 Rigoberto Salado-Castillo1
Rigoberto Salado-Castillo1 Manuel Sánchez-Alavéz2 Gina L. Quirarte3 María Isabel Martínez García4
Manuel Sánchez-Alavéz2 Gina L. Quirarte3 María Isabel Martínez García4 Roberto A. Prado-Alcalá3*
Roberto A. Prado-Alcalá3*- 1Facultad de Psicología, Universidad de Panamá, Panamá City, Panamá
- 2Department of Molecular and Integrative Neurosciences, The Scripps Research Institute, La Jolla, CA, USA
- 3Departamento de Neurobiología Conductual y Cognitiva, Instituto de Neurobiología, Universidad Nacional Autónoma de México, Querétaro, México
- 4Facultad de Ciencias Químicas, Benémerita Universidad Autónoma de Puebla, Puebla, México
Memory is markedly impaired when normal activity of any of a number of cerebral structures is disturbed after a learning experience. A growing body of evidence indicates, however, that such interference with neuronal function becomes negligible when the learning experience is significantly enhanced. We now report on the effects of enhanced training on retention after temporary inactivation of cerebral nuclei known to be involved in memory, namely the substantia nigra (SN), striatum (STR), and amygdala (AMY). When training was conducted with a relatively low intensity of footshock (1.0 mA), post-training infusion of lidocaine into the SN, STR, or AMY produced a marked memory deficit. Increasing the aversive stimulation to 2.0 mA protected memory from the amnesic effect of intranigral lidocaine, but there was still a deficit after its infusion into the STR and AMY. Administration of lidocaine into each of these nuclei, in the groups that had been trained with 3.0 mA, was completely ineffective in producing alterations in memory consolidation. Simultaneous infusion of lidocaine into STR + SN, AMY + SN, or AMY + STR was also ineffective in altering memory formation when the highest footshock intensity was used for training. To our knowledge, this is the first demonstration that an enhanced learning experience guards against memory deficits after simultaneous temporary interruption of neural activity of brain nuclei heretofore thought to be necessary for memory formation. These findings support the proposition that brain structures involved in memory processing are functionally connected in series during memory consolidation and that, after an enhanced learning experience, these structures become functionally connected in parallel.
Introduction
Ample evidence indicates that interference with normal activity of any one of a number of cerebral structures brings about deficiencies in memory consolidation and has led to the assumption that a set of cerebral nuclei is essential to establish memory for particular types of tasks. It has been shown, for example, that interference with the activity of the amygdala (AMY) (Liang et al., 1982; Power et al., 2000; Roesler et al., 2000; McGaugh, 2002; Roozendaal et al., 2009), striatum (STR) (Prado-Alcalá et al., 1972, 1973; Sanberg et al., 1978; Prado-Alcalá, 1995; Salado-Castillo et al., 1996; Ambrogi-Lorenzini et al., 1999; Packard and Knowlton, 2002; White and McDonald, 2002; Izquierdo et al., 2006; White, 2009; Miyoshi et al., 2012), and substantia nigra (SN) (Routtenberg and Holzman, 1973; Kim and Routtenberg, 1976; Ambrogi-Lorenzini et al., 1994; Da Cunha et al., 2001, 2003; Díaz del Guante et al., 2004) causes significant deficiencies in retention of inhibitory avoidance (IA). There are data that indicate, however, that treatments which normally induce amnesia become ineffective when infused into these structures in rats that have been subjected to enhanced IA training (Giordano and Prado-Alcalá, 1986; Pérez-Ruíz and Prado-Alcalá, 1989; Parent et al., 1992, 1994, 1995; Parent and McGaugh, 1994; Prado-Alcalá, 1995; Cobos-Zapiaín et al., 1996). This protective effect of enhanced training has also been observed in cats (Prado-Alcalá and Cobos-Zapiaín, 1977, 1979) and rats (Prado-Alcalá et al., 1980) trained to perform operant responses in order to obtain positive reinforcers.
These findings have led to two models that postulate that those cerebral structures involved in memory consolidation in standard conditions of training are functionally connected in series, that is, the neural activity derived from the learning experience must flow through all of them before reaching a hypothetical integrative “center” whose activation is necessary for consolidating memory. This flow is halted when any component of this ensemble of structures is not functional and thus, consolidation is not achieved. The nature of the integrative center is unknown—it may be one particular cerebral structure, a fixed system of structures, or a number of structures involved in a probabilistic fashion. The second model hypothesizes that, in conditions of learning mediated by enhanced training (relatively high levels of positive or negative reinforcers, a high number of trials or training sessions, or some combination of these factors), those structures that were originally connected in series undergo a functional change whereby they become functionally reconnected in parallel (additional structures may become involved in this process). Consequently, even when one or several components of this circuit are damaged or do not function normally, the neural activity produced by the learning experience will be able to reach the putative integrative center, thus allowing for memory consolidation to occur (Prado-Alcalá, 1995). This model makes an important prediction, namely, that after an enhanced training experience, interference with the activity of one structure (as previously shown), as well as simultaneous interference with several structures will not produce the expected mnemonic deficiencies that are commonly seen in conditions of regular training.
