 Marta Román1*
Marta Román1* Elsa Vázquez1
Elsa Vázquez1 Nicolás Weidberg1,2Rosa M. Viejo3Salvador Román1
Nicolás Weidberg1,2Rosa M. Viejo3Salvador Román1 Jesús S. Troncoso1
Jesús S. Troncoso1 David S. Wethey4Sarah A. Woodin4
David S. Wethey4Sarah A. Woodin4 Celia Olabarria1
Celia Olabarria1- 1Centro de Investigación Mariña, and Departamento de Ecoloxía e Bioloxía animal. Universidade de Vigo, EcoCost, Facultade de Ciencias do Mar, Edificio C. C. Experimentais, Vigo, Spain
- 2Departamento de Biología de Organismos y Sistemas, Universidad de Oviedo, Oviedo, Spain
- 3Área de Biodiversidad y Conservación, Universidad Rey Juan Carlos, Madrid, Spain
- 4Department of Biological Sciences, University of South Carolina, Columbia, SC, United States
More intense and frequent atmospheric heatwaves in the future may reduce the growth and survival of intertidal organisms. However, interspecific facilitative interactions can buffer legacy effects of thermal stress and favor recovery of the affected organisms. To test the role of facilitation, we exposed assemblages of Zostera noltei and juveniles of Venerupis corrugata, Ruditapes decussatus and R. philippinarum to a low-tide atmospheric heatwave in mesocosm experiments. A four-hour low-tide emersion period was applied daily for four consecutive days. During the heatwave, the temperatures of 108 experimental units were monitored withhermocouples. Assemblages were transplanted to two intertidal flats (Combarro and Noia) in NW Spain, and different temperature, salinity and nutrient concentrations were measured during the experiment. After two and a half months, there were no differences in shoot abundance or above-ground biomass between seagrass affected by legacy heat stress and control seagrass growing together with clams; however, the values of both variables were lower in the seagrass only scenario, suggesting clam-derived facilitation. At Combarro, characterized by higher temperature and salinity, a mutualistic interaction between clams and Z. noltei was observed, with greater above-ground biomass and more abundant shoots in the seagrass and enhanced growth of clams. No such interactions were observed at Noia, where the seagrass biomass was significantly lower than at Combarro. At Noia, characterized by higher nutrient concentrations in seawater, the presence of Ulva spp. canopies above the seagrass and higher levels of N in Z. noltei shoots indicated that eutrophic conditions hampered seagrass growth and mutualistic interactions with clams. The findings suggest that species-specific responses and abiotic context shape the magnitude and direction of ecological interactions.
1 Introduction
Record global surface temperatures in 2011–2024 indicate accelerated warming relative to the 1850–1900 period (ECMWF, 2025). Monthly anomalies beyond the 1.5°C threshold occurred throughout 2024 and are projected to continue in the future, thus exacerbating previously observed negative effects on natural systems (IPCC, 2023; Cannon, 2025). The human influence on climate has also increased the frequency of episodes of extreme heat, which are projected to become even more frequent and which can affect the growth, reproduction, survival and geographical distribution of species (IPCC, 2023; Stillman, 2019). These events can be particularly harmful to intertidal ecosystems, which are subjected to temperature and humidity oscillations (Rafaelli and Hawkins, 1996).
Recent studies of intertidal species have mainly focused on marine heatwaves (Wernberg et al., 2024). However, atmospheric heatwaves are of concern in relation to the composition and functioning of intertidal ecosystems, as they can affect the physiological performance and survival of sessile and sedentary emerged species (Southward, 1958). Although marine heatwaves are well defined (Hobday et al., 2016), there is no standardized definition of atmospheric heatwaves (AEMET, 2024). Thus, a variety of days and temperature percentile thresholds are applied. In general, atmospheric heatwaves consist of periods of three or more consecutive days when the maximum daily temperatures in summer months exceed a defined percentile (90th or higher) of the long-term records (years to decades) (Perkins and Alexander, 2013; AEMET, 2024).
More frequent and intense atmospheric heatwaves may be harmful to sessile intertidal organisms (Helmuth et al., 2006; Stillman, 2019), such as macroalgae, seagrass and invertebrates. Heat-stressed organisms experience body heating and water loss and spend less energy on feeding, growth and reproduction, at the expense of maintaining homeostasis for survival (Shick et al., 1988). Nonetheless, these physiological mechanisms cannot guarantee survival if the heat stress exceeds the tolerance thresholds of the organisms (Massa et al., 2009; Domínguez et al., 2021). Most research on intertidal heatwaves has focused on epibenthic organisms inhabiting rocky shores, while the effects on infaunal communities in soft-bottom substrates—where temperature fluctuations are dampened with depth— remain comparatively understudied. Recent literature focusing on these ecosystems has provided evidence of the negative effects of heatwaves on the behavior, growth, reproduction, and survival of bivalves (Vázquez et al., 2021; He et al., 2022; Luo et al., 2025) as well as on the photosynthetic efficiency, tissue integrity, and spatial cover of seagrasses (Massa et al., 2009; Danovaro et al., 2020; Román et al., 2023). These studies primarily examined impacts immediately after a heatwave or following a short recovery period of days. However, there remains a lack of research on legacy effects, i.e., the long-lasting impacts of a past disturbance after it has ceased (e.g., Vaz-Pinto et al., 2013; Reynolds et al., 2016). For instance, during the 2021 heat dome that affected Western North America, air temperatures near 40°C during low spring tides caused massive shellfish mortality, with unknown long-term consequences (Raymond et al., 2022). Assessing the legacy effects of atmospheric heatwaves on intertidal communities (e.g., Vaz-Pinto et al., 2013; Reynolds et al., 2016), is therefore crucial for understanding the potential long-term consequences.
Positive interactions between neighboring species, such as facilitation and mutualism (Bronstein, 2009), may be enhanced by abiotic stress (Bertness and Callaway, 1994; Chin et al., 2021; Cardini et al., 2022; Román et al., 2024). The stress-gradient hypothesis is exemplified by several cases in nature (Thompson, 1988). Generally, the positive reciprocal interactions between bivalves and angiosperms may be intensified by high water temperatures (Clemente and Thomsen, 2023). Specifically, recent studies have demonstrated that Zostera noltei Hornemann provided short-term facilitation to infaunal bivalves under low-tide thermal stress. After an atmospheric heatwave at low tide, photosynthetic efficiency was higher in Z. noltei growing with Ruditapes philippinarum (Adams & Reeve, 1850), probably due to increased amounts of phosphate in the sediment (Román et al., 2023). Growth of R. decussatus (Linnaeus, 1758) was also enhanced in sediment with Z. noltei relative to bare sediment when temperatures rose by 0.3–1.1°C, as shading by the plants kept the sediment cooler (Román et al., 2022). Despite such short-term positive interactions, sublethal effects can persist long after the stress has ceased and may be critical for the survival and physiological performance of species and the structure of intertidal communities (Raymond et al., 2022). Although the importance of heatwave legacy effects has been recognized, the effects have seldom been studied in clam-seagrass interactions.
The coast of the NW Iberian Peninsula is a suitable site for studying the legacy effects of heatwaves on ecological interactions in soft bottom intertidal communities, as extreme temperature events are likely to increase in this region (Gómez-Gesteira et al., 2011; Carvalho et al., 2022). In tidal flats in the inner parts of rias (sensu Von Richthofen, 1886), the sedimentary substrate is colonized by seagrass meadows composed by Z. noltei (25.52 km2) and Zostera marina Linnaeus (4.61 km2) (Cacabelos et al., 2015). Professional clam seeding and harvesting also take place in many of these areas (Frangoudes et al., 2008). Understanding responses of clam-seagrass interactions to legacy thermal stress will help to aid management decisions aimed at guaranteeing the sustainability of commercial clam stocks and the biologically important seagrass meadows that cohabit intertidal flats. Ensuring a proper balance between sustainable small-scale fisheries and seagrass conservation is essential to maintain ecosystem services provided by seagrass meadows. Seagrass functions comprise coastal protection against floodings and sea level rise, ecosystem engineering through substrate accretion and currents deceleration, atmospheric CO2 storage (Miyajima and Hamaguchi, 2019), natural removal of organic anthropogenic pollutants, nursery for larvae and/or juvenile stages of pelagic and benthic species, habitat and food for megafauna, and support for tourism and recreational activities (UNEP, 2020).
The aim of this study was to test whether reciprocal positive interactions between the commercial clam species Venerupis corrugata (Gmelin, 1791), R. decussatus and R. philippinarum and the temperate intertidal seagrass Z. noltei could buffer the negative legacy effects of thermal stress caused by an atmospheric heatwave during low tide. The following specific predictions were tested: 1) the presence of clams would ameliorate the negative effects of past low-tide heatwave stress on biomass, shoot abundance and carbon, nitrogen and carbohydrate contents of Z. noltei, 2) the presence of Z. noltei would ameliorate the negative effects of past low-tide heatwave stress on survival, growth and condition index of clams; and 3) the local environmental context would affect specific responses and interactions. To test these hypotheses, we carried out a mesocosm experiment in which assemblages of these organisms were exposed to a diurnal four-hour low tide period for four days, followed by four days of recovery. After the recovery period, the assemblages were transplanted (in summer) to two field sites characterized by different temperatures, salinity levels and nutrient concentrations for two and a half months.
2 Materials and methods
2.1 Heatwave in mesocosm
On 2 May 2022, sediment (1.9 m³) was collected in situ from Combarro (NW Spain; 42.437° N, 8.690° W), an intertidal flat partially covered by a Zostera noltei meadow where clam shell fishing is practiced (Figure 1), and sieved through a 5 mm mesh to prevent the collection of clam recruits. A sample of Z. noltei canopy and the rhizosphere sediment, was collected digging with a shovel to a depth of 10 cm. Care was taken to avoid damaging the rhizosphere. The average shoot density was 6520 ± 1459 shoots m−2 (Román et al., 2024). Samples were transported to the Toralla Marine Science Station (42.202° N, 8.801° W) (ECIMAT-University of Vigo). The mesocosm facility of ECIMAT consists of 12 tanks of capacity 800 L, supplied with running filtered (50 µm) seawater. The tanks are located outdoors below a methacrylate roof so that the system is lit directly by sunlight. During the experiment, a semi-diurnal tidal regime was performed in the tanks, simulating the natural conditions experienced by intertidal organisms.
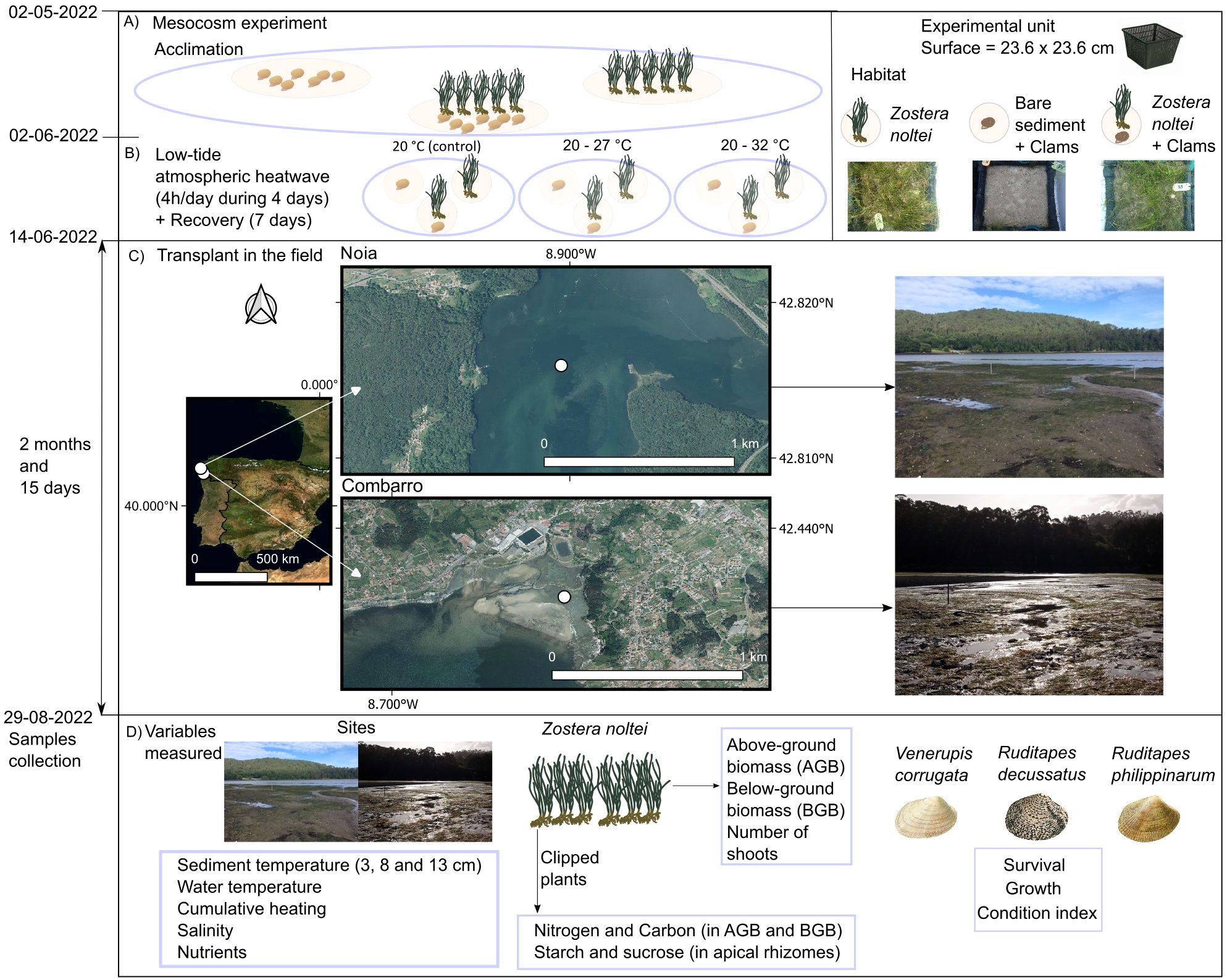
Figure 1. Chronology of experimental design. (A) Acclimation to the mesocosm and scheme of habitat treatments, (B) application of heatwave conditions during low tide at the mesocosm + recovery, (C) transplantation to the Noia and Combarro shellfishing beds for a period of two and a half months, (D) abiotic and biotic variables measured. Aerial orthophotographs of Figure 1C were obtained from Google satellite. Maps Data: Google (c) 2025/ Airbus. Maxar Technologies.
