 Yu-Chen Chen1,2
Yu-Chen Chen1,2 Huiyou Chen2Liang Jiang2Fan Bo2Jin-Jing Xu3Cun-Nan Mao2
Huiyou Chen2Liang Jiang2Fan Bo2Jin-Jing Xu3Cun-Nan Mao2 Richard Salvi4Xindao Yin2
Richard Salvi4Xindao Yin2 Guangming Lu1*Jian-Ping Gu2*
Guangming Lu1*Jian-Ping Gu2*- 1Department of Medical Imaging, Jinling Hospital, School of Medicine, Nanjing University, Nanjing, China
- 2Department of Radiology, Nanjing First Hospital, Nanjing Medical University, Nanjing, China
- 3Department of Otolaryngology, Nanjing First Hospital, Nanjing Medical University, Nanjing, China
- 4Center for Hearing and Deafness, University at Buffalo, State University of New York, Buffalo, NY, United States
Purpose: Presbycusis, age-related hearing loss, is believed to involve neural changes in the central nervous system, which is associated with an increased risk of cognitive impairment. The goal of this study was to determine if presbycusis disrupted spontaneous neural activity in specific brain areas involved in auditory processing, attention and cognitive function using resting-state functional magnetic resonance imaging (fMRI) approach.
Methods: Hearing and resting-state fMRI measurements were obtained from 22 presbycusis patients and 23 age-, sex- and education-matched healthy controls. To identify changes in spontaneous neural activity associated with age-related hearing loss, we compared the amplitude of low-frequency fluctuations (ALFF) and regional homogeneity (ReHo) of fMRI signals in presbycusis patients vs. controls and then determined if these changes were linked to clinical measures of presbycusis.
Results: Compared with healthy controls, presbycusis patients manifested decreased spontaneous activity mainly in the superior temporal gyrus (STG), parahippocampal gyrus (PHG), precuneus and inferior parietal lobule (IPL) as well as increased neural activity in the middle frontal gyrus (MFG), cuneus and postcentral gyrus (PoCG). A significant negative correlation was observed between ALFF/ReHo activity in the STG and average hearing thresholds in presbycusis patients. Increased ALFF/ReHo activity in the MFG was positively correlated with impaired Trail-Making Test B (TMT-B) scores, indicative of impaired cognitive function involving the frontal lobe.
Conclusions: Presbycusis patients have disrupted spontaneous neural activity reflected by ALFF and ReHo measurements in several brain regions; these changes are associated with specific cognitive performance and speech/language processing. These findings mainly emphasize the crucial role of aberrant resting-state ALFF/ReHo patterns in presbycusis patients and will lead to a better understanding of the neuropathological mechanisms underlying presbycusis.
Introduction
Presbycusis, or age-related hearing loss, is the most common hearing disorder and a major cause of chronic disability in the elderly, which can lead to social isolation, communication, language and speech processing problems and depression (Davis, 1991; Gates and Mills, 2005; Dubno et al., 2008; Sprinzl and Riechelmann, 2010; Rosenhall et al., 2011; Panza et al., 2015). Age-related hearing loss not only disrupts inputs to the primary auditory cortex and receptive language areas, but also language association areas that assign meaning to words and other meaningful sounds (Salvi et al., 2002; Ardila et al., 2016). Almost two-thirds of elderly Americans have age-related hearing loss consistent with comparable trends around the globe (Lin et al., 2011b; Feder et al., 2015). While age-related hearing loss results in large part from the loss of sensory hair cell and neurons in the cochlea, the sensory organ for hearing, there is growing awareness that presbycusis is also associated with structural and functional changes in the central auditory pathway as well as other regions in the central nervous system (Kazee et al., 1995; Spongr et al., 1997; Salvi et al., 2002; Ouda et al., 2015). In the auditory midbrain of rats with age-related hearing loss, sound evoked firing rates were reduced and tuning was broader (Palombi and Caspary, 1996). However, in primary auditory cortex of rats and nonhuman primates with age-related hearing loss, sound-evoked and spontaneous activity were increased, but temporal processing and response selectivity were degraded (Hughes et al., 2010; Chi-Wing and Recanzone, 2017). Changes in spontaneous rate are not uniform across the auditory cortex; while those in primary auditory cortex decrease following noise-induced hearing loss, those in the anterior auditory field did not change while those in secondary auditory cortex decreased (Kimura and Eggermont, 1999). Age-related hearing loss also disrupts the tonotopic organization of the central auditory pathway (Willott, 1991). Consistent with animal studies, sound-evoked functional magnetic resonance imaging (fMRI) response in auditory cortex of subjects with mild or more severe presbycusis were greater than young controls and these differences were more pronounced in the right temporal lobe of the elderly (Profant et al., 2015). Others have reported decreased activation of the contralateral auditory cortex and a reduced right-ear advantage with aging and presbycusis (Chen et al., 2016). Some fMRI studies have reported that age-related hearing loss increases the functional connectivity between the auditory cortex and right motion sensitive visual area during task and resting-state conditions (Puschmann and Thiel, 2017).
Data from large population-based longitudinal studies have shown that presbycusis is associated with an increased risk of cognitive impairment and incident dementia, which primarily manifests as declining executive function (Lin et al., 2011a, b; Gurgel et al., 2014), verbal memory (Tay et al., 2006; Lin et al., 2011a), episodic and semantic long-term memory (Ronnberg et al., 2011) and psychomotor processing (Lin et al., 2011b). However, the pathophysiological mechanisms underlying presbycusis and its association with cognitive impairments remain poorly understood.
Neuroimaging technique has been applied to investigate the anatomical and functional alterations in the brains of patients with presbycusis (Mudar and Husain, 2016). Gray matter (GM) atrophy and white matter (WM) lesions, common structural abnormalities observed in previous studies, are modestly linked with presbycusis-related cognitive decline (Peelle et al., 2011; Eckert et al., 2012; Lin et al., 2014; Profant et al., 2014; Ma et al., 2016). Gamma-aminobutyric acid (GABA) levels in the auditory cortex, an important inhibitory neurotransmitter that can be assessed by magnetic resonance spectroscopy (MRS), were negatively correlated with the degree of age-related hearing loss (Gao et al., 2015). Nonetheless, little is known about the complex neurophysiological changes in the central nervous system likely to be associated with presbycusis. Neural abnormalities have been detected in populations at risk for developing cognitive impairment (Machulda et al., 2011). Therefore, measures of neural activity could conceivably be used to detect and track the possible effects of presbycusis on brain function.
Most prior studies have used fMRI to examine the effects of hearing loss on the brain activity during auditory tasks in presbycusis patients (Peelle et al., 2011; Husain et al., 2014; Profant et al., 2015; Chen et al., 2016; Puschmann and Thiel, 2017). Resting-state fMRI (rs-fMRI) is a powerful tool for evaluating spontaneous neural activity (Mantini et al., 2007) and rs-fMRI has been used to investigate brain functional connectivity alterations in the default mode network and dorsal attention network in middle-aged participants with hearing loss compared to controls (Husain et al., 2014). A recent rs-fMRI study found that hearing loss modulated cross-modal functional connectivity between visual and auditory sensory cortices in presbycusis (Puschmann and Thiel, 2017), but did not evaluate other measures of resting-state neural activity. As the effects of hearing loss on the brain may be global, a whole-brain analysis of neural function would likely identify other central processing deficits linked to presbycusis.
Amplitude of low-frequency fluctuation (ALFF) and regional homogeneity (ReHo) are two important data-driven algorithms for analyzing global rs-fMRI signals. ALFF measures the amplitude of very low-frequency oscillations of the BOLD signal at the single-voxel level (Zang et al., 2007) while ReHo analyses the neural synchronization of a given voxel with its adjacent voxels, i.e., local neural synchrony (Zang et al., 2004). ReHo may be more sensitive at detecting regional brain abnormalities than ALFF. On the other hand, ALFF may be more useful than ReHo for investigating global changes in spontaneous neural activity (An et al., 2013; Zhang et al., 2015). Thus, the combination of ALFF and ReHo may provide a more comprehensive pathophysiological assessment of human brain dysfunction than either method alone. Therefore, we took advantage of the global and local analytical power of ALFF and ReHo to identify aberrant spontaneous neural activity in presbycusis patients. Based on prior human and animal studies noted above, we hypothesized that aberrant ALFF and ReHo values would be detected in specific brain regions involved in complex auditory processing (e.g., speech or language comprehension), attention and cognitive function.
