 Hongxi Ma1†
Hongxi Ma1† Cong Cao
Cong Cao- 1Clinical Laboratory, Wuzhou Gongren Hospital, Wuzhou, China
- 2Department of General Practice, The Affiliated Hospital of Youjiang Medical University for Nationalities, Baise, China
- 3Department of Ultrasonics, The Affiliated Hospital of Youjiang Medical University for Nationalities, Baise, China
Recent studies have reported a close association between circRNAs and cancer development. CircRNAs have been recognized to be involved in various biological processes. Up to now, the function of circRNAs in hepatocellular carcinoma (HCC) is still poorly known. qRT-PCR was used to test circ_0014717 expression in HCC tissue samples and cells was determined. It was shown that circ_0014717 was significantly decreased in HCC. Then, we observed overexpression of circ_0014717 obviously repressed HCC cell growth, migration and invasion. Next, we predicted circ_0014717 acted as a sponge of miR-668-3p. miR-668-3p has been reported to participate in several diseases. In our work, it was shown miR-668-3p was greatly increased in HCC and the direct binding sites between circ_0014717 and miR-668-3p were validated. In addition, B-cell translocation gene 2 (BTG2) is closely involved in cellular carcinogenic processes. BTG2 was predicted as a target for miR-668-3p. By performing rescue assays, we demonstrated that circ_0014717 repressed HCC progression via inhibiting BTG2 expression and sponging miR-668-3p. It was manifested loss of circ_0014717 induced HCC progression, which was reversed by BTG2 in Hep3B cells. In conclusion, our findings illustrated a novel circ_0014717/miR-668-3p/BTG2 regulatory signaling pathway in HCC.
Introduction
Recently, hepatocellular carcinoma (HCC) is becoming a prevalent cancer across the world (1, 2). Meanwhile, in China, HCC contribute a lot to a number of cancer-related death every year (3). In past few decades, great efforts and advances are made in order to improve the treatment for HCC. Despite those, the 5-year survival rate of HCC is still low (4). Herein, it is urgent to identify effective biomarkers for HCC and develop therapeutic targets.
In recent years, circRNAs are attracting increasing attention as a new significant member of non-coding RNAs (5, 6). CircRNAs are covalently closed transcripts derived from precursor mRNA back splicing (7). circRNAs may exhibit specific biological functions and they are significant mediators in tumors through various mechanisms (8–10). For example, circTP63 can act as a ceRNA to induce lung cancer progression via inducing FOXM1 (11). CircRNA_100782 contributes to pancreatic carcinoma development via activating the IL6-STAT3 pathway (12). CircRNA_5692 represses the progression of HCC via sponging miR-328-5p to induce DAB2IP level (13).
Previously, it has been shown that circ_0014717 is obviously reduced in gastric cancer tissues (14). Additionally, in colorectal cancer, circ_0014717 can exhibit tumor-suppressive roles (15). These implied circ_0014717 can serve as an important tumor inhibitor in various cancers. However, the detail role of circ_0014717 in HCC development is barely known.
Currently, the expression of circ_0014717 in HCC was determined. The underlying mechanism of circ_0014717 was investigated by evaluating its downstream microRNA target miR-668-3p and BTG2 in Hep3B and SMMC-7721 cells. In addition, we overexpressed circ_0014717 in HCC cell lines to elucidate in vitro and in vivo functions. Thus, we hypothesized down-regulated circ_0014717 expression may act as a promising biomarker for HCC via regulating miR-668-3p and BTG2.
Materials and Methods
Clinical Specimens
30 pairs of HCC and tumor-adjacent tissues were obtained from patients with hepatectomy at The Affiliated Hospital of Youjiang Medical University for Nationalities. No operative treatments, including radiofrequency ablation, immunotherapy or targeted therapy were given to the patients before the surgery. Samples were maintained in liquid nitrogen for RNA and protein isolation. This research was approved by the Ethics Committee of The Affiliated Hospital of Youjiang Medical University for Nationalities, and all subjects gave the written informed consent.
Cell Culture
LO2, HepG2, Hep3B, Huh7, SMMC-7721, MHCC97L cells were purchased from ATCC (Manassas, VA, USA). DMEM medium (Gibco BRL, Grand Island, NY, USA) was used to incubate the cells. The medium was added with 10% FBS (Gibco BRL, Grand Island, NY, USA) and antibiotics (100 μg/ml streptomycin and 100 U/ml penicillin, Sigma, St-Louis, MO, USA) in a humidified incubator with 5% CO2 at 37°C.
Cell Transfection
Lentivector-mediated shRNA of circ_0014717 (LV-shcirc_0014717) and non-targeting sh-control were synthesized by GeneChem (Shanghai, China). The full-length of circ_0014717 were sub-cloned into the lentivirus vector (LV-circ_0014717) by GeneChem (Shanghai, China). Lentivirus infection was carried out under 8 ng/ml Polybrene. pcDNA3.1-BTG2 and the empty plasmid pcDNA3.1 were obtained from GeneChem (Shanghai, China). miR-668-3p mimics, inhibitors and negative controls were obtained from RiboBio (Guangzhou, China). Lipofectamine 3000 (Invitrogen, Carlsbad, CA, USA) was employed to do cell transfection.
CCK8 Assay
To carry out CCK8 assay, the transfected cells were seeded into 96-well plates with 3,000 cells in each well. Ten microliters of CCK 8 solution (KeyGene, BioTECH, China) was added to the cells and they were maintained at 37°C for 2 h. The OD values were tested at 450 nm using a SpectraMax microtiter plate reader.
EdU Assay
EdU assay was carried out using EdU kit (Roche, Indianapolis, IN, USA). Results were obtained using Zeiss fluorescence photomicroscope (Carl Zeiss, Oberkochen, Germany).
Clone Formation Assay
To carry out clone formation experiment, the transfected cells were seeded in six-well plates. Then, after cells were cultured for 2 weeks, cells were fixed using 30% formaldehyde for 15 min and stained using 0.1% crystal violet (Beyotime Biotechnology, Shanghai, China).
Flow Cytometry Assay
To carry out cell cycle analysis, cells were fixed by 70% ethanol. 1 ×106 cells were resuspended using 500 μl PI/RNase Staining Buffer. Then, the cells were incubated for 15min with no light. A FACSCanto II flow cytometer (BD biosciences, San Jose, CA, USA) was utilized to analyze cell cycle. To perform cell apoptosis assay, the PE Annexin V Apoptosis Detection Kit I was used. After cells were washed using pre-cold PBS buffer, 5 μl PE Annexin V and 5 μl FITC solution were added to the cells.
Transwell Assay
To perform the migration assay, cells were trypsinized and then grown in the upper chamber of each insert (Corning, Cambridge, USA) with non-coated membrane with 1% FBS (600 μl). After 24 h, the upper surface of the membrane was removed by using a cotton tip. Cells on the lower surface were stained for half an hour with 0.1% crystal violet. To carry out the invasion assay, matrigel chambers (BD Biosciences, San Jose, CA, USA) were performed. Briefly, cells were collected, re-suspended in medium without serum, and shifted to the hydrated matrigel chambers. The bottom chambers were incubated in 500 μl culture medium with 10% FBS. Then, we scraped the cells on the upper surface, whereas the invasive cells on the lower surface were fixed and colored using 0.1% crystal violet for half an hour.