The sediment was placed in 108 plastic pond planting baskets (Figure 1, “experimental unit”, dimensions: 15 cm × 23.6 × 23.6 cm) lined with thick garden mesh that allowed the water to circulate. One third of the baskets (n=36) were filled with sediment only, and the remaining baskets (n = 72) were filled with sediment at the same height and portions of the Z. noltei canopy with the associated rhizosphere (mean density of 529 ± 180 shoots, and biomass of 5.9 ± 1.53 g dw per basket). Nine baskets (6 with Z. noltei and 3 with bare sediment) were haphazardly placed in each tank and left acclimatizing to the mesocosm conditions for one month (2 May - 2 June 2022). During this period, the salinity was 34.93 ± 0.004 and seawater temperature was 14.63 ± 0.03°C in the mesocosm (n=542). Mean daily solar radiation (W m-2) during the acclimatization period was 258 ± 98 (n = 32) at the weather station closest to the mesocosm unit (Vigo port). The tanks were covered with greenhouse plastic during two days of intense precipitation to prevent any decrease in salinity.
After acclimatization of the seagrass, juvenile clams of each species (V. corrugata, R. decussatus and R. philippinarum), of sizes between 15 and 18 mm, were obtained from hatcheries and transported to the mesocosm. We used juvenile clams because the effects of environmental stress on early life stages of marine invertebrates are less studied, despite their higher sensitivity (Pineda et al., 2012). The three clam species were mixed to replicate the typical conditions in seagrass-vegetated shellfish beds in NW Spain. The clams were submersed for 2 hours in ambient seawater containing 250 mg L–1 of the fluorochrome calcein (Sigma, CAS 1461-15-0), to mark the edges of the shells and thus enable subsequent measurement of shell growth. Eleven juvenile clams of each species (33 clams per basket) were placed on the surface of the sediment in 6 baskets per tank (three with Z. noltei and three with bare sediment), and the remaining 3 baskets with Z. noltei were left without clams. The clams were allowed to burrow and acclimatize for 24 h (31 May - 1 June 2022). The 33 clams yielded a density of 589 ind. m −2, which is within the optimal range for seeding densities at the shellfish banks (300–800 ind. m–2; Navajas et al., 2003) and prevents the appearance of density-dependent effects on growth or mortality (Royo et al., 2002; Melià and Gatto, 2005). During the acclimation period, any dead individuals observed on the sediment surface were replaced. The clams were fed daily with a mixture of Isochrysis galbana (TISO), Tetraselmis suecica, Chaetoceros gracilis and Rodomonas lens (1% of microalgae per clam [DW]).
The experimental low tide heatwave (HW) treatments applied in the mesocosm facility (3–6 June 2022) consisted of increasing the sediment temperature to 3 target ranges of 20-20°C (control), 20-27°C and 20-32°C, each applied to 4 independent replicate tanks. The temperatures and treatment durations simulated warming events recorded in intertidal flats in northwestern Spain (Supplementary Figure 1) (Domínguez et al., 2021). Metallic structures with arrays of 12,150-W ceramic heating lamps (Ceramix FTE-150) were placed above the treatment tanks and attached to the mesocosm roof with chains, so that they could be lowered during the low tide heatwave periods and raised at other times. The treatments consisted of gradual increasing the sediment temperature during the 4-h diurnal low tide periods with infrared radiation; the process was repeated daily during 4 consecutive days. To reproduce the gradual heating experienced in the field the lamps were connected to digital controllers (Omega CN7853), which were attached to feedback thermocouples placed at 2 cm depth in the sediment of each basket. Field observations and previous research indicated that the rhizosphere reaches a depth of 2 cm (Román et al., 2023). The thermocouples recorded the sediment temperature in each basket every 15 seconds. To prevent wind-driven evaporative cooling, the open tops of the tanks, including controls, were covered with insect screening mesh (1.6 x 1.6 mm mesh size), only during application of HW treatments (4 hours per day for 4 days). The mesh reduced the photosynthetically active radiation (PAR) by 58%, which could have affected photosynthesis during the treatment period. After exposure to heatwave conditions, the tanks were held under ambient solar radiation, air and water temperatures for a 4-day recovery period, and the baskets from all tanks were then transported to the field for transplantation (Figure 1).
2.2 Transplantation in the field
The baskets were transplanted on 14 June 2022 at Combarro and on 15 June 2022 at Noia, both sites located in NW Spain (Figure 1). Noia (coordinates 42.81594°N, 8.90054°W) is located at the inner part of the Ria de Muros-Noia, and is colder, rainier, less saline and more nutrient-rich than the other site (Román et al., 2024). This experimental site was located on an island at the outflow of the Tambre, Donas and Santa Baia rivers that drain an extensive watershed of 1600 km2 (areas calculated from GIS layers, available at https://www.miteco.gob.es/es/cartografia-y-sig/ide/descargas/agua.html). The second transplant site was Combarro (the same site where the sediment and rhizosphere were collected). This site is exposed to higher temperatures and salinity and lower rainfall and the nutrient concentrations are lower than at Noia (Román et al., 2024). Combarro is located at the outflow of the Mouro creek, which drained a smaller watershed of 11 km2. The mean annual temperature in NW Spain is 12.5-15 °C, and the annual precipitation ranges between 1400–2000 mm year-1 (AEMET and IM, 2011).
The baskets were covered with a plastic mesh of 2.8 cm diagonal opening, to prevent access by large predators. In the baskets containing Z. noltei, a square area of 10 x 10 cm was marked on the mesh with cable ties, and the seagrass leaves within this area were cut above the sheath. New tissue that grew in these squares was collected after the transplantation period for biochemical analysis to determine the nutrient uptake and physiological processes that occurred exclusively during the transplant period.
Half of the baskets (54) were transported to each site. The baskets without seagrass were placed in areas of bare sediment, whereas those with seagrass were placed within Z. noltei patches. All baskets were distributed at random in each habitat and buried at the same level as the surrounding sediment. The baskets were left for two and a half months in the field and retrieved on 29 August from Noia and on 30 August from Combarro. On the same day that the baskets were retrieved, the clipped plants in the 10 x 10 cm squares were removed and stored separately from the rest of the seagrass, before the clams were removed. The sediment was sieved in situ through a 5-mm mesh to prevent collection of clam recruits, and the clams and seagrass samples were stored separately. At Noia, the seagrass meadow where the baskets were placed was found to be covered by Ulva spp. canopies; the above-ground seagrass biomass in the baskets was scarce and damaged. All samples were placed in zip lock plastic bags and transported to the laboratory where they were frozen at -20°C.
The temperature in bare sediment outside the baskets at each site was recorded every 60 minutes during the transplant period with ibutton® dataloggers placed at depths of 3, 8 and 13 cm on two acrylic sticks buried vertically in the sediment. The weekly concentrations of nitrate, nitrite, ammonium and phosphate in surface waters (0–5 m depth) during the transplant were extracted from data published by the Technological Institute for the Marine Control in Galicia (Intecmar; http://www.intecmar.gal/PDFs/EvolFito/Nutrientes/Galego/Nutrientes_2022.pdf) by using the WebPlotDigitizer (https://automeris.io/). The irradiation at the sites during the field transplantation was recorded at the Marin port weather station (3 km from Combarro) and the Lesende weather station (~5 km from Noia) (https://www.meteogalicia.gal/web/observacion/rede-meteoroloxica/historico).
2.3 Zostera noltei metrics
In the laboratory, the total number of shoots in each basket was counted, including clipped and non-clipped plants, and the above-ground biomass and below-ground biomass were weighed (dry weight, 60°C, 48 h). The clipped plants contained in the 10 x 10 cm squares in each basket, comprising shoots with the new leaves and the below-ground parts, were subsampled for determination of C:N and carbohydrate content. In each basket, five replicates of both above- and below-ground tissues were separated. Each replicate consisted of 15 shoots (approximately 20 mg dw). Replicates were dried (60°C, 48h) and ground to a fine powder for analysis of the C and N contents, in a Fisons Carlo Erba EA1108 elemental microanalyzer, in the analytical facilities at the University of Vigo (CACTI-UVIGO). The first two internodes of the apical rhizomes were separated, dried (60°C, 48h) and ground to produce approximately 50 mg of sample, which was used to determine carbohydrate contents (starch and sucrose). The starch and sucrose analyses were based on resorcinol and anthrone assays, respectively, both standardized to sucrose (Olivé et al., 2007). The carbohydrate contents of the clipped plants from Noia could not be determined due to insufficient sample weights.
2.4 Survival, growth and condition index of clams
The final number of live clams (i.e. with valves closed during collection and freezing) in the transplant baskets was counted. To quantify shell growth in the retrieved 1,612 clams, the valve periostracum was removed by immersion in a solution of H2O2 (35 %) for 24 hours, and observed under a microscope (Leica MZ125) with a fluorescence filter (Leica SFL 100) at an excitation wavelength of 470 nm. The distance between the calcein mark and the shell edge was measured in triplicate in each shell with the Leica Application Suite V4 image analysis software. The dry weight of the flesh and shell of all the clams collected was quantified after drying each part separately in an oven (60°C, 48 h). The condition index (CI) was calculated using Equation 1 (Walne and Mann, 1975).
2.5 Data calculation
Thermocouple readings in each basket during the mesocosm heatwave revealed that the target temperatures were not reached in some baskets in each treatment (Supplementary Figure 2). Thus, to capture the thermal conditions experienced by bivalves and seagrass more precisely, cumulative heating was calculated for each basket during both the heatwave (HW) treatment and the field transplantation phase. The average burrowing depths of each species were calculated in bare sediment and seagrass. The sediment temperatures were then corrected by the burrowing depths, and cumulative heating (legacy degree hours and degree days) was computed (Supplementary Text 1). Hourly salinity and water levels at Noia and Combarro during the transplantation period were obtained from MeteoGalicia MOHID Water Modelling System Ocean forecasts (Supplementary Text 2).
To determine the presence of legacy effects, clam responses must be assessed long after the stressor has ceased. An additional short-term experiment was conducted simultaneously in the same tanks, in which the same treatments were applied to another batch of baskets (n= 72, 6 per tank), for evaluation of the short-term (4 days) response of clams and Z. noltei to heatwave conditions (Román et al., 2025). Clam responses measured after the short-term experiment were used to estimate responses attributable exclusively to the field transplantation phase. Clam mortality during field transplantation was calculated by adjusting the final mortality observed at the end of the experiment, based on clam abundance after the mesocosm heatwave (HW) treatment (Supplementary Text 3). To assess the legacy effects of the mesocosm heatwave (HW), shell growth attributable to the field transplantation phase was estimated by subtracting post-HW mesocosm growth from the final shell growth measurements (Supplementary Text 4).
2.6 Statistical analysis
Environmental conditions were compared between the two study sites. The 90th percentiles of daily temperatures at 3cm depth (emersion temperatures) in bare sediment and in sediment below Z. noltei were calculated. The mean between-site differences in the 90th percentiles of daily temperatures, night-time averages of daily water temperature, hourly salinity at high tide and weekly nutrient concentrations were determined by paired t-tests.
