1
Department of Behavioural and Molecular Neuroendocrinology, University of Regensburg, Regensburg, Germany
2
Department of Psychology, Center for Neuroendocrine Studies, University of Massachusetts Amherst, Amherst, MA, USA
Psychopathologies such as anxiety- and depression-related disorders are often characterized by impaired social behaviours including excessive aggression and violence. Excessive aggression and violence likely develop as a consequence of generally disturbed emotional regulation, such as abnormally high or low levels of anxiety. This suggests an overlap between brain circuitries and neurochemical systems regulating aggression and anxiety. In this review, we will discuss different forms of male aggression, rodent models of excessive aggression, and neurobiological mechanisms underlying male aggression in the context of anxiety. We will summarize our attempts to establish an animal model of high and abnormal aggression using rats selected for high (HAB) vs. low (LAB) anxiety-related behaviour. Briefly, male LAB rats and, to a lesser extent, male HAB rats show high and abnormal forms of aggression compared with non-selected (NAB) rats, making them a suitable animal model for studying excessive aggression in the context of extremes in innate anxiety. In addition, we will discuss differences in the activity of the hypothalamic–pituitary–adrenal axis, brain arginine vasopressin, and the serotonin systems, among others, which contribute to the distinct behavioural phenotypes related to aggression and anxiety. Further investigation of the neurobiological systems in animals with distinct anxiety phenotypes might provide valuable information about the link between excessive aggression and disturbed emotional regulation, which is essential for understanding the social and emotional deficits that are characteristic of many human psychiatric disorders.
Every year, more than 700,000 people worldwide die because of assault (Bartolomeos et al., 2007
), and many more become victims of aggressive behaviour including domestic violence, terrorism, and hooliganism, or get physically or psychologically injured. Besides the suffering of the affected persons and their families, a large financial burden for the society emerges. Several psychiatric disorders like depression (Fava, 1998
; Mammen et al., 2002
), anxiety disorders, antisocial personality disorder (Eronen et al., 1998
; Moran, 1999
), post-traumatic stress disorder (PTSD; Beckham and Moore, 2000
; Ohayon and Shapiro, 2000
), borderline syndrome (Sanislow et al., 2000
; Skodol et al., 2002
), and alcohol abuse (Giancola et al., 2009
) were associated not only with severe emotional disturbances, but also with impaired social functioning, like social withdrawal, impaired social cognition, and excessive aggression (for review see Kohn and Asnis, 2003
; Swann, 2003
). This is not surprising, given the fact that the neural circuits regulating emotions and social behaviours are highly interconnected. In fact, deficits in social behaviour leading to excessive aggression may develop as a consequence of disturbed emotional regulation (Davidson et al., 2000
). Accordingly, a better understanding of the link between social behaviours and emotional regulation and their neurobiological underpinnings is essential to improve the treatment of many psychopathologies. In this review, we will discuss different forms of male aggression, rodent models of high and/or abnormal aggression, and neurobiological mechanisms underlying male aggression in the context of anxiety. In particular, we will focus on our own attempts to reveal a possible association between male aggression and innate anxiety using rats selectively bred for high (HAB) and low (LAB) anxiety-related behaviour.
Two main forms of human excessive/violent aggression are described, namely impulsive-reactive-hostile-affective aggression and controlled-proactive-instrumental-predatory aggression (Vitiello and Stoff, 1997
). Whereas the first form of aggression is seen in patients with depression, PTSD, or intermittent explosive disorder, the second type of aggression can be revealed in patients with personality disorders (conduct, antisocial, and borderline disorder), but may also be found in individuals without noticeable emotional or social deficits (i.e. the “friendly neighbour” or the “polite and quiet young man”). In particular, cases of violent and criminal behaviour displayed by these “unlikely” individuals are difficult to understand and indicate the urgent necessity to reveal underlying neurobiological mechanisms of abnormal aggression in more detail. Both forms of excessive aggression are associated with distinct disturbances in the regulation of emotions and alterations in autonomic and neuroendocrine functioning. More specifically, impulsive-reactive-hostile-affective aggression is generally associated with high emotional reactions and autonomic responses including high glucocorticoid levels, whereas controlled-proactive-instrumental-predatory aggression is generally associated with low emotional, autonomic, and glucocorticoid responses, and low skin conductance (summarized in Haller and Kruk, 2006
). We hypothesize that the emotional responses underlying the two distinct forms of aggression are linked to differences in anxiety-mediating brain pathways.
Species-specific signals transmitting the emotional state of an individual are very prominent in humans and animals living in herds, cohorts, or any other kind of social community. The expression of emotions involving aggression and anxiety-related behaviours is essential for effective communication between individuals within a society.
Male Aggression
Male aggression is necessary for the acquisition and maintenance of nutrition, territory, and mating partners. Species-specific rules have to be strictly obeyed to guarantee effective and harmless communication. Thus, adaptive offensive aggression is comprised primarily of harmless threat behaviours allowing the opponent to escape or to switch to submissive behaviours in order to avoid direct physical confrontation. In rodents, such signs of offensive aggression include piloerection (intimidation of the opponent by larger appearance) and lateral threat (arched back and exposure of the flank). In case of an offensive attack, less vulnerable body parts of the opponent, such as those covered with muscles and a thick layer of skin, are targeted to avoid serious injuries (Blanchard and Blanchard, 1977
; Blanchard et al., 2003
). While offensive aggression is usually expressed during a fight for territory or exclusive mating, defensive aggression is mainly displayed in life-threatening situations and is linked to increased fear (Blanchard and Blanchard, 1981
). As opposed to offensive aggression, defensive aggression is less or not signalled in advance, and attack targets include more vulnerable body parts (such as the head, belly, and genitals) (Blanchard and Blanchard, 1977
; Blanchard et al., 2003
).
Abnormal Aggression
Although offensive aggression is adaptive and essential for reproductive success, it can become harmful and turn into abnormal forms of aggression when an individual (i) disregards species-specific rules, (ii) becomes insensitive towards social signals of the opponent (e.g. ignores submissiveness and continues attacking), or (iii) has a mismatched response to a provocation (Haller and Kruk, 2006
). In many species, attacks directed by a male towards female conspecifics or towards vulnerable body parts of a male opponent in a non life-threatening situation are considered to be abnormal (Haller et al., 2001
). Although abnormal forms of aggression are rarely seen in animals living in their natural environment, several factors may increase the appearance of exaggerated forms of aggression such as housing under stressful conditions, acute exposure to stress, and genetic risk factors (see also the sections Acute Interventions, Transgenic Mice, Early Life Stress, Selective Breeding).
The Resident-Intruder Test: A Test for Male Aggression
The resident-intruder (RI) test has been designed and validated for rodents to study offensive male aggression under standardized laboratory conditions (Koolhaas et al., 1980
). In preparation for the RI test, the experimental male rat is typically housed with a female for several days or weeks to stimulate territorial aggression (Flannelly and Lore, 1977
). The test is performed in the absence of the female in the early dark phase of the light/dark cycle, i.e. at a time of heightened activity. The resident male rat is confronted with a slightly smaller, unfamiliar male intruder mostly of the same strain, which is placed into the resident’s home-cage. Multiple aggression-related parameters such as the latency to the first attack and the frequency and duration of aggressive behaviours displayed by the resident as well as defensive or exploratory behaviour of the intruder can be quantified.
Anxiety-Related Behaviour
Anxiety may be interpreted as an emotional anticipation of an aversive situation and is reflected by species-specific behavioural fear responses to stressful and threatening stimuli characteristic for individual trait anxiety. Fear is not seen as basal state (as is anxiety), but as a complex behavioural response, such as startle or freezing. Further, in addition to factors which determine innate (trait) anxiety, several environmental or pharmacological factors may interact with the genetic background and determine the individual level of state anxiety and the final behavioural phenotype. Emotionality, often used as synonym for anxiety as well as fearfulness, may be seen in a broader sense, comprising both trait and state anxiety and stimulus-related fear. Emotionality is one of the major components underlying the ability of an organism to assess stressful stimuli and scenarios, and to adequately cope with them.
Although adequate levels of trait anxiety and subsequent fear responses are beneficial for an individual, exaggerated and uncontrolled forms of anxiety and fear are maladaptive in both animals and humans. The lifetime prevalence of anxiety disorders, like panic disorder or general anxiety disorder, is about 17% (Somers et al., 2006
) with social anxiety disorder, like social phobia, being the most common one (Kessler et al., 1994
). A variety of rodent models has been established in order to reveal the complex brain circuitries, and neuronal and genetic mechanisms underlying normal and extreme forms of trait or state anxiety (Gordon and Hen, 2004
; Bourin et al., 2007
). Such rodent models include selective breeding of rats or mice to augment naturally occurring differences in anxiety (Landgraf and Wigger, 2002
; Bunck et al., 2009
), exposure to early life stress (Wigger and Neumann, 1999
; Huot et al., 2001
), or to chronic stress in adulthood (Barrot et al., 2005
; Reber et al., 2007
), transgenic modification (Bale et al., 2000
; Mantella et al., 2003
; Müller et al., 2003
), and acute interventions that alter state anxiety (Neumann et al., 2000
; Millan, 2003
; Slattery and Neumann, 2010
). In this review, we will mainly focus on our model of genetic selection for differences in trait anxiety, i.e. on HAB and LAB rats.
