 Wayne M. Koster
Wayne M. Koster Håkan Westerberg
Håkan Westerberg David Dawson1
David Dawson1 Roland Kahsnitz
Roland Kahsnitz Kim Aarestrup
Kim Aarestrup Ben Fanson
Ben Fanson- 1Department of Energy, Environment, and Climate Action, Arthur Rylah Institute for Environmental Research, Heidelberg, VIC, Australia
- 2Department of Aquatic Resources, Institute of Freshwater Research, Swedish University of Agricultural Sciences, Drottningholm, Sweden
- 3Gippsland Eels P/L, Raymond Island, VIC, Australia
- 4National Institute of Aquatic Resources, Technical University of Denmark, Silkeborg, Denmark
This study presents the first investigation into the oceanic spawning migrations of Australian long-finned eels (Anguilla reinhardtii), a critical and previously undocumented stage in their life cycle. In autumn 2024, twenty adult eels were collected from two estuaries in southern Australia and tagged with pop-up satellite archival tags. Individuals were successfully tracked up to five months, with some migrating as far as ~2,500 km from their release sites to the tropical Coral Sea. Migration trajectories revealed relatively consistent and direct movement pathways, with eels remaining several hundred kilometers offshore for much of their journey before venturing farther offshore during later stages. Tag data provided clear evidence of predation events, indicating that escapement to the sea does not guarantee successful spawning and highlighting the potential ecological role of eels within marine food webs. Approximately 40% of migrations were prematurely terminated due to predation, with sharks likely responsible in nearshore environments and marine mammals in offshore regions. Additionally, consistent observations of diel vertical migration, characterized by nocturnal ascents toward the surface and diurnal descents to deeper waters, offer valuable insights into behavioral adaptations during the oceanic migration phase of this enigmatic life history. Tag endpoints and reconstructed migratory trajectories generally aligned with a region between the East Australian Current and the Lord Howe Seamount Chain. Several endpoints clustered within a small region of the northeastern Coral Sea, south of the Solomon Islands. These locations align with historical leptocephali collection sites, strengthening the hypothesis that the spawning area of long-finned eels lies in the northeastern Coral Sea.
Introduction
Animal migrations for breeding are among the most ecologically significant and enigmatic natural phenomena. These migrations can involve transitions between vastly different environments, such as from temperate to tropical regions, span immense distances and durations, and expose organisms to considerable natural risks and anthropogenic threats (Chapman et al., 2014; Dingle, 2014). Detailed knowledge of this crucial life-history stage is essential for understanding population dynamics, effects of changing environmental conditions, and informing conservation efforts (Lennox et al., 2016).
Among the most remarkable examples are the oceanic spawning migrations of anguillid eels (Tsukamoto, 2009). Following an extended growth phase in freshwater and estuarine habitats, adults migrate to offshore marine spawning grounds in tropical waters. These migrations can span thousands of kilometers across open ocean, culminating in a once-in-a-lifetime reproductive event (i.e., semelparity). Recent advances in biotelemetry, particularly the use of pop-up satellite archival tags, have significantly advanced our understanding of these migrations in species like the European eel (A. anguilla) (Aarestrup et al., 2009), American eel (A. rostrata) (Béguer-Pon et al., 2015), Japanese eel (A. japonica) (Manabe et al., 2011), New Zealand long-finned eel (A. dieffenbachii) (Franklin et al., 2023) and tropical Pacific eels (Anguilla spp.) (Schabetsberger et al., 2021). Nevertheless, for many other anguillid species, the oceanic spawning phase remains largely undocumented.
The Australian long-finned eel (A. reinhardtii) is broadly distributed along Australia’s east coast and also occurs in New Zealand, Lord Howe Island, Papua New Guinea, New Caledonia, and Vanuatu. Commonly exceeding one meter in length, this species is both commercially harvested and culturally significant. Kuroki et al. (2020) hypothesized that their spawning area lies in the northeastern Coral Sea, based on the collection of small numbers of long-finned eel leptocephali in this region (i.e., ~14–15.5°S, 160°E) (Kuroki et al., 2008, 2020). However, the oceanic migratory behavior, pathways, and spawning destinations of adults have remained unknown.
Here, we conducted the first direct investigation of the oceanic migrations of long-finned eels using pop-up satellite archival tags. Our findings provide novel insights into migratory routes, behavioral adaptations, threats encountered, and potential spawning areas in the western South Pacific, thereby advancing understanding of this species’ reproductive ecology.
Methods
Fish collection and tagging
In April and May 2024, twenty adult migrant long-finned eels (mean length: 1250 mm, range 1030–1390 mm; mean weight: 5990 g, range 2986–8155 g) were collected near the mouths of Lake King and Tamboon Inlet in southern Victoria, Australia using overnight-set fyke nets (Table 1, Figure 1). For tagging, eels were transferred to a 150 L aerated container filled with estuarine water and individually anaesthetized (0.09 mL AQUI-S per liter water) (AQUI-S, Lower Hutt, New Zealand), with induction times of approximately 8–10 minutes. Pop-up satellite archival tags (PSATs), either Microwave Telemetry X-tags or Wildlife Computers mrPATs, were externally attached using a three-point stainless steel wire configuration inserted dorsally beneath the skin, following the Westerberg method (Økland et al., 2013). The X-tags recorded depth and temperature at 15-minute intervals, with data transmitted via the Argos satellite system after the tag detaches from the eel and floats to the surface. In contrast, the mrPAT tags recorded only daily maximum and minimum temperatures. Throughout the tagging procedure, the head and gill regions of each eel remained submerged in aerated anesthetic solution. The entire tagging process, including biometric measurements, was completed within approximately six minutes per individual.
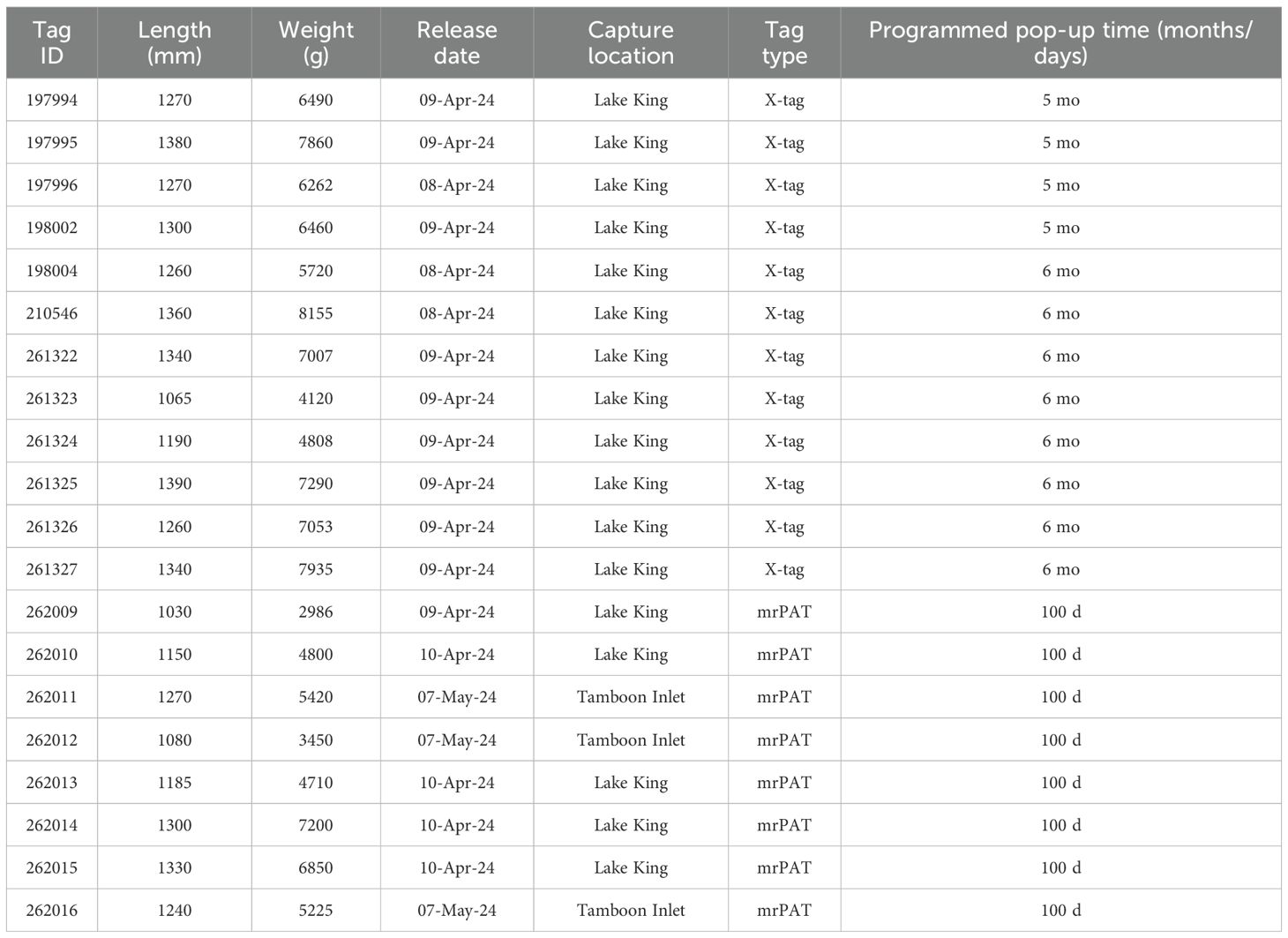
Table 1. Biological data for tagged adult long-finned eels from Gippsland Victoria, Australia.