As the target temperatures of 20, 27, and 32°C were not consistently reached in all baskets (Supplementary Figure 2), cumulative heating was modelled as a covariate for each basket to more accurately represent the thermal conditions experienced by the organisms. Legacy degree hours (LDH) above 20°C were defined as the cumulative thermal exposure experienced during the mesocosm treatments. The effects on Z. noltei of the fixed factors Habitat (Z. noltei and Z. noltei + clams), Site (Noia and Combarro), the covariate LDH, and their interactions, were tested by using linear models, with the lm function.
The effects on clams of the fixed factors Habitat (bare sediment and Z. noltei), Site, the covariate LDH, their interactions, and the random factor Basket (for growth and Condition index) were tested. Clam mortality, a binary response (dead/alive), was tested with generalized linear models by using a binomial distribution of errors and a “logit” (logistic cumulative distribution) function that allowed the probability of death to be modelled as a function of predictors; the models were implemented with the “glm” function in the base R package “stats” (R Core Team, 2023). Shell growth and condition index, both continuous variables, were analyzed using linear mixed-effects models to account for repeated measures within baskets. The models were implemented using the ‘lme’ function in the ‘nlme’ package (Pinheiro et al., 2022). Each clam species was analyzed separately, given that the responses of each species were not assumed to be independent. Species-specific differences were expected due to variations in burrowing depth (Macho et al., 2016; Domínguez et al., 2021) caused by their different siphon lengths (Gosling, 2015).
Prior to model application, extreme outliers for each response variable (except clam survival) were removed using a modified version of the method proposed by Mittaz and Harris (2011), to increase the reliability of the parametric statistical tests (Crawley, 2013). Extreme outliers were defined as values below or above the median ± 4 times the standard deviation, calculated as IQR/1.34. Factor 1.34 corresponds to the relationship between the interquartile range (IQR) and the standard deviation in a normal distribution. The outliers removed accounted for less than 10 % of the observations (Supplementary Table 2).
The global models were simplified through an information-theoretic selection approach. The corrected Akaike information criterion index (AICc) was calculated for the models with and without the random factor Basket, and those with the lowest AICc were retained (Supplementary Table 3). Subsets of the fixed part of the models with different link functions were generated in a last-wise order, and those with the lowest AICc and that did not have any other simpler model nested within were finally selected (Richards et al., 2011). AICc and model simplification were calculated with the AICc and dredge functions in the MumIn package (Bartoń, 2025) (Supplementary Tables 4; Supplementary Tables 5).
The normality of residuals and homogeneity of the variances of the models were checked with QQ plots and plots of standardized residuals vs. fitted values, respectively. If residuals were not homogeneously distributed and indicated heteroscedasticity, the models were adjusted by specifying different variances per factor level using the VarIdent function in the nlme package (Pinheiro et al., 2022). This adjustment typically improved the homogeneity and distribution of the residuals. The significant (p-value < 0.05) terms of the final models were obtained by computing a type 3 sum of squares analysis, with the Anova function in the car package (Fox and Weisberg, 2019). If more than two levels were present for the significant terms, post-hoc pairwise comparison of the means was performed with the emmeans function (Lenth, 2024). Tukey correction was applied to the p-values to reduce comparison-wise error rates to levels that constrain experiment-wise error rates to standard values (Underwood, 1997). All data processing and statistical tests were conducted with R v 4.3.2 (R Core Team, 2024). The significance threshold was established as p < 0.05. Environmental variables were expressed as mean ± SD and biological variables as mean ± SE.
3 Results
3.1 Temperature, salinity and nutrients at the transplant sites
The thermal amplitude was higher in the bare sediment than in sediment with Z. noltei in the baskets at both sites. The temperature during daytime low tides was lower in the sediment with Z. noltei, whereas the temperature during night-time low tides was lower in the bare sediment (Figure 2A). The number of degree days (DDs) experienced by clams was higher at Combarro (bare sediment = 141-151, seagrass =135-142) than at Noia (bare sediment = 43-50, seagrass = 40-44). In addition, within each site, the number of DDs was higher in bare sediment than in vegetated sediment, and within each habitat, R. decussatus experienced the lowest cumulative heating consistent with its greater burrowing depth (Figure 2B; Table 1).
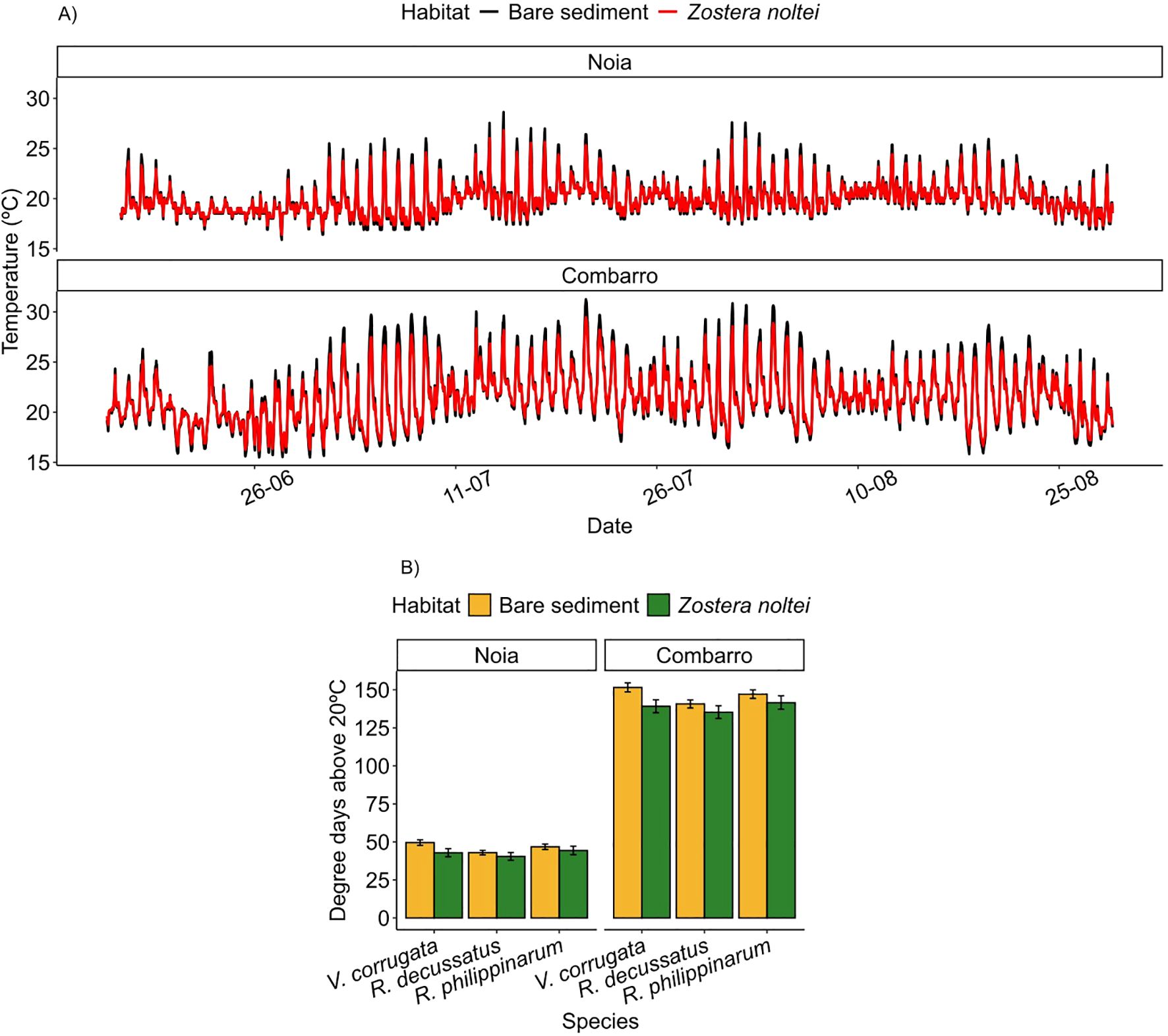
Figure 2. Sediment in baskets. (A) Combarro and Noia sediment temperatures at 3 cm depth during the transplantation period, measured in bare sediment and calculated for Z. noltei from water temperature, damping depths and bare sediment temperatures. (B) Field degree days above 20°C experienced by each clam species, calculated based on the different burrowing depths in vegetated and unvegetated sediment (Mean ± 95 % confidence intervals).

Table 1. Environmental variables during the field transplantation period (14/06/2022 – 30/08/2022).
The 90th percentile of sediment temperature at 3 cm depth (Table 1), the night-time averages of seawater temperature (°C) (Combarro = 20.14 ± 1.18, Noia = 18.82 ± 0.70) and hourly high tide salinities (Combarro = 34.41 ± 1.09, Noia = 30.28 ± 4.66) were higher at Combarro than at Noia (Figure 3; Table 1). The salinity of Noia presented greater oscillations than Combarro due to its location closer to a river outfall, that generated salinity drops due to frequent freshwater inputs. The concentrations (µmol L-1) of nitrate and nitrite in surface seawater were significantly higher (p < 0.05) at Noia (NO3- = 6.30 ± 3.43, NO2- = 0.26 ± 0.10) than at Combarro (NO3- = 0.72 ± 0.86, NO2- = 0.13 ± 0.09), whereas phosphate and ammonium concentrations did not differ significantly between the sites (Figure 3; Table 1). The mean daily irradiation in the field (W m-2) was 258 ± 73 at Noia (Lesende weather station) and 258 ± 66 at Combarro (Marín port weather station) (n=77).
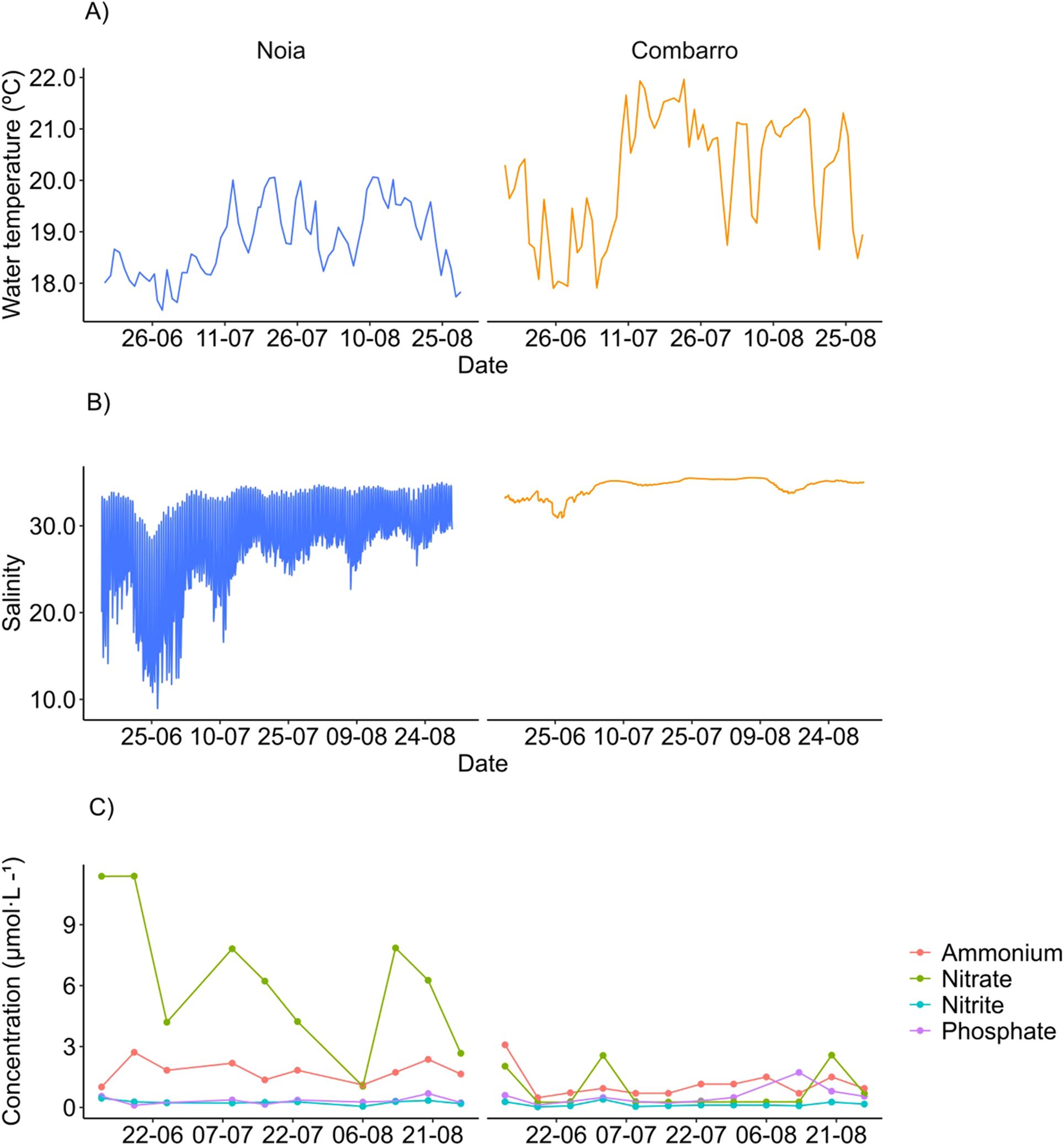
Figure 3. Surface water. Environmental variables during the transplant experiment. (A) Daily night-time average water temperature estimated from measurements made in the shellfishing bed, (B) Hourly salinity during high tide, retrieved from the MeteoGalicia MOHID model, (C) Weekly nutrient concentrations (0–5 m depth), retrieved from the Intecmar annual reports, recorded at coastal stations P0 (Combarro) and M7 (Noia).
