 Itxaso Álvarez-Rodríguez1,2
Itxaso Álvarez-Rodríguez1,2 Begoña Ugarte-Uribe1†
Begoña Ugarte-Uribe1† Igor de la Arada1,2
Igor de la Arada1,2 José Luis R. Arrondo1,2
José Luis R. Arrondo1,2 Carlos Garbisu3
Carlos Garbisu3 Itziar Alkorta1,2*
Itziar Alkorta1,2*- 1Department of Biochemistry and Molecular Biology, University of the Basque Country (UPV/EHU), Leioa, Spain
- 2Instituto Biofisika (UPV/EHU, CSIC), University of the Basque Country (UPV/EHU), Spanish Research Council (CSIC), Leioa, Spain
- 3NEIKER, Soil Microbial Ecology Group, Department of Conservation of Natural Resources, Derio, Spain
Type IV Coupling Proteins (T4CPs) are essential elements in many type IV secretion systems (T4SSs). The members of this family display sequence, length, and domain architecture heterogeneity, being the conserved Nucleotide-Binding Domain the motif that defines them. In addition, most T4CPs contain a Transmembrane Domain (TMD) in the amino end and an All-Alpha Domain facing the cytoplasm. Additionally, a few T4CPs present a variable domain at the carboxyl end. The structural paradigm of this family is TrwBR388, the T4CP of conjugative plasmid R388. This protein has been widely studied, in particular the role of the TMD on the different characteristics of TrwBR388. To gain knowledge about T4CPs and their TMD, in this work a chimeric protein containing the TMD of TraJpKM101 and the cytosolic domain of TrwBR388 has been constructed. Additionally, one of the few T4CPs of mobilizable plasmids, MobBCloDF13 of mobilizable plasmid CloDF13, together with its TMD-less mutant MobBΔTMD have been studied. Mating studies showed that the chimeric protein is functional in vivo and that it exerted negative dominance against the native proteins TrwBR388 and TraJpKM101. Also, it was observed that the TMD of MobBCloDF13 is essential for the mobilization of CloDF13 plasmid. Analysis of the secondary structure components showed that the presence of a heterologous TMD alters the structure of the cytosolic domain in the chimeric protein. On the contrary, the absence of the TMD in MobBCloDF13 does not affect the secondary structure of its cytosolic domain. Subcellular localization studies showed that T4CPs have a unipolar or bipolar location, which is enhanced by the presence of the remaining proteins of the conjugative system. Unlike what has been described for TrwBR388, the TMD is not an essential element for the polar location of MobBCloDF13. The main conclusion is that the characteristics described for the paradigmatic TrwBR388 T4CP should not be ascribed to the whole T4CP family. Specifically, it has been proven that the mobilizable plasmid-related MobBCloDF13 presents different characteristics regarding the role of its TMD. This work will contribute to better understand the T4CP family, a key element in bacterial conjugation, the main mechanism responsible for antibiotic resistance spread.
Introduction
Type IV coupling proteins (T4CPs) are essential elements in the conjugative type IV secretion systems (T4SSs) and are also key elements in many pathogenic T4SSs. The members of this family display a high sequence, length, and domain architecture heterogeneity being the Nucleotide-Binding Domain (NBD) the only conserved domain in all T4CPs. For this reason, they are classified according to the different domain architectures: (i) VirD4-type subfamily that are integral membrane proteins; (ii) TraG-J pairs, which are also integral membrane proteins but additionally present a physical and functional association with another membrane protein of the T4SS; (iii) T4CPs without Transmembrane Domain (TMD), which could or could not interact with other T4SS membrane proteins creating a VirD4-type complex, like the pair TraJpIP501 and TraIpIP501; (iv) FtsK-like T4CPs; and (v) Archaeal T4CPs (Llosa and Alkorta, 2017).
The structural paradigm of this family is the T4CP of conjugative plasmid R388, TrwBR388. It is a VirD4-type protein composed of a TMD at the N-terminus (consisting of two transmembrane α-helices connected through a small periplasmic loop) and a bulky globular cytosolic domain (CD). TrwBR388 is the only full-length T4CP that has been successfully purified to date (Hormaeche et al., 2002; Redzej et al., 2017), while trials for purifying other membrane T4CPs have not rendered the sufficient amounts of high quality protein for performing in vitro assays (Chen et al., 2008). For this reason most of the in vitro studies of T4CPs have been achieved using deletion mutant proteins that lack the TMD (Schroder and Lanka, 2003; Tato et al., 2007; Larrea et al., 2017). In this regard, the TMD deletion mutant protein of TrwBR388, TrwBΔN70, was resolved by X-ray crystallography, showing that the CD of TrwBR388 contains an NBD with the Walker A and Walker B motifs and a small membrane-distal All-Alpha Domain (AAD) (Gomis-Rüth et al., 2001).
Comparative studies of the properties of TrwBR388 and TrwBΔN70 showed significant differences regarding biological activity (such as in vivo function, in vitro nucleotide-binding properties, and in vitro ATPase activity), oligomerization pattern, subcellular location, and stability (Moncalián et al., 1999; Vecino et al., 2010, 2011; Hormaeche et al., 2002, 2004, 2006; Segura et al., 2013, 2014). For this reason it has been concluded that the TMD of TrwBR388 accomplishes a role beyond the anchorage of the protein to the membrane, influencing the location, stability, and activity of this protein.
To delve into the role of the TMD in T4CPs two different strategies have been followed. On the one hand, we have constructed a chimeric protein named TMDTraJCDTrwB composed of the TMD of TraJpKM101, the phylogenetically closest T4CP to TrwBR388 from the conjugative plasmid pKM101 (Paterson et al., 1999; Alvarez-Martinez and Christie, 2009) and the CD of TrwBR388. This strategy has already been used for the study of components of T4SSs, showing interesting results (Bourg et al., 2009). Specifically, through a chimeric protein approach the function of the AAD of VirD4At from the T-plasmid of Agrobacterium tumefaciens (Whitaker et al., 2016) and of the N-terminal HUH domain of TrwCR388 (Agúndez et al., 2018) have been analyzed. On the other hand, the T4CP from the mobilizable plasmid CloDF13, MobBCloDF13 and its deletion protein lacking the TMD, MobBΔTMD, have been constructed and studied. It is an interesting system to characterize since it is part of the rare MOBC1 plasmid family, which are mobilizable plasmids that encode their T4CP (Smillie et al., 2010). Additionally, MobBCloDF13 has been described as an atypical T4CP, due to its dual role in DNA transfer, since it acts as an accessory protein in CloDF13 relaxation process and also as a T4CP (Nuñez and de la Cruz, 2001).
We studied the functionality of these proteins in plasmid transfer, secondary structure, thermal stability, and subcellular localization. Our findings indicate that the TMD plays different roles in conjugative plasmid related and mobilizable plasmid related T4CPs. Specifically while the TMD could play a regulatory role in TrwBR388 this cannot be inferred from the results about MobBCloDF13.
Materials and Methods
Materials
n-dodecyl β-D maltoside (DDM) was purchased from Anatrace (Santa Clara, CA, United States). Mouse anti-His (C-term) monoclonal antibody and Alexa Fluor goat anti-mouse antibody were purchased from Invitrogen (Carlsbad, CA, United States) and Molecular Probes (Eugene, OR, United States), respectively. All buffers employed in this work are shown in Supplementary Table S1.
Bacterial Strains and Bacterial Growth Conditions
E. coli DH5α strain was used as host for plasmid constructions. This strain also served as the donor for mating experiments by hosting the plasmids of interest. E. coli UB1637 served as the recipient for mating experiments. E. coli Lemo21 (DE3), E. coli BL21 (DE3), and E. coli BL21C41 (DE3) strains were used for protein production, purification, and in vivo localization.
E. coli strains were grown in LB medium and when necessary antibiotics were added at the following final concentrations: ampicillin (100 μg/mL), streptomycin (50 μg/mL), kanamycin (50 μg/mL), chloramphenicol (12.5–25 μg/mL), and thrimethoprim (10 μg/mL).
Plasmids
The plasmids and oligonucleotides used in this study are listed in Tables 1, 2, respectively.

Table 1. Plasmids employed in this work.

Table 2. Oligonucleotides used in this work.
pSU4814 and pSU4833 plasmids were kindly provided by Fernando de la Cruz. pOPINE (Addgene plasmid # 26043; RRID: Addgene_26043)1 and pOPINE-3C-eGFP (Addgene plasmid # 41125; RRID: Addgene_41125)2 plasmids were a gift from Ray Owens. pKM101Spcr_ΔtraJ was kindly provided by Peter J. Christie. pKM101Ampr_ΔtraJ plasmid was obtained by cleavage of the spc cassette of pKM101Spcr_ΔtraJ plasmid using EcoRI restriction enzyme.
Cloning of T4CPs
To construct the chimeric protein TMDTraJCDTrwB and the transmembrane deletion mutant protein of MobBCloDF13, MobBΔTMD, the sequences of TrwBR388, TraJpKM101, and MobBCloDF13, were analyzed through bioinformatics tools. First, the different characteristics of the proteins, such as molecular weight and isoelectric point, were analyzed using ProtParam.3 Second, the topology of the membrane proteins was studied using Topcons4 (Tsirigos et al., 2015).
Then different constructions were achieved as follows:
TMDTraJCDTrwB chimeric protein consists of amino acids M1-D76 from TraJpKM101 followed by amino acids L71-I507 from TrwBR388. The tmdTraJcdTrwB sequence was synthesized de novo and inserted it into pET24a (+) plasmid vector using NdeI and XhoI restriction sites, rendering pUBQ4 plasmid to produce the chimeric protein. This construction was made by TOP Gene Technologies, Inc. (Saint-Lauren, QC-Canada). To study the role of the conserved lysine of the Walker A motif (K142), this residue was substituted with a threonine using the QuikChange II Site-Directed Mutagenesis Kit from Stratagene (San Diego, CA, United States) to obtain the TMDTraJCDTrwB (K142T) protein. Additionally, to clone the tmdTraJcdTrwB-eGFP gene tmdTraJcdTrwB sequence was inserted into the pWaldo-GFPe plasmid vector using XhoI and BamHI restriction sites (Segura et al., 2014).
To clone MobBCloDF13-related proteins, the cloning of mobB, MobBΔTMD, mob-eGFP, and MobBΔTMD-eGFP genes was performed in the Oxford Protein Production Facility (OPPF-UK) using the High-throughput protocol described by Bird (2011). Specifically, MobBΔTMD soluble mutant protein consists of amino acids D185-Y653 of MobBCloDF13 obtained after deletion of amino acids M1-A184 from wild type MobBCloDF13.
All the oligonucleotides employed in the aforementioned cloning experiments are specified in Table 2.
Overexpression and Purification
The same purification protocol was followed for TMDTraJCDTrwB and MobBCloDF13 proteins. Briefly, E. coli BL21C41 (DE3) cells freshly transformed with plasmids pUBQ4 for TMDTraJCDTrwB and pOPINE-mobB for MobBCloDF13 were grown overnight in LB (8 flasks of 10 mL) supplemented with the corresponding antibiotics at 37°C with continuous shaking. Then, cells were diluted 1:50 (v/v) with fresh LB supplemented with antibiotics (8 flasks of 500 mL) and were grown at 37°C with continuous shaking until an OD600 of 0.4–0.5 was achieved. Next, overexpression was induced by the addition of 1 mM isopropyl α-D-thiogalactopyranoside (IPTG) and cells were grown with continuous shaking at 25°C overnight. Cells were harvested by centrifugation at 8,000 g for 15 min at 4°C. The pellet was suspended in 80 mL of Cell buffer, frozen with liquid N2 and stored at −80°C.
