 Patrícia Gonçalves Barbalho1
Patrícia Gonçalves Barbalho1 Benilton de Sá Carvalho2
Benilton de Sá Carvalho2 Iscia Lopes-Cendes1
Iscia Lopes-Cendes1 Claudia Vianna Maurer-Morelli1*
Claudia Vianna Maurer-Morelli1*
- 1Department of Medical Genetics, School of Medical Sciences, University of Campinas, Sao Paulo, Brazil
- 2Department of Statistics, Institute of Mathematics, Statistics and Scientific Computing, University of Campinas, Sao Paulo, Brazil
Cyclooxygenases (COX)-1 and -2 are isoenzymes that catalyze the conversion of arachidonic acid into prostaglandins (PGs). COX-2 and PGs are rapidly increased following seizures and are known to play important roles in the neuroinflammatory process. COX-2 isoform has been predominantly explored as the most suitable target for pharmacological intervention in epilepsy studies, while COX-1 remains poorly investigated. In the present study, we evaluated the effects of selective COX-1 inhibitor or selective COX-2 inhibitor on seizure suppression in the zebrafish pentylenetetrazole (PTZ)-seizure model. Zebrafish larvae were incubated in 5 μM of SC-236 for 24 h or 2.8 μM of SC-560 for 30 min, followed by exposure to 15 mM PTZ for 60 min. Real-time quantitative PCR analysis was carried out to investigate transcription levels of cox1 (ptgs1), as well as to determine cfos levels, used as a marker for neuronal activity. Effects of selective COX-2 or COX-1 inhibitors on locomotor activity response (velocity and distance moved) during PTZ exposure were evaluated using the Danio Vision video-tracking system. Our results showed an inducible expression of the cox1 gene after 60 min of PTZ exposure. Cox1 mRNA levels were upregulated compared with the control group. We found that COX-2 inhibition treatment had no effect on zebrafish PTZ-induced seizures. On the other hand, COX-1 inhibition significantly attenuated PTZ-induced increase of locomotor activity and reduced the c-fos mRNA expression. These findings suggest that COX-1 inhibition rather than COX-2 has positive effects on seizure suppression in the zebrafish PTZ-seizure model.
Introduction
Seizures are conceptually defined as abnormal and hypersynchronous neuronal activity that is caused by an imbalance between excitatory and inhibitory neurotransmission (1). Thus, it is not surprising that the mechanisms of action of most antiepileptic drugs (AEDs) target neuronal mechanisms involving in the modulation of voltage-activated ion channels, inhibition of excitatory, or enhancement of inhibitory synaptic neurotransmission (2). However, approximately 30% of patients remain pharmacoresistant to conventional drug-therapy (3). Therefore, a better understanding of the molecular mechanisms involved in ictogenesis and epileptogenesis is crucial for the discovery of new drugs with anticonvulsant properties (4–6).
There is accumulated experimental evidence supporting the contribution of non-neuronal cells, astrocytes, and microglia, to the pathophysiology of epilepsy (7–12). It is known that astrocytes and microglia activation promotes the release of proinflammatory mediators, such as interleukin-1 beta (IL-1β), interleukin-6 (IL-6), tumor necrosis factor-alpha (TNF-α), cyclooxygenase (COX)-2, and prostaglandins (PGs) (13, 14). Experimental studies suggest that proinflammatory mediators can affect the physiological functions of glio-neuronal communications, thus contributing to neuronal hyperexcitability and seizure-related neuronal damage (15–17). Therefore, inflammatory mediators have been explored as alternative pharmacological targets for therapeutic intervention to treat epilepsy (11, 18, 19).
Cyclooxygenase-2 and PGs are rapidly increased following seizures and are known to play important roles in the neuroinflammatory process (19). Prostaglandin endoperoxide synthases (PTGS) or COX-1 and COX-2 are isoenzymes that catalyze the conversion of arachidonic acid into PGs (20). COX-1 and COX-2 have been traditionally classified into constitutive and inducible expression, respectively (20). Because COX-2 is considered the inducible expressed isoform responsible for propagating the inflammatory response, several studies have been predominantly exploring the COX-2 isoform as the most suitable target for pharmacological intervention in epilepsy studies (11, 21, 22). However, the role of COX-2 inhibition on epileptogenesis and/or seizure suppression remains controversial.
Dhir et al. (23, 24) reported that pretreatment with both, selective and non-selective COX-2 inhibitors in the pentylenetetrazole (PTZ)-kindling model in rats, decreased the behavioral and biochemical parameters used as kindling score. In contrast, Claycomb et al. (25) reported that the pretreatment with rofecoxib, a selective COX-2 inhibitor, did not attenuate kindling progress in the PTZ-kindled mice. Recently, Katyal et al. (26) showed that the selective COX-2 inhibitor, etoricoxib, presented an anticonvulsant effect in the PTZ-kindled rats. In addition, treatment with nimesulide, a COX-2 selective inhibitor, prior to electrical kindling, had antiepileptogenic effects in rodents (27, 28). Furthermore, it was reported that the treatment with selective COX-2 inhibitor (SC58236) administered during the latent period did not modify epileptogenesis or chronic seizure activity after electrically induced status epilepticus (SE) in rats (29). On the other hand, the treatment with the SC58236 administered prior to electrically induced or during the chronic period increased seizure frequency and mortality rate (30). Proconvulsant effects of COX-2 inhibitor administration have also been reported in kainic acid-induced seizure model (31, 32). Conflicting outcomes regarding selective COX-2 inhibition have been reported in the pilocarpine-induced SE model. It has been shown that the administration of celecoxib, a selective COX-2 inhibitor, after a pilocarpine-induced SE in rats had an antiepileptogenic effect (33). In contrast, Polascheck et al. (34) show that the selective COX-2 inhibitor, parecoxib, after a pilocarpine-induced SE in rats had no antiepileptogenic effect.