Western Blot
Cell lysates were extracted by RIPA buffer added with a protease inhibitor cocktail. Equal amounts of protein samples were separated on 10% SDS-polyacrylamide gel electrophoresis and then transferred onto PVDF membranes (Bio-Rad, CA, USA). After blocked in 5% non-fat dry milk, the PVDF membranes were incubated with the anti-human BTG2 antibody (dilution: 1:1,000, Cell Signaling Technology, MA, USA) and GAPDH antibody (dilution: 1:2,000, Cell Signaling Technology, MA, USA). The protein bonds were visualized by a chemiluminescent detection system (Millipore, Bedford, MA, USA). A FluroChem E Imager (Protein Simple, Santa Clara, CA, USA) was carried out to visualize the western blots.
Real−Time PCR Analysis
Total RNA of HCC cells and tissues was isolated using TRIzol. 1 µg DNase-treated RNA was reverse transcribed to cDNA using MMLV reverse transcriptase (Takara, Tokyo, Japan). Then, relative quantitative real-time PCR was conducted using SYBR Premix Ex Taq II (Takara, Tokyo, Japan) on LightCycler 96 (Roche, Penzberg, Germany). Then, the expression of target genes was detected using the formula 2^-ΔΔCt. The sequences of primers were demonstrated in Table 1.
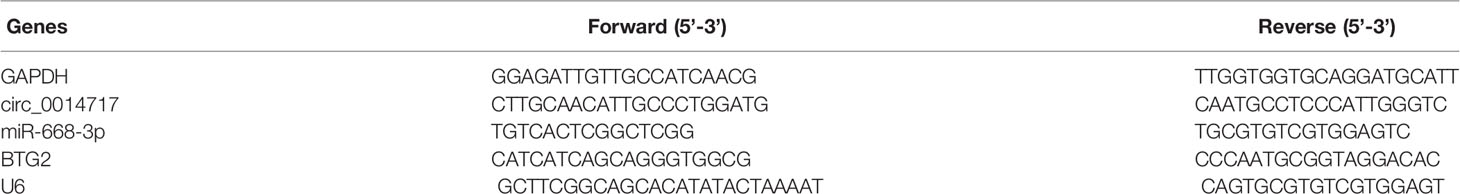
Table 1 Primers for real-time PCR.
RNA Pull-Down Assay
The biotinylated probe was constructed to bind to the junction area of circ_0014717. The circ_0014717 probe (Tsingke, Wuhan, China) was incubated with streptavidin magnetic beads (Life Technologies: Carlsbad, CA, USA). The bound miRNAs in the pull-down materials were extracted using Trizol reagent and qRT-PCR assay was carried out to detect miRNA expression.
Luciferase Activity Assays
The sequence of 3’-UTR of BTG2 or circ_0014717 were subcloned into pGL3 luciferase reporter vector (Promega, Madison, WI, USA). The WT/MUT 3’-UTR of BTG2 vector or WT/MUT circ_0014717 vector and control mimics or miR-668-3p mimics were co-transfected. The luciferase activity was normalized using a dual luciferase reporter assay system (Promega, Madison, WI, USA).
Tumor Xenograft
BALB/c nude mice (female, 4- to 5-week-old, 18–20g) were purchased from SLAC (Shanghai, China) and divided into two groups (n = 6 in each group). Hep3B cells (5 ×106 per injection) that were transfected with LV-circ_0014717 and LV-NC, respectively, were implanted into the right flank of the mice. After six weeks, mice were sacrificed under anesthesia. Tumor tissues were subjected to HE and Ki-67 staining. The animal experiments were approved by the Animal Care and Use Committee of The Affiliated Hospital of Youjiang Medical University for Nationalities.
Statistical Analysis
Data were analyzed using SPSS software 22.0 (Armonk, NY, USA). The Student’s t-test was carried out to analyze differences between two groups. Two-way ANOVA was carried out when more than two groups were compared. Differences were statistically significant if P <0.05.
Results
circ_0014717 Was Decreased in HCC
Firstly, we displayed that circ_0014717 in 30 pair of HCC tissues was greatly lower than that in matched tumor-adjacent tissues confirmed using qRT-PCR analysis in Figure 1A. Kaplan–Meier analysis revealed that the expression level of circ_0014717 was significantly associated with the overall survival (OS) and the time to tumor recurrence as displayed in Figures 1B, C. In addition, a decreased expression of circ_0014717 was observed in HCC cells (HepG2, Hep3B, SMMC-7721, Huh7 and MHCC97L) in comparison to LO2 cells in Figure 1D. We indicated that circ_0014717 down-regulation was a frequent event in HCC. The detailed clinical-pathological parameters of the obtained HCC samples in our study was provided in Table 2. Down-regulated circ_0014717 expression level was correlated with tumor size, TNM stage and metastasis.
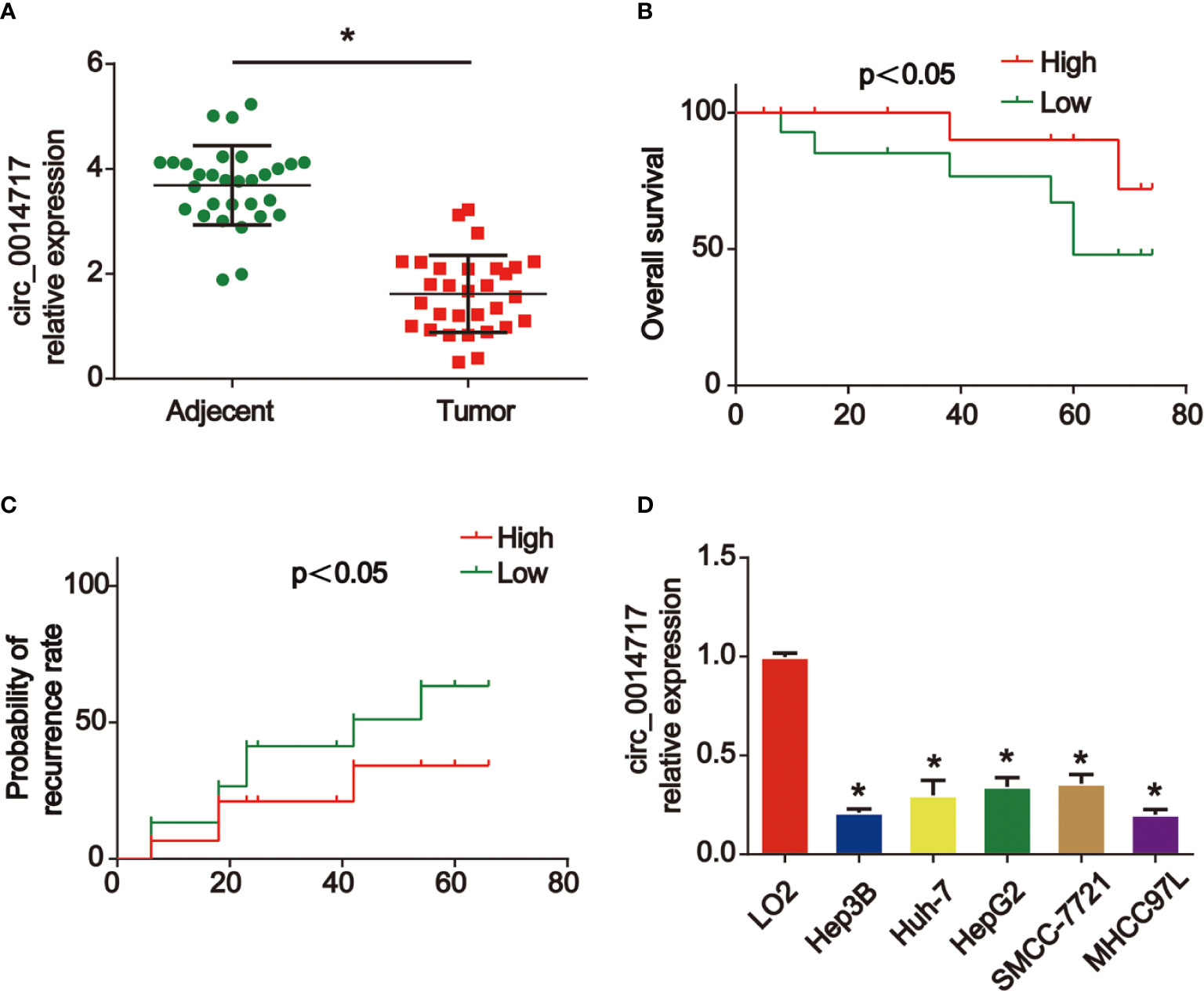
Figure 1 circ_0014717 was increased in hepatocellular carcinoma (HCC). (A) The expression of circ_0014717 was tested by real-time PCR in HCC carcinoma and normal adjacent tissues. (B, C) Kaplan-Meier analysis of the correlation between circ_0014717 expression and overall survival or recurrence in 30 HCC. (D) The expression of circ_0014717 in HCC cells (Huh-7, Hep-3B, HepG2, SMMCC-7721, MHCC97L) and LO2 cells. *P < 0.05.
