 Raffaele d'Isa1*
Raffaele d'Isa1* Robert Gerlai2*
Robert Gerlai2*- 1Institute of Experimental Neurology (INSPE), Division of Neuroscience (DNS), IRCCS San Raffaele Scientific Institute, Milan, Italy
- 2Department of Psychology, University of Toronto Mississauga, Mississauga, ON, Canada
Behavioral tests have three key elements: (1) a motivating factor (what motivates the animal in the test); (2) an observable behavior (which behaviors we may expect the animal exhibits in response to the test); (3) a measurable outcome (a quantifiable variable associated with the behavioral response).
For example, in the rodent step-through passive avoidance test (see d'Isa et al., 2014 for a brief history of the test), the animal is released into a strongly illuminated chamber connected to a dark zone. Being naturally photophobic and preferring dark areas, mice and rats will rapidly move from the well-illuminated zone into the dark zone, a behavior that in the wild is useful to avoid being seen by predators. When the animal enters the dark zone, it receives an electric shock. After the initial exposure to this apparatus (the training), the animal is released a second time in the same apparatus for a memory test, but this time without the shock deliverer being active. If the animal remembers receiving the shock, the dark zone should now be perceived as dangerous and hence avoided. In this test, the motivation is fear, the observable behavior is avoidance of a dangerous (dark) zone, and the quantifiable outcome is the latency to enter the dangerous zone, which thus serves as an index of memory. The longer is the latency, the stronger is the memory of the past electric shock exposure.
Behavioral testing of rodents in a laboratory setting started in the 1890's with the studies of Thomas Wesley Mills (1847–1915) from McGill University (Montreal, Canada) and of Linus Ward Kline (1866–1961) and Willard Stanton Small (1870–1943), both from Clark University (Worcester, Massachusetts, USA) (Mills, 1895, 1898; Kline, 1899a,b; Small, 1899, 1900, 1901). These studies were preceded by ethological, purely observational, studies on rodent behavior in the wild, e.g., Mills's studies on squirrel behavior (Mills, 1888, 1890, 1893), but it was only in the period 1895–1900 that behavioral tests for studying rodent behavior and psychology in a laboratory setting started to be designed. Mills also observed the behavior of two squirrels he captured and kept for few months, reporting, for example, how ethologically relevant behaviors could be observed, like nest-building and food storing, and how one of them learned to eat from his hand and enjoyed running on a running wheel that was installed in its home-cage (Mills, 1888). Although this was a first step for the study of rodent behavior in a controlled environment, the report with the findings was only anecdotal and appeared inserted within a paper on the behavior of squirrels in the wild. Two important elements of scientific testing were absent: systematic observations (i.e., observations at pre-set time-points according to a specific rationale) and choice of one or more quantitatively measurable behavioral outcomes as variables of interest. A systematic and quantitative study of rodent behavior in laboratory had yet to come.
Mills was the first, in the mid 1890's, to introduce for rodents the ontogenetic diary method, which consisted of observing and describing step by step the different developmental stages of a species, starting from the day of birth (Mills, 1895). Applying this method, he studied systematically the physical and psychological development of guinea pigs through a daily monitoring in a laboratory setting. In addition to purely observational studies of development, he also used some basic behavioral tests, e.g., reflex tests or taste reactivity tests, in which only qualitative responses were recorded (Mills, 1895, 1898). On the other hand, Kline and Small introduced behavioral tests aimed at specifically evaluating cognition. In 1898, a Clark University colleague of Kline and Small, Colin Campbell Stewart (1873–1944), published a study on the effects of alcohol, barometric pressure and type of diet on rat daily voluntary wheel running activity, as assessed by a revolving drum connected to automatic counters recording the total number of revolutions (Stewart, 1898). Stewart was the first to perform a quantitative rodent test of motor activity. The experiments of Stewart, from the Biology Faculty, inspired Kline, from the Psychology Faculty, the idea to choose rats as animal models for his research on learning processes (Kline, 1928; Miles, 1930). Kline designed a rat problem box (Kline, 1899a), while Small, who worked in the same laboratory as Kline, was the first to use a maze in the history of behavioral neuroscience (Small, 1901). In Kline's test, the task was finding the entrance of a box and retrieving the food contained inside it. Time to retrieve the food was recorded over multiple trials to assess learning. In Small's test, the task was finding food placed in the central zone of a complex maze inspired by the design of Hampton Court Maze, the well-known hedge labyrinth in England. Time to find the food and number of errors (entering a blind alley) were recorded over multiple trials. The ones performed by Kline and Small were the first quantitative rodent cognitive tests in the history of behavioral neuroscience. Regarding the motivating factor, both Kline and Small employed hungry rats in their cognitive tests, and food deprivation became a widely employed protocol in the subsequent studies using appetitive (reward-based) learning tasks.
However, since the dawn of behavioral neuroscience, researchers have been aware of the fact that hunger is only one possible motivator. Small himself wrote: “I trust the reader will not “jump” to the conclusion that no other motive would be workable. Hunger is merely the most fundamental” (Small, 1900). Noticing that well-fed rats still retrieved food during the task, he added: “Their performance of the task without the incitement of hunger can hardly be accounted for except upon the basis of a hoarding instinct almost as imperative as hunger” (Small, 1900). Nevertheless, these specifications seem to have been scarcely considered by the following researchers. Indeed, most behavioral tests designed up to the 1950's were based on rewards or punishments and commonly employed food deprivation or electric shocks, respectively.
Another classical avoidance task is, for example, the shuttle-box active avoidance, first conceived by Lucien Warner in the early 1930's (Warner, 1932), where instead of having to stay in the illuminated zone to avoid the shock, i.e., instead of not moving and being passive, the appropriate response is to actively move over to an opposite zone to avoid the shock when a stimulus (a tone or a light) announces its imminent release. In this task, the experimenter may have to employ a large number of shocks (even hundreds) over several days of training before animals reach high rates of shock avoidance and cognitively impaired animals could still show low rates of shock avoidance even at the end of the training (Montag-Sallaz and Montag, 2003; Cain, 2019). Painful stimulation, along with being ethically undesirable when dealing with any sentient organism, also leads to methodological complications for the experimenters. Indeed, pain generates stress, which is a major confounding factor in animal research. Still today, classical avoidance tests employ electric shocks, while many other traditional tests, although not utilizing painful stimuli, feature highly stressful conditions including starvation, water deprivation or pharmacologically induced sickness to motivate the animals to perform a task. However, an increasingly accepted view is that absence of pain and reduction of stress during behavioral testing are fundamental for both animal welfare and reproducibility of experimental results, and unless pain or stress is the main focus of the study, these conditions should be avoided as much as possible. In order to respond to such ethical and methodological concerns, several animal-friendly tests have been designed. However, since not all behavioral domains of investigation currently have such animal-friendly options, the development of new animal-friendly tests is an important goal for modern behavioral neuroscience.
How to design an animal-friendly behavioral test
Compared to shuttle-box active avoidance, step-through passive avoidance, introduced in the 1960's (Kopp et al., 1966; Jarvik and Kopp, 1967), is considerably less stressful and in the test session no shock delivery is actually present. Nevertheless, the training session still features a brief painful stimulus. In an ideal animal-friendly behavioral test, the motivating factor should not be painful or stressful. Furthermore, the observable behavior should be natural (i.e., an ethologically relevant species-specific motor or postural pattern). Finally, the outcome variable associated with this behavioral response should be practical to measure in a laboratory setting through a method that is safe for the animals (e.g., direct observation, videorecording, videotracking, audiorecording of ultrasonic vocalizations, photocell actimetry, weight sensors, infrared thermometry and other non-invasive methods). These are the three main characteristics that a behavioral test should have to be qualified as animal-friendly. Let us first focus on the motivational aspect of behavioral tests, the first of the key components mentioned above.
Ethologists have been stressing the argument that motivating factors are species-specific (Gerlai and Clayton, 1999a; Gerlai, 2021). A stimulus that is appetitive (rewarding) or aversive (punishing) for one species, may be neutral, or may have the opposite reinforcing value to another. Even among closely related species, the rewarding value of a stimulus may be remarkably different. Among felids, for instance, tigers like to bath in water, while lions do not. Hence a swimming pool may be a reward for the former, but not for the latter species, as found, for example, by Allison Hedgecoth who provided a water pool to a lioness and a tigress living together in the same environment in the Noah's Ark Animal Sanctuary of Locust Grove, Georgia, United States (Harries et al., 2020). The main issue, however, in the behavioral neuroscience literature is that systematic analysis of what motivates animals used in laboratory settings is often lacking, or that ethology research often does not intersect with biomedical studies.