Regarding COX-1 studies in animal models of seizures/epilepsy, Tanaka and colleagues reported that the selective COX-1 inhibitor slowed the development of epilepsy in electrical amygdala kindling in mouse model (35). Moreover, NSAIDs as indomethacin and aspirin, which inhibit the activity of both COX-1 and COX-2, reduced seizures in the absence epilepsy model, in the zebrafish seizure model, and in the pilocarpine-induced SE model (35–37). Nonetheless, the pharmacological inhibition of COX-1 isoform in acute and chronic epilepsy models remains poorly investigated. Therefore, in the present study we evaluated the effects of selective COX-1 inhibitor or selective COX-2 inhibitor on seizure suppression by evaluating c-fos mRNA expression and locomotor activity response in the zebrafish seizure model by evaluating the c-fos mRNA expression and locomotor activity response.
Materials and Methods
Animals
Wild-type adult fish were housed in 30–50 l tanks (two animals per liter) filled with non-chlorinated water cleared with mechanical and chemical filtration. Adult fish were maintained at 26 ± 2°C and in a simulated photoperiod cycle of 10 h dark/14 h light. Adult fish were fed twice a day with commercial flake fish food (Tetramin, Tetra, Blacksburg, VA, USA) and once a day with artemia; larvae were fed with paramecium and artemia twice a day. Fertilized eggs were collected after natural spawning. Embryos and larvae were housed using Petri dishes filled with water in an incubator system at the same temperature and photoperiods that were used for maintaining the adults. All experimental protocols used in this study were reviewed and approved by the Ethical Committee for Animal Research of the University of Campinas (#3098-1 and #4081-1).
Chemicals
5-(4-Chlorophenyl)-1-(4-methoxyphenyl)-3-trifluoromethyl pyrazole (SC-560), 4-[5-(4-Chlorophenyl)-3-(trifluoromethyl)-1H-pyrazol-1-yl]-benzenesulfonamide (SC-236), and PTZ were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Pentylenetetrazole Treatment
Detailed experimental procedures have been described previously (37). Briefly, 7 days post fertilization (dpf) larvae were placed in a 24-well plate (one larva per well) containing 15 mM PTZ [seizure group (SG)] or PTZ-free water [control group (CG)] for 60 min. Following, animals were cryoanaesthetized and their heads were isolated, quickly frozen in liquid nitrogen, and stored at −80°C until further processing. A total of five samples (n = 5) were used for each CG and SG, and each sample was composed by pooling five larval heads.
Pharmacological Pretreatment
Zebrafish larvae were incubated in 5 μM of SC-236 containing DMSO >0.01% or 2.8 μM of SC-560 containing >0.01% DMSO in Petri dishes for 24 h and 30 min, respectively. Those concentrations were selected based on the study of Teraoka et al. (38). After the incubation, they were exposed to 15 mM PTZ for 60 min as described above.
Real-time Quantitative PCR
Real-time quantitative PCR (qPCR) analysis was carried out to investigate the transcript levels of cox1 (ptgs1) and cfos (fos), the last one used as a marker for neuronal activity. Total RNA extraction, reverse transcription, and qPCR were performed as previously reported (37). Briefly, total RNA was extracted from each group samples (control; seizure; SC-236; vehicle (DMSO)-treated controls; and SC-560 groups) using TRIzol® (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions, and its concentration and quality were determined with the Epoch™ spectrophotometer (BioTek, Winooski, VT, USA) and electrophoresis using agarose gels. cDNA was generated using the High Capacity first-strand synthesis system for RT-PCR (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. Relative mRNA quantification was performed using the ABI 7500 Real-Time PCR system (Applied Biosystems, Foster City, CA, USA) with LuminoCt® qPCR ReadyMix (Sigma-Aldrich, St. Louis, MO, USA) and TaqMan® Gene Expression Assay (Invitrogen, Carlsbad, CA, USA). Runs were carried out in triplicate using the housekeeping gene eef1a1l1 (Dr03432748_m1) to normalize the mRNA level of ptgs1 (Dr03087197_m1) and c-fos (fos) (Dr03100809_g1). Data were analyzed using the SDS 7500 software (Applied Biosystems) to estimate qPCR efficiency and quantify the relative gene expression. A total of five samples (n = 5) were used for each group and each sample comprising a pool of five larval heads.
Behavioral Seizure Locomotor Activity
At 7 dpf, zebrafish larvae were transferred to 96-well plate in a semi-randomized distribution (in order to have larvae distributed in lines and columns along the plate) using a micropipette in a volume of 50 μl and acclimated for 30 min to minimize any interference in the test. Following acclimation, 50 μl of aquarium water or 50 μl of a 30 mM PTZ solution (dissolved in aquarium water) was added to obtain final volume and concentration of 15 mM. After that, the locomotor activity for each larva was recorded during 30 min using an automated computerized video-tracking system DanioVision (Noldus, Wageningen, The Netherlands). A total of seven zebrafish larvae (n = 7) were used to compose each experimental group: (i) control, (ii) seizure, (iii) vehicle (DMSO)-treated controls, (iv) SC-236, or (v) SC-560 PTZ-treated groups. The total distance moved and the speed (velocity) for each larva from each group was quantified using the EthoVision XT locomotion tracking software (Noldus, Wageningen, The Netherlands).
Statistical Analysis
Data are presented as mean values ± SEM. Statistical analysis was performed using the GraphPad Prism version 5.0 (GraphPad Software, San Diego, CA, USA). In all the analyses, the significance level was set at p ≤ 0.05. Statistical comparisons between two groups were performed using the Mann–Whitney test. Statistical comparisons between more than three groups were assessed by one-way analysis of variance (ANOVA) followed by Bonferroni’s post hoc test.