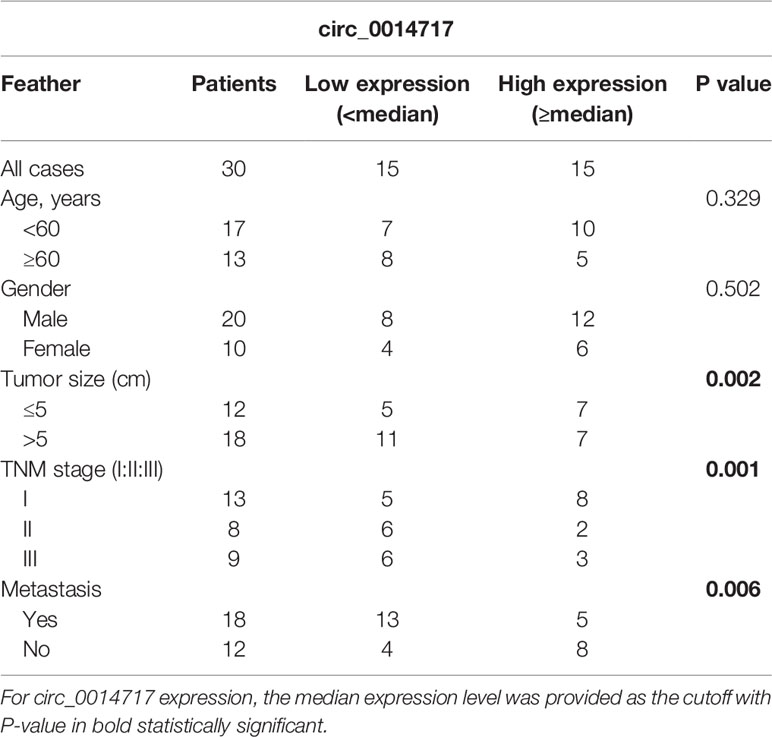
Table 2 Clinicopathological relevance analysis of circ_0014717 in hepatocellular carcinoma (HCC) patients.
Overexpression of circ_0014717 Suppressed Cell Growth, Migration, and Invasion of HCC Cells
Next, we studied the roles of circ_0014717 in HCC cell growth. Circ_0014717 was stably increased in Hep3B and SMMC7721 cells as shown in Figure 2A. CCK-8 assays indicated overexpression of circ_0014717 repressed the proliferation of Hep3B and SMMC7721 cells (Figures 2B, C). EdU incorporation assays also proved that circ_0014717 prominently restrained the growth of Hep3B and SMMC7721 cells (Figures 2D–F). Moreover, in Figures 2G–I, flow cytometry assays revealed that the cell cycle was blocked by circ_0014717 in G1 phase. Then, in Figures 2J–L, transwell migration assay indicated that increased circ_0014717 elevated the migration capacity of HCC cells. In addition, transwell invasion assay was conducted and we found that Hep3B and SMMC7721 cell invasion was significantly repressed by circ_0014717 as demonstrated in Figures 2M–O.
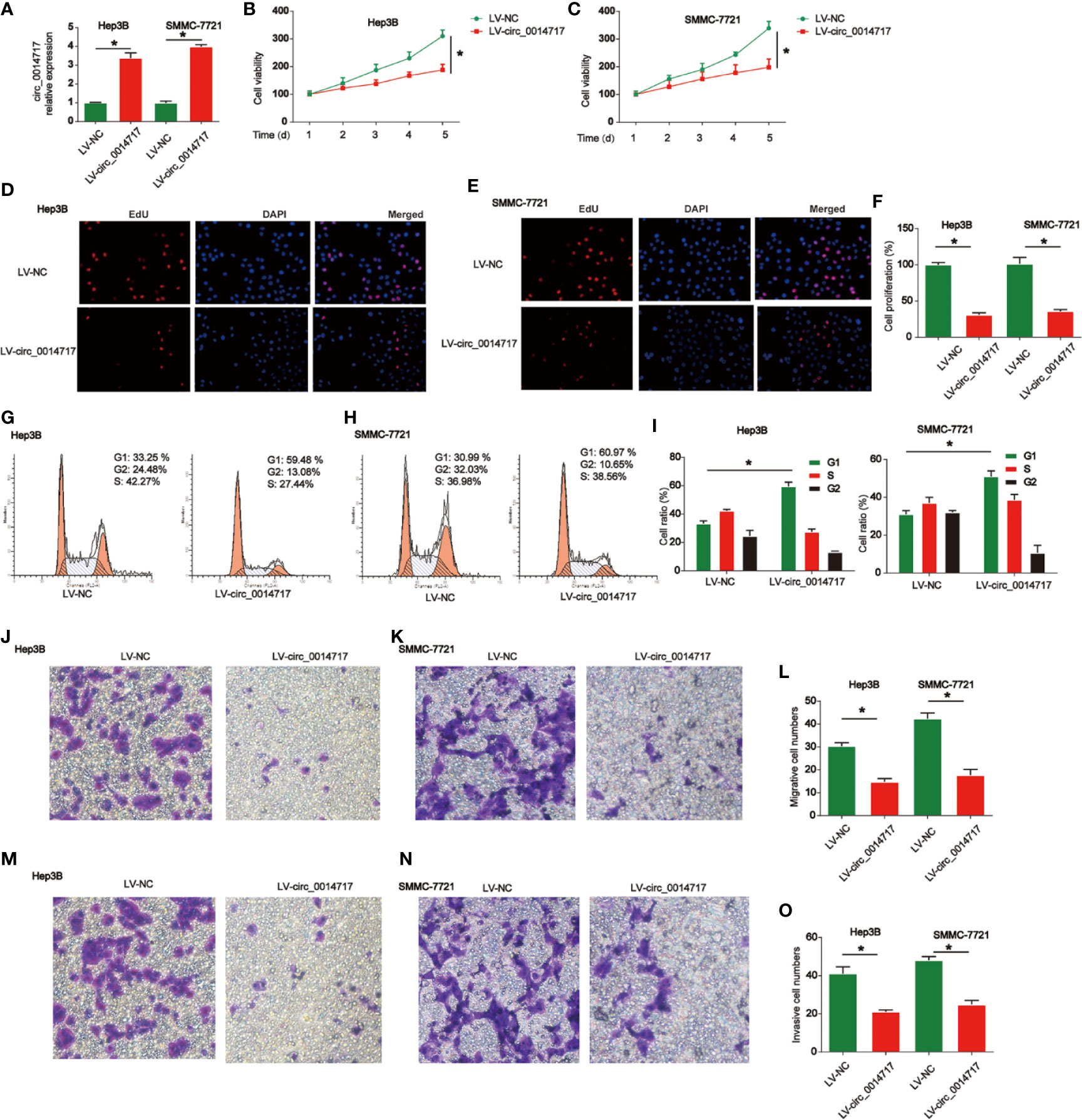
Figure 2 Effects of circ_0014717 overexpression on hepatocellular carcinoma (HCC) cells. (A) Expression level of circ_0014717 in HCC cells. Cells were infected with LV-circ_0014717 or LV-NC. (B, C) CCK-8 assay was conducted to test cell viability. (D–F) Effects of LV-circ_0014717 on HCC cell proliferation evaluated using EdU assay. (G–I) Effects of LV-circ_0014717 on HCC cell cycle distribution. Flow cytometry assay was carried out to detect cell cycle. (J–L) Effects of LV-circ_0014717 on HCC cell migration. Transwell migration assay was carried out to assess cell migration capacity. (M–O) Effects of LV-circ_0014717 on HCC cell invasion. Transwell invasion assay was utilized to detect cell invasion capacity. *P < 0.05.
Increased circ_0014717 Depressed Tumorigenesis of HCC In Vivo