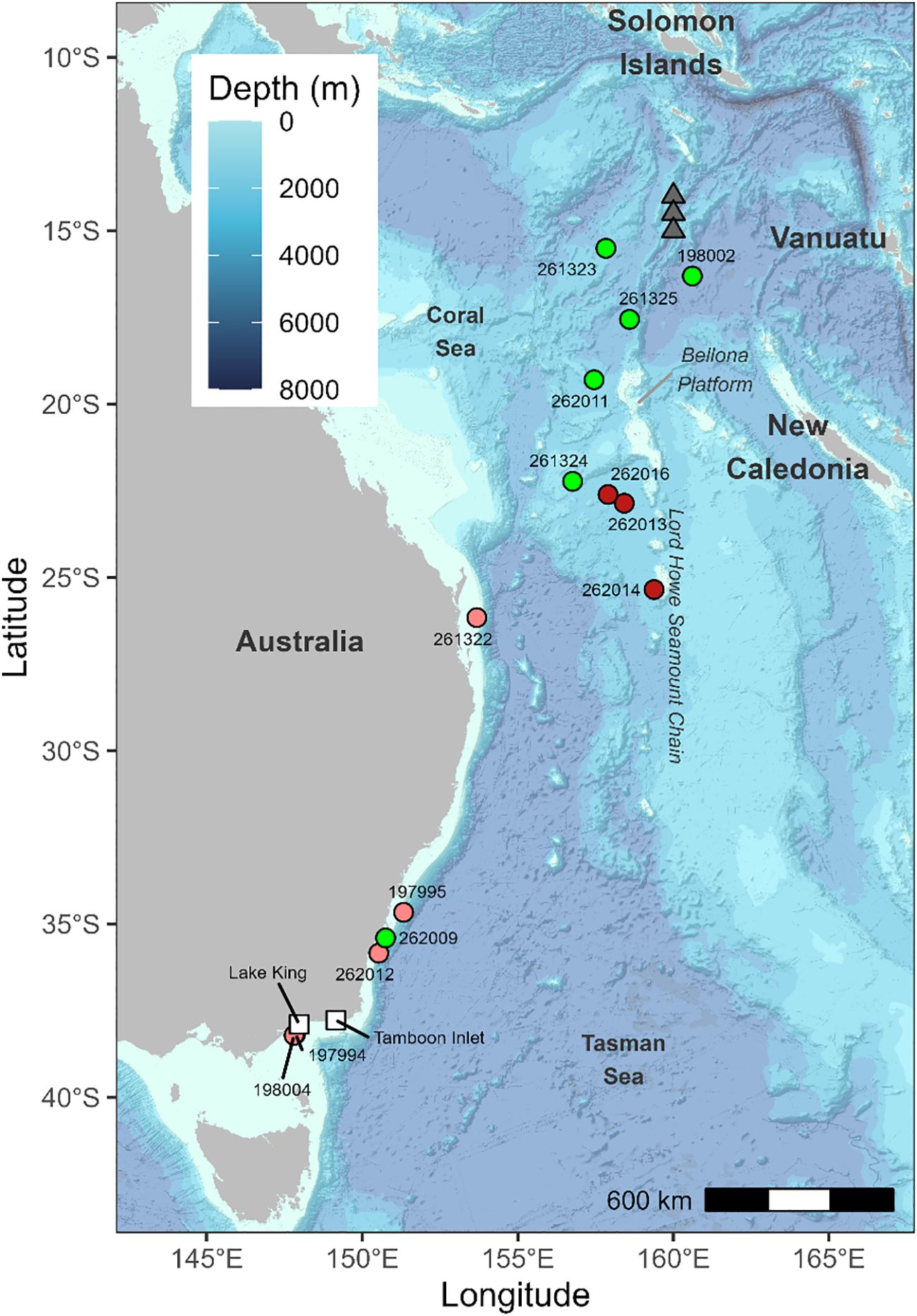
Figure 1. Pop-up locations (coloured circles) of tagged long-finned eels. White squares: release locations. Red circles: mammal predation, pink circles: non-mammal predation. Grey triangles: historical leptocephali collection sites (from Kuroki et al., 2008, 2020).
Following tagging, eels were placed in a 150 L recovery container with estuarine water. Once they regained equilibrium and began to swim freely, they were released either from a nearby beach or offshore via boat. The mrPAT tags were programmed to detach and surface 100 days after release, reflecting the maximum number of days for which data can be recorded. In contrast, X-tags were programmed to detach and surface five to six months post-release. This staggered approach enabled a broad tracking window, which was deemed important due to uncertainty surrounding the duration of migration to the spawning grounds.
Mapping migrations
Diel vertical migration (DVM) is a widespread behavioral pattern among migrating anguillid eels, characterized by nocturnal ascent to shallower depths and descent to deeper waters during daylight hours. This behavior is closely synchronized with local sunrise and sunset times and has been utilized to estimate the position of the eels, particularly daily longitude, by identifying the timing of local noon (Righton et al., 2016; Chang et al., 2020).
In Australian long-finned eels, DVM is marked by rapid vertical transitions, which can introduce distortion in depth profiles due to the data compression algorithms used by X-tags. Actual measured values of temperature and depth are replaced by approximations, called delta limited values, which give underestimates of the increase or decrease between successive datapoints. This distortion results in a systematic westward bias in the estimated longitudes when the sunrise and sunset times are calculated from the time of crossing of a depth or temperature reference level. Different reference levels were tried but all were unable to converge to the known popup longitude. The method adopted was to use the daily mean depth of the eel as a reference level. To correct for the bias, daily longitude estimates were first smoothed using a locally weighted scatterplot smoothing (LOWESS) approach (Cleveland and Devlin, 1988). Typical differences between the daily estimates and the smoothed values were less than 1° longitude. If DVM is a stereotype behavior triggered by light the longitude bias will be constant once the eel has left the shelf and settled with a constant DVM amplitude. With this assumption, the difference between the smoothed longitude estimate at the start and popup longitudes was used as a fixed number correction on the smoothed time-series. This procedure was applied to tag 198002, 261323 and 261325. The correction varied between the individual eels from 2 to 7° longitude. The fourth eel with a long travel distance, tag 261324, had very low data return (Tabel 2) and just 6 randomly distributed days with hourly data was available. In this case the uncorrected estimates based on temperature data was used.
Given temporal variation in sunrise and sunset over the study period, fixed time windows were defined for analysis: 00:00–06:00 UTC as “day” and 12:00–18:00 UTC as “night.” During those periods the vertical activity of the eels is low, and measurements are not distorted by delta limited values. Mean swimming depth and ambient temperature were calculated daily for each period and used to estimate latitude. Oceanographic reference data were obtained from the Copernicus Global Ocean Physics Reanalysis product (1/12° horizontal resolution, 50 vertical levels; Copernicus Marine Service). These data were visualized using Ocean Data View (Schlitzer, 2015; https://odv.awi.de). Modelled ocean temperature profiles were extracted at 10-day intervals following tag deployment, covering the region from 40°S to 10°N and 145°E to 170°E. Latitude estimates were derived by comparing observed night-time mean temperatures and depths with the modelled temperatures at corresponding depths along the corrected longitude. A second latitude estimate was obtained using daytime averages. When the two estimates differed by less than 1°, their mean was taken as the final latitude estimate for that day.
All field work was performed in accordance with the relevant guidelines and regulations under Victorian Flora and Fauna Guarantee Permit 10005451 and Fisheries Victoria Research Permit RP-827. This study was approved and conducted under ethics permit 18-006 (Arthur Rylah Institute for Environmental Research Animal Ethics Committee).
Results
Data were obtained from 14 of the 20 eels tagged (Table 2). Only one mrPAT tag (262011) reached its programmed release date. Of the 14 tags that returned data, the average deployment duration was 60 d (maximum 164 d), and the average data return (i.e., amount of data transmitted) was 66% (range 2–100%).
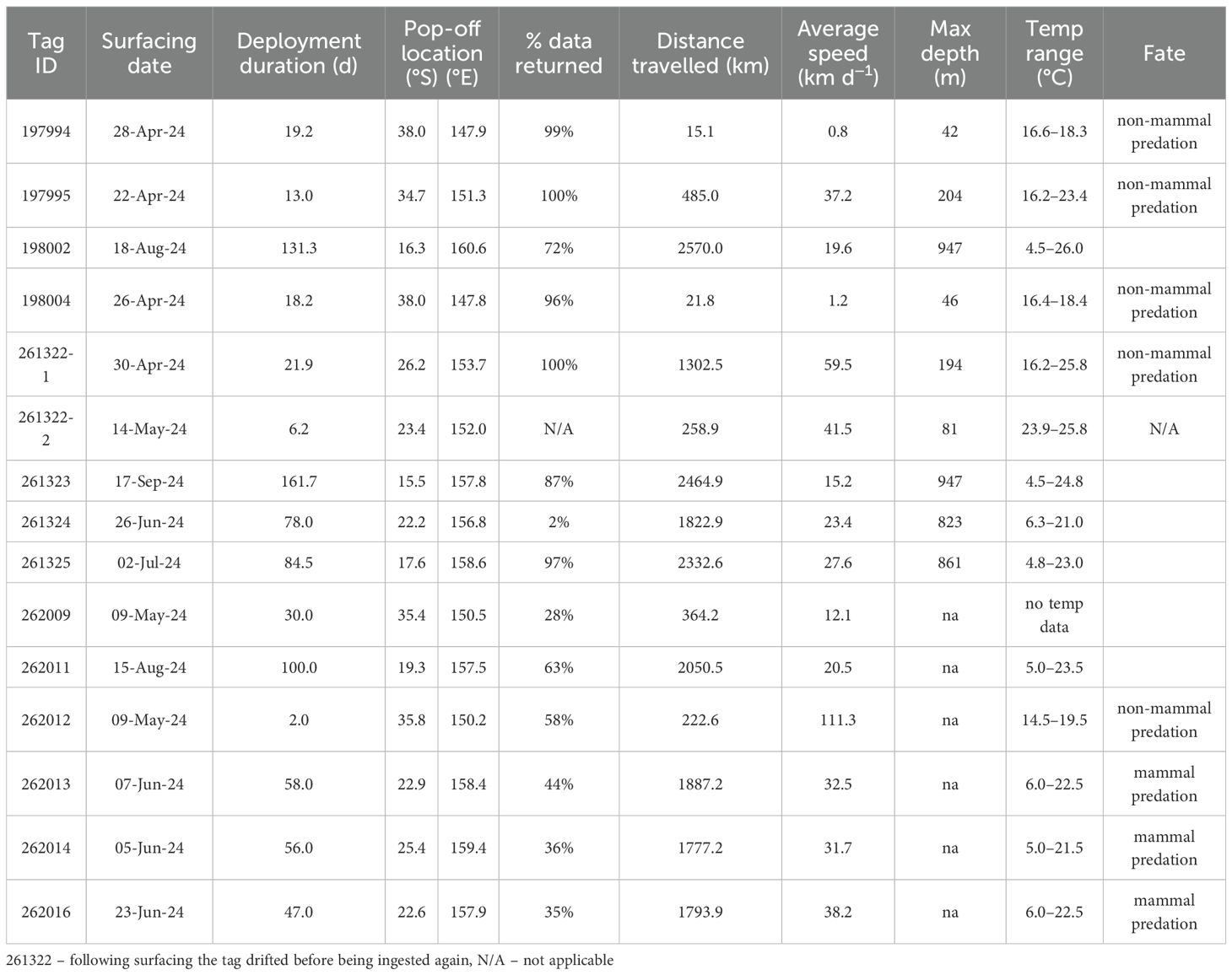
Table 2. Summary of data for the 14 tags attached to long-finned eels that returned data.
