 Sandeep Gawdiya
Sandeep Gawdiya Dinesh Kumar
Dinesh Kumar Ramandeep Kumar Sharma
Ramandeep Kumar Sharma- 1School of Agriculture, Galgotias University, Greater Noida, Uttar Pradesh, India
- 2Division of Agronomy; Indian Council of Agricultural Research (ICAR)-Indian Agricultural Research Institute (IARI), New Delhi, India
- 3Rutgers, The State University of New Jersey, New Brunswick, NJ, United States
Nutrients uptake by plants from the soil depends on the fertilizers applied, the physical and chemical properties of the soil, and various environmental and biological factors. Each nutrients have a positive or negative interaction with other nutrients in terms of their availability in the soil and their uptake in plants. The purpose of this study is to investigate the effects of successive increases in nitrogen (N) fertilizers on macronutrient uptake, system productivity (SP), and wheat equivalent yield (WEY) by wheat. This study was carried out in a split plot design with three distinct N input (N0, N75, and N150) in the main plot and ten wheat cultivars in the sub-plot over two consecutive years (2020-21 and 2021-22) in New Delhi, India. The highest SP of 9.85 t/ha-1, P & K uptake in grain (PUG) of 21.6 & 23.8 kg/ha-1, straw (PUS) of 13 & 106.4 kg/ha-1, and total phosphorus uptake (TPU) of 34.6 & 130.4 kg/ha-1 were obtained by the ‘HD 3249’ cultivar, followed by ‘HD 3117’. The application of N75 and N150 increased SP by 57.9% and 99.2%, WEY by 45.2% and 61.5%, PUG by 105.2% and 227%, PUS by 94% and 182%, and TPU by 100.5% and 208.7%, respectively, over N0. The study findings indicate that N fertilization positively influences macronutrient uptake in wheat, with cultivars ‘HD 3249’ and ‘HD 3117’ emerging as efficient candidates for optimizing macronutrient utilization. These cultivars hold significant potential for wheat breeding programs aimed at enhancing nutrient uptake while maintaining system productivity. Furthermore, incorporating biological nitrification inhibition traits into these cultivars is recommended to develop climate-smart wheat varieties.
Introduction
Wheat ranks as the second most important staple food crop globally (Sendhil et al., 2022). Since the Green Revolution era, the adoption and improvement of high-yielding wheat varieties, combined with the widespread use of synthetic nitrogen (N) fertilizers, have significantly enhanced productivity over the past five decades (John and Babu, 2021; Hamdan et al., 2022). Nitrogen inputs are central to plant growth, development, and overall productivity in agroecosystems (Zayed et al., 2023a). However, declining N sequestration in soils reduces N availability, making them insufficient to sustain higher yields in improved wheat cultivars. Unfortunately, over half of the N fertilizer used in agri-food systems becomes immobilized in the soil or lost to the environment (Anas et al., 2020; Menegaz et al., 2025). This results in a significant gap of approximately 50% between N losses and its efficient utilization in agroecosystems (Govindasamy et al., 2023). This significantly contributes to soil reactive nitrogen (Nr) losses, which in turn lead to negative environmental consequences that drive climate change and nutrient imbalances that contribute to biodiversity decline (Compton et al., 2011; Liu et al., 2018). Hence, the careful and efficient application of N fertilizer is crucial for maximizing productivity while maintaining long-term sustainability in agroecosystems (Govindasamy et al., 2023; Swathy et al., 2024).
In the global context, wheat-based cropping systems rank as the second-largest consumer of N, accounting for approximately 18% of total N fertilizer consumption among all crops (Ladha et al., 2016; Fernanda et al., 2022). Similarly, in India, wheat is one of the highest N-consuming crops. The N budget of Indian croplands has increased from 4.87 million tons (Mt) in 1961 to 24.08 Mt in 2017 (Velayudhan et al., 2024). As fertilizer prices rise relative to purchasing power parity, these places increasing pressure on the Indian economy. Additionally, excessive N use continues to result in low nitrogen use efficiency (NUE), leading to negative environmental consequences through multiple pathways such as leaching, volatilization, and denitrification (Anas et al., 2020; Yadav et al., 2023). The NUE of wheat in India ranges between 32–35%, primarily due to continuously increasing subsidies and the imbalanced use of fertilizer nutrients (Singh, 2022; Sapkota and Takele, 2023). Therefore, there is an urgent need to implement targeted agronomic interventions to enhance NUE. This can be achieved by identifying high-NUE cultivars and developing N-efficient wheat varieties with biological nitrification inhibition (BNI) properties, which should be a key research priority (Gawdiya and Kumar, 2025).
Global researchers have proposed various strategies to mitigate soil reactive nitrogen (Nr) losses and improve NUE. However, while many of these strategies aim to reduce specific Nr components, they often inadvertently lead to the release of other N forms or increase trace gas emissions (Xia et al., 2018; Zhang et al., 2022). To date, enhancing NUE through screened cultivars has proven to be an effective approach up to a certain level. However, beyond this, breeding N-efficient cultivars presents a viable strategy to overcome existing limitations and achieve further improvements in NUE (Gawdiya et al., 2023c). This climate-smart approach has gained momentum in the current scenario (Raghuram and Sharma, 2019) by offering a “triple-win” trade-off—enhancing yields, reducing N losses, and improving macronutrient uptake. Screening cultivars based on their field performance and identifying key physiological and biochemical processes that regulate N utilization are essential steps toward improving NUE (Flores-Saavedra et al., 2024; Gawdiya and Kumar, 2025). Synchronizing N supply with crop demand and optimizing biological N transformations in soils under various N-smart approaches are crucial for improving utilization of N (Lam et al., 2022; Govindasamy et al., 2023). The NUE is regulated by a complex interplay of N-responsive mechanisms and biogeochemical processes, which must be fully characterized to develop context-dependent strategies capable of improving NUE beyond 50% (Mandal et al., 2022; Gawdiya et al., 2023c).
Macronutrients are the primary limiting factors regulating plant growth and development (Sarmiento et al., 2006; Elser et al., 2007; Agren et al., 2012; Touhami et al., 2022; Ye et al., 2022). The synergistic interactions among macronutrients at optimal N rates are well-documented. In both soil and plants, these interactions depend on specific N:P/K ratios, which play a critical role in facilitating metabolic reactions and synthesizing essential compounds (Yuan and Chen, 2015a; Jiang et al., 2019). Multiple studies have reported that optimal N fertilization stimulates root growth, reduces soil nitrate-N accumulation, improves nutrient distribution, and enhances macronutrient uptake in wheat (Reganold, 1995; Hao et al., 2005; Kaiser et al., 2010; Wen et al., 2016; Kabato et al., 2022). Therefore, determining the optimal N dose is crucial for enhancing nutrient uptake and agroecosystem productivity (Roy et al., 2006). However, the low availability of macronutrients in the soil, coupled with the misapplication of N fertilizers, disrupts nutrient balance, compromising soil fertility and crop yields while posing significant environmental challenges (Asner et al., 2001; He et al., 2002; Vargas et al., 2022; Zhang et al., 2025). To address these issues, we evaluated the performance of ten wheat cultivars under graded N levels over two growing seasons. The study aimed to assess the effects of N levels on wheat equivalent yield, system productivity, and macronutrient uptake in different cultivars.
Materials and methods
Site description
Field experiments were conducted over two consecutive years (2020–2021 and 2021–2022, corresponding to the first and second winter seasons) at the research farm located in Block 14C MB of the ICAR-Indian Agricultural Research Institute, New Delhi, India (28°38′ N, 77°10′ E). Prior to the on-station genotype evaluation experiment, soil samples were collected from the top 0.02 m of the soil profile for initial characterization. The experimental site features sandy clay loam soil (USDA soil taxonomy) with a pH of 7.8 (1:2.5, soil:water), 0.4% organic carbon (Walkley and Black, 1934), 221 kg ha-¹ of available nitrogen (Subbiah and Asija, 1956), 15.23 kg ha-¹ of Olsen phosphorus (Olsen, 1954), and 252.2 kg ha-¹ of available potassium (Hanway, 1952). The field conditions were characterized by a cool, temperate climate. The recorded average maximum and minimum temperatures varied from 27.1°C and 10.6°C in the first season to 26.9°C and 11.5°C in the second season, with an overall temperature range of 17–38°C (maximum) and 7–17°C (minimum). The average rainfall during the cropping season was 74.3 mm in the first season and 181.5 mm in the second season (Figures 1, 2).
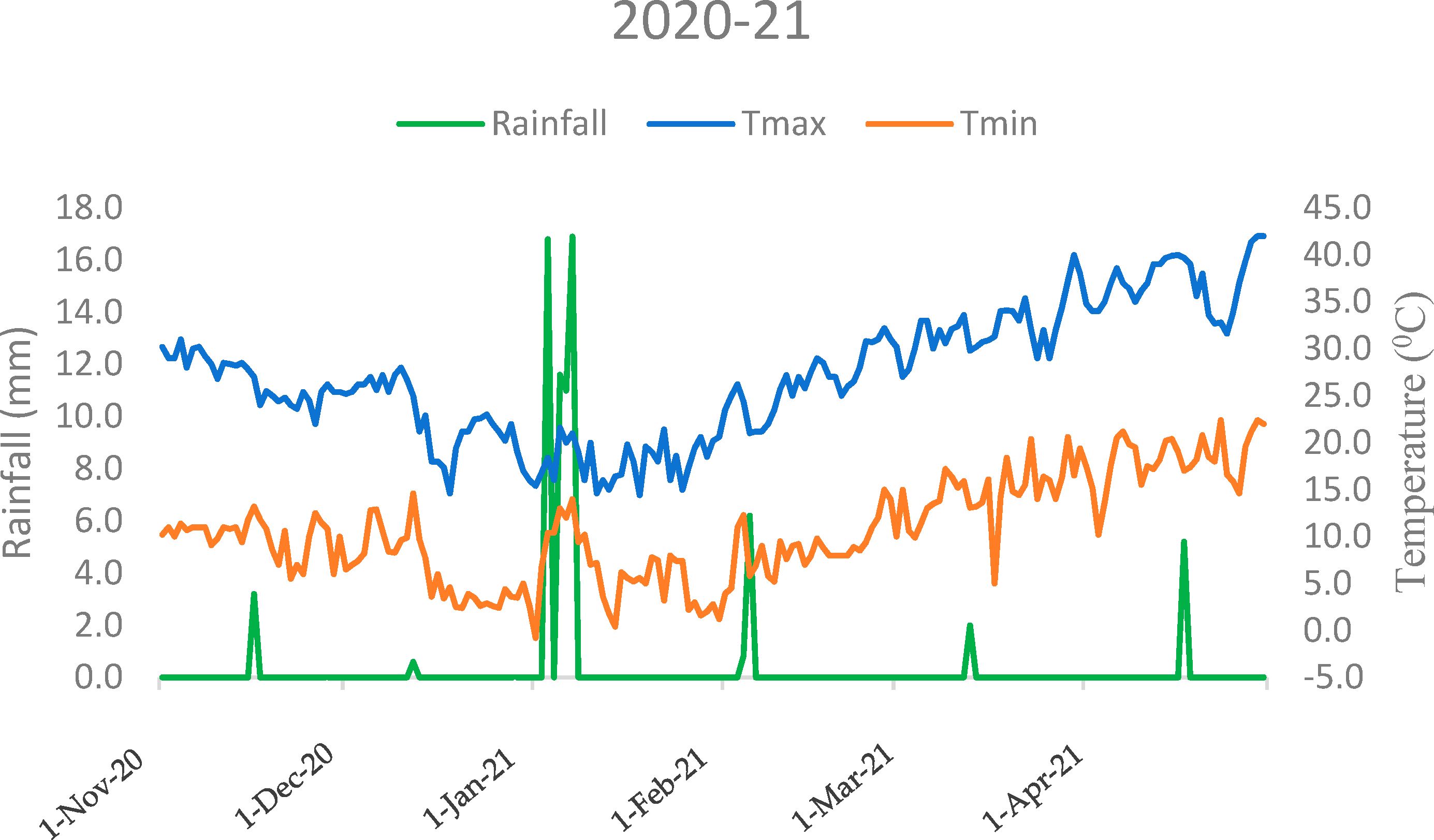
Figure 1. Seasonal trends in mean temperature and rainfall: monthly variations during the crop growing period (2020–21) at ICAR-IARI, New Delhi (Nov, November; Dec, December; Jan, January; Feb, February; Mar, March; Apr, April).

