 Brian A. Corgiat
Brian A. Corgiat Jacob C. Nordman
Jacob C. Nordman Nadine Kabbani
Nadine Kabbani- Department of Molecular Neuroscience, Krasnow Institute for Advanced Study, George Mason University, Fairfax, VA, USA
Receptor function is dependent on interaction with various intracellular proteins that ensure the localization and signaling of the receptor. While a number of approaches have been optimized for the isolation, purification, and proteomic characterization of receptor–protein interaction networks (interactomes) in cells, the capture of receptor interactomes and their dynamic properties remains a challenge. In particular, the study of interactome components that bind to the receptor with low affinity or can rapidly dissociate from the macromolecular complex is difficult. Here we describe how chemical crosslinking (CC) can aid in the isolation and proteomic analysis of receptor–protein interactions. The addition of CC to standard affinity purification and mass spectrometry protocols boosts the power of protein capture within the proteomic assay and enables the identification of specific binding partners under various cellular and receptor states. The utility of CC in receptor interactome studies is highlighted for the nicotinic acetylcholine receptor as well as several other receptor types. A better understanding of receptors and their interactions with proteins spearheads molecular biology, informs an integral part of bench medicine which helps in drug development, drug action, and understanding the pathophysiology of disease.
Crosslinking in the Analysis of Receptor Interactomes: To Link or not to Link?
To biochemically study receptor–protein interactions, one must be able to isolate receptors and their interactomes from the lipid plasma membrane. This process is challenging because of the need to keep diverse protein–protein interactions intact during the extraction and purification of the protein complex. Additionally, the structure and subcellular localization of the receptor in the membrane dictates the chemical conditions required for protein extraction (Thomas and McNamee, 1990). Large polypeptide membrane spanning receptors, such as ligand gated ion channels, demand strong detergent-based solubilization in order to ensure extraction of all receptor subunits. The stringency of these detergent conditions, however, can lead to a loss in numerous receptor–protein associations. Receptors that are embedded in lipid-rich and cholesterol heavy regions of the plasma membrane (such as rafts) require unconventional solubilization methods since these areas are resistant to detergents (Li et al., 2004; Sot et al., 2006). The isolation of membrane bound receptors and their interacting proteins is not trivial and requires extensive optimization on a receptor-by-receptor basis (Kalipatnapu and Chattopadhyay, 2005; Sarramegn et al., 2006).
Cell-based imaging strategies for the study of receptor–protein interactions, such as fluorescence resonance energy transfer (FRET), circumvent this problem by examining the interaction within an intact cell (Truong and Ikura, 2001). FRET analysis, however, only measures protein–protein interaction at relatively large distances [~100 Angstroms (Å)] and therefore may not be very informative about direct protein coupling (Pollok, 1999). Biochemical assessment of receptor–protein interaction using standard affinity purification methods alone also suffers from several drawbacks. First, the chemical stringency of the biochemical processing steps likely compromises and/or interferes with some receptor–protein interactions. Second, common affinity purification methods such as immunoprecipitation (IP) or pulldown assays heavily rely on the specificity of the antibody or capturing bait and may bias toward abundant proteins and stable protein–protein interactions. In the absence of stringent controls, standard IP experiments can produce substantial false positive results. Finally, current biochemical methods used to detect protein interactions lack cellular spatial specificity; consequently, when a true interaction is discovered the subcellular localization of the interaction is unknown.
New strategies have emerged for enhancing the detection of protein interactions. Methods such as protein fragment complementation and chemical crosslinking (CC) can stabilize transient or labile protein interactions in vivo and in vitro (Box 1), and therefore enable the identification of many proteins within the interactome (Kluger and Alagic, 2004; Morell et al., 2007). Conventionally CC has been used in the study of extracellular interactions of the receptor such as ligand binding (Gronemeyer and Govindan, 1986; Fanger et al., 1989; Boudreau et al., 2012; Kim et al., 2012), studies now reveal however a utility for cell permeable CC in the identification of the receptor interactome (Figure 1A; Guerrero et al., 2006; Nordman and Kabbani, 2012). In particular, dynamic changes in protein–protein associations within receptor interactomes appear better detected by CC at various stages of the receptor preparation and purification method (Vasilescu et al., 2004). Interactions that are generally too weak or too transient to be discovered in standard pulldown or IP assays alone, can be stabilized by covalent crosslinkers during the membrane solubilization process (Bond et al., 2009; Nordman and Kabbani, 2012). The common use of stringent chemical detergents such as radio-immunoprecipitation assay (RIPA) buffers, which interfere with many types of protein–protein interactions, can also benefit from the addition of covalent crosslinkers which are generally unperturbed by the RIPA reagent. Moreover, CC can be effectively combined with affinity purification protocols such as the IP prior to the mass spectrometry analysis (Vasilescu et al., 2004). To eliminate non-specific interactions of proteins during CC, the assay requires optimization before the start of the study. It is also not uncommon to run non-crosslinked samples in parallel during the course of a study (Kim et al., 2012).