Materials and Methods
Subjects
All the subjects provided written informed consent before their participation in the study protocol, which was approved by the Research Ethics Committee of the Nanjing Medical University. A total of 22 presbycusis patients with disease duration of 3–10 years were recruited for this study at the Department of Otolaryngology of Nanjing First Hospital. Hearing thresholds were determined by pure tone audiometry (PTA) at the frequencies of 0.25, 0.5, 1, 2, 4 and 8 kHz. The criteria for the estimation of the disease duration is the results of average PTA. Average PTA values of ≤25 dB hearing level (HL) were regarded as within the limit for normal hearing thresholds (Lin et al., 2011b). For the presbycusis group, audiometric thresholds inclusion criteria were average PTA >25 dB HL in the better hearing ear and age ≧60 years. Twenty-three age, sex, and education-matched healthy subjects with no hearing loss (average PTA ≦25 dB in the better-hearing ear) were recruited through community health screenings or newspaper advertisements. All subjects were right-handed and completed at least 8 years of education. Tympanometry was performed with a Madsen Electronics Zodiac 901 Middle Ear Analyzer (GN Otometrics) to confirm normal middle-ear function. None of the participants were excluded from the study because of excessive head movement during MR scanning. Exclusion criteria were ear diseases that affect hearing thresholds and sensorineural hearing loss other than presbycusis. Participants were excluded from the study if they suffered from tinnitus, hyperacusis, Meniere’s diseases, or had a past history of otologic surgery, ototoxic drug therapy, noise exposure, or hearing aid use severe smoking, stroke, alcoholism, brain injury, Parkinson’s disease, Alzheimer’s disease (AD), epilepsy, major depression, neurological or psychiatric disorders that could affect cognitive function, major medical illness (e.g., anemia, thyroid dysfunction and cancer), MRI contraindications, or severe visual loss. The characteristics of the presbycusis patients and healthy subjects are summarized in Table 1. All the participants were randomly assigned under double-blind conditions for further data analysis.
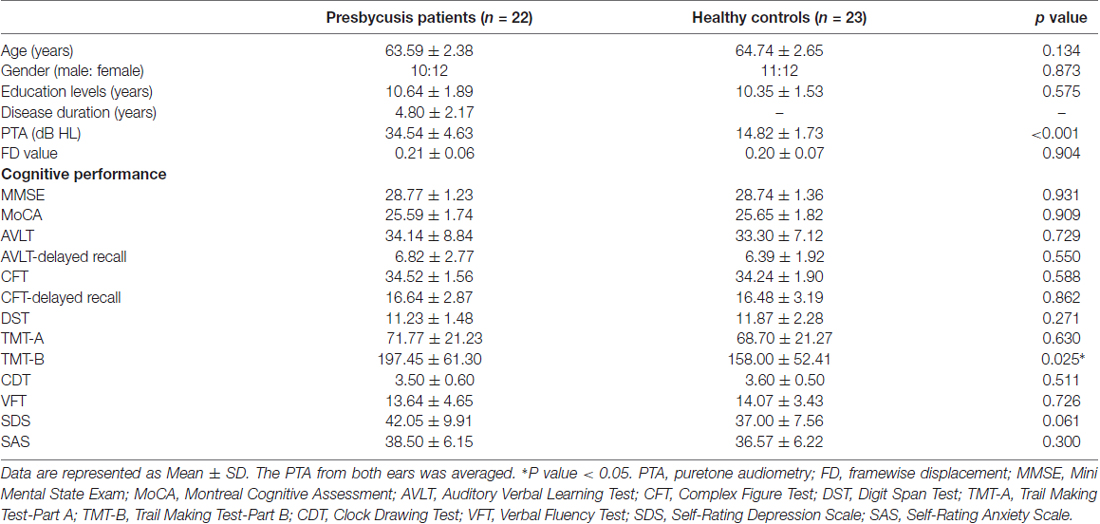
Table 1. Demographics, clinical and cognitive characteristics of the presbycusis patients and healthy controls.
Neuropsychological Assessment
All participants underwent a battery of neuropsychological tests that covered relevant cognitive domains. The neuropsychological status of the participants was established using the Mini Mental State Exam (MMSE; Galea and Woodward, 2005), Montreal Cognitive Assessment (MoCA; Nasreddine et al., 2005), Auditory Verbal Learning Test (AVLT; Schmidt, 1996), Complex Figure Test (CFT; Shin et al., 2006), Digit Span Test (DST; Hale et al., 2002), Trail-Making Test (TMT) A and B (Bowie and Harvey, 2006), Clock-Drawing Test (CDT; Samton et al., 2005) and Verbal Fluency Test (VFT; Brucki and Rocha, 2004). The tests provided an assessment reflecting the general cognitive function, episodic verbal and visual memory, semantic memory, attention, psychomotor speed, executive function and visuospatial skills. Depression and anxiety status were assessed according to the Self-Rating Depression Scale (SDS) and Self-Rating Anxiety Scale (SAS; Zung, 1971, 1986). It took approximately 60 min for each individual to complete all the tests in a fixed order.
MRI Acquisition
MRI data were acquired at using a 3.0 T MRI scanner (Ingenia, Philips Medical Systems, Netherlands) with an 8-channel receiver array head coil. Head motion and scanner noise were reduced using foam padding and earplugs. The earplugs (Hearos Ultimate Softness Series, USA) were used to attenuate scanner noise by approximately 32 dB based on the manufacture’s data sheet. Subjects were instructed to lie quietly with their eyes closed without falling asleep, not think of anything in particular, and avoid any head motion during the scan. Functional images were obtained axially using a gradient echo-planar imaging sequence as follows: repetition time (TR) = 2000 ms; echo time (TE) = 30 ms; slices = 36; thickness = 4 mm; gap = 0 mm; flip angle (FA) = 90°; field of view (FOV) = 240 mm × 240 mm; acquisition matrix = 64 × 64; voxel size = 3.75 × 3.75 × 4.0 mm3. The fMRI sequence took 8 min and 8 s. Structural images were acquired with a three-dimensional turbo fast echo (3D-TFE) T1WI sequence with high resolution as follows: TR/TE = 8.2/3.8 ms; slices = 170; thickness = 1 mm; gap = 0 mm; FA = 8°; acquisition matrix = 256 × 256; FOV = 256 mm × 256 mm. The structural sequence took 5 min and 29 s.
Functional Data Preprocessing
Functional data analyses were conducted using Data Processing Assistant for Resting-State fMRI (DPARSF) programs (Yan et al., 2016) based on statistical parametric mapping (SPM8)1 and resting-state fMRI data analyses toolkits (REST)2. A total of 240 volumes were scanned, and the first 10 volumes were discarded to allow for signal equilibrium of the initial magnetic resonance signals and adaptation of the subjects to scanner. The remaining 230 consecutive volumes were used for data analysis. Afterwards, the following procedures were carried out as follows: slice-timing adjustment, realignment for head-motion correction, spatial normalization to the Montreal Neurological Institute (MNI) template (resampling voxel size = 3 × 3 × 3 mm3) as well as linear trending and band-pass filtering (0.01–0.08 Hz). Any subjects with a head motion >2.0 mm translation or 2.0° rotation in any direction were excluded.
ALFF and ReHo Analyses
For ALFF analysis, the images were first smoothed with a Gaussian kernel of 6 mm full-width at half maximum (FWHM). Next, the time courses were first transformed to the frequency domain using a Fast Fourier Transform algorithm. The square root was then computed at each frequency of the power spectrum. ALFF values were obtained after calculation at each frequency of the power spectrum across 0.01–0.08 Hz at each voxel. For standardization purposes, the ALFF of each voxel was divided using the global mean ALFF value. ALFF was finally calculated using the REST software through the procedure described in previous studies (Zang et al., 2007).
For ReHo analysis, the images were calculated for Kendall’s coefficient of concordance of the time series of a given voxel with its 26 nearest neighboring voxels. The ReHo maps were then spatially smoothed with a Gaussian kernel of 6 mm. The ReHo value of each voxel was standardized by dividing the raw value by the global mean ReHo value, which was obtained with a similar calculation used to determine the global man ALFF value. ReHo was also calculated using the REST software through the procedure described in previous studies (Zang et al., 2004).
Structural Analysis
Structural images were processed using the VBM toolbox software in SPM81. Briefly, the structural images were normalized and segmented into GM, WM and cerebrospinal fluid (CSF) using the unified segmentation model in SPM8. Brain parenchyma volume was calculated as the sum of GM and WM volumes. The GM images were spatially smoothed using a Gaussian kernel of 8 mm FWHM. The voxel-wise GM volume was used in the following statistical analysis as covariates for ALFF and ReHo calculations.
Statistical Analysis
Demographic and clinical variables and cognitive performance scores were compared between the two groups using SPSS software (version 20.0; SPSS, Inc., Chicago, IL, USA). An independent two-sample t test was used for continuous variables, and a χ2 test was used for proportions. p < 0.05 were considered to be statistically significant.
Within-Group Analysis
For within-group whole brain ALFF and ReHo patterns, one-sample t-tests were performed on the individual ALFF and ReHo maps in a voxel-wise manner for patient and healthy control group. Significant thresholds were corrected using false discovery rate (FDR) criterion and the significance set at p < 0.001. The group-level ALFF and ReHo maps were then visualized with the REST Slice Viewer in REST software.
Between-Group Analysis
To investigate between-group differences of ALFF and ReHo values, two-sample t-tests were performed with REST software (within a GM mask). Age, sex, education levels and mean hearing thresholds were imported as covariates. To exclude potential effects of GM volume differences, the voxel-wise GM volume maps were also obtained as covariates. Multiple comparison correction was performed using FDR criterion and the significance set at p < 0.001 according to the suggestion from the prior study (Eklund et al., 2016). The results were visualized using the REST Slice Viewer.
