 Jan Tekülve
Jan Tekülve Adrien Fois
Adrien Fois Yulia Sandamirskaya
Yulia Sandamirskaya Gregor Schöner
Gregor Schöner- 1Institute for Neural Computation, Ruhr-University Bochum, Bochum, Germany
- 2Lorraine Research Laboratory in Computer Science and its Applications, Vandœuvre-lès-Nancy, France
- 3Institute for Neuroinformatics, University of Zürich and ETZ Zürich, Zurich, Switzerland
Neurally inspired robotics already has a long history that includes reactive systems emulating reflexes, neural oscillators to generate movement patterns, and neural networks as trainable filters for high-dimensional sensory information. Neural inspiration has been less successful at the level of cognition. Decision-making, planning, building and using memories, for instance, are more often addressed in terms of computational algorithms than through neural process models. To move neural process models beyond reactive behavior toward cognition, the capacity to autonomously generate sequences of processing steps is critical. We review a potential solution to this problem that is based on strongly recurrent neural networks described as neural dynamic systems. Their stable states perform elementary motor or cognitive functions while coupled to sensory inputs. The state of the neural dynamics transitions to a new motor or cognitive function when a previously stable neural state becomes unstable. Only when a neural robotic system is capable of acting autonomously does it become a useful to a human user. We demonstrate how a neural dynamic architecture that supports autonomous sequence generation can engage in such interaction. A human user presents colored objects to the robot in a particular order, thus defining a serial order of color concepts. The user then exposes the system to a visual scene that contains the colored objects in a new spatial arrangement. The robot autonomously builds a scene representation by sequentially bringing objects into the attentional foreground. Scene memory updates if the scene changes. The robot performs visual search and then reaches for the objects in the instructed serial order. In doing so, the robot generalizes across time and space, is capable of waiting when an element is missing, and updates its action plans online when the scene changes. The entire flow of behavior emerges from a time-continuous neural dynamics without any controlling or supervisory algorithm.
1. Introduction
Neurally inspired robotics already has a long history. To position our work in this history and review our conceptual commitments, we discuss three strands of neurallly inspired robotics.
1.1. Reactive Behaviors
One strand goes back to Grey's electronic turtle (Grey, 1950) and Braitenberg's thought experiments on vehicles (Braitenberg, 1984). This line of work reached maturity in behavior-based robotics (Brooks, 1991; Mataric, 1998) in which flexibility emerges from the coordination of elementary behaviors, each establishing a direct link from sensory inputs to actuators, in the manner of reflex loops. This is particularly suited to conceptual “vehicles,” robotic systems in which the sensors are mounted on the moving actuator. This enables closed loop situations that greatly reduce the demands on representation and abstraction. For instance, a visual sensor mounted in a robot hand makes it possible to achieve reaching by visual servoing without an explicit representation of objects in the world (Ruf and Horaud, 1999).
By organizing closed action-perception loops in architectures, most famously the subsumption architecture (Brooks, 1986), this form of reactive robotics may generate behaviors of a certain complexity (Proetzsch et al., 2010). The behavior is generated autonomously in the sense that sensory information from a structured environment may trigger the activation of elementary behaviors, which may lead to chains of activation and deactivation events through the architecture, inducing sequences of behavioral decisions, without the need for an explicit internal plan, schedule, or program. The organization of such behaviors is implicitly encoded in the architecture itself.
Avoiding representation and abstraction is a feature of the approach (Brooks, 1990), but also points to a limitation of this line of neurally inspired robotics: Behavior-based robots are not very good at cognition. Minimally, cognition is engaged when the link between sensing and acting becomes less direct. Building and exploiting memory is an example (Engels and Schöner, 1995). So when an action is based on sensory information that is no longer directly available on the sensory surface at the time the action unfolds, relevant information must be represented in memory. Memories are useful only if they are represented in a form in which they remain invariant under changes the system experiences between the acquisition of the memory and its use. For instance, the memory representation of a movement target for a vehicle needs to be invariant under rotation of the vehicle (Bicho et al., 2000). A more demanding form of cognition is the capacity to perceive sequences of events and store them in a memory for serial order so that a sequence with a matching serial order can then be acted out (such as hearing a phone number and then dialing it). Again, the information needs to abstract from the sensor data to be useful for the required actions.
Our approach is historically based on behavior-based thinking, which we extended by adding neural memory representations and neural mechanism of decision making (Schöner et al., 1995). Here we will study how memory for serial order can be built and used to act sequentially in new environments.
1.2. Neuronal Oscillators and Pattern Generators
A second strand of neurally inspired robotics is based on the idea that neural oscillators may generate rhythmic movement patterns. That idea has been used to generate legged locomotion in biologically inspired robots (Holmes et al., 2006; Ijspeert, 2008). Such neural oscillator ideas can be integrated with the dynamics of limbs and muscles and their interaction with the ground, enabling stable locmotion patterns (Full and Koditschek, 1999; Ghigliazza et al., 2003). Neural oscillators are one important class of neural networks in which recurrent connections are strong enough to induce endogenous patterns of neural activation that are not mere transformations of input. That class can be extended to neural timers that generate complex temporal patterns that may be the basis for certain motor skills (Buonomano and Laje, 2010). Coupling neural oscillators provides an account for coordination (Schöner and Kelso, 1988) and adaptation enables the modulation of rhythmic movement patterns (Aoi et al., 2017).
Typically, however, these kinds of models do not address how movement may be directed at targets in the world, such as when reaching for an object or intercepting a ball. A related class of neural models going back, perhaps, to Bullock and Grossberg (1988), generates time courses by integrating neural activity toward an end-point that may ultimately be determined by perceptual processes. This is the basis of the notion of dynamic movement primitives (Schaal et al., 2003), which is still broadly neurally inspired although it is typically implemented in a mathematical form that does not explicitly reference neural processing principles (see Ijspeert et al., 2013 for an excellent review). The dynamical systems framework for reaching toward objects can address how such movement is directed at objects in the world (Hersch and Billard, 2008). Typically, however, the representation of the object's pose and kinematic state remains clearly outside the neural metaphor (while achieving superhuman performance in skills such as catching, Kim et al., 2014).
Our approach builds on this tradition of using neural oscillators for timing. We generate individual goal-directed reaches from an active transient solution of a recurrent neural dynamics. We extend this tradition by providing a neural dynamic architecture that obtains from the visual array a neural representation of the targets of a reaching movement. This requires that an object's visual coordinates are transformed into coordinates anchored in the initial position of the hand (Schöner et al., 2019). We show how such a neural representation of movement targets may be linked to the visual array, enabling online updating of movement generation when the scene changes (see Knips et al., 2017 for an earlier version of such online updating).
1.3. Neural Networks for Perception
A third strand of neural inspiration for embodied cognitive systems is, of course, the use of neural networks to extract relevant information about the environment from sensory (e.g., image, sound) data (Kriegeskorte, 2015). This strand is currently undergoing explosive growth as the scaling of deep neural networks in size and learning examples enables superhuman performance in certain classification and detection tasks (Lecun et al., 2015; Schmidhuber, 2015). These neural networks essentially serve as intelligent filters of sensory information, a critical function when robot cognition is to be linked to the world.
While these networks by themselves do not perform cognitive functions, they may provide outputs that enable cognition. For instance, networks may deliver labels for a relational description of a visual scene (e.g., Kelleher and Dobnik, 2017). In most cases, the actual reasoning about spatial or other relations is, however, performed outside a neural processes model, based on algorithms and probabilistic inference. First steps are being made, however, toward such models generating the sequential attentional selection on which human visual cognition is centrally based (Ba et al., 2015).
Our approach is based on the classical notion of feature extraction along the visual pathway, the simplest step in these kinds of systems (e.g., Serre et al., 2007). As we do not address object recognition, we limit ourselves to very simple features here (see Lomp et al., 2017 for how the approach may link to object recognition). Instead, we demonstrate how a neural dynamic system may autonomously generate the sequence of attentional selections to build a visual scene memory that is intermittently coupled to the visual array, and thus is sensitive to change and capable of updating in response to such change.
1.4. Goals
In this paper, we integrate these three strands of neurally inspired robotics which requires us to extend each of them. Our emphasis is on how the integrated system—essentially a network of neural dynamic populations—is continuously or intermittently coupled to sensory information, while at the same time being capable of autonomously generating sequences of decisions, actions, and events. Neural activation is thus generated endogenously is this system, while retaining the coupling to the sensory surfaces.
The system addressed four key elements of grounded cognition: (1) It autonomously builds scene memory, a neural map of locations and feature values bound to those locations. Different objects are sequentially brought into the attentional foreground, in each case creating an entry into scene memory, which can be updated if change is detected. (2) The system learns the serial order of events that occur in its visual array. Each time an attended object changes, the system registers the transition and learns the new feature value as associated with its serial position. This provides a possible interface through which a human user can interact with the system. (3) The system generates a sequence of actions oriented at objects in the world in the learned serial order. At any point in the sequence, this entails finding an object in the visual surrounding that matches the feature values currently sought, generating the action, and then transitioning to the next sub-task within the learned sequence. This exemplifies the capacity of the system to autonomously generate organized behavior that is not merely reactive but reflective of a learned plan. (4) Each action consists of a pointing gesture oriented at an attended object. The action is initiated once the object has been brought into the attentional foreground, but may be updated any time if object shifts to a new location. This is a minimal instantiation of object-oriented action that any form of cognitive robotics must be capable of.
Key to this demonstration is the notion of neural dynamics, in which strongly recurrent neural networks, approximated as spatially and temporally continuous neural fields, evolve primarily under the influence of their internal interaction that sets up attractor states. Inputs induce instabilities that bring about switches of neural states from which sequences of cognitive or motor states emerge. Such neural dynamics are capable of making decisions, building working memories, and organizing sequential transitions (Schöner et al., 2016). Because their neural states are stable, neural fields retain their functional properties when they are coupled to other fields. Fields may thus serve as building blocks of networks of fields, which could be thought of as neural dynamic architectures. These networks may be coupled to sensory inputs, while evolving under their own, endogenous dynamics, resolving the tension between reactive and cognitive systems.
To make the ideas accessible, we restrict the demonstration to a very simple scenario. A robot observes a table top on which a human user places and removes colored objects in a particular serial order. The user then builds a new visual scene, that includes the objects with colors contained in the taught series. The robot points at these objects in the order defined by the human teacher. When an object of the next required color is not available, the system waits until such a color is presented. When the visual array changes, the robot updates its reaching plans. This may happen online if the change occurs while the robot is already attempting to point at the object. All action and observation run autonomously in neural dynamics. There is no control algorithm outside the neural dynamics. See Supplementary Videos 1 and 2 for exemplary demonstrations of teaching and executing the series.