For purification, cells were thawed at 37°C and 0.02 mg/mL DNase I, 1 mM dithiothreitol (DTT), 0.07% (w/v) lysozyme, 1 mM MgCl2, 1 mM phenylmethanesulfonyl fluoride (PMSF) and two tablets of cOmpleteTM EDTA-free Protease Inhibitor Cocktail from Sigma-Aldrich (San Luis, MO, United States) were added. From this point onward, the whole process was performed at 4°C to avoid aggregation of the proteins. After 45 min of incubation with agitation, cells were disrupted by sonication and centrifuged at 8,000 g for 15 min to remove non-lysed cell. The supernatant, containing the broken cells, was centrifuged at 138,000 g for 45 min to pellet the membrane fraction which was subsequently carefully resuspended in 30 mL of Cell buffer. Then DDM and NaCl were added to final concentrations of 19.6 mM and 600 mM, respectively, and the volume was adjusted to 40 mL. The mixture was incubated for 90 min with continuous stirring and then centrifuged at 138,000 g for 1 h.
The supernatant containing the protein to be purified was mixed 1:1 (v/v) with MP1 buffer to decrease the concentration of DDM and NaCl to 8.3 mM and 300 mM, respectively. Then, the sample was supplemented with 50 mM imidazole and loaded onto a 5 mL HisTrapTM FF (GE Life Sciences; Marlborough, MA, United States) column previously equilibrated with MP2 buffer. To increase the binding of the protein, the sample was left recirculating overnight. Next, it was connected to an ÄKTA-FPLC system and it was washed with 50 mL of MP2 buffer at a flow rate of 2 mL/min until the absorbance at 280 nm reached the baseline. Bound proteins were eluted with MP3 buffer, at a flow rate of 1.5 mL/min and fractions of 1 mL were collected. Obtained samples were analyzed by SDS-PAGE and the ones containing each target protein were pulled-down and concentrated using a centrifugal filter with a MWCO of 100 kDa. Then, 5 mL of the resulting samples were separately loaded onto a Superdex 200 HR 16/60 column and the size-exclusion chromatography (SEC) was performed in MP purification buffer at 0.5 mL/min. The fractions corresponding to each target protein were pulled-down and concentrated as explained before. Glycerol was added to a final concentration of 20% (v/v) and protein concentration was determined by measuring absorption at 280 nm. Finally, aliquots were stored at −80°C.
When TMDTraJCDTrwB was purified with the aim of performing infrared spectroscopy (IR) assays the DDM and NaCl concentrations of the MP purification buffer were changed to the ones described in the previously published purification protocols of TrwBR388 and TrwBΔN50 (i.e., 0.2 mM DDM and 200 mM NaCl instead of 0.6 mM DDM and 300 mM NaCl) (Vecino et al., 2011, 2012).
For MobBΔTMD purification, E. coli Lemo21 (DE3) cells freshly transformed with pOPINE-MobBΔTMD plasmid were grown in 4 L of LB supplemented with ampicillin at 37°C with continuous shaking until an OD600 of 0.5–0.6 was reached. Expression was induced with 1 mM IPTG and performed for 20 h at 25°C. Cells were harvested and stored as explained previously.
For protein purification, the cell suspension was thawed and the lysis protocol previously described for TMDTraJCDTrwB and MobBCloDF13 was followed. Then, the sample was centrifuged at 138,000 g for 45 min to pellet the membrane fraction and the inclusion bodies. The supernatant with the soluble proteins was supplemented with 50 mM imidazole and loaded onto a 5 mL HisTrapTM FF (GE Life Sciences; Marlborough, MA, United States) column, previously equilibrated with MobBΔTMD1 buffer. Affinity chromatography was performed as previously described for MobBCloDF13, but using MobBΔTMD1 buffer for washing and MobBΔTMD2 buffer for elution. Fractions containing the target protein were pulled-down and concentrated using a Centricon YM-30 to a final volume of 600 μL. The resulting sample was centrifuged to remove aggregates and loaded onto a Superdex 200 HR 10/30 column. The SEC was performed in Cell buffer at 0.3 mL/min and 0.5 mL fractions were collected. Fractions that contained the protein of interest were pulled-down and the sample was centrifuged to discard the aggregates. Glycerol was added to a final concentration of 20% (v/v) and aliquots were made.
Mating Assays
Mating assays were performed as described by Llosa et al. (2003) with small modifications. Briefly, donors (E. coli DH5α co-transformed with the appropriate plasmids) and recipient cells (E. coli UB1637) were grown in LB supplemented with the corresponding antibiotics overnight at 37°C. For each mating assay 100 μL of the donor and the recipient cells were mixed, centrifuged, resuspended in 50 μL LB and placed onto a GS Millipore filter (0.22 μm pore size) settled on a pre-warmed LB-agar plate. After 1 h incubation at 37°C bacteria were washed from the filters in 2 mL LB by shaking at 450 rpm for 20 min and vortexing for 30 s. Then, 100 μL of the appropriate dilutions were plated on selective media for donors and transconjugants. The plates were incubated overnight at 37°C and the colonies were counted, normalizing the conjugation frequency as the number of transconjugants per donor cell.
Infrared Spectroscopy
To accomplish IR studies of different T4CPs and their variants (i.e., TMDTraJCDTrwB, MobBCloDF13, and MobBΔTMD) purification of each protein was carried out as previously described. The H-D exchange protocol was performed at 4°C and adapted for each protein. Briefly, TMDTraJCDTrwB was diluted with the IR buffer, dialyzed against the same buffer using a D-TubeTM Dialyzer Midi (MWCO 3.5 kDa) (Merck; Darmstadt, Germany), diluted again in IR buffer and concentrated using an Amicon Ultra-0.5 mL centrifugal filter (MWCO: 100 kDa). A similar process was followed for MobBCloDF13 except for the dialysis step. Regarding MobBΔTMD, sample was diluted, dialyzed and concentrated as described for TMDTraJCDTrwB but using in MobBΔTMD IR buffer. Final protein samples were always above 1 mg/mL protein concentration.
Infrared spectra were recorded in a Thermo Nicolet Nexus 5700 (Thermo Fisher Scientific; Waltham, MA, United States) spectrometer equipped with a liquid nitrogen-refrigerated mercury-cadmium-telluride detector using a Peltier-based temperature controller (TempCompTM, BioTools; Wauconda, IL, United States), and a 25 μm optical path. Typically 370 scans for each, background and sample, were collected at 2 cm–1 resolution and averaged after each minute. Spectra were collected with OMNIC software (Nicolet) and data processing was performed with OMNIC and SpectraCalc, following previously resolved methods (Arrondo et al., 1993; Arrondo and Goñi, 1999).
The information about the secondary structure and about the thermal denaturation of T4CPs and their derivatives was obtained by IR spectroscopy through analysis of the infrared amide I band that corresponds mainly to the C = O stretching vibrations of the peptide bonds and which is located between 1700 and 1600 cm–1 region of the IR spectrum. Amide I band is conformationally sensitive and can be used to monitor the protein secondary structure composition and changes induced by thermal denaturation. Secondary structure studies were made at 20°C and band decomposition of the amide I was performed as previously reported (Vecino et al., 2011, 2012). For thermal stability studies samples were heated from 20 to 80°C at a rate of 1°C/min.
Subcellular Location of T4CPs
Subcellular location of different T4CPs and their variants (i.e., TrwBR388, TMDTraJCDTrwB, MobBCloDF13, and MobBΔTMD) was achieved by confocal fluorescence microscopy. To do so, two different approaches were used: (i) eGFP-labeling (Cormack et al., 1996) and (ii) immunofluorescence. Since the eGFP moiety only emits fluorescence when properly folded (Drew et al., 2005), the eGFP based approach allowed visualizing only properly folded proteins.
Prior to localization assays, the in vivo activity of the T4CP-eGFP fusion-proteins was proved by mating assays as previously described (Supplementary Table S2). Afterward, T4CP-eGFP fusion-proteins were expressed in E. coli BL21C41 (DE3) strain, except for MobBΔTMD that was expressed in BL21 (DE3) strain. To do so cells were transformed with pUBQ4, by induction with 1 mM IPTG at OD600 0.4–0.5 for the membrane proteins and OD600 0.5–0.6 for MobBΔTMD. Protein expression was performed for 4 and 20 h at 25°C. Additionally, the subcellular location of TrwBR388-related proteins was determined in the presence of pSU1456 plasmid, which codes for all the R388 conjugative proteins except TrwBR388. Similarly, MobBCloDF13 was also expressed in the presence of plasmid pSU1456 and to mimic the in vivo transfer of CloDF13, its location was additionally studied in the presence of plasmids pSU1456 (R388 plasmid that lacks TrwBR388 protein) and pSU4833 (CloDF13 plasmid that contains its mobilization region except for MobBCloDF13 protein). Sample handling was performed as described by Segura et al. (2014). The images were acquired in a Leica TCS SP5 confocal fluorescence microscope, with a 60× oil immersion objective. Sample excitation was performed with 488 nm wavelength, while fluorescence emission was measured between 505 and 525 nm. The images were analyzed using Huygens and ImageJ softwares. To ease the counting process and better distinguish the different locations the images were treated with the preset ICE filter of ImageJ software; in this manner five different locations for the T4CP-eGFP fusion-proteins were described (Supplementary Figure S1).
For immunofluorescence assays protein expression was performed as with the eGFP fusion-proteins. Sample collection and handling was performed as described by Segura et al. (2014). Cells were immunostained with mouse anti-His (C-term) monoclonal antibody as primary antibody, and Alexa Fluor goat anti-mouse as secondary antibody. Image acquisition was performed in an Olympus FluoviewTM 500 confocal fluorescence microscope at the “Analytical and high-resolution microscopy in biomedicine” facility (SGIker, UPV/EHU).
Results
Bacterial conjugation is one of the main processes responsible for the horizontal dissemination of antibiotic resistance genes among bacteria. One of the essential proteins in this process is the T4CP, which is ubiquitous in all conjugative systems. Despite its importance, the only widely studied T4CP is TrwBR388. Given its central role in bacterial conjugation, detailed knowledge of the T4CP family could contribute to the development of new strategies against the spread of antibiotic resistance among bacteria.
Previously published papers have highlighted the role of the TMD on different characteristics of TrwBR388, such as plasmid conjugation (Moncalián et al., 1999), subcellular localization (Segura et al., 2014), nucleotide-binding (Hormaeche et al., 2006), hexamerization (Hormaeche et al., 2002; Matilla et al., 2010), protein stability (Hormaeche et al., 2004), interaction with other proteins of the T4SS of R388 (Segura et al., 2013), and ATP hydrolase activity (Tato et al., 2005, 2007). From all these studies it was inferred that the TMD of TrwBR388 has a role beyond the mere anchorage in the membrane.
To gain more knowledge about different T4CPs, and in particular about the role of their TMD in T4CP features, in this work a TrwBR388 chimeric protein that combines its CD with the TMD of its phylogenetically closest T4CP, TraJpKM101, has been studied. Also one of the few T4CPs of mobilizable plasmids, MobBCloDF13, and its TMD deletion mutant protein, MobBΔTMD, have been studied. Plasmid transfer, secondary structure, thermal stability, and subcellular location studies have been carried out to shed light on the functioning of this protein family and in the role of their TMD.