Box 1. Technical Toolbox.
• To crosslink solubilized membrane proteins in vitro with BS3, add 2 mM BS3 to the enriched receptor fraction for 2 h at 4°C and mix (Figure 1B).
• To crosslink proteins in vivo, add 2.5 mM DSP to cultured cells for 2 h at 4°C (Figure 1C). The chemical reaction with DSP can be terminated by the addition of 50 mM Tris-HCI (pH 7.5) at 4°C.
• Clever use of DSP crosslinking enables a range of experiments on receptor–protein interactions including an analysis of receptor interactomes at the cell surface, inside recycled vesicles, or in response to ligand stimulation (Figures 2A,B).
• A crosslinked receptor interactome can be purified using standard methods for immunoprecipitation and mass spectrometry (Figure 2C).
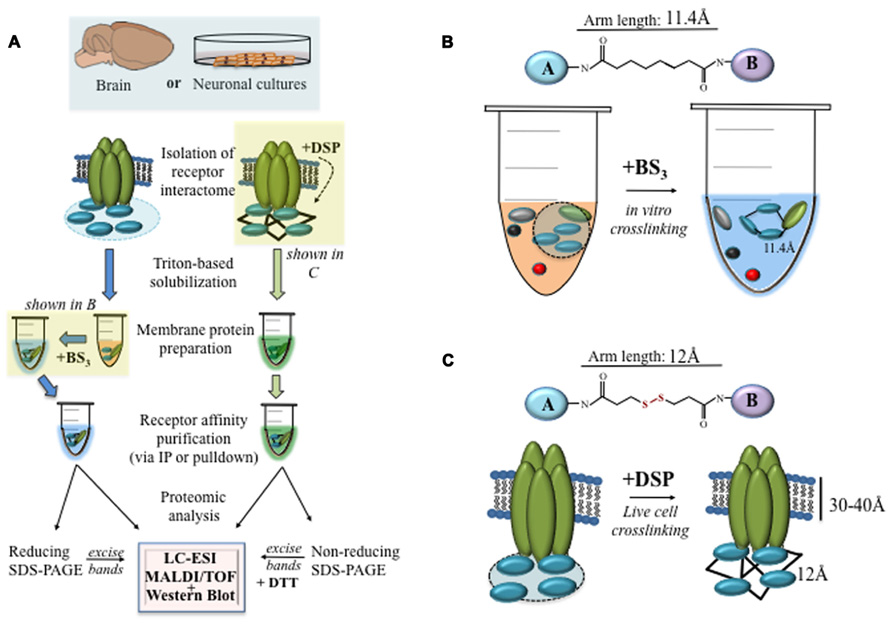
FIGURE 1. Crosslinking the nAChR interactome. (A) A flow chart showing the methods for isolation, crosslinking, and proteomic analysis of nAChR interactomes from brain tissue or neural cells. The experimental design should take into the consideration the choice of the crosslinker as well as the spacer arm. (B) The irreversible crosslinker BS3 is effective for crosslinking nAChR interactomes after membrane protein solubilization with Triton-X. (C) The membrane permeable crosslinker DSP, on the other hand, can be used to crosslink receptors and their interacting proteins in living cells.
A number of crosslinkers have been used to study receptor–protein interactions in cells (Brenner et al., 1985; Shinya et al., 2010; Miteva et al., 2013). These compounds are characterized by differences in their spacer arm as well as the composition of the two amine binding groups that recognize and covalently bind specific functional groups on target proteins (Figures 1B,C; Sinz, 2003; Trakselis et al., 2005). Table 1 lists crosslinkers that have been used to study receptor binding to intracellular proteins. The choice of a spacer arm length, between 5 and 25 Å, is experimentally important because it enables the identification of receptor–protein interactions at specific distances. A recent study utilized agarose beads whose surface was covalently linked with a cleavable chemical crosslinker by spacers of varied lengths to study the interactome of the post-synaptic density isolated from the rodent cortex (Yun-Hong et al., 2011). Experiments successfully demonstrate assemblages of proteins at various subcellular distances and compartments, including the post-synaptic density, thus underscoring the utility of the approach in the characterization of interactomes based on spacer arm properties. However, a possible disadvantage of CC is that some antibodies are no longer able to recognize their target protein after crosslinking (Boudreau et al., 2012).