2. Dynamic Field Theory
We use Dynamic Field Theory (DFT) (Schöner et al., 2016) as a conceptual framework. DFT provides neural process accounts for elementary cognitive functions such as decision making, memory creation, or the generation of sequences. The core elements of DFT are neural populations which may generate activation patterns that are not primarily dictated by input. This is based on structured and strong recurrent connectivity within the population. Excitatory recurrent connectivity enables detection decisions in which neural activation is induced by input, but then stabilized against decay even as input may weaken again. The initial detection occurs through an instability, in which the resting state becomes unstable. The detection is reversed when the activated state becomes unstable, typically at a lower level of input than needed for initial detection. If the excitatory recurrency is sufficiently strong, the reverse detection instability does not happen, leading to activation that is sustained even when the inducing input is removed entirely. This is the basis for working memory.
Inhibitory recurrent connectivity enables selection decisions, in which one sub-population becomes activated even if multiple sub-populations receive supra-threshold input. Such selection decisions are also stabilized so that the selection of a sub-population may persist even as inputs to other sub-populations become stronger (up to a limit, when the selection instability is encountered). So even though neural populations may be driven by input, they may realize non-unique mappings from input to activation states based on their activation history.
When different populations are coupled, they may induce these kinds of instabilities in each other. This is the basis for generating sequences of neural activation states. When the coupling occurs between excitatory and inhibitory sub-populations, the instabilities may trigger active transients, well-defined time courses of neural activation from temporally unstructured input. Neural oscillations are another possible dynamic regime.
Through their connectivity to sensory or motor surfaces, neural populations may effectively represent continuous feature dimensions, x. This leads to the notion of neural dynamic fields, u(x). We employ a particular mathematical formalization of the dynamics of such neural populations that goes back to Amari (1977),
where τ describes the field's relaxation time, h < 0 the field's resting level, s the sum of input stimuli, and ω the field's interaction kernel that defines the pattern of recurrent connectivity within the field. Only sufficiently activated field locations contribute to interaction or project onto other fields, as formalized by the sigmoidal non-linearity, σ(u). Thus, one may think of the activation variable, u, as something like a population-level membrane potential that reflects how close neurons in the population are to the firing threshold [other formalizations use the firing rate as a population variable, see Wilson and Cowan (1973)]. In the meantime, there is a large literature on the mathematics of such fields (Coombes et al., 2014).
The kernel, ω, combines short-range excitatory coupling with long-range inhibitory coupling. This leads to localized peaks of activation as the activation states that emerge from the instability of the resting state when localized input reaches a threshold (Figure 1). These peaks are the units of representation in DFT that specify through their locations particular values along the represented dimension.
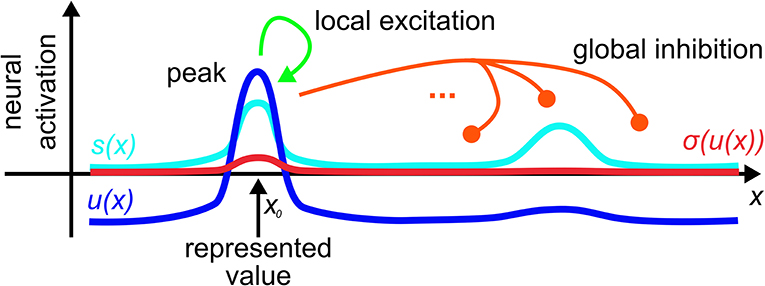
Figure 1. A dynamic neural field spanning a metric dimension, x, represents a specific value, x0, along that dimension through a supra-threshold activation peak that is stabilized by local excitatory and global inhibitory interactions.
Fields may represent low-dimensional metric spaces. When their dimensionality grows, the binding problem arises and can be solved, see Chapter 5 of Schöner et al. (2016). A limit case are zero-dimensional fields which can be thought of as populations of neurons that represent categorical states. These may arise from larger populations through inhomogoneities in the input or output connectivity. We sometimes call such zero-dimensional fields neural dynamic nodes and model them by single activation variables, u(t), subject to a neural dynamics analogous to Equation (1).
When fields of different dimensionality are coupled, new functions emerge (Zibner et al., 2011, see also Chapter 9 of Schöner et al., 2016). In projecting from a higher to a lower dimensional field, certain dimensions may be marginalized, which effectively probes for the existence of a peak anywhere along the marginalized dimensions. In projecting from a lower to a higher dimensional field, a boost may be given to a subspace, enabling locations within the subspace to reach the detection instability. This is the basis for visual search. The control of peak formation in a field through homogeneous boosting of its activation level is a mechanism of control that may effectively gate particular projections by enabling or disabling peak formation. This mechanism is also central to sequence generation through the condition of satisfaction (CoS) (Sandamirskaya and Schöner, 2010) that will play a central role in our sequence representation model. The neural representation of the CoS is a neural field or a neural node that is pre-activated by the currently active behavior. That behavior predicts the sensory or internal state that will indicate its successful completion. When a signal matching that prediction is received from sensory inputs or from other neural processes, the CoS goes through a detection instability. It then inhibits the current behavior in a reverse detection instability and enables the activation of a new behavior (Sandamirskaya and Schöner, 2010).
3. Model
The neural dynamic architecture described here is a network of neural fields that are coupled to a camera and a robotic arm. These links enable online connection to a changing visual scene and online control of the arm. Three sub-networks (Figure 2) autonomously organize sequences of activation states to build visual representations, learn or perform serially ordered sequences, and generate object-oriented movements.
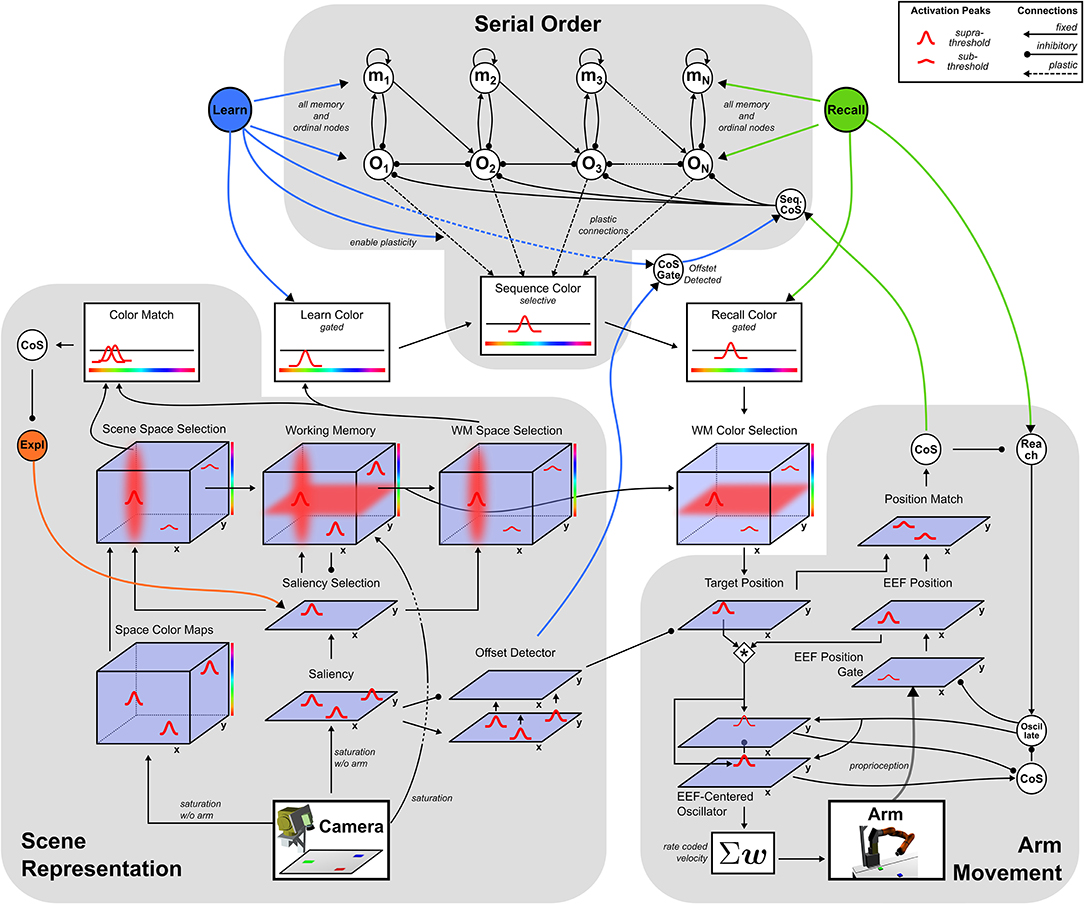
Figure 2. Sketch of the dynamic field network with its three sub-networks.
The perceptual sub-network, connected to the camera, creates a working memory representation of the visual scene through autonomous shifts of attention. A motor sub-network drives an oscillator generating velocity commands for the robotic arm. The cognitive sub-network represents ordinal positions in a sequence and may autonomously shift from one ordinal position to the next. The ordinal system may be used in two different manners, sequence learning and sequence recall, controlled by the activation of one of two different task nodes. These task nodes activate behaviors by boosting fields' resting levels and enabling fields to generate task relevant attractor states.
The following sections describe for each sub-network the states that drive behavior and the mechanism for how the system switches between those states. The last section addresses the integration of all three sub-networks for the two tasks Learn and Recall.
3.1. Perception: Scene Representation
The scene representation sub-network is based on Grieben et al. (2018) and creates three-dimensional (2D space and 1D color) working memory representations of objects in the visual scene captured by the camera. Each entry into the representation is created sequentially as the sub-network autonomously shifts attention across different objects in the scene.
The network's attention is modeled through peaks of activation in the two-dimensional Saliency Selection field that arise at salient locations in the scene. These locations are represented in the Saliency field which receives input directly from the camera. Based on their distinctive colors, the table and the robot's own arm are subtracted from the image in a pre-processing step. The saturation channel of the resulting HSV-image serves as input amplitude at each location.
Combined with a homogeneous boost of its resting level from the Exploration intention node, activation in the Saliency field is sufficient to create a single peak in the Saliency Selection field. Attentional shifts occur whenever the Exploration node deactivates and subsequently reactivates, causing a destabilization of the present peak in the Selection field followed by the emergence of a new peak at a new location. Previously unattended locations are more likely to be selected because inhibitory influence from the working memory gives them a competitive advantage.