Pop-off locations and fate
Eight tags showed signs of predation: three by mammalian predators and five by non-mammalian predators (see below). Two X-tags (197994 and 198004) surfaced within 25 km of their release site, approximately 20–21 days after deployment (Figure 1). Tag 197994 remained at shallow depths (<50 m) and exhibited irregular movement patterns (Figure 2). Tag 198004 also stayed near the surface but displayed a DVM pattern, typically descending to 40–50 m below surface during the day and ascending to 5–10 m below surface at night. A combination of near-constant temperatures (17–18°C) and absence of light during daytime suggests that both eels were ingested by an ectothermic predator, such as a shark (Figure 2).
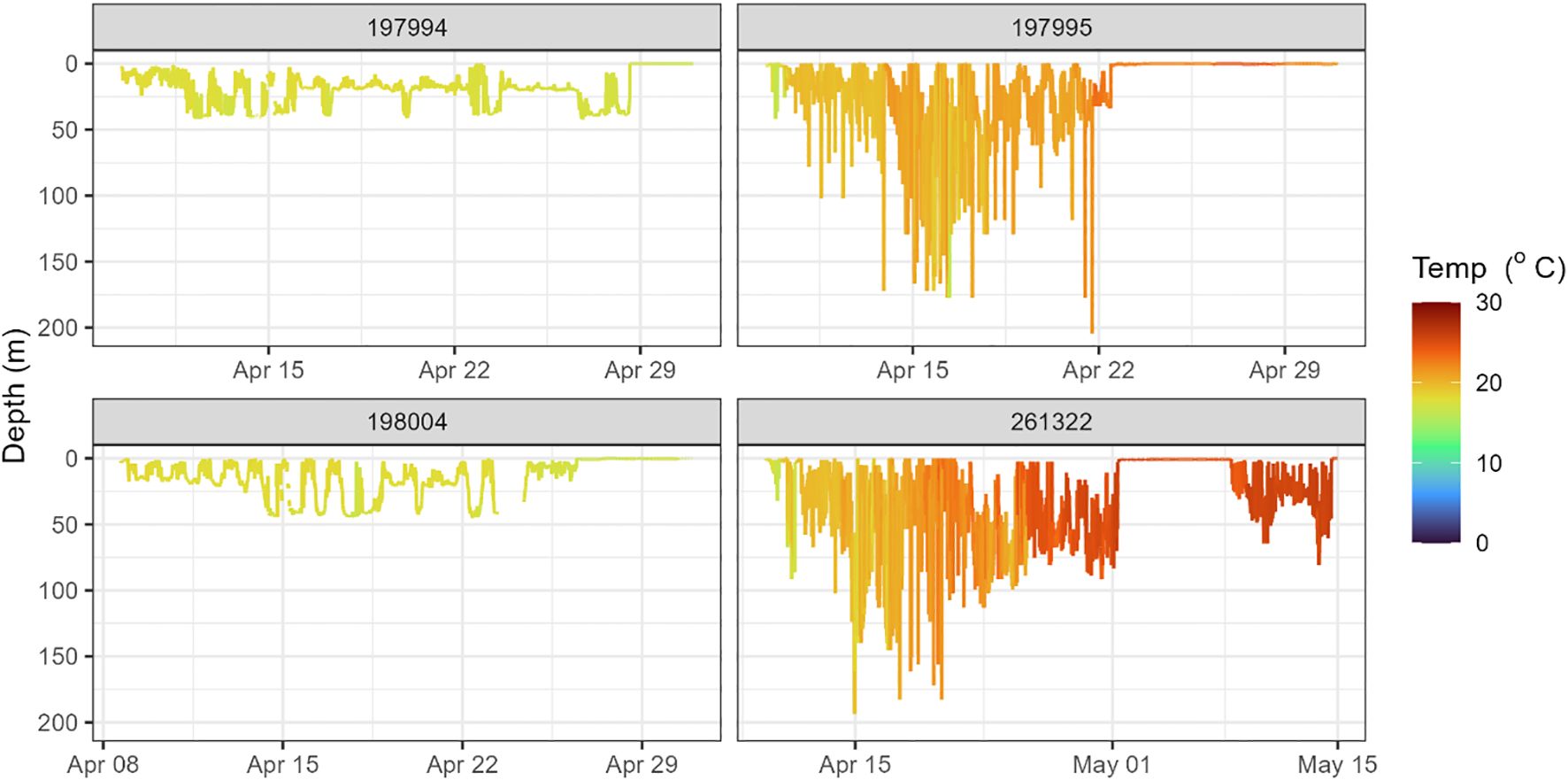
Figure 2. Daily depth and temperature profiles of individual tagged eels, indicating predation events shortly after deployment.
Three tags (197995, 262009, and 262012) surfaced off the southern coast of New South Wales, 230–550 km from their release sites, between 2 and 30 days after deployment (Figure 1). The mrPAT 262012 recorded a displacement of 230 km in just two days, an implausible migration rate for an anguillid eel (e.g., Aarestrup et al., 2009; Righton et al., 2016), strongly suggesting predation and subsequent northward transport before surfacing.
The X-tag 197995 remained near the surface and exhibited a shallow diel vertical movement pattern, descending to 100–150 m during the day and ascending to 10–30 m at night (Figure 2). Over time, recorded temperatures increased from 17 to 23 °C, consistent with a northward movement into progressively warmer waters (Figure 2). An absence of deep dives and light data suggests this eel was ingested by a predator shortly after deployment; however, the tag returned insufficient data to identify the predator (Figure 2). The mrPAT 262009 also returned insufficient data to determine the fate of the eel.
The X-tag 261322 surfaced on the continental shelf near Brisbane, approximately 1,357 km from its release site, just 23 days after deployment (Figure 1). This corresponds to an average displacement rate of 62 km d−1, which is inconsistent with anguillid eel movement rates. The rapid transit suggests the tag was ingested by a predator and transported northward. During this period, there was no light recorded during daytime, while recorded temperatures gradually increased from 17°C to 25°C, consistent with the tag being inside an ectothermic predator, likely a non-lamnid shark, shortly after deployment (Figure 2). Depth data revealed an irregular vertical movement pattern, lacking the DVM typically observed in anguillid eels. Following surfacing, the tag drifted for approximately one week before being ingested again and transported for an additional six days (Figure 2).
Eight tags surfaced in the Coral Sea region, around the northern and northwestern flank of the Lord Howe Seamount Chain, between 1,666 and 2,567 km from their release locations (Figure 1). Of these, four tags were mrPATs (261324, 262013, 262014, and 262016) and surfaced in June, 47–78 days post-release, approximately 500 km west/southwest of New Caledonia. Three of these tags (262013, 262014, 262016) exhibited clear signs of mammalian predation with a distinct temperature spike to 36.5°C prior to surfacing (Figure 3). Without detailed depth data the species cannot be determined, but the high temperature precludes an endothermic fish. The remaining four tags were X-tags (262011, 261323, 261325, and 198002) and surfaced between July and September, 85–162 days post-release, 600–700 km northwest of New Caledonia (Figure 1). Three of these tags (261323, 261325, and 198002) surfaced in close proximity to one another (i.e., within 200–300 km) in the northeastern Coral Sea (15.5–17.6°S; 157.8–160.6°E), south of the Solomon Islands. All those tags surfaced prematurely, without sign of predation. Track reconstructions indicated that the eels were actively migrating northward at the time of tag release. Migration speeds during the final 10–20 days prior to surfacing were consistent with the overall average speeds, and no significant changes in DVM behavior were observed.