3.2 Morphological and biochemical metrics of Zostera noltei
During retrieval of the baskets, the seagrass patches at the Noia intertidal flat were found to be covered by continuous and extensive canopies of Ulva spp., and Ulva fronds were also trapped by the basket mesh. The above-ground biomass of Z. noltei was sparse and visibly degraded at this site. At Combarro, the seagrass was in an overall better condition and was not covered by macroalgae.
At both sites, a significant Habitat x Legacy Degree Hours (LDH) interaction was observed for above-ground biomass and number of shoots. In plants subjected to higher heating in the mesocosm, above-ground biomass and number of shoots decreased in Z. noltei alone and were constant in Z. noltei with clams (Figure 4; Table 2). The significant Habitat x Site interaction for number of shoots and above-ground biomass (g dw) indicated that both variables were greater when co-existing with clams than when growing alone at Combarro (AGB: seagrass + clams = 0.72 ± 0.10 vs. seagrass alone = 0.51 ± 0.09; shoots: seagrass + clams = 91.9 ± 12.5 vs. seagrass alone = 38.4 ± 11.8), but not at Noia. The below-ground biomass (g) decreased at both sites as LDH increased. Overall, seagrass biomass (g dw) was greater at Combarro (1.39 ± 0.135) than at Noia (0.65 ± 0.13) (Figure 4; Table 2).
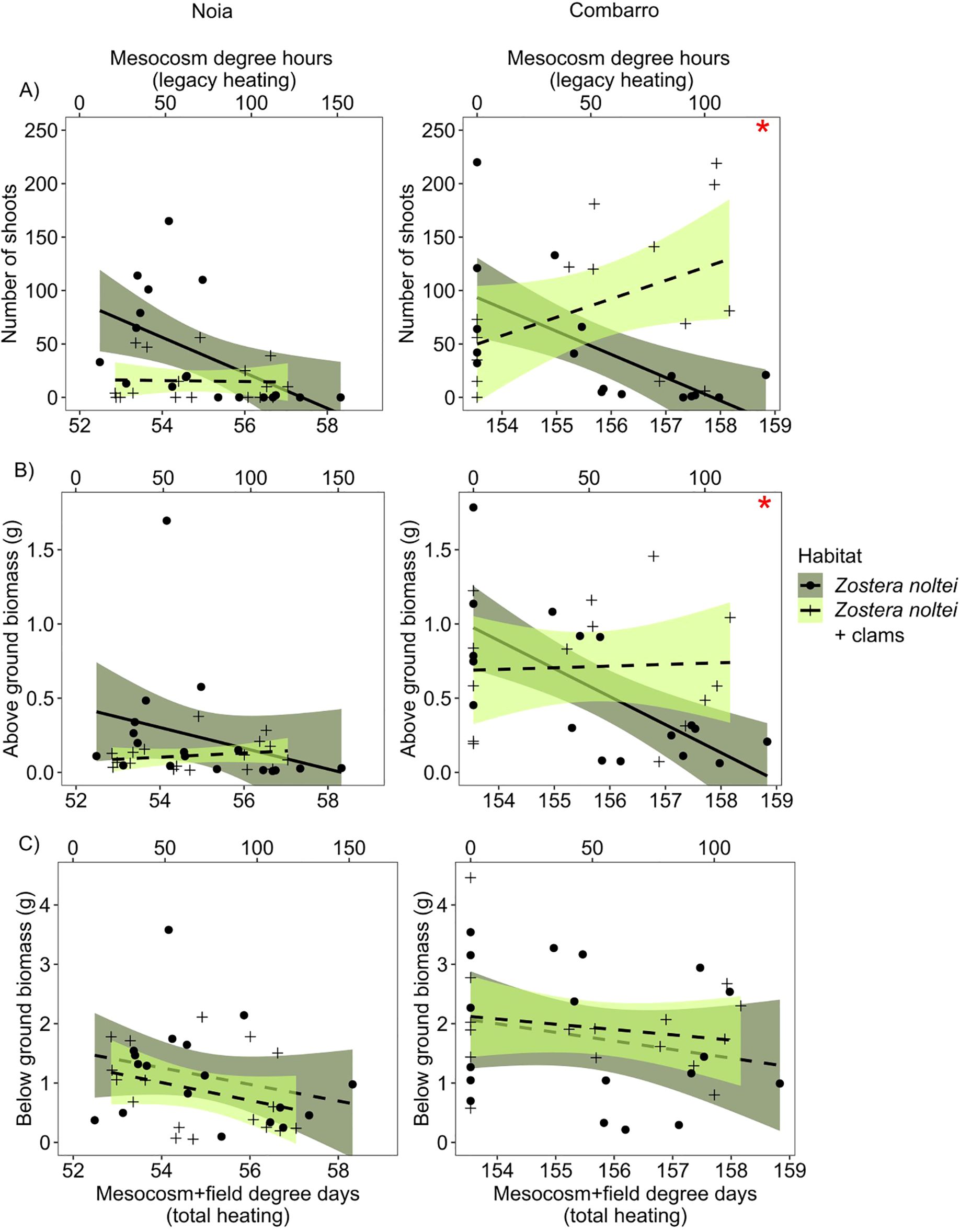
Figure 4. Zostera noltei. Responses of morphological variables measured in the baskets in each transplant site to increasing cumulative heating (above 20°C), in sediment with and without clams. Upper x axis indicates legacy heating after mesocosm treatments and before transplantation (legacy degree hours); the bottom x axis indicates total cumulative heating (total degree days). (A) Number of shoots. (B, C) Above- and below ground biomass. Points/crosses: Each observation per basket; lines: data fitted by linear models; shaded areas: 95% confidence intervals. Dashed and continuous lines represent non-significant and significant effects, respectively, of past cumulative heating. Red asterisks in the upper right side of the panels indicate the positive effects of clams at Combarro.
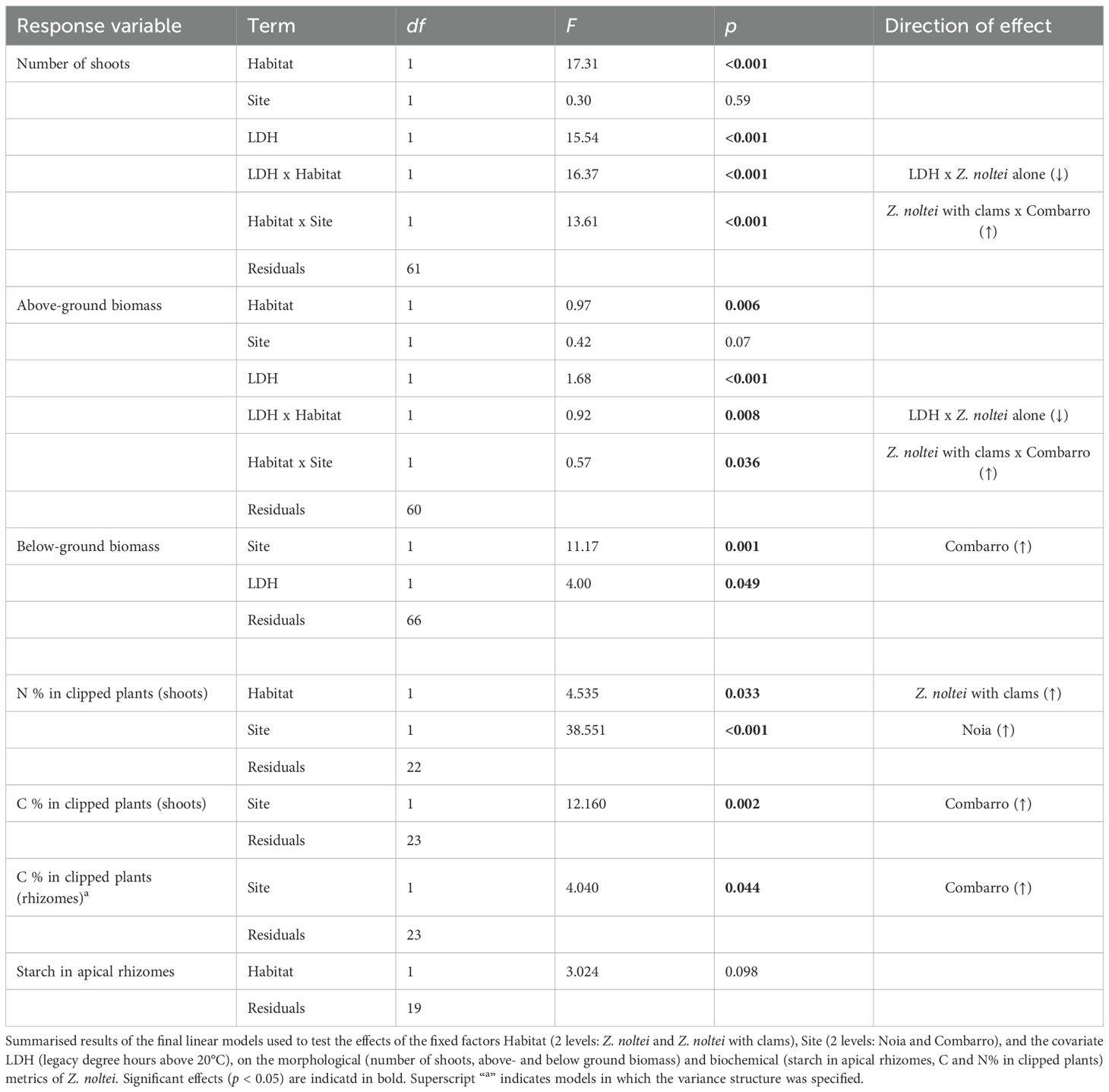
Table 2. Zostera noltei.
The % N in shoots of clipped plants was greater in the seagrass growing together with clams than in seagrass growing alone, at both sites (Figure 5). Independently of habitat, % N in shoots was greater at Noia (4.03 ± 0.11) than at Combarro (3.16 ± 0.09). By contrast, the % of C in shoots and in rhizomes of clipped plants was greater at Combarro than at Noia (Figure 5; Table 2). The concentrations of sucrose or starch in apical rhizomes at the Combarro site were not affected by the experimental treatments (Supplementary Figure 4; Table 2; Supplementary Table 4).