Figure 2. Seasonal trends in mean temperature and rainfall: monthly variations during the crop growing period (2021–22) at ICAR-IARI, New Delhi (Nov, November; Dec, December; Jan, January; Feb, February; Mar, March; Apr, April).
Design of experiment and crop management
The experiment was conducted in a split-plot design with three distinct nitrogen (N) input: N0 (0 kg N/ha-1), N75 (75 kg N/ha-1), and N150 (150 kg N/ha-1-recommended dose of N) are in main plots and ten different wheat cultivars (HD-3226, HDCSW-18, HD-2967, HD-3086, HD-3249, HD-2733, PBW-550, PBW-343, HD-3117, HD-3298) were grown in subplots. The plot size was 5 m × 2.5 m, and seed sowing was done with a row-to-row spacing of 22.5 cm and a plant-to-plant spacing of 5 cm. In the experimental field, a basal application of 60 kg P2O5/ha-1 (through single super phosphate) and 60 kg K2O/ha-1 (through muriate of potash) was applied uniformly. However, N was applied in three phases: an initial half dose as the basal application, followed by 25% during the crown root initiation stage (20-25 days after sowing, DAS) and the remaining 25% during the tillering stage (40-45 DAS) of the wheat crop. The source of N input was Neem-oil-coated urea (containing 46.6% N content; one ton of neem-coated urea contained 0.5 kg of neem oil). Irrigation was applied using the flooding method to ensure adequate soil moisture for crop growth. Further details about the experimental protocols and practices followed in the study are provided in Gawdiya et al. (2023 a, b, c, d).
Chemical analysis of phosphorus and potassium content in plant samples
At the harvest stage, wheat samples were collected from each plot for analysis of phosphorus (P) and potassium (K). For the estimation of P in plant samples, a concentrated nitric acid predigestion protocol was used in a 0.5 g plant sample (grain or straw) over an extended duration of overnight incubation. The sample was treated with 5 ml of diacid mixture (9:4; HNO3: HCIO4) until a white residue was produced by means of the digestion process. The resulting residue was solubilized in 6 N hydrochloric acid and subsequently transferred to a 100 ml volumetric flask, which was then diluted with distilled water. This solution was utilized to quantify the concentration of P in the plant samples. A simultaneous blank sample was also prepared. The P content within the plant sample was assessed using a spectrophotometer, using the yellow colorimetric technique (Jackson, 1973). Similarly, in plant digest K content in plant tissues was determined by using flame photometer (Jackson, 1973).
Calculation of wheat equivalent yield and system productivity
Wheat equivalent yield (WEY- t ha-1) = (rice yield (t/ha) × MSP of rice crop (INR/t)/MSP of wheat crop (INR/t)
MSP = market sale price.
System productivity (t ha-1) = wheat yield (t ha-1) + [rice yield (t ha-1) + MSP of rice (t ha-1)/MSP of wheat (t ha-1)]
In the kharif season, rice cultivars grown on plots are replaced with wheat cultivars during the winter season. Instead of a plot, rice cultivars sown are in kharif that plot wheat cultivars sown are G1-Tella Hamsa/HD-3226, G2-Vasumati/HDCSW-18, G3-VL Dhan-209/HD-2967, G4-Daya/HD-3086, G5-PB1728/HD-3249, G6-Anjali/HD-2733, G7-Heera/PBW-550, G8-Birupa/PBW-343, G9-Nagina-22/HD-3117’ G10-Nidhi/HD-3298.
Data analysis
The statistical analysis was performed using the open-source R programming language in R Studio. The agricolae package in R (Mendiburu, 2021) was utilized for data analysis, including analysis of variance (ANOVA) to assess treatment effects. Multiple mean comparisons were performed using Duncan’s Multiple Range Test (DMRT) at a significance level (α = 0.05) to determine statistically significant differences among treatments.
Results
Wheat equivalent yield and system productivity
The two-year pooled analysis data shows that wheat equivalent yield (WEY) was significantly influenced by cultivars (G) and nitrogen (N) input (Table 1). The highest WEY was observed in cultivars ‘HD 3298’ (5.53 t/ha) and ‘HD 3086’ (5.05 t/ha). The order of cultivars significantly decreasing WEY are HD 3298 > HD 3086 > PBW 343 ≥ HD 3249 > HDCSW 18 ≥ HD 3117 > HD 2967 ≥ HD 3226 ≥ PBW 550 > HD 2733. Similarly, the interaction of G×N on system productivity (SP) was significant (Table 2). Among all cultivars, the highest SP was observed in cultivar HD 3298’ (9.85 t/ha) and ‘HD 3086’ (9.69 t/ha), and the second-best cultivar was ‘HD-3117’. The three N inputs, ‘HD-3249’, ‘HD-3117’, and ‘HD-3298’, performed well compared to other cultivars. The order of cultivars in terms of decreasing SP was HD 3249 ≥ HD 3298 ≥ HD 3086 > HD 3117 > PBW 550 ≥ HD 2967 ≥ PBW 343 ≥ HDCSW 18 ≥ HD 2733 > HD 3226.
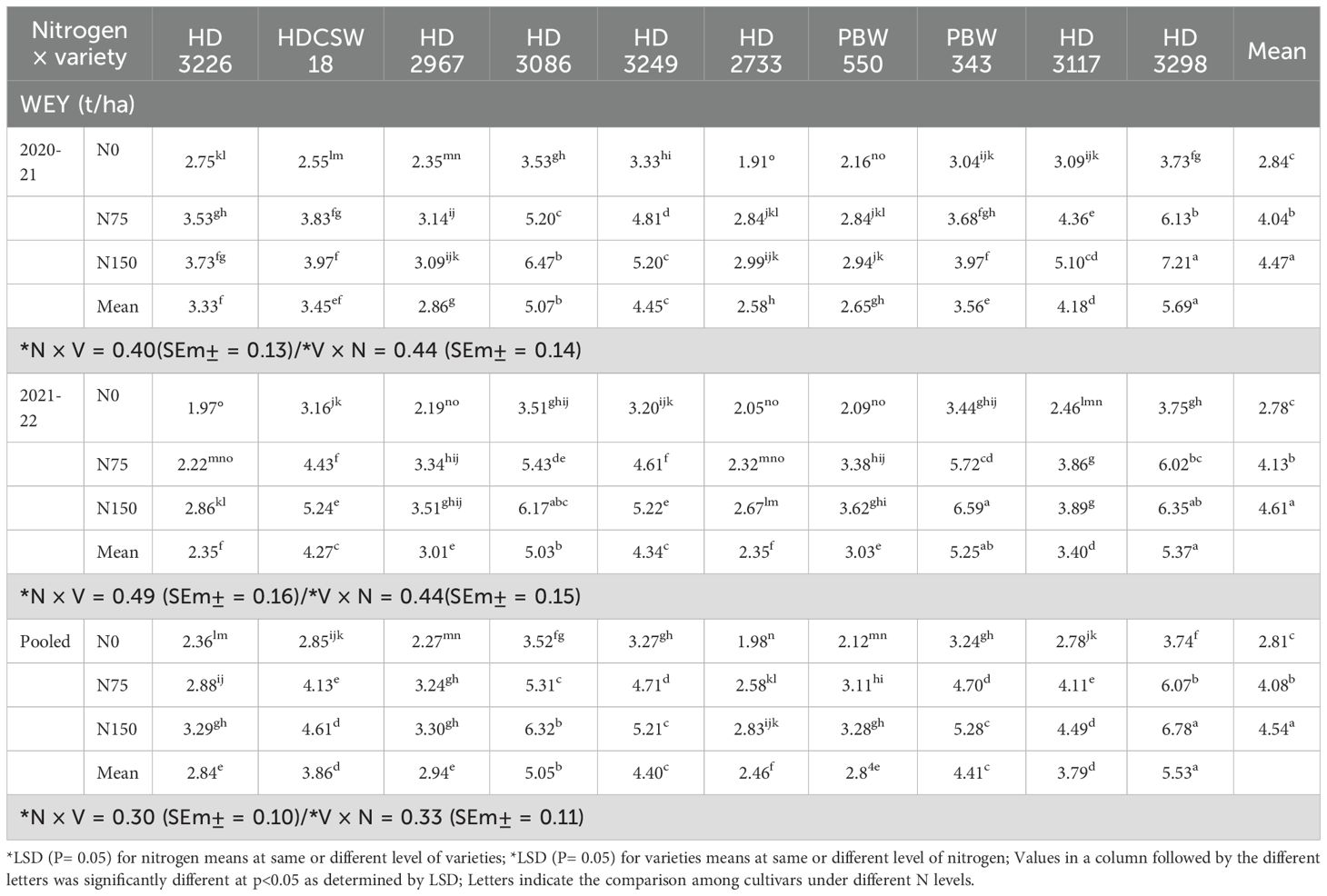
Table 1. Effect of nitrogen × cultivar interaction on wheat equivalent yield (WEY).