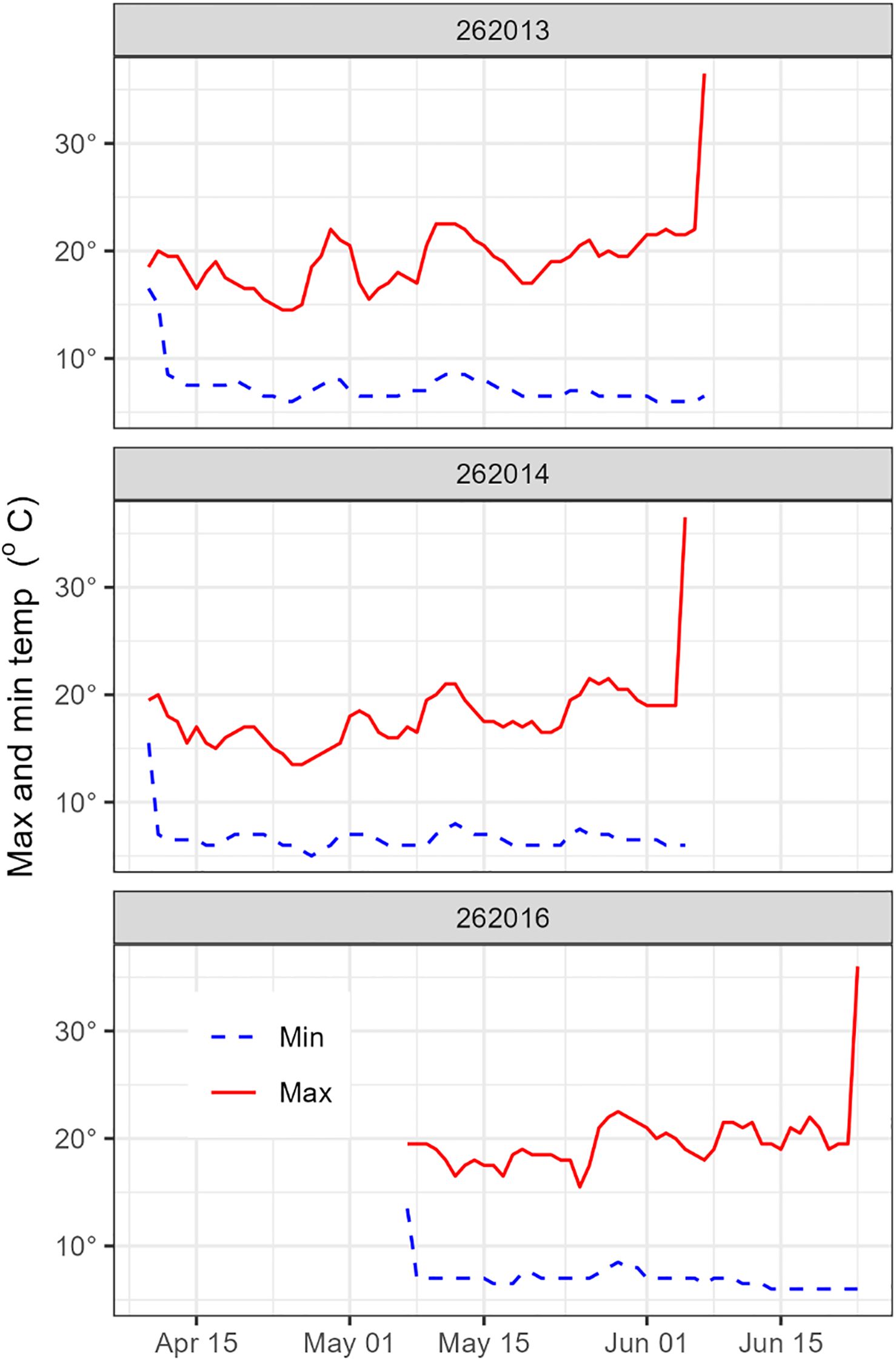
Figure 3. Minimum and maximum daily temperatures profiles showing examples of predation (likely by a whale) characterized by a distinct temperature spike to 36.5°C.
Reconstructed trajectories from the X-tag data revealed total migration distances ranging from approximately 2100 to 3400 km, with a mean trajectory length of 2900 km. The average migration speed across all individuals was 19.6 km d-1. However, migration speed between reconstructed positions exhibited substantial variability, with coefficients of variation ranging from 0.6 to 0.8, reflecting inherent uncertainties in geopositioning estimates. Migration speeds for two individuals (X-tags 261323 and 261325) decreased notably around 25–27°S, potentially corresponding to the region where the South Caledonian Jet intersects the Lord Howe Seamount Chain (Kessler and Cravatte, 2013, Oke et al., 2019).
Reconstruction of migratory pathways suggests that eels travelled several hundred kilometers offshore from the Australian coastline throughout much of their oceanic migration, venturing even farther offshore during the later stages of their journey (Figure 4). The reconstructed trajectories from two eels (198002 and 261323) suggest that they crossed or navigated around the Bellona Platform, a submarine carbonate platform located approximately 600 km west of New Caledonia, approximately 100 to 120 days after their release.
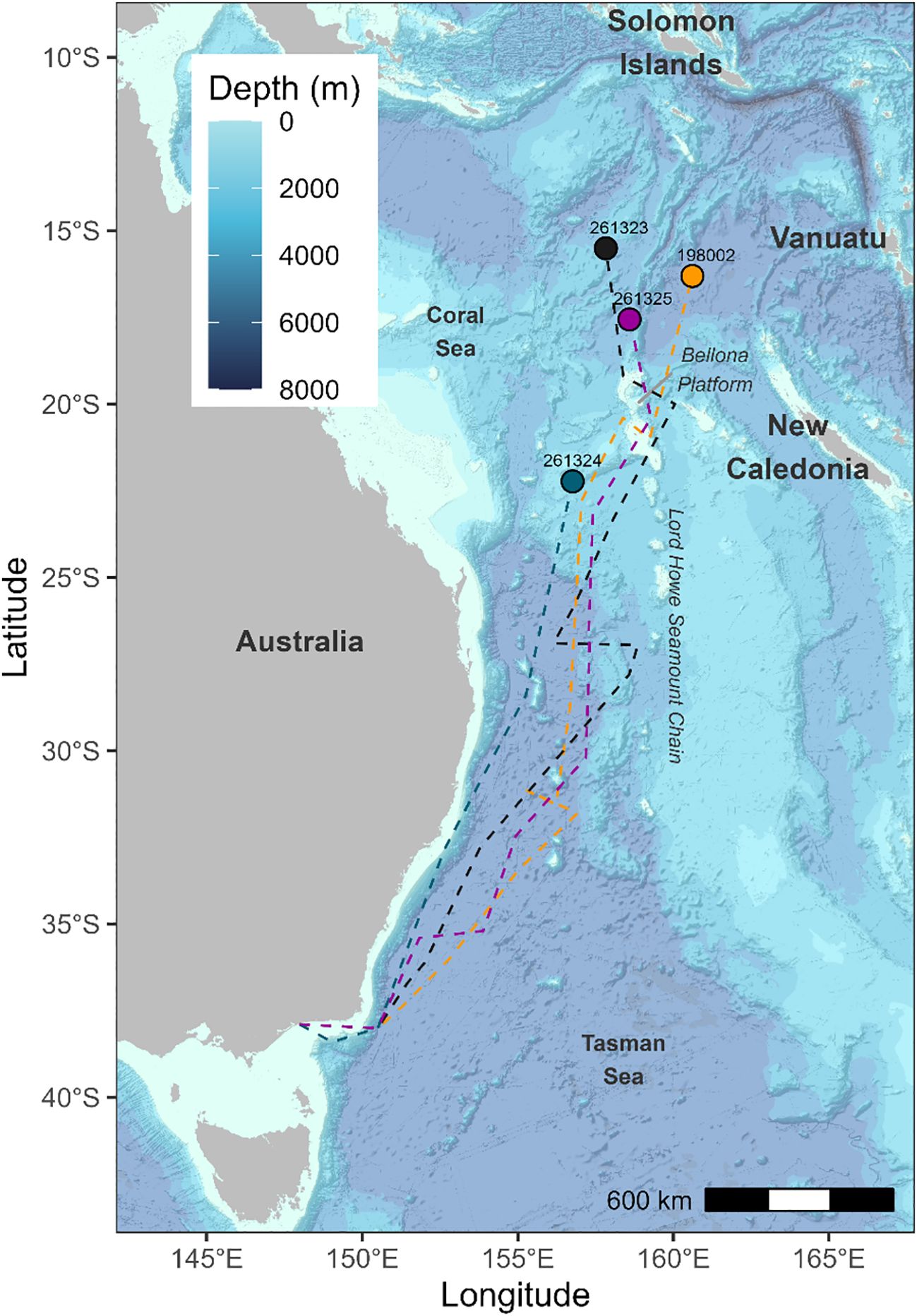
Figure 4. Reconstructed migratory pathways (dashed lines) and pop-up locations (coloured circles) of tagged long-finned eels.
Vertical movement behavior
Data from four of the eels (198002, 261323, 261324, and 261325) equipped with X-tags showed rapid descents to depths exceeding 650 meters within 1–5 days post-release (Figure 5). Shortly thereafter, they began exhibiting consistent DVM behavior, which characterized their oceanic journey. This pattern involved ascents to shallower depths (100–300 m) around sunset and descents to deeper waters (700–900 m) at sunrise (Figure 5). The duration of DVM ranged from 65 to 160 days, with a median of 106.5 days (Table 2). Daily maximum and minimum temperatures recorded by the tags attached to these eels closely align with data from individuals equipped with mrPAT tags (262011, 262013, 262014, and 262016), revealing highly similar thermal patterns (Figure 6). This consistency suggests that the tagged eels likely exhibited similar DVM behavior and followed comparable movement.