Correlation Analysis
To identify associations between regional ALFF and ReHo abnormalities, a bivariate correlation was performed between these two measurements. The average ALFF and ReHo values of brain regions with significant differences were individually extracted and correlated with one another.
To investigate the relationship among ALFF/ReHo values of the peak voxels, neuropsychological performances and HLs, Pearson’s correlative analyses were performed using SPSS software. Partial correlations were calculated using the same covariates as those controlled in the two-sample t-tests. p < 0.05 was considered to be statistically significant. Bonferroni correction was used for multiple comparisons in the correlation analyses.
Since micromovements from volume to volume can influence the spontaneous neuronal activity (Power et al., 2012), framewise displacement (FD) values were computed for each subject to reflect the temporal derivative of the movement parameters. No subjects had FD > 0.5 mm on greater than 35 volumes in this study. No significant difference was found in the mean FD values between presbycusis patients and controls (Table 1).
Results
Demographic and Neuropsychological Characteristics
Table 1 summarizes the demographic measures and neuropsychological test results of presbycusis patients and healthy controls. Since no significant differences in PTA between the left and right ears were observed in the presbycusis group and the control group, the thresholds of both ears were averaged and are presented for each group. The PTA was significantly higher in presbycusis patients than in healthy controls (p < 0.001, 1–8 kHz). In the presbycusis group, the mean hearing thresholds were >20 dB HL at 1 kHz and reached 38.0 dB HL at 2 kHz; 48.5 dB HL at 4 kHz and 58.9 dB HL at 8 kHz (Figure 1). In the control group, the mean hearing thresholds were <20 dB HL at all frequencies. All patients had a type-A curve on tympanometry, which indicated normal middle-ear function. On cognitive tests, presbycusis patients performed significantly poorer on Trail-Making Test B (TMT-B; p < 0.05). On the other neuropsychological tests, presbycusis patients have slightly lower scores than controls, but the differences were not significantly different.
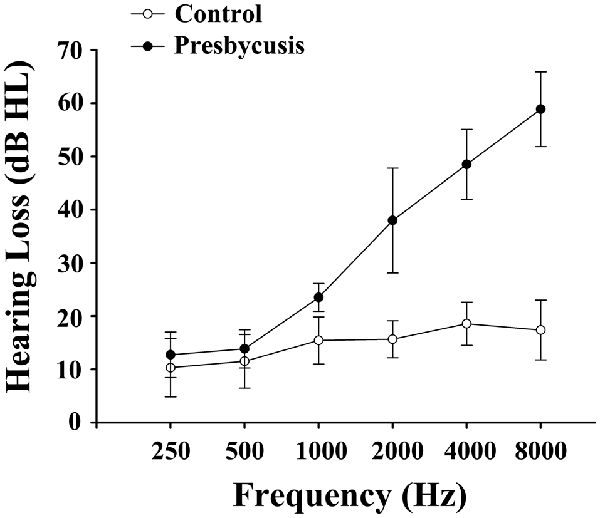
Figure 1. Average hearing thresholds of the presbycusis and control groups. The hearing thresholds were significantly higher in presbycusis patients than in healthy controls (p < 0.001, 1–8 kHz). In the presbycusis group, the mean hearing thresholds were >20 dB hearing level (HL) at 1 kHz and reached 38.0 dB HL at 2 kHz; 48.5 dB HL at 4 kHz and 58.9 dB HL at 8 kHz. Data are presented as mean ± SD.
Structural Results
Table 2 shows no significant difference in GM, WM or brain parenchyma volume in patients with presbycusis compared to healthy controls. After Monte Carlo simulation correction, no suprathreshold voxel-wise difference in the GM and WM volume between the presbycusis patients and healthy controls was observed. None of the participants in this study were excluded due to severe atrophy.

Table 2. Comparisons of the brain volumes between tinnitus patients and healthy controls.
ALFF and ReHo Analyses
Figure 2 shows the standardized ALFF (Figure 2A) and ReHo (Figure 2B) heat map for the control group and presbycusis group. ALFF and ReHo values were significantly higher than the global means values mainly in the superior temporal gyrus (STG), precuneus, posterior cingulate cortex (PCC), cuneus and medial prefrontal gyrus. Compared with healthy controls, presbycusis patients had significantly decreased ALFF values in the left STG, right parahippocampal gyrus (PHG) and right precuneus, but increased ALFF values in the left middle frontal gyrus (MFG), left cuneus and left postcentral gyrus (PoCG; Figure 3A, Table 3). Compared to healthy controls, ReHo values in presbycusis patients were significantly decreased in the left STG, right PHG and right inferior parietal lobule (IPL), but significantly increased in the left MFG, left cuneus and right PoCG (Figure 3B, Table 3).
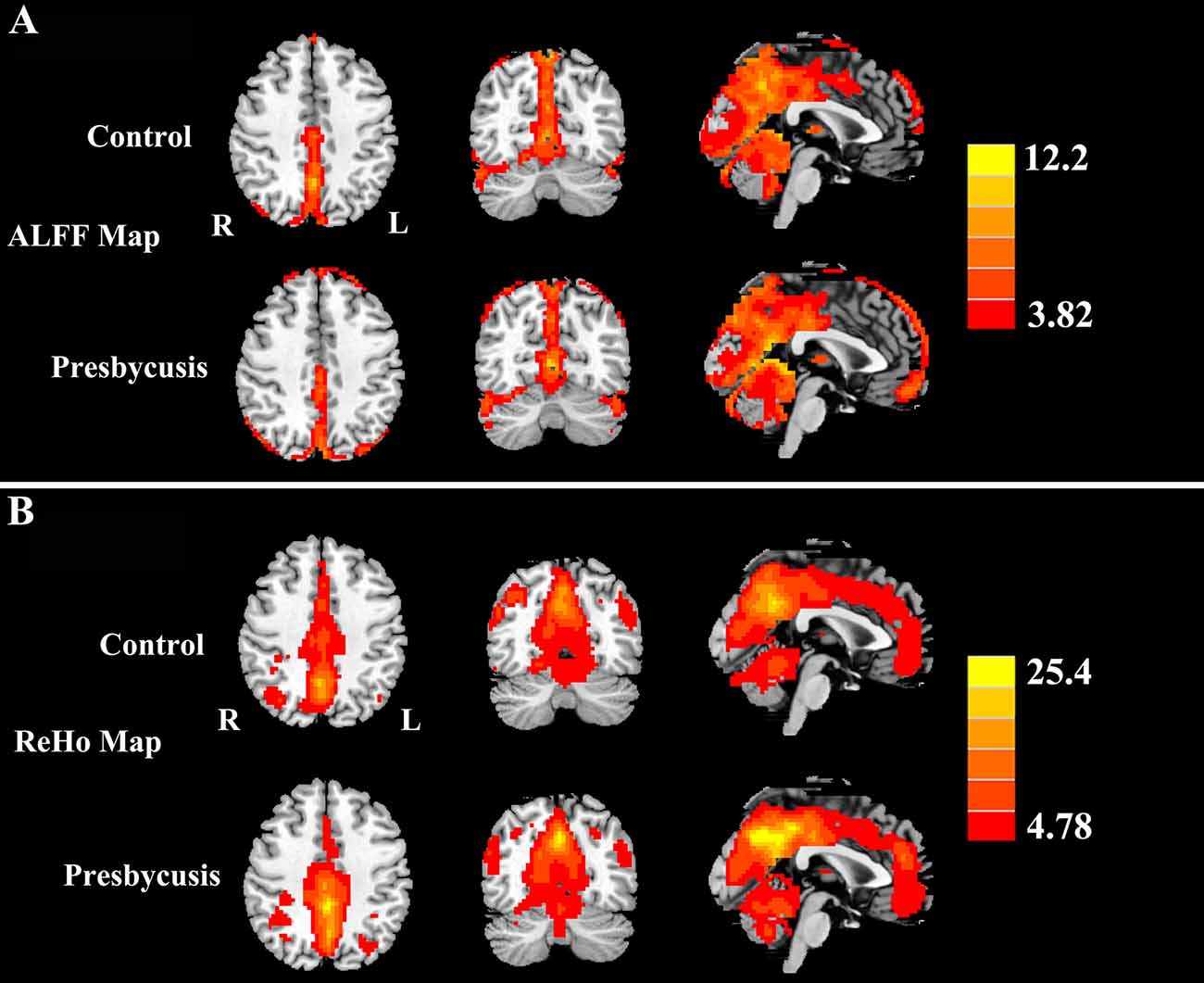
Figure 2. (A) One-sample t-test results of amplitude of low-frequency fluctuations (ALFF) maps in presbycusis and controls groups (p < 0.001 corrected by false discovery rate, FDR). (B) One-sample t-test results of regional homogeneity (ReHo) maps in presbycusis and controls groups (p < 0.001 corrected by FDR). The ALFF and ReHo values were significantly higher than the global means values mainly in the superior temporal gyrus (STG), precuneus, posterior cingulate cortex (PCC), cuneus and medial prefrontal gyrus. Left side corresponds to the right hemisphere.