For the quantification of anxiety-related behaviour in rodents several laboratory tests have been successfully validated (for review see File et al., 2004
). The black-white box or the elevated plus-maze (EPM), for example, are the most frequently used non-conditioned tests for anxiety based on the conflict between the rat’s exploratory drive and its innate fear of bright and open spaces (Crawley and Goodwin, 1980
). A high level of anxiety is expressed by avoidance of these aversive zones, i.e. the open, bright or unprotected compartments/arms of the test apparatus, which can be validated with established anxiolytic as well as anxiogenic substances (Pellow et al., 1985
).
Link between Aggression and Anxiety
Findings from both preclinical and clinical studies concerning the link between anxiety and aggression are conflicting, as anxiolytic drugs, for example, can reduce or potentiate male aggressive behaviour in several species including rodents (Malick, 1978
; Wongwitdecha and Marsden, 1996
; Kudryavtseva and Bondar, 2002
; Haller et al., 2007
). Besides their anxiolytic, tranquilizing and sedative effects, benzodiazepines generally reduce aggressive behaviour in humans, although, they may induce aggression in certain patients suffering from panic or conduct disorders (DiMascio, 1973
; Bond et al., 1995
; Cherek and Lane, 2001
). Other drugs used for the treatment of aggression-related disorders exert their effects via the dopaminergic or the serotonergic systems, which are both known to affect anxiety (Millan, 2003
; de Almeida et al., 2005
). Thus, although co-regulation of aggression and anxiety is likely based on the involvement of overlapping brain pathways, their complex interplay is far from being clear due to the limited availability of relevant animal models combining aggression and anxiety.
Animal models are essential for revealing neurobiological mechanisms of complex behaviours like anxiety, depression, and aggression and their associated psychopathologies. Due to the tendency towards an increase in excessive aggression and violence in industrialized societies, there is an urgent need to understand these abnormal forms of social behaviour in more detail and, subsequently, to develop more efficient therapeutic strategies. Despite a high need, and in contrast to rodent models of anxiety and depression, it is remarkable that the number of rodent models of aggression in combination with anxiety is extremely limited. Here, we briefly describe potential rodent models characterized by alterations in both aggression and anxiety-related behaviours.
Acute Interventions
Several acute manipulations have been established to induce a state of hyper-aggression or abnormal aggression, such as frustration (deprivation of an expected reward), instigation (pre-exposure to a possible opponent), alcohol (Miczek et al., 2003
; de Almeida et al., 2005
), glucocorticoid-deficiency (Haller et al., 2001
), and acute pharmacological approaches (de Almeida et al., 2005
; Ferris et al., 2006
). In some of these models, both aggression and anxiety-related behaviour were altered, making these acute models suitable for further detailed investigation into the link between aggression and anxiety.
Transgenic Mice
Our understanding of the impact of genetic factors shaping aggression has been significantly improved by the use of transgenic animal models. Reduced offensive aggression has been found in male knockout mice lacking the long form of the dopamine D2 receptor (Vukhac et al., 2001
), the α-isoform of the oestrogen receptor (Ogawa et al., 1997
; Scordalakes and Rissman, 2003
), the arginine vasopressin (AVP) V1b receptor (Wersinger et al., 2002
), or the enzyme dopamine β-hydroxylase (Marino et al., 2005
). In contrast, elevated offensive aggression is seen in mice lacking serotonin (5-HT) 1B receptors (Saudou et al., 1994
), dopamine transporters (Rodriguiz et al., 2004
), nitric oxide synthase (Nelson et al., 1995
), or monoamine oxidase A (Cases et al., 1995
). Interestingly, in most of these transgenic animals, concurrent alterations in anxiety and/or stress responsiveness were found, indicating a link between anxiety and aggressive responses. For example, mice lacking the regulators of G protein signalling 2 show a low level of aggression and a high level of anxiety (Oliveira-Dos-Santos et al., 2000
). In contrast, the elevated levels of aggression found in monoamine oxidase A knockout mice is accompanied by a lower level of anxiety-related behaviour (Cases et al., 1995
). Enkephalin knockout mice (König et al., 1996
) and α-calcium-calmodulin kinase II knockout mice (Chen et al., 1994
) show an increase in both aggression and anxiety/fear, whereas male oxytocin knockout mice have low levels of both aggression and anxiety (DeVries et al., 1997
; Winslow et al., 2000
). However, there are a few studies suggesting that anxiety and aggression can be altered independently. For example, male mice lacking the AVP V1b receptor are less aggressive than wild-type mice without a change in anxiety-related behaviour (Wersinger et al., 2002
).
Early Life Stress
Early life stress (e.g. emotional neglect, loss of parents, child abuse) is a major risk factor for the development of anxiety disorders and depression as well as excessive aggression and violence in adulthood (Dodge et al., 1990
; Loeber and Stouthamer-Loeber, 1998
; Heim and Nemeroff, 2001
; Barnow and Freyberger, 2003
). This has been confirmed by several rodent models of early life stress, including prenatal and postnatal stress, which have been – in the first instance – developed for studying neurobiological mechanisms of anxiety- and depression-related disorders (Levine et al., 1991
; Suchecki and Palermo Neto, 1991
; Plotsky and Meaney, 1993
; Rice et al., 2008
; Veenema, 2009
). Generally, early life stress in animals enhances anxiety- and depression-like behaviour, and is accompanied by functional changes of the hypothalamic–pituitary–adrenal (HPA) axis, one of the major stress systems of the organism. In addition, early life stress alters social and aggressive behaviours as well (Veenema et al., 2006
, 2007b
; Toth et al., 2008
; Veenema and Neumann, 2009
). The fact that concomitant changes in both anxiety/fear-related responses and aggression are a consequence of negative early life experiences further underscores the close interplay between aggression and anxiety. Therefore, early life stress is an important model in the study of mechanisms of excessive aggressive behaviour and their link to anxiety.
Selective Breeding
Breeding for a selected behavioural phenotype is an important approach to reveal neurochemical correlates of aggressive behaviour. Several lines of mice have been selected for differences in attack latency, including the SAL and LAL mice (van Oortmerssen and Bakker, 1981
), the Turku aggressive and non-aggressive mice (Sandnabba, 1996
), and the North Carolina 900 and 100 mice (Gariepy et al., 1996
). Among these selection lines, the SAL mice show the most excessive and abnormal forms of aggression (Sluyter et al., 2003
; Haller and Kruk, 2006
; Natarajan et al., 2009
). However, a significant difference in anxiety-related behaviours between SAL and LAL mice has not been consistently found (Hogg et al., 2000
; Veenema et al., 2003a
). In contrast, high aggression in the Turku mice is associated with low levels of anxiety (Nyberg et al., 2003
), whereas high aggression in the North Carolina mice is accompanied by high levels of anxiety (Nehrenberg et al., 2009
). Interestingly, both aggression and anxiety were reduced by acute diazepam treatment in the aggressive North Carolina 900 mice (Nehrenberg et al., 2009
). These examples demonstrate that selective breeding for aggression can result in concomitant changes in anxiety-related behaviours, albeit in different directions.
In rats, selective breeding for aggression is less common. The Novosibirsk Norway rats are selected for high and low aggression towards humans and, therefore, differ in defensive, rather than offensive, aggression (Naumenko et al., 1989
; Plyusnina and Oskina, 1997
). Nevertheless, this model is of interest as the high aggressive Novosibirsk rats show a high level of anxiety (Popova et al., 1999
). In an out-bred laboratory strain of wild-type rats, a wide range of aggressive behaviours was observed, although individuals with extremes in aggressive behaviour did not differ in anxiety-related behaviours (de Boer et al., 2003
).
In our laboratory, we selectively breed HAB and LAB Wistar rats for differences in trait anxiety as opposed to aggression. These rat lines have been extensively characterized in terms of emotionality and general stress coping strategies, as well as the underlying brain and neuroendocrine mechanisms (Landgraf and Wigger, 2002
; Landgraf et al., 2007
; Veenema and Neumann, 2007
). Here, we will present evidence that the HAB and LAB lines provide a promising model for studying brain mechanisms of high and abnormal forms of aggression and their link to trait anxiety.