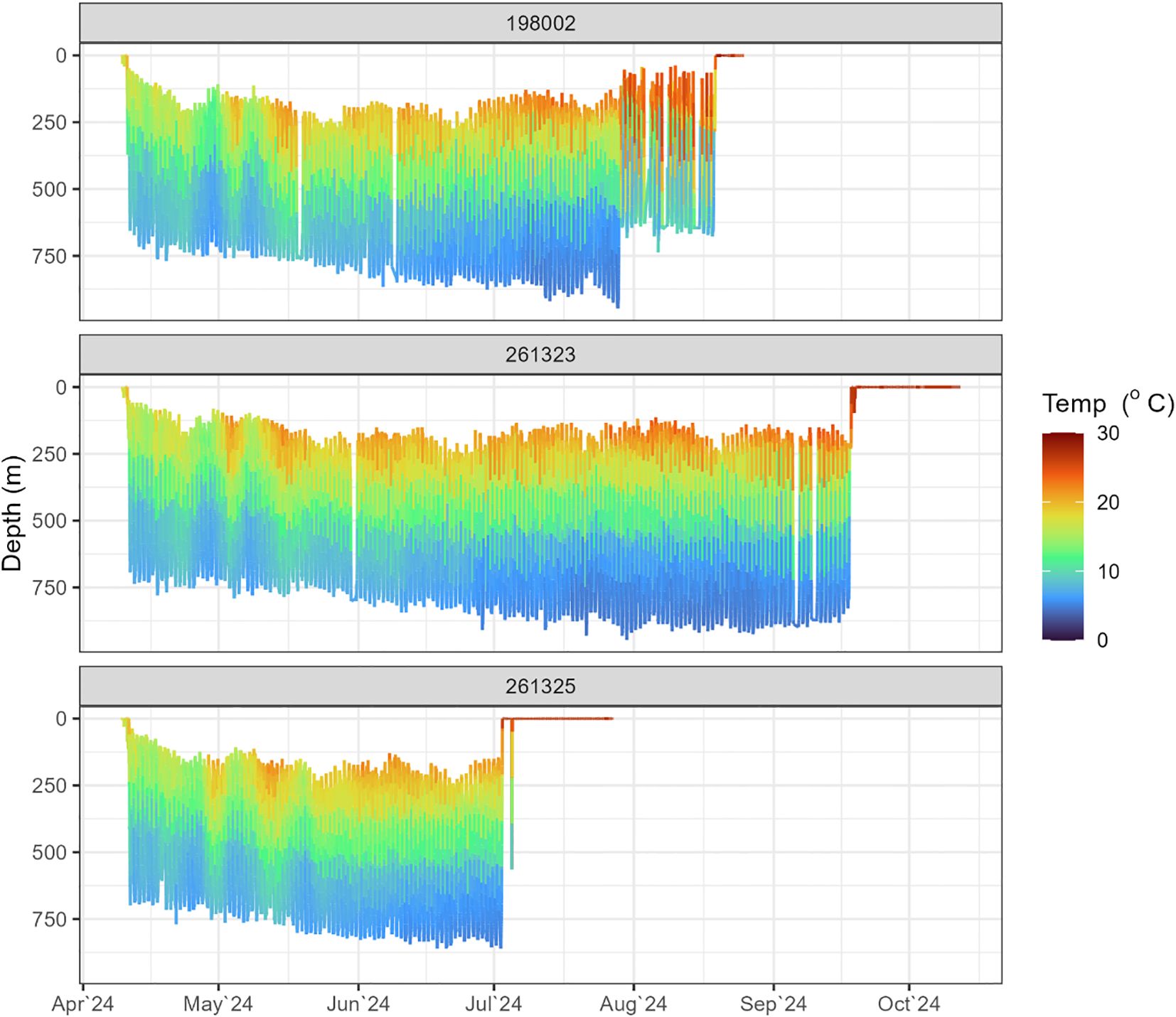
Figure 5. Examples of daily depth and temperature profiles of individual tagged long-finned eels, showing alternation between the warmer euphotic zone at night and the cooler mesopelagic zone during the day. Values around ‘0’ m depth indicate tag drifting on the surface.
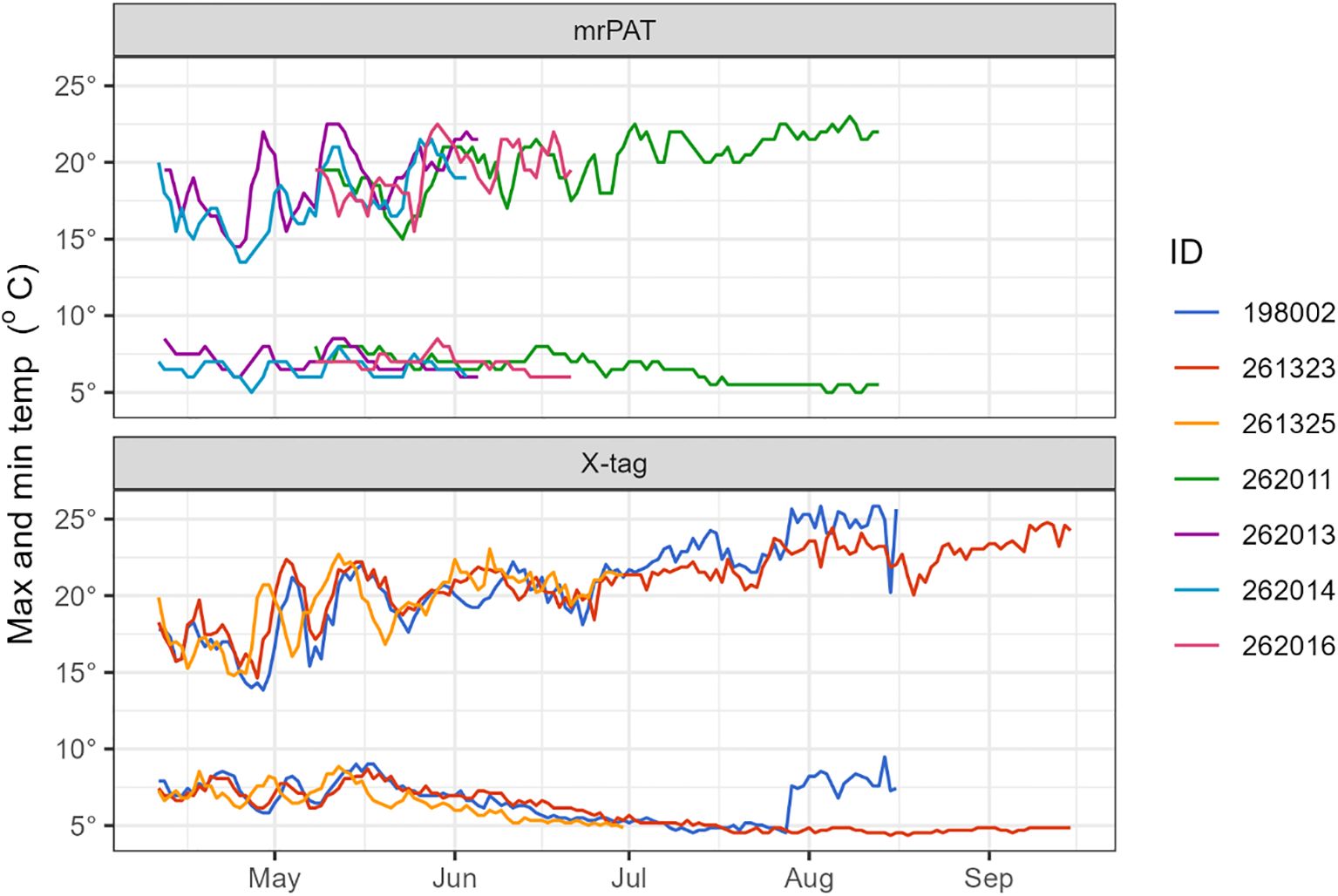
Figure 6. Daily maximum and minimum temperatures recorded by long-finned eels equipped with X-tags and mrPAT tags showing highly similar thermal patterns.
Mean night-time (2100−0400 h) swimming depth across eels equipped with X-tags (i.e., 198002, 261323, 261324, and 261325) was 194.0 ± 117.2 m (SD), and ranged from 168 m (261324) to 209 m (261325). Mean daytime (0800−1500 h) depth was 751.2 ± 158.3 m (SD) and ranged from 709 m (261324) to 797 m (261323) (Figure 7). Mean night-time temperature was 18.1 ± 4.3°C and ranged from 17.6°C (261325) to 19.2°C (261323). Mean daytime temperature was 6.7 ± 2.4 °C and ranged from 6.1 °C (261323) to 7.2°C (261324). Night-time temperatures increased as the eels migrated northward, whereas daytime temperatures typically show little variation regardless of location (Figure 5). Night-time swimming depth of long-finned eels increased with increased moonlight (Figure 8). For example, mean nighttime depth across eels 198002, 261323, and 261325 was 236.3 ± 83.2 m (SD) during the full moon compared to 184.3 ± 78.6 m (SD) during the new moon. The deepest recorded depth was 952 m by eel 198002. This individual also exhibited a notable shift in night-time swimming depth coinciding with its crossing of the Bellona Platform, occupying shallower depths (~600 m) during the final 50 days of its journey compared to the earlier phase (~800–900 m) (Figure 5).