Although there is a fair amount of literature germane to the participation of AMY, STR, and SN in memory processes, there are, to our knowledge, no data that determine the relative importance of these three regions for memory formation or if they participate differentially in these processes. If it turned out that there was a differential effect of typical amnesic treatments on retention, or if enhanced training did not protect against amnesia after simultaneous interference with two structures known to participate in memory, then a major revision of the serial/parallel model described would be in order. Thus, one objective of this work was to test if under identical conditions of training with low, medium, and high aversive stimulation, temporary inactivation of AMY, STR, and SN would produce the same retention deficits of IA. A second goal was to determine if concurrent inactivation of two cerebral areas (AMY + STR, AMY + SN, and STR + SN) would interfere with retention of enhanced training of IA.
Materials and Methods
All experimental procedures were approved by the Animal Ethics Committee of the Instituto de Neurobiología, Universidad Nacional Autónoma de México and complied with “Principles of laboratory animal care” of the National Institutes of Health.
Animals
The subjects were naive male Wistar rats (250–350 g) maintained in a room with a 12 h/12 h light-dark cycle (lights on at 7:00 h) and housed individually in acrylic cages with food and tap water ad libitum. The temperature of the various rooms in which they were reared and maintained was a constant 21°C. The subjects were randomly assigned to each group; sample size ranged from 10 to 12 rats per group.
Surgical Procedure
The rats were adapted to the laboratory vivarium for at least one week before surgery. They were anesthetized with sodium pentobarbital (50 mg/kg, i.p.), given atropine sulphate (1 mg/kg, i.p.), and positioned in a stereotaxic instrument (Stoelting, Co.; IL). Stainless steel guide cannulae (23 gauge) were bilaterally implanted in the STR (A = Bregma, L = ± 3.0, V = −4.0), AMY (A = −2.8, L = ± 4.5, V = −8.0), or SN (A = −5.3, L = ± 2.0, V = −7.5), or in two of these structures (Paxinos and Watson, 1997). The cannulae were fixed to the skull using two screws and dental acrylic, and a stylet was inserted into each cannula and retained there at all times except during the injections. Immediately after surgery, the animals received an intramuscular injection of penicillin (6000 units). The rats were allowed one week to recover from the surgical procedures before behavioral training, and they were handled on each of the three days preceding training to habituate them to the infusion procedure and maintain patent cannulae.
Apparatus
Training and testing were carried out in an apparatus specifically designed to study one-trial step-through IA training. The apparatus is an alley with two distinct compartments, separated by a guillotine door. The safe compartment (30 × 30 × 30 cm) had walls and lid of red-colored acrylic with a floor of aluminium bars (6 mm in diameter, 9 mm apart). This compartment was illuminated by a 10-Watt light bulb located in the center of its lid. The other, unlit compartment has side walls and floor made of stainless steel with end walls and lid made of red-colored acrylic. The compartment is 30 cm long and 25 cm deep. The walls and floor are shaped as a trough, 20 cm wide at the top and 8 cm wide at the bottom. In the middle of the floor, a 1.5 cm slot separates the two stainless steel plates that make up the walls and floor. When in this compartment, the rats are in contact with both plates, which can be electrified and, thereby, deliver aversive stimulation consistently to every subject. A square-pulse stimulator (Grass model No. S-48), in series with a constant current unit (Grass model No. CCU-1A), generated the electric shock. The duration of shock and the measurement of latencies to cross from one compartment to the other were accomplished with automated equipment. The conditioning box was located inside a dark, sound-proof room, provided with background masking noise.
Training and Testing
On the day of training, each animal was put inside the safe compartment; 10 s later the door between compartments was opened, and the latency to cross to the other compartment was measured (training latency). When the animals crossed to this compartment the door was closed and a footshock was delivered; intensities will be specified below. Five seconds later the door was reopened allowing the animal to escape to the safe compartment, and the stimulator was turned off; this latency was also measured (escape latency). After 30 s in the safe compartment the animal was put back in its home cage. Twenty-four hours later, during the retention test, the same procedure was followed except that the footshock was not delivered; if the animal did not cross to the second compartment within 600 s, a retention latency score of 600 was assigned and the session ended.