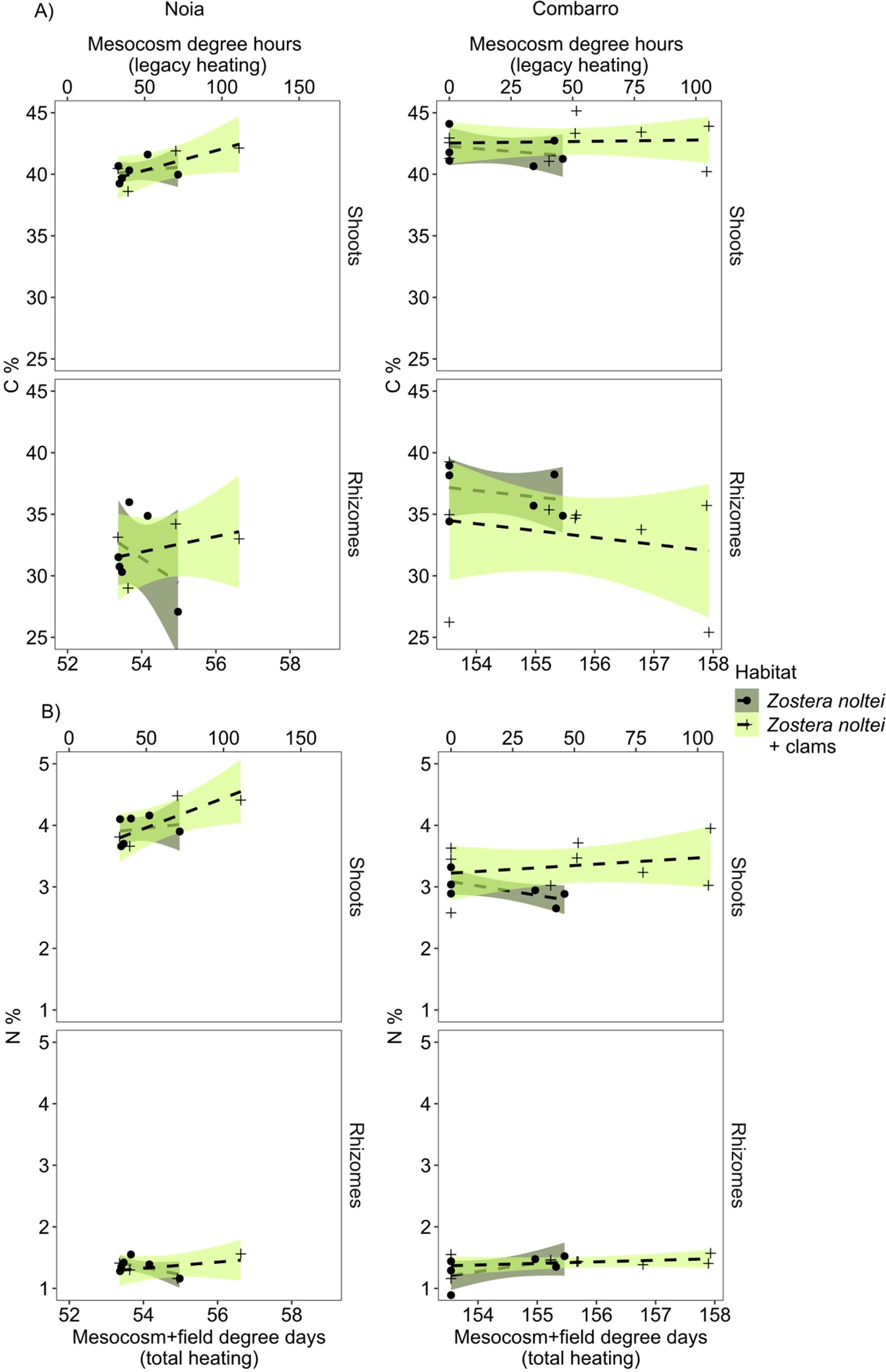
Figure 5. Zostera noltei. (A) Carbon content and (B) nitrogen content in the shoots and rhizomes of clipped plants (in 10 x 10 cm squares within each basket) and in transplanted plants in the Noia and Combarro sites after being exposed to increasing cumulative heating (above 20°C) in sediment with and without clams. Upper x axis indicates legacy heating after mesocosm treatmens and before transplant (legacy degree hours), bottom x axis indicates total cumulative heating (total degree days). Points: Mean values of the replicates per basket; lines: data fitted by linear models; shaded areas: 95% confidence intervals. Dashed lines represent non-significant effects of past cumulative heating.
3.3 Mortality, growth and CI of clams
Clam survival was proportional to total number of DD of exposure in all three species (Figure 6), and mortality was highest at Combarro. The effects were consistent across different habitats, with stronger impacts observed in V. corrugata and R. decussatus, as indicated by the steeper negative slopes.
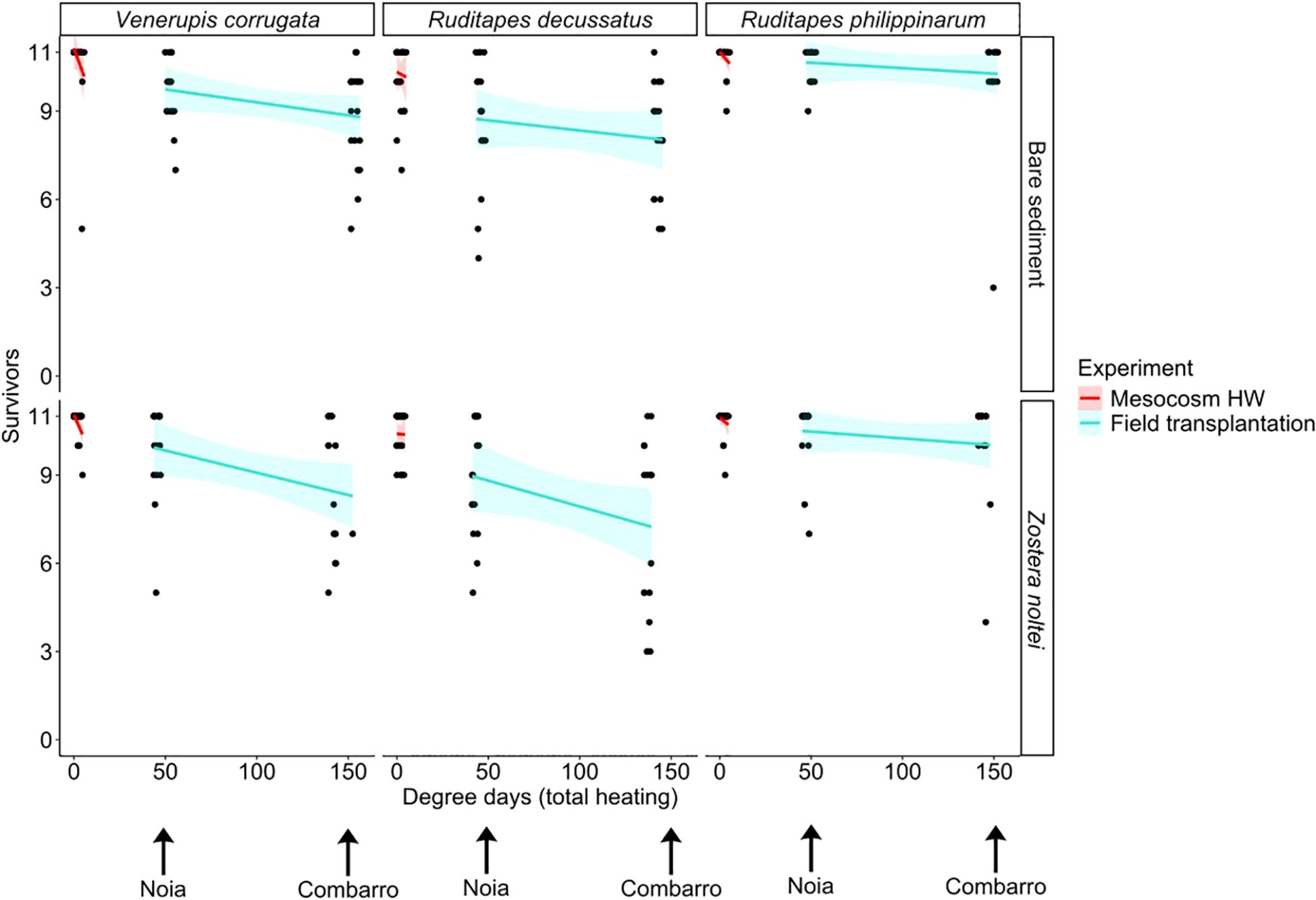
Figure 6. Number of surviving clams per basket as a function of total degree day exposure (above 20°C) after the mesocosm heatwave and the field transplantation. Points around 50 DD correspond to Noia site and those around 150 DD to Combarro. Lines: data fitted by binomial generalized linear models; shaded areas: 95% confidence intervals. Continuous lines represent significant effects of total heating (implicit in the site factor).
Shell growth (mm) in all three bivalve species was greater at Noia than at Combarro (Figure 7).The significant Habitat x Site interaction indicated that growth was greater in sediment with Z. noltei than in bare sediment at Combarro (V. corrugata: seagrass = 1.50 ± 0.13 vs. bare sediment = 0.73 ± 0.11; R. decussatus: seagrass = 2.57 ± 0.15 vs. bare sediment = 1.91 ± 0.11; R. philippinarum: seagrass = 4.87 ± 0.18 vs. bare sediment = 4.34 ± 0.17), and this pattern was not observed at Noia (Figure 7; Table 3). The effect of LDH x Site interaction on shell growth of V. corrugata and R. decussatus indicated that V. corrugata grew less at Noia when exposed to greater LDH, whereas R. decussatus at Combarro grew more after exposure to greater LDH (Figure 7; Table 3).
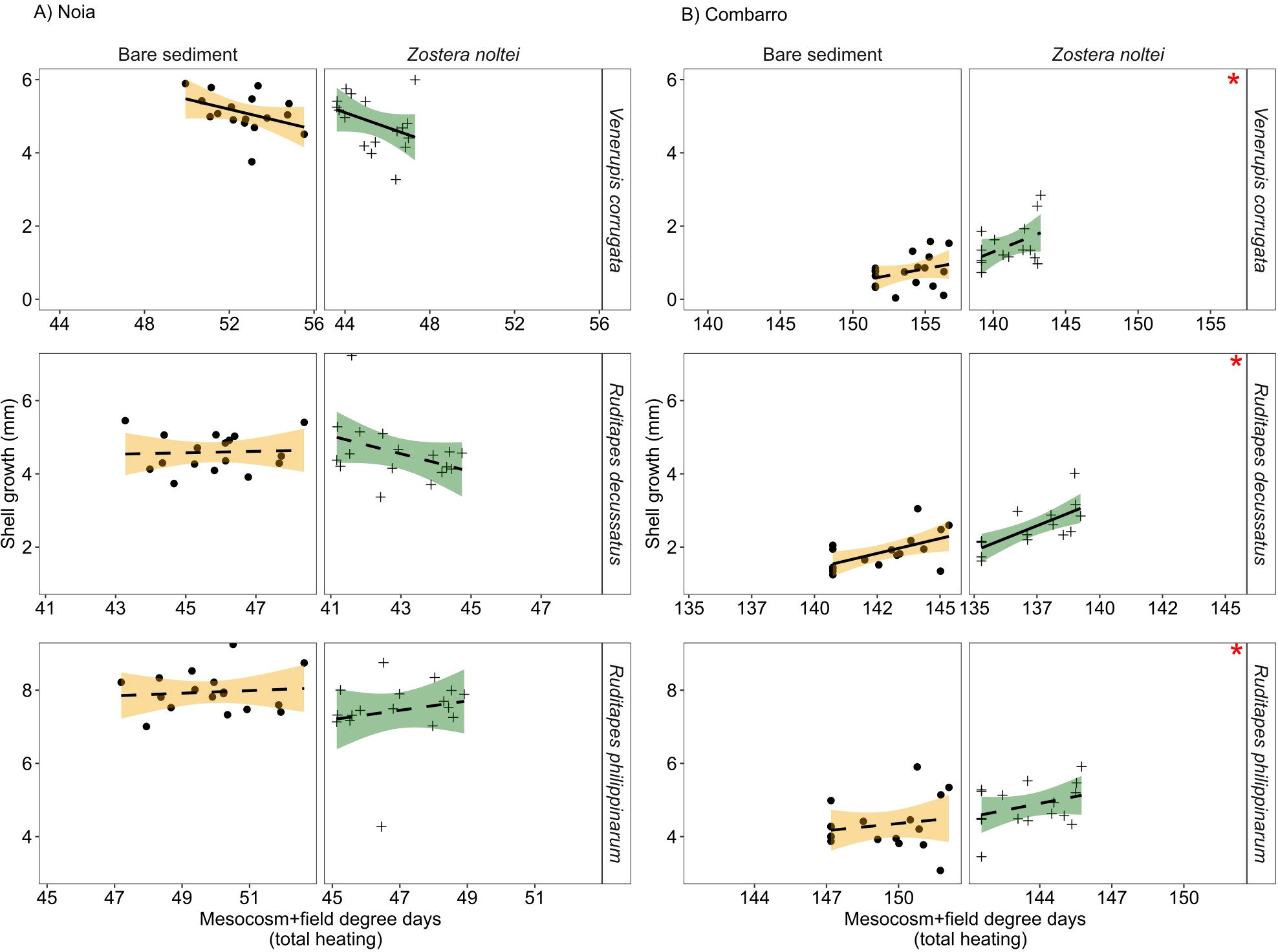
Figure 7. Shell growth in clams in bare sediment and in sediment with Zostera noltei at (A) Noia and (B) Combarro transplant sites, after being exposed to cumulative heating during low tide in the mesocosm and in field transplantation (total degree days above 20°C). Points/crosses: mean values per basket; lines: data fitted by linear models; shaded areas: 95% confidence intervals. Dashed and continuous lines represent non-significant and significant effects, respectively, of past cumulative heating. Red asterisks in the upper right side of the panels indicate the positive effects of Z. noltei at Combarro.