HAB and LAB rats were originally bred for extremes in anxiety-related behaviour with the primary goal of establishing a robust behavioural phenotype (HAB) that mimics as many symptoms of human anxiety- (and depression-) related disorders as possible. This approach enabled us to reveal neuroendocrine, neuronal, and neurogenetic parameters directly linked to differences in trait anxiety. HAB and LAB rats are particularly useful as they (i) allow the identification of intracerebral pathways underlying trait anxiety, (ii) show differential sensitivity to anxiolytic/antidepressive substances with high responsiveness in HAB and almost no responsiveness in LAB rats (Keck et al., 2003
, 2005
), and (iii) enable the identification of candidate genes and gene polymorphisms underlying trait anxiety (Murgatroyd et al., 2004
; Landgraf et al., 2007
). Over the last several years, we have used HAB and LAB rats to identify possible links between innate anxiety and various aspects of complex social behaviours including social interaction (Henniger et al., 2000
), maternal care (Neumann et al., 2005a
; Bosch and Neumann, 2008
), maternal aggression (Bosch et al., 2005
), and intermale aggression (Beiderbeck et al., 2007
; Veenema et al., 2007a
). During the course of these studies, the male LAB rat has emerged as an interesting model for abnormal social behaviour, including excessive aggression (Veenema and Neumann, 2007
) (Figure 1
).
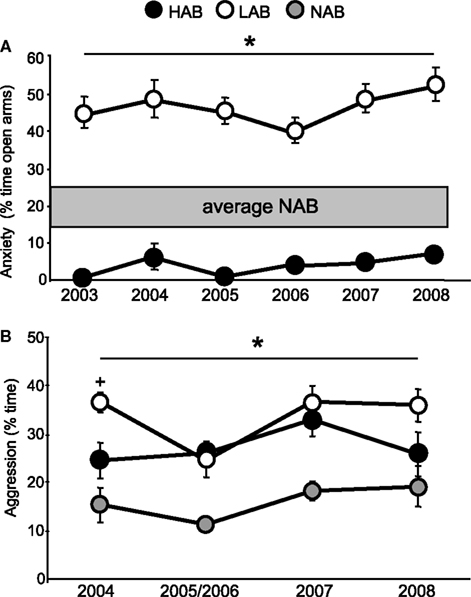
Figure 1. (A) Anxiety-related behaviour on the elevated plus-maze in 9-week-old HAB and LAB rats between 2003 and 2008; *p < 0. 05 LAB vs. HAB rats (2003–2008); (B) Aggressive behaviour during the RI test in 16- to 22-week-old male HAB, LAB, and NAB rats between 2004 and 2008. *p < 0.05 NAB vs. HAB (2005–2007) and LAB (2004–2008), +p < 0.05 LAB vs. HAB. Data are presented as mean ± SEM.
History of HAB and LAB Rats
Starting in 1993, commercially available Wistar rats (Charles River, Sulzfeld, Germany) were selected and further bred for extremes in anxiety-related behaviour as measured on the EPM resulting in the HAB and LAB breeding lines (Liebsch et al., 1998b
). One year later, Wistar rats bred for low vs. high behavioural performance in an active avoidance task (i.e. the Y-maze) at the University of Leipzig, Germany (Hess et al., 1992
), were crossbred into the HAB and LAB lines, respectively, in order to improve the fitness of these out-bred lines. Between 1993 and 2003, breeding of HAB and LAB rats was performed exclusively at the Max Planck Institute for Psychiatry in Munich. In 2003, the HAB and LAB breeding lines were transferred to our laboratory at the University of Regensburg, which is currently the only location maintaining these lines. Throughout their existence, the HAB and LAB rat lines have been treated identical in terms of housing, animal care, mating and behavioural testing, and, importantly, sibling mating has been strictly avoided. The bidirectional breeding and selection procedure of HAB and LAB rats for either further breeding or experimental use is based on their performance on the EPM, which is tested between 9 and 10 weeks of age. The line difference in anxiety-related behaviour is stable (Figure 1
A) and present over all seasons under our controlled laboratory conditions (12/12 h light/dark cycle with lights on at 6:00 a.m., 21 ± 1°C, 60% humidity, standard rat chow and tap water ad libitum). However, LAB rats have even lower levels of anxiety during the summer season (Figure 2
A). Male and female HAB and LAB rats are selected for further breeding only if the percentage of time spent on the open arms of the maze is below 5% or above 45%, respectively. For experimental purposes, HAB rats with less then 10% and LAB rats with more then 35% time on the open arms are used. These strict criteria are met by approximately 80–90% of all offspring.
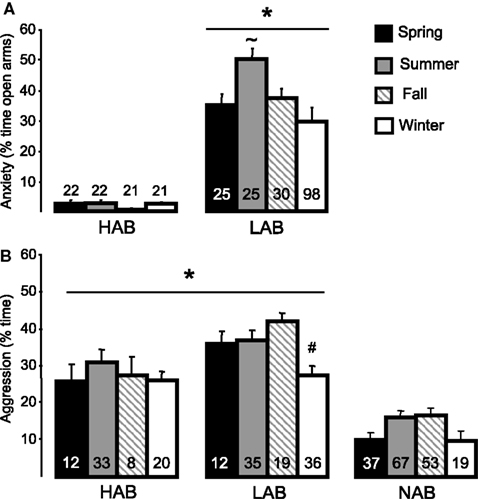
Figure 2. (A) Anxiety-related behaviour on the EPM as reflected by the percentage of time on the open arms measured in 9-week-old male HAB and LAB rats (*p < 0.05 vs. HAB at any time, ∼p < 0.05 vs. other 3 seasons), and (B) aggressive behaviour of 16- to 22-week-old male HAB, LAB, and NAB rats (*p < 0.05 vs. NAB, #p < 0.05 vs. summer and fall) during the four seasons. Numbers in columns indicate group size. Data are presented as mean + SEM.
Each behavioural test established to characterize a particular anxiety-related phenotype has its own limitations. Therefore, it is important that the differences in anxiety-related behaviour between HAB and LAB rats are not restricted to the selection criterion, i.e. their behaviour on the EPM. In the case of HAB and LAB rats, differences have also been confirmed in other validated tests of anxiety, including the open field, the black-white box (Henniger et al., 2000
; Slattery and Neumann, 2010
), the modified hole board (Ohl et al., 2001
), and in tests for conditioned anxiety (Muigg et al., 2008
). Moreover, the HAB/LAB differences in anxiety-related behaviour are (i) independent of gender, as both male and female offspring meet the selection criteria (Bosch et al., 2005
; Bosch and Neumann, 2008
), and (ii) independent of age, as HAB pups show more ultrasound vocalization when separated from their mother during the first week of life, and differences in EPM behaviour are consistent between 9 weeks and 19 months of age (Landgraf and Wigger, 2002
), and (iii) robust as behavioural outcomes have been consistent over the last 6 years (Figure 1
A) and across several European laboratories (Salome et al., 2002
; Veenema et al., 2007a
). Importantly, both HAB and LAB rats significantly differ from non-selected rats purchased from Charles River (NAB), which show intermediate behavioural and emotional responses in a variety of tests (Ohl et al., 2001
; Bosch et al., 2005
; Veenema et al., 2007a
). Taken together, these facts suggest some success in our goal to bidirectionally breed for extremes in anxiety-related behaviour with HAB rats showing the highest and LAB rats showing the lowest avoidance of an aversive environment.
It is well accepted that anxiety and depression represent overlapping symptoms which cannot be easily dissociated (Cryan and Holmes, 2005
; Grillon et al., 2005
; Landgraf et al., 2007
). In this context it is important to note that the selection for extremes in trait anxiety is paralleled by a divergence in depression-related behaviour. Thus, HAB rats float more and struggle less during a single 10-min exposure to the forced swim test (21–23°C) indicative of depression-like behaviour, whereas LAB rats show the opposite and are more active (Liebsch et al., 1998b
; Wiehager et al., 2009
; Slattery and Neumann, 2010
). In addition, long-term treatment with the serotonin-reuptake inhibitor paroxetine, an antidepressant, reversed the passive, depression-like behaviour of male HAB rats to the level of active coping strategy seen in LABs (Keck et al., 2003
). Other findings in HAB rats demonstrate a hyper-responsiveness of the HPA axis to a mild emotional stressor (Landgraf et al., 1999
) and a pathological adrenocorticotropic hormone (ACTH) response during the combined dexamethasone/corticotropin-releasing hormone challenge test (Keck et al., 2003
), a diagnostic marker for major depression (Holsboer et al., 1994
). Together, these results indicate the high potential of HAB rats as a model for anxiety with co-morbid depression.