Results
Evaluation of cox1 mRNA Expression after PTZ-Induced Seizures
We previously reported that cox2b mRNA expression was upregulated after PTZ-induced seizures in 7 dpf zebrafish larva (37). To determine whether the cox1 transcription is affected following PTZ-induced seizure, we used real-time qPCR to measure the relative quantification of the cox1 gene. Our results showed an inducible expression of cox1 after 60 min of PTZ exposure. Cox1 mRNA levels were upregulated compared with the control group (p = 0.004; Figure 1). The mean ± SEM of control and PTZ groups were 1.1 ± 0.07 and 1.5 ± 0.07, respectively.
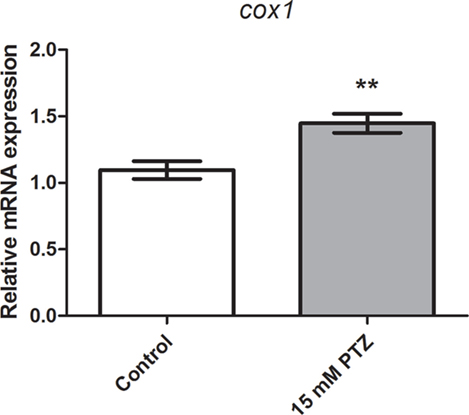
Figure 1. Evaluation of cox1 mRNA expression after PTZ-induced seizures. Relative quantification of cyclooxygenase-1 transcript levels following 60 min of pentylenetetrazole (PTZ)-induced seizures in the zebrafish brain at 7 days post fertilization. Seizure group was exposed to 15 mM PTZ, and the control group was handled identically, but in PTZ-free water (n = 5 per group). Data are presented as mean ± SEM. Statistical differences between control and seizure groups were tested using the Mann–Whitney test. Two asterisks (**) indicated that p ≤ 0.01.
Effects of Selective COX-1 or COX-2 Inhibitors on c-fos mRNA Expression
We previously showed that the treatment with indomethacin, a non-selective COX-1 and COX-2 inhibitor, prior to PTZ-induced seizures suppresses the c-fos mRNA upregulation (37). Here, we sought to address the contribution of either a COX-1 or COX-2 inhibition on c-fos mRNA expression. Pretreatment with the selective COX-1 inhibitor, SC-560 (2.8 μM), significantly decreased the c-fos mRNA expression compared to PTZ group (Figure 2). The mean ± SEM obtained were (i) control group: 0.82 ± 0.08; (ii) seizure group (PTZ): 57.4 ± 1.77; (iii) vehicle (DMSO)-treated control (>0.01%) group: 0.57 ± 0.09; and (iv) SC-560 group: 27.72 ± 1.9 (Figure 2). However, our results showed that the selective COX-2 inhibitor, SC-236, treatment had no effect on c-fos mRNA expression (Figure 3). The mRNA levels of SC-236 group were similar with those found in the PTZ group (Figure 3). The mean ± SEM obtained were (i) control group: 0.82 ± 0.08; (ii) seizure group (PTZ): 57.4 ± 1.77; (iii) vehicle (DMSO)-treated control (>0.01%) group: 0.57 ± 0.05; and (iv) SC-236 group: 59.16 ± 2.9.
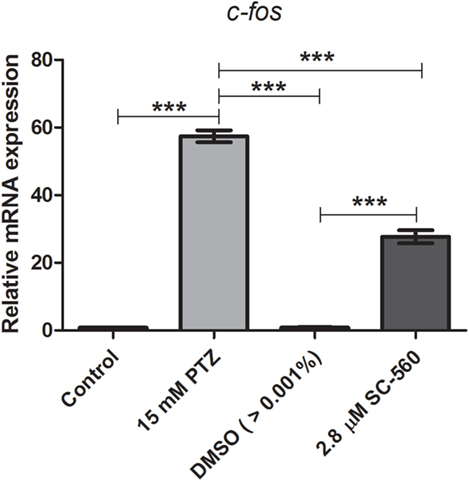
Figure 2. Effect of selective COX-1 inhibitor (SC-560) on c-fos mRNA expression. Seizure group (SG) was composed of animals exposed to 15 mM PTZ for 60 min. The SC-560 group (2.8 μM) was composed of animals that received SC-560 treatment for 30 min prior to PTZ exposure. Animals of the control group (CG) were handled identically but included exposure to water [no PTZ, DMSO (>0.01%), or SC-560 treatments; n = 5 per group]. Data are presented as mean ± SEM. One-way ANOVA followed by Bonferroni’s post hoc test was performed to determine statically significant differences between groups. Three asterisks (***) indicated that that p ≤ 0.001.
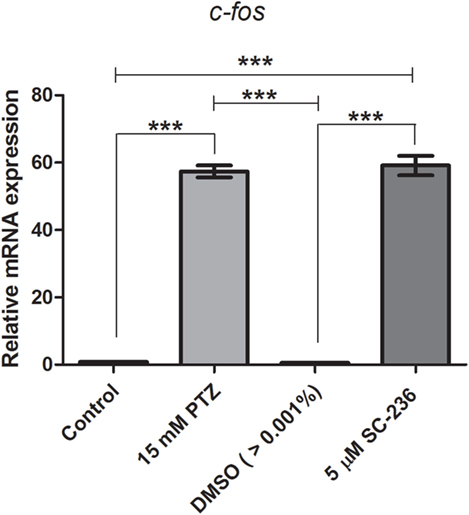
Figure 3. Effect of selective COX-2 inhibitor (SC-236) on c-fos mRNA expression. Relative quantification of c-fos transcript levels immediately after pentylenetetrazole (PTZ)-induced seizures in zebrafish brain at 7 days post fertilization. Seizure group (SG) was composed of animals exposed to 15 mM PTZ for 60 min. The SC-236 group (5 μM) was composed of animals that received SC-236 treatment prior to PTZ exposure. Animals in the control group (CG) were handled identically but included exposure to water [no PTZ, DMSO (>0.01%), or SC-560 treatments; n = 5 per group]. Data are presented as mean ± SEM. One-way ANOVA followed by Bonferroni’s post hoc test was performed to determine statically significant differences between groups. Three asterisks (***) indicated that that p ≤ 0.001.