Considering, for example, laboratory rodents, the house mouse (Mus musculus) and the common rat (Rattus norvegicus) are the two most widely used species in biomedical research. Their employment is almost universal in translational research studying mechanisms of central nervous system disorders. Most studies that require aversive stimuli with rodents use electric shocks. But electric shocks are rather unnatural stimuli. However, almost no one considers what consequences may result from the unnatural aspect of this stimulus. It is just assumed that pain is pain, and that electric shock-induced pain is relevant and strongly motivating. Most scientists do not even consider what complication this electricity passively running through the body of the animal, including its brain, may cause with respect to neuronal activity: such electric currents may alter synaptic function and numerous underlying molecular mechanisms. Similarly, studies that employ appetitive stimuli, almost always use food that the experimenter picks out based on tradition, habit or just personal preference. Comparative analyses of what food types, food quantities, food textures and food sizes are most preferred by rats or mice are often not considered, or have not even been conducted. Briefly, as animals are, through evolutionary processes, adapted to their natural environment, they possess species-specific characteristics that represent genetic predispositions, instincts in colloquial terms, that determine, or at least heavily influence what they like, what they dislike, and how they respond to these stimuli. Taking these species-specific features into account is thus a must in animal-friendly experimental designing (Gerlai and Clayton, 1999a,b). In rodents, a typical animal-friendly motivating factor is neophilia (attraction for novelty), which drives, for instance, object exploration behavior in the object recognition test (d'Isa et al., 2014), head-dipping in the hole-board test (d'Isa et al., 2021a) and arm alternation in the spontaneous alternation T-maze (d'Isa et al., 2021b). A similar example is the continuous spontaneous alternation test using a T-maze, which utilizes novel place preference to study short-term spatial memory in rodents (Gerlai, 1998).
Regarding the second key element mentioned above, the observable behavior in an animal-friendly test should be naturally displayed by the animal (e.g., should be part of the ethogram). Preferably, it should be a spontaneous behavior, an instinctive response that requires no pre-training, during which typically punishments and rewards are used by the experimenter to lead to a target behavior. Punishments are commonly painful stimuli (as electric shocks), while rewards, as food or liquids, are often associated with food-deprivation or water-deprivation, in order to use hunger or thirst as motivating factors. Lack of the need for pre-training makes the test more animal-friendly because it avoids punishments and deprivations, and is also time-saving for the experimenter. It is, however, also possible to use conditioned behaviors in an animal-friendly way, if certain conditions are respected. In particular, rewards should not be associated with a previous aversive state. Chow and colleagues, for instance, designed a reward-based cognitive test for gray squirrels in which no food-deprivation or water-deprivation was employed (Chow et al., 2017). The motivation of the rodents was ensured simply by using food rewards (hazelnuts) that were different from their daily diet (seeds, fresh fruit and vegetables), i.e., novelty alone was sufficient to motivate the animals. Novelty-seeking and exploratory drive (i.e., the motivation to learn about new places and/or new inanimate or animate components of the environment) are almost universal among animal species, and certainly have been shown for laboratory rodents (Gerlai et al., 1990; Crusio, 2001). In fact, stabilizing natural selection has been inferred for exploratory behaviors from fish to mammals, as it leads to an optimal level of activity ensuring the ability of the animal to find resources, including food, water and mates, as well as escape routes leading away from predators (Gerlai et al., 1990; Crusio, 2001). The use of novelty as a motivator may not be appropriate in some research contexts and, for certain studies, aversive stimulation may be required. However, even in such cases, painful punishments could be and should be substituted with non-painful aversive alternatives, for example, air-puffs. Indeed, air-puffs have been efficiently employed to elicit robust conditioned place avoidance negating the need for using any painful stimuli (d'Isa et al., 2011). Even for studies specifically focused on fear reactions, alternatives to painful stimulation are available. Odor of predators (e.g., fox's urine, or an extract from it) has been efficiently used to induce avoidance reactions and fear without previous painful stimulation (Blanchard et al., 2003).
The main steps for designing an animal-friendly test can be summarized as follows: (a) prepare a list of behaviors typical of the species (the ethogram), along with what stimuli may induce these behaviors, i.e., the motivating forces; (b) exclude behaviors induced by pain, physical suffering or psychological stress; (c) from the remaining, choose a behavior that can be studied through an apparatus that can be used in a laboratory setting; (d) choose which outcomes could be measured, safely for the animals, in the most efficient and precise way in order to provide quantitative experimental data. Let us examine an experimental example of how these steps may be accomplished.
A typical behavior of rodents is food hoarding, that is, collecting and hiding food as supply storage for times of food scarcity. This behavior can be observed in more than 180 rodents (Zhang et al., 2022). This is an adaptive behavior that is observable both in nature and in the laboratory setting. It is an instinctive behavior that does not require pre-training. Two main strategies are adopted by food hoarding rodents. Scatter hoarders, as gray squirrels, hide food in many dispersed small hoards. On the other hand, larder hoarders, as hamsters, store food in one large hoard, named the larder. A classification of the hoarding strategies of 183 rodents is provided by Zhang and colleagues (Zhang et al., 2022). These hoarding behaviors may be utilized by the experimenter to devise behavioral tests of motivation (during the food accumulation phase) or of spatial memory (during the subsequent phase of food retrieval from the spatially separated hoards). For motivation tests, easier to study in larder hoarding rodents, the measurable outcome could be the total weight of the seeds or pellets collected and stored in a fixed amount of time. For spatial memory tests, which would be best studied with scatter hoarding rodents, the recorded outcome could be the number of errors in finding the hoarding sites containing the previously stored food. Alternatively, spatial memory could be studied also in larder hoarders if, during the accumulation phase, the sources of food are multiple. Number of errors (returning to an already depleted food site) would serve as memory index. An apparatus for the testing of food hoarding behavior in a laboratory setting has been realized, for example, by Robert Deacon at Oxford University (Deacon, 2006).
An ethological approach may be useful to devise animal-friendly behavioral tests for two reasons. On the one hand, it may help researchers to choose among the elements of the ethogram a behavior that does not require painful or stressful motivating factors. On the other hand, among a taxonomical family of species (for example rodents), it may help researchers to select the most suitable species for a certain test. Let us return to the example we mentioned above. Laboratory mice are larder hoarders, just like hamsters, but their propensity to hoard is relatively low under baseline conditions. In order to avoid food-deprivation, long testing sessions may be required to obtain replicable results, including, e.g., overnight testing sessions (Deacon, 2006). Hamsters, on the other hand, have a high propensity to hoard (Vander Wall, 1990; Harris, 2017). Up to 90 kg of food have been found in hamster burrows (Nowak and Paradiso, 1983). Among food hoarders, they display a specific behavior known as cheek pouching, that is accumulating food in cheek pouches, specialized pockets that allow food transportation. Instead of eating the food items, hamsters keep the food items in their mouth to carry them to a safe place for storage (the larder). Importantly, hamsters easily show this behavior even when they are not hungry, with a latency to hoard within 2 min (Montoya and Gutiérrez, 2016). This peculiarity of hamsters makes them particularly suitable as animal models for scientists who want to design an animal-friendly reward-based memory test that does not require any previous starvation.
Another rodent, the chinchilla (Chinchilla lanigera), displays a peculiar behavior known as sand-bathing: when presented with a box full of sand, it will readily start rolling in the box, rotating along its longitudinal axis, to rub its fur in the sand (Stern and Merari, 1969). This natural and spontaneous behavior can be easily elicited in a laboratory setting and sand could be used as an animal-friendly reward in instrumental learning tests without the need of any previous deprivation condition (Redman, 1974).
Eastern woodrats (Neotoma floridana), also known as pack rats, have a special attraction for shiny objects, which they readily approach, pick up and bring to their nest, where they collect them (Bradley et al., 2022). This natural tendency of woodrats could be used in behavioral tests, employing small metal objects, as stripes or balls of aluminum foil, as motivators (Kaufman and Kaufman, 1984).