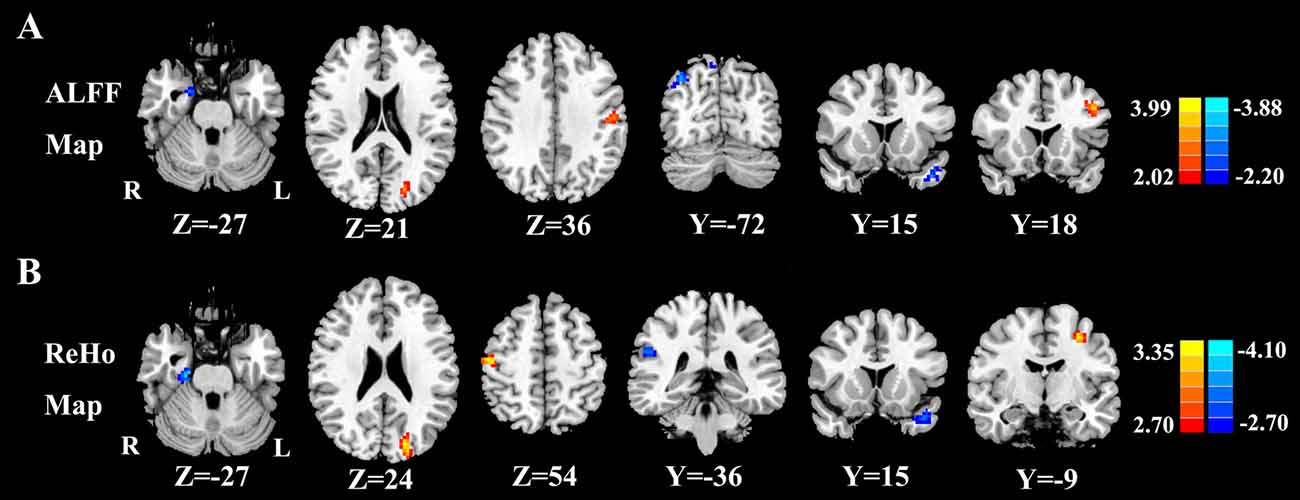
Figure 3. (A) Compared with healthy controls, presbycusis patients showed significantly decreased ALFF values in the left STG, right parahippocampal gyrus (PHG) and right precuneus, but increased ALFF values in the left middle frontal gyrus (MFG), left cuneus and left postcentral gyrus (PoCG; p < 0.001 corrected by FDR). (B) Compared with healthy controls, presbycusis patients had significantly decreased ReHo values in the left STG, right PHG and right inferior parietal lobule (IPL), but increased ReHo values in the left MFG, left cuneus and right PoCG. Left side corresponds to the right hemisphere.
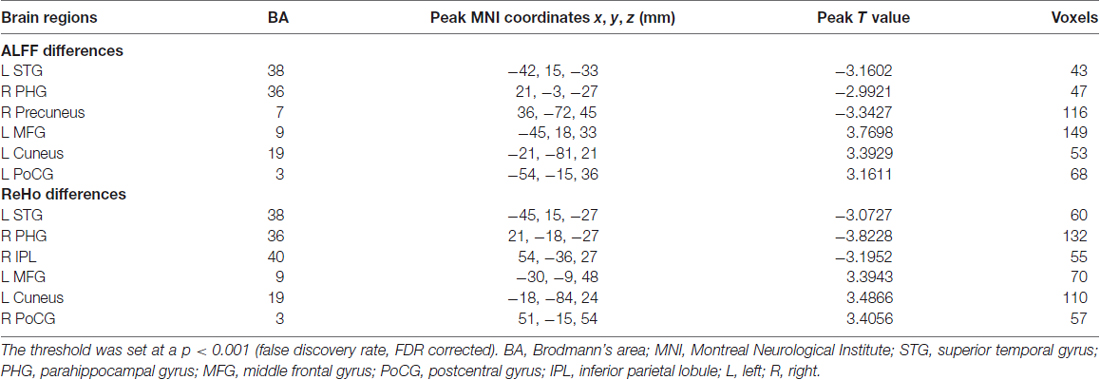
Table 3. Regions showing significant differences in amplitude of low-frequency fluctuations (ALFF) and regional homogeneity (ReHo) values between presbycusis patients and healthy controls.
Correlation Analyses
A bivariate correlation analysis indicated that regional ALFF and ReHo values extracted from the left STG (r = 0.654, p < 0.001), right PHG (r = 0.605, p = 0.003), and left cuneus (r = 0.770, p < 0.001) were strongly associated with each other in the presbycusis patients (Figure 4). After correcting for age, sex, education and GM volume, the ALFF (Figure 5A) and ReHo (Figure 5B) values in the left STG were negatively correlated with the mean PTA values in the presbycusis patients (r = −0.520, p = 0.023; r = −0.487, p = 0.035, respectively). Among the cognitive measures, TMT-B scores were positively correlated with the ALFF (Figure 5C) and ReHo (Figure 5D) values in the left MFG (r = 0.538, p = 0.021; r = 0.645, p = 0.004, respectively), corrected for age, sex, education, GM volume and mean PTA values. The other significant decreases or increases in ALFF/ReHo values were independent (i.e., no significant correlations) of hearing loss or any other cognitive tests.
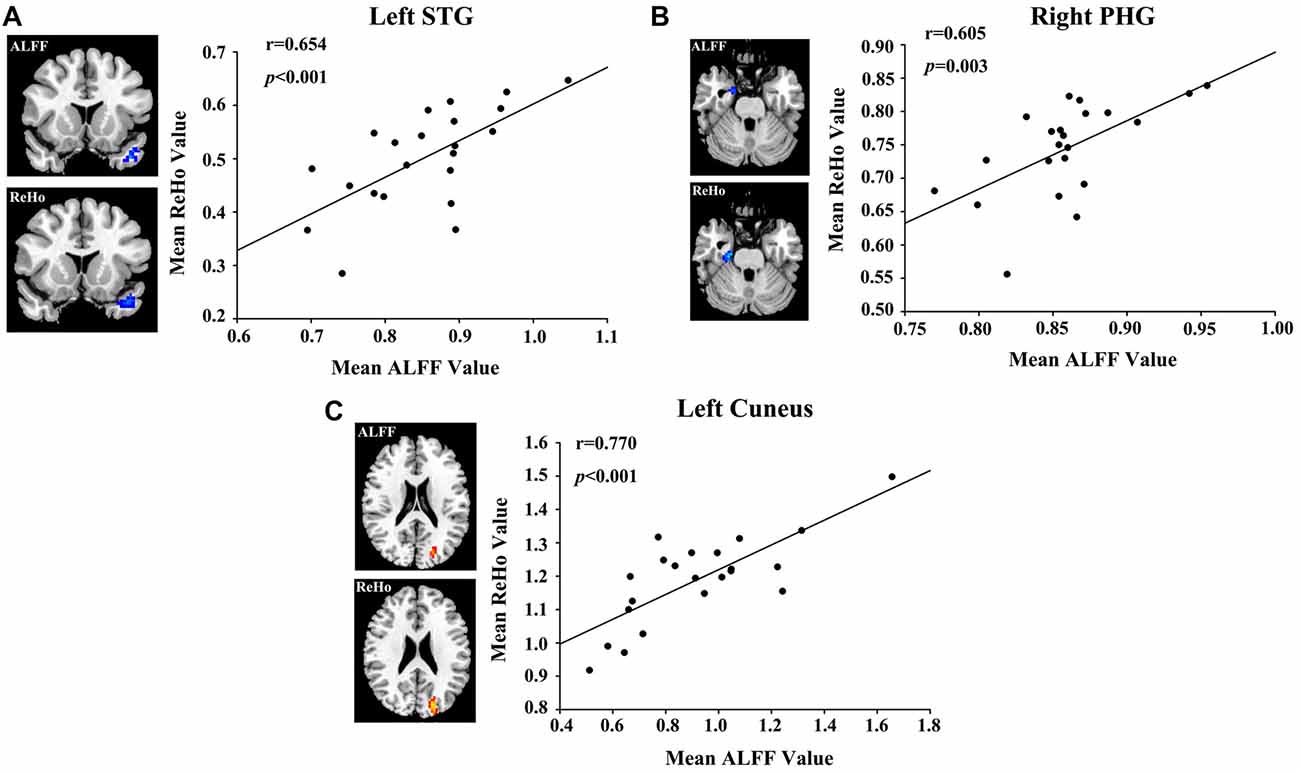
Figure 4. Correlations between the ALFF and ReHo values in (A) left STG (r = 0.654, p < 0.001), (B) right PHG (r = 0.605, p = 0.003) and (C) left cuneus (r = 0.770, p < 0.001).