Effects of Selective COX-1 or COX-2 Inhibitors on Locomotor Activity Response during PTZ Exposure
Our initial pilot experiment revealed that after 1 h of PTZ exposure there was no difference in velocity and distance moved between PTZ and the control groups. As pointed out by Afrikanova et al. (39), the decline of the locomotor activity observed in longer periods of PTZ exposure might be due to the fact that the larvae begin to undergo seizures with more frequency and spending more time on seizure stage III (loss of posture), which, as consequence, promotes a decrease of the locomotor activity. Therefore, we choose to evaluate the locomotor activity during 30 min of PTZ exposure. Our results have shown that SC-560 treatment inhibits the PTZ-induced increase of locomotor activity (Figure 4). The velocity and distance moved of the SC-560 group were lower in comparison to the PTZ group (Figures 4A,B). For velocity, the mean ± SEM of the control, seizure (PTZ), vehicle (DMSO)-treated control (>0.01%), and SC-560 groups were, respectively, 1.4 ± 0.16, 2.96 ± 0.20, 1.35 ± 0.26, and 1.55 ± 0.20. For distance moved, the mean ± SEM of control, seizure (PTZ), vehicle (DMSO)-treated control (>0.01%), and SC-560 were, respectively, 2536 ± 297.7, 5311 ± 342.5, 2432 ± 460.6, and 2802 ± 360.7. On the other hand, SC-236 treatment did not reduce PTZ-induced increase of locomotor activity (Figures 5A,B). Velocity and distance moved of the SC-236 group were similar to those found to the PTZ group (Figures 5A,B). For velocity, the mean ± SEM of control, seizure (PTZ), vehicle (DMSO)-treated control (>0.01%), and SC-236 were, respectively, 1.3 ± 0.16, 1.26 ± 0.18, 2.22 ± 0.22, and 2.20 ± 0.14. For distance moved, the mean ± SEM of control, seizure (PTZ), vehicle (DMSO)-treated control (>0.01%), and SC-236 groups were, respectively, 2365 ± 285.6, 2258 ± 322.9, 3995 ± 403.0, and 3965 ± 257.0.
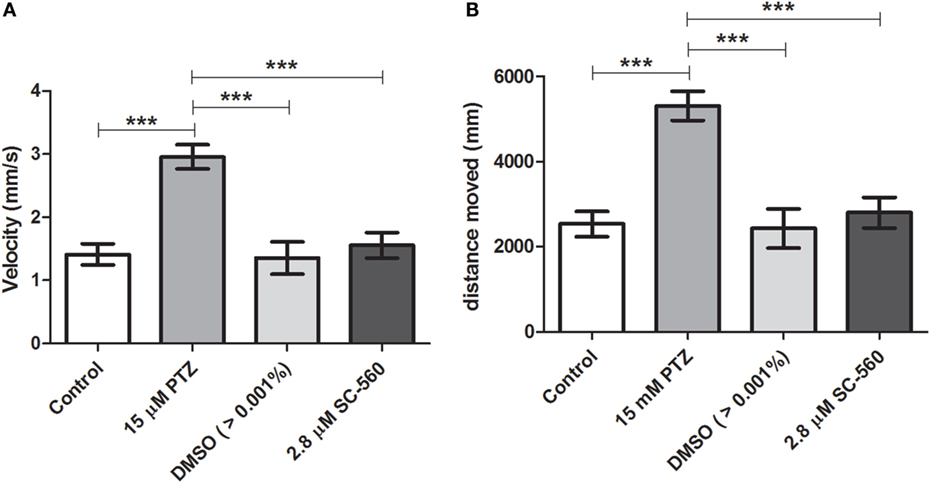
Figure 4. Effect of the selective COX-1 inhibitor on locomotor activity response during PTZ exposure. (A) Velocity and (B) distance moved. Animals were exposed to 2.8 μM of SC-560 for 30 min prior to pentylenetetrazole (15 mM) exposure. Animals in the control group (CG) were handled identically but included exposure to water [no PTZ, DMSO (>0.01%), or SC-560 treatments; n = 7 per group]. At 7 days post fertilization, larval zebrafish were transferred to 96-well plate and acclimated for 30 min to minimize any disturbance related to handling and transport from the Petri dishes to 96-well plate. Following acclimation, locomotor activity for each treatment group was recorded for 30 min using an automated video-tracking system Danio Vision (Noldus, Wageningen, The Netherlands). The distance moved and the velocity for each larva from each group was performed using EthoVision XT locomotion tracking software (Noldus, Wageningen, The Netherlands). Data are presented as mean ± SEM. One-way ANOVA followed by Bonferroni’s post hoc test was performed to determine statically significant differences between groups. Three asterisks (***) indicated that that p ≤ 0.001.