Microinjection Procedure
The bilateral infusions of 2% lidocaine or isotonic saline solution were made through 30 gauge injection needles connected to a Hamilton microsyringe by polyethylene tubing (1 μl into the STR and 0.5 μl into the AMY and SN; at the dose and volumes used here, lidocaine suppresses neuronal activity for less than two hours; Martin, 1991). The injection needles were inserted into the guide cannulae and protruded 1 mm beyond the tip of the cannulae. The infusion rate was 1.0 μl/min and was controlled by an automated microinfusion pump (WPI, model 220i). At the end of the infusion, the injection needles remained inside the guide cannulae for 60 s to insure diffusion away from the injector tip. The injection procedure was carried out within 1 min after training in a different room from that in which training and testing took place.
Histology
Upon completion of testing, all rats were anesthetized and perfused, intracardially, with isotonic saline followed by a 10% formalin solution (FS), and their brains were removed and post-fixed with a 10% FS for two days, at which time the solution was replaced with a 10% FS-30% sucrose solution until sectioning. Sections (50 μm thick) were cut on a cryostat and stained with cresyl-violet. Data from animals with cannulae tips outside the target areas were not included in the statistical analyses. Figure 1 depicts the location of cannulae tips of all successfully implanted rats.
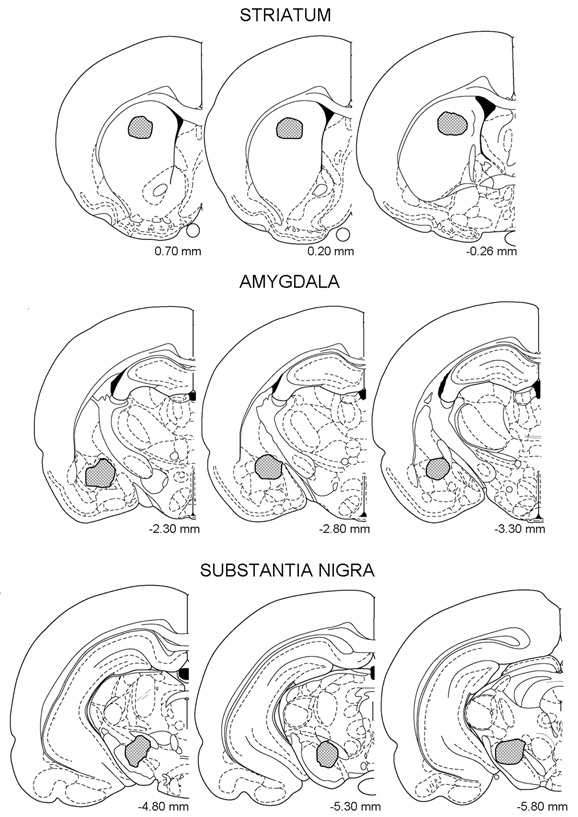
Figure 1. Hatched areas represent location of cannula tips within the striatum, amygdala, and substantia nigra. Redrawn from Paxinos and Watson, (1997).
Statistics
Because the measurement of retention was truncated at 600 s, non-parametric statistics were used in analyzing the results. Independent Kruskal–Wallis analyses of variance were computed for training, escape, and retention latencies. When appropriate, the Mann–Whitney U test was used to make comparisons between any two groups.
Experiment 1
As stated above, post-training interference with activity of STR, SN, or AMY produces marked retention deficits of IA, but enhanced training protects against these deficits (Thatcher and Kimble, 1966; Giordano and Prado-Alcalá, 1986; Pérez-Ruíz and Prado-Alcalá, 1989; Parent et al., 1992, 1994, 1995; Parent and McGaugh, 1994; Prado-Alcalá, 1995; Cobos-Zapiaín et al., 1996). It is not known, however, whether these structures are equally important for retention of IA. To put it another way, does inactivation of each of these structures produce the same degree of memory impairment? Our results indicated that such is not the case: a greater deficit in retention is produced by temporary inactivation of SN than that produced by inactivation of AMY or STR, although enhanced training overcomes such deficits, as shown below.
The rats of three independent groups that had been implanted in the STR, AMY, or SN, received a 1.0 mA footshock during training; half the rats were microinjected with lidocaine and the other half with isotonic saline.