To investigate the biological roles of circ_0014717 in HCC in vivo, Hep3B cells with increased circ_0014717 were implanted into nude mice. The findings of tumor growth curves and tumor weight demonstrated circ_0014717 obviously suppressed tumor growth in mice (Figures 3A–C). Subsequently, tumor tissues were harvested for HE and IHC staining in Figures 3D, E. Ki-67 assay evidenced that up-regulated circ_0014717 repressed Ki-67 positive cells in vivo. Altogether, these suggested that overexpression of circ_0014717 suppressed HCC tumorigenesis in vivo.
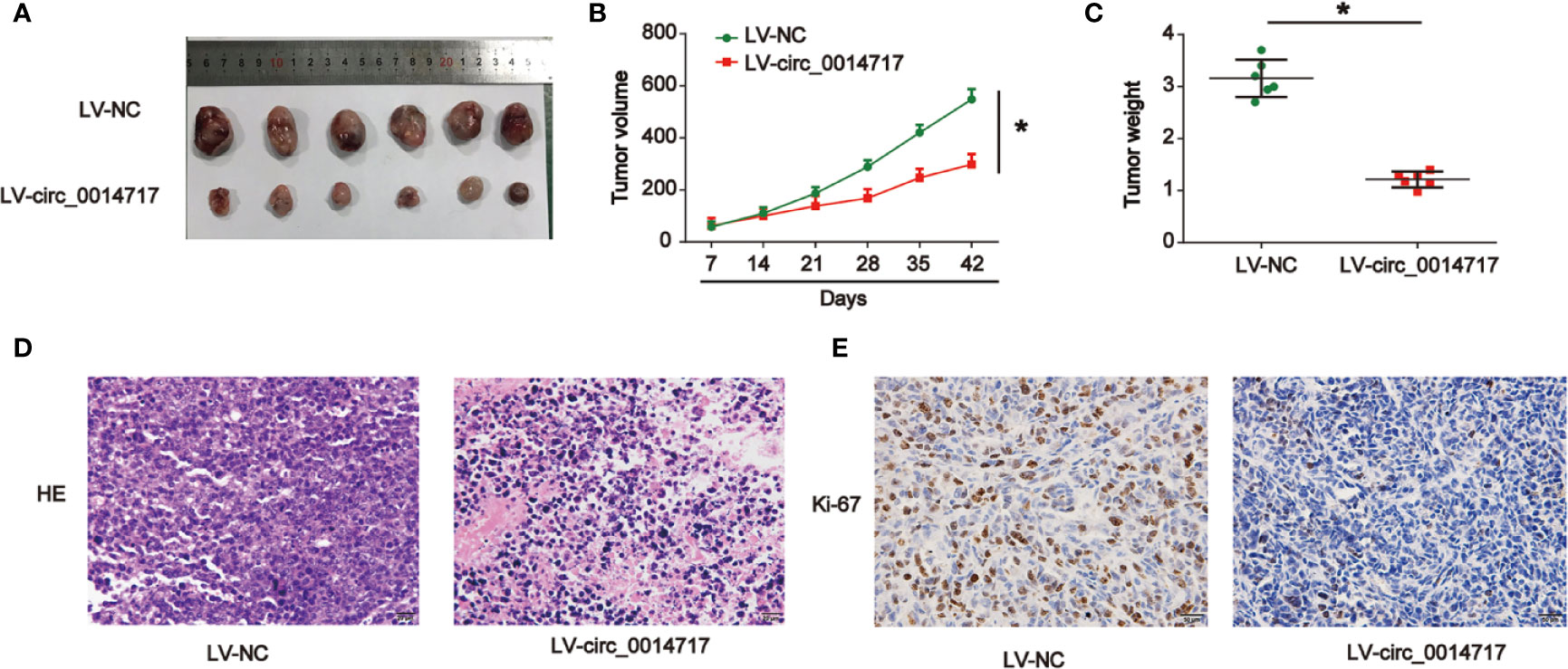
Figure 3 Up-regulation of circ_0014717 restrained hepatocellular carcinoma (HCC) cell growth in vivo. Twelve 6-week old female BALB/c nude mice were injected with Hep3B cells infected with LV-circ_0014717 or LV-NC. Six mice were used in each group. (A) Tumors were peeled from the mice. (B) Tumor volume. (C) Tumor weight. (D) H & E staining. (E) IHC staining of Ki-67 in tumor tissues. *P < 0.05.
Circ_0014717 Acted as a Sponge for miR-668-3p
To explore the mechanisms of the role of circ_0014717 in HCC, as displayed in Figures 4A, B, miR-668-3p was most abundantly pulled down by circ_0014717 in HCC cells via using https://circinteractome.nia.nih.gov/. Thus, we tested the expression level of miR-668-3p in Hep3B and SMMC7721 cells after circ_0014717 overexpression. Our data revealed that circ_0014717 significantly decreased miR-668-3p expression in HCC cells in Figure 4C. Furthermore, the binding sites between circ_0014717 and miR-668-3p was exhibited in Figure 4D. Luciferase reporter assay revealed miR-668-3p inhibitors down-regulated the luciferase activity of vectors containing WT-circ_0014717 rather than the vectors containing MUT-circ_0014717 in HCC cells in Figures 4E, F. Expression of miR-668-3p in HCC was significantly higher in Figures 4G, H.