TABLE 1. A summary of crosslinkers used in the analysis of receptor protein interactions.
Crosslinking Enables Detection of Proteins that Bind the Receptor in Various States
Interaction with trafficking and chaperone proteins is important for directing the localization and function of the receptor (Jeanclos et al., 2001; Lin et al., 2002; Xu et al., 2006; Kabbani et al., 2007; Nordman and Kabbani, 2012; Colombo et al., 2013). Protein associations may also contribute to receptor conformation at the cell surface (Giniatullin et al., 2005). While the ability to detect receptors at the plasma membrane and in the cytosol has been traditionally reliant on epitope tagging, live cell stain, and cell surface labeling methods such as biotinylation, CC has emerged as a complementary tool in the study of interacting proteins responsible for receptor cellular trafficking and localization. Numerous examples exist in the literature, however, work on β2 adrenergic receptor internalization has been central to understanding mechanisms in GPCR internalization (Lefkowitz and Shenoy, 2005). Key findings on β2 adrenergic receptor internalization have come from experiments using various crosslinkers to detect the association of the receptor with the endocytosis machinery of the cells. First, IP was used in conjunction with DSP [dithiobis(succinimidyl propionate)] crosslinking in human embryonic kidney (HEK) 293 cells to quantify changes in β2 adrenergic receptor internalization and demonstrate that the internalized receptor was bound to β-arrestin, which functions as an endocytic adaptor of the receptor complex (Shenoy et al., 2006). In a second similar study, also performed in HEK 293 cells that stably express the β2 adrenergic receptor, DSP was used to show that the β2 adrenergic receptor binds the trafficking and regulatory proteins β-arrestin and c-Src in its “desensitized” state (Luttrell et al., 1999). The study interestingly demonstrates that receptor signaling is sustained even in the absence of ligand binding. A third study on β2 adrenergic receptor internalization employed covalent protein crosslinking with DSP for the detection of transient, agonist-promoted association of dynamin and c-Src, showing how these protein interactions can alter the rate of receptor internalization in the cell (Ahn, 1999).
Crosslinking has also enabled detection of changes in receptor function for the angiotensin receptor (Quian et al., 2001) and has been useful in determining interactions impacted by post-translational modification (Cao et al., 1999; Connolly, 1999; Ehlers, 2000). When combined with cell surface labeling, CC has been effective in determining changes in receptor glycosylation. Differential glycosylation of cell surface human and rat (r) calcitonin (CT) receptor-like receptors (CRLR) as a result of interactions with accessory receptor activity-modifying proteins (RAMPs)-1 or -2 was confirmed by CC using BS3 [bis(sulfosuccinimidyl) suberate] in Drosophila S2 cells (Aldecoa et al., 2000). In this study CC revealed receptor components with the size of rCRLR, increased by the molecular weights of the corresponding RAMP – suggestive of a direct association between the receptor and the accessory protein during ligand activation.
Nicotinic Receptor Interactomes Defined by Crosslinking
Nicotinic acetylcholine receptors (nAChRs) are a family of ligand gated ion channels expressed throughout the nervous system contributing to learning, memory, and goal driven behavior (Changeux, 2012). Recent evidence also reveals that nAChRs operate by coupling to intracellular proteins such as heterotrimeric G proteins (Kabbani et al., 2013). Chronic nicotine exposure gives rise to neural adaptations such as an up-regulation of specific nAChRs through cell-delimited post-translational mechanisms (Sallette et al., 2005; Colombo et al., 2013). These receptor mechanisms are a hallmark of nicotine addiction yet it is still unclear which signaling pathways and mechanism regulate nAChR assembly and trafficking inside the cell. Proteomic studies, based on yeast-two-hybrid as well as conventional IP experiments have led to the identification of several intracellular proteins that bind nAChR subunits in the brain (Kabbani et al., 2007; Paulo et al., 2009; Nordman and Kabbani, 2012; McClure-Begley et al., 2013). Directed protein interaction screens have also enabled discovery of proteins responsible for nAChR trafficking and assembly (Lin et al., 2002; Lansdell et al., 2005; Kabbani, 2008; Rezvani et al., 2009).