Aggression and Other Social Behaviours in HAB and LAB Rats
The bidirectional breeding for high and low anxiety has also resulted in distinct phenotypes with respect to various aspects of social behaviour summarized in Table 1
. For example, differences in the overall level of social interaction, social discrimination abilities, maternal behaviour and intermale aggression could be revealed. In the modified hole board test, LAB rats spent less time in social contact with their cage mates than HAB rats (Ohl et al., 2001
). LAB males also have a slightly impaired ability to discriminate between a novel and a familiar juvenile to which they were exposed 30 min before (Landgraf and Wigger, 2002
). In a social preference test, which consists of measuring the time the experimental animal spends in contact with a caged male rat vs. the time the animal spends with an empty cage, LAB rats do not show social preference, whereas both HAB and NAB rats do (Lukas et al., 2008
). These data suggest that LAB rats are likely to have a lower “need” for social support in a novel environment, no preference for social vs. non-social stimuli or, alternatively, even an aversion to social interaction in general (social phobia). In fact, it cannot be ruled out that social avoidance may influence the results of discrimination tasks. Conversely, in a different social interaction test, pairs of male LAB rats spent more time in active social interaction compared with pairs of HAB males (Henniger et al., 2000
), but an increased display of aggression in an unavoidable social interaction might have contributed to the increased time spent in active social interactions. Indeed, breeding line-dependent differences in intermale aggression have been found under varying experimental conditions such as among cage-mates in the home cage (Henniger et al., 2000
), towards an intruder in the RI test (see below), and towards a resident in the social defeat test (Frank et al., 2006
). In each case, LAB rats were consistently more aggressive. Interestingly, in the RI test, LAB males strongly differ from HAB and NAB rats not only in aggression (see below), but also in non-aggressive social behaviour, as they spend less time in non-aggressive social interaction with the intruder (Beiderbeck et al., 2007
; Veenema et al., 2007a
). The behavioural differences in intermale aggression as seen in the RI test will be discussed in detail below.
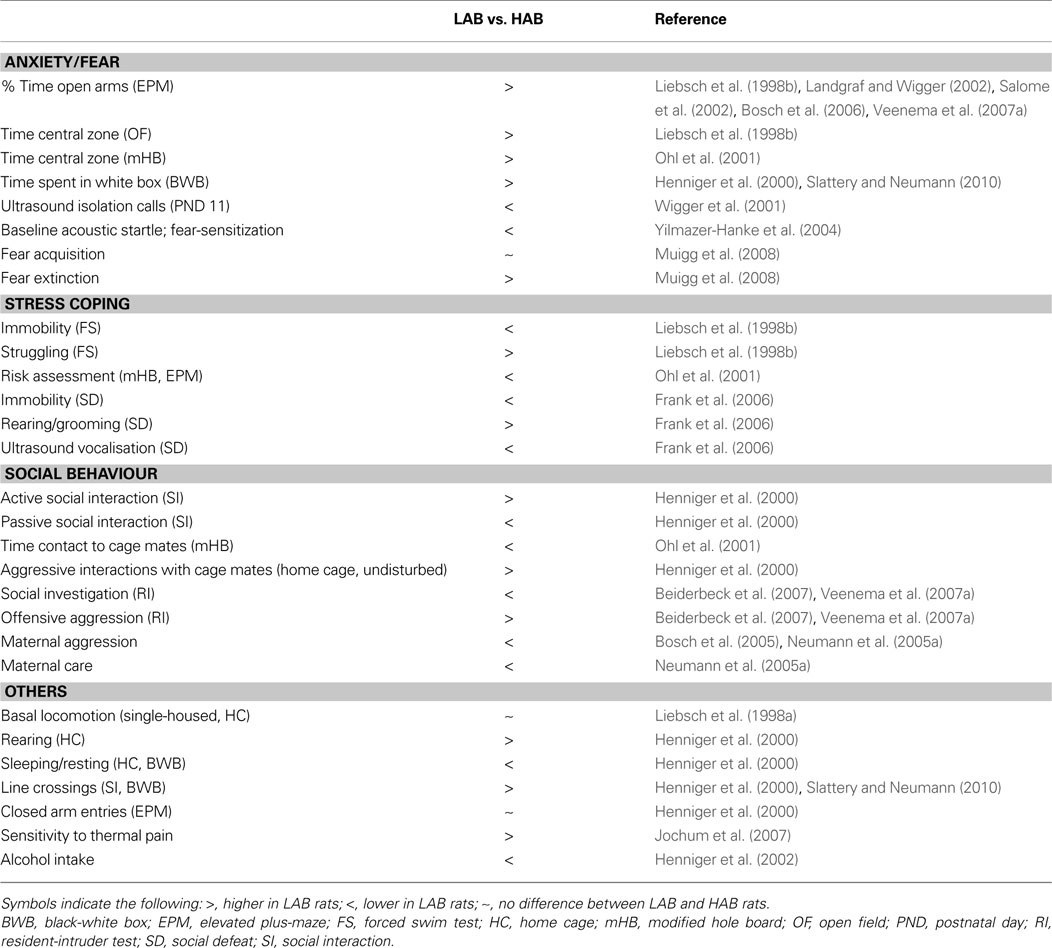
Table 1. Overview of the behavioural differences between LAB and HAB rats.
Profound differences in social and aggressive behaviours are also observed in female HAB and LAB rats, especially during lactation. HAB dams generally display a more protective style of maternal behaviour, as they leave the nest less often and spend more time on the offspring in an arched back nursing position (Bosch et al., 2005
; Neumann et al., 2005a
; Bosch and Neumann, 2008
). The protective behaviour of HAB dams is also reflected by an increased level of maternal aggression towards a female intruder rat during the maternal defence test (Bosch et al., 2005
; Bosch and Neumann, 2008
). Differences in the brain oxytocin and AVP systems between HAB and LAB dams were found to contribute to these distinct forms of maternal behaviour (Bosch et al., 2005
; Bosch and Neumann, 2008
).
LAB Rats as a Model for High and Abnormal Aggression
Although LAB rats originally served as breeding controls for HABs, continuous bidirectional breeding for anxiety-related behaviour resulted in two lines of Wistar rats with distinct behavioural phenotypes compared with NAB rats. Accordingly, LAB rats by themselves are of potential interest as an animal model because of their altered phenotype, which includes extremely low anxiety and fear responses, reduced risk assessment, high exploration, active stress coping style, and high and abnormal forms of aggressive behaviours in different social settings. Of particular interest for this review is the finding of a high level of intermale aggression in LAB residents, which remained consistent over the last 6 years (Figure 1
B), and an intermediate level of aggression in HAB residents during the RI test as compared with NAB residents (Figure 1
B).
In LAB rats, there was only a small seasonal effect with a reduction in the level of aggression during the winter season under the experimental conditions in our laboratory with constant light/dark cycle, room temperature, and humidity (Figure 2
B). A detailed analysis of the behavioural profile during the RI test (using an unfamiliar NAB male rat as standard intruder) indicates that LAB rats consistently show the highest level of aggression as indicated by more time spent with total aggressive behaviour (Figure 3
A), lateral threat (Figures 3
C,E), and offensive upright (Figures 3
D,F). Lateral threat, in particular, is a major element of aggression which often precedes an attack and comprises almost half of the total aggression time in both HAB and LAB rats. Moreover, male LAB rats spend less time with social investigation compared with HAB and NAB rats (Figure 3
B). The excessive aggression of LAB males is also reflected by the fact that almost 80% of all LAB rats tested over the last 6 years showed aggressive behaviour for more than 15% of the time of the RI test, and almost 10% of LABs showed aggressive behaviour for more than 55% of the time (Figure 4
). In contrast, none of the HAB or NAB rats reached this high level of aggression. In fact, more than 50% of the NAB rats did not reach the 15% of aggression level (Figure 4
). Thus, the LAB, and to a lesser extent the HAB, breeding line are characterized by remarkable aggressive behaviour compared with non-selected Wistar rats.