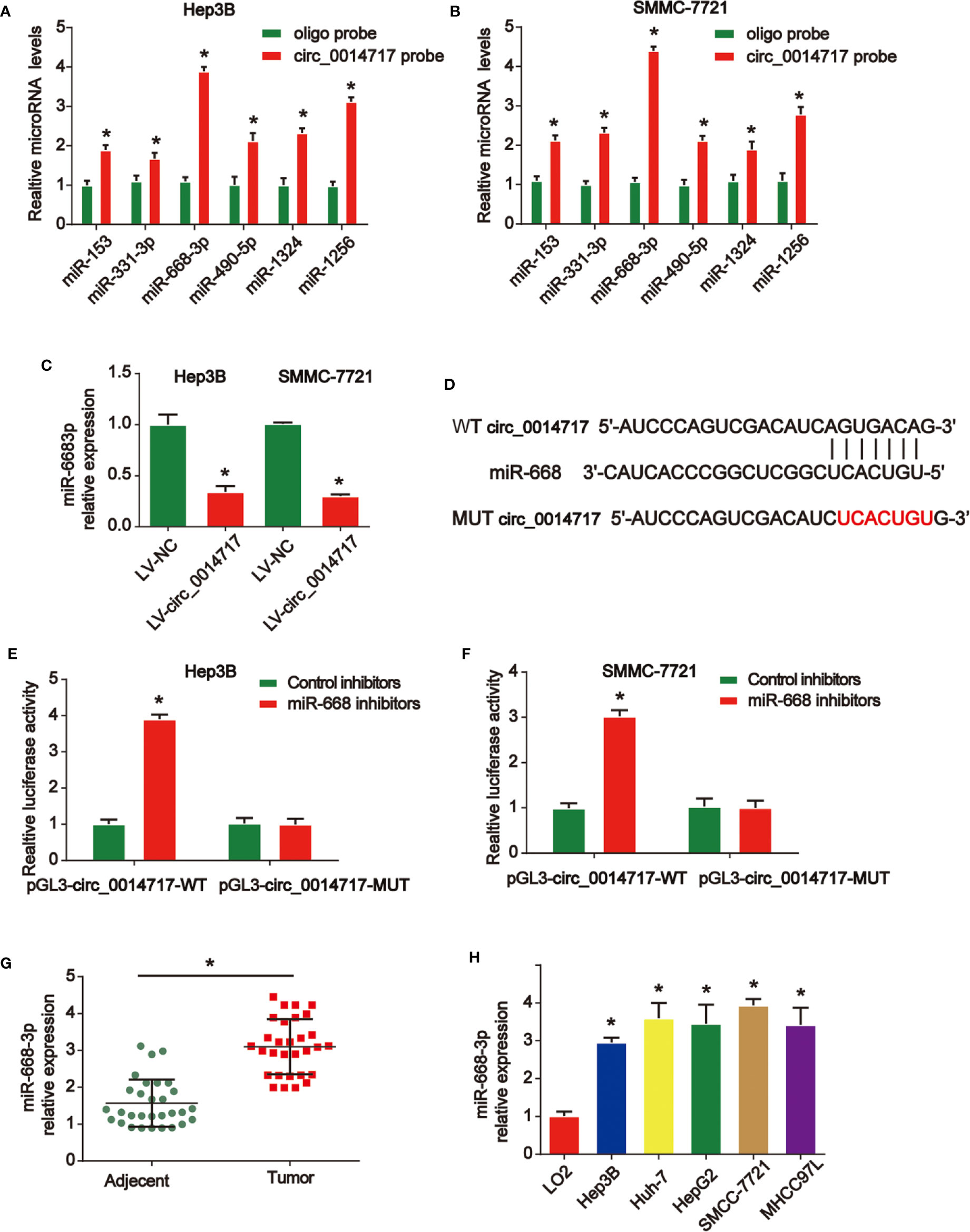
Figure 4 circ_0014717 sponged miR-668-3p in hepatocellular carcinoma (HCC) cells. (A, B) Top six miRNA candidates in HCC cell lysates were tested by real-time PCR. Multiple miRNAs were pulled down using circ_0014717 probe. (C) The expression level of miR-668-3p in HCC cells infected with LV-circ_0014717 or LV-NC. (D) The putative binding sites between circ_0014717 and miR-668-3p. (E, F) Luciferase activity was evaluated in HCC cells co-transfected with circ_0014717-WT or circ_0014717-MUT reporter and miR-668-3p inhibitors or its scramble control (NC). (G) The expression of miR-668-3p in HCC carcinoma and normal adjacent tissues. (H) The expression of miR-668-3p in HCC cells (Huh-7, Hep-3B, HepG2, SMMCC-7721, MHCC97L) and LO2 cells. *P < 0.05.
BTG2 Was A Novel Target of miR-668-3p
Then, we demonstrated the mRNA levels of top six predicted genes after the up-regulation of miR-668-3p in Hep3B and SMMC7721 cells in Figures 5A, B via consulting online bioinformatics analysis (http://starbase.sysu.edu.cn/). miR-668-3p overexpression depressed the expression of BTG2 in HCC cells most significantly. Then, it was indicated that BTG2 expression in Hep3B and SMMC7721 cells was reduced by miR-668-3p mimics in Figures 5C–F. In Figure 5G, the putative binding sites between miR-668-3p and BTG2 were exhibited. Then, luciferase activity was evaluated in Hep3B cells co-transfected with BTG2-WT or BTG2-MUT reporter and miR-668-3p mimics. In Figure 5H, miR-668-3p mimics induced the luciferase activity of vectors containing WT-BTG2 in Hep3B cells. Next, in Figures 5I–K, we verified the correlation of circ_0014717, miR-668-3p, and BTG2 in HCC tissues.