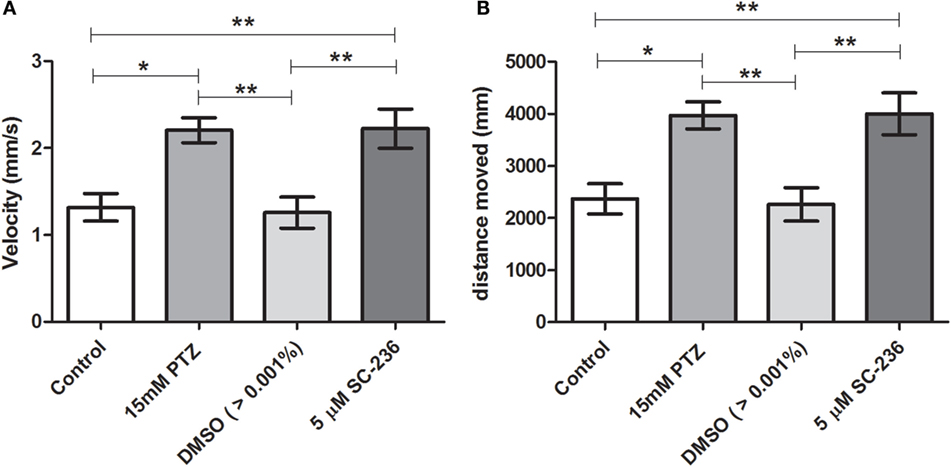
Figure 5. Effect of selective COX-2 inhibitor (SC-236) on locomotor activity response during PTZ exposure. (A) Velocity and (B) distance moved. Animals were exposed to 5 μM of SC-236 for 24 h prior to pentylenetetrazole (15 mM) exposure. Animals in the control group (CG) were handled identically but included exposure to water [no PTZ, DMSO (>0.01%), or SC-236 treatments; n = 7 per group]. At 7 days post fertilization, larval zebrafish were transferred to a 96-well plate and acclimated for 30 min to minimize any disturbance related to handling and transport from the Petri dishes to the plate. Following acclimation, locomotor activity for each treatment group was recorded for 30 min using an automated video-tracking system Danio Vision (Noldus, Wageningen, The Netherlands). The distance moved and the velocity for each larva from each group was performed using EthoVision XT locomotion tracking software (Noldus, Wageningen, The Netherlands). Data are presented as mean ± SEM. One-way ANOVA followed by Bonferroni’s post hoc test was performed to determine statically significant differences between groups. One asterisk (*) indicated that p ≤ 0.05; two asterisks (**) indicated that p ≤ 0.01.
Discussion
In the central nervous system (CNS), both COX-1 and COX-2 are constitutively expressed in neurons, astrocytes, and microglial cells, and both COXs lead to PG productions, which are important inflammatory mediators (20). Experimental evidence has shown that PGs are markedly increased following seizures and may contribute to epileptogenesis and reduction in the seizure threshold (11, 17–19). Traditionally, COX-1 and COX-2 isoforms have been considered constitutive and inducible expression, respectively (40–42). Therefore, most experimental epilepsy research reports have focused on the COX-2 isoform because of its role in the inflammatory response through PGs production (18, 19). Several studies were designed to interfere with the COX-2 enzyme without affecting the homeostatic function of COX-1, by using selective COX-2 inhibitors. However, both proconvulsant and anticonvulsant effects of COX-2 inhibition have been reported, and their role still remains controversial (11, 19, 20).
It has been reported that the therapeutic outcome of COX-2 inhibition on epileptogenesis and/or seizure suppression appears to depend on many factors including the type of inhibitor used (non-selective or selective) and the timing of the treatment administration (prior or after to seizure onset) (11, 36). Moreover, comparisons between these conflicting reports are difficult to be interpreted because they used different methods for seizure-induction, were evaluated in mature or immature brain, were assessed in different phase of epileptogenesis as well as were performed in different species (11, 36). Nonetheless, studies using non-selective COX inhibitors in the zebrafish seizure model or in the absence epilepsy rodent model as well as following pilocarpine-induced SE, showed anticonvulsant effects (35–37). Thus, these data indirectly suggest the involvement of the COX-1 isoform on seizure inhibition in these models.
Notably, experimental studies have reported neuroprotective effects of selective COX-1 inhibition and NSAIDs, with higher selectivity for COX-1, in models of neurodegenerative diseases such as Alzheimer disease and traumatic brain injury (41–49). It is noteworthy, however, that continuous NSAIDs use is associated with gastrointestinal side effects due to COX-1-derived PGs inhibition (42, 50). On the other hand, the continuous use of selective COX-2 inhibitors (coxibs) has been associated with increased risk of cardiovascular side effects (42, 50). Therefore, the benefit–risk evaluation of the administration of selective or non-selective COX inhibitors, as for many other drugs, should be considered.
Although several studies have suggested that COX-1 might have an important role in the neuroinflammation of neurodegenerative diseases (40–42), only few studies have investigated the association between COX-1 and epilepsy (35, 36). Notably, COX-1 expression has increased following kindling progression and in the SE induced by pilocarpine (35, 36). Moreover, treatment with selective COX-1 inhibitor reduced the epileptogenesis in the electrical kindling model (35). Based on this background, we sought to investigate the contribution of each COX isoforms on seizure suppression in the zebrafish PTZ-seizure model.
Although it is considered as an acute seizure model, the zebrafish is a powerful model in neuroscience research since it has many advantages when performing imaging studies, genetic manipulation, and modeling different disease mechanisms. In addition, it is a convenient model for drug screening, making it suitable for translational studies in epilepsy (51–53).
The zebrafish has homologs for both COX-1 and COX-2 human genes named cox1 (ptgs1) and cox2 (ptgs2), respectively (54). Furthermore, the zebrafish PTZ-seizure model has been widely used as an in vivo high-throughput screening method for antiepileptic drug discovery (55–57). Screening of new compounds with anticonvulsants’ properties is performed by evaluating the inhibition of the seizure-like behavior (i.e., by assessing the locomotor activity), as well as by the suppression of the increased neuronal activity, through the c-fos quantification in the brain and/or by the inhibition of epileptiform electrographic activity (39, 55, 58).
In this study, we showed that the cox1 mRNA expression was increased after PTZ-induced seizure (Figure 1). In addition, we found that the selective COX-1 inhibitor, SC-560, significantly decreased the c-fos mRNA expression (Figure 2) and inhibited the PTZ-induced increase of locomotor activity (Figure 4). Taking all together, these findings suggest that COX-1 inhibition has an important role on seizure suppression in the zebrafish seizure model, whereas the selective COX-2 inhibitor (SC-236) had no effect, producing any changes in the c-fos mRNA expression (Figure 3) and locomotor activity (Figure 5).