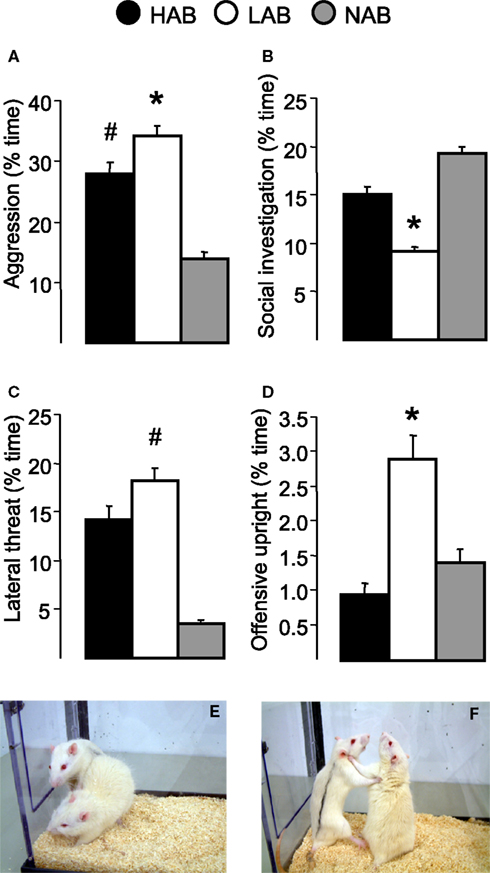
Figure 3. Social behaviours of HAB, LAB and NAB male rats during the 10-min RI test: (A) aggressive behaviour, (B) social investigation, (C) lateral threat, and (D) offensive upright. (E) Lateral threat and (F) offensive upright displayed by the resident towards the intruder (intruder is marked with black lines). *p < 0.05 vs. NAB and HAB, #p < 0.05 vs. NAB. Data are presented as mean + SEM.
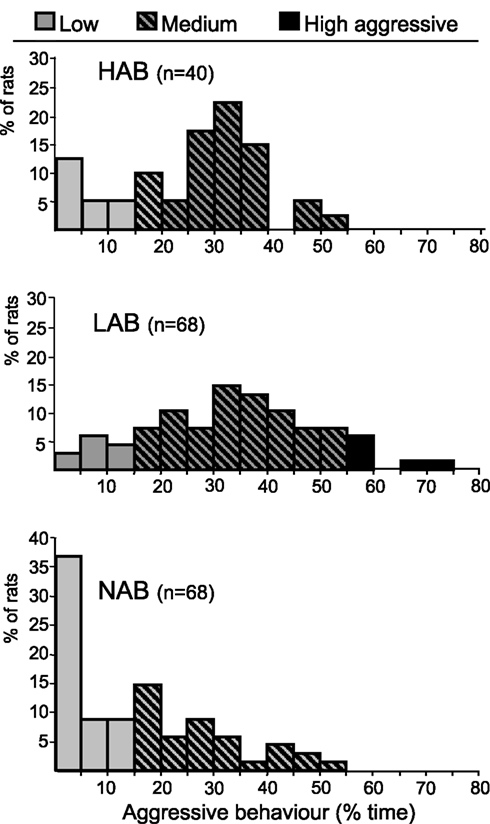
Figure 4. Allocation of HAB, LAB, and NAB male in low-, medium- and high-aggressive. Rats are categorized according to their level of aggressive behaviour expressed as percentage of time in the RI test (low-aggressive: less than 15% aggressive behaviour; medium-aggressive: between 15 and 55% aggressive behaviour; high-aggressive: more than 55% aggressive behaviour).
Is the high level of aggressive behaviour in male LAB rats accompanied by the display of abnormal forms of aggression? According to the definition given above, abnormal aggression occurs when species-specific rules are disregarded, which, e.g., would be reflected in the RI test by attacks directed towards vulnerable (head, belly, genitals) as opposed to less vulnerable (flanks and back) body parts of the male intruder. Indeed, in a pilot study we could observe that 90% of LAB and 70% of the HAB residents, but only 20% of NAB rats attacked vulnerable body parts of a male intruder rat (Figure 5
). Similarly, 70% of resident LAB males attacked a non-oestrus female intruder, whereas only 10% of HAB and NAB residents did attack the female (Figure 5
). Also, all LAB (100%), but only 38% of NAB males attacked a narcotized male intruder. We speculate that, in the case of the anesthetized male intruder, the lack of defensive signalling (due to the lack of vibrissae activity) (Blanchard et al., 1977
) may have triggered aggressive behaviour in LAB rats. Taken together, our behavioural observations demonstrate that both an extremely low and a high level of innate anxiety can be associated with high and abnormal forms of aggressive behaviour. As LAB rats display high and abnormal forms of aggression and avoid non-aggressive social interactions, they emerge as an interesting model for pathological social behaviours in general.
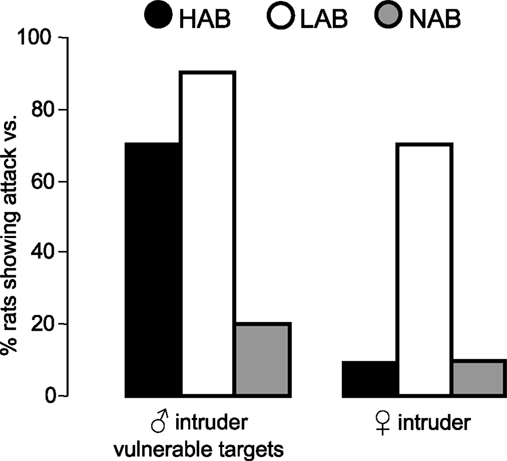
Figure 5. Line-specific differences in abnormal aggression as indicated by the percentage of HAB, LAB and NAB male rats attacking vulnerable body parts of a male intruder (left) or a female intruder (right) during the RI test in their home cage.
Correlation between Aggression and Anxiety in LAB and HAB Males
The unique phenotypes of LAB and HAB rats with consistent differences in both innate anxiety and intermale aggression provide the chance to correlate parameters of anxiety- and aggression-related behaviours. Both LAB and HAB rats show a higher level of aggression than NAB rats indicating that both high as well as low trait anxiety can be associated with relatively high aggression scores (Figures 1 and 2
), whereas the intermediate level of anxiety-related behaviour as found in NAB rats was accompanied by a low level of intermale aggression. In a previous analysis performed on a small sample of rats (n = 7–9 in the LAB, HAB, and NAB group) we reported a negative linear correlation between anxiety and intermale aggression (Veenema et al., 2007a
). In detail, the highest aggression was found in LAB rats that had the lowest level of anxiety. We are currently investigating a possible correlation between aggression and anxiety in a larger number of LAB, HAB and NAB rats, as our recent data indicate that a U-shaped correlation may emerge, i.e., that both HAB and LAB males show a higher level of aggression than NAB males. This may indicate that both high and low trait anxiety can be associated with relatively high aggression scores (Figure 1
).
The complex and fine-tuned emotional and social responsiveness of an individual is regulated by numerous neurochemicals including classical neurotransmitters and neuromodulators like neuropeptides and hormones. Preclinical research on high and abnormal aggression has focused mainly on the role of HPA axis hormones, the neuropeptide AVP, and neurotransmitters like 5-HT and gamma-aminobutyric acid (GABA; for reviews see de Almeida et al., 2005
; Haller et al., 2005
). Using LAB and HAB rats, we have found experimental evidence that differences in HPA axis responsiveness and the AVP and 5-HT systems do not only contribute to the extremes in anxiety-related behaviours of these animals (Landgraf et al., 2007
), but also to their aggression phenotypes.
HPA Axis
The HPA axis is one of the major stress systems and is regulated by the release of corticotropin-releasing hormone and AVP from the paraventricular nucleus (PVN) into the pituitary, which stimulates the secretion of ACTH into the blood. ACTH, in turn, triggers the adrenal release of glucocorticoids (cortisol in humans, corticosterone in rats), which facilitate physiological and behavioural adaptations to a stressor at multiple peripheral and brain levels. Dysregulation of the HPA axis has been frequently linked to anxiety- and depression-related disorders, as well as to violence and abnormal aggression (Plotsky et al., 1998
; Mello et al., 2003
; de Kloet et al., 2005
). Although data from preclinical studies are conflicting, they generally implicate glucocorticoids as important regulators of aggressive behaviour (Haller et al., 1998
, 2001
; Summers et al., 2005b
). Paradoxically, both acute high and chronically low levels of circulating glucocorticoids have been associated with high aggression (see above; Hayden-Hixson and Ferris, 1991
; Haller et al., 2000
; Kruk et al., 2004
; Summers and Winberg, 2006
). For example, an acute rise in glucocorticoids promotes intermale aggression in rodents (Mikics et al., 2004
) and might be associated with a state of hyperarousal and sudden outbursts of aggression. Acute blockade of glucocorticoid synthesis results in reduced aggression in male rats, an effect which is reversible by corticosterone treatment (Mikics et al., 2004
). In contrast to these findings, low HPA axis reactivity (Veenema et al., 2003b
, 2005
) or chronically low glucocorticoid levels (Haller et al., 2001
, 2005
; Kim and Haller, 2007
) also result in excessive aggression and a higher frequency of attacks directed towards vulnerable body parts and might underlie pathological forms of aggression including violence.