In the hippocampus, α7 nAChRs are expressed pre- and post-synaptically, contributing to GABA and glutamate neurotransmission (Liu et al., 2006; Lozada et al., 2012). α7 receptors are also found to mediate the growth of axons (Hancock et al., 2008; Nordman and Kabbani, 2012) and dendrites (Campbell et al., 2011) in the developing hippocampus. Using the membrane impermeable and irreversible crosslinker BS3, we have defined dynamic changes in α7 interaction within solubilized membrane fractions from differentiated PC12 cells and hippocampal neurons (Figure 1B; Nordman and Kabbani, 2012). We show that α7 receptors are directly coupled to a G protein pathway consisting of Gαo, Gprin1, and GAP-43 in growing cells (Nordman and Kabbani, 2012; Figure 1C). In these studies CC was vital to the detection of changes in receptor interaction with signaling molecules and heterotrimeric G proteins. The CC method was also able to enhance the detection of small signaling molecules such as receptor kinases in both Western blots and mass spectrometry experiments (Hu et al., 2010; Nordman and Kabbani, 2012). For example, using BS3 to crosslink the α7 nAChR network after nicotine activation, we identified rapid changes to the calcium-mediated signaling pathway of the receptor, which consisted of a dynamic association between GAP-43 and calmodulin (CaM) in the growing neurite (Figure 2C; Nordman and Kabbani, 2012). In particular, activation of the α7 nAChR was found to promote a rapid association between the receptor and CaM bound GAP-43. This interaction was rapidly reversed by ligand inactivation of the α7 nAChR, showing that receptor association with CaM bound GAP-43 was driven by nAChR channel function and calcium elevation in the cell (Nordman and Kabbani, 2012). These findings on dynamic associations of CaM and GAP-43 within the α7 nAChR interactome could not have been detected using standard IP assays alone underscoring the utility of the crosslinker in the study of protein interactions under physiological conditions. In a similar study, CC with disuccinimidyl suberate (DSS) was used in identifying dynamic changes in calcium bound CaM kinase II and subunits of the NMDA glutamate receptor within the post-synaptic density of hippocampal neurons (Gardoni et al., 2002), thus underscoring the utility of the method in the study of rapid calcium driven changes in protein coupling in cells.
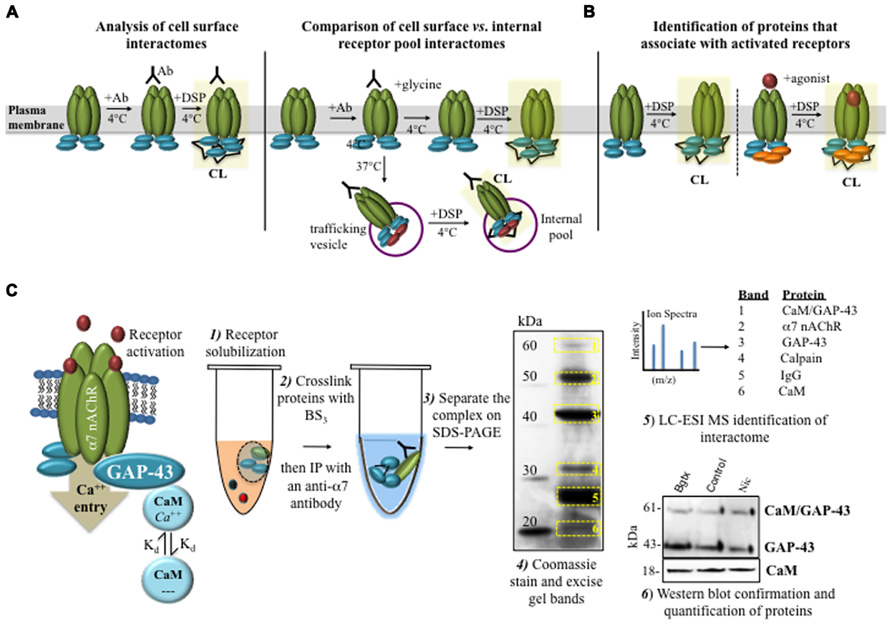
FIGURE 2. Identifying protein interactions that mediate nAChR trafficking, localization, and signaling. (A) Cell surface receptors can be selectively labeled with an anti-α7 nAChR monoclonal Ab. Cell surface labeling (at 4°C) can be combined with DSP in order to crosslink the receptor interactome. Alternatively, antibody labeling and crosslinking can be used to examine changes in the receptor interactome between internalized and cell surface nAChRs. (B) Chemical crosslinking can be used to study the dynamics of nAChR–protein interactions under various ligand treatment conditions. (C) Experimental evidence on α7 nAChR interactions with GAP-43 and CaM in developing neural cells. BS3 was used to crosslink the α7 nAChR interactome from differentiating cells. An IP was utilized to purify the receptor, which was visualized by SDS-PAGE. Protein identity was confirmed using LC-ESI MS and Western blot. These experiments demonstrate dynamic changes in CaM/GAP-43 association with α7 nAChR in response to nicotine activation (Nordman and Kabbani, 2012).