Of course, the issue is that quite often neurobiological, genetic, or other methods may not be as readily available, or as sophisticated, for such species as hamsters and chinchillas as for the favorites of biomedical research, mice and rats. How can we solve this conundrum? Firstly, we could improve biotechnological methods for the so called “alternative” species. Secondly, we could improve our understanding of the ethology, the natural species-specific behavioral characteristics, of the preferred model organisms, e.g., of mice and rats. Certainly, advances in both of these areas have been made during the past few decades. Regarding the first area, numerous novel techniques may now be equally useable with mice and hamsters (and many other species). The CRISPR/Cas technology is a clear example (Kampmann, 2020). Concerning the second area, there have been research efforts adopting ethological approaches in mouse neurobehavioral genetics, as for instance testing mouse mutants in the wild (Dell'Omo et al., 2000; Vyssotski et al., 2002) or in laboratory environments more closely resembling a natural habitat, like Eco-HAB (Puścian et al., 2016; Winiarski et al., 2022). Anders Ågmo's research group at University of Tromsø recreated a seminatural environment in the laboratory for the evaluation of rat behavior (Chu and Ågmo, 2014), a method that has been employed in several subsequent studies (Chu et al., 2015; Houwing et al., 2019; Le Moëne et al., 2020; Heinla et al., 2021). In the testing sessions, which may last days, rat behavior is continuously video-recorded and subsequently scored off-line by the researchers. Notably, another important ethological approach of the new century is testing the animals not in a setting designated uniquely for testing sessions, but rather in the permanent housing environment in which they commonly live (Mingrone et al., 2020; Voikar and Gaburro, 2020). In nature, most rodents build burrows (or occupy pre-existing holes or burrows) to use them as homes (long-term inhabiting spaces), in which they return to sleep, store food, seek shelter from the elements, keep warm, hide from predators, give birth, raise the pups and share a social life with conspecifics. Rodents develop a strong bond with their home and show a territorial behavior toward it, actively defending it from possible invaders. In the laboratory, if an unfamiliar conspecific is placed in the home-cage of mice or rats, the intruder will rapidly be attacked by the resident animal (Koolhaas et al., 2013; Ruzza et al., 2015). The home-cage is the place where laboratory rodents feel safest and where they are more likely to display spontaneous natural behaviors. Thus, the idea of testing in the home-cage has been gaining considerable attention in the past few years, and several home-cage automated multi-variable recording systems have been developed, e.g., the IntelliCage (Galsworthy et al., 2005; Kiryk et al., 2020; Iman et al., 2021), PhenoMaster (Urbach et al., 2008; König et al., 2020), Actual-HCA (Bains et al., 2016; Mitchell et al., 2020) and SmartKage (Ho et al., 2022). Automated home-cage testing systems have several advantages: (a) they allow behavioral phenotyping without human interference and without the consequent handling-related stress; (b) the animals are not tested in an external apparatus but in their familiar and well-known housing environment, which eliminates confounds arising from anxiety; (c) data collection is not restricted to a specific moment of the day, but can be performed continuously, 24 h a day, 7 days a week, allowing a more precise and realistic assessment of behavior; d) long longitudinal studies (lasting weeks, months or years), or even life-long studies, can be performed on the same animals with a continuous behavioral assessment, which is particularly relevant for developmental neuroscience and aging neuroscience; (e) interactive elements (e.g., levers, nose-poking ports, motorized doors and running wheels) may be installed in these home-cages, allowing not only detailed motor assessment, but also complex cognitive testing; (f) animals are tested in a natural social context while living together with other conspecifics, thus providing motor and cognitive measurements with a higher ethological validity and allowing additionally to monitor and analyze complex social interactions. Some of these automated home-cage testing systems are modular, allowing the connection of multiple cages to create a more complex envinronment. For instance, IntelliCage can be connected to two social boxes containing different social stimuli (Mitjans et al., 2017), while in ColonyRack mice can freely roam across 70 cages, arranged in a two-sided rack with fivs columns and seven rows, in which the cages are connected both horizontally and vertically (Zocher et al., 2020; Kempermann et al., 2022). The most recent innovation within this automated behavioral testing approach is connecting home-cages to mazes (Mei et al., 2020; Kohler et al., 2022), granting the experimental subjects free access to the novel test environment. This allows the animals to decide voluntarily when and for how long they explore the maze, similarly to what would happen in nature when rodents decide to leave their burrow for external exploratory excursions.
We believe that bringing closer the fields of ethology and neurobehavioral genetics or behavioral neuroscience will be the solution and will lead to cross-fertilization of these fields. Similarly to how the application of neuroscience-related knowledge to ethology led to the birth of neuroethology (i.e., the study of the neural basis of natural behaviors), the reverse could lead to an ethologically based neuroscience, or ethological neuroscience, which can be defined as the employment of knowledge of the natural behavior of animals in the wild to develop animal models of behavior and behavioral tests for neuroscience research. This ethologically based neuroscience can lead to animal-friendly testing approaches that will not only be more oriented toward the welfare of the animals involved, but also will provide more reliable and more replicable results for the experimenters.
Concluding remarks: Reproducibility, replicability and refinement
Reproducibility is when we obtain the same results repeatedly by using identical methods (Kafkafi et al., 2018; Gerlai, 2019), whereas replicability is when we reach similar conclusions by adopting different methodologies (Kafkafi et al., 2018; Gerlai, 2019). Minimizing stress of the tested animals is a value in itself from an ethical point of view. However, since stress is a confounding factor that increases variability of experimental outcomes, minimizing stress is also fundamental to achieve methodologically sound scientific research. Why does research that ignores species-specific features lead to increased variability? Why is stress a confounding factor that reduces reproducibility? These are intriguing questions that would deserve specific research. The answer may lay in the fact that stress causes activation of the hypothalamic–pituitary–adrenal (HPA) axis, which in turn alters physiological processes regulating cognition and behavior (Moreira et al., 2016). HPA reactivity depends on genetic, epigenetic and environmental factors (Holmes et al., 2005), which makes it more difficult to predict than instinctive responses. Let us make some overarching theoretical points. Most animal research includes human handling. Human handling is extremely difficult to standardize (Crabbe et al., 1999). Even if handling was perfectly standardized, stress reactivity of the animals would not. Animals experiencing more stress due to the experimental procedures will be more responsive to human handling, which then will lead to elevated error variation in the behavioral test. In order to maximize experiment reproducibility, the best option is to minimize handling-related stress (Gouveia and Hurst, 2017). Considering, for instance, mice, although tail picking is the most commonly employed method of handling (Ueno et al., 2020), this method features tail lifting, tail suspension and swinging the animal over a void, which are highly stressful for the mice. Indeed, tail lifting, compared with alternative handling methods that do not require tail lifting, increases anxiety in the open-field test (Gouveia and Hurst, 2019) and elevated plus maze (Hurst and West, 2010), and it has been shown to reduce exploratory activity (Gouveia and Hurst, 2017), to increase aversion for the human handler in voluntary interaction test (Hurst and West, 2010) and to impair responsiveness to sucrose reward, indicating a reduction of reward's hedonic value (Clarkson et al., 2018). Several animal-friendly approaches are now available to avoid the negative impact of human handling on mice: (a) adopting non-aversive manual handling techniques, as open-hand retrieval through the cupping method (Hurst and West, 2010; Gouveia and Hurst, 2017, 2019; d'Isa et al., 2021b; Davies et al., 2022); (b) employing a tool to handle the mice, as a plastic handling tunnel (Hurst and West, 2010; Gouveia and Hurst, 2013, 2017, 2019; Sensini et al., 2020; Davies et al., 2022); (c) using automated home-cage testing systems in which behavioral outcomes are recorded without physical interaction with the human experimenter (Kiryk et al., 2020; König et al., 2020; Mitchell et al., 2020; Ho et al., 2022; Kohler et al., 2022; Winiarski et al., 2022).
Furthermore, not knowing the species-specific characteristics of the studied organism, for example, applying inappropriate motivators, forcing the animal to exhibit behavioral responses it would not normally perform, and measuring the behavior under artificial conditions that do not have much to do with the natural environment in which the animal evolved, all can elevate random error, simply because the individuals tested this way may have to find unique solutions to the problems, considerably increasing individual differences in the study (Gerlai and Clayton, 1999a,b). To put it in the words of the aforementioned pioneer of experimental behavioral research Willlard Stanton Small, “the experiments must conform to the psycho-biological character of an animal if sane results are to be obtained” (Small, 1901).
Animal-friendly tests utilizing species-specific features of the studied organism may not be always available or applicable, but, when they are, they should be employed as a first option, in order to maximize both animal welfare and repeatability of experimental results. When fully animal-friendly tests are not available, then the least stressful available test should be employed. In Table 1 we present a rating scale for behavioral tests based on their impact on animal welfare. This rating is not meant to be final, but rather a starting point to stimulate reflection and discussion on the differential stress impact of behavioral tests. We hope that in future an increasing number of studies will employ tests of class A (animal-friendly) and B (minimally stressful) and that, in accordance with a progressive refinement principle, new animal-friendly tests will be designed to substitute the more stressful alternatives.