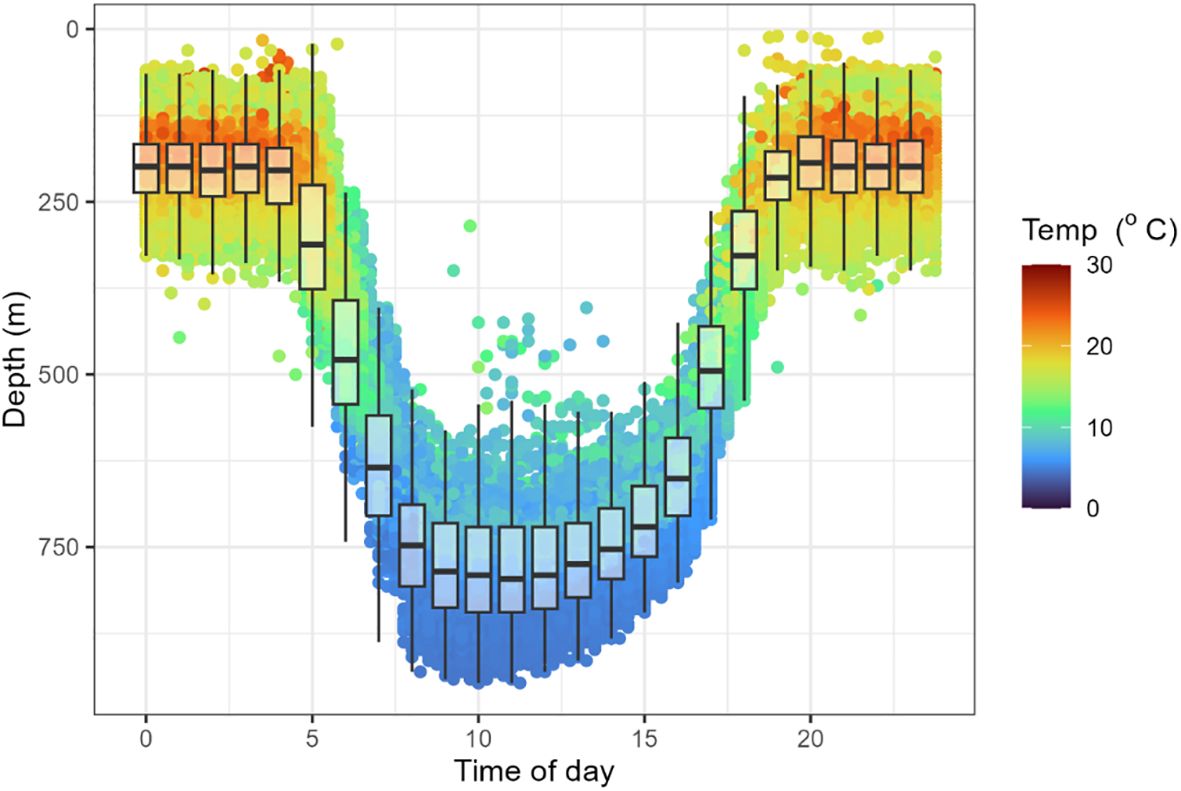
Figure 7. Diel depth and temperature data for tagged long-finned eels, showing occupation of warmer shallower environments at night and cooler deeper environments during the day. Plot shows raw values (dots), median (midline), lower and upper quartiles (ends of the box), and 1.5 x interquartile range (whiskers).
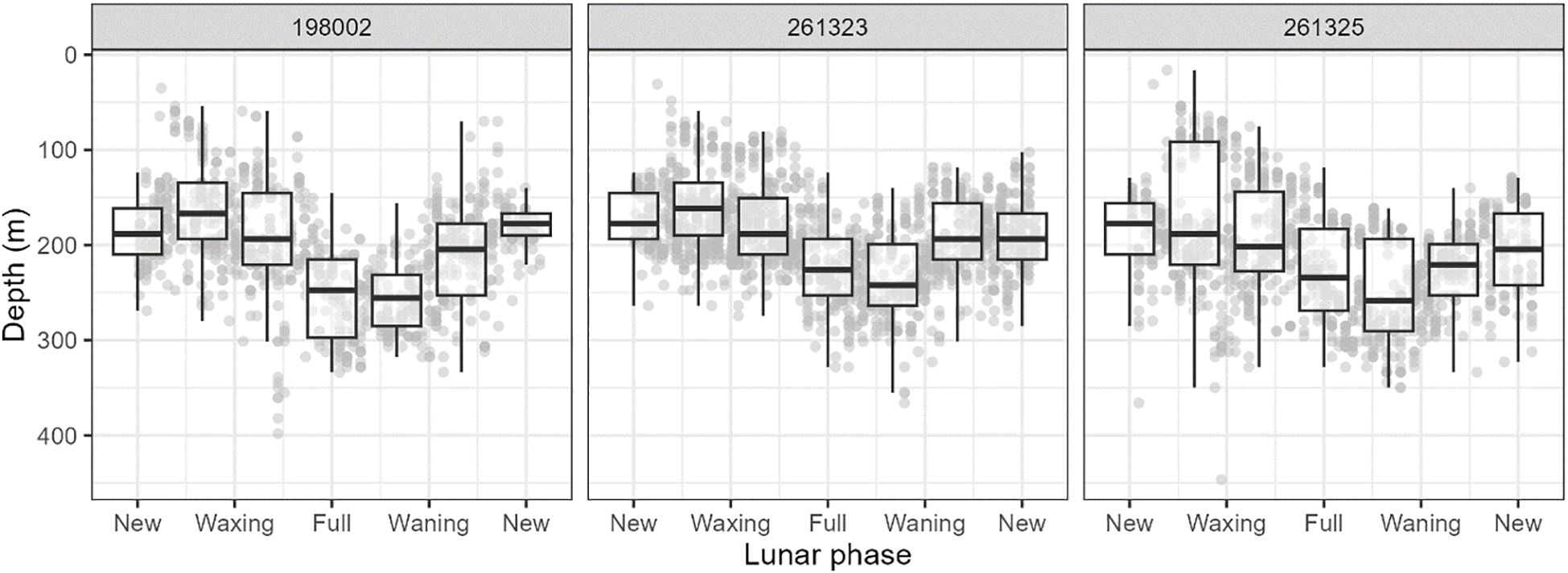
Figure 8. Examples of depth data for tagged long-finned eels as a function of lunar phase, showing use of deeper environments at night with increasing moonlight. Refer to Figure 7 for legend.
Discussion
This study has, for the first time, documented the long-distance migrations of Australian long-finned eels, from continental habitats in southern Australia to the tropical Coral Sea, located approximately 2,500 km away. Notably, the tags from the individuals that travelled the furthest surfaced within a relatively confined region in the northeastern Coral Sea (15.5–17.6°S; 157.8–160.6°E), south of the Solomon Islands. These tag endpoints align with the location of leptocephali collected by Kuroki et al. (2008, 2020), strengthening the hypothesis that the northeastern Coral Sea serves as a spawning area for this species. Reconstructed migratory trajectories indicate that long-finned eels follow a relatively consistent and predominantly direct route, generally tracking along the western slope of the Lord Howe Rise. The study also highlights the substantial risks encountered during migration, with tag data providing clear evidence of predation, underscoring that escapement to the sea does not necessarily equate to successful spawning. Furthermore, the consistent observation of DVM, characterized by nighttime ascents toward the surface and daytime descents to deeper waters, yields valuable information on behavioral adaptations during the oceanic phase of migration. This behavior, observed across all studied anguillid species during their spawning migrations, is likely a deeply rooted evolutionary trait.
Tag endpoints
The tags from the three eels that migrated the greatest distances (198002, 261323, 261325) surfaced between July and September 2024, clustering within a confined region of the northeastern Coral Sea (15.5–17.6°S; 157.8–160.6°E), south of the Solomon Islands. The estimated spawning period for long-finned eels is mid-June to August, based on back-calculated hatching dates derived from otolith analyses of leptocephali collected in a nearby region (~14–15.5°S, 160°E) during September 1995 and July 2016 (Kuroki et al., 2008, 2020). Although the precise timing and location of spawning remain unresolved, the spatial and temporal alignment of tag endpoints with historical leptocephali collection sites suggest that the probable spawning area of long-finned eels is located east of the tag endpoints in the north-eastern Coral Sea (Kuroki et al., 2020). Nevertheless, further research, including additional data from pop-up satellite archival tags and larval distribution surveys, are needed to delineate the precise spawning sites of long-finned eels. For example, the tags from the eels that travelled the furthest surfaced prematurely, suggesting they had not yet reached their final destinations.
Migratory pathways
The reconstructed paths of the X-tagged eels suggest that individuals travel several hundred kilometers offshore from the Australian coastline, generally following the western slope of the Lord Howe Rise. The surfacing positions of the mrPAT tags support this pattern. Upon reaching the Bellona Platform, the eels appear to disperse northward into the deeper waters of the Coral Sea. Consequently, the reconstructed trajectories of long-finned eels place them well east of the main axis of the southward-flowing East Australian Current (EAC). A recent study on the oceanic migration of short-finned eels (Anguilla australis) similarly found that individuals travelled east of the EAC en route to their presumed spawning grounds, although their migratory paths exhibited greater individual variability, particularly during the early stages of migration (Koster et al., 2021). More broadly, the migratory routes of anguillid eels toward their spawning areas appear to span a spectrum from relatively direct paths to more meandering trajectories, as documented in other species and regions (Righton et al., 2016, e.g., Chang et al., 2020; Franklin et al., 2023).
Tag endpoints and reconstructed migratory trajectories generally align with a region between the EAC and the Lord Howe Seamount Chain. Although this study did not identify a direct association with specific seamounts, their potential influence on migratory behavior and spawning site selection remains plausible. In the Japanese eel, seamounts have been linked to spawning activity, with eggs, leptocephali, and adults in spawning-condition collected near the West Mariana Ridge (Chow et al., 2009; Tsukamoto et al., 2011; Aoyama et al., 2014). Seamounts may act as navigational landmarks or aggregation points during migration (Righton et al., 2012), although their possible role in the spawning migrations of long-finned eel remains unresolved.