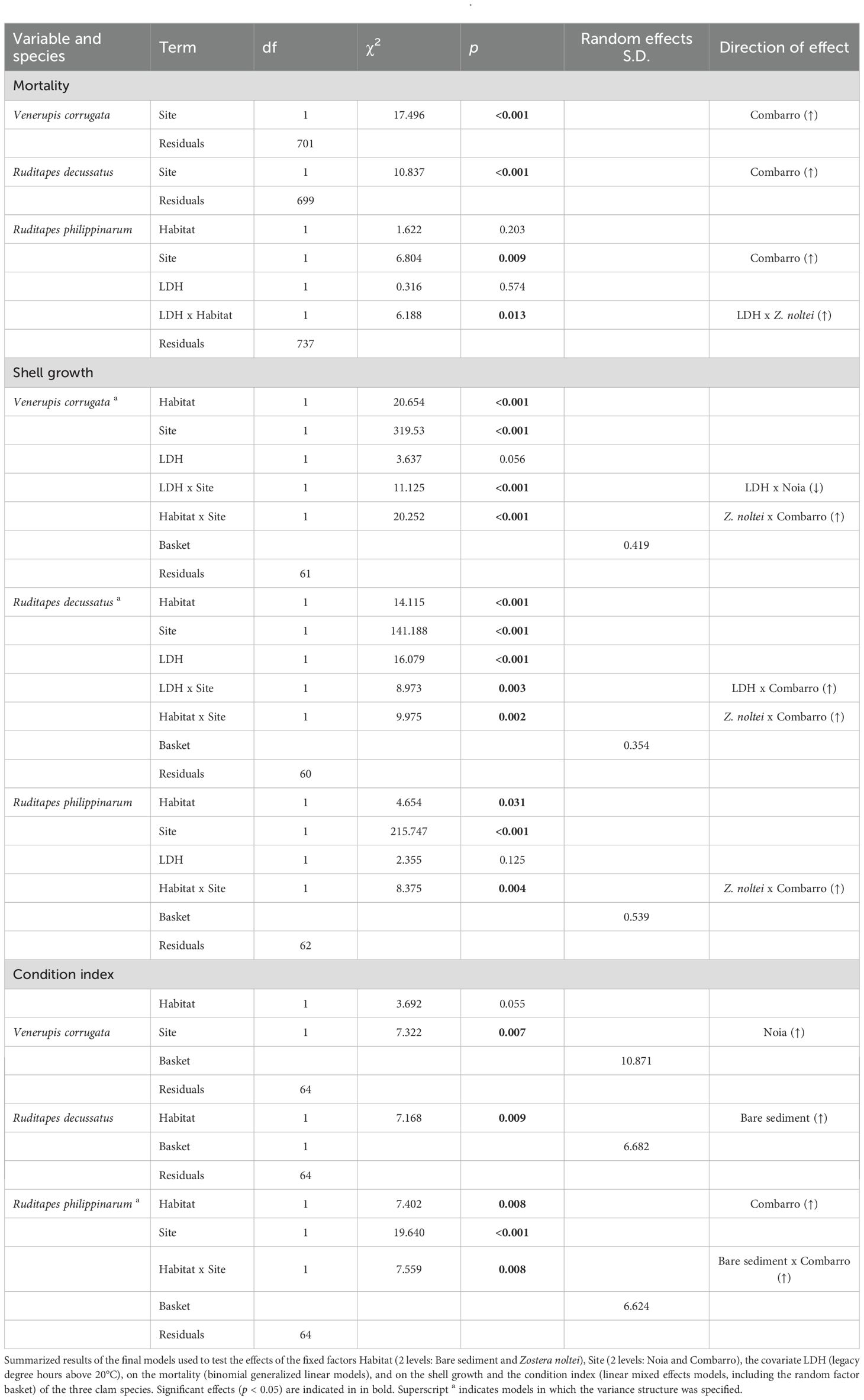
Table 3. Venerupis corrugata, Ruditapes decussatus and Ruditapes philippinarum.
The Condition Index (CI, ‰) of V. corrugata was significantly higher at Noia (138 ± 2) than at Combarro (129 ± 2). The CI of R. decussatus was greater in individuals placed in bare sediment than those placed in sediment with Z. noltei at both sites, whereas the CI of R. philippinarum only showed this pattern at Combarro (Figure 8; Table 3).
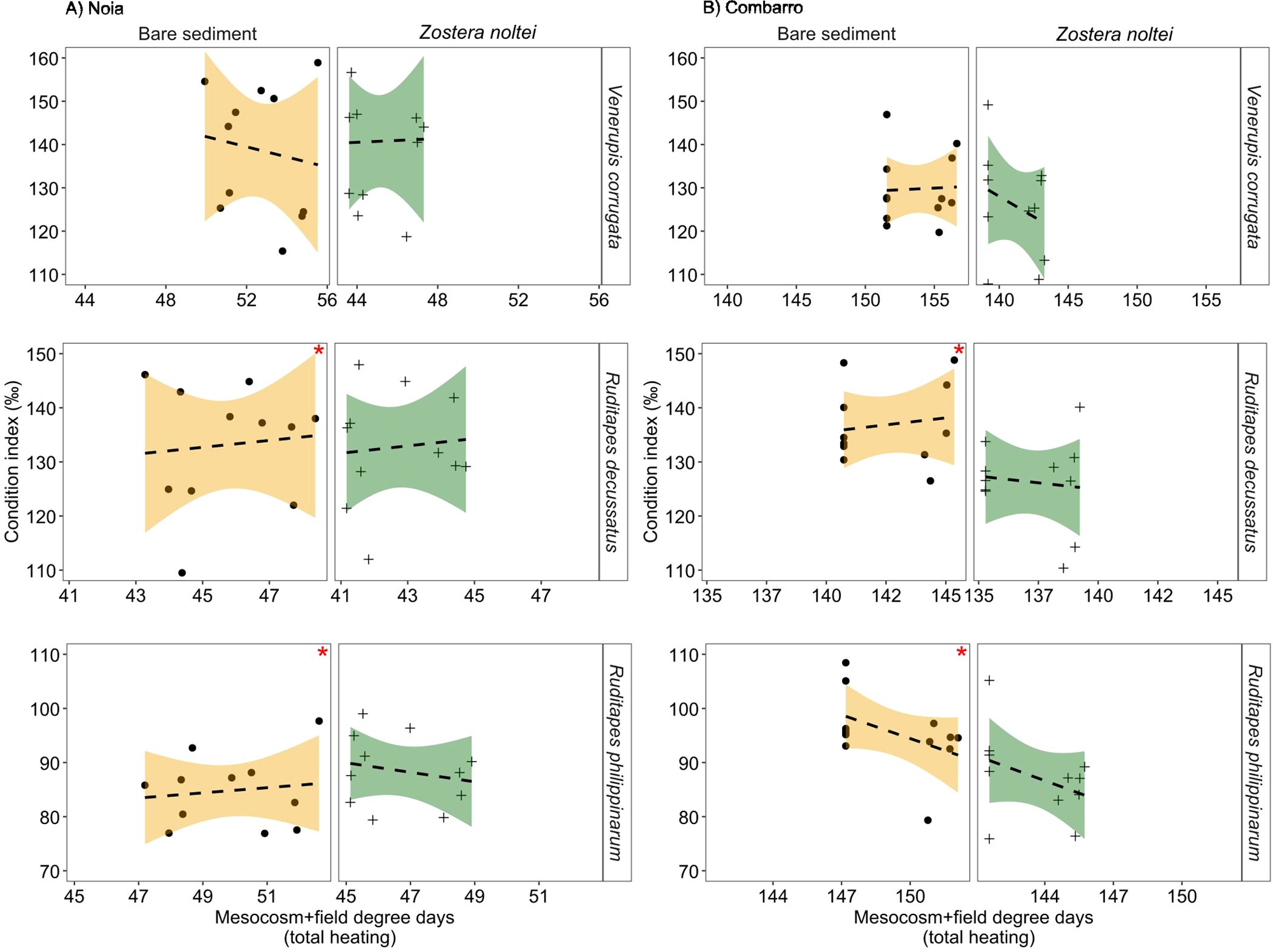
Figure 8. Condition index of clams in bare sediment and in sediment with Zostera noltei at (A) Noia and (B) Combarro transplant sites after being exposed to cumulative heating during low tide in the mesocosm and in field transplantation (total degree days above 20°C). Points: mean values per basket; lines: data fitted by linear models; shaded areas: 95% confidence intervals. Dashed lines represent non-significant effects of past cumulative heating. Red asterisks in the upper right side of the panels indicate the positive effects of bare sediment.
4 Discussion
4.1 Effects of clam presence on the recovery of Zostera noltei after heat stress
The presence of clams was associated with a mitigation of the negative effects of past low-tide thermal stress on Zostera noltei, as indicated by the significant Habitat × LDH interaction (Table 2; Figure 4), thereby supporting the first hypothesis of the study. This may be because clams enhance sediment fertilization and reworking, which favor irrigation and transport of solutes (Volkenborn et al., 2012) into and from the rhizosphere (Kristensen et al., 2012), promoting nutrient absorption by below-ground biomass (Hemminga, 1998). Moreover, filter feeding bivalves can reduce the epiphytic load of seagrass (Peterson and Heck, 2001) and improve light penetration, thereby promoting seagrass growth (Wall et al., 2008 and references therein). This clam-derived facilitation of seagrass observed after harsh environmental conditions is consistent with the stress gradient hypothesis (Bertness and Callaway, 1994), which states that positive interactions are enhanced under stressful conditions and aligns with studies on clam–seagrass interactions that support this theory (Chin et al., 2021; Cardini et al., 2022; Román et al., 2024).
Independently of legacy heating, the significant Habitat x Site interaction in the seagrass models showed that the shoot abundance and the above-ground biomass of Z. noltei was greater in plants growing together with clams at Combarro, characterized by lower nutrient availability and higher temperatures, (Table 2; Figure 4). Similar enhancements of seagrass growth by neighboring bivalves has been reported, including larger leaves of T. testudinum growing together with Modiolus americanus (Leach, 1815) (Peterson and Heck, 2001), increased above-ground biomass of Z. noltei growing with Loripes orbiculatus (Poli, 1795) (De Fouw et al., 2022) and greater patch size and plant biomass in seed sowing restoration (Zhang et al., 2021). No such facilitation was observed at the Noia site, where the seagrass was degraded and covered by Ulva spp. We suggest that growth of the ephemeral opportunistic macroalgae may have hampered growth of the seagrass canopy by reducing light penetration (Brun et al., 2003; Lee et al., 2007 and references therein). Moreover, the higher nutrient availability at Noia may have made the seagrass less reliant on clams for input of nutrients for growth.
The significant effect of Habitat indicated that the % N in shoots of clipped plants was higher in seagrass growing together with clams. This observed facilitation by clams on seagrass may be related to the acquisition of limiting nutrients from bivalve excretions (Bronstein, 2009). The ammonium excreted by clams increases N input into sediment and water and can be assimilated by below-ground biomass (Touchette and Burkholder, 2000; Peterson and Heck, 2001; Meysick et al., 2020).
4.2 Effects of the presence of Zostera noltei on clam recovery after heat stress
The absence of LDH effects in the mortality models indicated that the mesocosm heatwave did not have a strong legacy effect on clam mortality (Table 3; Supplementary Table 5; Supplementary Figure 5). The significant effect of Site in the mortality models suggested that the increased clam mortality at Combarro may be related to higher field temperatures, possibly in combination with other site-specific factors (Figure 6). Additionally, the significant Habitat x Site interaction in the clam growth models indicated that the presence of seagrass enhanced growth only at Combarro, the hottest site. Under higher temperatures, which are more stressful for clams (Rato et al., 2022), the presence of seagrass had a positive effect on clam growth, irrespective of legacy heat stress (Table 3; Figure 7). The seagrass presence did not have a positive effect on clam growth at Noia. We hypothesize that the degradation of seagrass was partly due to the small and degraded patches at Noia, which likely hampered the facilitation of clam growth. Moreover, the milder temperatures at Noia made the clams less reliant on facilitative interactions to maintain survival and growth. Thus, the second hypothesis was not supported, as the presence of seagrass did not ameliorate negative effects of legacy low-tide heatwave stress on juvenile clams.
The most plausible mechanism explaining the enhanced growth of clams in vegetated sediment compared to bare sediment in Combarro is seagrass facilitation. Specifically, clams likely expended less energy maintaining homeostasis, primarily through body temperature regulation, when shaded by Zostera noltei (García-Souto et al., 2024). The role of seagrass as a thermal buffer was demonstrated by the lower cumulative DD and mean temperatures in vegetated sediment than in bare sediment at both sites. The seagrass-vegetated sediments are frequently finer and have lower porosity than unvegetated sediments, which enhances water retention (Maxwell et al., 2017; Miyajima and Hamaguchi, 2019). This slowed heat diffusion, leading to a shallower damping depth, and made the vegetated sediment much harder to heat, and thus the sediment below Z. noltei was cooler than bare sediment after the low tides.
4.3 Effects of local abiotic conditions on interspecific interactions
There was a significant effect of Site (alone or in interaction with the factors LDH or Habitat) in all models tested. Thus, the third hypothesis was supported, as the environmental context affected specific responses and interactions differently. At Noia, under conditions of lower temperatures and higher nutrient availability, clam survival and growth were favored, but not seagrass growth (Figures 4, 6, 7). We observed that greater availability of nutrients hampered seagrass growth, negatively affecting the facilitation effect on clams.