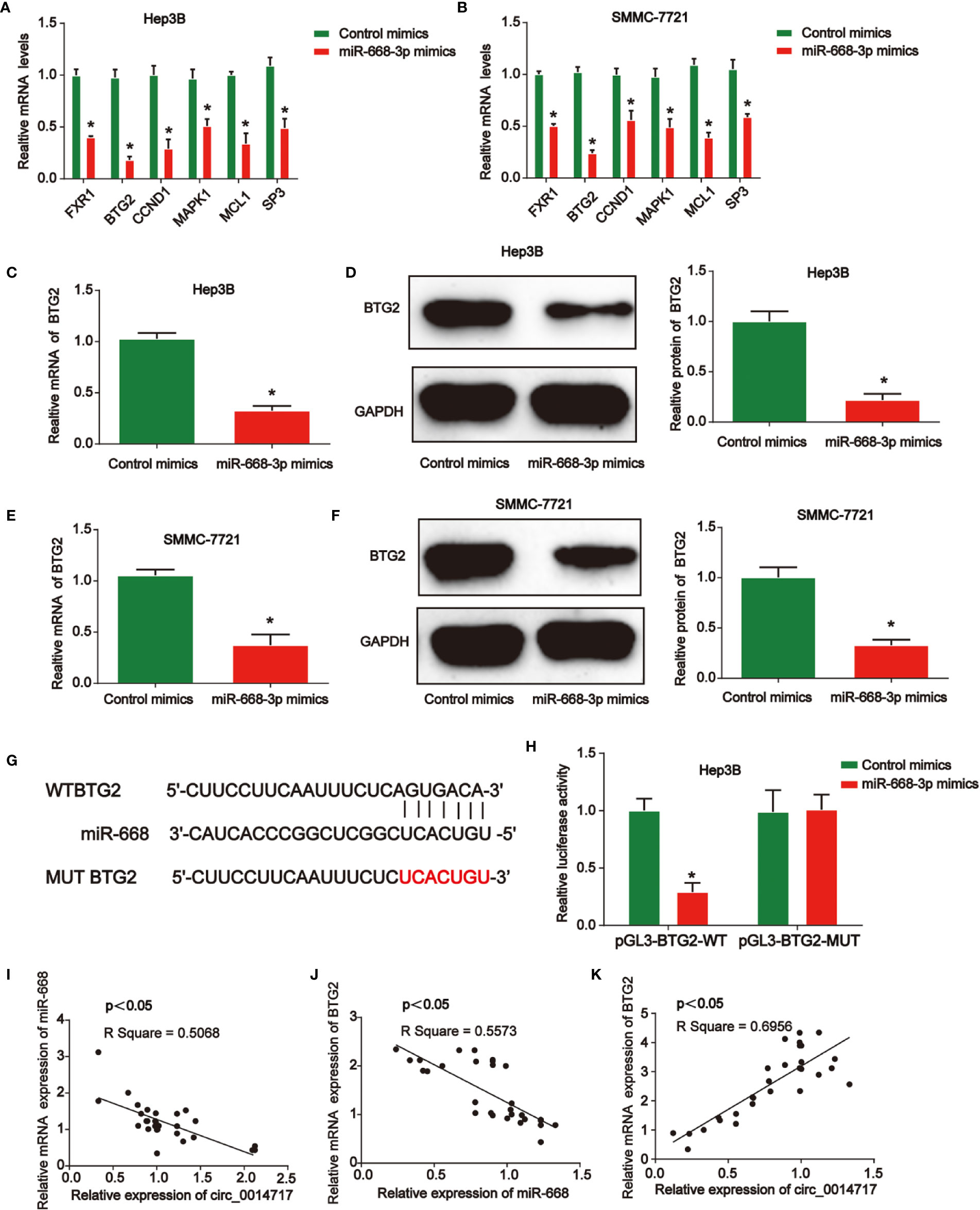
Figure 5 B-cell translocation gene 2 (BTG2) was a direct target of miR-668-3p in hepatocellular carcinoma (HCC) cells. (A, B) Top six mRNA candidates in HCC cell lysates regulated by miR-668-3p mimics were tested by real-time PCR. (C, D) BTG2 expression in Hep3B cells transfected with miR-668-3p mimics. (E, F) BTG2 expression in SMMC-7721 cells transfected with miR-668-3p mimics. (G) The putative binding sites between miR-668-3p and BTG2. (H) Luciferase activity was evaluated in Hep3B cells co-transfected with BTG2-WT or BTG2-MUT reporter and miR-668-3p mimics or its scramble control (NC). (I–K) The correlation of circ_0014717, miR-668-3p, and BTG2 in HCC tissues. *P < 0.05.
Restoration of BTG2 Reversed the Effect of circ_0014717 Down-Regulation on HCC Cells
To study whether BTG2 was critical for HCC cell proliferation, apoptosis and invasion upon loss of circ_0014717, Hep3B cells with circ_0014717 knockdown were transfected with BTG2 expression vectors. In Figure 6A, we confirmed the effect of LV-shcirc_0014717-1 and it exhibited the best knockdown effect in Hep3B cells. LV-shcirc_0014717-1 was used in the following assays. Moreover, we found that BTG2 expression was obviously decreased by circ_0014717 shRNA as exhibited in Figures 6B–D. In Figure 6E, CCK8 assays proved BTG2 reduced the pro-proliferation role of circ_0014717 knockdown in Hep3B cells. Flow cytometry assays indicated BTG2 overexpression induced apoptosis in Hep3B cells with circ_0014717 knockdown in Figure 6F. Furthermore, colony formation also indicated that overexpression of BTG2 reduced the colony formation induced by loss of circ_0014717 as exhibited in Figure 6G. Subsequently, Hep3B cell invasion was increased by lack of circ_0014717, which was obviously repressed by the up-regulation of BTG2 in Figure 6H. Thus, these results suggested circ_0014717 knockdown triggered HCC cell growth was partially mediated by the repression of BTG2 in HCC. Subsequently, we demonstrated the mechanism diagram of our entire study, which implied that circ_0014717 act as a ceRNA sponge of miR-668-3p to regulate BTG2 expression in HCC progression (Figure 6I).
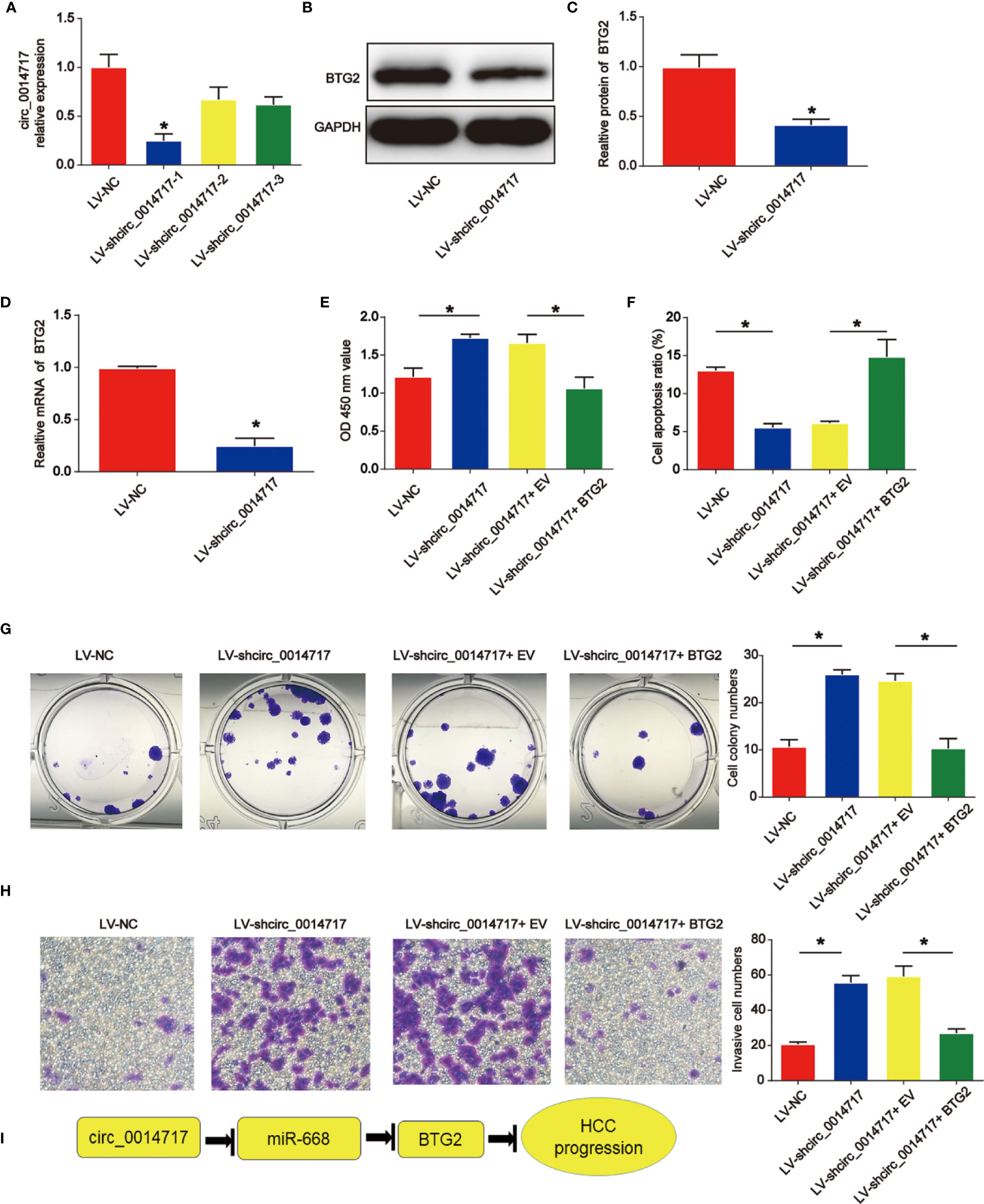
Figure 6 Effects of circ_0014717 shRNA on hepatocellular carcinoma (HCC) cell growth was reversed by B-cell translocation gene 2 (BTG2). (A) Circ_0014717 expression in Hep3B cells. Hep3B cells were infected with circ_0014717 shRNA-1, 2 or 3 for 48 h. (B–D) BTG2 protein and mRNA expression in Hep3B cells. (E) CCK-8 assay was carried out to evaluate Hep3B cell viability. Hep3B cells with decreased circ_0014717 were transfected with BTG2 vector or EV. (F) Apoptosis assay was used to assess cell apoptosis. (G) Colony formation assay was conducted to test Hep3B cell colony formation capacity. (H) Transwell invasion assay was used to detect Hep3B cell invasion. (I) Mechanism diagram of circ_0014717/miR-668-3p/BTG2 in HCC progression. *P < 0.05.
Discussion
CircRNAs exist in a wide range among various organisms, such as human cells (16). Importantly, since circRNAs are disease-specific, they can function as a potential biomarker for many diseases (17, 18). CircRNAs can manifest many functions, including microRNA sponges, RBP sponges or mRNA masters (19–21).