Results
Because all the groups shared identical conditions during the training (microinjections were delivered after training), data of training and escape latencies of the six groups were analysed with separate Kruskall–Wallis analysis of variance tests, and no significant differences among the groups were found (H[5] = 3.81, p = 0.58 and H[5] = 3.645, p = 0.60, respectively). The lack of significant differences in training and escape latencies among the groups was also found in the remaining experiments.
Evaluation of the retention latencies also indicated that there were no significant differences among the saline groups (H[2] = 0.20, p = 0.90) nor among the lidocaine groups (H[2] = 0.08, p = 0.96). When pairwise comparisons were made (Mann–Whitney U test), however, highly significant differences became evident between the saline- and lidocaine-treated rats of the SN, STR, and AMY groups; Figure 2 shows these results.
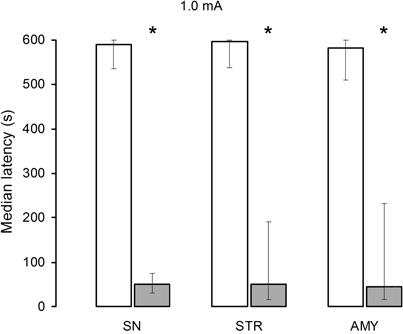
Figure 2. Median retention scores (with interquartile range) of groups of rats trained in one-trial inhibitory avoidance and injected with either isotonic saline solution (open bars) or 2% lidocaine (filled bars). Lidocaine infusion into the substantia nigra (SN), striatum (STR), or amygdala (AMY) produced amnesia in animals trained with 1.0 mA. *Denotes p < 0.05 between saline and lidocaine.
Experiment 2
Experiment 1 showed that lidocaine infusions produced a marked impairment of memory consolidation, regardless of whether the infusions had been made in the SN, STR, or AMY. The aim of this experiment was, as stated earlier, to find out if increasing the intensity of the training experience, by increasing the magnitude of the aversive stimulation, would produce a differential effect on retention of the task, depending on the cerebral structure whose neural activity had been hindered by the lidocaine. The same experimental design as that of Experiment 1 was followed, except that the intensity of the footshock delivered during training was 2.0 mA.
Results
While no significant differences in retention scores among the three saline groups were found (H[2] = 0.05, p = 0.97), the retention scores of the lidocaine groups yielded a significant treatment effect (H[2] = 9.14, p = 0.01). The U tests revealed that there were no significant differences between the effects of saline and lidocaine administration into the SN. In contrast, lidocaine produced a significant retention deficit as compared with the saline treatment in both the STR (p < 0.001) and AMY (p < 0.005) groups. Finally, pairwise comparisons between the lidocaine groups showed that the AMY and STR groups had significantly lower retention scores than the SN group (p < 0.01 and p < 0.05, respectively) while the AMY and STR groups did not differ from each other (Figure 3).
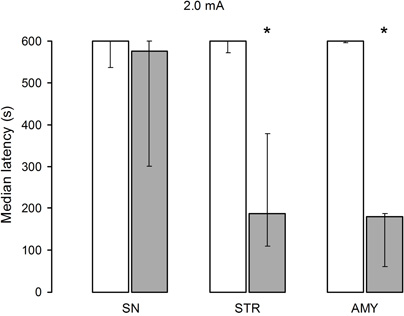
Figure 3. Median retention scores (with interquartile range) of groups of rats trained in one-trial inhibitory avoidance and injected with either isotonic saline solution (open bars) or 2% lidocaine (filled bars). Training with 2.0 mA protected memory in animals infused with lidocaine into the substantia nigra (SN), but not in those infused into the striatum (STR), or amygdala (AMY). *Denotes p < 0.05 between saline and lidocaine.
Experiment 3
In Experiment 2 it was found that a relatively high footshock during training prevented the amnesic effect of lidocaine infusion into the SN, but that amnesia was still observed when the lidocaine was infused into the STR and AMY. Previous publications had shown that enhanced training of IA protected against the effects of amnesic treatments applied to the SN (Cobos-Zapiaín et al., 1996), STR (Giordano and Prado-Alcalá, 1986; Pérez-Ruíz and Prado-Alcalá, 1989; Prado-Alcalá, 1995), and AMY (Thatcher and Kimble, 1966; Parent et al., 1992, 1994, 1995; Parent and McGaugh, 1994). It was of interest, therefore, to assess the effects of interference with the neural activity of these structures on retention after training with higher footshock intensity, as measured under the same experimental conditions. To this end, rats of three independent groups that had been implanted in the STR, AMY, or SN, received a 3.0 mA footshock during training; half the rats were microinjected with lidocaine and the other half with isotonic saline.