Cloning of Soluble Mutant and Chimeric Proteins
The membrane protein topologies obtained after the bioinformatic analysis performed with Topcons software of TrwBR388, TraJpKM101, and MobBCloDF13 are shown in Figure 1A. TrwBR388 and TraJpKM101 have similar size and organization of their TMDs that consist of about 70 residues and contain two α-helixes connected by a small periplasmic loop. In contrast, the TMD of MobBCloDF13 is larger (about 150 amino acids) and is organized into three α-helixes. This information was used to design the chimeric and mutant proteins studied in this work (Figure 1B). The chimeric protein TMDTraJCDTrwB was made by combination of the TMD of the T4CP TraJpKM101 and the CD of TrwBR388. In addition, in this work the T4CP of the mobilizable plasmid CloDF13, MobBCloDF13, and its TMD-deletion protein MobBΔTMD, were constructed (Figure 1B). The theoretical molecular weights of these proteins, necessary for their purification process, were calculated using ProtParam3 bioinformatic tool. The estimated molecular weights were 58.28, 73.95, and 53.13 kDa for TMDTraJCDTrwB, MobBCloDF13, and MobBΔTMD, respectively. Finally, the eGFP fusion-proteins (i.e., TMDTraJCDTrwB-eGFP, MobBCloDF13-eGFP, and MobBΔTMD-eGFP) were constructed and since they emitted a fluorescent signal, it was deduced that they were correctly folded (Drew et al., 2006).
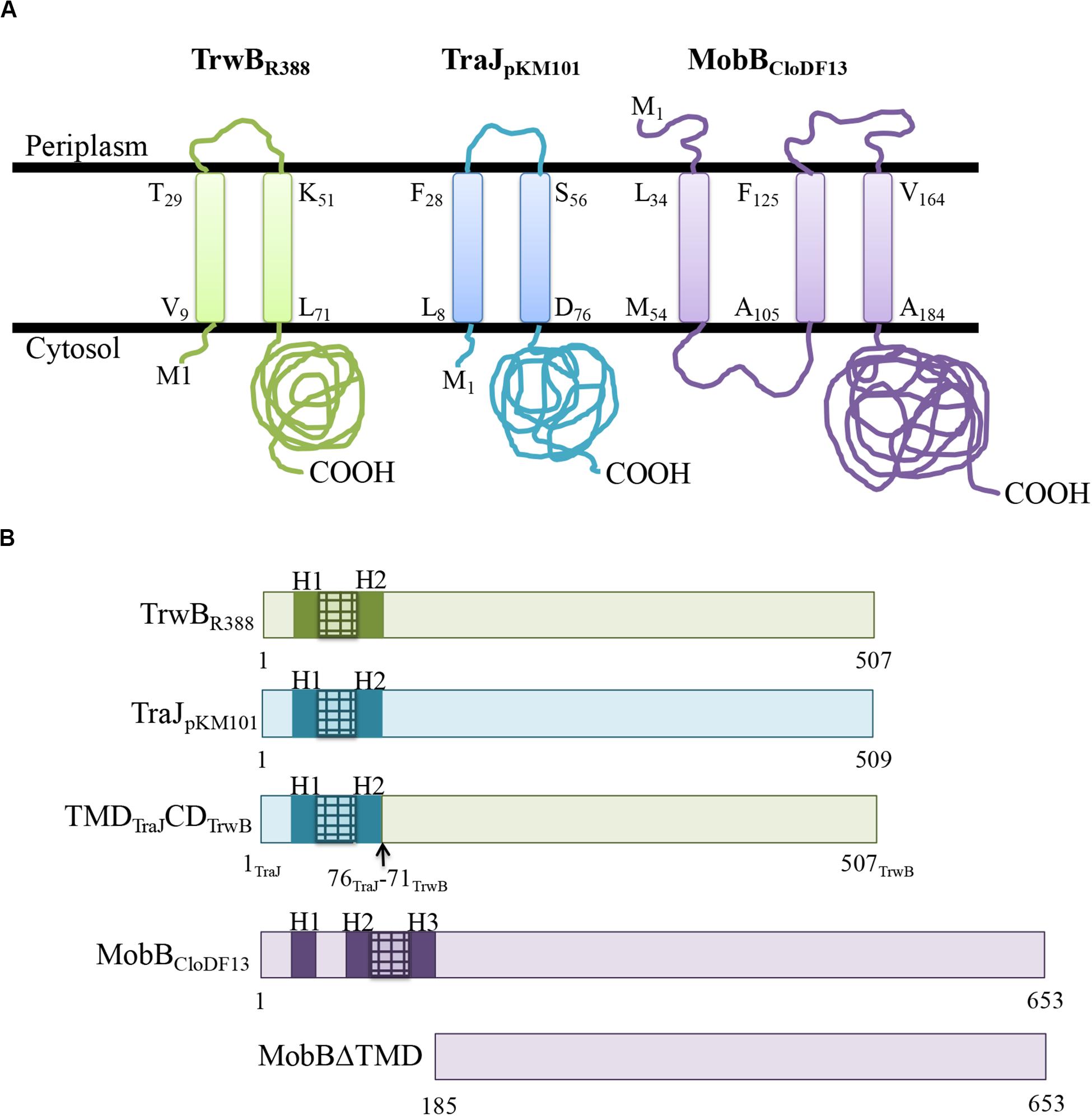
Figure 1. (A) Predicted membrane topology of TrwBR388, TraJpKM101, and MobBCloDF13 proteins. Membrane topology of the different T4CPs was predicted using Topcons software. The black lines represent the inner bacterial membrane. M1, amino-terminus; COOH, carboxy-terminus. The first and last residues of each transmembrane helix are shown indicating their position in the sequence. Proteins from R388, pKM101, and CloDF13 plasmids are shown in green, blue, and purple, respectively. (B) Schematic representation of the different T4CPs and their variants used in the present study. Proteins from R388, pKM101, and CloDF13 plasmids are shown in green, blue and purple, respectively. The transmenbrane α-helices (H) and the small periplasmic loops connecting α-helices are indicated in dark boxes and stripped boxes, respectively.
Functionality and Dominance Experiments
Through mating assays two different properties of TMDTraJCDTrwB were analyzed: (i) its capacity to complement the conjugative process in the absence of another T4CP (functionality studies) and (ii) its effect on each native conjugative system (R388 or pKM101 plasmids), being the corresponding T4CP present (TrwBR388 or TraJpKM101, respectively) (dominance studies). Results obtained in mating assays are summarized in Table 3.

Table 3. Conjugation and dominance experiments with TMDTraJCDTrwB. Transfer frequencies of plasmids pSU1456 and pKM101ΔtraJ complemented with TMDTraJCDTrwB protein have been studied.
Our results showed that TMDTraJCDTrwB efficiently complemented the ΔtrwB mutation in R388 transfer but to a lower rate than native R388 (0.21 vs. 1.82 10–4 transconjugants per donor, respectively). On the contrary, TMDTraJCDTrwB was unable to complement the ΔtraJ mutation in pKM101 transfer (Table 3). These results are in agreement with the necessary specific interactions between the CD of the T4CP and its cognate relaxase for transfer to happen as reported previously (Cabezón et al., 1997; Hamilton et al., 2000; Llosa et al., 2003).
It has been reported that mutation of the conserved lysine in the Walker A motif rendered a transfer deficient mutant protein TrwBK136T (Hormaeche et al., 2006). Similarly, an equivalent mutant of the soluble protein TrwBΔN70, TrwBΔN70 (K136T), lacked ATPase activity (Moncalián et al., 1999), underlying the essential role of this amino acid in the activity of TrwBR388. Here we studied the effect of the equivalent point mutation in the Walker A domain, TMDTraJCDTrwB (K142T), on the transfer capacity of chimeric protein. As expected, TMDTraJCDTrwB (K142T) was unable to complement the ΔtrwB mutation in R388 plasmid transfer (Table 3). This is in agreement with the crucial role of the K residue as it has been reported with homologous mutants in other T4CPs (Moncalián et al., 1999; Kumar and Das, 2002; Gunton et al., 2005).
Next, to accomplish dominance assays, the transfer frequencies of plasmids R388 or pKM101 in the presence of the cognate T4CP and the chimeric protein were measured. It was observed that TMDTraJCDTrwB reduced the transfer frequency of R388 or pKM101 by an order of magnitude (Table 3).
Mobilization Experiments
CloDF13 mobilization experiments were achieved to know whether the TMD of MobBCloDF13 was essential for the mobilization of the plasmid as happens with TrwBR388 or not as it has been described for TcpApcW3, whose TMD-less mutant can perform conjugation although at a lower frequency (Parsons et al., 2007). First of all, the transfer frequency of the mobilizable region of CloDF13 (plasmid pSU4814) mediated by the T4SS of R388 was analyzed. Afterwards, the complementation experiments were performed in the presence of both pSU4833 (the mobilizable region of CloDF13 without functional MobBCloDF13) and pSU1456 plasmid (encoding for R388 conjugative system except for TrwBR388). Results obtained in mobilization assays are summarized in Table 4. It was observed that cloned MobBCloDF13 was functional when the T4SS of R388 was used, in agreement with what has been previously published for TrwBR388 (Hormaeche et al., 2006). Similarly, the deletion of the TMD, MobBΔTMD mutant, rendered a non-functional phenotype as it occurs with other T4CP mutants that lack the TMD, such as TrwBΔN70 and PcfCΔN103 (Moncalián et al., 1999; Chen et al., 2008).

Table 4. Mobilization experiments.
Secondary Structure of T4CPs and Their Variants
Analysis of the secondary structure components of TMDTraJCDTrwB, MobBCloDF13, and MobBΔTMD were performed by IR spectroscopy through analysis of the IR amide I band.
The secondary structure of TMDTraJCDTrwB was compared to those of the native TrwBR388 and its mutants TrwBΔN50 and TrwBΔN70 (Vecino et al., 2012). Figure 2A shows the original spectra and the curve-fitting decomposition corresponding to TMDTraJCDTrwB purified in the presence of detergent. Band position, percentage area, and structure assignation corresponding to the deconvolved spectrum of the amide I region are summarized in Table 5 together with those previously reported of TrwBR388, TrwBΔN50, and TrwBΔN70 (Vecino et al., 2012).