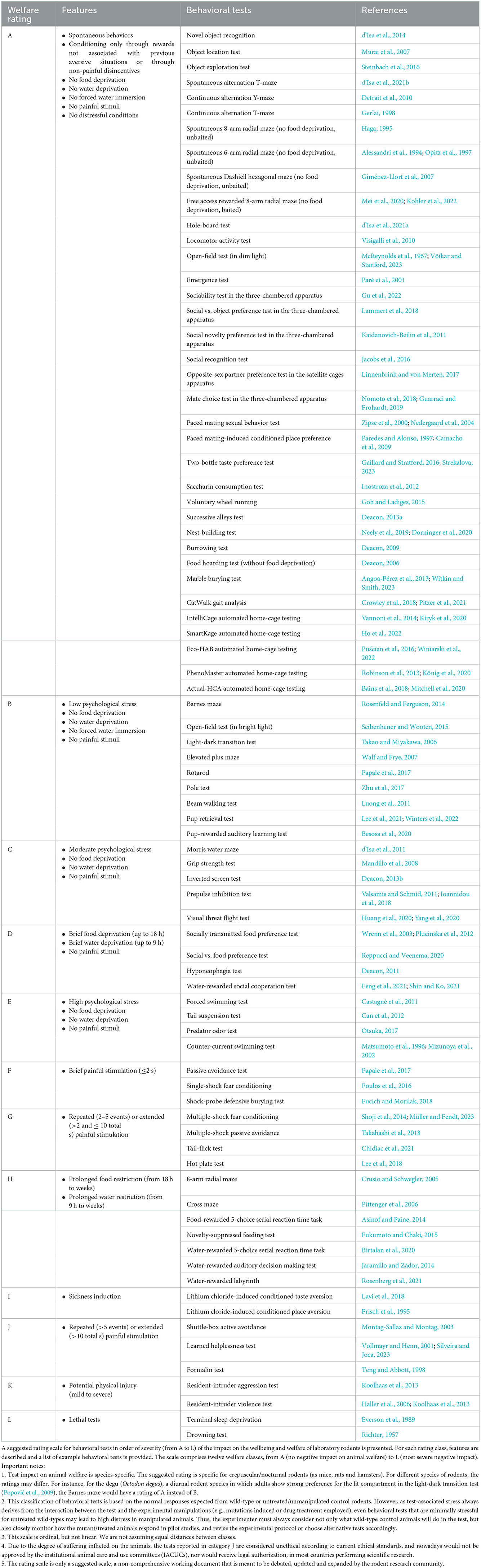
Table 1. Rating of the impact of behavioral tests on animal welfare.
Author contributions
Rd'I and RG: conceptualization, writing, revision, and final approval of the manuscript. Both authors provided funding. Both authors contributed to the article and approved the submitted version.
Funding
The present work was funded by the Natural Science and Engineering Research Council (NSERC) of Canada (Discovery Grant #311637 to RG), by University of Toronto Mississauga Distinguished Professorship Award to RG and by Rd'I.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Alessandri, B., FitzGerald, R. E., Schaeppi, U., Krinke, G. J., and Classen, W. (1994). The use of an unbaited tunnel maze in neurotoxicology: I. Trimethyltin-induced brain lesions. Neurotoxicology 15, 349–357
Angoa-Pérez, M., Kane, M. J., Briggs, D. I., Francescutti, D. M., and Kuhn, D. M. (2013). Marble burying and nestlet shredding as tests of repetitive, compulsive-like behaviors in mice. J. Vis. Exp. 82, 50978. doi: 10.3791/50978
Asinof, S. K., and Paine, T. A. (2014). The 5-choice serial reaction time task: a task of attention and impulse control for rodents. J. Vis. Exp. 90, e51574. doi: 10.3791/51574
Bains, R. S., Cater, H. L., Sillito, R. R., Chartsias, A., Sneddon, D., Concas, D., et al. (2016). Analysis of individual mouse activity in group housed animals of different inbred strains using a novel automated home cage analysis system. Front. Behav. Neurosci. 10, 106. doi: 10.3389/fnbeh.2016.00106
Bains, R. S., Wells, S., Sillito, R. R., Armstrong, J. D., Cater, H. L., Banks, G., et al. (2018). Assessing mouse behaviour throughout the light/dark cycle using automated in-cage analysis tools. J. Neurosci. Methods 300, 37–47. doi: 10.1016/j.jneumeth.2017.04.014
Besosa, C. I., Dunlap, A. G., Lu, K., and Liu, R. C. (2020). Using the pup retrieval instinct as reinforcement for efficient auditory learning in mice. MethodsX 7, 101051. doi: 10.1016/j.mex.2020.101051
Birtalan, E., Bánhidi, A., Sanders, J. I., Balázsfi, D., and Hangya, B. (2020). Efficient training of mice on the 5-choice serial reaction time task in an automated rodent training system. Sci. Rep. 10, 22362. doi: 10.1038/s41598-020-79290-2
Blanchard, D. C., Griebel, G., and Blanchard, R. J. (2003). Conditioning and residual emotionality effects of predator stimuli: some reflections on stress and emotion. Prog. Neuropsychopharmacol. Biol. Psychiatry 27, 1177–1185. doi: 10.1016/j.pnpbp.2003.09.012
Bradley, R. D., Edwards, C. W., Lindsey, L. L., Bateman, J. R., Cajimat, M. N. B., Milazzo, M. L., et al. (2022). Reevaluation of the phylogenetic relationships among Neotomini rodents (Hodomys, Neotoma, and Xenomys) and comments on the woodrat classification. J. Mammal. 103, 1221–1236. doi: 10.1093/jmammal/gyac031
Cain, C. K. (2019). Avoidance problems reconsidered. Curr. Opin. Behav. Sci. 26, 9–17. doi: 10.1016/j.cobeha.2018.09.002
Camacho, F. J., García-Horsman, P., and Paredes, R. G. (2009). Hormonal and testing conditions for the induction of conditioned place preference by paced mating. Horm. Behav. 56, 410–415. doi: 10.1016/j.yhbeh.2009.07.007
Can, A., Dao, D. T., Terrillion, C. E., Piantadosi, S. C., Bhat, S., and Gould, T. D. (2012). The tail suspension test. J. Vis. Exp. 59, e3769. doi: 10.3791/3769
Castagné, V., Moser, P., Roux, S., and Porsolt, R. D. (2011). Rodent models of depression: forced swim and tail suspension behavioral despair tests in rats and mice. Curr. Protoc. Neurosci. 55, 8.10A1–8.10A14. doi: 10.1002/0471142301.ns0810as55
Chidiac, C., Xue, Y., Muniz Moreno, M. D. M., Bakr Rasheed, A. A., Lorentz, R., Birling, M. C., et al. (2021). The human SCN10AG1662S point mutation established in mice impacts on mechanical, heat, and cool sensitivity. Front. Pharmacol. 12, 780132. doi: 10.3389/fphar.2021.780132
Chow, P. K. Y., Lea, S. E. G., Hempel de Ibarra, N., and Robert, T. (2017). How to stay perfect: the role of memory and behavioural traits in an experienced problem and a similar problem. Anim. Cogn. 20, 941–952. doi: 10.1007/s10071-017-1113-7
Chu, X., and Ågmo, A. (2014). Sociosexual behaviours in cycling, intact female rats (Rattus norvegicus) housed in a seminatural environment. Behaviour 151, 1143–1184. doi: 10.1163/1568539X-00003177
Chu, X., Guarraci, F. A., and Ågmo, A. (2015). Sociosexual behaviors and reproductive success of rats (Rattus norvegicus) in a seminatural environment. Physiol. Behav. 151, 46–54. doi: 10.1016/j.physbeh.2015.07.005
Clarkson, J. M., Dwyer, D. M., Flecknell, P. A., Leach, M. C., and Rowe, C. (2018). Handling method alters the hedonic value of reward in laboratory mice. Sci. Rep. 8, 2448. doi: 10.1038/s41598-018-20716-3
Crabbe, J. C., Wahlsten, D., and Dudek, B. C. (1999). Genetics of mouse behavior: interactions with laboratory environment. Science 284, 1670–1672. doi: 10.1126/science.284.5420.1670
Crowley, S. T., Kataoka, K., and Itaka, K. (2018). Combined catwalk index: an improved method to measure mouse motor function using the automated gait analysis system. BMC Res. Notes 11, 263. doi: 10.1186/s13104-018-3374-x
Crusio, W. E. (2001). Genetic dissection of mouse exploratory behaviour. Behav. Brain Res. 125, 127–132. doi: 10.1016/S0166-4328(01)00280-7
Crusio, W. E., and Schwegler, H. (2005). Learning spatial orientation tasks in the radial-maze and structural variation in the hippocampus in inbred mice. Behav. Brai. Funct. 1, 3. doi: 10.1186/1744-9081-1-3
Davies, J. R., Purawijaya, D. A., Bartlett, J. M., and Robinson, E. S. J. (2022). Impact of refinements to handling and restraint methods in mice. Animals 12, 2173. doi: 10.3390/ani12172173
Deacon, R. M. (2006). Assessing hoarding in mice. Nat. Protoc. 1, 2828–2830. doi: 10.1038/nprot.2006.171
Deacon, R. M. (2009). Burrowing: a sensitive behavioural assay, tested in five species of laboratory rodents. Behav. Brain Res. 200, 128–133. doi: 10.1016/j.bbr.2009.01.007
Deacon, R. M. (2011). Hyponeophagia: a measure of anxiety in the mouse. J. Vis. Exp. 51, 2613. doi: 10.3791/2613
Deacon, R. M. (2013a). The successive alleys test of anxiety in mice and rats. J. Vis. Exp. 76, 2705. doi: 10.3791/2705
Dell'Omo, G., Ricceri, L., Wolfer, D. P., Poletaeva, I. I., and Lipp, H. (2000). Temporal and spatial adaptation to food restriction in mice under naturalistic conditions. Behav. Brain Res. 115, 1–8. doi: 10.1016/S0166-4328(00)00234-5