In our current work, we first reported that circ_0014717 was greatly decreased in HCC tissues and cells. Low circ_0014717 expression levels were associated with significantly reduced overall survival and an increased risk of tumor recurrence. Circ_0014717 might represent an independent prognostic biomarker in HCC patients. Then, overexpression of circ_0014717 was induced in Hep3B and SMMC-7721 cells. We found that circ_0014717 repressed the progression of HCC significantly. Previous studies have confirmed that circ_0014717 is reduced in gastric and colorectal cancer (14, 15). Then, we explored the detailed mechanism of circ_0014717 in HCC progression.
Many studies have investigated the roles of microRNAs in cancers. Many aberrantly expressed miRNAs are reported in the progression of HCC (22–24). As reported, most circRNA molecules have the miRNA response elements, which enable them bind to the corresponding miRNAs. Circ_001569 can induce colorectal cancer growth through targeting miR-145 (25). Circ_RNA circNRIP1 can sponge miRNA-149-5p to promote gastric cancer progression via regulating AKT1/mTOR signaling (26). Here, in our work, miR-668-3p was predicted as the target for circ_0014717. miR-668 has been reported to enhance the radio-resistance of breast cancer cell through targeting IκBα expression (27). Here, miR-668-3p was greatly induced in HCC. Result indicated that miR-668-3p and circ_0014717 exhibit opposite expression trends in HCC tissues. Then, we elucidated the direct biological function of the circ_0014717/miR-668-3p axis. We found overexpression of circ_0014717 obviously reduced miR-668-3p expression in HCC cells. This suggested that circ_0014717 may negatively regulate miR-668-3p. Their direct association between them was confirmed in our study for the first time.
Next, BTG2 was found to be the downstream target of miR-668-3p. The function of BTG2 in tumor growth is well explored in several reports (28–30). For instance, BTG2 as a tumor suppressor gene, is upregulated by p53 and PTEN in bladder carcinoma cells (31). In addition, BTG2 is reduced and HCC cancer stem cell-like features are inhibited by BTG2 (32). In our work, we confirm that increased BTG2 reversed the effect of circ_0014717 shRNA on HCC cell growth.
Our investigation has several limitations. The function of the circ_0014717-miR-668-3p-BTG2 axis is warranted to be verified in vivo. Although several circ_0014717 and miR-668-3p target genes were predicted using bioinformatics, whether these represent bona fide targets in HCC is still to be studied, which requires more experimental analyses.
In summary, the expression level of circ_0014717 was reduced in HCC. Meanwhile, expression of the downstream target miR-668-3p was negatively correlated with circ_0014717. Additionally, BTG2 was confirmed as the direct target for miR-668-3p. These data uncover a novel circ_0014717-miR-668-3p-BTG2 signaling in HCC cell growth.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
The studies involving human participants were reviewed and approved by the Medical Ethics Committee of The Affiliated Hospital of Youjiang Medical University for Nationalities. The patients/participants provided their written informed consent to participate in this study. The animal study was reviewed and approved by the Medical Ethics Committee of The Affiliated Hospital of Youjiang Medical University for Nationalities.