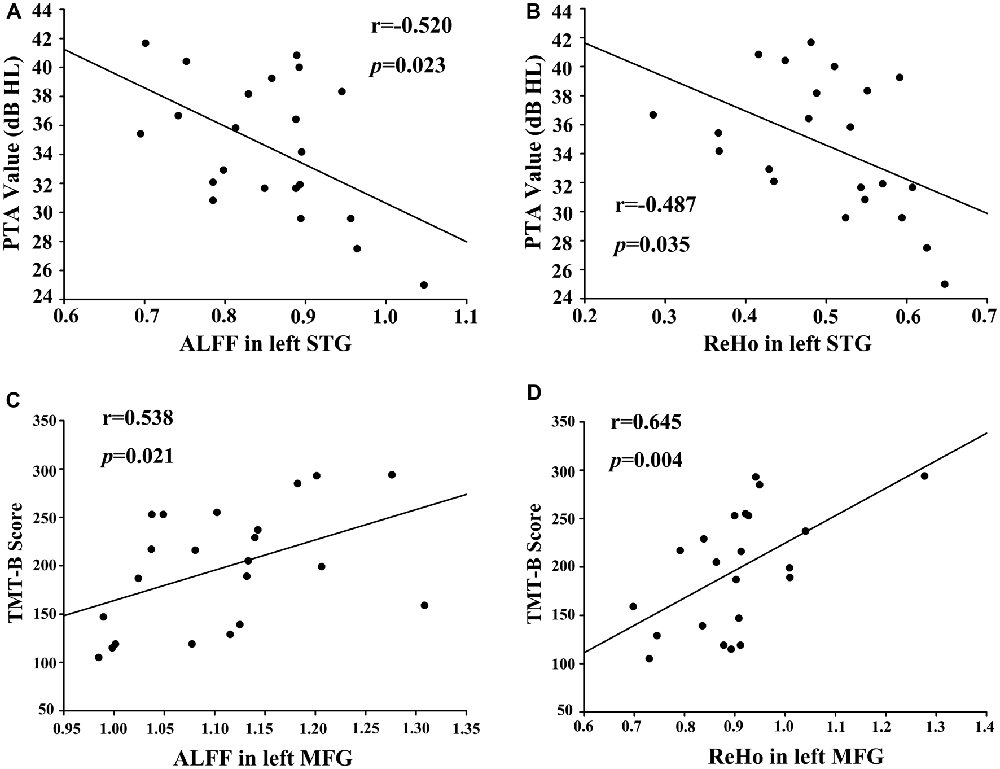
Figure 5. (A) Correlations between the mean pure tone audiometry (PTA) values and decreased ALFF values in left STG (r = −0.520, p = 0.023). (B) Correlations between mean PTA values and decreased ReHo values in left STG (r = −0.487, p = 0.035). (C) Correlations between the Trail-Making Test B (TMT-B) scores and increased ALFF in left MFG (r = 0.538, p = 0.021). (D) Correlations between TMT-B scores and increased ReHo in left MFG (r = 0.645, p = 0.004).
Discussions
To our knowledge, this is the first fMRI study to use both ALFF and ReHo to determine if spontaneous neural activity is disrupted in presbycusis patients and to ascertain if the disruptions in spontaneous activity are correlated with executive or cognitive decline. Increased or decreased neuronal changes were detected primarily in the left STG (BA 38), prefrontal cortex and portions of the DMN (e.g., precuneus and IPL). Moreover, a significant correlation was found between ALFF and ReHo values in the left STG (BA 38) and average PTA values, specifically ALFF and ReHo values decreased with increasing hearing loss in presbycusis patients. The left STG plays an important role in speech comprehension (Giraud et al., 2004) and auditory temporal processing, a psychoacoustic property that is disrupted by presbycusis and high frequency hearing loss (Salvi and Arehole, 1985; Zatorre and Belin, 2001; Gordon-Salant, 2005; Ozmeral et al., 2016). In addition, ALFF and ReHo activity in the left MFG increased significantly with decline in executive function as reflected in increasing TMT-B test scores, which implicates executive dysfunction as a putative mechanism involved in presbycusis.
ALFF and ReHo have been widely used to explore the pathogenesis of various neuropsychiatric disorders (Zang et al., 2004, 2007; Wang et al., 2011; An et al., 2013; Zhang et al., 2015). ALFF measures the intensity of spontaneous low-frequency oscillations (Zang et al., 2007) whereas ReHo reflects the local neural coherence of the underlying spontaneous neural activity (Zang et al., 2004). In the current study, ALFF and ReHo values were significantly higher than the global means values mainly in the DMN regions. Previous studies using positron emission tomography (PET) have confirmed that the metabolism of the DMN is higher than the global means values in the whole brain, which is also observed in our study utilizing the ALFF and ReHo methods, reflecting that neural activity in the DMN is most active at rest as distinguished from regions that are transiently activated in support of varying goal-directed activities (Raichle et al., 2001; Raichle and Snyder, 2007). In presbycusis patients, the increase in ALFF intensity was strongly correlated with increased ReHo coherence values in three regions, the left STG, left cuneus and right PHG. The simultaneous increase of both amplitude and coherence may reflect a more severe or complex dysfunction in these three regions. Surprisingly, the changes in the PoCG were different for ALFF and ReHo in our results, which could probably be due to the different mathematical algorithms between these two methods (Zang et al., 2004, 2007). Moreover, An et al. also suggested that there existed several abnormal brain regions that did not overlap in the same lateralization although the same brain area was detected by using ALFF and ReHo algorithms separately (An et al., 2013).
Elderly patients with presbycusis and other forms of hearing loss often suffer from tinnitus (Baracca et al., 2011). One major finding of this study was that presbycusis patients showed decreased ALFF and ReHo values in the left STG (BA 38), which were correlated with the mean PTA values. Interestingly, in profoundly deaf patients with tinnitus, activation of their cochlear implant temporarily suppresses tinnitus (residual inhibition) and activates BA 38. One interpretation of these results is that cochlear deafness suppresses spontaneous activity in BA 38, creating conditions that may allow the phantom sound of tinnitus to emerge, i.e., a certain level of spontaneous activity in BA 38 may be needed to suppress phantom sound memories (Osaki et al., 2005).
The increase in PTA thresholds are likely due to peripheral hearing loss and cochlear pathologies; however these change may contribute to central auditory disturbance that can contribute to presbycusis (Willott et al., 1993; Vasama and Mäkelä, 1997; Kraus et al., 2010; Newman et al., 2015). Impairments in peripheral hearing and central auditory processing have been linked to accelerated cognitive decline, incident cognitive impairment and AD (Gates and Mills, 2005; Panza et al., 2015). In elderly, non-demented subjects, Alzheimer neurofibrillary tangles are most often seen in the hippocampus and entorhinal cortex. However, in some cases they neurofibrillary tangles were also observed in BA 38 of the STG, suggestive of subclinical Alzheimer pathology (Vermersch et al., 1995).
fMRI has been used to detect the relationship between brain function in older adults during sentence processing and hearing ability; differences in hearing ability could predict the degree of neural recruitment in the bilateral STG during sentence comprehension (Peelle et al., 2011). Pink noise stimulation during fMRI imaging was used to examine the neural response in mild presbycusis patients and normal-hearing young controls (Profant et al., 2015). Presbycusis patients showed greater response to acoustical stimuli in the STG region compared with young subjects, indicative of sound evoked hyperactivity. Others used fMRI to explore the effect of hearing loss on engagement of the auditory cortex during the processing of monosyllabic words in presbycusis (Chen et al., 2016). Compared with age-matched healthy controls, bilateral activation in the auditory cortex was reduced in the presbycusis group. One MRS study also found reduced GABA concentrations in auditory regions of presbycusis patients and suggested that loss of GABA-medicated inhibition could contribute significantly to the deterioration of hearing function in the elderly (Gao et al., 2015). Our study extends these earlier reports by showing that presbycusis decreases the amplitude and local synchrony of spontaneous activity in the left STG (BA 38), which lies outside the primary auditory cortex. It is unclear if these decreases are mainly due to cochlear hearing loss, central auditory dysfunction or some combination of the two. Surprisingly, we did not observe a change in ALFF or ReHo in primary auditory cortex (BA 41, 42), regions in which electrophysiological studies in animal models showed increased spontaneous activity and neural synchrony with presbycusis or cochlear hearing loss degraded (Seki and Eggermont, 2003; Hughes et al., 2010; Chi-Wing and Recanzone, 2017). The reason for the difference between fMRI measures in humans and electrophysiological measure in primary auditory cortex of animals are clear, but could be due to methodological differences (e.g., anesthesia), species differences or duration of hearing loss or layer(s) of the cortex being evaluated (Novák et al., 2016).
In presbycusis patients, increased spontaneous activity and local synchrony in the MFG were associated with impaired cognitive performance on the TMT-B test, which assesses cognitive impairment related to executive function and damage to the frontal lobe (Tombaugh, 2004). The prefrontal cortex is mainly responsible for executive and cognitive functions (Roberts et al., 1998). Increased neuronal activity in the MFG may therefore have contributed to patients’ poor performance of the TMT-B test, which has been used to evaluate the executive dysfunction in central presbycusis (Gates et al., 2010). Husain et al. (2014) also observed increased connectivity between left MFG and seed region for the DMN in middle-aged patients with hearing loss. The aberrant spontaneous activity in the MFG play a pivotal role in presbycusis-related cognitive dysfunction related to executive function.