The activation of the Saliency Selection field, usel, is governed by the following neural dynamics:
where hsel describes the field's resting level, uexp the homogeneous boost activation from the Explore intention node, usal(x, y) the activation of the Saliency field, ∫ σ(umem(x, y, c))dc the activation of the Working Memory projected onto the two spatial dimensions, x and y, and ωsel the field's selective lateral interaction kernel. Each input, σuin, to the field is weighted by a specific weight, win. The same notation is used in all following equations and the concrete parameter values can be found in the Appendix.
The currently attended location achieves spatial feature binding: It is forwarded to the three 3D space-color fields, Scene Space Selection, Working Memory, and WM Space Selection, ensuring that the color features represented in those fields originate from the same location. The Scene Space Selection field combines sub-threshold activation from the Space Color Maps field, that represents color information in the scene, with spatial sub-threshold activation from the Saliency Selection field. Together, these inputs induce a single peak in three dimensions that represents the attended spatial location and the color perceived at that location.
That color is extracted and combined with spatial information from the Saliency Selection field to add a peak in the 3D-Working Memory field.1 The Working Memory field receives additional input directly from the camera image that includes the robot's arm. That input is proportional to the saturation channel from which the table color saturation has been subtracted. This mask, seen, for instance, in the Working Memory row of Figure 4, makes it possible to sustain peaks of activation anywhere where the camera picks up visual structure. Peaks representing objects can thus remain stable in working memory when they become occluded by the robot's arm, but are removed from working memory when they disappear from the scene at any other location.
The activation, umem(x, y, c), of the Working Memory field is governed by the following dynamics:
where hmem describes the field's resting level, usel(x, y) the activation from the Saliency Selection field, inbk(x, y) the saturation channel from the camera image, ∫ σ (ussl (x, y, c))dxdy the activation of the Scene Space Selection field projected onto the color dimension, c, and ωmem the field's lateral interaction kernel.
Supra-threshold activation of the Working Memory field is forwarded to the Memory Space Selection field, which works analogously to the Scene Space Selection field and thus forms a single 3D activation peak, representing color and spatial location of the attended location in working memory.
Color information represented in the Scene and Memory Space Selection fields is forwarded to the Color Match field. That field forms a peak only when the input from the scene overlaps in location and color with one of the peaks in the memory field. A peak in the match field thus signals successful entry of an item into the Working Memory at the currently attended location. Supra-threshold activation in the match field projects onto the CoS Explore node, which in turn inhibits the Explore intention node. Deactivation of the Explore intention node removes the resting level boost from the Saliency Selection field inducing a reverse detection instability that propagates to the Scene and Memory Space Selection fields, the Color Match field and ultimately to the CoS Explore node. The newly created peak in the Working Memory field is sustained and the Explore intention node is released from inhibition enabling attentional selection of a new location.
3.1.1. Offset Detector
The scene representation sub-network is capable of detecting sudden object movement with the help of a two-layer offset detector connected to the Saliency field. Both layers, udfa and udsl, are two-dimensional fields over image space that are governed by the following dynamics with timescales, τdfa < τdsl:
where hdet describes the common resting level, and σ(usal(x, y)) the thresholded activation of the Saliency field which excites the slower layer, udsl, and inhibits the faster layer, udfa. Because inhibitory input is stronger than excitatory input (wsin > wdsl), static visual structure induces supra-threshold activation in the slow layer, udsl, not in the fast layer, udsl.
Once an object is removed from the scene, the inhibitory influence, wsin, vanishes faster than the excitatory influence from the slow layer, udsl, leading to the formation of a peak in udfa that represents the detection of an object that moves away from the location of the peak.
3.2. Motor: Arm Movement
The sub-network responsible for reaching movements, based on Schöner et al. (2019), autonomously drives an oscillator that creates velocity commands which move a robotic arm to a given target in two-dimensional space. A hierarchy of intention and CoS nodes governs the behavior: The Reach intention node activates the Oscillate intention node, which initiates an active transient (see Figure 3). The Cos Oscillate node is activated once the transient reaches a new steady state, while the CoS Reach is activated when the representations of target and end-effector position match. Thus, multiple active transients (oscillations) are generated until the arm reaches the represented target.
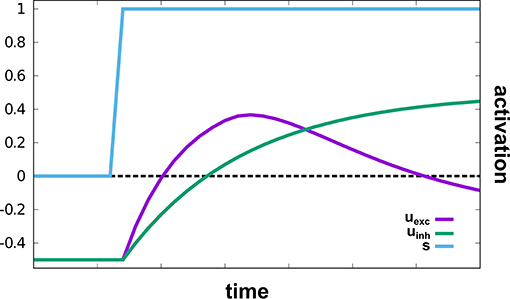
Figure 3. An active transient in uexc generated by a two-layer oscillator in response to an input stimulus, s. The supra-threshold activation level leads to a bell-shaped velocity profile on read-out. This illustration is zero-dimensional while in the model, a two-dimensional field of identical oscillators is used.
Target and end-effector (EEF) are both represented as peaks of activation in two-dimensional fields defined over image space, the Target Position and the EEF Position DNFs, respectively. Activation originating from working memory causes the creation of a peak in the Target Position field. Proprioceptive information from the current arm configuration is mapped through a forward kinematics into end-effector space and then transformed from rate to space code inducing a peak in a two-dimensional EEF Position field. Target and end-effector representations are cross-correlated with each other to create an end-effector centered representation of the target position. This representation is input into a two-layer field of neural oscillators, uexc and uinh. The faster excitatory layer, uexc, generates an active transient illustrated in Figure 3: Its input first drives up excitation, which is then suppressed by inhibition from the slower inhibitory layer, uinh:
where τexc < τinh are the different relaxation times, hosc the resting level, σ(ucct(x, y)) the end-effector-centered target representation, σ(uosc) the homogeneous resting level boost from the Oscillate intention node, and θ – a semi-linear threshold function.
The thresholded activation, θ(uexc(x, y)), is transformed into a rate coded Cartesian velocity vector, v, using a set of feed-forward weights, wvel(x, y):
The weights, wvel(x, y), describe a linear distance function in the end-effector centered representation of the target position. For different movement distances, (x, y), these weights are tuned such that the arm reaches the target position within a fixed movement time. The velocity vector, v, is transformed into a joint velocity vector, , using the pseudo-inverse of the arm's Jacobian, J+, which depends on the current joint configuration λ(t):
For more details on the generated velocity profile see Schöner et al. (2019).
While the oscillator is going, its input is not updated, because the connection from proprioception to the EEF-Position field is gated by the Oscillate intention node. The EEF-Position field thus effectively represents the initial position of the hand. Termination of the transient is detected by the CoS Oscillate node, which receives excitatory activation from uinh and inhibitory activation from uexc. Activation of CoS Oscillate inhibits the Oscillate intention node, which resets the oscillator, and releases the EEF Position Gate from inhibition so that the end-effector position is updated. When the target representation overlaps sufficiently with the updated EEF Position, a peak forms in the Position Match field and activates the CoS Reach, which terminates the reach.
3.3. Cognition: Serial Order
The serial order sub-network, based on Sandamirskaya and Schöner (2010), allows for the autonomous storage and recall of a sequence of activation patterns. Each activation pattern is represented through learned inhomogeneous connections between an ordinal node and a feature field, here the one-dimensional Sequence Color field. Supra-threshold activation in a particular ordinal node thus induces a peak in the Sequence Color field that represents the color associated with that particular stage in the sequence.
The sub-network consisting of ordinal nodes, memory nodes and a single CoS node enforces the sequential activation of ordinal nodes in a fixed order:
An active ordinal node, oi, representing the ith position in the sequence, inhibits all other ordinal nodes, oj, and activates its own self-sustained memory node, mi. The memory node pre-activates the next ordinal node, oi+1, through an excitatory connection and inhibits its own ordinal node, oi, to prevent it from becoming reactivated after completion of the stage. While activated, an ordinal node's self excitation, woi,oi, is sufficient to overcome inhibition from its memory node, wmi,oi. An ordinal node remains active until the CoS node, uCoS, is activated and destabilizes all ordinal nodes, which, in turn, removes input from the CoS node that deactivates. The self-sustained memory nodes are unaffected, so that upon release from inhibition by the CoS, the pre-activated ordinal node of the next element in the sequence is activated. Recurring activation and deactivation of the CoS node thus creates a sequence of autonomous transitions between sequence elements in the order of ascending i. Ordinal and memory nodes can become activated only in the presence of an excitatory boost, wh+, from one of the task nodes, Learn (ulrn) or, Recall (urcl). Deactivation of an active task node leads to deactivation of all memory and ordinal nodes, effectively resetting the entire system.
Connection weights, woi,ucol, between the active ordinal node, oi, and the active region in the Sequence Color field, ucol, are strengthened according to a dynamic version of the Hebbian learning rule:
where η describes the learning rate and ulrn the activation of the Learn task node that gates the learning process.
Before learning, peaks in the Sequence Color field arise when a color attended in the Working Memory Selection field is input through the gate field, Learn color, ulcol:
After learning, peaks in the Sequence color field may arise from previously learned connections, woi,ucol(c), of an ordinal node, oi. The selective kernel, ωcol, ensures that only a single color is represented at all times.
3.4. Task Integration: Learn and Recall
The full network may operate in two different regimes: In the learning regime, a sequence of colors is presented to the system and learned. In the recall regime, a learned sequence of colors is reproduced by pointing at colored objects in a specific order. Each regime is evoked by the activation of its corresponding task node, Learn and Recall, which alter the resting level of certain sub-sets of fields.
Both task nodes boost the resting level of all ordinal and memory nodes to allow supra-threshold activation. When task nodes are deactivated, the removal of the corresponding boost causes activation of all self-sustained nodes to decay, effectively resetting the system. This happens, for instance, at the end of the sequence due to activation of the sequence's condition of satisfaction.
The Learn node acts as a gate between the Scene Representation and the Serial Order sub-networks. By boosting the Learn Color field, the Learn node enables that field to form supra-threshold peaks. At which color such a peak is erected is controlled by input from the Memory Space Selection field that represents the color at the currently attended location. That color is then imprinted in the connections to the currently active ordinal node through the learning dynamics (Equation 9). The Learn node pre-activates the Offset Detected node, which connects to the Sequence CoS. Thus, whenever a single object is presented in the learning regime, its color is associated with the currently active ordinal node and its removal from the scene causes a transition in which the active ordinal node is replaced by the next ordinal node.