Detrait, E., Brohez, C., Hanon, E., and De Ryck, M. (2010). Automation of continuous spontaneous alternation to increase the throughput for in vivo screening of cognitive enhancers. Optimization of the Ethovision system for the Y-maze test in mice. Proceed. Meas. Behav. 2010, 141–144
d'Isa, R., Brambilla, R., and Fasano, S. (2014). Behavioral methods for the study of the Ras-ERK pathway in memory formation and consolidation: passive avoidance and novel object recognition tests. Methods Mol. Biol. 1120, 131–156. doi: 10.1007/978-1-62703-791-4_9
d'Isa, R., Clapcote, S. J., Voikar, V., Wolfer, D. P., Giese, K. P., Brambilla, R., et al. (2011). Mice lacking ras-GRF1 show contextual fear conditioning but not spatial memory impairments: convergent evidence from two independently generated mouse mutant lines. Front. Behav. Neurosci. 5, 78. doi: 10.3389/fnbeh.2011.0007
d'Isa, R., Comi, G., and Leocani, L. (2021a). The 4-hole-board test for assessment of long-term spatial memory in mice. Curr. Protoc. 1, e228. doi: 10.1002/cpz1.228
d'Isa, R., Comi, G., and Leocani, L. (2021b). Apparatus design and behavioural testing protocol for the evaluation of spatial working memory in mice through the spontaneous alternation T-maze. Sci. Rep. 11, 21177. doi: 10.1038/s41598-021-00402-7
Dorninger, F., Zeitler, G., and Berger, J. (2020). Nestlet shredding and nest building tests to assess features of psychiatric disorders in mice. Bio-Protoc. 10, e3863. doi: 10.21769/BioProtoc.3863
Everson, C. A., Bergmann, B. M., and Rechtschaffen, A. (1989). Sleep deprivation in the rat: III. Total sleep deprivation. Sleep 12, 13–21. doi: 10.1093/sleep/12.1.13
Feng, W., Zhang, Y., Wang, Z., Wang, T., Pang, Y., Zou, Y., et al. (2021). A water-reward task assay for evaluating mouse mutualistic cooperative behaviour. bioRxiv [preprint]. doi: 10.1101/2021.02.06.430037
Frisch, C., Hasenöhrl, R. U., Mattern, C. M., Häcker, R., and Huston, J. P. (1995). Blockade of lithium chloride-induced conditioned place aversion as a test for antiemetic agents: comparison of metoclopramide with combined extracts of Zingiber officinale and Ginkgo biloba. Pharmacol. Biochem. Behav. 52, 321–327. doi: 10.1016/0091-3057(95)00073-6
Fucich, E. A., and Morilak, D. A. (2018). Shock-probe defensive burying test to measure active versus passive coping style in response to an aversive stimulus in rats. Bio-Protoc. 8, e2998. doi: 10.21769/BioProtoc.2998
Fukumoto, K., and Chaki, S. (2015). Involvement of serotonergic system in the effect of a metabotropic glutamate 5 receptor antagonist in the novelty-suppressed feeding test. J. Pharmacol. Sci. 127, 57–61. doi: 10.1016/j.jphs.2014.09.003
Gaillard, D., and Stratford, J. M. (2016). Measurement of behavioral taste responses in mice: two-bottle preference, lickometer, and conditioned taste-aversion tests. Curr. Protoc. Mouse Biol. 6, 380–407. doi: 10.1002/cpmo.18
Galsworthy, M. J., Amrein, I., Kuptsov, P. A., Poletaeva, I. I., Zinn, P., Rau, A., et al. (2005). A comparison of wild-caught wood mice and bank voles in the intellicage: assessing exploration, daily activity patterns and place learning paradigms. Behav. Brain Res. 157, 211–217. doi: 10.1016/j.bbr.2004.06.021
Gerlai, R. (1998). A new continuous alternation task in T-maze detects hippocampal dysfunction in mice. A strain comparison and lesion study. Behav. Brain Res. 95, 91–101. doi: 10.1016/S0166-4328(97)00214-3
Gerlai, R. (2019). Reproducibility and replicability in zebrafish behavioral neuroscience research. Pharmacol. Biochem. Behav. 178, 30–38. doi: 10.1016/j.pbb.2018.02.005
Gerlai, R. (2021). From genes to behavior: the question of evolutionary conservation and the role of ethology in the analysis of the zebrafish. Front. Neuroanat. 15, 809967. doi: 10.3389/fnana.2021.809967
Gerlai, R., and Clayton, N. S. (1999a). Analysing hippocampal function in transgenic mice: an ethological perspective. Trends Neurosci. 22, 47–51. doi: 10.1016/S0166-2236(98)01346-0
Gerlai, R., and Clayton, N. S. (1999b). Tapping artificially into natural talents – reply. Trends Neurosci. 22, 301–302. doi: 10.1016/S0166-2236(99)01421-6
Gerlai, R., Crusio, W. E., and Csányi, V. (1990). Inheritance of species specific behaviors in the paradise fish (Macropodus opercularis): a diallel study. Behav. Genet. 20, 487–498. doi: 10.1007/BF01067715
Giménez-Llort, L., Schiffmann, S. N., Shmidt, T., Canela, L., Camón, L., Wassholm, M., et al. (2007). Working memory deficits in transgenic rats overexpressing human adenosine A2A receptors in the brain. Neurobiol. Learn. Mem. 87, 42–56. doi: 10.1016/j.nlm.2006.05.004
Goh, J., and Ladiges, W. (2015). Voluntary wheel running in mice. Curr. Protoc. Mouse Biol. 5, 283–290. doi: 10.1002/9780470942390.mo140295
Gouveia, K., and Hurst, J. L. (2013). Reducing mouse anxiety during handling: effect of experience with handling tunnels. PLoS ONE 8, e66401. doi: 10.1371/journal.pone.0066401
Gouveia, K., and Hurst, J. L. (2017). Optimising reliability of mouse performance in behavioural testing: The major role of non-aversive handling. Sci. Rep. 7, 44999. doi: 10.1038/srep44999
Gouveia, K., and Hurst, J. L. (2019). Improving the practicality of using non-aversive handling methods to reduce background stress and anxiety in laboratory mice. Sci. Rep. 9, 20305. doi: 10.1038/s41598-019-56860-7
Gu, Y., Han, Y., Ren, S., Zhang, B., Zhao, Y., Wang, X., et al. (2022). Correlation among gut microbiota, fecal metabolites and autism-like behavior in an adolescent valproic acid-induced rat autism model. Behav. Brain Res. 417, 113580. doi: 10.1016/j.bbr.2021.113580
Guarraci, F. A., and Frohardt, R. J. (2019). “What a girl wants”: what can we learn from animal models of female sexual motivation? Front. Behav. Neurosci. 13, 216. doi: 10.3389/fnbeh.2019.00216
Haga, Y. (1995). Effects of food deprivation and food reward on the behavior of rats in the radial- arm maze. Jpn. Psychol. Res. 37, 252–257. doi: 10.4992/psycholres1954.37.252
Haller, J., Tóth, M., Halasz, J., and De Boer, S. F. (2006). Patterns of violent aggression-induced brain c-fos expression in male mice selected for aggressiveness. Physiol. Behav. 88, 173–182. doi: 10.1016/j.physbeh.2006.03.030
Harries, C., Mourgue, G., and Purefoy, J. (2020). Amazing Animal Friends. Documentary Series, Episode 1-03. London: Oxford Scientific Films.
Harris, R. B. S. (2017). “The hamster as a model for human ingestive behavior”, in Appetite and Food Intake: Central Control, 2nd edn, eds R. B. S. Harris (Boca Raton, FL: CRC Press/Taylor and Francis), Chapter 3.