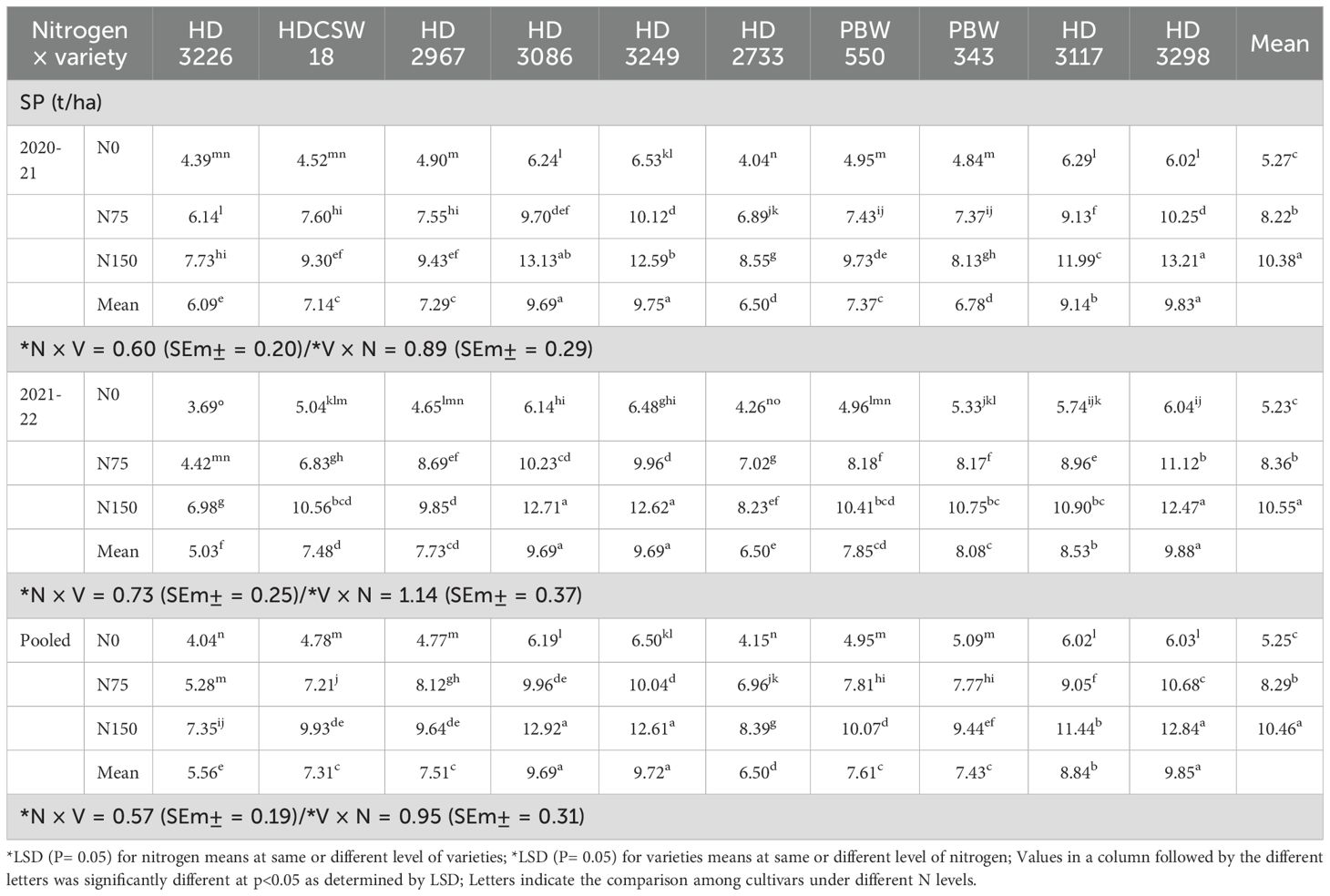
Table 2. Effect of nitrogen × cultivar interaction on wheat system productivity (SP).
Phosphorus and potassium concentration and uptake
The two-year pooled data shows significant interactions between N inputs and cultivars for P uptake in grain (PUG), straw (PUS), and total P uptake (TPU), but in case of percentage P content in grain (PPCG) and straw (PPCS) was not significant (Figures 3–8). Among all cultivars with the highest PPCG, PPCS was observed in cultivar ‘HD 3249’, which is statistically on par with ‘HD 3117’. Similarly, the highest mean PUG was observed in cultivar ‘HD 3239’ and the second-best cultivar was ‘HD 3117’, but in the case of PUS and TPU the highest value was observed in cultivar ‘HD 3249’, which is equal to ‘HD 3117’ and the second-best cultivar was ‘HD 3298’. The order of the cultivars in decreasing PUG and PUS was as follows: HD 3249 > HD 3117 > HD 3086 > PBW 550 ≥ HD 3298 ≥ HD 2733 ≥ HD 2967 ≥ HDCSW 18 > PBW 343 > HD 3226 and HD 3249 ≥ HD 3117 > HD 3298 ≥ HD 2967 ≥ HD 2733 ≥ HD 3086 ≥ PBW 550 ≥ HDCSW 18 ≥ PBW 343 > HD 3226. Similarly, the order of cultivars with significantly decreasing TPU was as follows: HD 3249 ≥ HD 3117 > HD 3298 ≥ HD 2733 ≥ HD 3086 ≥ HD 2967 ≥ PBW 550 > HDCSW 18 ≥ PBW 343 > HD 3226.The data pertaining to potassium (K) concentration in grain, straw, and total K uptake was available in Tables 3–6. Different N levels significantly influenced K concentration and uptake in grain and straw. The highest percentage K concentration in grain was observed in two cultivars ‘HD 3249’ (0.429%), and ‘HD 3117’ (0.427%) and these two cultivars were at par with cultivar ‘HD 3298’. Similarly, the highest percentage of K concentration in straw was observed in cultivars ‘HD 3249’ (1.52%), and ‘HD 3117’ (1.52%). Both the year’s highest K uptake in grain and straw were observed in cultivar ‘HD 3249’ (23.8 and 23.9 kg ha-1), (106.7 kg ha-1). Similarly, the highest total K uptake was observed in cultivar ‘HD 3249’ (130.3 and 103.5 kg ha-1). The orders of cultivars decreasing uptake of K in grain and straw are same as trends follows on phosphorus.
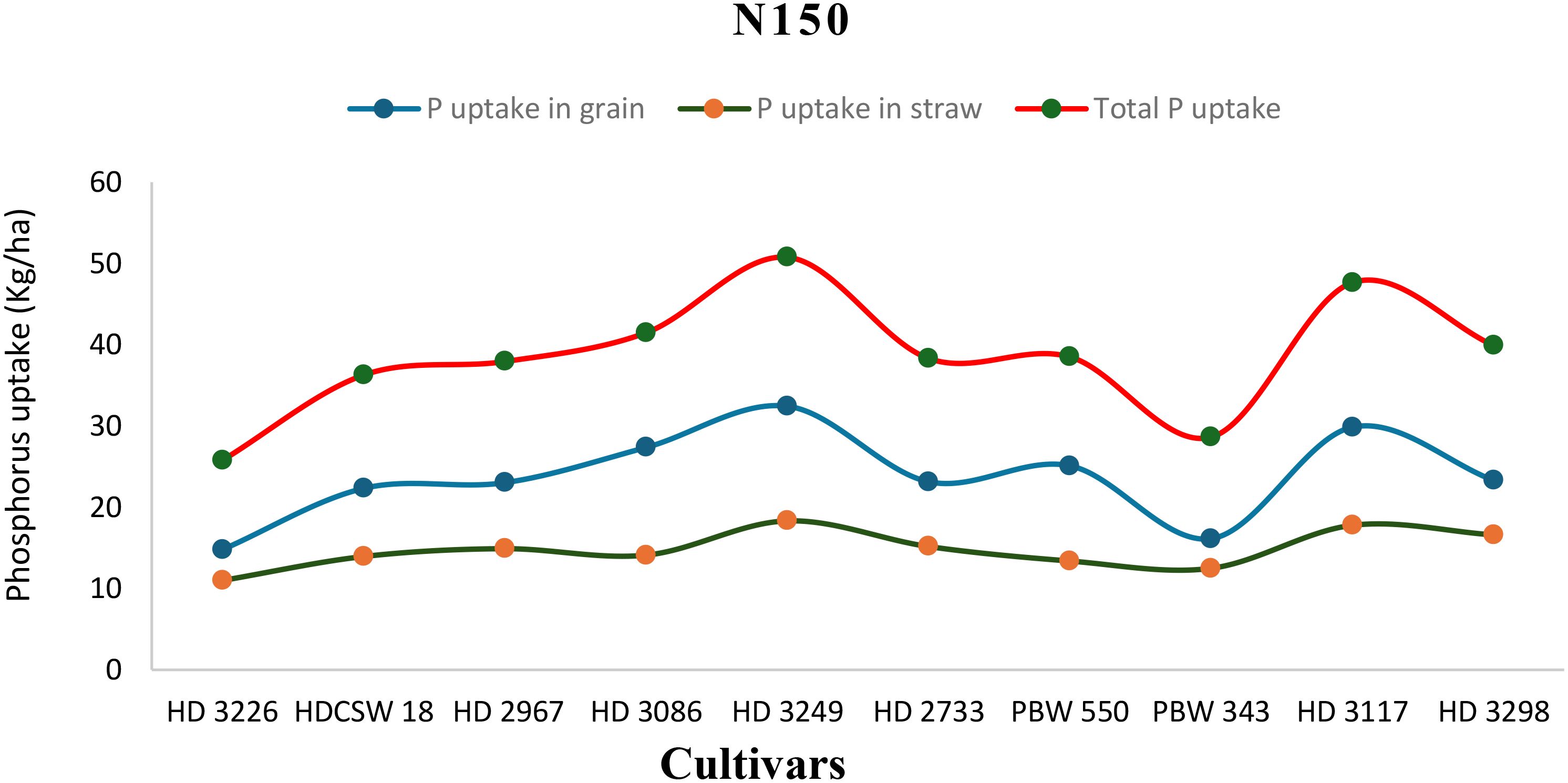
Figure 3. Phosphorus uptake in grain, straw, total P uptake and their trends of ten wheat cultivars over two years under N150-HN (High-nitrogen) conditions.