The Recall node is a gate between the sequence generation and the arm movement sub-networks. It boosts the Recall Color gating field so that the color represented in the Sequence Color field is passed on to the three-dimensional Memory Color Selection field. If an object in working memory overlaps with that color, a peak forms in the Memory Color Selection field. The peak's spatial position is forwarded to the Target Position field of the Arm Movement sub-network, which initiates a reaching movement. Once a reach has been successfully performed, the Reach CoS is activated, which triggers the Sequence CoS, causing the transition to the next ordinal node. In the recall regime, the arm will thus move autonomously to colored objects in the learned order, as long as appropriately colored objects are visible in the scene.
4. Results
In this section we show how activation within the network unfolds in time during the learn and recall tasks. We visualize relevant activation fields to illustrate how the network's autonomy enables it to cope with variable timing during learning and with changes of the scene during recall.
The network is effectively a large dynamical system. We solved it numerically on digital computers, and that numerical solution was the only form in which algorithms intervened in the system. The numerical implementation of the model made use of CEDAR (Lomp et al., 2016), an open source framework in which DFT models can be graphically assembled and interactively tuned. Cedar can be used to simulate robotic behavior, which was done for the results illustrated in this paper. The visual scene, camera, and robot arm were simulated using WEBOTS (Michel, 2004) that can be coupled into Cedar. The same Cedar code can also link to real sensors and robots. We did this, driving the model from a real camera and manipulating the visual scene by placing colored objects on a white table top. We also controlled a lightweight KUKA arm from the same Cedar code to verify its capacity to act out the planned movements. These informal robotic experiments are not further documented in this paper.
4.1. Scene Representation: Autonomous Build-up of Visual Working Memory
The build-up of the scene working memory is an ongoing process that provides visual information to the network irrespective of the currently active task node. In Figure 4 we show activation snapshots of different points in time during working memory build-up in an exemplary scene containing three objects and the arm's end-effector.
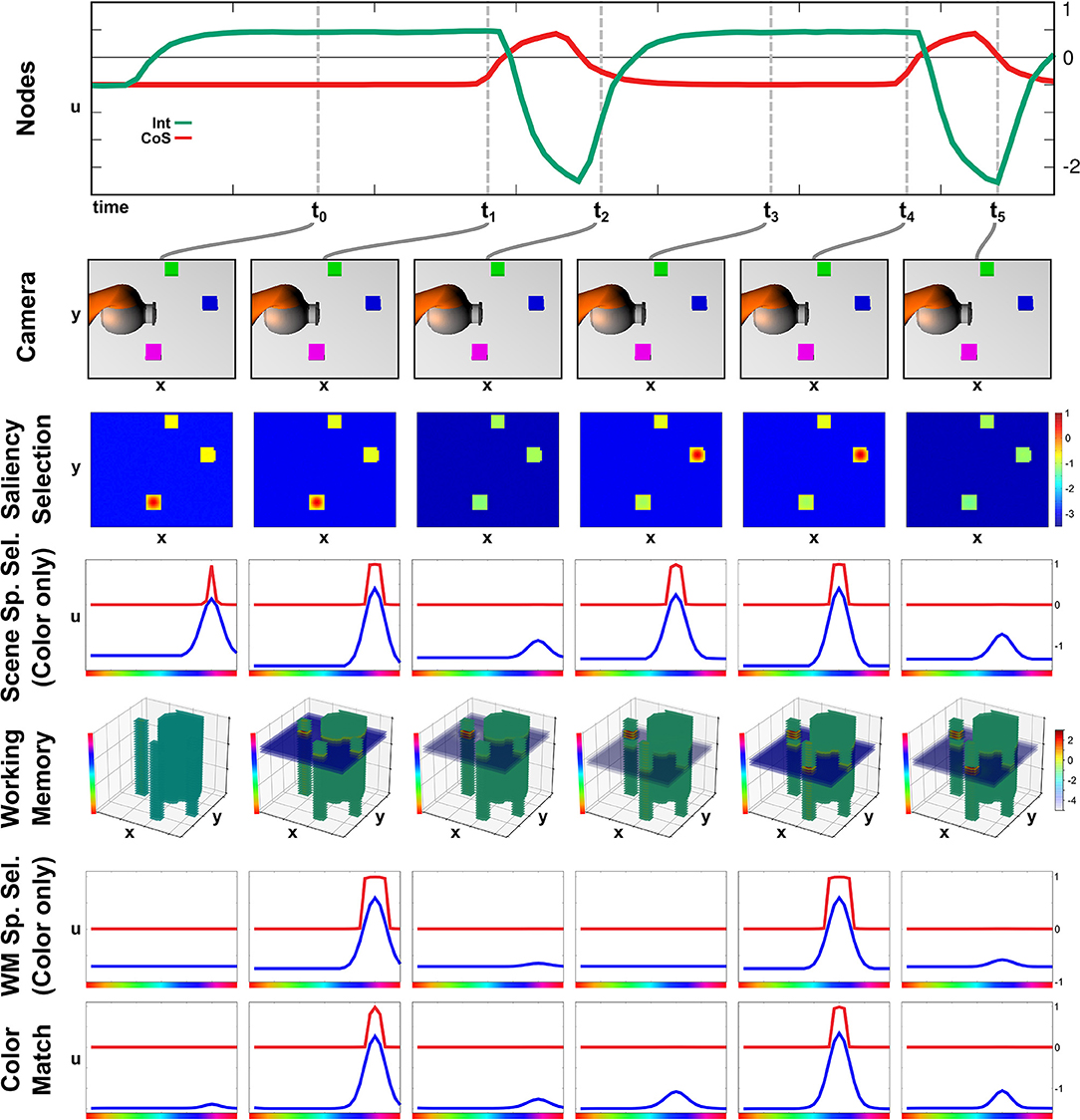
Figure 4. Time course of building a scene memory.
At point t0, the Exploration intention node provides a homogeneous boost to the Saliency Selection field leading to an activation peak at the location of the purple object. This causes the emergence of a three-dimensional peak in the Scene Selection field, of which the color dimension is shown in the third row. The Working Memory field contains no supra-threshold activation yet but, at the locations of the non-background objects, the resting level is increased across the whole color dimension.
Once the peak in the Scene Selection field has fully emerged at t1, its color component is forwarded as a slice toward the Working Memory, where it overlaps with the tube originating from the Saliency Selection field and forms a three-dimensional peak. Subsequently a peak also forms in the Memory Spatial Selection field, which shares the same color as the peak in the Scene Space Selection causing an overlap in the Color Match field.
The peak forming in the Color Match field activates the CoS Explore node, which inhibits the Explore intention node. Thus the resting level boost is removed from the Saliency Selection field, which subsequently falls down to sub-threshold activation at point t2. Only the self-sustained peak in the Working Memory field remains.
The absence of a peak in the Color Match field causes the CoS node to fall below threshold again, bringing the sub-network to its initial state. The following activation of the Explore intention node, depicted from t3 until t5, follows the same temporal activation pattern as the previous one with different feature values for spatial location and color. The spatial location in the Saliency Selection field differs due to the inhibitory influence from the Working Memory field. See Supplementary Video 3 for a different example of autonomous build-up of visual working memory in continuous time.
4.2. Learning Demonstration
A particular color sequence is taught to the network in its learning regime by presenting objects of a certain color one after another. In Figure 5 activation snapshots of some points in time during an exemplary learning episode are shown. The top row depicts the temporal evolution of activation of the ordinal nodes and the Sequence CoS node, while each snapshot column shows the camera image, the activation of the Saliency field, activation of the fast layer of the Offset Detector, activation of the Sequence Color field, and the weight values, woi,ucol, for each ordinal node at one particular point in time.
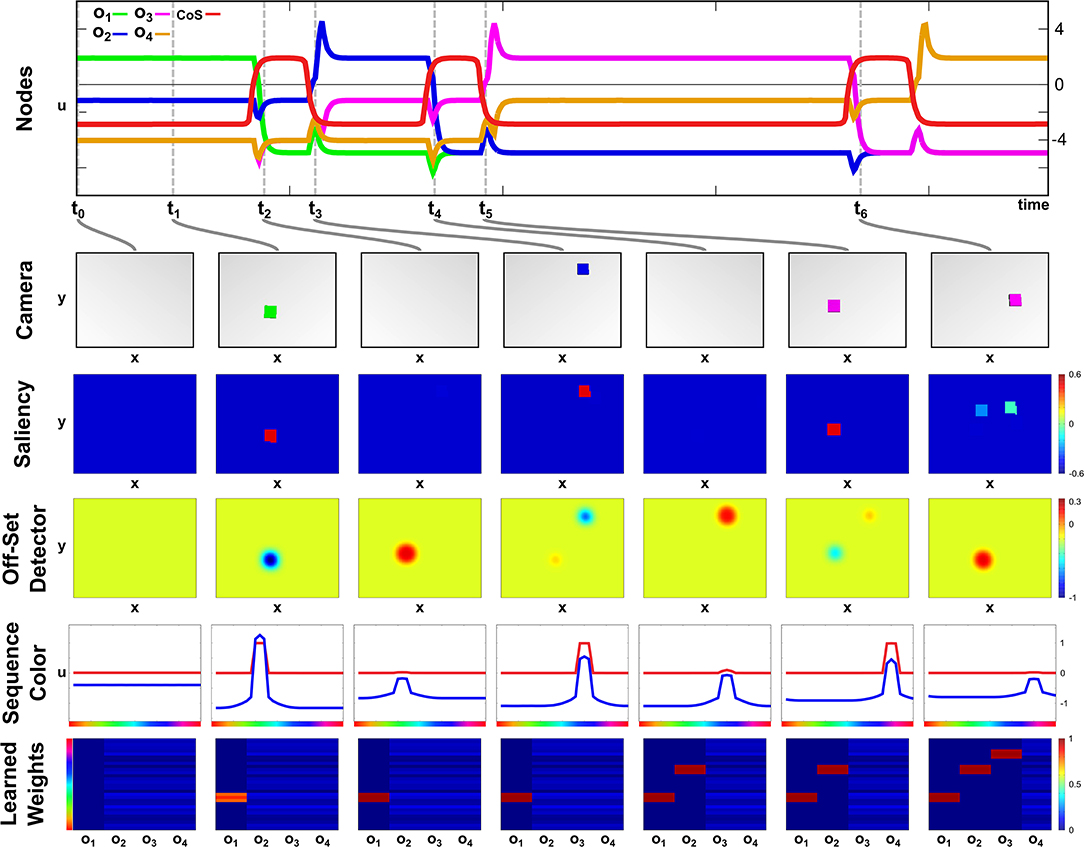
Figure 5. Time course of learning a three element sequence with varying presentation time.