The distinct behavioural phenotypes of the LAB and HAB breeding lines are accompanied by stressor-dependent differences in neuroendocrine responses. When exposed to a non-social stressor, e.g. a novel environment, HAB rats show a more pronounced ACTH and corticosterone secretory response (Landgraf et al., 1999
; Neumann et al., 2005b
) and elevated neuronal responsiveness in the PVN, the medial preoptic area and the locus coeruleus (Salome et al., 2004
). In contrast, exposure to a social conflict situation seems more threatening to LAB rats (Veenema and Neumann, 2007
). When exposed as an intruder during the social defeat test, LAB males show a higher HPA axis response despite showing fewer signs of emotional distress like ultrasound vocalization and freezing (Frank et al., 2006
). Moreover, higher ACTH and corticosterone responses are found in LAB compared with HAB and NAB rats when exposed as resident to the RI test (Veenema et al., 2007a
). The elevated neuroendocrine responses are also accompanied by higher neuronal activation of the PVN and the central amygdala in LAB compared with HAB resident males (Veenema et al., 2007a
). These results suggest a generally higher responsiveness to social stressors in LAB males. However, as LABs generally display a higher level of aggression, two possibilities may explain the elevated stress responsiveness to social stimuli in LAB males: (i) their active behavioural coping style increases metabolic demands or (ii) social stimuli are perceived as more of a threat, which is translated into higher levels of aggression when the stimulus can not be avoided. In support for the latter possibility, an acute rise in glucocorticoids is required to initiate aggressive behaviour in rodents as discussed above (Clarke and File, 1983
; Haller et al., 2000
; Kruk et al., 2004
; Mikics et al., 2004
). In general, LAB and HAB rats mimic clinical reports in which high and abnormal aggression have been linked to either a state of high or low glucocorticoid function making them an interesting tool to further explore the link between HPA axis dysregulation and high and abnormal forms of aggression, and distinct anxiety levels.
Brain AVP System
More than any other class of neuroactive substances, brain neuropeptides are critically involved in emotionality and complex social behaviours. In particular, the nonapeptide AVP and the closely related oxytocin have been extensively characterized with respect to their ability to modulate anxiety- and depression-related behaviours and a broad variety of social behaviours including social cognition, pair-bonding, maternal behaviour, and aggression. With respect to intermale aggression and anxiety-related behaviour, brain AVP seems to play an especially important role. AVP is synthesized in the hypothalamic PVN and supraoptic nucleus as part of the hypothalamo-neurohypophysial system, and in the bed nucleus of the stria terminalis (BST), medial amygdala and suprachiasmatic nucleus. AVP can be released within distinct brain regions as a neurotransmitter, but generally may act more like a neuromodulator. Via the latter type of information processing, the brain is liberated from the constraints of wiring, as locally released neuropeptides can reach any neighbouring brain region by diffusion thereby increasing the regulatory capacity of neuropeptidergic neurons (Landgraf and Neumann, 2004
). Several stress-related, fear-provoking and social stimuli were found to trigger local release of AVP within distinct brain regions, such as the PVN, where AVP can be released from dendrites and perikarya, as well as other regions outside the hypothalamus (Engelmann et al., 2004
; Landgraf and Neumann, 2004
; Veenema and Neumann, 2008
).
Brain AVP has been implicated in aggressive behaviour in several species including rats, mice, hamsters, birds, and humans (Koolhaas et al., 1990
; Ferris, 1992
; Albers and Bamshad, 1998
; Coccaro et al., 1998
; Veenema and Neumann, 2007
; Goodson, 2008
). However, conflicting data have been reported on AVP as a potential modulator of intermale aggression. In hamsters, local injections of AVP into brain regions relevant for aggression like the anterior hypothalamus, BST, and lateral septum promote aggressive behaviour (Ferris and Potegal, 1988
; Irvin et al., 1990
). Moreover, a higher density of AVP immunoreactivity and V1a receptors were found in the lateral septum of highly vs. less aggressive Peromyscus mice (Bester-Meredith et al., 1999
). In rats, testosterone-dependent AVP pathways, which originate in the BST/medial amygdala and project to the lateral septum (de Vries and Buijs, 1983
), are diminished after castration (de Vries et al., 1984
) as are levels of aggression (Koolhaas et al., 1990
). Injection of AVP into the medial amygdala or lateral septum of castrated male rats reverses (i.e., increases) the aggression level to that seen in intact males (Koolhaas et al., 1990
, 1991
). In humans, AVP concentration in the cerebrospinal fluid correlates positively with the level of aggression in personality-disordered subjects (Coccaro et al., 1998
). Recently, intranasal AVP has been shown to alter aspects of social communication in men, as a decrease in the perception of friendly faces and an increase in the perception of threat and anger when exposed to neutral faces were found in AVP-treated healthy men (Thompson et al., 2004
). Together, these data provide evidence for a positive link between brain AVP and aggression, but there are also reports demonstrating the opposite. For example, a lower AVP cell number in the BST and a lower density of AVP fibres within the lateral septum were found in two animal models, i.e. in SAL vs. LAL mice and in high vs. low aggressive wild-type rats (Compaan et al., 1993
; Everts et al., 1997
). These contradictory findings led us to question the role of endogenous brain AVP within the lateral septum during the expression of intermale aggression.
There is also substantial evidence that AVP is an important anxiogenic factor (for reviews see Ring, 2005
; Landgraf, 2006
; Landgraf et al., 2007
). Accordingly, dysregulation of the brain AVP system, especially over-activity of AVP neurons, is likely to contribute to the etiology and symptomatology of anxiety- and depression-related disorders (Murgatroyd et al., 2004
; Simon et al., 2008
; Surget and Belzung, 2008
). Based on the behavioural and neuroendocrine phenotype of LAB and HAB rats, together with the anxiogenic effects of AVP, the AVP gene has been considered a candidate gene for anxiety (for review see Landgraf et al., 2007
). Indeed, characterization of the activity of the brain AVP system in LAB and HAB rats has revealed an elevated expression of the AVP gene in male (Keck et al., 2002
; Wigger et al., 2004
) and female (Bosch and Neumann, 2010
) HAB rats without changes in central AVP V1a receptor expression or binding (Wigger et al., 2004
; Bosch and Neumann, 2010
). Sequence analysis of the AVP gene promoter revealed 10 single nucleotide polymorphisms within the AVP promoter of HAB rats, one of which was localized in a transcription factor DNA-binding site known as CArG box (Murgatroyd et al., 2004
). Mutation within the CArG box impairs the binding of a transcriptional repressor, i.e. the CArG box binding factor A, resulting in disinhibition of AVP gene expression. Overexpression of the AVP gene and subsequent elevated levels of local AVP release within the PVN could contribute not only to the high level of anxiety (Keck et al., 2003
), but also to the elevated HPA axis response to non-social stimuli observed in HABs (Keck et al., 2002
).
Moreover, the differences in brain AVP system activity and, consequently, AVP availability in LAB and HAB rats together with their distinct behavioural phenotypes provided the opportunity to reveal the contribution of endogenous AVP to a given aggression phenotype. Thus, we compared local AVP release patterns between the breeding lines during the RI test using intracerebral microdialysis. Within the lateral septum of LAB males, there was a significant decrease in AVP release during the display of intermale aggression accompanied by a lower neuronal activation compared with HAB males, in which local AVP release tended to increase (Beiderbeck et al., 2007
). In contrast to our expectation, increasing the availability of AVP in the extracellular fluid within the lateral septum of LAB rats by application of synthetic AVP via reversed microdialysis did not alter the level of aggressive behaviour, but resulted instead in an increase in anxiety on the EPM (Beiderbeck et al., 2007
). Further, blockade of local V1a receptors by the selective AVP V1a receptor antagonist d(CH2)5Tyr(Me)AVP (kindly provided by Dr. Maurice Manning, Toledo, USA) did not affect aggressive behaviour of HAB rats. Interestingly, however, the V1a receptor antagonist reduced the duration of general social investigation during the RI test in HAB rats to the low level seen in LAB rats.
Taken together, despite differences in AVP release patterns within the septum found in LAB and HAB rats during the RI test, a causal role of locally released AVP in the expression of intermale aggression could not be confirmed. Therefore, we recently suggested that the aggression-induced change in septal AVP release may modulate closely related adaptive behaviours like anxiety- and fear-responses, stress coping, or social cognition relevant for future behavioural strategies rather than aggression itself (Veenema and Neumann, 2008
). In this context it is of interest to note that differences in local AVP release within the septum and within the BST are also associated with differences in aggression in NAB rats (Veenema et al., 2010
). In NAB males classified as being aggressive during the RI test, AVP release within the septum significantly increased, whereas it remained unchanged in non-aggressive NAB rats. However, local application of synthetic AVP to non-aggressive NAB rats did not alter their respective aggression level, even though the anxiogenic effect of AVP could be confirmed on the EPM. Local application of the V1a receptor antagonist to aggressive NAB rats did reduce their aggression level.