Migration speeds
The migration speeds of the eight long-finned eels with a long travel distance before premature loss or predation was 29 ± 6.5 km d-1. The coefficient of variation of the individual daily displacement rate was between 0.6 and 0.9. The average migration speed, and the degree of variation, are broadly consistent with those reported for other anguillid species (e.g., Aarestrup et al., 2009; Béguer-Pon et al., 2015; Righton et al., 2016; Franklin et al., 2023). Apart from the uncertainty in geolocation, which is reflected in the daily displacement data, the variability may also reflect factors such as physiological differences among individuals (e.g., body size or condition), as well as environmental factors encountered during migration, including ocean currents (Béguer-Pon et al., 2015; Righton et al., 2016). Notably, two individuals (261323 and 261325) showed a marked decrease in migration speed near 27°S, potentially corresponding to the interaction zone between the South Caledonian Jet and the Lord Howe Ridge, a region characterized by complex oceanographic dynamics (Kessler and Cravatte, 2013; Oke et al., 2019). A more detailed understanding of the factors influencing migration speed would be valuable, particularly in the context of environmental change. These include shifts in ocean circulation patterns and stressors in freshwater environments, such as habitat degradation, pollution, diseases and migration barriers that may impair body condition (Durif et al., 2006; Belpaire et al., 2009; Fenkes et al., 2016), and consequently affect swimming performance during the marine migration phase.
Vertical movement behavior
Long-finned eels exhibited regular DVM in deep oceanic waters. At night, they ascend to the warmer euphotic zone (~100–300 m, 17–19 °C), while during the day, they descend into the cooler mesopelagic zone (~700–900 m, 6–8 °C). DVM behavior is characteristic of Anguilla species and may serve multiple ecological functions. The daytime descent may reduce exposure to visual predators (Jellyman and Tsukamoto, 2005, Aarestrup et al., 2009). Alternating between shallow, warm waters and deeper, cooler waters has been hypothesized to influence thermal regulation and the timing of sexual maturation (Jellyman and Tsukamoto, 2005; Aarestrup et al., 2009; Jellyman and Tsukamoto, 2010). However, the significance of these factors in driving DVM remains undetermined (Righton et al., 2021).
The night-time swimming depth of long-finned eels increased with increased moonlight. As the water column becomes more brightly illuminated, the risk of detection by nocturnal predators may increase, prompting eels to descend to deeper waters during these periods (Schabetsberger et al., 2013). Similar lunar-associated increases in swimming depth have been observed in other anguillid species, including the giant mottled eel (A. marmorata) (Schabetsberger et al., 2013), Japanese eel (Chow et al., 2015), and New Zealand long-finned eel (Franklin et al., 2023). However, this behavior is not consistent across the genus; for instance, no significant correlation between depth and moonlight has been reported in the American eel (Béguer-Pon et al., 2015).
The depth and temperature ranges observed during DVM in long-finned eels were comparable to those reported for other anguillid species (e.g., Aarestrup et al., 2009; Schabetsberger et al., 2013; Béguer-Pon et al., 2015; Watanabe et al., 2020). However, one individual (198002) exhibited a distinct pattern in the later stages of migration, characterized by shallower daytime depths (approximately 500–600 m) and associated elevated temperatures (approximately 8–10°C). The sudden shift in DVM range and amplitude for this individual appears to coincide with crossing the Bellona Platform. Further telemetry data would be valuable to assess whether this pattern is more widespread among migrating eels. Such changes in vertical movement and thermal exposure for instance may have implications for reproductive maturation and spawning (Dou et al., 2008).
Predation of migrating eels
This study demonstrates that the oceanic migration of long-finned eels entails substantial predation risk. Almost half (8 out of 20) of the tagged eels showed signs of predation, and we cannot exclude that others were predated given the nature of the satellite telemetry data. Moreover, our findings reveal that these risks vary spatially and temporally, with different predators prevalent at distinct stages. In nearshore environments, sharks appear to be the primary predators. As the eels move into offshore waters, the nature of predation shifts, with whales emerging as a significant threat during the later stages of migration. Similar patterns of high and variable predation pressure have been reported in other anguillid eel species (e.g., European eel, New Zealand long-finned eel) (Righton et al., 2016; Westerberg et al., 2021; Jellyman et al., 2024). It is acknowledged pop-up satellite archival tags may increase susceptibility to predation or impair swimming performance (Burgerhout et al., 2011; Methling et al., 2011; Verhelst et al., 2022). However, a comparison of average migration speeds from geolocation data across the first and second halves of the three longest tracks revealed no decline in migration speed (28.2 ± 6.1 and 31.3 ± 8.7 km d-1 respectively). It can also be noted the average migration speed of eight A. australis tracked in the same area was essentially the same (29.3 ± 13 km d-1) (Koster et al., 2021) in spite of the fact that the tag to bodyweight ratio of A. australis was three times larger than that of the A. reinhardtii eels. This and the observation that many individuals travelled over 1500 km prior to predation suggests that swimming performance was not significantly impaired.
Despite these insights, our broader understanding of the ecological consequences of eel predation during oceanic migration remains limited. For example, to what extent does marine predation influence spawning stock size and reproductive success of anguillid eels (Righton et al., 2021; Stuart et al., 2024)? Could the recovery of marine mammal populations (e.g., whales) from historical over-exploitation (Whitehead and Shin, 2022) lead to increased predation on eels? Does mortality risk differ among eels migrating from different geographic regions, and could such variation inform the spatial prioritization of conservation efforts in continental habitats? Addressing such knowledge gaps is essential for developing more comprehensive and effective conservation strategies. The predation of ocean-migrating eels also illustrates their potential importance within marine food webs and raises additional ecological questions. For example, white sharks are known to undertake seasonal migrations to exploit temporally abundant prey resources (Semmens et al., 2013; Lipscombe et al., 2024). Whether the foraging behaviors of predators such as sharks are influenced by the timing and distribution of migrating eels is unknown.
Conclusions
This study provides the first direct evidence of the long-distance oceanic migrations of Australian long-finned eels. The convergence of tag endpoints within a confined area south of the Solomon Islands, closely aligned with historical leptocephali collections, supports the hypothesis that their probable spawning area lies in the northeastern Coral Sea (Kuroki et al., 2020). However, additional data are needed to delineate the precise spawning sites of the species. The findings highlight substantial and variable predation risks during migration and underscore the potential ecological role of eels in marine food webs. Observed diel vertical migration patterns, including lunar influences, provide insight into behavioral adaptations during the oceanic phase. Collectively, these results advance our understanding of anguillid eel migrations and underscore the critical importance of continued research into this crucial life-history stage. Such insights are vital for informing effective conservation strategies for this enigmatic and globally threatened genus (Jellyman, 2022).
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by Arthur Rylah Institute for Environmental Research Animal Ethics Commitee. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
WK: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Visualization, Writing – original draft, Writing – review & editing. HW: Data curation, Formal Analysis, Methodology, Visualization, Writing – original draft, Writing – review & editing. DD: Investigation, Methodology, Writing – original draft, Writing – review & editing. RK: Investigation, Methodology, Resources, Writing – original draft, Writing – review & editing. KA: Data curation, Resources, Writing – original draft, Writing – review & editing. BF: Data curation, Formal Analysis, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. Funding for this project was provided to Gunaikurnai Land and Waters Aboriginal Corporation (GLaWAC) by the Department of Energy, Environment, and Climate Action.
Acknowledgments
The authors thank Judy Stewart and Matt Holland from the GLaWAC for the opportunity to undertake the project. Thanks to Bryce Baxter, and Will Ingram and Paul Bodsworth from the Victorian Fisheries Authority, for providing field work support. Rowan Calder from Wildlife Computers is acknowledged for help with interpreting the data from PSATs. We thank Frank Amtstaetter and David Crook for constructive comments on earlier versions of this manuscript.