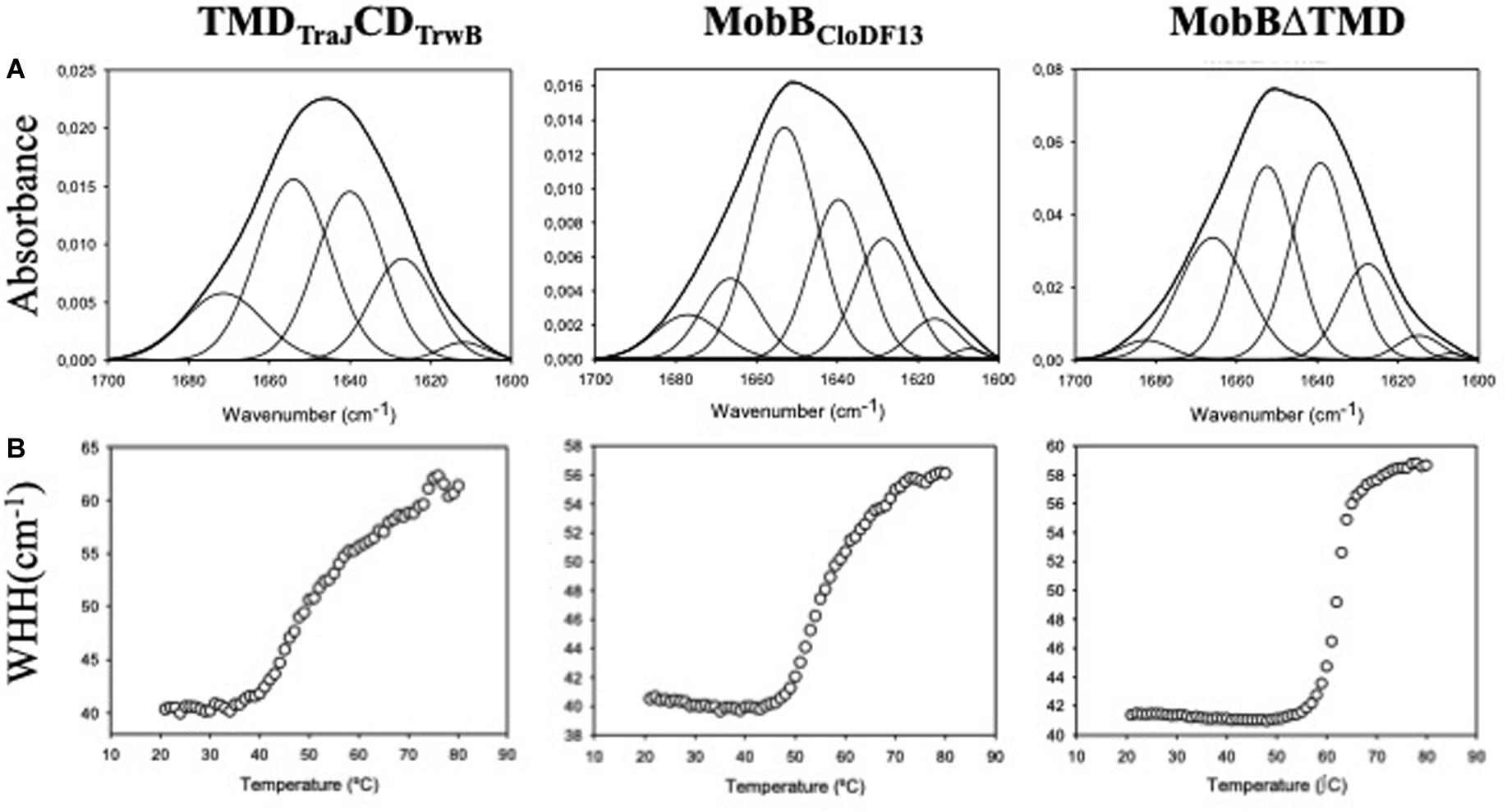
Figure 2. (A) Amide I region of the infrared spectra of TMDTraJCDTrwB, MobBCloDF13, and MobBΔTMD. Proteins were purified, dialyzed against the corresponding buffer in D2O and analyzed by IR spectroscopy as explained in “Materials and Methods” section. Obtained spectra were curve-fitted to show the different secondary structure components as detailed in Table 5. (B) Thermal denaturation of TMDTraJCDTrwB, MobBCloDF13, and MobBΔTMD as seen by IR spectroscopy. The widths at half-height (WHH) of the amide I bands are plotted as a function of temperature (°C). Thermal denaturation is marked by an abrupt increase in bandwidth. Mid-point denaturation temperature (Tm) values corresponding to each protein are detailed in Table 5.
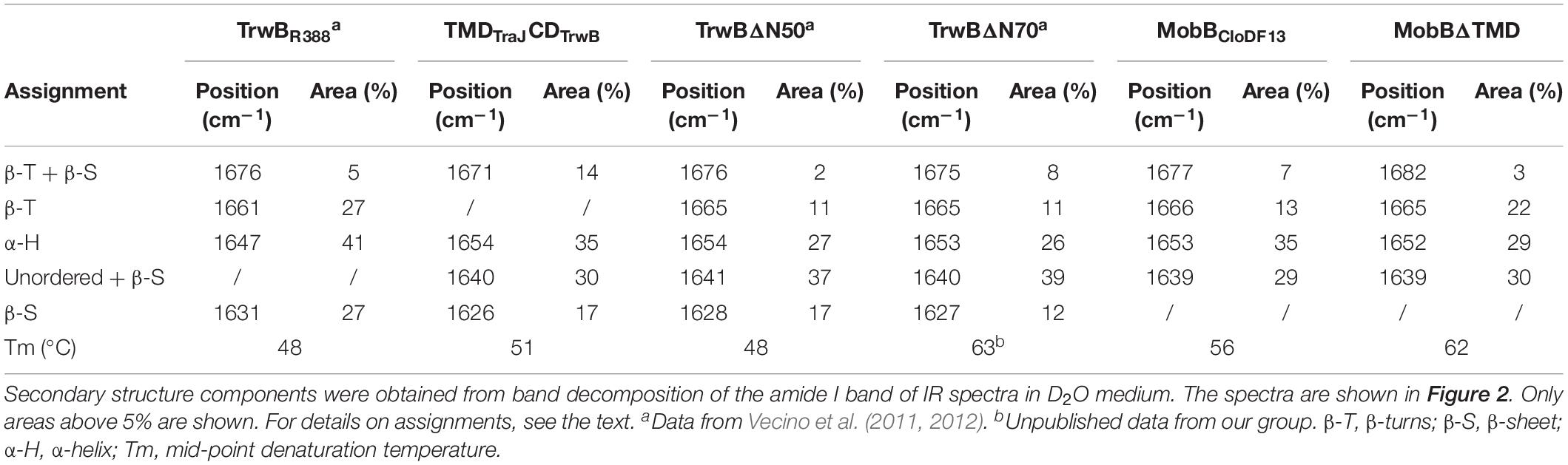
Table 5. Secondary structure components and mid-point denaturation temperatures (Tm) of TrwBR388, TMDTraJCDTrwB, TrwBΔN50, TrwBΔN70, MobBCloDF13, and MobBΔTMD.
The spectrum of TMDTraJCDTrwB purified in the presence of detergent exhibited four bands related to protein structure at 1671, 1654, 1640, and 1626 cm–1 (Table 5). Interpretation of the results must be done taking into account that band assignation is not always a straightforward process since its position can be altered by the environment (Arrondo and Goñi, 1999). The component at 1654 cm–1 was assigned to α-helix, the bands at 1626 and 1671 cm–1 were associated with the low and high-frequency vibrations of β-sheet, respectively, although it should be noted that the later is also assigned to β-turns. And the band at 1640 cm–1 was assigned to flexible, non-periodic elements.
When these results were compared to previously reported ones (i.e., TrwBR388 TrwBΔN50, and TrwBΔN70) (Vecino et al., 2012) it can be observed that the proportion of the α-helix (35%) is lower than the one observed in the native protein TrwBR388 purified in detergent (41%) and higher than the one shown in the deletion mutant proteins (26%). This result can be directly associated to the presence of a TMD both in TrwBR388 and TMDTraJCDTrwB, even if in the later belongs to another T4CP such as TraJpMK101. Regarding bands associated to β-sheet and β-turns, it is remarkable the absence of a band around 1661–1665 cm–1 in TMDTraJCDTrwB, as it was observed in TrwBR388, TrwBΔN50, and TrwBΔN70. Nevertheless the total proportion of the different bands associated to β-sheet and β-turns of TMDTraJCDTrwB (31%) is similar to the proportion seen in the soluble mutant proteins and significantly lower than that of TrwBR388 (59%). Finally, a sizeable proportion (35%) of the structure of TMDTraJCDTrwB gave off a signal centered at 1640 cm–1 (assigned to flexible, non-periodic elements) as seen in the deletion mutant proteins TrwBΔN50 and TrwBΔN70 but not in TrwBR388 (Vecino et al., 2012). Previous studies about TrwBΔN70 and TrwBΔN50 showed that this band at 1640 cm–1 also had a β-sheet component (Vecino et al., 2012). And it was published that at higher temperatures the band at 1640 cm–1 split showing a β-sheet related band, which would not happen if the band was purely composed of unordered structures (Andraka et al., 2017). To elucidate if this also happened in TMDTraJCDTrwB, IRS experiments at different temperatures were performed (20, 40, 60, and 80°C). With the increase of temperature, the 1640 cm–1 band shifts to 1645 cm–1 and that at 1626 cm–1 increases both its contribution and width, suffering also a shift to higher wavenumbers. This behavior indicates that although those two not resolved bands included in that at 1640 cm–1 do not directly split into two bands, there is a transfer of the β-sheet contribution to the 1626 cm–1 only β-sheet band, confirming that the 1640 cm–1 band in the chimeric protein was composed of both unordered and β-sheet elements.
With the aim of further studying if the deletion of the TMD has the same effect in all T4CPs, the same study was carried out with MobBCloDF13 and its deletion mutant MobBΔTMD to be compared with TrwBR388 and its variants. Figure 2A shows the original spectra and their curve-fitting decomposition corresponding to MobBCloDF13 purified in the presence of detergent and its deletion mutant MobBΔTMD. In this case, both proteins exhibited four main bands. Band position, percentage area, and structure assignation corresponding to the deconvolved spectrum of the amide I region are summarized in Table 5. It can be observed that the component at 1653 and 1652 cm–1 was assigned to α-helix in MobBCloDF13 and MobBΔTMD, respectively. But it should be pointed out that the proportion of the α-helix in MobBΔTMD (29%) was lower than in native protein (35%), as expected taking into account the three transmembrane α-helices postulated for the native protein (Figure 1A). The bands at 1666 or 1665 cm–1 were associated with β-turns which were also observed in the proteins studied in Vecino et al. (2012), but not in TMDTraJCDTrwB. Nevertheless, proportions were slightly different (13 and 22% for MobBCloDF13 and MobBΔTMD, respectively). On the contrary, the signal centered at 1677 or 1682 cm–1 assigned to low-frequency vibrations of β-sheet and β-turns showed similar proportions in MobBCloDF13, MobBΔTMD (7 and 3%, respectively) and in the proteins studied in Vecino et al. (2012) but not in TMDTraJCDTrwB where the proportion of this component was 14%. Finally, as observed in TMDTraJCDTrwB, TrwBΔN50 and TrwBΔN70, both MobBCloDF13 and MobBΔTMD had a band at 1639 cm–1 assigned to unordered structures that represented 30% of the structure, a band that is missing in TrwBR388.
Thermal Stability of T4CPs and Their Variants
The information regarding the denaturation of TMDTraJCDTrwB, MobBCloDF13, and MobBΔTMD was obtained through analysis of the amide I band (Figure 2B). Specifically, two bands appear at 1615–1620 cm–1 and 1680 cm–1 when the protein aggregates. The appearance of these bands allows monitoring the denaturation process of the protein and the calculation of the mid-point denaturation temperature (Tm) (Table 5). Data treatment and band decomposition of the original amide I have been described previously (Vecino et al., 2011, 2012).
The thermal stability of TMDTraJCDTrwB purified in detergent was compared to TrwBR388, TrwBΔN50, and TrwBΔN70 (Vecino et al., 2012). As observed in Figure 2B, the thermal denaturation of TMDTraJCDTrwB started at 35°C showing a mid-point denaturation temperature of 51.1°C at the tested conditions. This result is similar to TrwBR388 and to the TrwBΔN50 (Vecino et al., 2012; Table 5).
Thermal stability of MobBCloDF13 and MobBΔTMD was also studied by IR spectroscopy. As depicted in Figure 2B, the denaturation of the native protein starts at 47°C, achieving its Tm at 56°C, while the denaturation of MobBΔTMD starts at 55°C, achieving its Tm at 62°C (Table 5).
Subcellular Location
Studies about subcellular location of different T4CPs reported up to now have rendered ambiguous results (Kumar and Das, 2002; Gunton et al., 2005; Segura et al., 2014). To gain a deeper knowledge of this matter, specifically regarding the role of the TMD, we have studied the subcellular location of TMDTraJCDTrwB, MobBCloDF13, and MobBΔTMD under different experimental conditions. Subcellular location was analyzed by confocal fluorescence microscopy using eGFP-labeling and immunofluorescence techniques. Since eGFP proteins are only fluorescent when they are properly folded, their visualization ensures the analysis of functional proteins, excluding those that are denatured or included in inclusion bodies (Drew et al., 2006). In both studies similar results were obtained but eGFP-labeling rendered better quality images (Supplementary Figure S2).