In the initial phase of the learning at point t0 no objects are in the scene, but the Learning task node has been activated leading to supra-threshold activation in the first ordinal node. All other ordinal nodes are below threshold activity with a slight advantage for o2, which already receives an excitatory bias through the active memory node, m1.
At t1, a green object is inserted into the scene, which forms a peak in the Saliency field leading to a localized inhibition in the fast OffSet Detector field. It is also committed to working memory and leads to the emergence of a peak in the Sequence Color field encoding the green color. Due to present supra-threshold activation in the Sequence Color field and the ordinal node o1, the Hebbian learning rule strengthens weights between the ordinal node and the green color feature values.
The object is removed from the scene at t2, which destabilizes the peak in the Saliency field removing the inhibition from the fast layer of the Offset Detector. The slow layer (not depicted) still carries supra-threshold activation, exciting the fast layer leading to the formation of a peak, which will subsequently activate the Sequence CoS node inhibiting all ordinal nodes. This deactivates o1 and causes the color peak in the Sequence Color field to vanish as it is no longer supported by either learned connections nor color input from the scene. The missing input in the scene will also ultimately lead to a decay of activation in the slow Offset Detector layer and subsequently cause a reverse-detection instability in the fast layer and the Sequence CoS node.
The deactivation of the Sequence CoS node is followed by an activation of the next ordinal node o2 at t3. Between t2 and t3 a blue object has been added to the scene, whose color is then connected to the freshly activated ordinal node via the Hebbian learning rule. Removal of the object at t4 triggers the Offset Detector and the CoS node enabling the activation of the next ordinal node o3 at t5. The presented purple object is kept in the scene for a longer time span than the green or blue one, which does not influence the learning as the transition to the next sequence element at t6 is based on the removal event rather than timing.
4.3. Recall Demonstration
We demonstrate successful sequence recall through a pointing task, where the network moves the arm to an object in the scene matching the color of the current sequence element. Only a successful reach toward that object allows a progress to the next sequence element. An exemplary recall of three sequence elements is depicted in Figure 6, which demonstrates the temporal evolution of the activation of ordinal nodes as well as the field activity of the Sequence Color, the Target Position, and the Position Match field at discrete points during the sequence recall.
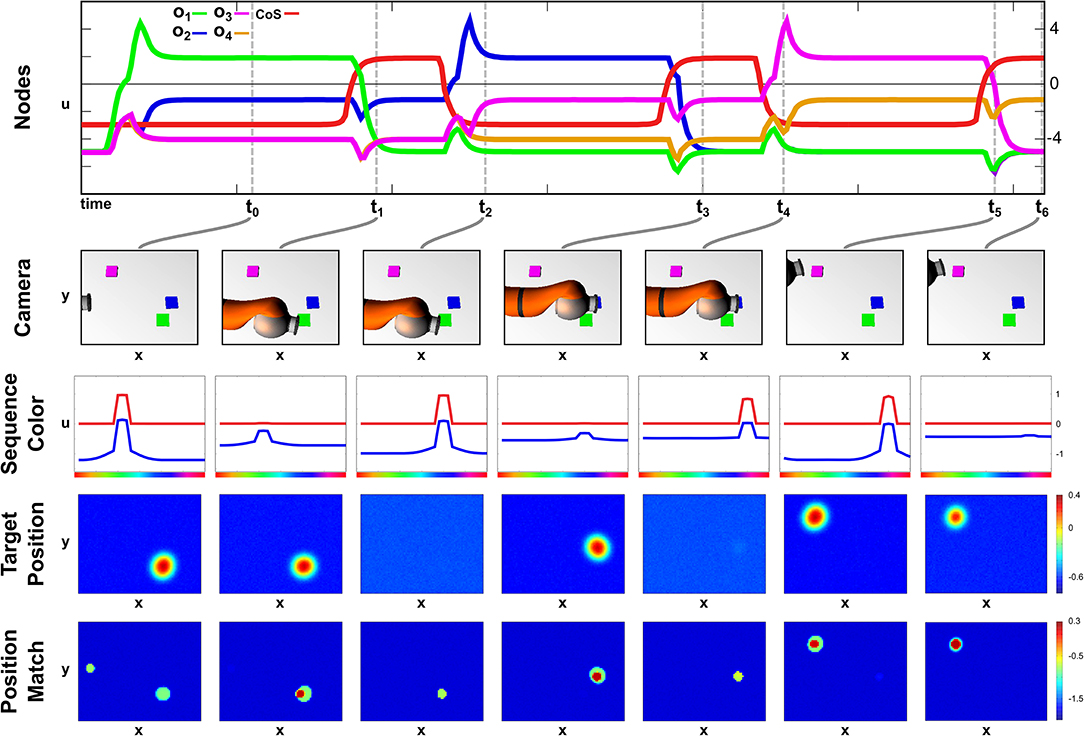
Figure 6. Time course of recalling a three element sequence through pointing at colored objects.
At point t0, the Recall task node has been activated, which lead to the activation of the first ordinal node and the emergence of a peak in the Sequence Color field at the green location due to the learned connections, wo1,ucol. The color information converges with the content of the Working Memory field in the Memory Color Selection field to form a three dimensional peak specifying position and color. Positional information is projected to the Target Position field of the Arm Movement sub-network, where it is forwarded to the movement generating oscillator and the Position Match field, which compares the current end effector position (center/left) with the current target position (bottom/right).
Due to a successful arm movement both positions match at point t1, which is represented through a peak in the Position Match field that activates the Sequence CoS deactivating the current ordinal node. The CoS node itself falls below threshold activity as soon as the peak in the Position Match field destabilizes through a missing target representation that vanished through insufficient color input from the Sequence Color field.
The missing inhibition from the CoS causes an activation of the next ordinal node o2, which is associated with blue color. At t2 however the blue peak has emerged in the Sequence Color field, but the target position has not yet been extracted from working memory. The column of point t3 depicts the end of the movement, where the overlap of end effector and target cause a peak that triggers the Sequence CoS. In this particular configuration the match representation is only possible due to the self-sustaining working memory representation that shields the blue object representation from the occlusion through the arm.
The movement toward the purple object depicted from t4 until t6 follows an analog activation pattern in which the ordinal node causes the formation of a purple peak in the Sequence Color field, which causes an extraction of the target position, leading to movement that terminates due to an represented match of positions. The movement times of all three movements are roughly the same despite their differences in distance, which results from the movement oscillator that enforces the same movement timing for all movements. See Supplementary Video 4 for another sequence recall demonstration showing the activation development of selected fields in continuous time.
4.3.1. Recall With a Moving Object
The autonomy of all three parts of the field network makes the execution of the recall task robust against unforeseen changes in the scene. We demonstrate this in an exemplary recall episode, where one of the objects in the scene is moved while its color corresponds to the active sequence element. The episode is depicted in Figure 7, which shows activation snapshots analog to Figure 6. Additionally activation of the Intention and CoS node driving the two-layer oscillator are shown as well as snapshots of the Memory Color Selection field.
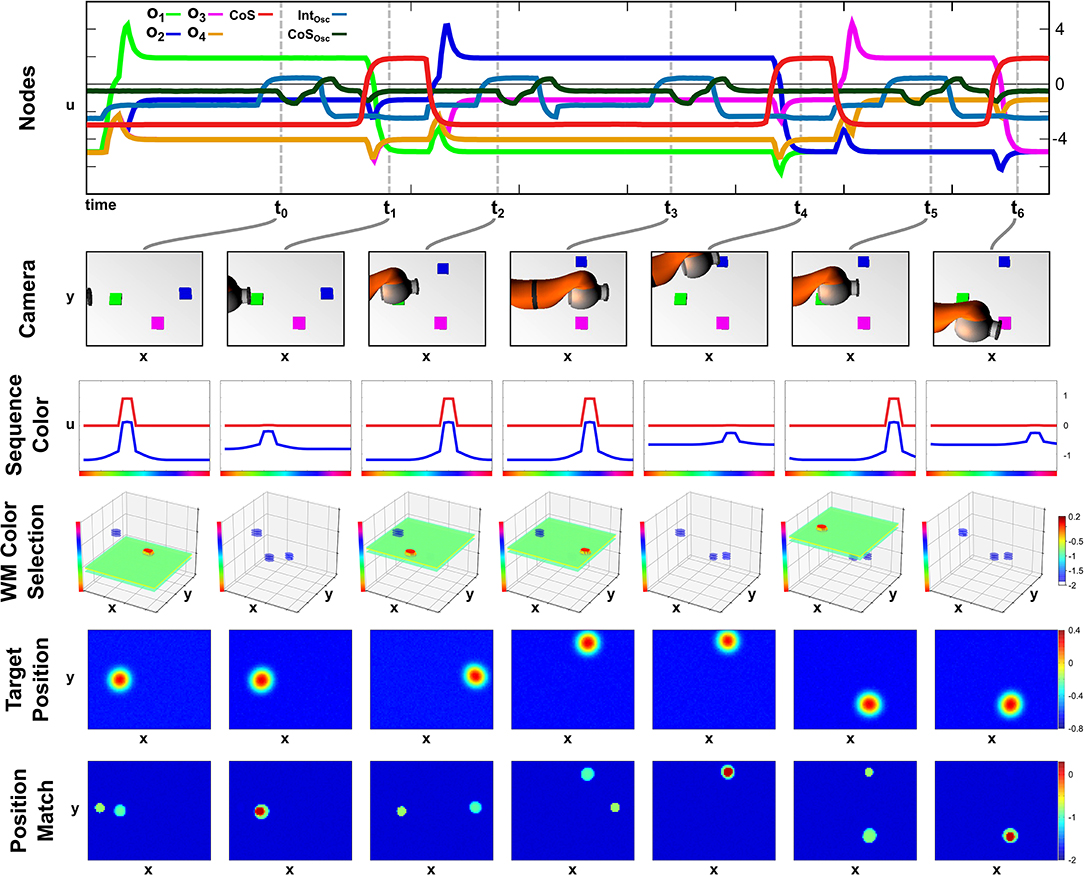
Figure 7. Online updating of the movement during sequence recall.