Results
All groups of rats displayed perfect or near-perfect retention performances, as depicted in Figure 4. Independent comparisons of retention latencies among the saline groups and of the lidocaine groups yielded non-significant differences (H[2] = 0.16, p = 0.92) and (H[2] = 0.71, p = 0.70, respectively).
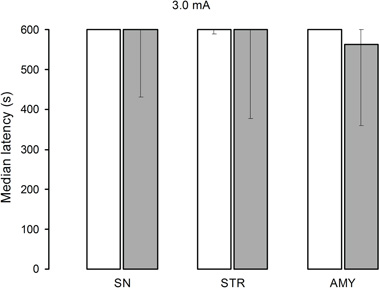
Figure 4. Median retention scores (with interquartile range) of groups of rats trained in one-trial inhibitory avoidance and injected with either isotonic saline solution (open bars) or 2% lidocaine (filled bars) into the nigra (SN), striatum (STR), or amygdala (AMY). Lidocaine was totally ineffective in altering memory when training was conducted with 3.0 mA, regardless of infusion site.
Experiment 4
An important goal of this experimental series was to empirically test the prediction of the proposed “parallel model” (Prado-Alcalá, 1995) that after an enhanced training experience, simultaneous interference with several structures will not produce the expected mnemonic deficiencies that are commonly seen in conditions of regular training. Accordingly, rats were fixed with bilateral cannulae in two structures: STR + SN, AMY + SN, or AMY + STR, and then submitted to the IA task using a 3.0 mA footshock.
Results
Just as in Experiment 3, no significant differences in retention were found among the saline (H[2] = 3.79, p = 0.15) and lidocaine (H[2] = 0.34, p = 0.84) groups. The U tests revealed that there were no significant differences between the effects of saline and lidocaine administration in any of the structures (Figure 5). It is worth noting that the retention latencies of the AMY + SN and AMY + STR groups treated with saline or lidocaine were somewhat lower than those of the saline groups of Experiments 1, 2, and 3 and the lidocaine groups trained with 3.0 mA (Figure 5). These results support the proposition that the STR and AMY are more important for long-term memory formation than the SN.
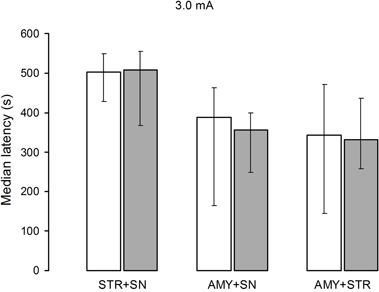
Figure 5. Median retention scores (with interquartile range) of groups of rats trained in one-trial inhibitory avoidance and injected with either isotonic saline solution (open bars) or 2% lidocaine (filled bars). No significant differences among the groups trained with 3.0 mA were found after simultaneous infusion of lidocaine or isotonic saline solution STR + SN, AMY + SN, or AMY + STR.
Discussion
The main finding of this work is that enhanced training protected memory against the amnesic effect of post-training, simultaneous inactivation of any two of three cerebral nuclei known to be involved in memory, namely the AMY, STR, and SN.
Infusion of lidocaine into the SN, STR, or AMY produced a marked memory deficit when training of IA was carried out with the lowest footshock (Figure 2). The fact that there were no significant differences in training latencies among the groups indicates that the bilateral implants of cannulae in different cerebral regions did not interfere with the motor and perceptual mechanisms necessary for performance during training. Furthermore, the lack of differences in escape latencies argues against deficiencies in sensitivity to the footshock. These latencies were not significantly different from those of a group of unimplanted rats (data not shown). Therefore, the good retention scores of the saline groups and the amnesia shown by the lidocaine groups are attributed to the differential effects of these treatments.
Previous reports had shown, independently, that relatively high intensity of footshock during IA training prevented the effect of a number of amnesic treatments administered to the SN (Cobos-Zapiaín et al., 1996), STR (Giordano and Prado-Alcalá, 1986; Pérez-Ruíz and Prado-Alcalá, 1989; Prado-Alcalá, 1995), and AMY (Thatcher and Kimble, 1966; Parent et al., 1992, 1994, 1995; Parent and McGaugh, 1994). However, there are no direct comparisons among these structures of the effect of interference with their neural activity, using different levels of aversive stimulation in training. In the present work, we made these comparisons under identical experimental conditions. We found that when the aversive stimulation was increased to 2.0 mA, memory was protected when lidocaine was injected into the SN, but there was still a significant deficit after its infusion into the STR or AMY (Figure 3). These results indicate that the STR and AMY are more critically involved than the SN in consolidation of the IA task, and that for any given structure the proposed functional transfer from serial to parallel connectivity (Prado-Alcalá, 1995) depends upon the impact of the learning experience.