To study the effect of expression times in the absence of the rest of T4SS proteins, the subcellular location of each protein was visualized after induction with 1 mM IPTG for 4 or 20 h. Different patterns were observed in the location of each protein at different expression times (Figure 3 and Table 6). After 4 h of expression, TrwBR388 and TMDTraJCDTrwB were mainly located along the whole cell membrane and switched to a single-pole after 20 h, being this change less pronounced in the case of the chimeric protein. MobBCloDF13 was predominantly located at both poles both at 4 and 20 h, showing a little increase in one pole location after 20 h (Figure 4 and Table 6). On the contrary, most of the cells (95%) showed MobBΔTMD located on a single pole in the cytosol at both tested times.
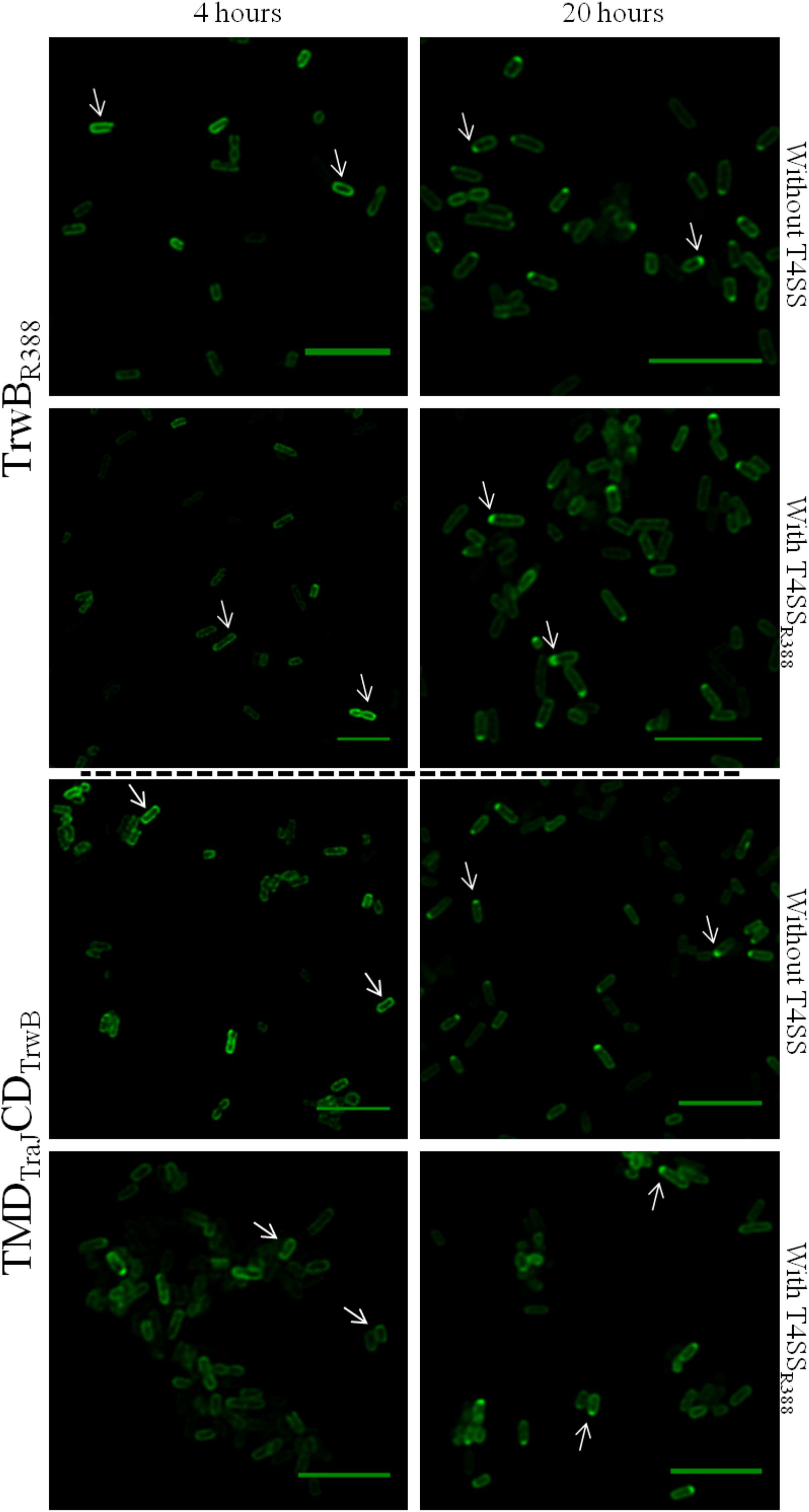
Figure 3. Subcellular location of TrwBR388GFP and TMDTraJCDTrwBGFP fusion-proteins by confocal fluorescence microscopy. Proteins were expressed in E. coli BL21C41 (DE3) strain by induction with 1 mM IPTG for 4 (left panels) and 20 h (right panels) at 25°C. Subcellular location of TrwBR388GFP and TMDTraJCDTrwBGFP was determined in E. coli strains containing plasmid pSU1456 that expresses all R388 conjugative proteins except TrwBR388, or without plasmid pSU1456. The images were acquired in a Leica TCS SP5 confocal fluorescence microscope, with a 60× oil immersion objective. Sample excitation was performed with 488 nm wavelength, while fluorescence emission was measured between 505 and 525 nm. The images were analyzed using Huygens and ImageJ softwares. Arrrowheads indicate the eGFP fluorescence through the periphery (1st column) and at a single cell pole (2nd column). Scale bar: 5 μm.
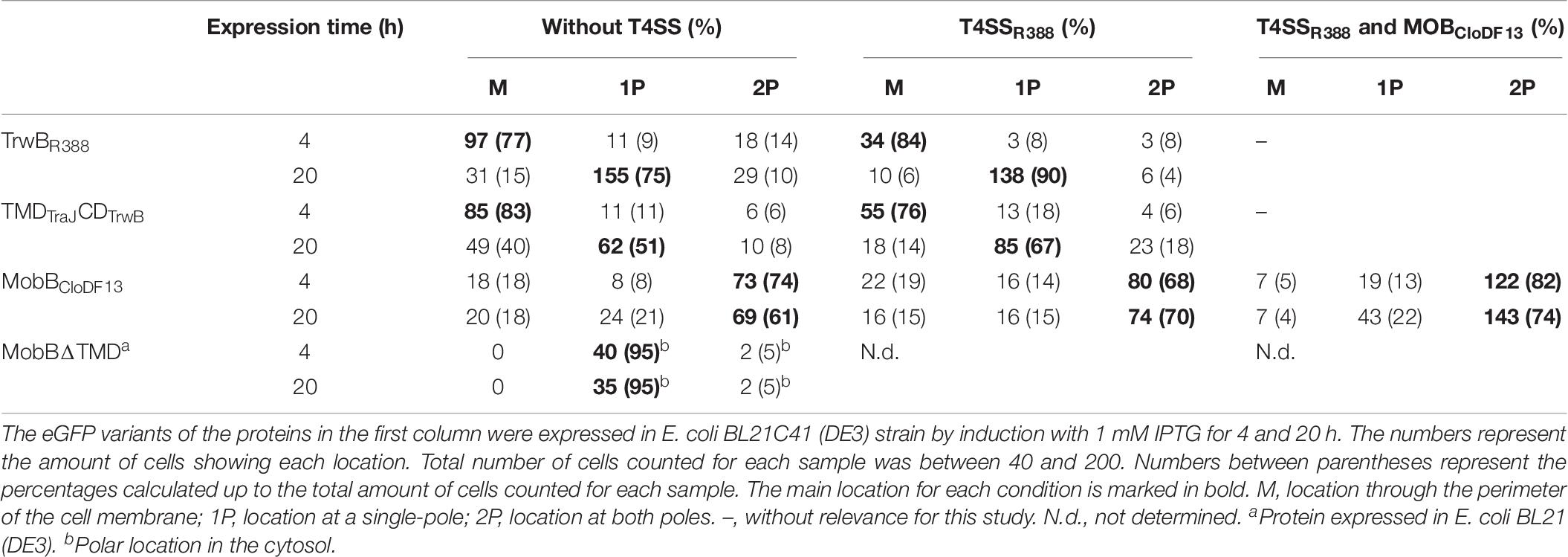
Table 6. Subcellular location of different T4CP-eGFP fusion-proteins at different expression times in the absence or presence of T4SSR388.
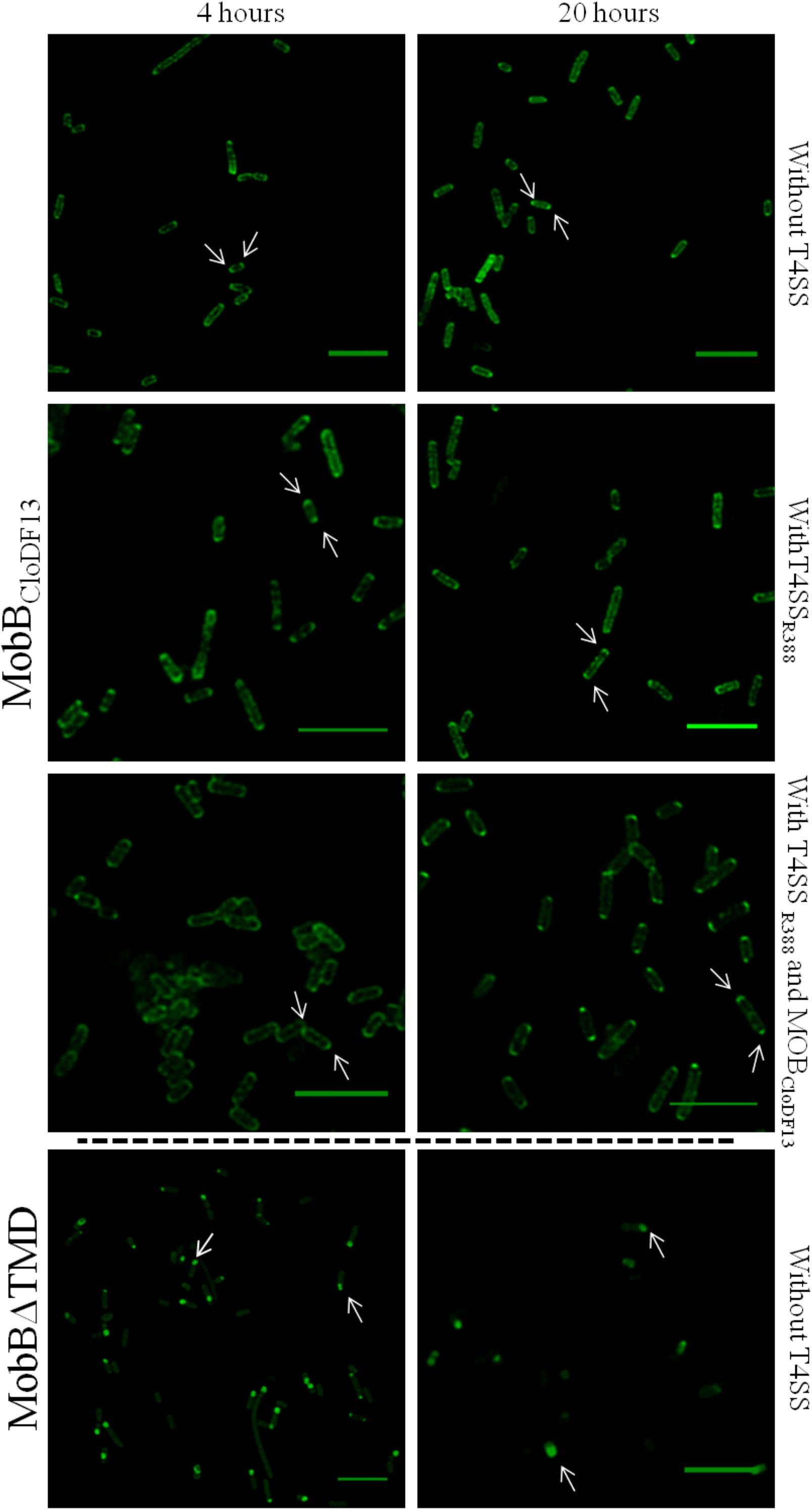
Figure 4. Subcellular location of MobBCloDF13GFP and MobBΔTMDGFP fusion-proteins by confocal fluorescence microscopy. MobBCloDF13GFP and MobBΔTMDGFP proteins were expressed in E. coli BL21C41 (DE3) and E. coli BL21 (DE3) strains, respectively. Subcellular location of these proteins was determined by induction with 1 mM IPTG after 4 (left panels) and 20 h (right panels) at 25°C. Additionally, subcelular location was analyzed in the presence of pSU1456 plasmid, which expresses all R388 conjugative proteins except TrwBR388, or without plasmid pSU1456. MobBCloDF13 was also expressed in the presence of both pSU1456 and pSU4833 that codes for the mobilization proteins of plasmid CloDF13 (MOBCloDF13), except for MobBCloDF13. The images were acquired in a Leica TCS SP5 confocal fluorescence microscope, with a 60× oil immersion objective. Sample excitation was performed with 488 nm wavelength, while fluorescence emission was measured between 505 and 525 nm. The images were analyzed using Huygens and ImageJ software. Arrrowheads indicate the eGFP fluorescence foci at both cell poles (MobBCloDF13) and at a single cell pole (MobBΔTMD). Scale bar: 5 μm.