Heinla, I., Chu, X., Ågmo, A., and Snoeren, E. (2021). Rat ultrasonic vocalizations and novelty-induced social and non-social investigation behavior in a seminatural environment. Physiol. Behav. 237, 113450. doi: 10.1016/j.physbeh.2021.113450
Ho, H., Kejzar, N., Sasaguri, H., Saito, T., Saido, T. C., De Strooper, B., et al. (2022). A novel fully-automated system for lifelong continuous phenotyping of mouse cognition and behaviour. bioRxiv [preprint]. doi: 10.1101/2022.06.18.496688
Holmes, A., le Guisquet, A. M., Vogel, E., Millstein, R. A., Leman, S., and Belzung, C. (2005). Early life genetic, epigenetic and environmental factors shaping emotionality in rodents. Neurosci. Biobehav. Rev. 29, 1335–1346. doi: 10.1016/j.neubiorev.2005.04.012
Houwing, D. J., Heijkoop, R., Olivier, J. D. A., and Snoeren, E. M. S. (2019). Perinatal fluoxetine exposure changes social and stress-coping behavior in adult rats housed in a seminatural environment. Neuropharmacology 151, 84–97. doi: 10.1016/j.neuropharm.2019.03.037
Huang, Y., Li, L., Dong, K., Tang, H., Yang, Q., Jia, X., et al. (2020). topological shape changes weaken the innate defensive response to visual threat in mice. Neurosci. Bull. 36, 427–431. doi: 10.1007/s12264-019-00454-w
Hurst, J. L., and West, R. S. (2010). Taming anxiety in laboratory mice. Nat. Methods 7, 825–826. doi: 10.1038/nmeth.1500
Iman, I. N., Yusof, N. A. M., Talib, U. N., Ahmad, N. A. Z., Norazit, A., Kumar, J., et al. (2021). The intellicage system: a review of its utility as a novel behavioral platform for a rodent model of substance use disorder. Front. Behav. Neurosci. 15, 683780. doi: 10.3389/fnbeh.2021.683780
Inostroza, M., Cid, E., Menendez de la Prida, L., and Sandi, C. (2012). Different emotional disturbances in two experimental models of temporal lobe epilepsy in rats. PLoS ONE 7, e38959. doi: 10.1371/journal.pone.0038959
Ioannidou, C., Marsicano, G., and Busquets-Garcia, A. (2018). Assessing prepulse inhibition of startle in mice. Bio-Protoc. 8, e2789. doi: 10.21769/BioProtoc.2789
Jacobs, S. A., Huang, F., Tsien, J. Z., and Wei, W. (2016). Social recognition memory test in rodents. Bio-Protoc. 6, e1804. doi: 10.21769/BioProtoc.1804
Jaramillo, S., and Zador, A. M. (2014). Mice and rats achieve similar levels of performance in an adaptive decision-making task. Front. Syst. Neurosci. 8, 173. doi: 10.3389/fnsys.2014.00173
Jarvik, M. E., and Kopp, R. (1967). An improved one-trial passive avoidance learning situation. Psychol. Rep. 21, 221–224. doi: 10.2466/pr0.1967.21.1.221
Kafkafi, N., Agassi, J., Benjamini, Y., Chesler, E., Crabbe, J., Crusio, W., et al. (2018). Reproducibility and Replicability of mouse phenotyping in pre-clinical studies. Neurosci. Biobehav. Rev. 87, 218–232. doi: 10.1016/j.neubiorev.2018.01.003
Kaidanovich-Beilin, O., Lipina, T., Vukobradovic, I., Roder, J., and Woodgett, J. R. (2011). Assessment of social interaction behaviors. J. Vis. Exp. 48, 2473. doi: 10.3791/2473
Kampmann, M. (2020). CRISPR-based functional genomics for neurological disease. Nat. Rev. Neurol. 16, 465–480. doi: 10.1038/s41582-020-0373-z
Kaufman, D. M., and Kaufman, D. W. (1984). Size preference for novel objects by the eastern woodrat (Neotoma floridana) under field conditions. Trans. Kans. Acad. Sci. 87, 129–131. doi: 10.2307/3627848
Kempermann, G., Lopes, J. B., Zocher, S., Schilling, S., Ehret, F., Garthe, A., et al. (2022). The individuality paradigm: automated longitudinal activity tracking of large cohorts of genetically identical mice in an enriched environment. Neurobiol. Dis. 175, 105916. doi: 10.1016/j.nbd.2022.105916
Kiryk, A., Janusz, A., Zglinicki, B., Turkes, E., Knapska, E., Konopka, W., et al. (2020). IntelliCage as a tool for measuring mouse behavior - 20 years perspective. Behav. Brain Res. 388, 112620. doi: 10.1016/j.bbr.2020.112620
Kline, L. W. (1899a). Methods in animal psychology. Am. J. Psychol. 10, 256–279. doi: 10.2307/1412481
Kline, L. W. (1899b). Suggestions toward a laboratory course in comparative psychology. Am. J. Psychol. 10, 399–430. doi: 10.2307/1412142
Kohler, J., Mei, J., Banneke, S., Winter, Y., Endres, M., and Emmrich, J. V. (2022). Assessing spatial learning and memory in mice: classic radial maze versus a new animal-friendly automated radial maze allowing free access and not requiring food deprivation. Front. Behav. Neurosci. 16, 1013624. doi: 10.3389/fnbeh.2022.1013624
König, C., Plank, A. C., Kapp, A., Timotius, I. K., von Hörsten, S., and Zimmermann, K. (2020). Thirty mouse strain survey of voluntary physical activity and energy expenditure: influence of strain, sex and day-night variation. Front. Neurosci. 14, 531. doi: 10.3389/fnins.2020.00531
Koolhaas, J. M., Coppens, C. M., de Boer, S. F., Buwalda, B., Meerlo, P., and Timmermans, P. J. (2013). The resident-intruder paradigm: a standardized test for aggression, violence and social stress. J. Vis. Exp. 77, e4367. doi: 10.3791/4367
Kopp, R., Bohdanecky, Z., and Jarvik, M. E. (1966). Long temporal gradient of retrograde amnesia for a well-discriminated stimulus. Science. 153, 1547–1549. doi: 10.1126/science.153.3743.1547
Lammert, C. R., Frost, E. L., Bolte, A. C., Paysour, M. J., Shaw, M. E., Bellinger, C. E., et al. (2018). Cutting edge: critical roles for microbiota-mediated regulation of the immune system in a prenatal immune activation model of autism. J. Immunol. 201, 845–850. doi: 10.4049/jimmunol.1701755
Lavi, K., Jacobson, G. A., Rosenblum, K., and Lüthi, A. (2018). Encoding of conditioned taste aversion in cortico-amygdala circuits. Cell Rep. 24, 278–283. doi: 10.1016/j.celrep.2018.06.053
Le Moëne, O., Hernández-Arteaga, E., Chu, X., and Ågmo, A. (2020). Rapid changes in sociosexual behaviors around transition to and from behavioral estrus, in female rats housed in a seminatural environment. Behav. Processes 174, 104101. doi: 10.1016/j.beproc.2020.104101
Lee, Y. C., Lu, S. C., and Hsieh, Y. L. (2018). Establishing a mouse model of a pure small fiber neuropathy with the ultrapotent agonist of transient receptor potential vanilloid type 1. J. Vis. Exp. 132, 56651. doi: 10.3791/56651
Lee, Y. J., Lin, H. T., Chaudhary, M. A., Lee, Y. C., and Wang, D. C. (2021). Effects of prenatal phthalate exposure and childhood exercise on maternal behaviors in female rats at postpartum: a role of oxtr methylation in the hypothalamus. Int. J. Mol. Sci. 22, 9847. doi: 10.3390/ijms22189847
Linnenbrink, M., and von Merten, S. (2017). No speed dating please! Patterns of social preference in male and female house mice. Front. Zool. 14, 38. doi: 10.1186/s12983-017-0224-y
Luong, T. N., Carlisle, H. J., Southwell, A., and Patterson, P. H. (2011). Assessment of motor balance and coordination in mice using the balance beam. J. Vis. Exp. 49, 2376. doi: 10.3791/2376
Mandillo, S., Tucci, V., Hölter, S. M., Meziane, H., Banchaabouchi, M. A., Kallnik, M., et al. (2008). Reliability, robustness, and reproducibility in mouse behavioral phenotyping: a cross-laboratory study. Physiol. Genomics 34, 243–255. doi: 10.1152/physiolgenomics.90207.2008
Matsumoto, K., Ishihara, K., Tanaka, K., Inoue, K., and Fushiki, T. (1996). An adjustable-current swimming pool for the evaluation of endurance capacity of mice. J. Appl. Physiol. (1985) 81, 1843–1849. doi: 10.1152/jappl.1996.81.4.1843
McReynolds, W. E., Weir, M. W., and DeFries, J. C. (1967). Open-field behavior in mice: effect of test illumination. Psychon. Sci. 9, 277–278. doi: 10.3758/BF03332220
Mei, J., Kohler, J., Winter, Y., Spies, C., Endres, M., Banneke, S., et al. (2020). Automated radial 8-arm maze: a voluntary and stress-free behavior test to assess spatial learning and memory in mice. Behav. Brain Res. 381, 112352. doi: 10.1016/j.bbr.2019.112352
Miles, W. R. (1930). On the history of research with rats and mazes: a collection of notes. J. Gen. Psychol. 3, 324–337. doi: 10.1080/00221309.1930.9918210
Mills, T. W. (1888). Squirrels: their habits and intelligence, with special reference to feigning. Proceed. Trans. Royal Soc. Canada (1st Series) 5, 175–188.