Administration of lidocaine into the STR, AMY, or SN of the groups trained with 3.0 mA had no effect on long-term memory (Figure 4). More importantly, simultaneous infusion of lidocaine into the STR + SN, AMY + SN, or AMY + STR was also ineffective in altering memory formation when a 3.0 mA footshock was administered (Figure 5). These results agree well with one of the predictions of the proposed parallel model of enhanced learning (Prado-Alcalá, 1995), namely that after an enhanced training experience, simultaneous interference with several structures will not produce the expected mnemonic deficiencies commonly seen in conditions of regular training.
Not only the overall results of this study support the models described above, but their implications also go to the heart of the current notion about memory formation. The vast majority of experiments dealing with the effects on memory of interference with normal activity of the brain support the century-old theory of memory consolidation (Müller and Pilzecker, 1900). Consistently, it has been found that post-training administration of a variety of treatments shortly after a learning experience produces amnesia. This detrimental effect diminishes as the interval between learning and treatment increases, until the treatments become ineffective (McGaugh, 1966, 2000; McGaugh and Herz, 1972; Weingartner and Parker, 1984; Lechner et al., 1999). However, evidence has accumulated that does not fit the consolidation theory (Prado-Alcalá, 1995). Post-training treatments that produce amnesia of training mediated by aversive stimulation have no effect after enhanced training, independent of the amnesic agents used, and the mode of their administration. It might be argued that these agents do not produce amnesia after enhanced training because memory consolidation has been accelerated and the maximal effect of the pharmacologic agents now occurs after consolidation has taken place. Nonetheless, we have found that pre-training infusion of tetrodotoxin into the hippocampus produces amnesia of IA trained with low footshock, but long-term memory was normal when a relatively high footshock was used. These data confirm the protective effect of over-learning against experimentally induced amnesia, and suggest that the establishment of long-term memory was not due to accelerated consolidation (Garín-Aguilar et al., 2003) and that the period of consolidation may be absent in the case of enhanced training. It remains to be determined if this result can be generalized to a condition where infusions of lidocaine are made into the SN, STR, and AMY before training.
More experiments are under way to test the validity of these models. If the new data are consistent with the predictions that can be derived from them, then it will be possible to think about the brain as having at least two different ways to store learned information, depending on whether it is dealing with normal or enhanced learning.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Ángel Méndez, Andrea C. Medina, Norma Serafín, Omar González, Martín García, and Ramón Martínez Olvera for their excellent technical assistance; and Dr. Dorothy Plerss for reviewing and helping us to improve the manuscript. Supported by the National Council of Science and Technology of México (Grant CONACYT 128259).
References
Ambrogi-Lorenzini, C. G., Baldi, E., Bucherelli, C., Sacchetti, B., and Tassoni, G. (1999). Neural topography and chronology of memory consolidation: a review of functional inactivation findings. Neurobiol. Learn. Mem. 71, 1–18.
Ambrogi-Lorenzini, C. G., Baldi, E., Bucherelli, C., and Tassoni, G. (1994). Passive avoidance response distribution by post-training substantia nigra functional tetrodotoxin inactivation in the rat. Arch. Ital. Biol. 132, 85–92.
Cobos-Zapiaín, G. G., Salado-Castillo, R., Sánchez-Alavez, M., Quirarte, G. L., Roldán-Roldán, G., Díaz del Guante, M. A., and Prado-Alcalá, R. A. (1996). High level of footshock during inhibitory avoidance training prevents amnesia induced by intranigral injection of GABA antagonists. Neurobiol. Learn. Mem. 65, 202–206.
Da Cunha, C., Gevaerd, M. S., Vital, M., Miyoshi, E., Andreatini, R., Silveira, R., Takahashi, R. N., and Canteras, N. S. (2001). Memory disruption in rats with nigral lesions induced by MPTP: a model for early Parkinson's disease amnesia. Behav. Brain Res. 124, 9–18.
Da Cunha, C., Wietzikoski, S., Wietzikoski, E. C., Miyoshi, E., Ferro, M. M., Anselmo-Franci, J. A., and Canteras, N. S. (2003). Evidence for the substantia nigra pars compacta as an essential component of a memory system independent of the hippocampal memory system. Neurobiol. Learn. Mem. 79, 236–242.
Díaz del Guante, M. A., Rivas, M., Prado-Alcalá, R. A., and Quirarte, G. L. (2004). Amnesia produced by pre-training infusion of serotonin into the substantia nigra. Neuroreport 15, 2527–2529.