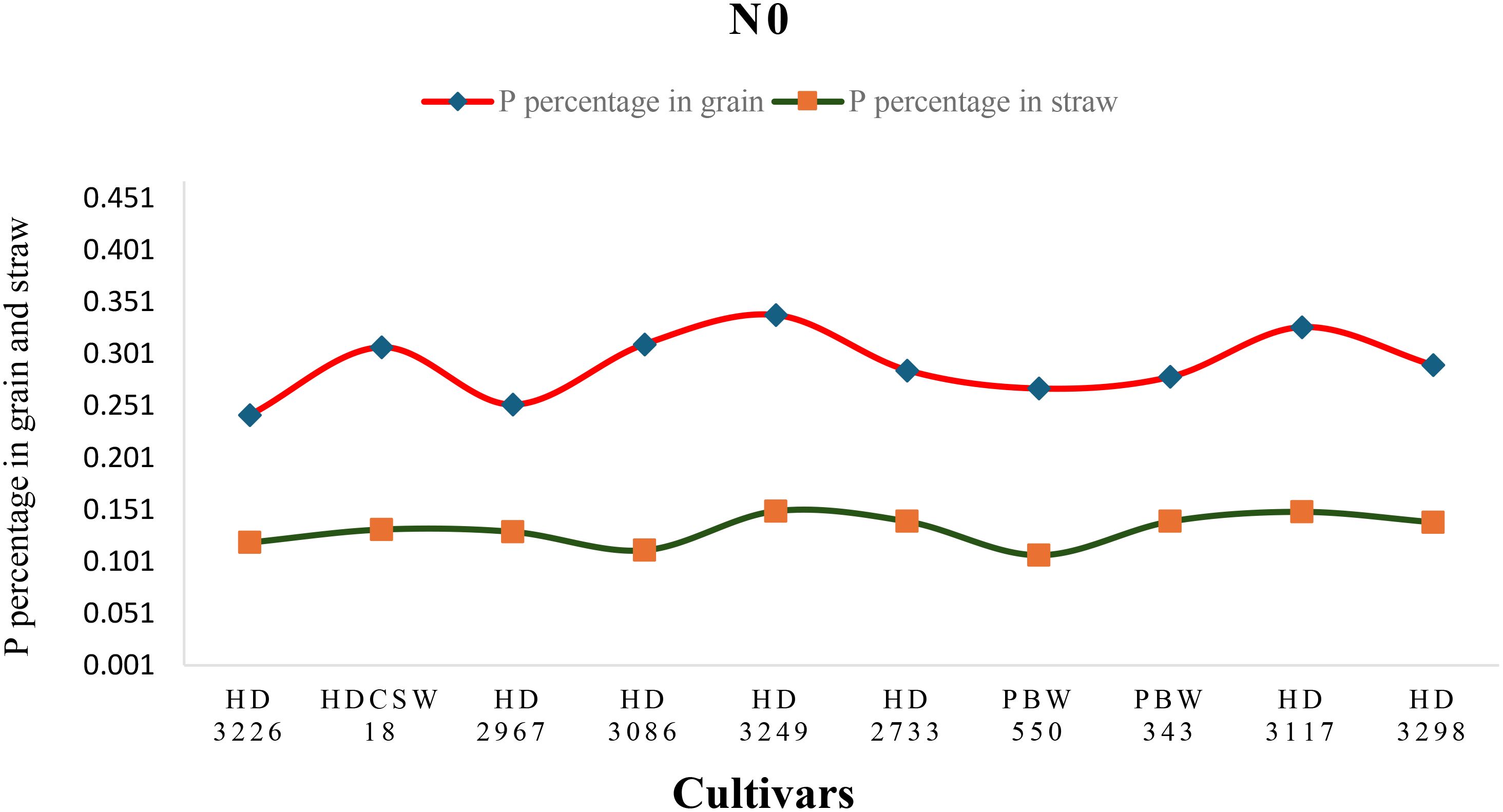
Figure 4. Phosphorus (P) concentration (percentage) in grain, straw, and their trends of ten wheat cultivars over two years under N0- (No-nitrogen) conditions.
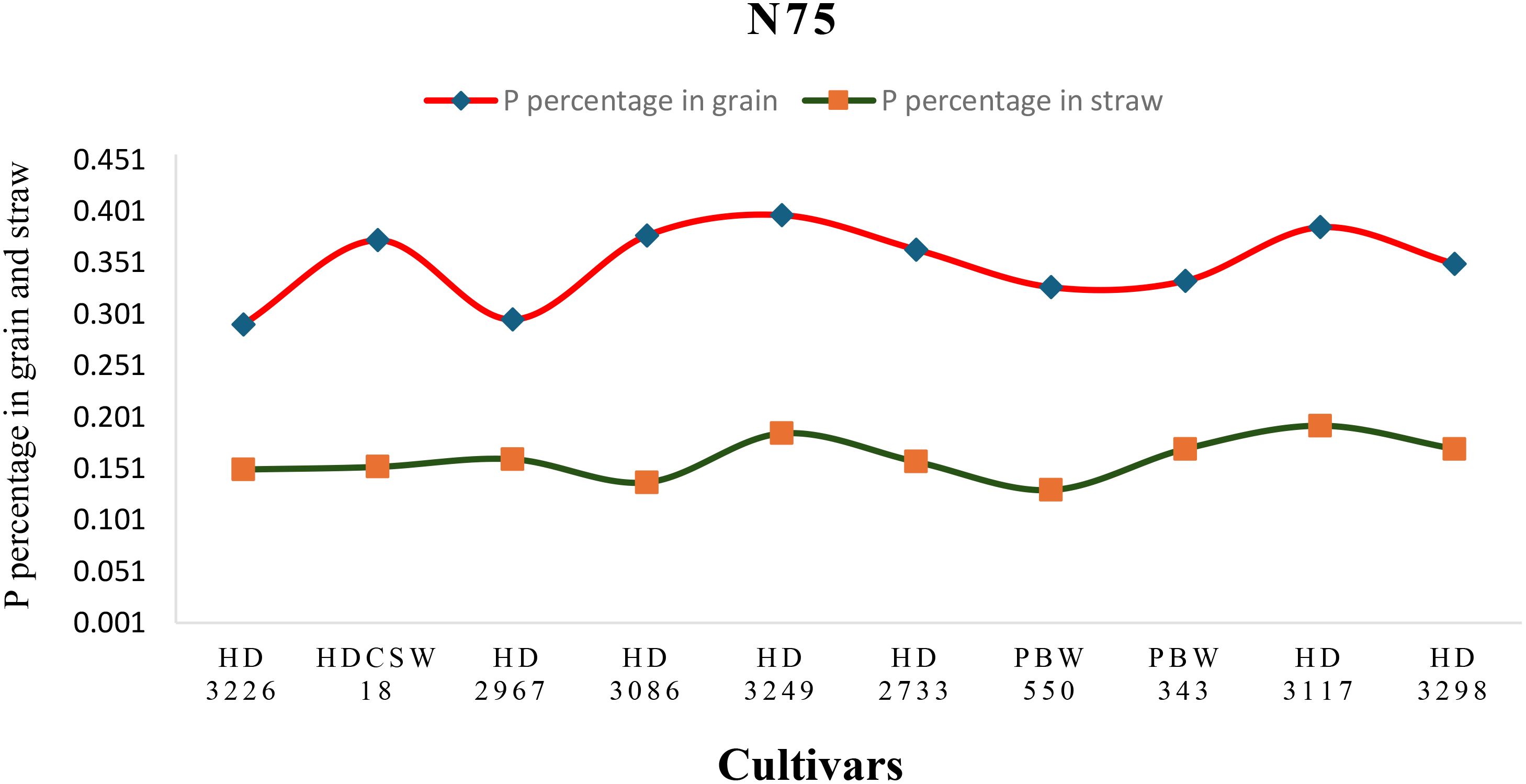
Figure 5. Phosphorus concentration (percentage) in grain, straw, and their trends of ten wheat cultivars over two years under N75-LN (low nitrogen) conditions.
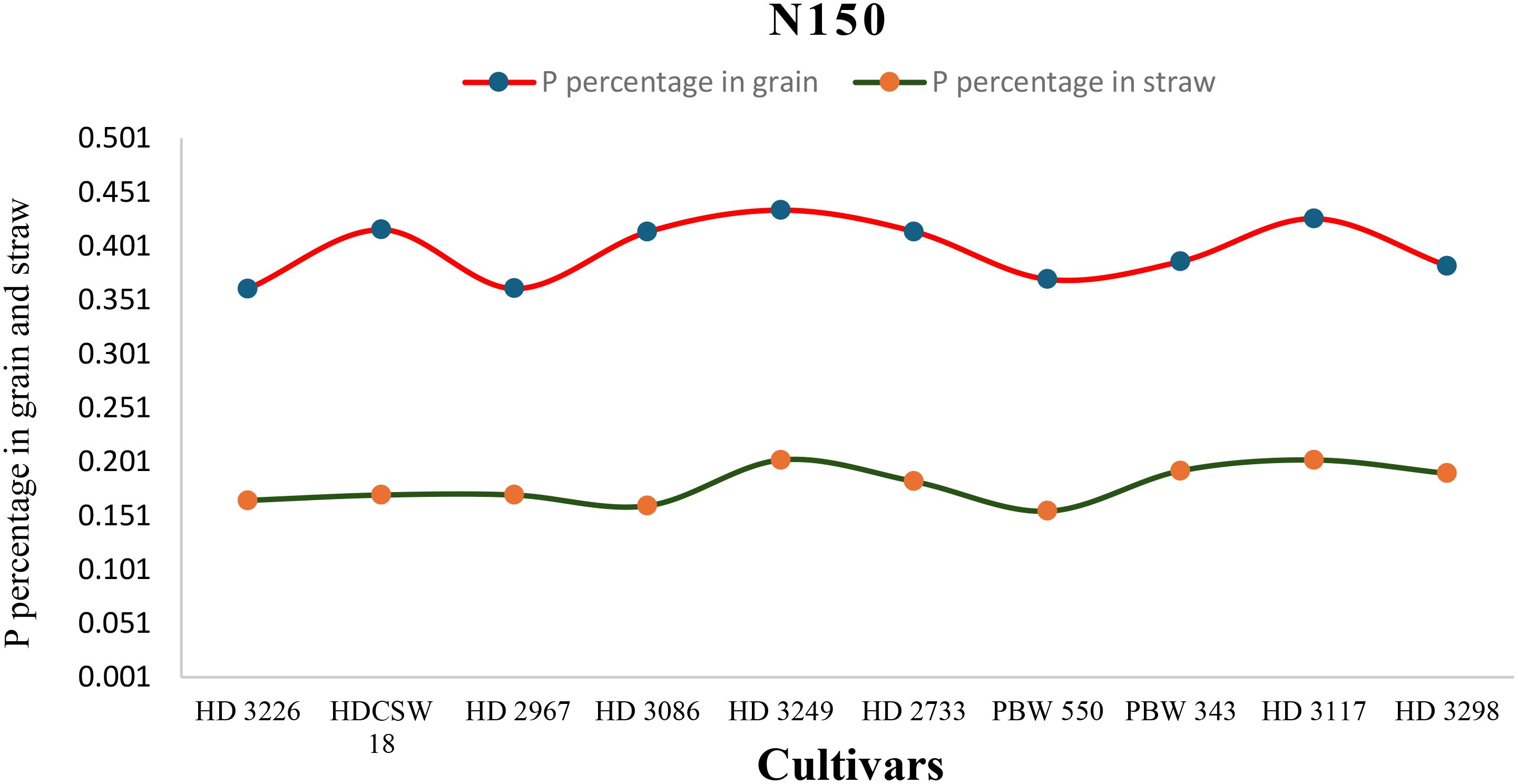
Figure 6. Phosphorus concentration (percentage) in grain, straw, and their trends of ten wheat cultivars over two years under N150-HN (High-nitrogen) conditions.