Since the interaction with other conjugative proteins of the T4SS could modify the subcellular location pattern (Segura et al., 2014), the proteins that were active in vivo (i.e., TrwBR388, TMDTraJCDTrwB, and MobBCloDF13) were also observed in the presence of a T4SS lacking a functional T4CP that could interfere with the studied eGFP variant. On the one hand, TrwBR388 and TMDTraJCDTrwB were analyzed in the presence of the remaining conjugative proteins of R388 (i.e., in the presence of plasmid pSU1456 that lacks functional TrwBR388 but contains the remaining conjugative proteins). On the other hand, since CloDF13 needs the T4SS of a co-resident conjugative plasmid to be mobilized, the subcellular location of MobBCloDF13 was studied in the presence of R388 lacking functional TrwBR388 (plasmid pSU1456) and also in the presence of both, R388 lacking functional TrwBR388 (plasmid pSU1456) and the mobility region of CloDF13 lacking functional MobBCloDF13 (plasmid pSU4833) (Table 6). In these experiments (Figures 3, 4) the location pattern of each protein was the same in the absence or presence of a conjugative system (Table 6). However, although MobBCloDF13 kept two poles as its main location in all the tested conditions, in the presence of R388 lacking functional TrwBR388 it did not partially switch to a single-pole after 20 h, as in the absence of it.
The general observed pattern for all studied T4CPs was that the presence of T4SS enhanced the percentage of cells with the T4CP at the predominant location shown in the absence of the T4SS for each protein after 20 h (Figures 3, 4). Specifically, the percentage of cells showing TrwBR388 and TMDTraJCDTrwB at a single pole increased and so did the percentage of cells showing MobBCloDF13 at both poles. Moreover, regarding MobBCloDF13, the additional presence of its cognate mobilization region enhanced the effect produced by T4SSR388.
Discussion
The increase of multidrug-resistant bacteria has become one of the major health concerns in our society (World Health Organization [WHO], 2019), being bacterial conjugation one of the key mechanisms responsible for the spread of antibiotic resistance genes among bacteria (Bello-López et al., 2019). This process is performed through a T4SS, a multiprotein complex that transfers the nucleoprotein substrate from a donor into a recipient bacterium (Waksman, 2019). T4CPs are essential proteins during conjugation, as they connect the substrates to be transferred in the cytosol with the secretion machinery in the membrane (Gomis-Ruth et al., 2005). Despite their importance, as membrane proteins are challenging to be studied, their characterization has been mostly accomplished using mutants that lack their TMD (Schroder and Lanka, 2003; Tato et al., 2007; Larrea et al., 2017). However, several studies performed with TrwBR388, the full-length T4CP of the conjugative plasmid R388, have proven that the TMD is more than a mere anchor to the membrane and that it has a role in protein activity, stability, oligomerization and subcellular localization (Moncalián et al., 1999; Hormaeche et al., 2002, 2004, 2006; Vecino et al., 2010, 2011; Segura et al., 2013, 2014).
The aim of this work has been to provide new data about different T4CPs that will contribute to infer general conclusions on their functioning to develop strategies to inhibit them and control the spread of antibiotic resistance genes. To do so, we studied the in vivo functionality, secondary structure, thermal stability, and subcellular location of TrwBR388, its chimeric protein TMDTraJCDTrwB, and the T4CP of the mobilizable plasmid CloDF13, MobBCloDF13, and its TMD-less mutant, MobBΔTMD.
In vivo Functionality
TMDTraJCDTrwB was able to complement the ΔtrwB mutation for R388 transfer, although with a lower transfer frequency than TrwBR388. This decrease in conjugation frequency may be due to a combination of effects such as conformational changes in the CD of TrwBR388 due to its chimeric nature that render a less active protein and the heterologous interaction between TMDTraJ and the T4SS of R388 (Llosa et al., 2003). Thus, TMDTraJCDTrwB-mediated transfer of R388 would occur through a specific interaction between CDTrwB with its cognate relaxase, TrwCR388, and an unspecific interaction of TMDTraJ with the heterologous T4SS from R388. This could explain why TMDTraJCDTrwB did not complement the ΔtraJ mutation (pKM101ΔtraJ) since CDTrwB could not recognize the heterologous TraIpKM101 relaxase, even if TMDTraJ interacted with its cognate T4SSpKM101.
Moreover, TMDTraJCDTrwB showed negative dominance in the presence of the native T4CPs TrwBR388 or TraJpKM101. Since it has been reported that TMDTraJ can interact with both T4SSR388 and T4SSpKM101 (Llosa et al., 2003; De Paz et al., 2010; Celaya et al., 2017), a possible explanation for the observed negative dominance could be that competition for the T4SS occurred. According to this hypothesis, TMDTraJCDTrwB would have interacted with the secretion channel, sequestering it from interacting with the native T4CPs and reducing the conjugative rate of the wild type system. This hypothesis comes into agreement with the fact that the transfer of pKM101 using TraJpKM101 and T4SSR388 is one order of magnitude lower than using T4SSpKM101 (Llosa et al., 2003). Another explanation compatible with dominance experiments with both systems would be that non-functional heteroligomers were made between the native and the chimera proteins, competing for the conjugative machinery and therefore lowering the transfer frequency. Any of these alternatives or a combination of them would have caused a decrease in the plasmid transfer rate, as observed in the dominance mating assays. It must be underlined that a point mutation in the Walker A domain, TMDTraJCDTrwB (K142T), resulted in a non-functional phenotype. Hence, the K mutation totally arrested transfer capacity as previously reported for other T4CPs (Moncalián et al., 1999; Kumar and Das, 2002; Gunton et al., 2005), suggesting that concerning its NBD TMDTraJCDTrwB is functionally similar to TrwBR388 despite the results obtained in the complementation and dominance studies.
Regarding CloDF13 system, it was observed that the deletion of the TMD rendered a non-functional MobBΔTMD as it occurs with other TMD-less mutants such as TrwBΔN70 and PcfCΔN103 (Chen et al., 2008). However, a TcpA mutant lacking the TMD, but not the N-term cytosolic residues, TcpAΔ46–104, was able to perform conjugation, although at a frequency lower than the wild type plasmid. In this regard, unpublished experiments with TrwBΔN8, which lacks the N-term cytosolic eight residues, showed a decrease in the transfer rate of more than three orders of magnitude (Vecino, 2009). Taken together these results, it seems that not only the TMD but also the N-term cytosolic residues have an important role in the transfer capacity of T4CPs. In this context, the importance of this small region in specific interactions with the relaxosome has already been described (Llosa et al., 2003; Schroder and Lanka, 2003). In the case of MobBCloDF13 its N-term is located in the periplasm (Figure 1A), which could be an important feature for recognition and interaction with conjugative secretion channels.
Secondary Structure and Thermal Stability
As it has been described that the TMD influences the secondary structure of the CD and the thermal stability of TrwBR388 (Hormaeche et al., 2004; Vecino et al., 2011, 2012), in this work we have studied whether this behavior can be observed in TMDTraJCDTrwB and in MobBCloDF13.
Regarding TMDTraJCDTrwB, one of the most important differences in comparison with TrwBR388 was the appearance of a band at 1640 cm–1, mainly assigned to flexible structures (non-periodic elements) related to a less compact overall structure (Echabe et al., 1998; Agopian et al., 2016). As the crystal structure of TrwBΔN70 shows flexible loops (Gomis-Rüth et al., 2002), the presence of the 1640 cm–1 band present at similar percentages (30–37%) in TMDTraJCDTrwB, TrwBΔN50, and TrwBΔN70 mutant proteins but missing in TrwBR388, could be explained as the loss of the compact structure of TrwBR388 due to the deletion of its cognate TMD (Hormaeche et al., 2004).
Taking into account the values related to all the bands associated to β-sheet elements (1671, 1640, and 1626 cm–1) it could be concluded that the total percentage of β-sheets in TMDTraJCDTrwB is just slightly smaller to that of TrwBR388. Concerning β-turns, the band assigned to them in the TrwBR388 related proteins, 1661–1665 cm–1, was not observed in TMDTraJCDTrwB. However, it can be postulated that the increase seen in TMDTraJCDTrwB of the band at 1671 cm–1, was partially related to the β-turns component seen in the mutants at 1665 cm–1. These would imply that the β-turns component of the chimeric protein is lower than the one of the native protein and similar to the TMD deletion mutants. Finally, TMDTraJCDTrwB shows a decrease in α-helix percentage (35%) in comparison with TrwBR388 (41%), but an increase in comparison to the mutants (26%).
Therefore it seems that TMDTraJCDTrwB presents qualitative and quantitative features in between the native protein and the deletion mutants. These results suggest that the presence of a heterologous full-length TMD does provide a more compact and ordered structure to the T4CP in comparison to the TMD-less mutants, even if it does not reach the level of the native protein. This result comes in agreement with the transfer capacity reduction of the chimeric protein that could be explained partially by the observed structural changes reported here.
To test if the results described for TrwBR388 could be extrapolated to other T4CPs, the secondary structures of MobBCloDF13 and MobBΔTMD were studied (Table 5). Surprisingly, MobBΔTMD presented an IR spectrum quite similar to the one obtained for MobBCloDF13. It presented smaller helical structure percentages and higher β-turns percentages but both showed similar unordered and β-sheet percentages. This suggests that in MobBCloDF13 the presence of the TMD does not have an effect on the structure of the CD as it does in TrwBR388. As MobBCloDF13 has to interact with heterologous T4SSs, it could be that its TMD has to interact with heterologous T4SSs without altering the structure of its CD where the specific interaction with its cognate relaxosome occurs.