In addition to the medio-lateral septum, the BST has been suggested in the regulation of intermale aggression (Hines et al., 1992
). Local microdialysis revealed differences in AVP release between aggressive and non-aggressive NAB rats (Veenema et al., 2010
). In contrast to the lateral septum, local administration of synthetic AVP into the BST of aggressive NAB rats significantly reduced their aggression level without affecting anxiety-related behaviours. These findings highlight brain-region specific functions of AVP in relation to the regulation of aggressive and anxiety-related behaviours. Further microdialysis studies combined with local pharmacological or genetic manipulations are necessary in order to gain a better understanding of the region-dependent role of AVP in the regulation of intermale aggression and its interplay with anxiety. Because the extrahypothalamic AVP system is under significant influence of gonadal steroids, and testosterone, in particular, modulates AVP mRNA expression in the medial amygdala and the BST (de Vries et al., 1992
), the role of sexual steroids in the control of aggression deserves closer attention.
Testosterone
Testosterone, the principal male sex steroid, has long been implicated in the regulation of aggression in both animals and humans (Dijkstra et al., 1992
; Banks and Dabbs Jr., 1996
; Lucion et al., 1996
; Wingfield et al., 2001
), although varieties between species, social status, type of aggression and type of social encounter make a simple causal relationship rather unlikely (Archer, 2006
). Evidence exists for a relationship between circulating testosterone and aggression, for example during puberty, when testosterone and aggression levels simultaneously rise in most mammalian species (Koolhaas et al., 1980
). Also, castration reduces aggressive behaviour, an effect which is reversible by testosterone administration (Beeman, 1947
; Barfield et al., 1972
; Luttge, 1972
; Barr et al., 1976
). The reduction in aggression after castration is accompanied by testosterone-dependent reduction in AVP immunoreactivity in the septum (de Vries et al., 1992
; de Vries and Miller, 1998
). Thus, testosterone effects on intermale aggression may, at least in part, be mediated by changes in the limbic AVP system. However, testosterone-independent aggressive behaviour has been described in birds in the non-breeding season (Logan and Carlin, 1991
; Wingfield, 1994a
,b
). In humans, aggression correlates with elevated levels of plasma testosterone in adolescent men (Mattsson et al., 1980
; Dabbs Jr. et al., 1991
), but not in prepubertal boys (Susman et al., 1987
; Constantino et al., 1993
). This is confirmed in a more recent study that further demonstrates a positive relationship between testosterone and delinquency behaviour (van Bokhoven et al., 2006
). Testosterone was also associated with antisocial personality in a small sample of violent men (Aromäki et al., 1999
). However, others found that a combination of testosterone and low socioeconomic status was a better predictor of violence, than was testosterone alone (Dabbs Jr. and Morris, 1990
; Mazur, 1995
). We could recently show that basal plasma testosterone did not differ between LAB and HAB males, but a significant increase in plasma testosterone in response to the RI test was only found in LAB residents (Veenema et al., 2007a
). Thus, the possibility exists that the acute rise in testosterone contributes to the modulation of neuronal networks regulating aggression (Delville et al., 1996
).
Brain 5-HT System
5-HT is synthesized in the midbrain raphe nuclei and released from projections throughout the brain, and is significantly involved in the regulation of emotionality (Lucki, 1998
; Olivier and van Oorschot, 2005
). Hence, 5-HT likely plays a prominent role in the pathophysiology of anxiety, depression, and aggression (Linnoila and Virkkunen, 1992
; Baldwin and Rudge, 1995
; Ressler and Nemeroff, 2000
). Clinical and animal studies suggest that excessive aggression and violence are associated with reduced 5-HT function. For example, low cerebrospinal fluid levels of the 5-HT metabolite 5-hydroxyindoleacetic acid were found in patients with high levels of aggression and violent behaviour (Berman et al., 1997
). Second, a direct link between a polymorphism in the microRNA target region of the 5-HT1B receptor gene and a history of aggression-related conduct-disorder behaviour was recently reported in young men (Jensen et al., 2009
). This polymorphism determines the efficacy of direct interactions with the microRNA target region of the 5-HT1B receptor gene and, thus, regulates 5-HT1B receptor mRNA expression. Furthermore, variations in the 5-HT reuptake transporter gene are known to influence emotional and social behaviours (Lesch et al., 1996
, 1997
). For example, female rhesus monkeys carrying the short allele of the 5-HT reuptake transporter initiate more aggressive and submissive behaviour than females carrying the long variant (Jarrell et al., 2008
). Several rodent studies also point towards an inhibitory role of 5-HT in intermale aggression: 5-HT acting through 5-HT1B receptors in the anterior hypothalamus reduces offensive aggression in male hamsters (Ferris, 1996
; Ferris et al., 1997
, 1999
). In support, an increase in intermale aggression in adult male rats exposed to early life stress, i.e. maternal separation, correlates negatively with 5-HT immunoreactivity within the anterior hypothalamus (Veenema et al., 2006
). Finally, abnormal forms of aggression were associated with chronically reduced 5-HT activity (Miczek et al., 2002
). In contrast, other studies demonstrate that the display of aggression is associated with an acute increase in 5-HT activity (van der Vegt et al., 2003a
,b
; Summers et al., 2005a
), indicating a possible difference in the role of 5-HT in adaptive forms of aggression, such as social dominance (high 5-HT neurotransmission) vs. excessive or abnormal forms of aggression (low 5-HT neurotransmission).
In comparing LAB and HAB rats, distinct differences in the activity of the brain 5-HT system were found. Such differences include the release of 5-HT within the hippocampus, amygdala and lateral septum in response to non-social stressors (higher in LAB rats), hippocampal 5-HT1A receptor expression (higher in LAB rats) and 5-HT transporter binding sites (lower in LAB rats) (Keck et al., 2005
). Thus, in comparison with HAB rats, LAB rats generally seem to show higher 5-HT neurotransmission. Chronic treatment with the 5-HT reuptake inhibitor paroxetine elicited the stress-induced rise in hippocampal 5-HT only in HAB rats (Keck et al., 2005
), suggesting that low 5-HT neurotransmission in HABs might contribute to their enhanced anxiety- and depression-related behaviour.
Evidence for breeding line differences in local 5-HT release during social stressors or during the display of aggression is lacking so far. Nevertheless, we have preliminary evidence for a crucial involvement of the 5-HT system in high and abnormal forms of aggression in LAB rats. Blocking postsynaptic 5-HT release by administration of a preferential 5-HT1A autoreceptor agonist selectively reduced the level of intermale aggression towards an intact intruder and decreased the number of attacks directed towards vulnerable body part of an anesthetized intruder (unpublished). This suggests that a high 5-HT neurotransmission in LABs is associated with the display of high and abnormal aggressive behaviours.
Brain GABA System
Based on the findings that elevated aggression is likely to be linked to both extremely low and high levels of trait anxiety as found in LAB and HAB rats, respectively, the question arises, whether the level of intermale aggression can be altered by acute manipulation of state anxiety. Our preliminary results indicate that anxiogenic or anxiolytic substances acting on GABA receptors affect the level of aggression in LAB and HAB males (unpublished). LAB rats treated with the anxiogenic substance pentylenetetrazole – a GABAA receptor antagonist – showed higher anxiety scores on the EPM, and this result was accompanied by a dramatic reduction in aggression and social investigation during the RI test. It has been shown that anxiogenic treatment increases neuronal activation in brain regions including the central amygdala, BST, lateral septum, and PVN that are associated with stress-, fear- and aggression-related behaviour (Singewald et al., 2003
). Thus, treatment-induced alterations in local activity could directly affect offensive and non-offensive social approach as seen in pentylenetetrazole-treated LAB rats. Interestingly, acute manipulation of state anxiety by anxiogenic or anxiolytic substances in NAB rats with an overall low level of aggression did not change any behavioural parameter of aggressive behaviour (unpublished). This indicates that LAB and HAB rats may be a better model of high and abnormal aggression than commercially available rats given their sensitivity to psychopharmacological manipulation.
The striking evidence for an overlap in neuroendocrine and neurochemical systems regulating aggression as well as anxiety suggests a strong correlation between these two behaviours. Here, examples of co-morbid effects on aggression and anxiety as observed in several different rodent models are discussed, including those found after acute pharmacological interventions, exposure to early life stress, in transgenic mice, or in rodents selectively bred for extremes in emotionality. Most of the evidence to date provides support for a link between aggression and anxiety, although not always in a uni-directional manner. In this context, we could demonstrate that male LAB and HAB rats, characterized by distinct anxiety phenotypes, are a suitable model to (i) correlate parameters of aggression and anxiety-related behaviour and (ii) study underlying neuronal, neuroendocrine and neurochemical substrates. Both LAB and HAB males display elevated and abnormal forms of aggression in various tests, in which the LAB males are the most excessive. Microdialysis revealed significant differences in the release patterns of septal AVP between LAB and HAB rats, during the RI test, although such locally released AVP was not causally involved in regulating intermale aggression. Additionally, LAB rats generally seem to have a higher 5-HT neurotransmission compared with HAB rats. These and other preliminary findings lead us to conclude that LAB and HAB rats are a promising rodent model to study the link between aggression and anxiety on a behavioural, neuronal, molecular, and genetic level in more detail.