In our study, significant hypoactivity was found in subdivision of the DMN such as precuneus and IPL. The DMN, consisting of nodes in the precuneus/PCC, bilateral IPL, medial temporal gyrus and medial prefrontal gyrus, is most active at rest and shows reduced activity when a subject enters a task-based state involving attention or goal-directed behavior (Raichle et al., 2001; Mantini et al., 2007). Alterations to resting-state functional connectivity within the DMN have been detected in age-related hearing loss (Husain et al., 2014). The source or type of aberrant neural activity within specific DMN regions due to presbycusis remains unclear. Our results suggest that decreased ALFF/ReHo activity in precuneus and IPL may be responsible for disrupting the DMN in presbycusis patients.
Structural and functional alterations have been observed in the limbic system of presbycusis patients (Husain et al., 2014; Lin et al., 2014; Mudar and Husain, 2016). We observed a decrease in spontaneous activity and local connectivity in the right PHG, a primary node of the limbic system that plays a central role in memory encoding and recognition of environmental scenes and transferring information from the hippocampus to association areas (Diederen et al., 2010). Lin et al. reported atrophy of the PHG in older adults with hearing impairment compared with those with normal hearing (Lin et al., 2014). Moreover, Husain et al. (2014) used task-fMRI to demonstrate that positively and negatively valent sounds caused greater engagement of the limbic system compared to neutral sounds in normal-hearing individuals whereas older adults with hearing loss exhibited a decreased response in the PHG. Our results presumably indicate that these changes may be linked to disrupted resting-state neuronal activity in limbic system due to presbycusis.
The decline in ALFF/ReHo values in the STG, an auditory area, was associated with a complementary increase in the cuneus, a visual processing area and the PoCG, a somesthesis processing region. Decreased spontaneous activity in the auditory cortex due to hearing loss may promote spontaneous activity and/or cross-modal plasticity in visual and somatic processing areas consistent with other sound-activated imaging studies (Hadjikhani and Roland, 1998; Sadato et al., 2002; Zhang et al., 2006). A recent study utilizing rs-fMRI found that hearing loss modulated cross-modal functional connectivity between visual and auditory sensory cortices in presbycusis (Puschmann and Thiel, 2017). Auditory functional reorganization partly compensates for the needs of the visual system. Several studies observed enhanced visual and somatosensory projections into the auditory cortex, indicating that cross-modal plasticity as potential compensatory mechanism for presbycusis and/or hearing loss (Kok et al., 2014; Wong et al., 2015). Functional alterations observed in other sensory areas plus limbic and cognitive suggest that presbycusis is not limited to the classical auditory pathway, but involves sharing of multiple neurocognitive resources (Wayne and Johnsrude, 2015).
Since age-related hearing loss has been associated with structural changes, we compared GM and WM volumes but did not detect any differences between our presbycusis patients and matched controls. Previous studies have reported decreased GM volume associated with age-related hearing impairment in the central auditory and cerebral system (Peelle et al., 2011; Eckert et al., 2012; Lin et al., 2014). However, consistent with our results, Profant et al. reported no association between age-related hearing loss and GM volume (Profant et al., 2014). It is possible that the inherent heterogeneity of the hearing loss population may be one reason for the different results. Moreover, the MR technique and analytical method may also contribute to the differences.
Several limitations must be acknowledged in the current study. First, our cross-sectional study involved a relatively small sample size. Thus, it is difficult to make direct causal inferences regarding the relationships between the aberrant ALFF/ReHo and age-associated hearing loss. Further longitudinal fMRI studies are needed to establish to confirm the current findings. Second, we attempted to exclude subjects with tinnitus or hyperacusis from our study because subjects with tinnitus or hyperacusis exhibited resting-state brain abnormalities in auditory and non-auditory regions (Gu et al., 2010; Chen et al., 2017). However, it would be useful to include patients with tinnitus and hyperacusis in further studies in order to determine if spontaneous neural activity is disrupted in a similar manner to that observed in the current study. Finally, although we attempted to minimize the scanner noise with earplugs, we cannot rule out the possibility that scanner noise affected our results to some degree. This confounding factor should be taken into consideration in future studies.
Conclusions
In this study, our combined ALFF and ReHo analyses demonstrated disrupted spontaneous neural activity in specific brain regions in presbycusis patients in the absence of any major structural changes. Aberrant spontaneous neural activity mainly occurred in the auditory cortex, prefrontal cortex and some parts of the DMN. Age-related hearing loss was associated with a decline in spontaneous activity of the auditory cortex whereas impaired cognitive/executive function was associated with increased spontaneous activity in the prefrontal cortex.
Author Contributions
Y-CC designed the experiment, collected the data, performed the analysis and wrote the manuscript. HC, LJ, FB, J-JX, C-NM and XY collected the data. RS, GL and J-PG contributed to the discussion and manuscript revision.
Funding
This work was supported by a grant from the National Natural Science Foundation of China (Nos. 81601477, 81600638), Natural Science Foundation of Jiangsu Higher Education Institutions (No. 16KJB320001), Youth Medical Talents of Jiangsu Province (No. QNRC2016062), China Postdoctoral Science Foundation (No. 2017M610337), Jiangsu Postdoctoral Science Foundation (No. 1701007A), and 14th “Six Talent Peaks” Project of Jiangsu Province (No. YY-079) and Nanjing Outstanding Youth Fund (No. JQX17006).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Footnotes
References
An, L., Cao, Q.-J., Sui, M.-Q., Sun, L., Zou, Q.-H., Zang, Y.-F., et al. (2013). Local synchronization and amplitude of the fluctuation of spontaneous brain activity in attention-deficit/hyperactivity disorder: a resting-state fMRI study. Neurosci. Bull. 29, 603–613. doi: 10.1007/s12264-013-1353-8
Ardila, A., Bernal, B., and Rosselli, M. (2016). How localized are language brain areas? A review of Brodmann areas involvement in oral language. Arch. Clin. Neuropsychol. 31, 112–122. doi: 10.1093/arclin/acv081
Baracca, G., Del Bo, L., and Ambrosetti, U. (2011). “Tinnitus and hearing loss,” in Textbook of Tinnitus, eds A. R. Møller, B. Langguth, D. De Ridder and T. Kleinjung (New York, NY: Springer), 285–291.
Bowie, C. R., and Harvey, P. D. (2006). Administration and interpretation of the Trail Making Test. Nat. Protoc. 1, 2277–2281. doi: 10.1038/nprot.2006.390
Brucki, S. M., and Rocha, M. S. (2004). Category fluency test: effects of age, gender and education on total scores, clustering and switching in Brazilian Portuguese-speaking subjects. Braz. J. Med. Biol. Res. 37, 1771–1777. doi: 10.1590/s0100-879x2004001200002
Chen, X., Wang, M., Deng, Y., Liang, Y., Li, J., and Chen, S. (2016). Language processing of auditory cortex revealed by functional magnetic resonance imaging in presbycusis patients. Acta Otolaryngol. 136, 113–119. doi: 10.3109/00016489.2015.1049662
Chen, Y. C., Wang, F., Wang, J., Bo, F., Xia, W., Gu, J. P., et al. (2017). Resting-state brain abnormalities in chronic subjective tinnitus: a meta-analysis. Front. Hum. Neurosci. 11:22. doi: 10.3389/fnhum.2017.00022
Chi-Wing, N., and Recanzone, G. H. (2017). Age-related changes in temporal processing of rapidly-presented sound sequences in the macaque auditory cortex. Cereb. Cortex doi: 10.1093/cercor/bhx240 [Epub ahead of print].
Davis, A. C. (1991). Epidemiological profile of hearing impairments: the scale and nature of the problem with special reference to the elderly. Acta Otolaryngol. Suppl. 476, 23–31. doi: 10.3109/00016489109127252
Diederen, K. M., Neggers, S. F., Daalman, K., Blom, J. D., Goekoop, R., Kahn, R. S., et al. (2010). Deactivation of the parahippocampal gyrus preceding auditory hallucinations in schizophrenia. Am. J. Psychiatry 167, 427–435. doi: 10.1176/appi.ajp.2009.09040456
Dubno, J. R., Lee, F.-S., Matthews, L. J., Ahlstrom, J. B., Horwitz, A. R., and Mills, J. H. (2008). Longitudinal changes in speech recognition in older persons. J. Acoust. Soc. Am. 123, 462–475. doi: 10.1121/1.2817362
Eckert, M. A., Cute, S. L., Vaden, K. I. Jr., Kuchinsky, S. E., and Dubno, J. R. (2012). Auditory cortex signs of age-related hearing loss. J. Assoc. Res. Otolaryngol. 13, 703–713. doi: 10.1007/s10162-012-0332-5
Eklund, A., Nichols, T. E., and Knutsson, H. (2016). Cluster failure: why fMRI inferences for spatial extent have inflated false-positive rates. Proc. Natl. Acad. Sci. U S A 113, 7900–7905. doi: 10.1073/pnas.1602413113
Feder, K., Michaud, D., Ramage-Morin, P., McNamee, J., and Beauregard, Y. (2015). Prevalence of hearing loss among canadians aged 20 to 79: audiometric results from the 2012/2013 canadian health measures survey. Health Rep. 26, 18–25.