Looking Ahead
Proteomic and yeast-two-hybrid studies on receptor interactions have enabled a broad understanding on the diversity and function of receptor–protein interactions in cells. These studies have enabled an interaction-based framework for defining the mechanisms of receptor signaling. Receptor–protein interaction identification however is not sufficient for understanding how receptors operate in cells. In particular, important questions remain on the spatial specificity and temporal aspects of receptor expression and signaling in cells. For multi-subunit channel receptors such as the glutamate AMPA receptor, the addition of the membrane impermeable linker BS3 has proven effective in the analysis of receptor subunit composition at the cell surface (Boudreau et al., 2012). Similar approaches with the aim of detecting protein–protein interaction in living cells are now necessary. Advancement in the design and experimental utility of CC such as photo-reactive amino acid analogs (Suchanek et al., 2005) promises to enhance the study of receptor–protein interactions in vivo.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
This work was supported by a Wings for Life Spinal Cord Research Grant to Nadine Kabbani.
References
Ahn, S. (1999). Src-mediated tyrosine phosphorylation of dynamin is required for beta 2-adrenergic receptor internalization and mitogen-activated protein kinase signaling. J. Biol. Chem. 274, 1185–1188. doi: 10.1074/jbc.274.3.1185
Aldecoa, A., Gujer, R., Fischer, J. A., and Born, W. (2000). Mammalian calcitonin receptor-like receptor/receptor activity modifying protein complexes define calcitonin gene-related peptide and adrenomedullin receptors in Drosophila Schneider 2 cells. FEBS Lett. 471, 156–160. doi: 10.1016/S0014-5793(00)01387-9
Bond, C. E., Zimmermann, M., and Greenfield, S. A. (2009). Upregulation of alpha7 nicotinic receptors by acetylcholinesterase c-terminal peptides. PLoS ONE 4:e4846. doi: 10.1371/journal.pone.0004846
Boudreau, A. C., Milovanovic, M., Conrad, K. L., Nelson, C., Ferrario, C. R., and Wolf, M. E. (2012). A protein cross-linking assay for measuring cell surface expression of glutamate receptor subunits in the rodent brain after in vivo treatments. Curr. Protoc. Neurosci. Chapter 5, Unit 5.30.1–19. doi: 10.1002/0471142301.ns0530s59
Brenner, M. B., Trowbridge, I. S., and Strominger, J. L. (1985). Cross-linking of human T cell receptor proteins: association between the T cell idiotype β subunit and the T3 glycoprotein heavy subunit. Cell 40, 183–190. doi:10.1016/0092-8674(85)90321-6
Campbell, N. R., Fernandes, C. C., John, D., Lozada, A. F., and Berg, D. K. (2011). Nicotinic control of adult-born neuron fate. Biochem. Pharmacol. 82, 820–827. doi: 10.1016/j.bcp.2011.06.021
Cao, T. T., Deacon, H. W., Reczek, D., Bretscher, A., and von Zastrow, M. (1999). A kinase-regulated PDZ-domain interaction controls endocytic sorting of the beta2-adrenergic receptor. Nature 401, 286–290. doi: 10.1038/45816
Changeux, J.-P. (2012). The nicotinic acetylcholine receptor: the founding father of the pentameric ligand-gated ion channel superfamily. J. Biol. Chem. 287, 40207–40215. doi: 10.1074/jbc.R112.407668
Chu, F., Shan, S., Moustakas, D. T., Alber, F., Egea, P. F., Stroud, R. M., et al. (2004). Unraveling the interface of signal recognition particle and its receptor by using chemical cross-linking and tandem mass spectrometry. Proc. Natl. Acad. Sci. U.S.A. 101, 16454–16459. doi: 10.1073/pnas.0407456101
Colombo, S. F., Mazzo, F., Pistillo, F., and Gotti, C. (2013). Biogenesis, trafficking and up-regulation of nicotinic ACh receptors. Biochem. Pharmacol. 86, 1063–1073. doi:10.1016/j.bcp.2013.06.023
Connolly, C. N. (1999). Cell surface stability of gamma-aminobutyric acid type A receptors. Dependence on protein kinase C activity and subunit composition. J. Biol. Chem. 274, 36565–36572. doi: 10.1074/jbc.274.51.36565
Ehlers, M. D. (2000). Reinsertion or degradation of AMPA receptors determined by activity-dependent endocytic sorting. Neuron 28, 511–525. doi: 10.1016/S0896-6273(00)00129-X
Fanger, B. O., Stephens, J. E., and Staros, J. V. (1989). High-yield trapping of EGF-induced receptor dimers by chemical cross-linking. FASEB J. 3, 71–75.