Garín-Aguilar, M. E., Quirarte, G. L., and Prado-Alcalá, R. A. (2003). Effect of Pre-training Intrahippocampal TTX on Overlearning. Program No. 938.8. Neuroscience Meeting Planner, New Orleans, LA: Soc. for Neurosci. Online.
Giordano, M., and Prado-Alcalá, R. A. (1986). Retrograde amnesia induced by post-trial injection of atropine into the caudate-putamen. Protective effect of the negative reinforcer. Pharmacol. Biochem. Behav. 24, 905–909.
Izquierdo, I., Bevilaqua, L. R., Rossato, J. I., Bonini, J. S., Da Silva, W. C., Medina, J. H., and Cammarota, M. (2006). The connection between the hippocampal and the striatal memory systems of the brain: a review of recent findings. Neurotox. Res. 10, 113–121.
Kim, H. J., and Routtenberg, A. (1976). Retention disruption following post-trial picrotoxin injection into the substantia nigra. Brain Res. 113, 620–625.
Lechner, H. A., Squire, L. R., and Byrne, J. H. (1999). 100 years of consolidation – remembering Müller and Pilzecker. Learn. Mem. 6, 77–87.
Liang, K. C., McGaugh, J. L., Martinez, J. L., Jensen, R. A., Vasquez, B. J., and Messing, R. B. (1982). Post-training amygdaloid lesions impair retention of an inhibitory avoidance response. Behav. Brain Res. 4, 237–249.
Martin, J. H. (1991). Autoradiographic estimation of the extent of reversible inactivation produced by microinjection of lidocaine and muscimol in the rat. Neurosci. Lett. 127, 160–164.
McGaugh, J. L. (2002). Memory consolidation and the amygdala: a systems perspective. Trends Neurosci. 25, 456–461.
McGaugh, J. L., and Herz, M. J. (1972). Memory Consolidation. San Francisco, CA: Albion Publishing Company.
Miyoshi, E., Wietzikoski, E. C., Bortolanza, M., Boschen, S. L., Canteras, N. S., Izquierdo, I., Da Cunha, C. (2012). Both the dorsal hippocampus and the dorsolateral striatum are needed for rat navigation in the Morris water maze. Behav. Brain Res. 226, 171–178.
Müller, G. E., and Pilzecker, A. (1900). Experimentelle beiträge zur lehre vom gedächtnis. Z. Psychol. 1, 1–288.
Packard, M. G., and Knowlton, B. J. (2002). Learning and memory functions of the basal ganglia. Annu. Rev. Neurosci. 25, 563–593.
Parent, M. B., and McGaugh, J. L. (1994). Posttraining infusion of lidocaine into the amygdala basolateral complex impairs retention of inhibitory avoidance training. Brain Res. 661, 97–103.
Parent, M. B., Quirarte, G. L., Cahill, L., and McGaugh, J. L. (1995). Spared retention of inhibitory avoidance learning after posttraining amygdala lesions. Behav. Neurosci. 109, 803–807.
Parent, M. B., Tomaz, C., and McGaugh, J. L. (1992). Increased training in an aversively motivated task attenuates the memory-impairing effects of posttraining N-methyl-D-aspartate-induced amygdala lesions. Behav. Neurosci. 106, 789–797.
Parent, M. B., West, M., and McGaugh, J. L. (1994). Memory of rats with amygdala lesions induced 30 days after footshock-motivated escape training reflects degree of original training. Behav. Neurosci. 108, 1080–1087.
Paxinos, G., and Watson, C. (1997). The Rat Brain in Stereotaxic Coordinates. San Diego, CA: Academic Press.
Pérez-Ruíz, C., and Prado-Alcalá, R. A. (1989). Retrograde amnesia induced by lidocaine injection into the striatum: protective effect of the negative reinforcer. Brain Res. Bull. 22, 599–603.
Power, A. E., Roozendaal, B., and McGaugh, J. L. (2000). Glucocorticoid enhancement of memory consolidation in the rat is blocked by muscarinic receptor antagonism in the basolateral amygdala. Eur. J. Neurosci. 12, 3481–3487.
Prado-Alcalá, R. A. (1995). “Serial and parallel processing during memory consolidation,” in Plasticity in the Central Nervous System: Learning and Memory, eds McGaugh, J. L., Bermúdez-Rattoni, F., Prado-Alcalá, R. A. (New Jersey: Erlbaum), 57–65.