At Noia, the highest nutrient concentrations in seawater, the proliferation of ephemeral macroalgae above the seagrass, the lower biomass and the higher % N in shoots suggested early eutrophic conditions (Burkholder et al., 2007). The Noia estuary may be subjected to fluvial inputs of land-derived organic matter enriched with anthropogenic nitrogen (Roca et al., 2016). High N availability in the water column promotes growth of phytoplankton, epiphytes and macroalgae, which reduce light penetration and restrain growth of phanerogams (Lee et al., 2007; Smetacek and Zingone, 2013).
By contrast, at Combarro C fixation and growth of seagrass could be favored by the absence of shading by ephemeral macroalgae canopies (Hemminga, 1998), although there was a greater cumulative heating effect. High water and sediment temperatures inhibit photosynthetic performance in Z. noltei (Massa et al., 2009; Román et al., 2023), what can reduce the shoot density and length of Zostera spp (Berger et al., 2024). Thus, harsher environmental conditions at Combarro may have made Z. noltei more reliant on interactions with clams to sustain shoot biomass and density.
Several studies in both terrestrial and marine ecosystems have shown that environmental conditions can shift interactions from mutualism to antagonism (Thompson, 1988; Cheney and Côté 2005) and that mutualistic interactions are more frequent and/or stronger under adverse environmental conditions (Bertness and Callaway, 1994). Similar patterns have been observed in clam–seagrass interactions (Meysick et al., 2020). Nonetheless, the present findings suggest that facilitation strengthening only occurs when the growth and survival of the facilitator species are not critically affected by abiotic stress. This is consistent with a reported breakdown of clam–seagrass mutualism following seagrass degradation after a drought event (De Fouw et al., 2016).
4.4 Legacy effects of the experimental heatwave
Overall, the lack of effect of LDH in the models revealed that the mesocosm heatwave did not have a legacy effect on clam mortality or condition index. Clam growth did not respond overall to legacy heat stress, and the observed trends depended on the species and habitat. By contrast, elevated temperatures in the field had a greater impact on clams than the heatwave simulated in the mesocosm. The significant Site effect indicated that mortality was highest at Combarro, where cumulative heating (around 150 degree days) was much higher than at Noia (around 50 degree days), and in the mesocosm (around 6 degree days).
However, the mesocosm heatwave had a measurable impact on seagrass, which was buffered by the interaction with clams. After the mesocosm heatwave, biomass and shoot density was negatively affected only in plants without clams, suggesting that the clams exerted a facilitation effect under high temperatures (Clemente and Thomsen, 2023). The decreased below-ground biomass with increasing legacy degree hours (LDH) indicated an incomplete recovery after two and a half months. Accordingly, previous research has shown legacy impacts of heat stress on intertidal macrophytes. A decrease in the biomass and growth of Z. marina was noted 5 weeks after exposure to moderate warming (Reynolds et al., 2016), and the structure of macroalgae assemblages changed 6 months after exposure to high temperatures (Vaz-Pinto et al., 2013).
5 Conclusions
The above-ground biomass and shoot abundance measured in Z. noltei growing together with clams were less affected by legacy effects of a low-tide heatwave than those measured in seagrass without clams. This was consistent with the stress gradient hypothesis and implied that facilitation not only occurs during environmental stress, but also after the stress ceases. By contrast, the interaction between legacy heatwave stress and habitat on the growth and condition index of clams proved to be species- and site-specific. Local environmental conditions at transplant sites led to contrasting responses in the growth and survival of seagrass and clams and in their interactions. The findings suggest that facultative mutualism between the temperate seagrass Z. noltei and the three commercial clam species was strengthened under higher temperatures and weakened under eutrophic conditions. This study provides evidence of the complex relationship between local abiotic factors and biological interactions that shape the structure and organization of intertidal communities exposed to climatic stressors.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
MR: Formal analysis, Investigation, Visualization, Writing – original draft, Writing – review & editing. EV: Conceptualization, Funding acquisition, Investigation, Project administration, Resources, Supervision, Writing – review & editing. NW: Investigation, Methodology, Writing – review & editing. RV: Formal analysis, Investigation, Methodology, Visualization, Writing – review & editing. SR: Investigation, Methodology, Writing – review & editing. JT: Investigation, Methodology, Writing – review & editing. DW: Conceptualization, Formal analysis, Investigation, Methodology, Visualization, Writing – review & editing. SW: Conceptualization, Formal analysis, Investigation, Methodology, Visualization, Writing – review & editing. CO: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Visualization, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was supported by the Spanish Ministerio de Ciencia e Innovación/Agencia Estatal de Investigación (10.13039/501100011033) through the grants RTI2018-095583-B-I00 and TED2021-129524B-I00 to the projects ZEUS and RECOBI, respectively. Salvador Román was supported by a PhD fellowship from the Xunta de Galicia (ED481A-2020/199). Marta Román was supported by a Post-doc fellowship from the Xunta de Galicia (ED481B-2023-006).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1706079/full#supplementary-material
References
AEMET (2024). Olas de calor en España desde 1975. Available online at: https://www.aemet.es/es/conocermas/recursos_en_linea/publicaciones_y_estudios/estudios/detalles/olascalor (Accessed January 12, 2025).
AEMET, IM (2011). “Iberian climate atlas,” in Air temperature and precipitation, (1971-2000) (Agencia Estatal de Meteorología (España) and Instituto de Meteorologia, Portugal). doi: 10.31978/784-11-002-5
Bartoń K. (2025). “MuMIn:multi-model inference,” in MuMIn, R package version 1.48.11. doi: 10.32614/CRAN.package
Berger A. C., Berg P., McGlathery K. J., Aoki L. R., and Kerns K. (2024). Eelgrass meadow response to heat stress. II. Impacts of ocean warming and marine heatwaves measured by novel metrics. Mar. Ecol. Prog. Ser. 736, 47–62. doi: 10.3354/meps14588
Bertness M. and Callaway R. (1994). Positive interactions in communities. Trends Ecol. Evol. 9, 187–191. doi: 10.1016/0169-5347(94)90088-4
Bronstein J. L. (2009). The evolution of facilitation and mutualism. J. Ecol. 97, 1160–1170. doi: 10.1111/j.1365-2745.2009.01566.x
Brun F. G., Vergara J. J., Navarro G., Hernández I., and Pérez-Lloréns J. L. (2003). Effect of shading by Ulva rigida canopies on growth and carbon balance of the seagrass Zostera noltii. Mar. Ecol. Prog. Ser. 265, 85–96. doi: 10.3354/meps265085
Burkholder J. M., Tomasko D. A., and Touchette B. W. (2007). Seagrasses and eutrophication. J. Exp. Mar. Bio. Ecol. 350, 46–72. doi: 10.1016/j.jembe.2007.06.024
Cacabelos E., Quintas P., Troncoso J. S., Bárbara I., García V., Cremades J., et al. (2015). “Las praderas marinas de España: una visión general. La biodiversidad de las praderas españolas: Atlántico norte,” in Atlas de las praderas marinas de España. Eds. Ruiz ,. J. M., Guillén J. E., Ramos Segura A., and Otero M. M. (IEO/IEL/UICN, Murcia-Alicante-Málaga), 87–91. Available online at: https://www.ieo.es/es/atlas-praderas-marinas (Accessed January 12, 2025).
Cannon A. J. (2025). Twelve months at 1.5 °C signals earlier than expected breach of Paris Agreement threshold. Nat. Clim. Change 15, 266–269. doi: 10.1038/s41558-025-02247-8
Cardini U., Marín-Guirao L., Montilla L. M., Marzocchi U., Chiavarini S., Rimauro J., et al. (2022). Nested interactions between chemosynthetic lucinid bivalves and seagrass promote ecosystem functioning in contaminated sediments. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.918675
Carvalho D., Cardoso Pereira S., and Rocha A. (2022). Future surface temperature changes for the Iberian Peninsula according to EURO-CORDEX climate projections. Climate Dyn. 56, 123–138. doi: 10.1007/s00382-020-05472-3
Cheney K. L. and Côté I. M. (2005). Mutualism or parasitism? The variable outcome of cleaning symbioses. Biol. Lett. 1, 162–165. doi: 10.1098/rsbl.2004.0288
Chin D. W., de Fouw J., van der Heide T., Cahill B. V., Katcher K., Paul V. J., et al. (2021). Facilitation of a tropical seagrass by a chemosymbiotic bivalve increases with environmental stress. J. Ecol. 109, 204–217. doi: 10.1111/1365-2745.13462
Clemente K. J. E. and Thomsen M. S. (2023). High temperature frequently increases facilitation between aquatic foundation species: A global meta-analysis of interaction experiments between angiosperms, seaweeds and bivalves. J. Ecol. 111, 1340–1361. doi: 10.1111/1365-2745.14101
Crawley M. J. (2013). “Classical tests,” in The R book. Ed. Crawley M. J. (Wiley, United Kingdom), 344–387.
Danovaro R., Nepote E., Martire M., Carugati L., Da Ros Z., Torsani F., et al. (2020). Multiple declines and recoveries of Adriatic seagrass meadows over forty years of investigation. Mar. pollut. Bull. 161, 111804. doi: 10.1016/j.marpolbul.2020.111804
De Fouw J., Govers L. L., Van De Koppel J., Van Belzen J., Dorigo W., Sidi Cheikh M. A., et al. (2016). Drought, mutualism breakdown, and landscape-scale degradation of seagrass beds. Curr. Biol. 26, 1051–1056. doi: 10.1016/j.cub.2016.02.023
De Fouw J., Rehlmeyer K., van der Geest M., Smolders A. J. P., and van der Heide T. (2022). Increased temperature reduces the positive effect of sulfide-detoxification mutualism on Zostera noltii nutrient uptake and growth. Mar. Ecol. Prog. Ser. 692, 43–52. doi: 10.3354/meps14074
Domínguez R., Olabarria C., Woodin S. A., Wethey D. S., Peteiro L. G., Macho G., et al. (2021). Contrasting responsiveness of four ecologically and economically important bivalves to simulated heat waves. Mar. Env. Res. 164, 105229. doi: 10.1016/j.marenvres.2020.105229
ECMWF (2025). “The 2024 annual climate summary,” in Global climate highlights 2024. European Union's Earth Observation Programme. Available online at: https://climate.copernicus.eu/sites/default/files/custom-uploads/GCH-2024/GCH2024-PDF-1.pdf (Accessed January 12, 2025).
Fox J. and Weisberg S. (2019). An R companion to applied regression (Thousand Oaks, CA: Sage). Available online at: https://socialsciences.mcmaster.ca/jfox/Books/Companion/ (Accessed January 12, 2025).
Frangoudes K., Marugán-Pintos B., and Pascual-Fernández J. J. (2008). From open access to co-governance and conservation: The case of women shellfish collectors in Galicia (Spain). Mar. Pol. 32, 223–232. doi: 10.1016/j.marpol.2007.09.007
García-Souto D., Martínez-Mariño V., Morán P., Olabarria C., and Vázquez E. (2024). Hiding from heat: the transcriptomic response of two clam species is modulated by behaviour and habitat. J. Therm. Biol. 119, 103776. doi: 10.1016/j.jtherbio.2023.103776
Gómez-Gesteira M., Gimeno L., DeCastro M., Lorenzo M. N., Alvarez I., Nieto R., et al. (2011). The state of climate in NW Iberia. Clim. Res. 48, 109–144. doi: 10.3354/cr00967
Gosling E. (2015). Marine bivalve molluscs. 2nd ed. (United Kingdom: Wiley Blackwell). doi: 10.1002/9781119045212
He G., Zou J., Liu X., Liang F., Liang J., Yang K., et al. (2022). Assessing the impact of atmospheric heatwaves on intertidal clams. Sci. Total Environ. 841, 156744. doi: 10.1016/j.scitotenv.2022.156744
Helmuth B., Mieszkowska N., Moore P., and Hawkins S. J. (2006). Living on the edge of two changing worlds: forecasting the responses of rocky intertidal ecosystems to climate change. Annu. Rev. Ecol. Evol. Syst. 37, 373–404. doi: 10.1146/annurev.ecolsys.37.091305.110149
Hemminga M. A. (1998). The root/rhizome system of seagrasses: An asset and a burden. J. Sea Res. 39, 183–196. doi: 10.1016/S1385-1101(98)00004-5
Hobday A. J., Alexander L. V., Perkins S. E., Smale D. A., Straub S. C., Oliver E. C. J., et al. (2016). A hierarchical approach to defining marine heatwaves. Prog. Oceanogr. 141, 227–238. doi: 10.1016/j.pocean.2015.12.014
IPCC and Core Writing Team (2023). “Summary for policymakers,” in Climate change 2023: synthesis report. Contribution of working groups I, II and III to the sixth assessment report of the intergovernmental panel on climate change. Eds. Lee H. and Romero J. (IPCC, Geneva, Switzerland), 1–34. doi: 10.59327/IPCC/AR6-9789291691647.001
Kristensen E., Penha-Lopes G., Delefosse M., Valdemarsen T., Quintana C. O., and Banta G. T. (2012). What is bioturbation? The need for a precise definition for fauna in aquatic sciences. Mar. Ecol. Prog. Ser. 446, 285–302. doi: 10.3354/meps09506
Lee K. S., Park S. R., and Kim Y. K. (2007). Effects of irradiance, temperature, and nutrients on growth dynamics of seagrasses: A review. J. Exp. Mar. Bio. Ecol. 350, 144–175. doi: 10.1016/j.jembe.2007.06.016
Lenth R. (2024). “emmeans: Estimated marginal means, aka least-squares means,” in R package version 1.10.0. Available online at: https://CRAN.R-project.org/package=emmeans (Accessed January 12, 2025).