Author Contributions
HM, CH, and QH performed the experiments and prepared the manuscript. GL and JL collected and analyzed the data. BH and QZ supported administration, technique and materials. CC designed and supervised the study, and revised the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This study was supported by the National Natural Science Foundation of Guangxi Province (2020JJA140192), the Young and middle-aged teachers’ basic scientific research ability improvement project in Guangxi colleges and universities (No. 2020KY13020), and the Innovation Project of GuangXi Graduate Education (JGY2020166).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA: Cancer J Clin (2015) 65(2):87–108. doi: 10.3322/caac.21262
2. Sayiner M, Golabi P, Younossi ZM. Disease Burden of Hepatocellular Carcinoma: A Global Perspective. Digest Dis Sci (2019) 64(4):910–7. doi: 10.1007/s10620-019-05537-2
3. Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA: Cancer J Clin (2011) 61(2):69–90. doi: 10.3322/caac.20107
4. Oliveri RS, Wetterslev J, Gluud C. Hepatocellular carcinoma. Lancet (2012) 380(9840):470; author reply –1. doi: 10.1016/S0140-6736(12)61285-9
5. Consortium EP. An integrated encyclopedia of DNA elements in the human genome. Nature (2012) 489(7414):57–74. doi: 10.1038/nature11247
6. Sana J, Faltejskova P, Svoboda M, Slaby O. Novel classes of non-coding RNAs and cancer. J Trans Med (2012) 10:103. doi: 10.1186/1479-5876-10-103
7. Chen LL. The biogenesis and emerging roles of circular RNAs. Nat Rev Mol Cell Biol (2016) 17(4):205–11. doi: 10.1038/nrm.2015.32
8. Yao T, Chen Q, Fu L, Guo J. Circular RNAs: Biogenesis, properties, roles, and their relationships with liver diseases. Hepatol Res (2017) 47(6):497–504. doi: 10.1111/hepr.12871
9. Shang Q, Yang Z, Jia R, Ge S. The novel roles of circRNAs in human cancer. Mol Cancer (2019) 18(1):6. doi: 10.1186/s12943-018-0934-6
10. Li P, Chen S, Chen H, Mo X, Li T, Shao Y, et al. Using circular RNA as a novel type of biomarker in the screening of gastric cancer. Clinica chimica acta; Int J Clin Chem (2015) 444:132–6. doi: 10.1016/j.cca.2015.02.018
11. Cheng Z, Yu C, Cui S, Wang H, Jin H, Wang C, et al. circTP63 functions as a ceRNA to promote lung squamous cell carcinoma progression by upregulating FOXM1. Nat Commun (2019) 10(1):3200. doi: 10.1038/s41467-019-11162-4
12. Chen G, Shi Y, Zhang Y, Sun J. CircRNA_100782 regulates pancreatic carcinoma proliferation through the IL6-STAT3 pathway. OncoTargets Ther (2017) 10:5783–94. doi: 10.2147/OTT.S150678
13. Liu Z, Yu Y, Huang Z, Kong Y, Hu X, Xiao W, et al. CircRNA-5692 inhibits the progression of hepatocellular carcinoma by sponging miR-328-5p to enhance DAB2IP expression. Cell Death Dis (2019) 10(12):900. doi: 10.1038/s41419-019-2089-9
14. Shao Y, Li J, Lu R, Li T, Yang Y, Xiao B, et al. Global circular RNA expression profile of human gastric cancer and its clinical significance. Cancer Med (2017) 6(6):1173–80. doi: 10.1002/cam4.1055
15. Li Z, Ruan Y, Zhang H, Shen Y, Li T, Xiao B. Tumor-suppressive circular RNAs: Mechanisms underlying their suppression of tumor occurrence and use as therapeutic targets. Cancer Sci (2019) 110(12):3630–8. doi: 10.1111/cas.14211
16. Memczak S, Jens M, Elefsinioti A, Torti F, Krueger J, Rybak A, et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature (2013) 495(7441):333–8. doi: 10.1038/nature11928
17. Meng S, Zhou H, Feng Z, Xu Z, Tang Y, Li P, et al. CircRNA: functions and properties of a novel potential biomarker for cancer. Mol Cancer (2017) 16(1):94. doi: 10.1186/s12943-017-0663-2
18. Kristensen LS, Hansen TB, Veno MT, Kjems J. Circular RNAs in cancer: opportunities and challenges in the field. Oncogene (2018) 37(5):555–65. doi: 10.1038/onc.2017.361
19. Li X, Yang L, Chen LL. The Biogenesis, Functions, and Challenges of Circular RNAs. Mol Cell (2018) 71(3):428–42. doi: 10.1016/j.molcel.2018.06.034
20. Wilusz JE. A 360 degrees view of circular RNAs: From biogenesis to functions. Wiley Interdiscip Rev RNA (2018) 9(4):e1478. doi: 10.1002/wrna.1478
21. Zhang Z, Yang T, Xiao J. Circular RNAs: Promising Biomarkers for Human Diseases. EBioMedicine (2018) 34:267–74. doi: 10.1016/j.ebiom.2018.07.036
22. Tsuchiya N, Sawada Y, Endo I, Saito K, Uemura Y, Nakatsura T. Biomarkers for the early diagnosis of hepatocellular carcinoma. World J Gastroenterol (2015) 21(37):10573–83. doi: 10.3748/wjg.v21.i37.10573
23. Wei L, Wang X, Lv L, Liu J, Xing H, Song Y, et al. The emerging role of microRNAs and long noncoding RNAs in drug resistance of hepatocellular carcinoma. Mol Cancer (2019) 18(1):147. doi: 10.1186/s12943-019-1086-z
24. Xu J, Li J, Zheng TH, Bai L, Liu ZJ. MicroRNAs in the Occurrence and Development of Primary Hepatocellular Carcinoma. Adv Clin Exp Med (2016) 25(5):971–5. doi: 10.17219/acem/36460
25. Xie H, Ren X, Xin S, Lan X, Lu G, Lin Y, et al. Emerging roles of circRNA_001569 targeting miR-145 in the proliferation and invasion of colorectal cancer. Oncotarget (2016) 7(18):26680–91. doi: 10.18632/oncotarget.8589
26. Zhang X, Wang S, Wang H, Cao J, Huang X, Chen Z, et al. Circular RNA circNRIP1 acts as a microRNA-149-5p sponge to promote gastric cancer progression via the AKT1/mTOR pathway. Mol Cancer (2019) 18(1):20. doi: 10.1186/s12943-018-0935-5
27. Luo M, Ding L, Li Q, Yao H. miR-668 enhances the radioresistance of human breast cancer cell by targeting IkappaBalpha. Breast Cancer (2017) 24(5):673–82. doi: 10.1007/s12282-017-0756-1
28. Lim IK, Lee MS, Lee SH, Kim NK, Jou I, Seo JS, et al. Differential expression of TIS21 and TIS1 genes in the various organs of Balb/c mice, thymic carcinoma tissues and human cancer cell lines. J Cancer Res Clin Oncol (1995) 121(5):279–84. doi: 10.1007/BF01209594
29. Ryo A, Uemura H, Ishiguro H, Saitoh T, Yamaguchi A, Perrem K, et al. Stable suppression of tumorigenicity by Pin1-targeted RNA interference in prostate cancer. Clin Cancer Res (2005) 11(20):7523–31. doi: 10.1158/1078-0432.CCR-05-0457
30. Elmore LW, Di X, Dumur C, Holt SE, Gewirtz DA. Evasion of a single-step, chemotherapy-induced senescence in breast cancer cells: implications for treatment response. Clin Cancer Res (2005) 11(7):2637–43. doi: 10.1158/1078-0432.CCR-04-1462
31. Tsui KH, Chiang KC, Lin YH, Chang KS, Feng TH, Juang HH. BTG2 is a tumor suppressor gene upregulated by p53 and PTEN in human bladder carcinoma cells. Cancer Med (2018) 7(1):184–95. doi: 10.1002/cam4.1263
Keywords: hepatocellular carcinoma, circ_0014717, miR-668-3p, B-cell translocation gene 2, ceRNA
Citation: Ma H, Huang C, Huang Q, Li G, Li J, Huang B, Zhong Q and Cao C (2021) Circular RNA circ_0014717 Suppresses Hepatocellular Carcinoma Tumorigenesis Through Regulating miR-668-3p/BTG2 Axis. Front. Oncol. 10:592884. doi: 10.3389/fonc.2020.592884
Received: 08 August 2020; Accepted: 26 November 2020;
Published: 27 January 2021.
Edited by:
Ke Ai-wu, Fudan University, ChinaReviewed by:
Bai-Quan Qiu, Second Affiliated Hospital of Nanchang University, ChinaKam Hui, National Cancer Centre Singapore, Singapore
Copyright © 2021 Ma, Huang, Huang, Li, Li, Huang, Zhong and Cao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cong Cao, caocong281261971@163.com
†These authors have contributed equally to this work