Prado-Alcalá, R. A., and Cobos-Zapiaín, G. C. (1977). Learning deficits induced by cholinergic blockade of the caudate nucleus as a function of experience. Brain Res. 138, 190–196.
Prado-Alcalá, R. A., and Cobos-Zapiaín, G. G. (1979). Interference with caudate nucleus activity by potassium chloride. Evidence for a ‘moving’ engram. Brain Res. 172, 577–583.
Prado-Alcalá, R. A., Grinberg-Zylberbaum, J., Alvarez-Leefmans, J., and Brust-Carmona, H. (1973). Suppression of motor conditioning by the injection of 3 M KCl in the caudate nuclei of cats. Physiol. Behav. 10, 59–64.
Prado-Alcalá, R. A., Grinberg-Zylberbaum, J., Alvarez-Leefmans, J., Gómez, A., Singer, S., and Brust-Carmona, H. (1972). A possible caudate-cholinergic mechanism in two instrumental conditioned responses. Psychopharmacologia 25, 339–346.
Prado-Alcalá, R. A., Kaufmann, P., and Moscona, R. (1980). Scopolamine and KCl injections into the caudate nucleus. Overtraining-induced protection against deficits of learning. Pharmacol. Biochem. Behav. 12, 249–253.
Roesler, R., Vianna, M. R., De-Paris, F., Quevedo, J., Walz, R., and Bianchin, M. (2000). Infusions of AP5 into the basolateral amygdala impair the formation, but not the expression, of step-down inhibitory avoidance. Braz. J. Med. Biol. Res. 33, 829–834.
Roozendaal, B., McEwen, B. S., and Chattarji, S. (2009). Stress, memory and the amygdala. Nat. Rev. Neurosci. 10, 423–433.
Routtenberg, A., and Holzman, N. (1973). Memory disruption by electrical stimulation of substantia nigra, pars compacta. Science 181, 83–96.
Salado-Castillo, R., Díaz del Guante, M. A., Alvarado, R., Quirarte, G. L., and Prado-Alcalá, R. A. (1996). Effects of regional GABAergic blockade of the striatum on memory consolidation. Neurobiol. Learn. Mem. 66, 102–108.
Sanberg, P. R., Lehmann, J., and Fibiger, H. C. (1978). Impaired learning and memory after kainic acid lesions of the striatum: and a behavioral model of Huntington's disease. Brain Res. 149, 546–551.
Thatcher, R. W., and Kimble, D. P. (1966). Effect of amygdaloid lesions of retention of an avoidance response in overtrained and non-overtrained rats. Psychon. Sci. 6, 9–10.
Weingartner, H., and Parker, E. S. (1984). Memory Consolidation. Psychobiology of Cognition. Hillsdale, NJ: Lawrence Erlbaum Associates.
White, N. M. (2009). Some highlights of research on the effects of caudate nucleus lesions over the past 200 years. Behav. Brain Res. 199, 3–23.
Keywords: memory consolidation, amnesia, TTX, inhibitory avoidance, amygdala, striatum, substantia nigra
Citation: Salado-Castillo R, Sánchez-Alavéz M, Quirarte GL, Martínez García MI and Prado-Alcalá RA (2011) Enhanced training protects memory against amnesia produced by concurrent inactivation of amygdala and striatum, amygdala and substantia nigra, or striatum and substantia nigra. Front. Behav. Neurosci. 5:83. doi: 10.3389/fnbeh.2011.00083
Received: 28 October 2011; Paper pending published: 18 November 2011;
Accepted: 02 December 2011; Published online: 20 December 2011.
Edited by:
Antonella Gasbarri, University of l'Aquila, ItalyReviewed by:
Antonella Gasbarri, University of l'Aquila, ItalyIván Izquierdo, Pontifícia Universidade Católica do Rio Grande do Sul, Brazil
Selva Rivas-Arancibia, Universidad Nacional Autónoma de México, Mexico
Copyright: © 2011 Salado-Castillo, Sánchez-Alavez, Quirarte, Martínez García and Prado-Alcalá. This is an open-access article distributed under the terms of the Creative Commons Attribution Non Commercial License, which permits non-commercial use, distribution, and reproduction in other forums, provided the original authors and source are credited.
*Correspondence: Roberto A. Prado-Alcalá, Departamento de Neurobiología Conductual y Cognitiva, Instituto de Neurobiología, Universidad Nacional Autónoma de México, Campus UNAM-Juriquilla, Boulevard Juriquilla # 3001, Querétaro, Qro., 76230, México. e-mail:cHJhZG9Ac2Vydmlkb3IudW5hbS5teA==