Our findings suggest that inhibition of the COX-1 isoform has an important role on seizure suppression in the zebrafish PTZ-seizure model. Together, with the accumulating knowledge arising from other animal models of epilepsy and given the emerging importance of the zebrafish as a model for epilepsy investigations and drug screening, we believe these findings may contribute with new avenues of investigations toward seizure suppression.
Author Contributions
PB performed the experiments, participated in the design of the study, analyzed the data, and wrote the paper. BC analyzed the statistical data. IL-C contributed with the laboratory support and revised the manuscript. CM-M conceived the study, participated in its design, coordination, and revised the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the support from FAPESP.
Funding
This work was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) #2014/15640-8 and Brazilian Institute of Neuroscience and Neurotechnology (BRAINN) CEPID-FAPESP (#2013/07559-3). PB received fellowship from FAPESP (#2013/19151-9).
References
1. Fisher RS, Acevedo C, Arzimanoglou A, Bogacz A, Cross JH, Elger CE, et al. ILAE official report: a practical clinical definition of epilepsy. Epilepsia (2014) 55(4):475–82. doi:10.1111/epi.12550
2. Lasoń W, Chlebicka M, Rejdak K. Research advances in basic mechanisms of seizures and antiepileptic drug action. Pharmacol Rep (2013) 65(4):787–801. doi:10.1016/S1734-1140(13)71060-0
3. Laxer KD, Trinka E, Hirsch LJ, Cendes F, Langfitt J, Delanty N, et al. The consequences of refractory epilepsy and its treatment. Epilepsy Behav (2014) 37:59–70. doi:10.1016/j.yebeh.2014.05.031
4. Kobow K, Auvin S, Jensen F, Löscher W, Mody I, Potschka H, et al. Finding a better drug for epilepsy: antiepileptogenesis targets. Epilepsia (2012) 53(11):1868–76. doi:10.1111/j.1528-1167.2012.03716.x
5. Yu N, Liu H, Di Q. Modulation of immunity and the inflammatory response: a new target for treating drug-resistant epilepsy. Curr Neuropharmacol (2013) 11(1):114–27. doi:10.2174/157015913804999540
6. Doeser A, Dickhof G, Reitze M, Uebachs M, Schaub C, Pires NM, et al. Targeting pharmacoresistant epilepsy and epileptogenesis with a dual-purpose antiepileptic drug. Brain (2015) 138(Pt 2):371–87. doi:10.1093/brain/awu339
7. Devinsky O, Vezzani A, Najjar S, De Lanerolle NC, Rogawski MA. Glia and epilepsy: excitability and inflammation. Trends Neurosci (2013) 36(3):174–84. doi:10.1016/j.tins.2012.11.008
8. Amor S, Peferoen LA, Vogel DY, Breur M, van der Valk P, Baker D, et al. Inflammation in neurodegenerative diseases – an update. Immunology (2014) 142(2):151–66. doi:10.1111/imm.12233
9. Legido A, Katsetos CD. Experimental studies in epilepsy: immunologic and inflammatory mechanisms. Semin Pediatr Neurol (2014) 21(3):197–206. doi:10.1016/j.spen.2014.10.001
10. Bedner P, Dupper A, Hüttmann K, Müller J, Herde MK, Dublin P, et al. Astrocyte uncoupling as a cause of human temporal lobe epilepsy. Brain (2015) 138(Pt 5):1208–22. doi:10.1093/brain/awv067
11. Dey A, Kang X, Qiu J, Du Y, Jiang J. Anti-inflammatory small molecules to treat seizures and epilepsy: from bench to bedside. Trends Pharmacol Sci (2016) 37(6):463–84. doi:10.1016/j.tips.2016.03.001
12. Fakhoury M. Role of immunity and inflammation in the pathophysiology of neurodegenerative diseases. Neurodegener Dis (2015) 15(2):63–9. doi:10.1159/000369933
13. Aronica E, Ravizza T, Zurolo E, Vezzani A. Astrocyte immune responses in epilepsy. Glia (2012) 60(8):1258–68. doi:10.1002/glia.22312
14. Vezzani A, Aronica E, Mazarati A, Pittman QJ. Epilepsy and brain inflammation. Exp Neurol (2013) 244:11–21. doi:10.1016/j.expneurol.2011.09.033
15. Robel S, Sontheimer H. Glia as drivers of abnormal neuronal activity. Nat Neurosci (2015) 19(1):28–33. doi:10.1038/nn.4184
16. Robel S, Buckingham SC, Boni JL, Campbell SL, Danbolt NC, Riedemann T, et al. Reactive astrogliosis causes the development of spontaneous seizures. J Neurosci (2015) 35(8):3330–45. doi:10.1523/JNEUROSCI.1574-14.2015
17. Vezzani A. Anti-inflammatory drugs in epilepsy: does it impact epileptogenesis? Expert Opin Drug Saf (2015) 14(4):583–92. doi:10.1517/14740338.2015.1010508
18. Marchi N, Granata T, Janigro D. Inflammatory pathways of seizure disorders. Trends Neurosci (2014) 37(2):55–65. doi:10.1016/j.tins.2013.11.002
19. Rojas A, Jiang J, Ganesh T, Yang MS, Lelutiu N, Gueorguieva P, et al. Cyclooxygenase-2 in epilepsy. Epilepsia (2014) 55(1):17–25. doi:10.1111/epi.12461
20. Yagami T, Koma H, Yamamoto Y. Pathophysiological roles of cyclooxygenases and prostaglandins in the central nervous system. Mol Neurobiol (2015) 53(7):4754–71. doi:10.1007/s12035-015-9355-3