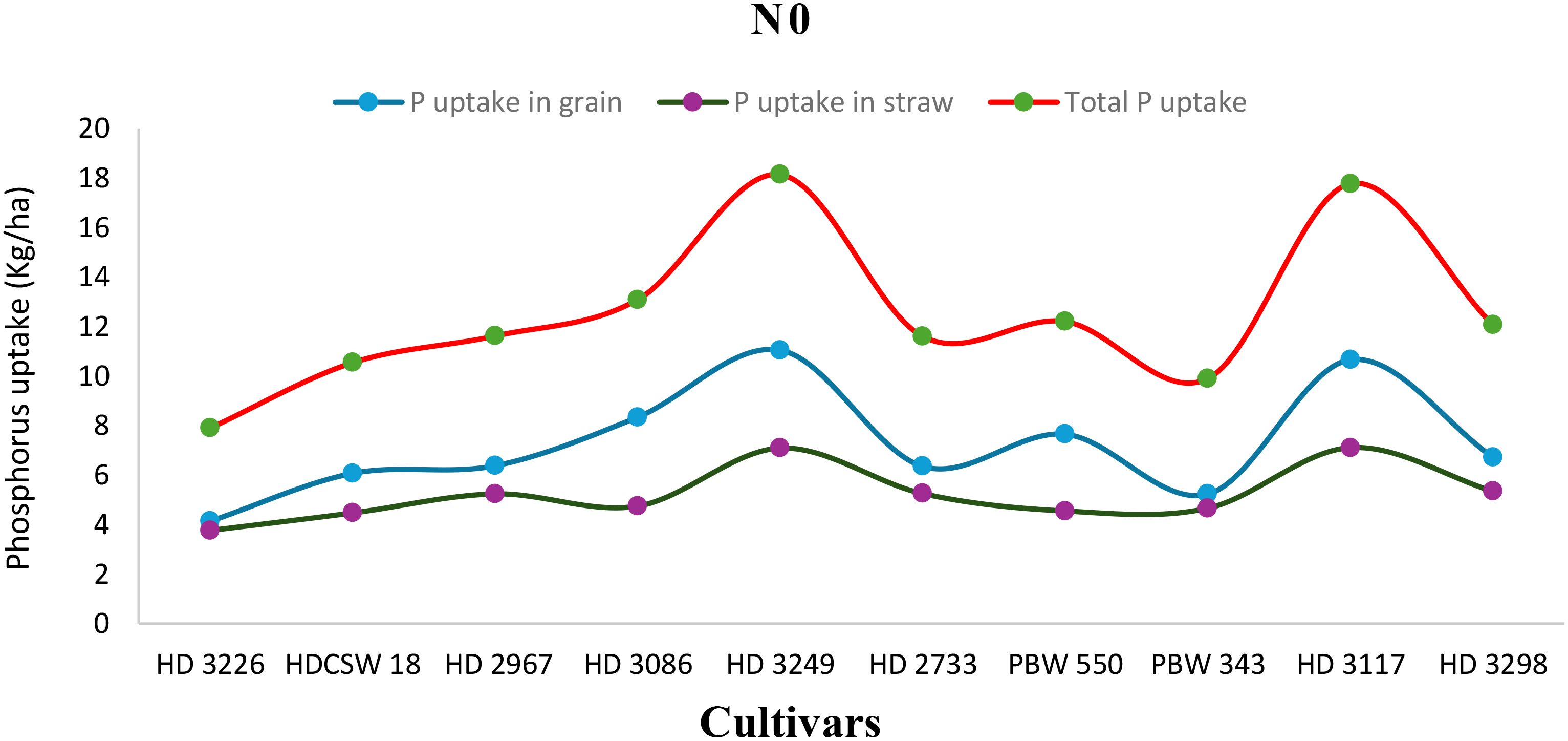
Figure 7. Phosphorus uptake in grain, straw, total P uptake and their trends of ten wheat cultivars over two years under N0- (no nitrogen) conditions.
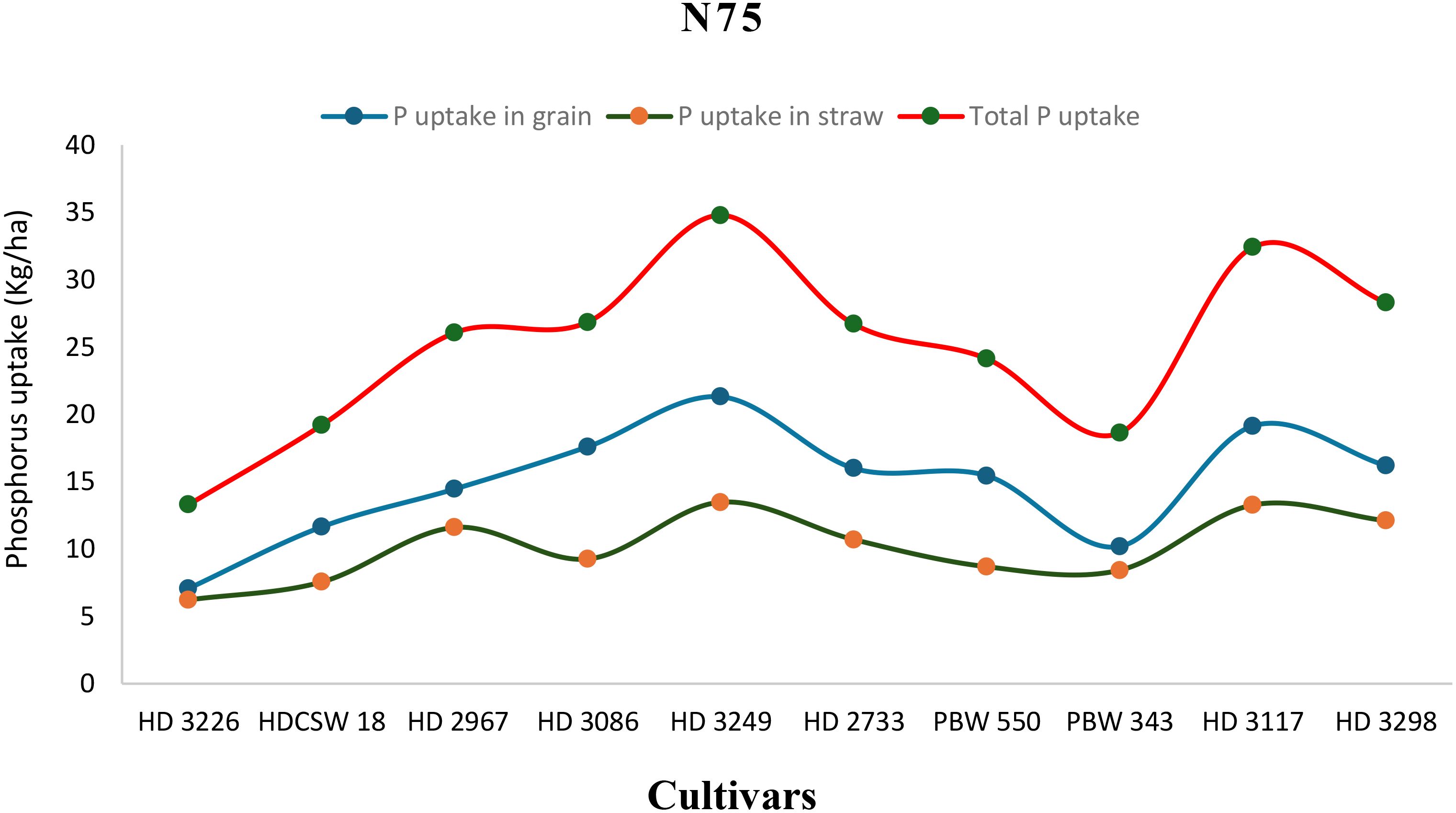
Figure 8. Phosphorus uptake in grain, straw, total P uptake and their trends of ten wheat cultivars over two years under N75-LN (low nitrogen) conditions.
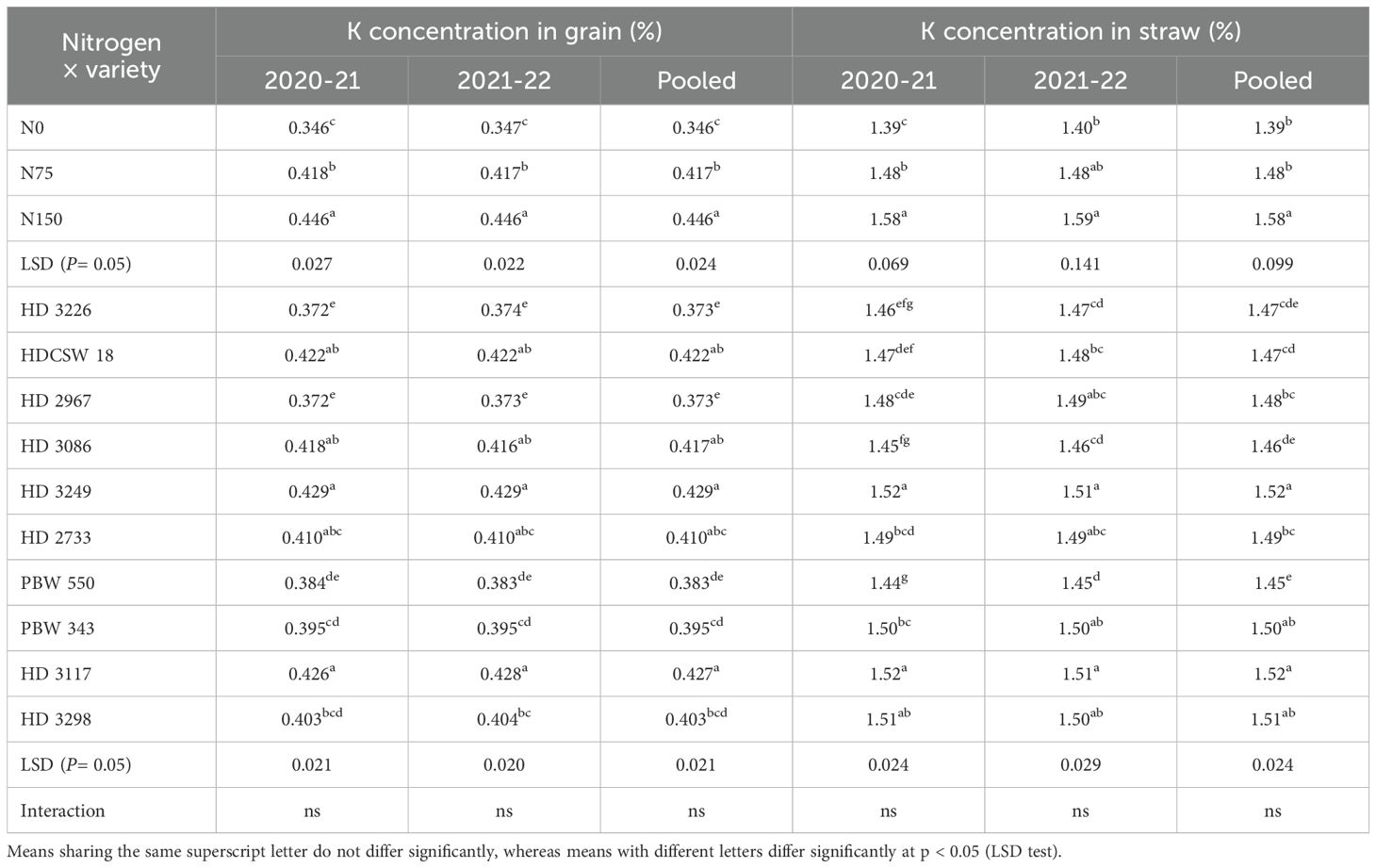
Table 3. Effect of nitrogen fertilization on potassium (K) concentration (%) in grain and straw of wheat at harvesting stage.