Galea, M., and Woodward, M. (2005). Mini-mental state examination (MMSE). Aust. J. Physiother. 51:198.doi: 10.1016/s0004-9514(05)70034-9
Gao, F., Wang, G., Ma, W., Ren, F., Li, M., Dong, Y., et al. (2015). Decreased auditory GABA+ concentrations in presbycusis demonstrated by edited magnetic resonance spectroscopy. Neuroimage 106, 311–316. doi: 10.1016/j.neuroimage.2014.11.023
Gates, G. A., Gibbons, L. E., McCurry, S. M., Crane, P. K., Feeney, M. P., and Larson, E. B. (2010). Executive dysfunction and presbycusis in older persons with and without memory loss and dementia. Cogn. Behav. Neurol. 23, 218–223. doi: 10.1097/WNN.0b013e3181d748d7
Gates, G. A., and Mills, J. H. (2005). Presbycusis. Lancet 366, 1111–1120. doi: 10.1016/S0140-6736(05)67423-5
Giraud, A., Kell, C., Thierfelder, C., Sterzer, P., Russ, M., Preibisch, C., et al. (2004). Contributions of sensory input, auditory search and verbal comprehension to cortical activity during speech processing. Cereb. Cortex 14, 247–255. doi: 10.1093/cercor/bhg124
Gordon-Salant, S. (2005). Hearing loss and aging: new research findings and clinical implications. J. Rehabil. Res. Dev. 42, 9–24. doi: 10.1682/jrrd.2005.01.0006
Gu, J. W., Halpin, C. F., Nam, E.-C., Levine, R. A., and Melcher, J. R. (2010). Tinnitus, diminished sound-level tolerance, and elevated auditory activity in humans with clinically normal hearing sensitivity. J. Neurophysiol. 104, 3361–3370. doi: 10.1152/jn.00226.2010
Gurgel, R. K., Ward, P. D., Schwartz, S., Norton, M. C., Foster, N. L., and Tschanz, J. T. (2014). Relationship of hearing loss and dementia: a prospective, population-based study. Otol. Neurotol. 35, 775–781. doi: 10.1097/MAO.0000000000000313
Hadjikhani, N., and Roland, P. E. (1998). Cross-modal transfer of information between the tactile and the visual representations in the human brain: a positron emission tomographic study. J. Neurosci. 18, 1072–1084.
Hale, J. B., Hoeppner, J.-A. B., and Fiorello, C. A. (2002). Analyzing digit span components for assessment of attention processes. J. Psychoeduc. Assess. 20, 128–143. doi: 10.1177/073428290202000202
Hughes, L. F., Turner, J. G., Parrish, J. L., and Caspary, D. M. (2010). Processing of broadband stimuli across A1 layers in young and aged rats. Hear. Res. 264, 79–85. doi: 10.1016/j.heares.2009.09.005
Husain, F. T., Carpenter-Thompson, J. R., and Schmidt, S. A. (2014). The effect of mild-to-moderate hearing loss on auditory and emotion processing networks. Front. Syst. Neurosci. 8:10. doi: 10.3389/fnsys.2014.00010
Kazee, A. M., Han, L. Y., Spongr, V. P., Walton, J. P., Salvi, R. J., and Flood, D. G. (1995). Synaptic loss in the central nucleus of the inferior colliculus correlates with sensorineural hearing loss in the C57BL/6 mouse model of presbycusis. Hear. Res. 89, 109–120. doi: 10.1016/0378-5955(95)00128-6
Kimura, M., and Eggermont, J. J. (1999). Effects of acute pure tone induced hearing loss on response properties in three auditory cortical fields in cat. Hear. Res. 135, 146–162. doi: 10.1016/s0378-5955(99)00104-5
Kok, M. A., Chabot, N., and Lomber, S. G. (2014). Cross-modal reorganization of cortical afferents to dorsal auditory cortex following early- and late-onset deafness. J. Comp. Neurol. 522, 654–675. doi: 10.1002/cne.23439
Kraus, K. S., Mitra, S., Jimenez, Z., Hinduja, S., Ding, D., Jiang, H., et al. (2010). Noise trauma impairs neurogenesis in the rat hippocampus. Neuroscience 167, 1216–1226. doi: 10.1016/j.neuroscience.2010.02.071
Lin, F., Ferrucci, L., An, Y., Goh, J., Doshi, J., Metter, E., et al. (2014). Association of hearing impairment with brain volume changes in older adults. Neuroimage 90, 84–92. doi: 10.1016/j.neuroimage.2013.12.059
Lin, F. R., Ferrucci, L., Metter, E. J., An, Y., Zonderman, A. B., and Resnick, S. M. (2011a). Hearing loss and cognition in the baltimore longitudinal study of aging. Neuropsychology 25, 763–770. doi: 10.1037/a0024238
Lin, F. R., Thorpe, R., Gordon-Salant, S., and Ferrucci, L. (2011b). Hearing loss prevalence and risk factors among older adults in the United States. J. Gerontol. A Biol. Sci. Med. Sci. 66, 582–590. doi: 10.1093/gerona/glr002
Ma, W., Li, M., Gao, F., Zhang, X., Shi, L., Yu, L., et al. (2016). DTI analysis of presbycusis using voxel-based analysis. Am. J. Neuroradiol. 37, 2110–2114. doi: 10.3174/ajnr.A4870
Machulda, M. M., Jones, D. T., Vemuri, P., McDade, E., Avula, R., Przybelski, S., et al. (2011). Effect of APOE ε4 status on intrinsic network connectivity in cognitively normal elderly subjects. Arch. Neurol. 68, 1131–1136. doi: 10.1001/archneurol.2011.108
Mantini, D., Perrucci, M. G., Del Gratta, C., Romani, G. L., and Corbetta, M. (2007). Electrophysiological signatures of resting state networks in the human brain. Proc. Natl. Acad. Sci. U S A 104, 13170–13175. doi: 10.1073/pnas.0700668104
Mudar, R. A., and Husain, F. T. (2016). Neural alterations in acquired age-related hearing loss. Front. Psychol. 7:828. doi: 10.3389/fpsyg.2016.00828
Nasreddine, Z. S., Phillips, N. A., Bédirian, V., Charbonneau, S., Whitehead, V., Collin, I., et al. (2005). The Montreal Cognitive Assessment, MoCA: a brief screening tool for mild cognitive impairment. J. Am. Geriatr. Soc. 53, 695–699. doi: 10.1111/j.1532-5415.2005.53221.x
Newman, A. J., Hayes, S. H., Rao, A. S., Allman, B. L., Manohar, S., Ding, D., et al. (2015). Low-cost blast wave generator for studies of hearing loss and brain injury: blast wave effects in closed spaces. J. Neurosci. Methods 242, 82–92. doi: 10.1016/j.jneumeth.2015.01.009
Novák, O., Zelenka, O., Hromádka, T., and Syka, J. (2016). Immediate manifestation of acoustic trauma in the auditory cortex is layer specific and cell type dependent. J. Neurophysiol. 115, 1860–1874. doi: 10.1152/jn.00810.2015
Osaki, Y., Nishimura, H., Takasawa, M., Imaizumi, M., Kawashima, T., Iwaki, T., et al. (2005). Neural mechanism of residual inhibition of tinnitus in cochlear implant users. Neuroreport 16, 1625–1628. doi: 10.1097/01.WNR.0000183899.85277.08
Ouda, L., Profant, O., and Syka, J. (2015). Age-related changes in the central auditory system. Cell Tissue Res. 361, 337–358. doi: 10.1007/s00441-014-2107-2
Ozmeral, E. J., Eddins, A. C., Frisina, D. R., and Eddins, D. A. (2016). Large cross-sectional study of presbycusis reveals rapid progressive decline in auditory temporal acuity. Neurobiol. Aging 43, 72–78. doi: 10.1016/j.neurobiolaging.2015.12.024
Palombi, P. S., and Caspary, D. M. (1996). Physiology of the aged Fischer 344 rat inferior colliculus: responses to contralateral monaural stimuli. J. Neurophysiol. 76, 3114–3125. doi: 10.1152/jn.1996.76.5.3114
Panza, F., Solfrizzi, V., and Logroscino, G. (2015). Age-related hearing impairment—a risk factor and frailty marker for dementia and AD. Nat. Rev. Neurol. 11, 166–175. doi: 10.1038/nrneurol.2015.12
Peelle, J. E., Troiani, V., Grossman, M., and Wingfield, A. (2011). Hearing loss in older adults affects neural systems supporting speech comprehension. J. Neurosci. 31, 12638–12643. doi: 10.1523/JNEUROSCI.2559-11.2011
Power, J. D., Barnes, K. A., Snyder, A. Z., Schlaggar, B. L., and Petersen, S. E. (2012). Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. Neuroimage 59, 2142–2154. doi: 10.1016/j.neuroimage.2011.10.018
Profant, O., Škoch, A., Balogová, Z., Tintěra, J., Hlinka, J., and Syka, J. (2014). Diffusion tensor imaging and MR morphometry of the central auditory pathway and auditory cortex in aging. Neuroscience 260, 87–97. doi: 10.1016/j.neuroscience.2013.12.010
Profant, O., Tintěra, J., Balogová, Z., Ibrahim, I., Jilek, M., and Syka, J. (2015). Functional changes in the human auditory cortex in ageing. PLoS One 10:e0116692. doi: 10.1371/journal.pone.0116692
Puschmann, S., and Thiel, C. M. (2017). Changed crossmodal functional connectivity in older adults with hearing loss. Cortex 86, 109–122. doi: 10.1016/j.cortex.2016.10.014
Raichle, M. E., MacLeod, A. M., Snyder, A. Z., Powers, W. J., Gusnard, D. A., and Shulman, G. L. (2001). A default mode of brain function. Proc. Natl. Acad. Sci. U S A 98, 676–682. doi: 10.1073/pnas.98.2.676
Raichle, M. E., and Snyder, A. Z. (2007). A default mode of brain function: a brief history of an evolving idea. Neuroimage 37, 1083–1090. doi: 10.1016/j.neuroimage.2007.02.041
Roberts, A. C., Robbins, T. W., and Weiskrantz, L. E. (1998). The Prefrontal Cortex: Executive and Cognitive Functions. New York, NY: Oxford University Press.