21. Kulkarni SK, Dhir A. Cyclooxygenase in epilepsy: from perception to application. Drugs Today (Barc) (2009) 45(2):135–54. doi:10.1358/dot.2009.45.2.1322481
22. Citraro R, Leo A, Marra R, De Sarro G, Russo E. Antiepileptogenic effects of the selective COX-2 inhibitor etoricoxib, on the development of spontaneous absence seizures in WAG/Rij rats. Brain Res Bull (2015) 113:1–7. doi:10.1016/j.brainresbull.2015.02.004
23. Dhir A, Naidu PS, Kulkarni SK. Effect of naproxen a non-selective cyclooxygenase inhibitor on pentylenetetrazol-induced kindling in mice. Clin Exp Pharmacol Physiol (2005) 32(7):574–84. doi:10.1111/j.1440-1681.2005.04233.x
24. Dhir A, Naidu PS, Kulkarni SK. Neuroprotective effect of nimesulide, a preferential COX-2 inhibitor, against pentylenetetrazol (PTZ)-induced chemical kindling and associated biochemical parameters in mice. Seizure (2007) 16(8):691–7. doi:10.1016/j.seizure.2007.05.016
25. Claycomb RJ, Hewett SJ, Hewett JA. Prophylactic, prandial rofecoxib treatment lacks efficacy against acute PTZ-induced seizure generation and kindling acquisition. Epilepsia (2011) 52(2):273–83. doi:10.1111/j.1528-1167.2010.02889
26. Katyal J, Kumar H, Gupta YK. Anticonvulsant activity of the cyclooxygenase-2 (COX-2) inhibitor etoricoxib in pentylenetetrazole-kindled rats is associated with memory impairment. Epilepsy Behav (2015) 44:98–103. doi:10.1016/j.yebeh.2014.12.032
27. Tu B, Bazan NG. Hippocampal kindling epileptogenesis upregulates neuronal cyclooxygenase-2 expression in neocortex. Exp Neurol (2003) 179:167–75. doi:10.1016/S0014-4886(02)00019-5
28. Takemiya T, Suzuki K, Sugiura H, Yasuda S, Yamagata K, Kawakami Y, et al. Inducible brain COX-2 facilitates the recurrence of hippocampal seizures in mouse rapid kindling. Prostaglandins Other Lipid Mediat (2003) 71(3–4):205–16. doi:10.1016/S1098-8823(03)00040-6
29. Holtman L, Vliet EA, Schaik R, Queiroz CM, Aronica E, Gorter JA. Effects of SC58236, a selective COX-2 inhibitor, on epileptogenesis and spontaneous seizures in a rat model for temporal lobe epilepsy. Epilepsy Res (2009) 84(1):56–66. doi:10.1016/j.eplepsyres.2008.12.006
30. Holtman L, Vliet EA, Edelbroek PM, Aronica E, Gorter JA. COX-2 inhibition can lead to adverse effect in a rat model for temporal lobe epilepsy. Epilepsy Res (2010) 91(1):49–56. doi:10.1016/j.eplepsyres.2010.06.011
31. Baik EJ, Kim EJ, Lee SH, Moon C. Cyclooxygenase-2 selective inhibitors aggravate kainic acid induced seizure and neuronal cell death in the hippocampus. Brain Res (1999) 843(1–2):118–29. doi:10.1016/S0006-8993(99)01797-7
32. Kim HJ, Chung JI, Lee SH, Jung YS, Moon CH, Baik EJ. Involvement of endogenous prostaglandin F2alpha on kainic acid-induced seizure activity through FP receptor: the mechanism of proconvulsant effects of COX-2 inhibitors. Brain Res (2008) 1193:153–61. doi:10.1016/j.brainres.2007.12.017
33. Jung KH, Chu K, Lee ST, Kim J, Sinn DI, Kim JM, et al. Cyclooxygenase-2 inhibitor, celecoxib, inhibits the altered hippocampal neurogenesis with attenuation of spontaneous recurrent seizures following pilocarpine-induced status epilepticus. Neurobiol Dis (2006) 23:237–46. doi:10.1016/j.nbd.2006.02.016
34. Polascheck N, Bankstahl M, Loscher W. The COX-2 inhibitor parecoxib is neuroprotective but not antiepileptogenic in the pilocarpine model of temporal lobe epilepsy. Exp Neurol (2010) 224:219–33. doi:10.1016/j.expneurol.2010.03.014
35. Tanaka S, Nakamura T, Sumitani K, Takahashi F, Konishi R, Itano T, et al. Stage- and region-specific cyclooxygenase expression and effects of a selective COX-1 inhibitor in the mouse amygdala kindling model. Neurosci Res (2009) 65(1):79–87. doi:10.1016/j.neures.2009.05.013
36. Ma L, Cui XL, Wang Y, Li XW, Yang F, Wei D, et al. Aspirin attenuates spontaneous recurrent seizures and inhibits hippocampal neuronal loss, mossy fiber sprouting and aberrant neurogenesis following pilocarpine-induced status epilepticus in rats. Brain Res (2012) 1469:103–13. doi:10.1016/j.brainres.2012.05.058
37. Barbalho PG, Lopes-Cendes I, Maurer-Morelli CV. Indomethacin treatment prior to pentylenetetrazole-induced seizures downregulates the expression of il1b and cox2 and decreases seizure-like behavior in zebrafish larvae. BMC Neurosci (2016) 17(1):12. doi:10.1186/s12868-016-0246-y
38. Teraoka H, Kubota A, Dong W, Kawai Y, Yamazaki K, Mori C, et al. Role of the cyclooxygenase 2-thromboxane pathway in 2,3,7,8-tetrachlorodibenzo-p-dioxin-induced decrease in mesencephalic vein blood flow in the zebrafish embryo. Toxicol Appl Pharmacol (2009) 234(1):33–40. doi:10.1016/j.taap.2008.09.021