In this episode, build-up of the scene memory starts simultaneously with activation of the recall task, which causes a delay between the activation of the first ordinal node and the first movement as the green object, which is the first sequence element, is the second object committed to memory. This can be observed at t0 in the Memory Color Selection field, where the green object forms a peak as it overlaps with the green color slice specified by the sequence color, while the purple object is present as a sub-threshold activation blob, and the blue object is entirely absent. As the first movement is finished at t1 all three objects are present in working memory as sub-threshold activation blobs.
Thus at t2, the second movement starts closely after the activation of the second ordinal node with the blue object as the target on the right side of the camera image. While the arm is moving the object is moved to the center/top position of the image, which results in a non-match between arm and target at the end of the movement, which can be seen at t3. Here working memory has updated the position of the blue object, which leads to an extraction of a different target position that does not match with the current position of the end effector. Only at t4 after a second movement was generated, the blue object and the end effector match, which concludes the recall of the second element of the sequence.
The last movement toward the purple object is then conducted without any further perturbations and terminates after a single movement at t6.
4.3.2. Recall With a Missing Object
In this second recall episode demonstrating the robustness of the field network we start the recall in a scene that lacks the second object of the sequence. In Figure 8, activation snapshots of the same sub-set of fields used in the previous perturbation episode are shown.
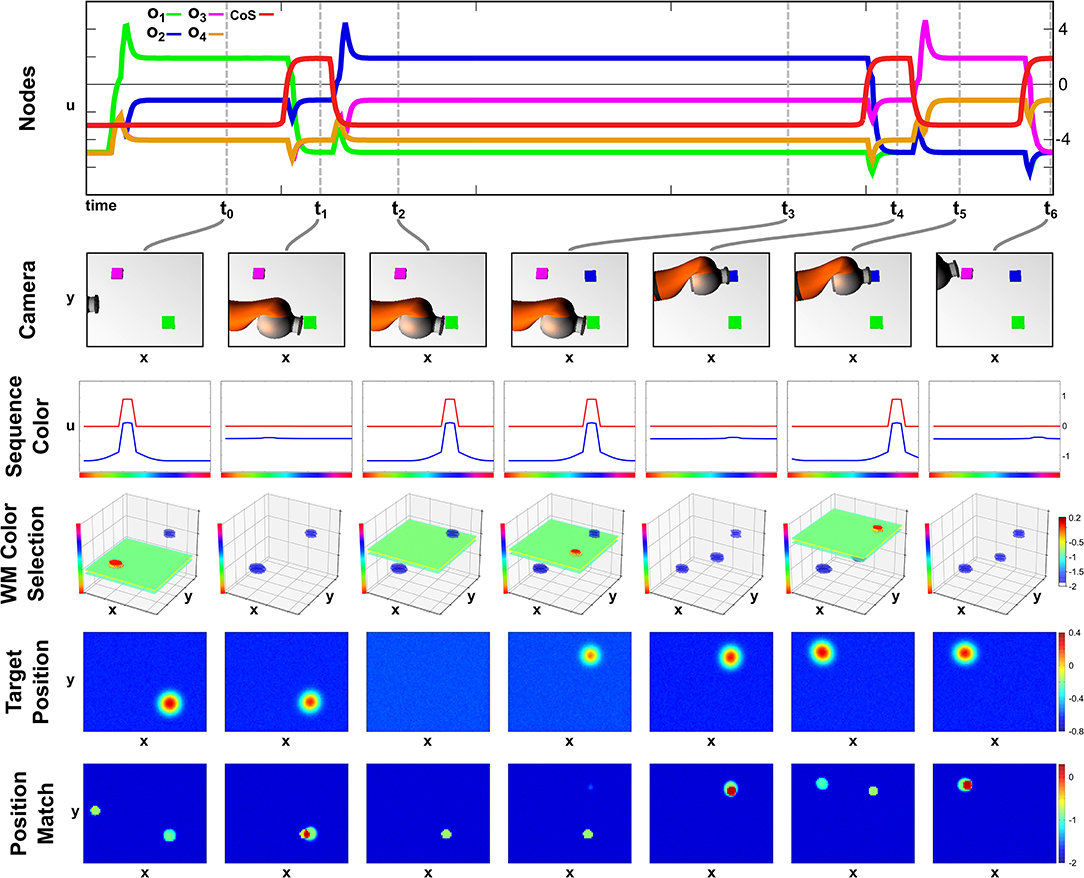
Figure 8. Recall with a delayed second object.
At points t0 and t1, the network's activation develops analog to the previous two recall examples with a color slice used to extract the target position and the position match to determine the successful termination of the movement. However as the second ordinal node activates at t2 no blue object is present in the scene, thus no sub-threshold activation blob overlaps with the blue color slice in the Memory Color Selection field and no peak forms.
At point t3, the blue object is added to the scene, which is committed to memory and afterwards extracted as a valid target position. The movement than concludes at t4 with the arm occluding the purple object, which is kept in working memory due to the self-sustaining kernel. The working memory information is then used in t5, when the third ordinal node specifies purple as the next sequence color. Thus the sequence ends at t6 with no further perturbations.
5. Discussion
We have presented a network of dynamic neural fields that integrates the complete pathway from the sensor surface (vision) to representations of higher cognition (serial order) and to the motor system (pointing). The network architecture enables a robotic agent to autonomously learn a sequence of colors from demonstration and then to act according to the defined serial order on a scene. Both during learning and while acting out the sequence, the transitions between elements of the sequence are detected without the need for an external control signal (The switch between learning and recall mode is not autonomous, however, reflecting a similar need for task instructions when a human operator performs such a task).
In each of the three sub-networks responsible for scene representation, the representation of serial order, and movement generation, sequential transitions between neural activation states are brought about through the mechanism of the condition of satisfaction. Thus, visual attention shifts only once a currently attended item has been committed to working memory. A transition to the next element in the serial order occurs only once the robot has successfully acted on the current element. And an arm movement terminates only once the desired movement target has been reached. The mechanism of the condition of satisfaction thus reconciles the capacity to autonomously act according to learned or structurally determined plans with the capacity to be responsive to sensory or internal information about the achievement of goals.
5.1. What the Scenario Stands for
The scenario was simple, but meant to demonstrate the fundamental components of any neurally grounded autonomous robot.
(1) A representation of the visual surround is the basis for any intelligent action directed at the world. It is also the basis for sharing an environment with a human user. We humans are particularly tuned to building scene representations which form the basis of much of our visual cognition (Henderson and Hollingworth, 1999). Scene representations need to include scene memory to deal with occlusions (e.g., by the agent's own body or body parts of a collaborating human user) and with a limited viewing range. Scene representations must also be open to updating, however, when the scene changes over time. Attentional selection is the key process that provides an interface between the scene and any action plan. So, while we stripped the system down to the bare essentials, the core processes of scene representation were covered.
(2) Directing action to objects in the world requires transforming attentionally selected scene information into a coordinate frame anchored in the initial position of the actuator. In that representation, motor plans can be framed as movement parameters (Erlhagen and Schöner, 2002) that characterize the movement as a whole. Movements must be initiatiated and terminated, and time courses of motor commands must be generated that take the effector to the target. In dynamic environments, such as when a human user interferes with objects, the movement parameters must be open to online updating. If movements still fail to reach the target, correction movements must be generated. Even in our extremely limited implementation, these core processes of movement generation were covered. Control issues, which are not trivial in human movement but are well-understood in robotics, were neglected.
(3) The cognition of goal-directed action was simplified to serial order. Serial order is a cognitive construct in that it abstracts from the contents (what is serially ordered) and from time (when is each item addressed). Based on these abstractions, a broad set of actions can be conceived of as serially ordered processing steps. For instance, assembling a piece of IKEA furniture could be described this way. Unlike many classical, disembodied cognitive tasks, real action sequences require the capacity to deal with variable and perhaps unpredictable amounts of time needed to achieve each processing step. Learning the—a priori—arbitrary contents of a serially ordered sequence makes this scenario quite powerful. It goes beyond, for instance, a mere capacity to imitate or emulate behavior, which would lead to the reproduction of the same movements or effects without generalization to new conditions. It also goes beyond the generation of sequences of behaviors that would be triggered by environmental conditions according to a fixed organizational scheme encoded in a behavior-based robotic architecture (Elements of fixed sequencing are contained in the present system such as when attentional selection always precedes pointing).
5.2. Scaling Beyond the Simplified Scenario
(1) The scene representation system was built on a single, trivial color feature. We have previously explored and demonstrated elsewhere how a neural dynamic system of the same kind can deal with multiple feature dimensions (e.g., Chapter 8 of Schöner et al., 2016 or Grieben et al., 2018). This entails a feature binding problem that can be solved through a shared spatial dimension across multiple low-dimensional feature/space fields. Binding occurs by attending selectively to a spatial location and transmitting feature information separately in the different feature/space fields. In this account, such binding through space is ultimately the reason why objects need to be attended sequentially. Active gaze shifts would be another extension, the basis for which has been outline in previous work (e.g., Wilimzig et al., 2006). A further generalization would be to extend the scene representation into a semantic map in which objects are also classified and their class labels are stored. There is, of course, a plethora of neurally based feedforward classification system (reviewed in the section 1), and their outputs could be treated as such labels. Such modules may need improvement in terms of object segmentation and pose estimation (as discussed in Lomp et al., 2017), which clearly requires attentional processes. Visual search for such labels is a challenge that may need some research attention.
(2) The motor domain is, of course, a much richer domain than we were able to demonstrate. There are myriad problems to be solved such as dealing with many degrees of freedom, dealing with obstacles, dealing with compliant actuators, controlling impedance, grasp planning and control, manipulating objects dynamically, and many more. For many of these, technical solutions are available or are being actively researched. Neurally grounded process accounts have not been developed as strongly as one would hope, however. The fundamental difference between human motor control and the control of current robot arms limits the extent to which approaches inspired by modern control theory and the theory of optimal control carry over to neurally inspired robotics (but see Driess et al., 2018).
(3) We have looked at the learning and recall of a single sequence. To learn multiple sequences, additional processing substrate must be introduced that represents activation of such learned sequences as well as selection of a neural population when a new sequence is added to the sequence memory. In principle, an approach inspired by Adaptive Resonance Theory (Carpenter and Grossberg, 2017) may achieve that. We have outlined such an approach in related work on contingency learning (Tekülve and Schöner, 2019), but important questions remain open such as how to align sequences of different lengths. The position encoding of serial order in the ordinal nodes makes it possible, however, to represent sequences that entail the same elements in different serial positions (Sandamirskaya and Schöner, 2010).