Table 4. Effect of nitrogen × cultivar interaction on K uptake in grain of wheat.
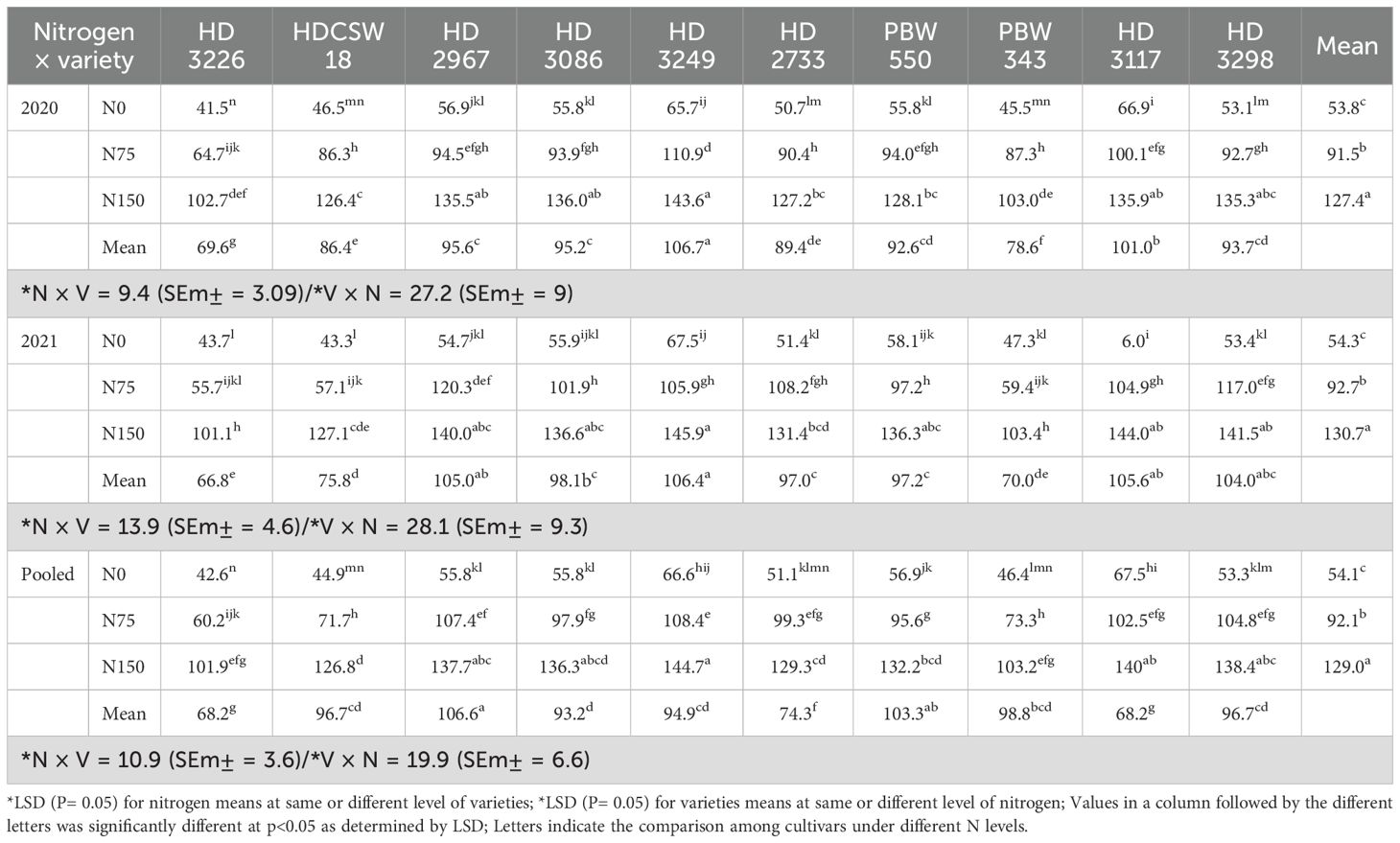
Table 5. Effect of nitrogen × cultivar interaction on K uptake in straw of wheat.
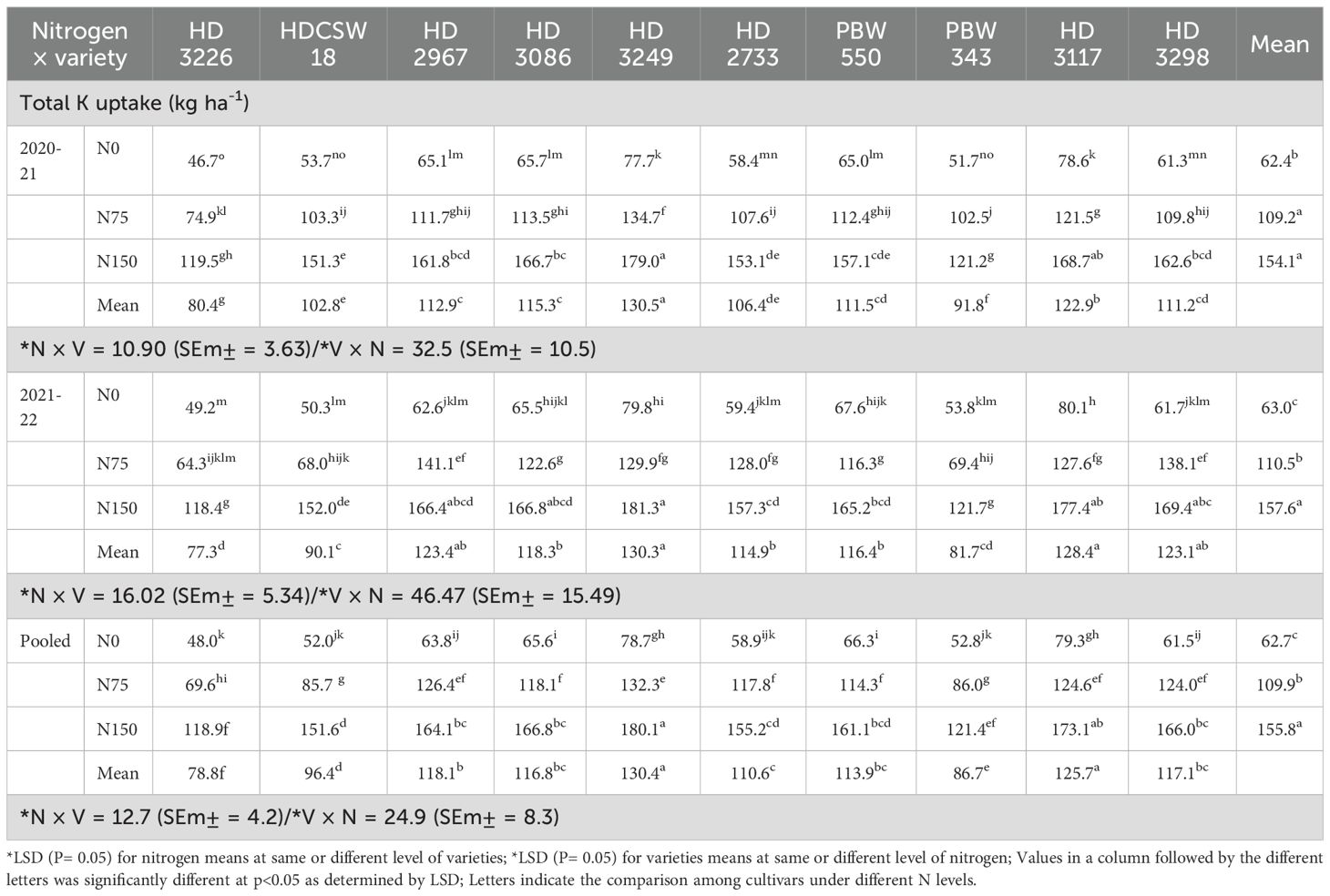
Table 6. Effect of nitrogen × cultivar interaction on total K uptake of wheat.
Discussion
The Indo-Gangetic Plains (IGP) of India are a hotspot for excessive N use, primarily due to limited access to advanced technology, inadequate extension services, and the absence of economic disincentives for overuse (Stringer et al., 2020; Khan et al., 2024). This over-reliance on N fertilizers leads to multiple negative agroecosystem outcomes. To address these challenges, there is a growing need for on-station and on-farm screening experiments to identify N-efficient cultivars and enhance their adoption rates in farming systems (Gawdiya et al., 2023a, Gawdiya et al., 2023b, Gawdiya et al., 2023c, Gawdiya et al., 2023e). Our findings highlight substantial opportunities to improve nutrient use efficiency (NUE) through two complementary pathways: identifying N-efficient cultivars (N-saving roadmap) and assessing yield responses to graded N nutrition (yield-gain roadmap).
Identifying N-efficient cultivars is one of the most sustainable approaches in the current agricultural context. Previous attempts to identify agronomically optimal cultivars have primarily relied on greenhouse and laboratory settings (Zhang et al., 2024), which often lead to generalized assumptions about the N levels and their interactions with macronutrients. Our study findings align with previous research, demonstrating a positive relationship between N application and phosphorus (P) and potassium (K) uptake in wheat (Godebo et al., 2021). However, these responses are highly dependent on multiple factors, including soil pH, microbial activity, and root system architecture of cultivars, which influence nutrient availability and uptake efficiency (Touhami et al., 2022). These insights provide a practical roadmap for an evidence-based, targeted governance approach to N-smart production systems. By moving away from uniform recommended fertilizer doses and embracing site-specific nutrient management, scientists and policymakers can enhance NUE and promote sustainable agricultural intensification.