39. Afrikanova T, Serruys AS, Buenafe OE, Clinckers R, Smolders I, de Witte PA, et al. Validation of the zebrafish pentylenetetrazol seizure model: locomotor versus electrographic responses to antiepileptic drugs. PLoS One (2013) 8(1):e54166. doi:10.1371/journal.pone.0054166
40. Choi SH, Aid S, Bosetti F. The distinct roles of cyclooxygenase-1 and -2 in neuroinflammation: implications for translational research. Trends Pharmacol Sci (2009) 30(4):174–81. doi:10.1016/j.tips.2009.01.002
41. Bosetti F, Choi SH. Rethinking the role of cyclooxygenase-1 in neuroinflammation: more than homeostasis. Cell Cycle (2010) 9(15):2919–20. doi:10.4161/cc.9.15.12715
42. Aïd S, Bosetti F. Targeting cyclooxygenases-1 and -2 in neuroinflammation: therapeutic implications. Biochimie (2011) 93(1):46–51. doi:10.1016/j.biochi.2010.09.009
43. Hoozemans JJ, Rozemuller AJ, Janssen I, De Groot CJ, Veerhuis R, Eikelenboom P. Cyclooxygenase expression in microglia and neurons in Alzheimer’s disease and control brain. Acta Neuropathol (2001) 101(1):2–8. doi:10.1007/s004010000251
44. McGeer PL, McGeer EG. NSAIDs and Alzheimer disease: epidemiological, animal model and clinical studies. Neurobiol Aging (2007) 28(5):639–47. doi:10.1016/j.neurobiolaging.2006.03.013
45. Frautschy SA. Thinking outside the box about COX-1 in Alzheimer’s disease. Neurobiol Dis (2010) 38(3):492–4. doi:10.1016/j.nbd.2010.02.009
46. Choi SH, Aid S, Caracciolo L, Minami SS, Niikura T, Matsuoka Y, et al. Cyclooxygenase-1 inhibition reduces amyloid pathology and improves memory deficits in a mouse model of Alzheimer’s disease. J Neurochem (2013) 124(1):59–68. doi:10.1111/jnc.12059
47. Shang JL, Cheng Q, Yang WF, Zhang M, Cui Y, Wang YF. Possible roles of COX-1 in learning and memory impairment induced by traumatic brain injury in mice. Braz J Med Biol Res (2014) 47(12):1050–6. doi:10.1590/1414-431X20143601
48. Doost Mohammadpour J, Hosseinmardi N, Janahmadi M, Fathollahi Y, Motamedi F, Rohampour K. Non-selective NSAIDs improve the amyloid-β-mediated suppression of memory and synaptic plasticity. Pharmacol Biochem Behav (2015) 132:33–41. doi:10.1016/j.pbb.2015.02.012
49. Heneka MT, Carson MJ, El Khoury J, Landreth GE, Brosseron F, Feinstein DL, et al. Neuroinflammation in Alzheimer’s disease. Lancet Neurol (2015) 14(4):388–405. doi:10.1016/S1474-4422(15)70016-5
50. Fries S, Grosser T. The cardiovascular pharmacology of COX-2 inhibition. Hematology Am Soc Hematol Educ Program (2005) 2005:445–51. doi:10.1182/asheducation-2005.1.445
51. Stewart AM, Desmond D, Kyzar E, Gaikwad S, Roth A, Riehl R, et al. Perspectives of zebrafish models of epilepsy: what, how and where next? Brain Res Bull (2012) 87(2–3):135–43. doi:10.1016/j.brainresbull.2011.11.020
52. Stewart AM, Braubach O, Spitsbergen J, Gerlai R, Kalueff AV. Zebrafish models for translational neuroscience research: from tank to bedside. Trends Neurosci (2014) 37(5):264–78. doi:10.1016/j.tins.2014.02.011
53. Grone BP, Baraban SC. Animal models in epilepsy research: legacies and new directions. Nat Neurosci (2015) 18(3):339–43. doi:10.1038/nn.3934
54. Grosser T, Yusuff S, Cheskis E, Pack MA, FitzGerald GA. Developmental expression of functional cyclooxygenases in zebrafish. Proc Natl Acad Sci U S A (2002) 99(12):8418–23. doi:10.1073/pnas.112217799
55. Baxendale S, Holdsworth CJ, Meza Santoscoy PL, Harrison MR, Fox J, Parkin CA, et al. Identification of compounds with anti-convulsant properties in a zebrafish model of epileptic seizures. Dis Model Mech (2012) 5(6):773–84. doi:10.1242/dmm.010090
56. Cunliffe VT. Building a zebrafish toolkit for investigating the pathobiology of epilepsy and identifying new treatments for epileptic seizures. J Neurosci Methods (2015) 260:91–5. doi:10.1016/j.jneumeth.2015.07.015
57. MacRae CA, Peterson RT. Zebrafish as tools for drug discovery. Nat Rev Drug Discov (2015) 14(10):721–31. doi:10.1038/nrd4627
Keywords: zebrafish model, seizures, neuroinflammation, cyclooxygenases, anticonvulsant
Citation: Barbalho PG, Carvalho BS, Lopes-Cendes I and Maurer-Morelli CV (2016) Cyclooxygenase-1 as a Potential Therapeutic Target for Seizure Suppression: Evidences from Zebrafish Pentylenetetrazole-Seizure Model. Front. Neurol. 7:200. doi: 10.3389/fneur.2016.00200
Received: 12 September 2016; Accepted: 31 October 2016;
Published: 15 November 2016
Edited by:
Jeremy Daniel Slater, University of Texas Medical School at Houston, USAReviewed by:
Dieter Schmidt, Epilpesy Research Group, GermanyLuciene Covolan, Federal University of São Paulo, Brazil
Copyright: © 2016 Barbalho, Carvalho, Lopes-Cendes and Maurer-Morelli. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Claudia Vianna Maurer-Morelli, Y21hdXJlckBmY20udW5pY2FtcC5icg==