Conflict of interest
Author RK was employed by Gippsland Eels P/L.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. AI was used to improve writing clarity.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1690502/full#supplementary-material
References
Aarestrup K., Økland F., Hansen M. M., Righton D., Gargan P., Castonguay M., et al. (2009). Oceanic spawning migration of the European eel (Anguilla Anguilla). Science 325, 1660–1660. doi: 10.1126/science.1178120
Aoyama J., Watanabe S., Miller M. J., Mochioka N., Otake T., Yoshinaga T., et al. (2014). Spawning sites of the Japanese eel in relation to oceanographic structure and the West Mariana Ridge. PloS One 9, e88759. doi: 10.1371/journal.pone.0088759
Béguer-Pon M., Castonguay M., Shan S., Benchetrit J., and Dodson J. J. (2015). Direct observations of American eels migrating across the continental shelf to the Sargasso Sea. Nat. Commun. 6, 8705. doi: 10.1038/ncomms9705
Belpaire C., Goemans G., Geeraerts C., Quataert P., Parmentier K., Hagel P., et al. (2009). Decreasing eel stocks: survival of the fattest? Ecol. Freshw. fish 18, 197–214. doi: 10.1111/j.1600-0633.2008.00337.x
Burgerhout E., Manabe R., Brittijn S. A., Aoyama J., Tsukamoto K., and van den Thillart G. E. (2011). Dramatic effect of pop-up satellite tags on eel swimming. Naturwissenschaften 98, 631–634. doi: 10.1007/s00114-011-0805-0
Chang Y.-L. K., Olmo G. D., and Schabetsberger R. (2020). Tracking the marine migration routes of South Pacific silver eels. Mar. Ecol. Prog. Ser. 646, 1–12. doi: 10.3354/meps13398
Chapman B. B., Hulthén K., Wellenreuther M., Hansson L.-A., Nilsson J-Å, and Brönmark C. (2014). Patterns of animal migration. Anim. movement across scales 1, 11–35. doi: 10.1093/acprof:oso/9780199677184.003.0002
Chow S., Kurogi H., Mochioka N., Kaji S., Okazaki M., and Tsukamoto K. (2009). Discovery of mature freshwater eels in the open ocean. Fisheries Sci. 75, 257–259. doi: 10.1007/s12562-008-0017-5
Chow S., Okazaki M., Watanabe T., Segawa K., Yamamoto T., Kurogi H., et al. (2015). Light-sensitive vertical migration of the Japanese eel Anguilla japonica revealed by real-time tracking and its utilization for geolocation. PloS One 10, e0121801. doi: 10.1371/journal.pone.0121801
Cleveland W. S. and Devlin S. J. (1988). Locally weighted regression: an approach to regression analysis by local fitting. J. Am. Stat. Assoc. 83, 596–610. doi: 10.1080/01621459.1988.10478639
Dou S.-Z., Yamada Y., Okamura A., Shinoda A., Tanaka S., and Tsukamoto K. (2008). Temperature influence on the spawning performance of artificially-matured Japanese eel, Anguilla japonica, in captivity. Environ. Biol. fishes 82, 151–164. doi: 10.1007/s10641-007-9268-8
Durif C. M., Dufour S., and Elie P. (2006). Impact of silvering stage, age, body size and condition on reproductive potential of the European eel. Mar. Ecol. Prog. Ser. 327, 171–181. doi: 10.3354/meps327171
Fenkes M., Shiels H. A., Fitzpatrick J. L., and Nudds R. L. (2016). The potential impacts of migratory difficulty, including warmer waters and altered flow conditions, on the reproductive success of salmonid fishes. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol. 193, 11–21. doi: 10.1016/j.cbpa.2015.11.012
Franklin P. A., Jellyman D., Baker C., Birnie-Gauvin K., Jellyman P., and Aarestrup K. (2023). Oceanic migration routes and behavior of the New Zealand longfin eel Anguilla dieffenbachii. Mar. Ecol. Prog. Ser. 724, 127–139. doi: 10.3354/meps14443
Jellyman D., Franklin P., Aarestrup K., Birnie-Gauvin K., Baker C., and Jellyman P. (2024). Evidence of marine predation on silver New Zealand longfin eels, Anguilla dieffenbachii. Fisheries Manage. Ecol. 31, e12695. doi: 10.1111/fme.12695
Jellyman D. and Tsukamoto K. (2005). Swimming depths of offshore migrating longfin eels Anguilla dieffenbachii. Mar. Ecol. Prog. Ser. 286, 261–267. doi: 10.3354/meps286261
Jellyman D. and Tsukamoto K. (2010). Vertical migrations may control maturation in migrating female Anguilla dieffenbachii. Mar. Ecol. Prog. Ser. 404, 241–247. doi: 10.3354/meps08468
Jellyman D. J. (2022). An enigma: how can freshwater eels (Anguilla spp.) be such a successful genus yet be universally threatened? Rev. Fish Biol. Fish. 32, 701–718.
Kessler W. S. and Cravatte S. (2013). Mean circulation of the coral sea. J. Geophysical Research: Oceans 118, 6385–6410. doi: 10.1002/2013JC009117
Koster W. M., Aarestrup K., Birnie-Gauvin K., Church B., Dawson D., Lyon J., et al. (2021). First tracking of the oceanic spawning migrations of Australasian short-finned eels (Anguilla australis). Sci. Rep. 11, 1–13. doi: 10.1038/s41598-021-02325-9
Kuroki M., Aoyama J., Miller M. J., Watanabe S., Shinoda A., Jellyman D. J., et al. (2008). Distribution and early life-history characteristics of anguillid leptocephali in the western South Pacific. Mar. Freshw. Res. 59, 1035–1047. doi: 10.1071/MF08041
Kuroki M., Miller M. J., Feunteun E., Sasal P., Pikering T., Han Y.-S., et al. (2020). Distribution of anguillid leptocephali and possible spawning areas in the South Pacific Ocean. Prog. oceanography 180, 102234. doi: 10.1016/j.pocean.2019.102234
Lennox R. J., Chapman J. M., Souliere C. M., Tudorache C., Wikelski M., Metcalfe J. D., et al. (2016). Conservation physiology of animal migration. Conserv. Physiol. 4, cov072. doi: 10.1093/conphys/cov072
Lipscombe R. S., Meyer L., Butcherine P., Morris S., Huveneers C., Scott A., et al. (2024). A taste of youth: Seasonal changes in the diet of immature white sharks in eastern Australia. Front. Mar. Sci. 11, 1359785. doi: 10.3389/fmars.2024.1359785
Manabe R., Aoyama J., Watanabe K., Kawai M., Miller M. J., and Tsukamoto K. (2011). First observations of the oceanic migration of Japanese eel, from pop-up archival transmitting tags. Mar. Ecol. Prog. Ser. 437, 229–240. doi: 10.3354/meps09266
Methling C., Tudorache C., Skov P. V., and Steffensen J. F. (2011). Pop up satellite tags impair swimming performance and energetics of the European eel (Anguilla Anguilla). PloS One 6, e20797. doi: 10.1371/journal.pone.0020797
Oke P. R., Pilo G. S., Ridgway K., Kiss A., and Rykova T. (2019). A search for the tasman front. J. Mar. Syst. 199, 103217. doi: 10.1016/j.jmarsys.2019.103217
Økland F., Thorstad E. B., Westerberg H., Aarestrup K., and Metcalfe J. D. (2013). Development and testing of attachment methods for pop-up satellite archival transmitters in European eel. Anim. Biotelemetry 1, 1–13. doi: 10.1186/2050-3385-1-3
Righton D., Aarestrup K., Jellyman D., Sébert P., van den Thillart G., and Tsukamoto K. (2012). The Anguilla spp. migration problem: 40 million years of evolution and two millennia of speculation. J. Fish Biol. 81, 365–386. doi: 10.1111/j.1095-8649.2012.03373.x
Righton D., Piper A., Aarestrup K., Amilhat E., Belpaire C., Casselman J., et al. (2021). Important questions to progress science and sustainable management of anguillid eels. Fish Fisheries 22, 762–788. doi: 10.1111/faf.12549
Righton D., Westerberg H., Feunteun E., Økland F., Gargan P., Amilhat E., et al. (2016). Empirical observations of the spawning migration of European eels: The long and dangerous road to the Sargasso Sea. Sci. Adv. 2, e1501694. doi: 10.1126/sciadv.1501694
Schabetsberger R., Chang Y.-L. K., and Miller M. J. (2021). Spawning migration and larval dispersal of tropical Pacific eels (Anguilla spp.) in the centre of their distribution ranges. Mar. Ecol. Prog. Ser. 670, 167–184. doi: 10.3354/meps13745
Schabetsberger R., Økland F., Aarestrup K., Kalfatak D., Sichrowsky U., Tambets M., et al. (2013). Oceanic migration behavior of tropical Pacific eels from Vanuatu. Mar. Ecol. Prog. Ser. 475, 177–190. doi: 10.3354/meps10254
Schlitzer R. (2015). Data analysis and visualization with Ocean Data View. CMOS Bull. SCMO 43, 9–13. doi: 10.1016/j.semcdb.2015.03.011
Semmens J. M., Payne N., Huveneers C., Sims D. W., and Bruce B. (2013). Feeding requirements of white sharks may be higher than originally thought. Sci. Rep. 3, 1471. doi: 10.1038/srep01471
Stuart R. E., Stockwell J. D., and Marsden J. E. (2024). Anguillids: Widely studied yet poorly understood—A literature review of the current state of Anguilla eel research. Rev. Fish Biol. Fisheries 34, 1637–1664. doi: 10.1007/s11160-024-09892-w
Tsukamoto K. (2009). Oceanic migration and spawning of anguillid eels. J. fish Biol. 74, 1833–1852. doi: 10.1111/j.1095-8649.2009.02242.x
Tsukamoto K., Chow S., Otake T., Kurogi H., Mochioka N., Miller M. J., et al. (2011). Oceanic spawning ecology of freshwater eels in the western North Pacific. Nat. Commun. 2, 179. doi: 10.1038/ncomms1174
Verhelst P., Aarestrup K., Hellström G., Jepsen N., Koed A., Reubens J., et al. (2022). The effect of externally attached archival data loggers on the short-term dispersal behavior and migration speed of European eel (Anguilla Anguilla L.). Anim. Biotelemetry 10, 9. doi: 10.1186/s40317-022-00280-4
Watanabe S., Higuchi T., Noshiro M., Manabe R., Miller M. J., Jellyman D. J., et al. (2020). Reexamination of the spawning migration of Anguilla dieffenbachii in relation to water temperature and the lunar cycle. New Z. J. Mar. Freshw. Res. 54, 131–147. doi: 10.1080/00288330.2019.1614075
Westerberg H., Amilhat E., Wahlberg M., Aarestrup K., Faliex E., Simon G., et al. (2021). Predation on migrating eels (Anguilla Anguilla L.) from the Western Mediterranean. J. Exp. Mar. Biol. Ecol. 544, 151613. doi: 10.1016/j.jembe.2021.151613
Keywords: anguillid, spawning migration, diel vertical migration (DVM), pop-up satellite archival tag (PSAT), catadromy
Citation: Koster WM, Westerberg H, Dawson D, Kahsnitz R, Aarestrup K and Fanson B (2025) Revealing the oceanic spawning migrations of Australian long-finned eels (Anguilla reinhardtii) via satellite telemetry. Front. Mar. Sci. 12:1690502. doi: 10.3389/fmars.2025.1690502
Received: 21 August 2025; Accepted: 24 October 2025;
Published: 10 November 2025.
Edited by:
Tomas Chalde, CONICET Centro Austral de Investigaciones Científicas (CADIC), ArgentinaReviewed by:
Martin C. Arostegui, Woods Hole Oceanographic Institution, United StatesPatricia Zarate, Instituto de Fomento Pesquero, Chile
Copyright © 2025 Koster, Westerberg, Dawson, Kahsnitz, Aarestrup and Fanson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Wayne M. Koster, d2F5bmUua29zdGVyQGRlZWNhLnZpYy5nb3YuYXU=