Ronnberg, J., Danielsson, H., Rudner, M., Arlinger, S., Sternang, O., Wahlin, A., et al. (2011). Hearing loss is negatively related to episodic and semantic long-term memory but not to short-term memory. J. Speech Lang. Hear. Res. 54, 705–726. doi: 10.1044/1092-4388(2010/09-0088)
Rosenhall, U., Hederstierna, C., and Idrizbegovic, E. (2011). Otological diagnoses and probable age-related auditory neuropathy in “younger” and “older” elderly persons. Int. J. Audiol. 50, 578–581. doi: 10.3109/14992027.2011.580786
Sadato, N., Okada, T., Honda, M., and Yonekura, Y. (2002). Critical period for cross-modal plasticity in blind humans: a functional MRI study. Neuroimage 16, 389–400. doi: 10.1006/nimg.2002.1111
Salvi, R. J., and Arehole, S. (1985). Gap detection in chinchillas with temporary high-frequency hearing loss. J. Acoust. Soc. Am. 77, 1173–1177. doi: 10.1121/1.392181
Salvi, R., Lockwood, A., Frisina, R., Coad, M., Wack, D., and Frisina, D. (2002). PET imaging of the normal human auditory system: responses to speech in quiet and in background noise. Hear. Res. 170, 96–106. doi: 10.1016/s0378-5955(02)00386-6
Samton, J. B., Ferrando, S. J., Sanelli, P., Karimi, S., Raiteri, V., and Barnhill, J. W. (2005). The clock drawing test: diagnostic, functional, and neuroimaging correlates in older medically ill adults. J. Neuropsychiatry Clin. Neurosci. 17, 533–540. doi: 10.1176/appi.neuropsych.17.4.533
Schmidt, M. (1996). Rey Auditory Verbal Learning Test: A Handbook. Los Angeles, CA: Western Psychological Services.
Seki, S., and Eggermont, J. J. (2003). Changes in spontaneous firing rate and neural synchrony in cat primary auditory cortex after localized tone-induced hearing loss. Hear. Res. 180, 28–38. doi: 10.1016/s0378-5955(03)00074-1
Shin, M.-S., Park, S.-Y., Park, S.-R., Seol, S.-H., and Kwon, J. S. (2006). Clinical and empirical applications of the Rey-Osterrieth complex figure test. Nat. Protoc. 1, 892–899. doi: 10.1038/nprot.2006.115
Spongr, V. P., Flood, D. G., Frisina, R. D., and Salvi, R. J. (1997). Quantitative measures of hair cell loss in CBA and C57BL/6 mice throughout their life spans. J. Acoust. Soc. Am. 101, 3546–3553. doi: 10.1121/1.418315
Sprinzl, G., and Riechelmann, H. (2010). Current trends in treating hearing loss in elderly people: a review of the technology and treatment options-a mini-review. Gerontology 56, 351–358. doi: 10.1159/000275062
Tay, T., Wang, J. J., Kifley, A., Lindley, R., Newall, P., and Mitchell, P. (2006). Sensory and cognitive association in older persons: findings from an older Australian population. Gerontology 52, 386–394. doi: 10.1159/000095129
Tombaugh, T. N. (2004). Trail Making Test A and B: normative data stratified by age and education. Arch. Clin. Neuropsychol. 19, 203–214. doi: 10.1016/s0887-6177(03)00039-8
Vasama, J.-P., and Mäkelä, J. P. (1997). Auditory cortical responses in humans with profound unilateral sensorineural hearing loss from early childhood. Hear. Res. 104, 183–190. doi: 10.1016/s0378-5955(96)00200-6
Vermersch, P., David, J.-P., Frigard, B., Fallet-Bianco, C., Wattez, A., Petit, H., et al. (1995). Cortical mapping of alzheimer pathology in brains of aged non-demented subjects. Prog. Neuropsychopharmacol. Biol. Psychiatry 19, 1035–1047. doi: 10.1016/0278-5846(95)00195-6
Wang, Z., Yan, C., Zhao, C., Qi, Z., Zhou, W., Lu, J., et al. (2011). Spatial patterns of intrinsic brain activity in mild cognitive impairment and alzheimer’s disease: a resting-state functional MRI study. Hum. Brain Mapp. 32, 1720–1740. doi: 10.1002/hbm.21140
Wayne, R. V., and Johnsrude, I. S. (2015). A review of causal mechanisms underlying the link between age-related hearing loss and cognitive decline. Ageing Res. Rev. 23, 154–166. doi: 10.1016/j.arr.2015.06.002
Willott, J. F. (1991). Central physiological correlates of ageing and presbycusis in mice. Acta Otolaryngol. Suppl. 111, 153–156. doi: 10.3109/00016489109127271
Willott, J. F., Aitkin, L. M., and McFadden, S. L. (1993). Plasticity of auditory cortex associated with sensorineural hearing loss in adult C57BL/6J mice. J. Comp. Neurol. 329, 402–411. doi: 10.1002/cne.903290310
Wong, C., Chabot, N., Kok, M. A., and Lomber, S. G. (2015). Amplified somatosensory and visual cortical projections to a core auditory area, the anterior auditory field, following early- and late-onset deafness. J. Comp. Neurol. 523, 1925–1947. doi: 10.1002/cne.23771
Yan, C.-G., Wang, X.-D., Zuo, X.-N., and Zang, Y.-F. (2016). DPABI: data processing and analysis for (resting-state) brain imaging. Neuroinformatics 14, 339–351. doi: 10.1007/s12021-016-9299-4
Zang, Y. F., He, Y., Zhu, C. Z., Cao, Q. J., Sui, M. Q., Liang, M., et al. (2007). Altered baseline brain activity in children with ADHD revealed by resting-state functional MRI. Brain Dev. 29, 83–91. doi: 10.1016/j.braindev.2006.07.002
Zang, Y., Jiang, T., Lu, Y., He, Y., and Tian, L. (2004). Regional homogeneity approach to fMRI data analysis. Neuroimage 22, 394–400. doi: 10.1016/j.neuroimage.2003.12.030
Zatorre, R. J., and Belin, P. (2001). Spectral and temporal processing in human auditory cortex. Cereb. Cortex 11, 946–953. doi: 10.1093/cercor/11.10.946
Zhang, J., Chen, Y.-C., Feng, X., Yang, M., Liu, B., Qian, C., et al. (2015). Impairments of thalamic resting-state functional connectivity in patients with chronic tinnitus. Eur. J. Radiol. 84, 1277–1284. doi: 10.1016/j.ejrad.2015.04.006
Zhang, Y.-T., Geng, Z.-J., Zhang, Q., Li, W., and Zhang, J. (2006). Auditory cortical responses evoked by pure tones in healthy and sensorineural hearing loss subjects: functional MRI and magnetoencephalography. Chin. Med. J. 119, 1548–1554.
Zung, W. W. (1971). A rating instrument for anxiety disorders. Psychosomatics 12, 371–379. doi: 10.1016/s0033-3182(71)71479-0
Keywords: presbycusis, ALFF, ReHo, resting-state fMRI, spontaneous activity
Citation: Chen Y-C, Chen H, Jiang L, Bo F, Xu J-J, Mao C-N, Salvi R, Yin X, Lu G and Gu J-P (2018) Presbycusis Disrupts Spontaneous Activity Revealed by Resting-State Functional MRI. Front. Behav. Neurosci. 12:44. doi: 10.3389/fnbeh.2018.00044
Received: 20 December 2017; Accepted: 26 February 2018;
Published: 13 March 2018.
Edited by:
Liana Fattore, Consiglio Nazionale delle Ricerche (CNR), ItalyReviewed by:
Josef Syka, Institue of Experimental Medicine, Academy of Sciences of the Czech Republic, CzechiaXiaoming Jin, Indiana University, Purdue University Indianapolis, United States
Copyright © 2018 Chen, Chen, Jiang, Bo, Xu, Mao, Salvi, Yin, Lu and Gu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Guangming Lu, Y2pyLmx1Z3VhbmdtaW5nQHZpcC4xNjMuY29t
Jian-Ping Gu, Y2pyLmd1amlhbnBpbmdAdmlwLjE2My5jb20=