Gardoni, F., Caputi, A., Cimino, M., Pastorino, L., Cattabeni, F., and Di Luca, M. (2002). Calcium/calmodulin-dependent protein kinase II is associated with NR2A/B subunits of NMDA receptor in postsynaptic densities. J. Neurochem. 71, 1733–1741. doi: 10.1046/j.1471-4159.1998.71041733.x
Giniatullin, R., Nistri, A., and Yakel, J. L. (2005). Desensitization of nicotinic ACh receptors: shaping cholinergic signaling. Trends Neurosci. 28, 371–378. doi:10.1016/j.tins.2005.04.009
Gronemeyer, H., and Govindan, M. V. (1986). Affinity labelling of steroid hormone receptors. Mol. Cell. Endocrinol. 46, 1–19. doi: 10.1016/0303-7207(86)90064-X
Guerrero, C., Tagwerker, C., Kaiser, P., and Huang, L. (2006). An integrated mass spectrometry-based proteomic approach: quantitative analysis of tandem affinity-purified in vivo cross-linked protein complexes (QTAX) to decipher the 26 S proteasome-interacting network. Mol. Cell. Proteomics 5, 366–378. doi: 10.1074/mcp.M500303-MCP200
Hancock, M. L., Canetta, S. E., Role, L. W., and Talmage, D. A. (2008). Presynaptic type III neuregulin1-ErbB signaling targets {alpha}7 nicotinic acetylcholine receptors to axons. J. Cell Biol. 181, 511–521. doi: 10.1083/jcb.200710037
Hu, J., Wang, Y., Zhang, X., Lloyd, J. R., Li, J. H., Karpiak, J., et al. (2010). Structural basis of G protein-coupled receptor-G protein interactions. Nat. Chem. Biol. 6, 541–548. doi: 10.1038/nchembio.385
Jeanclos, E. M., Lin, L., Treuil, M. W., Rao, J., DeCoster, M. A., and Anand, R. (2001). The chaperone protein 14-3-3eta interacts with the nicotinic acetylcholine receptor alpha 4 subunit. Evidence for a dynamic role in subunit stabilization. J. Biol. Chem. 276, 28281–28290. doi: 10.1074/jbc.M011549200
Kabbani, N. (2008). Proteomics of membrane receptors and signaling. Proteomics 8, 4146–4155. doi: 10.1002/pmic.200800139
Kabbani, N., Nordman, J. C., Corgiat, B. A., Veltri, D. P., Shehu, A., Seymour, V. A., et al. (2013). Are nicotinic acetylcholine receptors coupled to G proteins? Bioessays 35, 1025–1034. doi: 10.1002/bies.201300082
Kabbani, N., Woll, M. P., Levenson, R., Lindstrom, J. M., and Changeux, J.-P. (2007). Intracellular complexes of the beta2 subunit of the nicotinic acetylcholine receptor in brain identified by proteomics. Proc. Natl. Acad. Sci. U.S.A. 104, 20570–20575. doi: 10.1073/pnas.0710314104
Kalipatnapu, S., and Chattopadhyay, A. (2005). Membrane protein solubilization: recent advances and challenges in solubilization of serotonin1A receptors. IUBMB Life 57, 505–512. doi: 10.1080/15216540500167237
Kim, K. M., Yi, E. C., and Kim, Y. (2012). Mapping protein receptor-ligand interactions via in vivo chemical crosslinking, affinity purification, and differential mass spectrometry. Methods 56, 161–165. doi: 10.1016/j.ymeth.2011.10.013
Kluger, R., and Alagic, A. (2004). Chemical cross-linking and protein-protein interactions-a review with illustrative protocols. Bioorg. Chem. 32, 451–472. doi: 10.1016/j.bioorg.2004.08.002
Lansdell, S. J., Gee, V. J., Harkness, P. C., Doward, A. I., Baker, E. R., Gibb, A. J., et al. (2005). RIC-3 enhances functional expression of multiple nicotinic acetylcholine receptor subtypes in mammalian cells. Mol. Pharmacol. 68, 1431–1438. doi: 10.1124/mol.105.017459
Lau, A. G., and Hall, R. A. (2001). Oligomerization of NHERF-1 and NHERF-2 PDZ domains: differential regulation by association with receptor carboxyl-termini and by phosphorylation. Biochemistry 40, 8572–8580. doi: 10.1021/bi0103516