The sliver of cognition we have captured may be part of communication, showing each other what to do. If perception was better (e.g., recognizing events and perceiving relationships between actuators and objects), and if action was richer (e.g., the ability to use tools and manipulate objects), then the modeled interface would already make the robot quite useful. It would enable a robot to learn the solution of problems from a human user, as long as the perception system extracts the conceptual structure of the demonstrated action. A big extension would be the capacity of the system to solve problems by itself, devising the sequences of actions required to achieve a goal. This would require neural processes in new domains such as exploration, outcome representations, perhaps value systems. There is a growing literature on such models (Mnih et al., 2015), but their import for robotic learning is an open research problem.
5.3. Related Work
A number of groups have addressed object-directed action and the requisite perception in a similar neural-dynamic framework (Fard et al., 2015; Strauss et al., 2015; Tan et al., 2016). Serial order and the specific neural mechanism for sequencing neural activation patterns were not yet part of these efforts, which otherwise overlap with ours. A number of neural dynamic models of serial order or sequencing have been proposed (e.g., Deco and Rolls, 2005), but not been brought into robotic problems. One reason may be the lack of a control structure comparable to our condition of satisfaction, so that the sequences unfold in neural dynamics at a given rhythm that is not synchronized with perceptual events. Such systems would not remain tied to the actual performance of a sequence in the world.
Related attempts to model in neural terms the entire chain from perception to action have been made for robotic vehicles. For instance, Alexander and Sporns (2002) enabled a vehicle to learn from reward a task directed at objects that a robot vehicle was able to pick up. pick up. Gurney et al. (2004) realized a neurally inspired system the organized the organism (This paper is useful also for its careful discussion of different levels of descriptions for neurally inspired approaches to robotics). Both systems are conceptually in the fold of behavior-based robotics, in that the sequences of actions emerge from a neural architecture, modulated by adaptation. To our knowledge, systems of that kind have not yet been shown to be able to form serial order memories and acquire scene representations.
A different style of neural robotic model for cognition is SPAUN (Eliasmith et al., 2012). This is an approach based on the Neural Engineering Framework (Eliasmith, 2005), which is able to implement any neural dynamic model in a spiking neural network. Thus, models based on DFT may, in principle, be implemented within this framework. On the other hand, SPAUN has also been turned to approaches to cognition that may not be compatible with the principles of DFT, in particular, the Vector Symbolic Architecture (VSA) framework that goes back to Smolensky, Kanerva, Plate, and Gayler (see Levy and Gayler, 2008 for review). In VSA, concepts are mapped onto high-dimensional vectors, that enable processing these concepts in the manner of symbol manipulation. If this approach is entirely free of non-neural algorithmic steps is not clear to us.
6. Conclusion
We have shown, in a minimal scenario, how sequences of attentional shifts, of movements, and of serially ordered actions can be autonomously generated in a neural dynamic framework that is free of any non-neural algorithmic control. The continuous or intermittent coupling to sensory and motor systems is made possible by creating neural attractor states. Inducing instabilities in a controlled manner enables the system to make sequential transitions between such states. As a result, the neural dynamic robot demonstrates a minimal form of cognition, learning and acting out serially ordered actions. Much work remains to be done to scale such systems to the real world.
Data Availability Statement
All relevant data to reproduce the presented dynamic field network with our open-source software cedar (cedar.ini.rub.de) is contained within the manuscript. For technical questions regarding the software please contact the authors.
Author Contributions
JT and AF performed the work, and contributed to the writing of the manuscript. YS and GS conceived of the project, provided research supervision, and contributed to the writing of the manuscript.
Funding
This work was funded by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) - 402791869 (SCHO 336/12-1) within the SPP The Active Self (SPP 2134) and the SNSF grant PZOOP2_168183_1 Ambizione.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We acknowledge support by the DFG Open Access Publication Funds of the Ruhr-Universität Bochum.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnbot.2019.00095/full#supplementary-material
Footnotes
1. ^Spatial information is taken from the Saliency Selection field rather than directly from the Scene Space Selection field to allow for possible coordinate transforms between image and memory space. In the present scenario, the camera is not moved so that there is no need for a coordinate transform.
References
Alexander, W. H., and Sporns, O. (2002). An embodied model of learning, plasticity, and reward. Adapt. Behav. 10:143. doi: 10.1177/1059712302010003001
Amari, S.-I. (1977). Dynamics of pattern formation in lateral-inhibition type neural fields. Biol. Cybernet. 27, 77–87. doi: 10.1007/BF00337259
Aoi, S., Manoonpong, P., Ambe, Y., Matsuno, F., and Wörgötter, F. (2017). Adaptive control strategies for interlimb coordination in legged robots: a review. Front. Neurorobot. 11:39. doi: 10.3389/fnbot.2017.00039
Ba, J. L., Mnih, V., and Kavukcuoglu (2015). “Multiple object recognition with visual attention,” in IRLC (San Diego, CA), 1–10.
Bicho, E., Mallet, P., and Schöner, G. (2000). Target representation on an autonomous vehicle with low-level sensors. Int. J. Robot. Res. 19, 424–447. doi: 10.1177/02783640022066950
Brooks, R. A. (1986). A robust layered control system for a mobile robot. IEEE J. Robot. Automat. 2, 12–23. doi: 10.1109/JRA.1986.1087032
Brooks, R. A. (1990). Elephants don't play chess. Robot. Autonom. Syst. 6, 3–15. doi: 10.1016/S0921-8890(05)80025-9
Brooks, R. A. (1991). New approches to robotics. Science 253, 1227–1232. doi: 10.1126/science.253.5025.1227
Bullock, D., and Grossberg, S. (1988). Neural dynamics of planned arm movements: emergent invariants and speed-accuracy properties during trajectory formation. Psychol. Rev. 95, 49–90. doi: 10.1037/0033-295X.95.1.49
Buonomano, D. V., and Laje, R. (2010). Population clocks: motor timing with neural dynamics. Trends Cogn. Sci. 14, 520–527. doi: 10.1016/j.tics.2010.09.002
Carpenter G. A., and Grossberg S. (2017). “Adaptive resonance theory,” in Encyclopedia of Machine Learning and Data Mining, eds C. Sammut and G. I. Webb (Boston, MA: Springer).
Coombes, S., beim Graben, P., Potthast, R., and Wright, J., (eds.). (2014). Neural Fields: Theory and Applications. New York, NY: Springer Verlag.
Deco, G., and Rolls, E. T. (2005). Sequential memory: a putative neural and synaptic dynamical mechanism. J. Cogn. Neurosci. 17, 294–307. doi: 10.1162/0898929053124875
Driess, D., Zimmermann, H., Wolfen, S., Suissa, D., Haeufle, D., Hennes, D., et al. (2018). “Learning to control redundant musculoskeletal systems with neural networks and SQP: exploiting muscle properties,” in ICRA (Brisbane), 6461–6468.
Eliasmith, C. (2005). A unified approach to building and controlling spiking attractor networks. Neural Comput. 17, 1276–1314. doi: 10.1162/0899766053630332
Eliasmith, C., Stewart, T. C., Choo, X., Bekolay, T., DeWolf, T., Tang, C., et al. (2012). A large-scale model of the functioning brain. Science 338, 1202–1205. doi: 10.1126/science.1225266
Engels, C., and Schöner, G. (1995). Dynamic fields endow behavior-based robots with representations. Robot. Auton. Syst. 14, 55–77. doi: 10.1016/0921-8890(94)00020-3
Erlhagen, W., and Schöner, G. (2002). Dynamic field theory of movement preparation. Psychol. Rev. 109, 545–572. doi: 10.1037/0033-295X.109.3.545
Fard, F. S., Hollensen, P., Heinke, D., and Trappenberg, T. P. (2015). Modeling human target reaching with an adaptive observer implemented with dynamic neural fields. Neural Netw. 72, 13–30. doi: 10.1016/j.neunet.2015.10.003
Full, R. J., and Koditschek, D. E. (1999). Templates and anchors: neuromechanical hypotheses of legged locomotion on land. J. Exp. Biol. 2, 3–125.
Ghigliazza, R. M., Altendorfer, R., Holmes, P., and Koditschek, D. (2003). A simply stabilized running model. SIAM J. Appl. Dyn. Syst. 2, 187–218. doi: 10.1137/S1111111102408311
Grieben, R., Tekülve, J., Zibner, S. K. U., Schneegans, S., and Schöner, G. (2018). “Sequences of discrete attentional shifts emerge from a neural dynamic architecture for conjunctive visual search that operates in continuous time,” in CogSci 2018, eds C. Kalish, M. Rau, J. Zhu, and T. Rogers, Madison, WI: Cognitive Science Society, 427–432.
Gurney, K., Prescott, T. J., Wickens, J. R., and Redgrave, P. (2004). Computational models of the basal ganglia: from robots to membranes. Trends Neurosci. 27, 453–459. doi: 10.1016/j.tins.2004.06.003
Henderson, J. M., and Hollingworth, A. (1999). High-level scene perception. Annu. Rev. Psychol. 50, 243–271. doi: 10.1146/annurev.psych.50.1.243
Hersch, M., and Billard, A. G. (2008). Reaching with multi-referential dynamical systems. Auton. Robots 25, 71–83. doi: 10.1007/s10514-007-9070-7
Holmes, P., Full, R. J., Koditschek, D., and Guckenheimer, J. (2006). The dynamics of legged locomotion: models, analyses, and challenges. SIAM Rev. 48, 207–304. doi: 10.1137/S0036144504445133
Ijspeert, A. J. (2008). Central pattern generators for locomotion control in animals and robots: a review. Neural Netw. 21, 642–653. doi: 10.1016/j.neunet.2008.03.014
Ijspeert, A. J., Nakanishi, J., Hoffmann, H., Pastor, P., and Schaal, S. (2013). Dynamical movement primitives : learning attractor models for motor behaviors. Neural Comput. 25, 328–373. doi: 10.1162/NECO_a_00393
Kelleher, J. D, and Dobnik, S. (2017). “What is not where: the challenge of integrating spatial representations into deep learning architectures,” in CLASP Papers in Computational Linguistics Vol 1: Proceedings of the Conference on Logic and Machine Learning in Natural Language (LaML 2017), eds S. Dobnik and S. Lappin (Gothenburg), 41–52. Available online at: http://hdl.handle.net/2077/54911
Kim, S., Shukla, A., and Billard, A. (2014). Catching objects in flight. IEEE Trans. Robot. 30, 1049–1065. doi: 10.1109/TRO.2014.2316022
Knips, G., Zibner, S. K. U., Reimann, H., and Schöner, G. (2017). A neural dynamic architecture for reaching and grasping integrates perception and movement generation and enables on-line updating. Front. Neurorobot. 11:9. doi: 10.3389/fnbot.2017.00009
Kriegeskorte, N. (2015). Deep neural networks : a new framework for modeling biological vision and brain information processing. Annu. Rev. Vis. Sci. 1, 417–446. doi: 10.1146/annurev-vision-082114-035447
Lecun, Y., Bengio, Y., and Hinton, G. (2015). Deep learning. Nature 521, 436–444. doi: 10.1038/nature14539
Levy, S. D., and Gayler, R. (2008). “Vector symbolic architectures: a new building material for artificial general intelligence,” in Proceedings of the 2008 Conference on Artificial General Intelligence 2008: Proceedings of the First AGI Conference (Memphis, TN: IOS Press), 414–418.
