. (2009). Something in the way you move: dispersal pathways affect invasion success. Trends Ecol. Evol. 24, 136–144. doi: 10.1016/j.tree.2008.10.007
Wilson J. R. U., García-Díaz P., Cassey P., Richardson D. M., Pyšek P., and Blackburn T. M. (2016). Biological invasions and natural colonisations are different—the need for invasion science. NeoBiota 31, 87–98. doi: 10.3897/neobiota.31.9185
Keywords: invasive species, anadromous fish, ocean migration, leading-edge dispersal, river connectivity
Citation: Riva-Rossi C, Ciancio J, Gomez-Uchida D, Pascual M, Clarke R, Quiroga P, Di Prinzio C, Lázari C and Garza JC (2025) Between oceans: stepping-stone dispersal and the Pacific-to-Atlantic expansion of Chinook salmon across Patagonia. Front. Mar. Sci. 12:1662755. doi: 10.3389/fmars.2025.1662755
Received: 09 July 2025; Accepted: 15 September 2025;
Published: 08 October 2025.
Edited by:
Pedro Morais, California Department of Water Resources, United StatesReviewed by:
David José Nachón, Oceanographic Center of Vigo, Spanish Institute of Oceanography (IEO), SpainYan Gowert, University federal of Rio Grande (FURG), Brazil
Copyright © 2025 Riva-Rossi, Ciancio, Gomez-Uchida, Pascual, Clarke, Quiroga, Di Prinzio, Lázari and Garza. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Javier Ciancio, Y2lhbmNpb0BjZW5wYXQtY29uaWNldC5nb2IuYXI=