Concerning the thermal denaturation of TMDTraJCDTrwB, its Tm was similar to the ones described for TrwBR388 and the mutant lacking the first transmembrane helix, TrwBΔN50 (Table 5). This result suggests that although the secondary structures of the mutants differ from that of the native protein, their overall thermal stability is similar. Additionally, when comparing the results between both studied systems, as expected due to the high instability of purified membrane proteins (González Flecha, 2017) the soluble proteins showed higher mid-point denaturation temperatures than the full-length proteins. Specifically, MobBΔTMD and TrwBΔN70 showed similar Tm values (Tm 62 and 63°C, respectively); on the contrary, MobBCloDF13 was more stable than TrwBR388 against thermal denaturation (Tm 56 and 48°C, respectively). This could be related to different buffer compositions that had to be used when MobBCloDF13 and TrwBR388 were analyzed.
Subcellular Location
The polar location of proteins in bacteria underlines their sophisticated internal organization, being important in many processes like chemotaxis and cellular division (Howard, 2004). Similarly, subcellular location has been considered important in bacterial conjugation (Chen et al., 2008; Leonetti et al., 2015). Sequence analysis, cell fractionation, and protein purification experiments proved that TrwBR388, TMDTraJCDTrwB, and MobBCloDF13 are located in the bacterial membrane, while MobBΔTMD is located in the cytosol (data not shown). As studies in the literature do not show a consensus pattern either in the location of the T4CPS nor in the role of each domain in this property (Kumar and Das, 2002; Bauer et al., 2011; Segura et al., 2014), the subcellular location of TrwBR388, TMDTraJCDTrwB, MobBCloDF13, and MobBΔTMD was investigated.
In this work we observed TrwBR388 located along the membrane after 4 h of induction and only after 20 h it focused at single pole. These results differ from our previous results where the polar location of TrwBR388 was observed after 4 h (Segura et al., 2014). However, the induction OD600 values were different (0.4 vs. 0.7, this work and previous work, respectively), probably rendering populations at different growth phase. On the contrary MobBCloDF13 was located at both poles in the membrane at 4 and 20 h after induction.
Previous studies have reported that in the absence of other conjugative proteins, T4CPs that lacked the whole TMD or even the periplasmic loop located in the cytosol or at the membrane periphery, respectively (Kumar and Das, 2002; Segura et al., 2014). Moreover, the TMD alone of TrwBR388 located at the membrane poles without the need for the CD (Segura et al., 2014), suggesting a leading role for the TMD in the subcellular location of the T4CPs. Surprisingly, MobBΔTMD located at a single cell pole in the cytosol even in the absence of other conjugative proteins. These results suggest that mobilizable plasmid-related T4CPs could use different mechanisms than VirD4-type T4CPs for subcellular location. It has been speculated that the polar location of T4CPs could be related to interactions with the cardiolipin enriched membrane poles (Mileykovskaya and Dowhan, 2009; Segura et al., 2014), but at the same time mediated by complex and dynamic changes in transduction, cytoskeleton proteins, etc. (Shapiro et al., 2002). Since interactions between mobilizable plasmid-related T4CPs and T4SSs are not specific, these T4CPs could have evolved to develop different mechanisms to interact with the membrane and ensure their polar location.
Previous studies (Kumar and Das, 2002; Gunton et al., 2005; Segura et al., 2014) have reported that the location of native T4CPs is independent of the presence of the rest of the conjugative proteins. Our results suggest that the presence of a complete conjugative system (i.e., mobilizable region and secretion channel) seems to enhance the polar location of wild type T4CPs. Moreover, although the location of MobBCloDF13 at both poles was enhanced in the presence of T4SSR388, it was further enhanced when MOBCloDF13 was also present.
Taking all together, it seems that that no universal location patterns can be attributed to T4CPs. Nevertheless, three conclusions can be undertaken regarding subcellular location: (i) T4CPs localize either at a single pole or both poles, depending on the system; (ii) the presence of a TMD is not essential for the polar location of a mobilizable plasmid associated T4CP, and (iii) the presence of a conjugative system enhances the polar location of full-length T4CPs.
To sum up, the comparative study between the conjugative system related TrwBR388 and the mobilizable plasmid-related MobBCloDF13 and their variants has underlined that the characteristics described for the paradigmatic conjugative plasmid related VirD4-type T4CPs and their TMDs should not be ascribed to the whole T4CP family.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation, to any qualified researcher.
Author Contributions
IÁ-R, CG, and IAl contributed to the design of the work (text and figures) and the acquisition of the data, writing and revision of the content, approval of the last version, and ensuring accuracy and integrity of the work. IAr and JA contributed to acquisition of the data, revision of the content, and approval of the last version of the work. BU-U contributed to writing, revision of the content, and approval of the last version. All authors contributed to the article and approved the submitted version.
Funding
This work was in part supported by grants from the University of the Basque Country (GIU18/229 and COLAB19/08) and the Industry Department of the Basque Government (ELKARTEK 2020 KK-2020/00007). IÁ-R was a pre-doctoral student supported by the Basque Government.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to sincerely thank Dr. Lide Arana for her invaluable help and contribution to this work, to Unai Lorenzo for his help with the confocal microscope and image treatment, and to Dr. Louise Bird for her help with the high-throughput cloning at OPPF-UK.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmolb.2020.00185/full#supplementary-material
Footnotes
- ^ http://n2t.net/addgene:26043
- ^ http://n2t.net/addgene:41125
- ^ https://web.expasy.org/protparam/
- ^ http://topcons.cbr.su.se/
References
Agopian, A., Quetin, M., and Castano, S. (2016). Structure and interaction with lipid membrane models of Semliki Forest virus fusion peptide. Biochim. Biophys. Acta Biomembr. 1858, 2671–2680. doi: 10.1016/j.bbamem.2016.07.003
Agúndez, L., Zárate-Pérez, F., Meier, A. F., Bardelli, M., Llosa, M., Escalante, C. R., et al. (2018). Exchange of functional domains between a bacterial conjugative relaxase and the integrase of the human adeno-associated virus. PLoS One 13:e0200841. doi: 10.1371/journal.pone.0200841
Alvarez-Martinez, C. E., and Christie, P. J. (2009). Biological diversity of prokaryotic type IV secretion systems. Microbiol. Mol. Biol. Rev. 73, 775–808. doi: 10.1128/mmbr.00023-09
Andraka, N., Sánchez-Magraner, L., García-Pacios, M., Goñi, F. M., and Arrondo, J. L. R. (2017). The conformation of human phospholipid scramblase 1, as studied by infrared spectroscopy. Effects of calcium and detergent. Biochim. Biophys. Acta Biomembr. 1859, 1019–1028. doi: 10.1016/j.bbamem.2017.02.015
Arrondo, J. L., and Goñi, F. M. (1999). Structure and dynamics of membrane proteins as studied by infrared spectroscopy. Prog. Biophys. Mol. Biol. 72, 367–405. doi: 10.1016/s0079-6107(99)00007-3
Arrondo, J. L., Muga, A., Castresana, J., and Goñi, F. M. (1993). Quantitative studies of the structure of proteins in solution by Fourier-transform infrared spectroscopy. Prog. Biophys. Mol. Biol. 59, 23–56.
Bauer, T., Rösch, T., Itaya, M., and Graumann, P. L. (2011). Localization pattern of conjugation machinery in a gram-positive bacterium. J. Bacteriol. 193, 6244–6256. doi: 10.1128/jb.00175-11
Bello-López, J. M., Cabrero-Martínez, O. A., Ibáñez-Cervantes, G., Hernández-Cortez, C., Pelcastre-Rodríguez, L. I., Gonzalez-Avila, L. U., et al. (2019). Horizontal gene transfer and its association with antibiotic resistance in the genus Aeromonas spp. Microorganisms 7:363. doi: 10.3390/microorganisms7090363
Berrow, N. S., Alderton, D., Sainsbury, S., Nettleship, J., Assenberg, R., Rahman, N., et al. (2007). A versatile ligation-independent cloning method suitable for high-throughput expression screening applications. Nucleic Acids Res. 35, e45. doi: 10.1093/nar/gkm047
Bird, L. E. (2011). High throughput construction and small scale expression screening of multi-tag vectors in Escherichia coli. Methods 55, 29–37. doi: 10.1016/j.ymeth.2011.08.002
Bourg, G., Sube, R., O’Callaghan, D., and Patey, G. (2009). Interactions between Brucella suis VirB8 and its homolog TraJ from the plasmid pSB102 underline the dynamic nature of type IV secretion systems. J. Bacteriol. 191, 2985–2992. doi: 10.1128/jb.01426-08
Cabezón, E., Sastre, J. I, and de la Cruz, F. (1997). Genetic evidence of a coupling role for the TraG protein family in bacterial conjugation. Mol. Gen. Genet. 254, 400–406. doi: 10.1007/s004380050432
Celaya, G., Perales-Calvo, J., Muga, A., Moro, F., and Rodriguez-Larrea, D. (2017). Label-free, multiplexed, single-molecule analysis of protein-DNA complexes with nanopores. ACS Nano 11, 5815–5825. doi: 10.1021/acsnano.7b01434
Chen, Y., Zhang, X., Manias, D., Yeo, H.-J., Dunny, G. M., and Christie, P. J. (2008). Enterococcus faecalis PcfC, a Spatially Localized Substrate Receptor for Type IV Secretion of the pCF10 Transfer Intermediate. J. Bacteriol. 190, 3632–3645. doi: 10.1128/jb.01999-07
Cormack, B. P., Valdivia, R. H., and Falkow, S. (1996). FACS-Optimized Mutants of the Green Fluorescent Protein (GFP). Amsterdam: Elsevier, 33–38.