Lefkowitz, R. J., and Shenoy, S. K. (2005). Transduction of receptor signals by beta-arrestins. Science 308, 512–517. doi: 10.1126/science.1109237
Li, N., Shaw, A. R. E., Zhang, N., Mak, A., and Li, L. (2004). Lipid raft proteomics: analysis of in-solution digest of sodium dodecyl sulfate-solubilized lipid raft proteins by liquid chromatography-matrix-assisted laser desorption/ionization tandem mass spectrometry. Proteomics 4, 3156–3166. doi: 10.1002/pmic.200400832
Lin, L., Jeanclos, E. M., Treuil, M., Braunewell, K.-H., Gundelfinger, E. D., and Anand, R. (2002). The calcium sensor protein visinin-like protein-1 modulates the surface expression and agonist sensitivity of the alpha 4beta 2 nicotinic acetylcholine receptor. J. Biol. Chem. 277, 41872–41878. doi: 10.1074/jbc.M206857200
Liu, Z., Neff, R. A., and Berg, D. K. (2006). Sequential interplay of nicotinic and GABAergic signaling guides neuronal development. Science 314, 1610–1613. doi: 10.1126/science.1134246
Lozada, A. F., Wang, X., Gounko, N. V., Massey, K. A., Duan, J., Liu, Z., et al. (2012). Glutamatergic synapse formation is promoted by α7-containing nicotinic acetylcholine receptors. J. Neurosci. 32, 7651–7661. doi: 10.1523/JNEUROSCI.6246-11.2012
Luttrell, L. M., Ferguson, S. S. G., Daaka, Y., Miller, W. E., Maudsley, S., Della Rocca, G. J., et al. (1999). β-arrestin-dependent formation of β2 adrenergic receptor-src protein kinase complexes. Science 283, 655–661. doi: 10.1126/science.283.5402.655
McClure-Begley, T. D., Stone, K. L., Marks, M. J., Grady, S. R., Colangelo, C. M., Lindstrom, J. M., et al. (2013). Exploring the nicotinic acetylcholine receptor-associated proteome with iTRAQ and transgenic mice. Genomics Proteomics Bioinformatics 11, 218–207. doi: 10.1016/j.gpb.2013.05.005
Miteva, Y. V., Budayeva, H. G., and Cristea, I. M. (2013). Proteomics-based methods for discovery, quantification, and validation of protein-protein interactions. Anal. Chem. 85, 749–768. doi: 10.1021/ac3033257
Morell, M., Espargaró, A., Avilés, F. X., and Ventura, S. (2007). Detection of transient protein-protein interactions by bimolecular fluorescence complementation: the Abl-SH3 case. Proteomics 7, 1023–1036. doi: 10.1002/pmic.200600966
Nordman, J. C., and Kabbani, N. (2012). An interaction between α7 nicotinic receptors and a G-protein pathway complex regulates neurite growth in neural cells. J. Cell Sci. 125, 5502–5513. doi: 10.1242/jcs.110379
Paulo, J. A., Brucker, W. J., and Hawrot, E. (2009). Proteomic analysis of an alpha7 nicotinic acetylcholine receptor interactome. J. Proteome Res. 8, 1849–1858. doi: 10.1021/pr800731z
Pollok, B. (1999). Using GFP in FRET-based applications. Trends Cell Biol. 9, 57–60. doi: 10.1016/S0962-8924(98)01434–1432
Quian, H., Pipolo, L., and Thomas, W. G. (2001). Association of -arrestin 1 with the type 1A angiotensin II receptor involves phosphorylation of the receptor carboxyl terminus and correlates with receptor internalization. J. Mol. Endocrinol. 15, 1706–1719. doi: 10.1210/me.15.10.1706
Rezvani, K., Teng, Y., Pan, Y., Dani, J. A., Lindstrom, J., García Gras, E. A., et al. (2009). UBXD4, a UBX-containing protein, regulates the cell surface number and stability of alpha3-containing nicotinic acetylcholine receptors. J. Neurosci. 29, 6883–6896. doi: 10.1523/JNEUROSCI.4723-08.2009
Sallette, J., Pons, S., Devillers-Thiery, A., Soudant, M., Prado de Carvalho, L., Changeux, J.-P., et al. (2005). Nicotine upregulates its own receptors through enhanced intracellular maturation. Neuron 46, 595–607. doi: 10.1016/j.neuron.2005.03.029