However, several animal models discussed in this review lack an obvious link between aggression and anxiety, and in other models simultaneous alterations in these behaviours were not found after acute intervention. Thus, aggression and anxiety are not always co-regulated, but, under some circumstances, these behaviours may come under the control of the same genes and neuroactive substances including sexual steroids, neuropeptides and neuroamines within specific brain circuitries. Such a view is in agreement with clinical findings. On the one hand, excessive and violent behaviours are seen in humans exposed to adverse early life experiences and in patients with depression- and anxiety-related disorders, or PTSD. On the other hand, rather conflicting data exist on the effects of anxiolytic drugs on anti-social and aggressive behaviours. In future studies that focus on the neurobiological mechanisms of (co-)regulation of aggression and anxiety, epigenetic modifications need to be considered in addition to the neuronal and neuroendocrine parameters discussed above. Thus far, such studies are lacking. In contrast, a role of epigenetic modification of gene activity due to, for example, early life experiences or chronic psychosocial stress in adulthood were found to contribute to differences in adult stress coping responses (Weaver et al., 2004
; Tsankova et al., 2006
). Also, genetic variations, for example in the AVP V1a receptor (Young and Wang, 2004
) or the 5-HT transporter (Jarrell et al., 2008
), were shown to account for variations in both emotionality and complex social behaviours in animals and humans (for review see Shepard et al., 2009
). Together with our findings concerning the AVP system in the HAB/LAB rat model, a closer examination of this system in human populations with respect to co-morbid aggression and anxiety is warranted. The genetic/epigenetic predisposition of an individual is also likely to influence the behavioural effects of neurochemicals that regulate aggression and anxiety, which may explain the discrepant outcome of anxiolytic treatment on aggressive and violent behaviour in humans.
Taking into account that complex social behaviours like aggression are more than hormone- and neuropeptide-driven events triggered by social cues, future studies are needed to reveal the (epi)genetic factors driving the switch from normal to excessive aggression and/or anxiety. Thus, genetic association studies are required to reveal gene polymorphisms in the context of high and abnormal aggression. In combination, genetic manipulations and validation of selected genes (using gene silencing techniques like antisense oligonucleotides or siRNA) or amplification of gene activity (using viral gene transfer) will likely help to reveal new insights into the regulation of normal and abnormal aggressive behaviour. The use of animal models with genetic differences in relevant neuropeptide systems underlying extremes in social and/or emotional behaviours, such as LAB and HAB rats, are a promising starting point.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank Dr. Benjamin Rood (University of Massachusetts, Amherst, MA, USA) for the critical reading of the manuscript. Supported by BMBF (Inga D. Neumann), DFG (Inga D. Neumann; Ne465/17) and Bayerische Forschungsstiftung (Alexa H. Veenema). The experiments were approved by the Committee on Animal Health and Care of the Government of the Oberpfalz and are in accordance with the Guide for the Care and Use of Laboratory Animals by the National Institute of Health.
Bunck, M., Czibere, L., Horvath, C., Graf, C., Frank, E., Kessler, M. S., Murgatroyd, C., Muller-Myhsok, B., Gonik, M., Weber, P., Putz, B., Muigg, P., Panhuysen, M., Singewald, N., Bettecken, T., Deussing, J. M., Holsboer, F., Spengler, D., and Landgraf, R. (2009). A hypomorphic vasopressin allele prevents anxiety-related behavior. PLoS ONE 4, e5129. doi: 10.1371/journal.pone.0005129.
Keck, M. E., Sartori, S. B., Welt, T., Muller, M. B., Ohl, F., Holsboer, F., Landgraf, R., and Singewald, N. (2005). Differences in serotonergic neurotransmission between rats displaying high or low anxiety/depression-like behaviour: effects of chronic paroxetine treatment. J. Neurochem. 92, 1170–1179.
Keck, M. E., Welt, T., Muller, M. B., Uhr, M., Ohl, F., Wigger, A., Toschi, N., Holsboer, F., and Landgraf, R. (2003). Reduction of hypothalamic vasopressinergic hyperdrive contributes to clinically relevant behavioral and neuroendocrine effects of chronic paroxetine treatment in a psychopathological rat model. Neuropsychopharmacology 28, 235–243.
Keck, M. E., Wigger, A., Welt, T., Muller, M. B., Gesing, A., Reul, J. M., Holsboer, F., Landgraf, R., and Neumann, I. D. (2002). Vasopressin mediates the response of the combined dexamethasone/CRH test in hyper-anxious rats: implications for pathogenesis of affective disorders. Neuropsychopharmacology 26, 94–105.
Landgraf, R., Kessler, M. S., Bunck, M., Murgatroyd, C., Spengler, D., Zimbelmann, M., Nussbaumer, M., Czibere, L., Turck, C. W., Singewald, N., Rujescu, D., and Frank, E. (2007). Candidate genes of anxiety-related behavior in HAB/LAB rats and mice: focus on vasopressin and glyoxalase-I. Neurosci. Biobehav. Rev. 31, 89–102.
Lesch, K. P., Meyer, J., Glatz, K., Flugge, G., Hinney, A., Hebebrand, J., Klauck, S. M., Poustka, A., Poustka, F., Bengel, D., Mossner, R., Riederer, P., and Heils, A. (1997). The 5-HT transporter gene-linked polymorphic region (5-HTTLPR) in evolutionary perspective: alternative biallelic variation in rhesus monkeys. Rapid communication. J. Neural Transm. 104, 1259–1266.
Liebsch, G., Linthorst, A. C., Neumann, I. D., Reul, J. M., Holsboer, F., and Landgraf, R. (1998a). Behavioral, physiological, and neuroendocrine stress responses and differential sensitivity to diazepam in two Wistar rat lines selectively bred for high- and low-anxiety-related behavior. Neuropsychopharmacology 19, 381–396.
Muigg, P., Hetzenauer, A., Hauer, G., Hauschild, M., Gaburro, S., Frank, E., Landgraf, R., and Singewald, N. (2008). Impaired extinction of learned fear in rats selectively bred for high anxiety – evidence of altered neuronal processing in prefrontal-amygdala pathways. Eur. J. Neurosci. 28, 2299–2309.
Müller, M. B., Zimmermann, S., Sillaber, I., Hagemeyer, T. P., Deussing, J. M., Timpl, P., Kormann, M. S., Droste, S. K., Kuhn, R., Reul, J. M., Holsboer, F., and Wurst, W. (2003). Limbic corticotropin-releasing hormone receptor 1 mediates anxiety-related behavior and hormonal adaptation to stress. Nat. Neurosci. 6, 1100–1107.
Oliveira-Dos-Santos, A. J., Matsumoto, G., Snow, B. E., Bai, D., Houston, F. P., Whishaw, I. Q., Mariathasan, S., Sasaki, T., Wakeham, A., Ohashi, P. S., Roder, J. C., Barnes, C. A., Siderovski, D. P., and Penninger, J. M. (2000). Regulation of T cell activation, anxiety, and male aggression by RGS2. Proc. Natl. Acad. Sci. U.S.A. 97, 12272–12277.
Summers, C. H., Korzan, W. J., Lukkes, J. L., Watt, M. J., Forster, G. L., Overli, O., Hoglund, E., Larson, E. T., Ronan, P. J., Matter, J. M., Summers, T. R., Renner, K. J., and Greenberg, N. (2005a). Does serotonin influence aggression? Comparing regional activity before and during social interaction. Physiol. Biochem. Zool. 78, 679–694.
van Bokhoven, I., van Goozen, S. H., van Engeland, H., Schaal, B., Arseneault, L., Seguin, J. R., Assaad, J. M., Nagin, D. S., Vitaro, F., and Tremblay, R. E. (2006). Salivary testosterone and aggression, delinquency, and social dominance in a population-based longitudinal study of adolescent males. Horm. Behav. 50, 118–125.
Wigger, A., Sanchez, M. M., Mathys, K. C., Ebner, K., Frank, E., Liu, D., Kresse, A., Neumann, I. D., Holsboer, F., Plotsky, P. M., and Landgraf, R. (2004). Alterations in central neuropeptide expression, release, and receptor binding in rats bred for high anxiety: critical role of vasopressin. Neuropsychopharmacology 29, 1–14.