Nitrogen is absorbed by plant roots primarily in the form of nitrate (NO3-) and ammonium (NH4+) ions. Its interaction with macronutrient uptake is dynamic and complex, influenced by soil properties, microbial activity, rhizodeposition, root traits, and rhizosphere processes (; Treseder, 2004; Yuan and Chen, 2012b; Fan et al., 2019; Bicharanloo et al., 2020; Schleuss et al., 2020; He et al., 2020; Leptin et al., 2021). Nitrogen stimulates root and shoot growth, modifies plant metabolism, and influences soil pH through rhizosphere acidification, which can reduce Ca-bound phosphorus (Ca-P) and total phosphorus (P) levels in the soil, thereby facilitating soil-to-plant P translocation (Blair et al., 1971; Sharpley, 2000). Additionally, N enhances the mobility and accessibility of less-soluble inorganic macronutrients, improving their solubility and bioavailability for plant uptake (Sherman et al., 2006; Fan et al., 2019). These processes collectively contribute to greater nutrient acquisition efficiency, supporting optimal plant growth and productivity (Hinsinger, 2001; Zhang et al., 2004; Havlin et al., 2016). In our study, the cultivars ‘HD 3249’ and ‘HD 3117’ exhibited superior performance compared to other cultivars, primarily due to their greater soil nutrient acquisition by roots, ability to alter soil pH, and genetic traits that enhance stability and adaptability to the microclimate. This underscores the pivotal role of soil chemical processes, in conjunction with the genetic potential of superior cultivars, in regulating nutrient availability, uptake efficiency, and assimilation under varying N inputs.
In various cultivar-based rice and wheat cropping systems, ‘HD 3298’ (Nidhi-rice) and ‘HD 3086’ (Daya-rice) exhibit superior WEY due to their consistently higher grain productivity (Gawdiya et al., 2023a, Gawdiya et al., 2023b, Gawdiya et al., 2023c, –, Gawdiya et al., 2023e; Gawdiya et al., 2024). Similar findings were reported by Ramachandra et al. (2007) and Rao et al. (2014). To comprehensively assess treatment impacts, we quantified system productivity (SP) using WEY as a standardized metric. Notably, ‘HD 3249’, ‘HD 3117’, and ‘HD 3298’ demonstrated the highest SP, with their performance predominantly driven by higher WEY.
Macronutrient’s concentrations and uptake
Nitrogen-smart production systems that synchronize N supply with cultivar-specific demand through split-dose applications are essential for maximizing nitrogen use efficiency (NUE) in agricultural systems (Hu et al., 2021; Udvardi et al., 2021; Su et al., 2022). Our study highlights that N-efficient genotypes (‘HD 3249’ and ‘HD 3117’), exhibited superior morphophysiological and biochemical adaptations by optimize macronutrient acquisition and utilization across varying N availability. These genotypes also balanced trade-offs between yield potential and N dilution effects, enhancing nutrient uptake, assimilation, and remobilization (Uribelarrea et al., 2007; Móring et al., 2021; Zhao et al., 2022; He et al., 2023; Liu et al., 2024);. Their enhanced efficiency is rooted in robust root architecture, efficient metabolic pathways, symbiotic soil microbiomes, and organic exudation, enabling them to explore greater soil volumes and dynamically respond to edaphic and endogenous signals for optimized nutrient acquisition (Rengel and Marschner, 2005; Chen et al., 2019; Jia et al., 2022; Abdulraheem et al., 2024). This adaptive foraging strategy, coupled with increased expression and activity of N transporter proteins, enhances N uptake efficiency (Nacry et al., 2013). Under optimal N conditions, the low-affinity transport system becomes more active, facilitating efficient N assimilation (Nacry et al., 2013). Once absorbed, N is efficiently assimilated through nitrate reductase and glutamine synthetase, working in tandem with hormonal regulation to optimize root-to-shoot signaling and enhance nutrient uptake (Zörb et al., 2018; Zhang et al., 2017; Jia et al., 2022). These genotypic adaptations also enhance N-driven nutrient uptake under environmental stress, improving N assimilation and metabolic efficiency (Francis et al., 2023). A finely-tuned genetic regulation and partitioning mechanism directs N allocation toward chlorophyll synthesis, key transporters, and photosynthetic proteins such as Rubisco, while also supporting the synthesis of peptides and other N-containing compounds. This optimized allocation enhances macronutrient utilization, maximizing photosynthetic efficiency and biomass production per unit of N absorbed (Balyan et al., 2016; Evans and Clarke, 2019; Mălinaş et al., 2022; Aluko et al., 2023; Zayed et al., 2023b). As crops mature, N-efficient genotypes exhibit superior N remobilization, reallocating stored N from vegetative tissues to developing grains during senescence (Liang et al., 2023). Their phenotypic plasticity further enables them to maintain stable performance across varying N inputs (Lenoir et al., 2023). The intricate synergy among plant genetics, soil biology, and nutrient interactions underscores a transformative opportunity for advancing sustainable food systems in diverse agroecosystems.
Dominant influencing factors
Cultivars grown across diverse agroecological regions exhibit significant variation in macronutrient uptake, influenced by environmental conditions, genotype-environment interactions, and soil fertility status. Our analysis of diverse cultivars underscores the critical role of soil properties, nutrient availability (N, P, K), and their interactions in regulating nutrient acquisition efficiency. These insights provide a foundation for developing region-specific N management strategies, tailored to cultivar-specific nutrient demands while integrating site-specific agronomic practices and environmental constraints. Such approaches could enhance NUE, optimize yield potential, and promote sustainable agricultural intensification.
Implication and perspectives
The environmental costs associated with low NUE in global croplands pose a significant challenge due to increased N losses, greenhouse gas emissions, and soil and water contamination (Omara et al., 2019). Since the Green Revolution, agricultural practices have primarily emphasized high-yielding, N-intensive production systems. However, beyond the optimal N threshold, further increases in yield and phosphorus/potassium (P/K) concentrations can occur at the expense of elevated N losses, which ultimately disrupting nutrient balance and environmental sustainability. This imbalance contributes to increasing pressure on climate stability and biodiversity through negative N balances (Goyal et al., 2024; Srivastava et al., 2024). Therefore, there is a urgent need to enhance NUE to address global agricultural and environmental challenges. In response to these challenges and the impacts of climate variability, on-station and on-farm cultivar screening experiments offer a sustainable approach to identify resilient, N-efficient genotypes. Our study provides novel insights into the interactions between graded N levels and the assimilation, translocation, and utilization of essential macronutrients. These findings can enhance resource use efficiency while minimizing environmental impacts, particularly in intensive wheat-growing regions such as North-West India, where excessive N application leads to nutrient imbalances and environmental degradation. Furthermore, these insights open new avenues for breeding agronomically important cultivars with biological nitrification inhibition traits, facilitating the development of climate-smart cultivars that establish robust trade-offs between yield gains and reduced environmental footprints (Gawdiya et al., 2024; Leon and Nedumaran, 2024; Gawdiya and Kumar, 2025). To further enhance NUE, we recommend a strategy that includes the partial substitution of synthetic fertilizers with organic sources combined with foliar nutrient supplementation. These multidimensional strategies are essential for developing sustainable, resilient production systems that improve soil health, crop productivity, and long-term agricultural sustainability.
Conclusion
The present study highlights the genetic variability among wheat cultivars under graded nitrogen levels, with ‘HD 3249’ and ‘HD 3117’ demonstrating superior performance in WEY, SP, and macronutrient uptake. The adoption of these N-efficient cultivars is crucial for optimizing fertilizer use, reducing input costs, and mitigating environmental concerns. Their potential for biological nitrification inhibition could further enhance nitrogen use efficiency, making wheat production in the Indo-Gangetic Plains more economically viable and environmentally sustainable. These findings underscore the need for cultivar-specific nutrient management strategies and farmer awareness programs to maximize productivity while minimizing environmental impact.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
Author contributions
SG: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. DK: Data curation, Writing – review & editing, Project administration, Methodology, Resources. RKS: Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research experiment was financially supported by ICAR- Indian Agricultural Research Institute (IARI), New Delhi, India.
Acknowledgments
The lead author is very grateful to IARI, New Delhi, India for offering a senior research fellowship for the entire duration of his Ph.D. work.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abdulraheem M. I., Moshood A. Y., Li L., Taiwo L. B., Oyedele A. O., Ezaka E., et al. (2024). Reactivating the potential of lima bean (Phaseolus lunatus) for enhancing soil quality and sustainable soil ecosystem stability. Agriculture 14, 976. doi: 10.3390/agriculture14070976
Agren G. I., Wetterstedt J. A. M., and Billberger M. F. (2012). Nutrient limitation on terrestrial plant growth–modeling the interaction between nitrogen and phosphorus. New Phytol. 194, 953–960. doi: 10.1111/j.1469-8137.2012.04116.x
Aluko O. O., Kant S., Adedire O. M., Li C., Yuan G., Liu H., et al. (2023). Unlocking the potentials of nitrate transporters at improving plant nitrogen use efficiency. Front. Plant Sci. 14, 1074839. doi: 10.3389/fpls.2023.1074839
Anas M., Liao F., Verma K. K., Sarwar M. A., Mahmood A., Chen Z. L., et al. (2020). Fate of Nitrogen in agriculture and environment: agronomic, eco-physiological and molecular approaches to improve N use efficiency. Biol. Res. 53, 1–20. doi: 10.1186/s40659-020-00312-4
Asner G. P., Townsend A. R., Riley W. J., Matson P. A., Nef J. C., and Cleveland C. C. (2001). Physical and biogeochemical controls over terrestrial ecosystem responses to nitrogen deposition. Biogeochemistry 54, 1–39. doi: 10.1023/A:1010653913530
Balyan H. S., Gahlaut V., Kumar A., Jaiswal V., Dhariwal R., Tyagi S., et al. (2016). Nitrogen and phosphorus use efficiencies in wheat: physiology, phenotyping, genetics, and breeding. Plant Breed. Rev. 40, 167–234. doi: 10.1002/9781119279723.ch4
Bicharanloo B., Shirvan M. B., and Keitel C and Dijkstra F. A. (2020). 2019. Nitrogen and phosphorus availability affect wheat carbon allocation pathways: rhizodeposition and mycorrhizal symbiosis. Soil Res. 58, 125–136. doi: 10.1071/SR19183
Blair G. J., Mamaril C. P., and Miller M. H. (1971). Influence of Nitrogen Source on Phosphorus Uptake by Corn from Soils Differing in pH 1. Agron. J. 63 (2), 235–238.
Compton J. E., Harrison J., Dennis R. L., Greaver T., Hill B., Jordan S. J., et al. (2011). Ecosystem services altered by human changes in the Nitrogen cycle: a new perspective for US decision making. Ecology letters, 14 (8) 804–815. doi: 10.1111/j.1461-0248.2011.01631.x
Chen Y., Palta J. A., Wu P., and Siddique K. H. (2019). Crop root systems and rhizosphere interactions 370, 1–29. doi: 10.1111/j.1461-0248.2011.01631.x
Elser J. J., Bracken M. E. S., Cleland E. E., Gruner D. S., Harpole W. S., Hillebrand H., et al. (2007). Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 10, 1135–1142. doi: 10.1111/j.1461-0248.2007.01113.x
Evans J. R. and Clarke V. C. (2019). The nitrogen cost of photosynthesis. J. Exp. Bot. 70 (1), 7–15.