Sarramegn, V., Muller, I., Milon, A., and Talmont, F. (2006). Recombinant G protein-coupled receptors from expression to renaturation: a challenge towards structure. Cell. Mol. Life Sci. 63, 1149–1164. doi: 10.1007/s00018-005-5557-6
Shenoy, S. K., Drake, M. T., Nelson, C. D., Houtz, D. A., Xiao, K., Madabushi, S., et al. (2006). beta-arrestin-dependent, G protein-independent ERK1/2 activation by the beta2 adrenergic receptor. J. Biol. Chem. 281, 1261–1273. doi: 10.1074/jbc.M506576200
Shinya, T., Osada, T., Desaki, Y., Hatamoto, M., Yamanaka, Y., Hirano, H., et al. (2010). Characterization of receptor proteins using affinity cross-linking with biotinylated ligands. Plant Cell Physiol. 51, 262–270. doi: 10.1093/pcp/pcp185
Sinz, A. (2003). Chemical cross-linking and mass spectrometry for mapping three-dimensional structures of proteins and protein complexes. J. Mass Spectrom. 38, 1225–1237. doi: 10.1002/jms.559
Sot, J., Bagatolli, L. A., Goñi, F. M., and Alonso, A. (2006). Detergent-resistant, ceramide-enriched domains in sphingomyelin/ceramide bilayers. Biophys. J. 90, 903–914. doi: 10.1529/biophysj.105.067710
Suchanek, M., Radzikowska, A., and Thiele, C. (2005). Photo-leucine and photo-methionine allow identification of protein-protein interactions in living cells. Nat. Methods 2, 261–268. doi: 10.1038/nmeth752
Thomas, T. C., and McNamee, M. G. (1990). Purification of membrane proteins. Methods Enzymol. 182, 499–520. doi: 10.1016/0076-6879(90)82040-9
Trakselis, M. A., Alley, S. C., and Ishmael, F. T. (2005). Identification and mapping of protein-protein interactions by a combination of cross-linking, cleavage, and proteomics. Bioconjug. Chem. 16, 741–750. doi: 10.1021/bc050043a
Truong, K., and Ikura, M. (2001). The use of FRET imaging microscopy to detect protein–protein interactions and protein conformational changes in vivo. Curr. Opin. Struct. Biol. 11, 573–578. doi: 10.1016/S0959-440X(00)00249-9
Vasilescu, J., Guo, X., and Kast, J. (2004). Identification of protein-protein interactions using in vivo cross-linking and mass spectrometry. Proteomics 4, 3845–3854. doi: 10.1002/pmic.200400856
Wang, X., DeFrances, M. C., Dai, Y., Pediaditakis, P., Johnson, C., Bell, A., et al. (2002). A mechanism of cell survival: sequestration of Fas by the HGF receptor Met. Mol. Cell 9, 411–421. doi: 10.1016/S1097-2765(02)00439-2
Xu, J., Zhu, Y., and Heinemann, S. F. (2006). Identification of sequence motifs that target neuronal nicotinic receptors to dendrites and axons. J. Neurosci. 26, 9780–9793. doi: 10.1523/JNEUROSCI.0840-06.2006
Keywords: nicotinic acetylcholine receptor, chemical crosslinking, mass spectrometry, protein–protein interaction, signaling network, interactome
Citation: Corgiat BA, Nordman JC and Kabbani N (2014) Chemical crosslinkers enhance detection of receptor interactomes. Front. Pharmacol. 4:171. doi: 10.3389/fphar.2013.00171
Received: 16 October 2013; Accepted: 17 December 2013;
Published online: 07 January 2014.
Edited by:
Ruud Hovius, École Polytechnique Fédérale de Lausanne, SwitzerlandReviewed by:
Asha Suryanarayanan, Manchester College School of Pharmacy, USAGopalkumar Rakesh, National Institute of Mental Health and Neurosciences, India
Copyright © 2014 Corgiat, Nordman and Kabbani. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nadine Kabbani, Department of Molecular Neuroscience, Krasnow Institute for Advanced Study, George Mason University, 4400 University Drive, Mail Stop 2A1, Fairfax, VA 22030, USA e-mail:bmthYmJhbmlAZ211LmVkdQ==
† Brian A. Corgiat and Jacob C. Nordman have contributed equally to this work.