Luo X., Masanja F., Liu Y., and Zhao L. (2025). Behavioral responses of clams to recurrent marine heatwaves. Mar. pollut. Bull. 210, 117362. doi: 10.1016/j.marpolbul.2024.117362
Macho G., Woodin S. A., Wethey D. S., and Vázquez E. (2016). Impacts of sublethal and lethal high temperatures on clams exploited in European fisheries. J. Shell Res. 35, 405–419. doi: 10.2983/035.035.0215
Massa S. I., Arnaud-Haond S., Pearson G. A., and Serrao E. A. (2009). Temperature tolerance and survival of intertidal populations of the seagrass Zostera noltii (Hornemann) in southern Europe (Ria Formosa, Portugal). Hydrobiol. 619, 195–201. doi: 10.1007/s10750-008-9609-4
Maxwell P. S., Eklöf J. S., van Katwijk M. M., O’Brien K. R., de la Torre-Castro M., Boström C., et al. (2017). The fundamental role of ecological feedback mechanisms for the adaptive management of seagrass ecosystems – a review. Biol. Rev. 92, 1521–1538. doi: 10.1111/brv.12294
Melià P. and Gatto M. (2005). A stochastic bioeconomic model for the management of clam farming. Ecol. Modell. 184, 163–174. doi: 10.1016/j.ecolmodel.2004.11.011
Meysick L., Norkko A., Gagnon K., Gräfnings M., and Boström C. (2020). Context-dependency of eelgrass-clam interactions: Implications for coastal restoration. Mar. Ecol. Prog. Ser. 647, 93–108. doi: 10.3354/meps13408
Mittaz J. and Harris A. (2011). A physical method for the calibration of the AVHRR/3 thermal IR channels. Part II: An in-orbit comparison of the AVHRR longwave thermal IR channels on board MetOp-A with IASI. J. Atmos. Ocean Technol. 28, 1072–1087. doi: 10.1175/2011JTECHA1517.1
Miyajima T. and Hamaguchi M. (2019). “Carbon sequestration in sediment as an ecosystem function of seagrass meadows,” in Blue carbon in shallow coastal ecosystems. Eds. Kuwae T. and Hori M. (Springer, Singapore), 33–72. doi: 10.1007/978-981-13-1295-3_2
Navajas R., González J. L., and Royo A. (2003). Revisión del cultivo de la almeja japonesa (Ruditapes philippinarum, Adams & Reeve (1850) en España (Cádiz, Spain: IX Congreso Nacional de Acuicultura).
Olivé I., Brun F. G., Vergara J. J., and Pérez-Lloréns J. L. (2007). Effects of light and biomass partitioning on growth, photosynthesis and carbohydrate content of the seagrass Zostera noltii Hornem. J. Exp. Mar. Bio. Ecol. 345, 90–100. doi: 10.1016/j.jembe.2007.02.008
Perkins S. E. and Alexander L. (2013). On the measurement of heat waves. J. Clim. 26, 4500–4517. doi: 10.1175/JCLI-D-12-00383.1
Peterson B. J. and Heck K. L. (2001). Positive interactions between suspension-feeding bivalves and seagrass - a facultative mutualism. Mar. Ecol. Prog. Ser. 213, 143–155. doi: 10.3354/meps213143
Pineda M. C., McQuaid C. D., Turon X., López-Legentil S., Ordóñez V., and Rius M. (2012). Tough adults, frail babies: an analysis of stress sensitivity across early life-history stages of widely introduced marine invertebrates. PloS One 7, e46672. doi: 10.1371/journal.pone.0046672
Pinheiro J., Bates D., DebRoy S., Sarkar D., Heisterkamp S., Van Willingen B., et al. (2022). “nlme: Linear and nonlinear mixed effects models,” in R package version 3, 1–162. Available online at: https://CRAN.R-project.org/package=nlme (Accessed January 12, 2025).
Rafaelli D. and Hawkins S. (1996). “Causes of zonation,” in Intertidal ecology. Eds. Rafaelli D. and Hawkins S. (Kluwer Academic Publishers, Dordrecht, The Netherlands), 71–97.
Rato A., Joaquim S., Matias A. M., Roque C., Marques a., and Matias D. (2022). The Impact of Climate Change on Bivalve Farming: Combined Effect of Temperature and Salinity on Survival and Feeding Behavior of Clams Ruditapes decussatus. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.932310
Raymond W. W., Barber J. S., Dethier M. N., Hayford H. A., Harley C. D. G., King T. L., et al. (2022). Assessment of the impacts of an unprecedented heatwave on intertidal shellfish of the Salish Sea. Ecol. 103, e3798. doi: 10.1002/ecy.3798
R Core Team (2022). R: A language and environment for statistical computing (Vienna: R Foundation for Statistical Computing).
Reynolds L. K., DuBois K., Abbott J. M., Williams S. L., and Stachowicz J. J. (2016). Response of a habitat-forming marine plant to a simulated warming event is delayed, genotype specific, and varies with phenology. PloS One 11, 1–16. doi: 10.1371/journal.pone.0154532
Richards S. A., Whittingham M. J., and Stephens P. A. (2011). Model selection and model averaging in behavioural ecology: The utility of the IT-AIC framework. Behav. Ecol. Sociobiol. 65, 77–89. doi: 10.1007/s00265-010-1035-8
Roca G., Alcoverro T., Krause-Jensen D., Balsby T. J. S., van Katwijk M. M., Marbà N., et al. (2016). Response of seagrass indicators to shifts in environmental stressors: a global review and management synthesis. Ecol. Indicat. 63, 310–323. doi: 10.1016/j.ecolind.2015.12.007
Román M., Gilbert F., Viejo R. M., Román S., Troncoso J. S., Elsa V., et al. (2023). Are clam-seagrass interactions affected by heatwaves during emersion? Mar. Environ. Res. 186, 105906. doi: 10.1016/j.marenvres.2023.105906
Román S., Vázquez E., Román M., Viejo R. M., Weidberg N., Troncoso J. S., et al. (2025). Understanding the effects of recent atmospheric heatwaves on seagrass-inhabited intertidal shellfish beds: A mesocosm experiment. Mar. env. Res. 212, 107547. doi: 10.1016/j.marenvres.2025.107547
Román S., Vázquez E., Román M., Viejo R. M., and Woodin S. A. (2022). Effects of warming on biological interactions between clams and the seagrass Zostera noltei: A case study using open top chambers. Estuar. Coast. Shelf Sci. 276, 108027. doi: 10.1016/j.ecss.2022.108027
Román M., Vázquez E., Viejo R. M., Woodin S. A., Wethey D. S., Román S., et al. (2024). Context-dependent resilience of intertidal seagrass and venerid clams after hyposalinity stress. Mar. Ecol. Prog. Ser. 729, 117–133. doi: 10.3354/meps14516
Royo A., Quintero D., Hurtado Burgos M., and Hurtado Cancelo M. (2002). Cultivo de almeja japonesa Ruditapes philippinarum (Adams and Reeve 1850) a altas densidades de siembra y en zona intermareal) a altas densidades de siembra y en zona intermareal. Bol. Inst. Esp. Oceanogr. 18, 349–356. Available online at: http://hdl.handle.net/10261/313233.
Shick J. M., Widdows J., and Gnaiger E. (1988). Calorimetric studies of behavior, metabolism and energetics of sessile intertidal animals. Integr. Comp. Biol. 28, 161–181. doi: 10.1093/icb/28.1.161
Smetacek V. and Zingone A. (2013). Green and golden seaweed tides on the rise. Nat. 504 7478, 84–88. doi: 10.1038/nature12860
Southward A. J. (1958). Note on the temperature tolerances of some intertidal animals in relation to environmental temperatures and geographical distribution. J. Mar. Biol. Assoc. United Kingdom 37, 49–66. doi: 10.1017/S0025315400014818
Stillman J. H. (2019). Heat waves, the new normal: Summertime temperature extremes will impact animals, ecosystems, and human communities. Physiol. 34, 86–100. doi: 10.1152/physiol.00040.2018
Thompson J. N. (1988). Variation in interspecific interactions. Ann. Rev. Ecol. Syst. 19, 65–87. Available online at: https://www.jstor.org/stable/2097148 (Accessed January 12, 2025).
Touchette B. and Burkholder J. (2000). Review of nitrogen and phosphorus metabolism in seagrasses. J. Exp. Mar. Bio. Ecol. 250, 133–167. doi: 10.1016/S0022-0981(00)00195-7
Underwood A. J. (1997). “Factorial experiments,” in Experiments in ecology: Their logical design and interpretation using analyses of variance. Ed. Underwood A. J. ( Cambridge University Press)Cambridge, 298–358. doi: 10.1017/CBO9780511806407
United Nations Environment Programme (2020). Out of the blue: The value of seagrasses to the environment and to people (Nairobi: UNEP). Available online at: https://www.unep.org/resources/report/out-blue-value-seagrasses-environment-and-people (Accessed January 12, 2025).
Vaz-Pinto F., Olabarria C., Gestoso I., Cacabelos E., Incera M., and Arenas F. (2013). Functional diversity and climate change: Effects on the invasibility of macroalgal assemblages. Biol. Invasions 15, 1833–1846. doi: 10.1007/s10530-013-0412-4
Vázquez E., Woodin S. A., Wethey D. S., Peteiro L. G., and Olabarria C. (2021). Reproduction Under Stress: Acute Effect of Low Salinities and Heat Waves on Reproductive Cycle of Four Ecologically and Commercially Important Bivalves. Front. Mar. Sci. 8, 1–19. doi: 10.3389/fmars.2021.685282
Volkenborn N., Meile C., Polerecky L., Pilditch C. A., Norkko A., Norkko J., et al. (2012). Intermittent bioirrigation and oxygen dynamics in permeable sediments: An experimental and modeling study of three tellinid bivalves. J. Mar. Res. 70, 794–823. doi: 10.1357/002224012806770955
Von Richthofen F. (1886). Führer für forschungsreisende (Jenecke, Hannover: Verlag Robert Oppenheim), 743.
Wall C. C., Peterson B. J., and Gobler C. J. (2008). Facilitation of seagrass Zostera marina productivity by suspension-feeding bivalves. Mar. Ecol. Prog. Ser. 357, 165–174. doi: 10.3354/meps07289
Walne P. R. and Mann R. (1975). “Growth and biochemical composition of Ostrea edulis and Crassostrea gigas,” in Proceedings of the 9th european marine biology symposium oban. Ed. Barnes H.(Scotland: Aberdeen University Press), 587–607.
Wernberg T., Thomsen M. S., Baum J. K., Bishop M. J., Bruno J. F., and Coleman M. A. (2024). Impacts of climate change on marine foundation species. Ann. Rev. Mar. Sci. 16, 247–282. doi: 10.1146/annurev-marine-042023-093037
Keywords: atmospheric heatwave, low tide, intertidal, seagrass, clams, mutualistic interactions, shellfisheries
Citation: Román M, Vázquez E, Weidberg N, Viejo RM, Román S, Troncoso JS, Wethey DS, Woodin SA and Olabarria C (2025) Legacy effects of an atmospheric heatwave are buffered by mutualistic interactions between intertidal clams and seagrass. Front. Mar. Sci. 12:1706079. doi: 10.3389/fmars.2025.1706079
Received: 15 September 2025; Accepted: 17 October 2025;
Published: 07 November 2025.
Edited by:
Iñigo Muxika, Technological Center Expert in Marine and Food Innovation (AZTI), SpainReviewed by:
Andrea Augusto Sfriso, University of Ferrara, ItalyEttore Nepote, Marche Polytechnic University, Italy
Copyright © 2025 Román, Vázquez, Weidberg, Viejo, Román, Troncoso, Wethey, Woodin and Olabarria. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marta Román, bWFycm9tYW5AdXZpZ28uZXM=