Mills, T. W. (1893). Hibernation and allied states in animals. Proceed. Trans. Royal Soc. Canada (1st Series) 10, 49–66.
Mills, T. W. (1895). The psychic development of young animals and its physical correlation: V – the rabbit and the cavy or guinea pig. Proceed. Trans. Royal Soc. Canada (2nd Series) 1, 231–239.
Mingrone, A., Kaffman, A., and Kaffman, A. (2020). The promise of automated home-cage monitoring in improving translational utility of psychiatric research in rodents. Front. Neurosci. 14, 618593. doi: 10.3389/fnins.2020.618593
Mitchell, E. J., Brett, R. R., Armstrong, J. D., Sillito, R. R., and Pratt, J. A. (2020). Temporal dissociation of phencyclidine: induced locomotor and social alterations in rats using an automated homecage monitoring system - implications for the 3Rs and preclinical drug discovery. J. Psychopharmacol. 34, 709–715. doi: 10.1177/0269881120920455
Mitjans, M., Begemann, M., Ju, A., Dere, E., Wüstefeld, L., Hofer, S., et al. (2017). Sexual dimorphism of AMBRA1-related autistic features in human and mouse. Transl. Psychiatry 7, e1247. doi: 10.1038/tp.2017.213
Mizunoya, W., Oyaizu, S., Ishihara, K., and Fushiki, T. (2002). Protocol for measuring the endurance capacity of mice in an adjustable-current swimming pool. Biosci. Biotechnol. Biochem. 66, 1133–1136. doi: 10.1271/bbb.66.1133
Montag-Sallaz, M., and Montag, D. (2003). Severe cognitive and motor coordination deficits in tenascin-R-deficient mice. Genes Brain Behav. 2, 20–31. doi: 10.1034/j.1601-183X.2003.00003.x
Montoya, B., and Gutiérrez, G. (2016). Social context modulates food hoarding in Syrian hamsters. Int. J. Psychol. Res. 9, 61–69. doi: 10.21500/20112084.2646
Moreira, P. S., Almeida, P. R., Leite-Almeida, H., Sousa, N., and Costa, P. (2016). Impact of chronic stress protocols in learning and memory in rodents: systematic review and meta-analysis. PLoS ONE 11, e0163245. doi: 10.1371/journal.pone.0163245
Müller, I., and Fendt, M. (2023). Fear conditioning in laboratory rodents. Neuromethods 190, 119–160. doi: 10.1007/978-1-0716-2748-8_8
Murai, T., Okuda, S., Tanaka, T., and Ohta, H. (2007). Characteristics of object location memory in mice: behavioral and pharmacological studies. Physiol. Behav. 90, 116–124. doi: 10.1016/j.physbeh.2006.09.013
Nedergaard, P., Sanchez, C., and Mellerup, E. (2004). Different roles of 5-HT2A and 5-HT2C receptors in regulation of female rat paced mating behaviour. Behav. Brain Res. 149, 151–157. doi: 10.1016/S0166-4328(03)00215-8
Neely, C. L. C., Pedemonte, K. A., Boggs, K. N., and Flinn, J. M. (2019). Nest building behavior as an early indicator of behavioral deficits in mice. J. Vis. Exp. 152, e60139. doi: 10.3791/60139
Nomoto, K., Ikumi, M., Otsuka, M., Asaba, A., Kato, M., Koshida, N., et al. (2018). Female mice exhibit both sexual and social partner preferences for vocalizing males. Integr. Zool. 13, 735–744. doi: 10.1111/1749-4877.12357
Nowak, R. M., and Paradiso, J. L. (1983). Walker's Mammals of the World (4th Edition). Baltimore: The Johns Hopkins University Press.
Opitz, B., Mothes, H. K., and Clausing, P. (1997). Effects of prenatal ethanol exposure and early experience on radial maze performance and conditioned taste aversion in mice. Neurotoxicol. Teratol. 19, 185–190. doi: 10.1016/S0892-0362(96)00225-5
Otsuka, S. (2017). Predator odor-induced freezing test for mice. Bio-Protoc. 7, e2534. doi: 10.21769/BioProtoc.2534
Papale, A., d'Isa, R., Menna, E., Cerovic, M., Solari, N., Hardingham, N., et al. (2017). Severe intellectual disability and enhanced gamma-aminobutyric acidergic synaptogenesis in a novel model of rare rasopathies. Biol. Psychiatry 81, 179–192. doi: 10.1016/j.biopsych.2016.06.016
Paré, W. P., Tejani-Butt, S., and Kluczynski, J. (2001). The emergence test: effects of psychotropic drugs on neophobic disposition in Wistar Kyoto (WKY) and Sprague Dawley rats. Prog. Neuropsychopharmacol. Biol. Psychiatry 25, 1615–1628. doi: 10.1016/S0278-5846(01)00204-4
Paredes, R. G., and Alonso, A. (1997). Sexual behavior regulated (paced) by the female induces conditioned place preference. Behav. Neurosci. 111, 123–128. doi: 10.1037/0735-7044.111.1.123
Pittenger, C., Fasano, S., Mazzocchi-Jones, D., Dunnett, S. B., Kandel, E. R., and Brambilla, R. (2006). Impaired bidirectional synaptic plasticity and procedural memory formation in striatum-specific cAMP response element-binding protein-deficient mice. J. Neurosci. 26, 2808–2813. doi: 10.1523/JNEUROSCI.5406-05.2006
Pitzer, C., Kurpiers, B., and Eltokhi, A. (2021). Gait performance of adolescent mice assessed by the CatWalk XT depends on age, strain and sex and correlates with speed and body weight. Sci. Rep. 11, 21372. doi: 10.1038/s41598-021-00625-8
Plucinska, K., Strachan, L., Peeters, D., Platt, B., and Riedel, G. (2012). Social transmission of food preference in C57BL/6 mice. Proceed. Meas. Behav. 2012, 488–490.
Popović, N., Baño-Otálora, B., Rol, M. A., Caballero-Bleda, M., Madrid, J. A., and Popović, M. (2009). Aging and time-of-day effects on anxiety in female octodon degus. Behav. Brain 200, 117–121. doi: 10.1016/j.bbr.2009.01.001
Poulos, A. M., Mehta, N., Lu, B., Amir, D., Livingston, B., Santarelli, A., et al. (2016). Conditioning- and time-dependent increases in context fear and generalization. Learn. Mem. 23, 379–385. doi: 10.1101/lm.041400.115
Puścian, A., Łeski, S., Kasprowicz, G., Winiarski, M., Borowska, J., Nikolaev, T., et al. (2016). Eco-HAB as a fully automated and ecologically relevant assessment of social impairments in mouse models of autism. Elife 5, e19532. doi: 10.7554/eLife.19532.040
Redman, K. R. (1974). The Effectiveness of Dusting as a Reinforcer for the Chinchilla (Chinchilla Lanigera). University of North Dakota Theses and Dissertations 3448, 1–88 (Master's Degree Thesis). Grand Forks, ND: University of North Dakota.