Lomp, O., Faubel, C., and Schöner, G. (2017). A neural-dynamic architecture for concurrent estimation of object pose and identity. Front. Neurorobot. 11:23. doi: 10.3389/fnbot.2017.00023
Lomp, O., Richter, M., Zibner, S. K., and Schöner, G. (2016). Developing dynamic field theory architectures for embodied cognitive systems with cedar. Front. Neurorobot. 10:14. doi: 10.3389/fnbot.2016.00014
Mataric, M. J. (1998). Behavior-based robotics as a tool for synthesis of artificial behavior and analysis of natural behavior. Trends Cogn. Sci. 2, 82–87. doi: 10.1016/S1364-6613(98)01141-3
Michel, O. (2004). Cyberbotics Ltd. webots: professional mobile robot simulation. Int. J. Adv. Robot. Syst. 1:5. doi: 10.5772/5618
Mnih, V., Kavukcuoglu, K., Silver, D., Rusu, A. A., Veness, J., Bellemare, M. G., et al. (2015). Human-level control through deep reinforcement learning. Nature 518, 529–533. doi: 10.1038/nature14236
Proetzsch, M., Luksch, T., and Berns, K. (2010). Development of complex robotic systems using the behavior-based control architecture iB2C. Robot. Auton. Syst. 58, 46–67. doi: 10.1016/j.robot.2009.07.027
Ruf, A., and Horaud, R. (1999). Visual servoing of robot manipulators part I: projective kinematics. Int. J. Robot. Res. 18, 1101–1118. doi: 10.1177/02783649922067744
Sandamirskaya, Y., and Schöner, G. (2010). An embodied account of serial order: how instabilities drive sequence generation. Neural Netw. 23, 1164–1179. doi: 10.1016/j.neunet.2010.07.012
Schaal, S., Ijspeert, A., and Billard, A. (2003). Computational approaches to motor learning by imitation. Philos. Trans. R. Soc. Lond. B 358, 537–547. doi: 10.1098/rstb.2002.1258
Schmidhuber, J. (2015). Deep learning in neural networks : an overview. Neural Netw. 61, 85–117. doi: 10.1016/j.neunet.2014.09.003
Schöner, G., Dose, M., and Engels, C. (1995). Dynamics of behavior: {t}heory and applications for autonomous robot architectures. Robot. Auton. Syst. 16, 213–245. doi: 10.1016/0921-8890(95)00049-6
Schöner, G., and Kelso, J. A. S. (1988). Dynamic pattern generation in behavioral and neural systems. Science 239, 1513–1520. doi: 10.1126/science.3281253
Schöner, G., Spencer, J. P., and DFT Research Group, T. (2016). Dynamic Thinking: A Primer on Dynamic Field Theory. Oxford University Press.
Schöner, G., Tekülve, J., and Zibner, S. (2019). “Reaching for objects: a neural process account in a developmental perspective,” in Reach-to-Grasp Behavior, eds D. Corbetta and M. Santello (New York, NY: Routledge).
Serre, T., Wolf, L., Bileschi, S., Riesenhuber, M., and Poggio, T. (2007). Robust object recognition with cortex-like mechanisms. IEEE Trans. Pattern Anal. Mach. Intell. 29, 411–426. doi: 10.1109/TPAMI.2007.56
Strauss, S., Woodgate, P. J., Sami, S. A., and Heinke, D. (2015). Choice reaching with a LEGO arm robot (CoRLEGO): the motor system guides visual attention to movement-relevant information. Neural Netw. 72, 3–12. doi: 10.1016/j.neunet.2015.10.005
Tan, C. K., Plöger, P. G., and Trappenberg, T. P. (2016). “A neural field approach to obstacle avoidance,” in KI 2016: Advances in Artificial Intelligence; Lecture Notes in Computer Science, Vol. 9904, eds G. Friedrich, M. Helmert, and F. Wotawa (Springer International Publishing), 69–87.
Tekülve, J., and Schöner, G. (2019). “Autonomously learning beliefs is facilitated by a neural dynamic network driving an intentional agent,” in International Conference on Development and Learning and Epigenetic Robotics (Oslo).
Wilimzig, C., Schneider, S., and Schöner, G. (2006). The time course of saccadic decision making : dynamic field theory. Neural Netw. 19, 1059–1074. doi: 10.1016/j.neunet.2006.03.003
Wilson, H. R., and Cowan, J. D. (1973). A mathematical theory of the functional dynamics of cortical and thalamic nervous tissue. Kybernetik 13, 55–80. doi: 10.1007/BF00288786
Zibner, S. K. U., Faubel, C., Iossifidis, I., and Schöner, G. (2011). Dynamic neural fields as building blocks for a cortex-inspired architecture of robotic scene representation. IEEE Trans. Auton. Mental Dev. 3, 74–91. doi: 10.1109/TAMD.2011.2109714
Appendix
Network Parameters
The following tables list the field parameters of the different sub-networks. Lateral kernels are constructed according to the following equation: . The learning rate used in Equation (9) is η = 0.05.
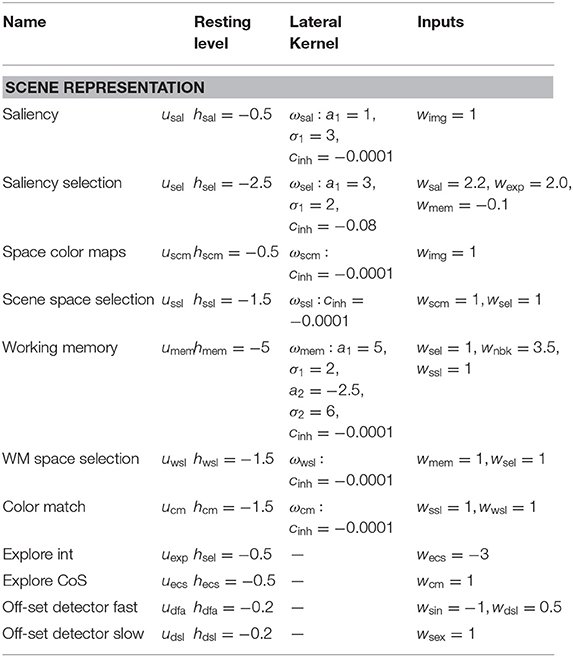
Table A1. Parameter values of the scene representation sub-network.
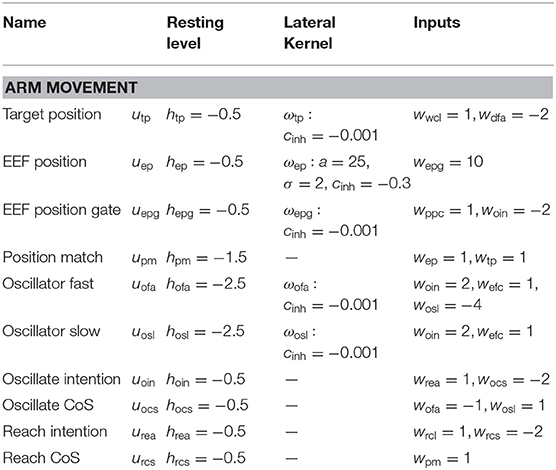
Table A2. Parameter values of the arm movement sub-network.
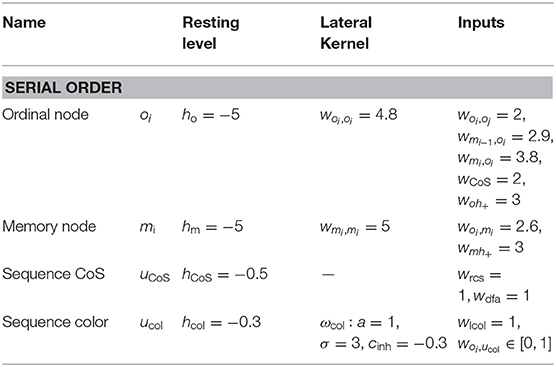
Table A3. Parameter values of the serial order sub-network.
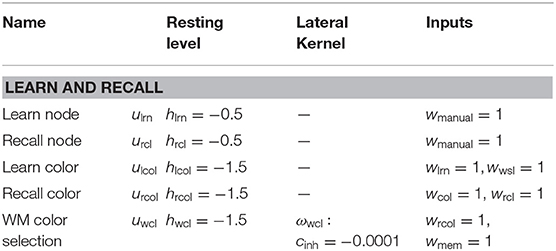
Table A4. Parameter values of task related fields of the dynamic field network.
Keywords: neural dynamic modeling, autonomous robot, sequence generation, scene perception, reaching movement
Citation: Tekülve J, Fois A, Sandamirskaya Y and Schöner G (2019) Autonomous Sequence Generation for a Neural Dynamic Robot: Scene Perception, Serial Order, and Object-Oriented Movement. Front. Neurorobot. 13:95. doi: 10.3389/fnbot.2019.00095
Received: 16 April 2019; Accepted: 28 October 2019;
Published: 15 November 2019.
Edited by:
Florian Röhrbein, Technical University of Munich, GermanyReviewed by:
Nicolas Cuperlier, Université de Cergy-Pontoise, FranceZhenshan Bing, Technical University of Munich, Germany
Dietmar Heinke, University of Birmingham, United Kingdom
Copyright © 2019 Tekülve, Fois, Sandamirskaya and Schöner. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jan Tekülve, amFuLnRla3VlbHZlQGluaS5ydWIuZGU=