Datta, N., and Hedges, R. W. (1972). Trimethoprim resistance conferred by W plasmids in Enterobacteriaceae. J. Gen. Microbiol. 72, 349–355. doi: 10.1099/00221287-72-2-349
De Paz, H. D., Larrea, D., Zunzunegui, S., Dehio, C., De La Cruz, F., and Llosa, M. (2010). Functional dissection of the conjugative coupling protein TrwB. J. Bacteriol. 192, 2655–2669. doi: 10.1128/jb.01692-09
Drew, D., Lerch, M., Kunji, E., Slotboom, D.-J., and de Gier, J.-W. (2006). Optimization of membrane protein overexpression and purification using GFP fusions. Nat. Methods 3, 303–313. doi: 10.1038/nmeth0406-303
Drew, D., Slotboom, D.-J., Friso, G., Reda, T., Genevaux, P., Rapp, M., et al. (2005). A scalable, GFP-based pipeline for membrane protein overexpression screening and purification. Protein Sci. 14, 2011–2017. doi: 10.1110/ps.051466205
Echabe, I., Prado, A., Goñi, F. M., Arrondo, J. L. R., and Dornberger, U. (1998). Topology of sarcoplasmic reticulum Ca2 + -ATPase: an infrared study of thermal denaturation and limited proteolysis. Protein Sci. 7, 1172–1179. doi: 10.1002/pro.5560070511
Gomis-Ruth, F., Sola, M., Cruz, F., and Coll, M. (2005). Coupling factors in macromolecular type-IV secretion machineries. Curr. Pharm. Des. 10, 1551–1565. doi: 10.2174/1381612043384817
Gomis-Rüth, F. X., Moncalián, G., de la Cruz, F., and Coll, M. (2002). Conjugative plasmid protein TrwB, an integral membrane type IV secretion system coupling protein. J. Biol. Chem. 277, 7556–7566. doi: 10.1074/jbc.m110462200
Gomis-Rüth, F. X., Moncalián, G., Pérez-Luque, R., González, A., Cabezón, E., de la Cruz, F., et al. (2001). The bacterial conjugation protein TrwB resembles ring helicases and F1-ATPase. Nature 409, 637–641. doi: 10.1038/35054586
González Flecha, F. L. (2017). Kinetic stability of membrane proteins. Biophys. Rev. 9, 563–572. doi: 10.1007/s12551-017-0324-0
Gunton, J. E., Gilmour, M. W., Alonso, G., and Taylor, D. E. (2005). Subcellular localization and functional domains of the coupling protein, TraG, from IncHI1 plasmid R27. Microbiology 151, 3549–3561. doi: 10.1099/mic.0.28255-0
Hamilton, C. M., Lee, H., Li, P.-L., Cook, D. M., Piper, K. R., von Bodman, S. B., et al. (2000). TraG from RP4 and TraG and VirD4 from Ti plasmids confer relaxosome specificity to the conjugal transfer system of pTiC58. J. Bacteriol. 182, 1541–1548. doi: 10.1128/jb.182.6.1541-1548.2000
Hormaeche, I., Alkorta, I., Moro, F., Valpuesta, J. M., Goni, F. M., and De La Cruz, F. (2002). Purification and properties of TrwB, a hexameric, ATP-binding integral membrane protein essential for R388 plasmid conjugation. J. Biol. Chem. 277, 46456–46462. doi: 10.1074/jbc.m207250200
Hormaeche, I., Iloro, I., Arrondo, J. L. R., Goñi, F. M., De La Cruz, F., and Alkorta, I. (2004). Role of the transmembrane domain in the stability of TrwB, an integral protein involved in bacterial conjugation. J. Biol. Chem. 279, 10955–10961. doi: 10.1074/jbc.m310422200
Hormaeche, I., Segura, R. L., Vecino, A. J., Goñi, F. M., de la Cruz, F., and Alkorta, I. (2006). The transmembrane domain provides nucleotide binding specificity to the bacterial conjugation protein TrwB. FEBS Lett. 580, 3075–3082. doi: 10.1016/j.febslet.2006.04.059
Howard, M. (2004). A mechanism for polar protein localization in bacteria. J. Mol. Biol. 335, 655–663. doi: 10.1016/j.jmb.2003.10.058
Kumar, R. B., and Das, A. (2002). Polar location and functional domains of the Agrobacterium tumefaciens DNA transfer protein VirD4. Mol. Microbiol. 43, 1523–1532. doi: 10.1046/j.1365-2958.2002.02829.x
Langer, P. J., and Walker, G. C. (1981). Restriction endonuclease cleavage map of pKM101: relationship to parental plasmid R46. Mol. Gen. Genet. 182, 268–272. doi: 10.1007/BF00269669
Larrea, D., de Paz, H. D., Matilla, I., Guzmán-Herrador, D. L., Lasso, G., de la Cruz, F., et al. (2017). Substrate translocation involves specific lysine residues of the central channel of the conjugative coupling protein TrwB. Mol. Genet. Genomics 292, 1037–1049. doi: 10.1007/s00438-017-1331-3
Leonetti, C. T., Hamada, M. A., Laurer, S. J., Broulidakis, M. P., Swerdlow, K. J., Lee, C. A., et al. (2015). Critical components of the conjugation machinery of the integrative and conjugative element ICEBs1 of Bacillus subtilis. J. Bacteriol. 197, 2558–2567. doi: 10.1128/jb.00142-15
Llosa, M., and Alkorta, I. (2017). Coupling proteins in type IV secretion. Curr. Top. Microbiol. Immunol. 413, 143–168. doi: 10.1007/978-3-319-75241-9_6
Llosa, M., Bolland, S., and de la Cruz, F. (1994). Genetic organization of the conjugal DNA processing region of the IncW plasmid R388. J. Mol. Biol. 235, 448–464. doi: 10.1006/jmbi.1994.1005
Llosa, M., Zunzunegui, S., and de la Cruz, F. (2003). Conjugative coupling proteins interact with cognate and heterologous VirB10-like proteins while exhibiting specificity for cognate relaxosomes. Proc. Natl. Acad. Sci. U.S.A. 100, 10465–10470. doi: 10.1073/pnas.1830264100
Matilla, I., Alfonso, C., Rivas, G., Bolt, E. L., de la Cruz, F., and Cabezon, E. (2010). The conjugative DNA translocase TrwB is a structure-specific DNA-binding protein. J. Biol. Chem. 285, 17537–17544. doi: 10.1074/jbc.m109.084137
Mileykovskaya, E., and Dowhan, W. (2009). Cardiolipin membrane domains in prokaryotes and eukaryotes. Biochim. Biophys. Acta Biomembr. 1788, 2084–2091. doi: 10.1016/j.bbamem.2009.04.003
Moncalián, G., Cabezón, E., Alkorta, I., Valle, M., Moro, F., Valpuesta, J. M., et al. (1999). Characterization of ATP and DNA binding activities of TrwB, the coupling protein essential in plasmid R388 conjugation. J. Biol. Chem. 274, 36117–36124. doi: 10.1074/jbc.274.51.36117
Nuñez, B., and de la Cruz, F. (2001). Two atypical mobilization proteins are involved in plasmid CloDF13 relaxation. Mol. Microbiol. 39, 1088–1099. doi: 10.1046/j.1365-2958.2001.02308.x
Parsons, J. A., Bannam, T. L., Devenish, R. J., and Rood, J. I. (2007). TcpA, an FtsK/SpoIIIE homolog, is essential for transfer of the conjugative plasmid pCW3 in Clostridium perfringens. J. Bacteriol. 189, 7782–7790. doi: 10.1128/jb.00783-07
Paterson, E. S., Moré, M. I., Pillay, G., Cellini, C., Woodgate, R., Walker, G. C., et al. (1999). Genetic analysis of the mobilization and leading regions of the IncN plasmids pKM101 and pCU1. J. Bacteriol. 181, 2572–2583. doi: 10.1128/jb.181.8.2572-2583.1999
Redzej, A., Ukleja, M., Connery, S., Trokter, M., Felisberto-Rodrigues, C., Cryar, A., et al. (2017). Structure of a VirD4 coupling protein bound to a VirB type IV secretion machinery. EMBO J. 36, 3080–3095. doi: 10.15252/embj.201796629
Schroder, G., and Lanka, E. (2003). TraG-like proteins of type IV secretion systems: functional dissection of the multiple activities of TraG (RP4) and TrwB (R388). J. Bacteriol. 185, 4371–4381. doi: 10.1128/jb.185.15.4371-4381.2003
Segura, R. L., Águila-Arcos, S., Ugarte-Uribe, B., Vecino, A. J., De La Cruz, F., Goñi, F. M., et al. (2013). The transmembrane domain of the T4SS coupling protein TrwB and its role in protein-protein interactions. Biochim. Biophys. Acta Biomembr. 1828, 2015–2025. doi: 10.1016/j.bbamem.2013.05.022
Segura, R. L., Águila-Arcos, S., Ugarte-Uribe, B., Vecino, A. J., de la Cruz, F., Goñi, F. M., et al. (2014). Subcellular location of the coupling protein TrwB and the role of its transmembrane domain. Biochim. Biophys. Acta Biomembr. 1838, 223–230. doi: 10.1016/j.bbamem.2013.08.016
Shapiro, L., McAdams, H. H., and Losick, R. (2002). Generating and exploiting polarity in bacteria. Science 298, 1942–1946. doi: 10.1126/science.1072163
Smillie, C., Garcillán-Barcia, M. P., Francia, M. V., Rocha, E. P. C., and de la Cruz, F. (2010). Mobility of plasmids. Microbiol. Mol. Biol. Rev. 74, 434–452.
Tato, I., Matilla, I., Arechaga, I., Zunzunegui, S., de la Cruz, F., and Cabezon, E. (2007). The ATPase activity of the DNA transporter TrwB is modulated by protein TrwA: implications for a common assembly mechanism of DNA translocating motors. J. Biol. Chem. 282, 25569–25576.
Tato, I., Zunzunegui, S., de la Cruz, F., and Cabezon, E. (2005). TrwB, the coupling protein involved in DNA transport during bacterial conjugation, is a DNA-dependent ATPase. Proc. Natl. Acad. Sci. U.S.A. 102, 8156–8161. doi: 10.1073/pnas.0503402102
Tsirigos, K. D., Peters, C., Shu, N., Käll, L., and Elofsson, A. (2015). The TOPCONS web server for consensus prediction of membrane protein topology and signal peptides. Nucleic Acids Res. 43, W401–W407.
Vecino, A. J. (2009). La Reconstitución en Proteoliposomas de TrwB, un Nanomotor que Transporta DNA, Revela la Importancia del Dominio Transmembrana de la Proteína. Ph.D. thesis. País Vasco: University of the Basque Country.
Vecino, A. J., de la Arada, I., Segura, R. L., Goñi, F. M., de la Cruz, F., Arrondo, J. L. R., et al. (2011). Membrane insertion stabilizes the structure of TrwB, the R388 conjugative plasmid coupling protein. Biochim. Biophys. Acta 1808, 1032–1039. doi: 10.1016/j.bbamem.2010.12.025
Vecino, A. J., de Segura, R. L., de la Arada, I., de la Cruz, F., Goñi, F. M., Arrondo, J. L., et al. (2012). Deletion of a single helix from the transmembrane domain causes large changes in membrane insertion properties and secondary structure of the bacterial conjugation protein TrwB. Biochim. Biophys. Acta Biomembr. 1818, 3158–3166. doi: 10.1016/j.bbamem.2012.08.015
Vecino, A. J., Segura, R. L., Ugarte-Uribe, B., Aguila, S., Hormaeche, I., de la Cruz, F., et al. (2010). Reconstitution in liposome bilayers enhances nucleotide binding affinity and ATP-specificity of TrwB conjugative coupling protein. Biochim. Biophys. Acta 1798, 2160–2169. doi: 10.1016/j.bbamem.2010.07.005
Waksman, G. (2019). From conjugation to T4S systems in Gram-negative bacteria: a mechanistic biology perspective. EMBO Rep. 20:e47012.
Waldo, G. S., Standish, B. M., Berendzen, J., and Terwilliger, T. C. (1999). Rapid protein-folding assay using green fluorescent protein. Nat. Biotechnol. 17, 691–695. doi: 10.1038/10904
Whitaker, N., Berry, T. M., Rosenthal, N., Gordon, J. E., Gonzalez-Rivera, C., Sheehan, K. B., et al. (2016). Chimeric coupling proteins mediate transfer of heterologous type IV effectors through the Escherichia coli pKM101-encoded conjugation machine. J. Bacteriol. 198, 2701–2718. doi: 10.1128/jb.00378-16
Keywords: coupling proteins, type IV secretion systems, bacterial conjugation, membrane proteins, antibiotic resistance spread
Citation: Álvarez-Rodríguez I, Ugarte-Uribe B, de la Arada I, Arrondo JLR, Garbisu C and Alkorta I (2020) Conjugative Coupling Proteins and the Role of Their Domains in Conjugation, Secondary Structure and in vivo Subcellular Location. Front. Mol. Biosci. 7:185. doi: 10.3389/fmolb.2020.00185
Received: 28 May 2020; Accepted: 14 July 2020;
Published: 11 August 2020.
Edited by:
Tatiana Venkova, Fox Chase Cancer Center, United StatesReviewed by:
Elisabeth Grohmann, Beuth Hochschule für Technik Berlin, GermanyGraciela Castro Escarpulli, National School of Biological Sciences, National Polytechnic Institute of Mexico (IPN), Mexico
Copyright © 2020 Álvarez-Rodríguez, Ugarte-Uribe, de la Arada, Arrondo, Garbisu and Alkorta. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Itziar Alkorta, aXR6aS5hbGtvcnRhQGVodS5ldXM=
†Present address: Begoña Ugarte-Uribe, EMBL Heidelberg Cell Biology and Biophysics, Heidelberg, Germany