Reppucci, C. J., and Veenema, A. H. (2020). The social versus food preference test: a behavioral paradigm for studying competing motivated behaviors in rodents. MethodsX 7, 101119. doi: 10.1016/j.mex.2020.101119
Richter, C. P. (1957). On the phenomenon of sudden death in animals and man. Psychosom. Med. 19, 191–198. doi: 10.1097/00006842-195705000-00004
Robinson, L., Plano, A., Cobb, S., and Riedel, G. (2013). Long-term home cage activity scans reveal lowered exploratory behaviour in symptomatic female Rett mice. Behav. Brain Res. 250, 148–156. doi: 10.1016/j.bbr.2013.04.041
Rosenberg, M., Zhang, T., Perona, P., and Meister, M. (2021). Mice in a labyrinth show rapid learning, sudden insight, and efficient exploration. Elife 10, e66175. doi: 10.7554/eLife.66175.sa2
Rosenfeld, C. S., and Ferguson, S. A. (2014). Barnes maze testing strategies with small and large rodent models. J. Vis. Exp. 84, e51194. doi: 10.3791/51194
Ruzza, C., Asth, L., Guerrini, R., Trapella, C., and Gavioli, E. C. (2015). Neuropeptide S reduces mouse aggressiveness in the resident/intruder test through selective activation of the neuropeptide S receptor. Neuropharmacology 97, 1–6. doi: 10.1016/j.neuropharm.2015.05.002
Seibenhener, M. L., and Wooten, M. C. (2015). Use of the open field maze to measure locomotor and anxiety-like behavior in mice. J. Vis. Exp. 96, e52434. doi: 10.3791/52434
Sensini, F., Inta, D., Palme, R., Brandwein, C., Pfeiffer, N., Riva, M. A., et al. (2020). The impact of handling technique and handling frequency on laboratory mouse welfare is sex-specific. Sci. Rep. 10, 17281. doi: 10.1038/s41598-020-74279-3
Shin J. and Ko, J.. (2021). Protocol for quantitative assessment of social cooperation in mice. STAR Protoc. 2, 100305. doi: 10.1016/j.xpro.2021.100305
Shoji, H., Takao, K., Hattori, S., and Miyakawa, T. (2014). Contextual and cued fear conditioning test using a video analyzing system in mice. J. Vis. Exp. 85, 50871. doi: 10.3791/50871
Silveira, K. M., and Joca, S. (2023). Learned helplessness in rodents. Neuromethods 190, 161–184. doi: 10.1007/978-1-0716-2748-8_9
Small, W. S. (1899). Notes on the psychic development of the young white rat. Am. J. Psych. 11, 80–100. doi: 10.2307/1412730
Small, W. S. (1900). An experimental study of the mental processes of the rat. Am. J. Psych. 11, 133–165. doi: 10.2307/1412267
Small, W. S. (1901). Experimental study of the mental process of the rat. II. Am. J. Psych. 12, 206–239. doi: 10.2307/1412534
Steinbach, J. M., Garza, E. T., and Ryan, B. C. (2016). Novel object exploration as a potential assay for higher order repetitive behaviors in mice. J. Vis. Exp. 114, 54324. doi: 10.3791/54324
Stern, J. J., and Merari, A. (1969). The bathing behavior of the chinchilla: effects of deprivation. Psychon. Sci. 14, 115. doi: 10.3758/BF03332734
Stewart, C. C. (1898). Variations in daily activity produced by alcohol and by changes in barometric pressure and diet, with a description of recording methods. Am. J. Physiol. 1, 40–56. doi: 10.1152/ajplegacy.1898.1.1.40
Strekalova, T. (2023). How the sucrose preference succeeds or fails as a measurement of anhedonia. Neuromethods 190, 89–102. doi: 10.1007/978-1-0716-2748-8_6
Takahashi, S., Ohmiya, M., Honda, S., and Ni, K. (2018). The KCNH3 inhibitor ASP2905 shows potential in the treatment of attention deficit/hyperactivity disorder. PLoS ONE 13, e0207750. doi: 10.1371/journal.pone.0207750
Takao, K., and Miyakawa, T. (2006). Light/dark transition test for mice. J. Vis. Exp. 1, 104. doi: 10.3791/104
Teng, C. J., and Abbott, F. V. (1998). The formalin test: a dose-response analysis at three developmental stages. Pain 76, 337–347. doi: 10.1016/S0304-3959(98)00065-7
Ueno, H., Takahashi, Y., Suemitsu, S., Murakami, S., Kitamura, N., Wani, K., et al. (2020). Effects of repetitive gentle handling of male C57BL/6NCrl mice on comparative behavioural test results. Sci. Rep. 10, 3509. doi: 10.1038/s41598-020-60530-4
Urbach, Y. K., Raber, K. A., Appl, T., Häberle, L., and von Horsten, S. (2008). Automated phenotyping using intra-home-cage technology. Proceed. Measur. Behav. 2008, 330.
Valsamis, B., and Schmid, S. (2011). Habituation and prepulse inhibition of acoustic startle in rodents. J. Vis. Exp. 55, e3446. doi: 10.3791/3446
Vannoni, E., Voikar, V., Colacicco, G., Sánchez, M. A., Lipp, H. P., and Wolfer, D. P. (2014). Spontaneous behavior in the social homecage discriminates strains, lesions and mutations in mice. J. Neurosci. Methods 234, 26–37. doi: 10.1016/j.jneumeth.2014.04.026
Visigalli, I., Delai, S., Politi, L. S., Di Domenico, C., Cerri, F., Mrak, E., et al. (2010). Gene therapy augments the efficacy of hematopoietic cell transplantation and fully corrects mucopolysaccharidosis type I phenotype in the mouse model. Blood 116, 5130–5139. doi: 10.1182/blood-2010-04-278234
Voikar, V., and Gaburro, S. (2020). Three pillars of automated home-cage phenotyping of mice: novel findings, refinement, and reproducibility based on literature and experience. Front. Behav. Neurosci. 14, 575434. doi: 10.3389/fnbeh.2020.575434
Võikar, V., and Stanford, S. C. (2023). The open field test. Neuromethods 190, 9–29. doi: 10.1007/978-1-0716-2748-8_2
Vollmayr, B., and Henn, F. A. (2001). Learned helplessness in the rat: improvements in validity and reliability. Brain Res. Brain Res. Protoc. 8, 1–7. doi: 10.1016/S1385-299X(01)00067-8
Vyssotski, A. L., Dell'Omo, G., Poletaeva, I. I., Vyssotsk, D. L., Minichiello, L., Klein, R., et al. (2002). Long-term monitoring of hippocampus-dependent behavior in naturalistic settings: mutant mice lacking neurotrophin receptor TrkB in the forebrain show spatial learning but impaired behavioral flexibility. Hippocampus 12, 27–38. doi: 10.1002/hipo.10002
Walf, A. A., and Frye, C. A. (2007). The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat. Protoc. 2, 322–328. doi: 10.1038/nprot.2007.44
Warner, L. (1932). The association span of the white rat. J. Genet. Psychol. 41, 57–90. doi: 10.1080/08856559.1932.9944143
Winiarski, M., Kondrakiewicz, L., Kondrakiewicz, K., Jedrzejewska-Szmek, J., Turzyński, K., Knapska, E., et al. (2022). Social deficits in BTBR T+ Itpr3tf/J mice vary with ecological validity of the test. Genes Brain Behav. 21, e12814. doi: 10.1111/gbb.12814
Winters, C., Gorssen, W., Ossorio-Salazar, V. A., Nilsson, S., Golden, S., and D'Hooge, R. (2022). Automated procedure to assess pup retrieval in laboratory mice. Sci. Rep. 12, 1663. doi: 10.1038/s41598-022-05641-w
Witkin, I. M., and Smith, J. L. (2023). Marble burying in mice. Neuromethods 190, 71–88. doi: 10.1007/978-1-0716-2748-8_5
Wrenn, C. C., Harris, A. P., Saavedra, M. C., and Crawley, J. N. (2003). Social transmission of food preference in mice: methodology and application to galanin-overexpressing transgenic mice. Behav. Neurosci. 117, 21–31. doi: 10.1037/0735-7044.117.1.21
Yang, X., Liu, Q., Zhong, J., Song, R., Zhang, L., and Wang, L. (2020). A simple threat-detection strategy in mice. BMC Biol. 18, 93. doi: 10.1186/s12915-020-00825-0
Zhang, Y., Yu, F., Yi, X., Zhou, W., Liu, R., Holyoak, M., et al. (2022). Evolutionary and ecological patterns of scatter- and larder-hoarding behaviours in rodents. Ecol. Lett. 25, 1202–1214. doi: 10.1111/ele.13992
Zhu, K., Sun, J., Kang, Z., Zou, Z., Wu, G., and Wang, J. (2017). Electroacupuncture promotes remyelination after cuprizone treatment by enhancing myelin debris clearance. Front. Neurosci. 10, 613. doi: 10.3389/fnins.2016.00613
Zipse, L. R., Brandling-Bennett, E. M., and Clark, A. S. (2000). Paced mating behavior in the naturally cycling and the hormone-treated female rat. Physiol. Behav. 70, 205–259. doi: 10.1016/S0031-9384(00)00242-0
Keywords: behavioral testing, ethology, refinement, animal welfare, animal-friendly, rodents
Citation: d'Isa R and Gerlai R (2023) Designing animal-friendly behavioral tests for neuroscience research: The importance of an ethological approach. Front. Behav. Neurosci. 16:1090248. doi: 10.3389/fnbeh.2022.1090248
Received: 05 November 2022; Accepted: 16 December 2022;
Published: 10 January 2023.
Edited by:
Valerie J. Bolivar, Wadsworth Center, United StatesReviewed by:
Hans-Peter Lipp, University of Zurich, SwitzerlandCopyright © 2023 d'Isa and Gerlai. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Raffaele d'Isa,  ZGlzYS5yYWZmYWVsZUBoc3IuaXQ=; Robert Gerlai, cm9iZXJ0LmdlcmxhaUB1dG9yb250by5jYQ==
ZGlzYS5yYWZmYWVsZUBoc3IuaXQ=; Robert Gerlai, cm9iZXJ0LmdlcmxhaUB1dG9yb250by5jYQ==