Fan Y., Zhong X., Lin F., Liu C., Yang L., Wang M., et al. (2019). Responses of soil phosphorus fractions after nitrogen addition in a subtropical forest ecosystem: Insights from decreased Fe and Al oxides and increased plant roots. Geoderma 337, 246–255. doi: 10.1016/j.geoderma.2018.09.028
Fernanda M., Golombek K., Hansen J., Iqbal A., Asif Ali M., Hahn E., et al. (2022). Fertilizer use by crop and country for the 2017–2018 period (Paris, France: International Fertilizer Association (IFA).
Flores-Saavedra M., Villanueva G., Gramazio P., Vilanova S., Mauceri A., Abenavoli M. R., et al. (2024). Nitrogen use efficiency, growth and physiological parameters in different tomato genotypes under high and low N fertilisation conditions. Plant Physiol. Biochem. 208, 108447. doi: 10.1016/j.plaphy.2024.108447
Francis B., Aravindakumar C. T., Brewer P. B., and Simon S. (2023). Plant nutrient stress adaptation: A prospect for fertilizer limited agriculture. Environ. Exp. Bot. 105431. doi: 10.1016/j.envexpbot.2023.105431
Gawdiya S. and Kumar D. (2025). Biological nitrification inhibition enabled climate smart nitrogen efficient genotypes for today and tomorrow. Indian J. Fertilisers 21, 112–119.
Gawdiya S., Kumar D., Ahmed B., Sharma R. K., Das P., Choudhary M., et al. (2024). Field scale wheat yield prediction using ensemble machine learning techniques. Smart Agric. Technol. 9, 100543. doi: 10.1016/j.atech.2024.100543
Gawdiya S., Kumar D., and Shivay Y. S. (2023c). Evaluation of wheat (Triticum aestivum) genotypes for higher yield and enhanced nitrogen use efficiency in Indo-Gangetic Plains. Indian J. Agric. Sci. 93, 715–719. doi: 10.56093/ijas.v93i7.136347
Gawdiya S., Kumar D., Shivay Y. S., Bhatia A., Mehrotra S., Chandra M. S., et al. (2023e). Field-based evaluation of rice genotypes for enhanced growth, yield attributes, yield and grain yield efficiency index in irrigated lowlands of the Indo-Gangetic plains. Sustainability 15, 8793. doi: 10.3390/su15118793
Gawdiya S., Kumar D., Shivay Y. S., Kour B., Kumar R., Meena S., et al. (2023a). Field screening of wheat cultivars for enhanced growth, yield, yield attributes, and nitrogen use efficiencies. Agronomy 13, 2011. doi: 10.3390/agronomy13082011
Gawdiya S., Kumar D., Shivay Y. S., Nayak S., Ahmed B., Kour B., et al. (2023b). Nitrogen-driven genotypic diversity of wheat (Triticum aestivum L.) genotypes. Agronomy 13, 2447. doi: 10.3390/agronomy13102447
Godebo T., Laekemariam F., and Loha G. (2021). Nutrient uptake, use efficiency and productivity of bread wheat (Triticum aestivum L.) as affected by nitrogen and potassium fertilizer in Keddida Gamela Woreda, Southern Ethiopia. Environ. Syst. Res. 10, 12. doi: 10.1186/s40068-020-00210-4
Govindasamy P., Muthusamy S. K., Bagavathiannan M., Mowrer J., Jagannadham P. T. K., Maity A., et al. (2023). Nitrogen use efficiency—a key to enhance crop productivity under a changing climate. Front. Plant Sci. 14, 1121073. doi: 10.3389/fpls.2023.1121073
Goyal S. S., Kumar R., and Bhatia U. (2024). Assessing temporal dynamics of nitrogen surplus in Indian agriculture: district scale data from 1966 to 2017. Sci. Data 11, 1191. doi: 10.1038/s41597-024-04023-3
Hamdan M. F., Mohd Noor S. N., Abd-Aziz N., Pua T. L., and Tan B. C. (2022). Green revolution to gene revolution: technological advances in agriculture to feed the world. Plants 11, 1297. doi: 10.3390/plants11101297
Hanway J. J. (1952). Soil analysis methods as used in the iowa state college soil testing laboratory. Iowa Agric. 57, 1.
Hao M. D., Jun F., Wei X., Pen L. F., and Lai L. (2005). Effect of fertilization on soil fertility and wheat yield of dryland in the Loess Plateau. Pedosphere 15, 189–195.
Havlin J. L., Tisdale S. L., Nelson W. L., and Beaton J. D. (2016). Soil fertility and fertilizers. Pearson Educ. India.
He W., Li J., Lu Y., Chen S., Deng L., Xu X., et al. (2023). Development of critical K dilution curves for diagnosing sweetpotato K status. Front. Plant Sci. 14, 1124328. doi: 10.3389/fpls.2023.1124328
He H., Wu M., Fan C., Zhang Z., Su R., Peng Q., et al. (2020). Release of tartrate as a major carboxylate by alfalfa (Medicago sativa L.) under phosphorus deficiency and the effect of soil nitrogen supply. Plant Soil 449, 169–178. doi: 10.1007/s11104-020-04481-9
He Y. Q., Zhu Y. G., and Smith S and Smith F. (2002). Interactions between soil moisture content and phosphorus supply in spring wheat plants grown in pot culture. J. Plant Nutr. 25, 913–925. doi: 10.1081/PLN-120002969
Hinsinger P. (2001). Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: a review. Plant Soil 237 (2), 173–195.
Hu C., Sadras V. O., Lu G., Zhang P., Han Y., Liu L., et al. (2021). A global meta-analysis of split nitrogen application for improved wheat yield and grain protein content. Soil Tillage Res. 213, 105111. doi: 10.1016/j.still.2021.105111
Jackson M. L. (1973). Soil chemical analysis, pentice hall of India Pvt. Ltd Vol. 498 (New Delhi, India), 151–154.
Jia Z., Giehl R. F., and von Wirén N. (2022). Nutrient–hormone relations: Driving root plasticity in plants. Mol. Plant 15, 86–103. doi: 10.1016/j.molp.2021.12.004
Jiang J., Wang Y. P., Yang Y., Yu M., and Wang C and Yan J. (2019). Interactive effects of nitrogen and phosphorus additions on plant growth vary with ecosystem type. Plant Soil 440, 523–537. doi: 10.1007/s11104-019-04119-5
John D. A. and Babu G. R. (2021). Lessons from the aftermaths of green revolution on food system and health. Front. Sustain. Food Syst. 5, 644559. doi: 10.3389/fsufs.2021.644559
Kabato W., Ergudo T., Mutum L., and Janda T and Molnár Z. (2022). Response of wheat to combined application of nitrogen and phosphorus along with compost. J. Crop Sci. Biotechnol. 25, 557–564. doi: 10.1007/s12892-022-00151-7
Kaiser D. E., Mallarino A. P., and Sawyer J. E. (2010). Utilization of poultry manure phosphorus for corn production. Soil Sci. Soc. America J. 74, 2211–2222. doi: 10.2136/sssaj2009.0419
Khan F. U., Nouman M., Negrut L., Abban J., Cismas L. M., and Siddiqi M. F. (2024). Constraints to agricultural finance in underdeveloped and developing countries: a systematic literature review. Int. J. Agric. Sustainability 22, 2329388. doi: 10.1080/14735903.2024.2329388
Ladha J. K., Tirol-Padre A., Reddy C. K., Cassman K. G., Verma S., Powlson D. S., et al. (2016). Global Nitrogen budgets in cereals: A 50-year assessment for maize, rice and wheat production systems. Sci. Rep. 6, 1–9. doi: 10.1038/srep19355
Lam S. K., Wille U., Hu H. W., Caruso F., Mumford K., Liang X., et al. (2022). Next-generation enhanced-efficiency fertilizers for sustained food security. Nat. Food 3, 575–580. doi: 10.1038/s43016-022-00542-7
Leon A. and Nedumaran S. (2024). Estimating the effect of biological nitrification inhibition-enabled sorghum on nitrogen fertilizer consumption, life cycle GHG emissions, farmer’s benefit and fertilizer subsidy from Indian sorghum production. Sci. Total Environ. 957, 177385. doi: 10.1016/j.scitotenv.2024.177385
Lenoir A., Slafer G. A., Siah A., and Dumont B. (2023). Plasticity of wheat yield components in response to N fertilization. Eur. J. Agron. 150, 126933.
Leptin A., Whitehead D., Anderson C. R., Cameron K. C., and Lehto N. J. (2021). Increased soil nitrogen supply enhances root-derived available soil carbon leading to reduced potential nitrification activity. Appl. Soil Ecol. 159, 103842. doi: 10.1016/j.apsoil.2020.103842
Liang H., Zhou G. P., Gao S. J., Nie J., Xu C. X., Wu J., et al. (2023). Exploring site-specific N application rate to reduce N footprint and increase crop production for green manure-rice rotation system in southern China. J. Environ. Manage. 347, 119033.
Liu J., Wang D., Yan X., Jia L., Chen N., Liu J., et al. (2024). Effect of nitrogen, phosphorus and potassium fertilization management on soil properties and leaf traits and yield of Sapindus mukorossi. Front. Plant Sci. 15, 1300683. doi: 10.3389/fpls.2024.1300683
Liu W., Yang H., Folberth C., M̈ller C., Ciais P., Abbaspour K. C., et al. (2018). Achieving high crop yields with low Nitrogen emissions in global agricultural input intensification. Environ. Sci. Technol. 52, 13782–13791. doi: 10.1021/acs.est.8b03610
Mălinaş A., Vidican R., Rotar I., Mălinaş C., Moldovan C. M., and Proorocu M. (2022). Current status and future prospective for nitrogen use efficiency in wheat (Triticum aestivum L.). Plants 11, 217. doi: 10.3390/plants11020217
Mandal V. K., Jangam A. P., Chakraborty N., and Raghuram N. (2022). Nitrate-responsive transcriptome analysis reveals additional genes/processes and associated traits viz. height, tillering, heading date, stomatal density and yield in japonica rice. Planta 255, 42. doi: 10.1007/s00425-021-03816-9
Mendiburu F. D. (2021). agricolae: Statistical Procedures for Agricultural Research. R package version. 1. Available online at: https://CRAN.R-project.org/package=agricolae (Accessed December 11, 2025).
Menegaz S. T., Fernández F. G., Venterea R. T., Helmers M. J., and Pagliari P. H. (2025). Nitrate, nitrous oxide, and ammonia loss mitigation with optimum rate of enhanced efficiency nitrogen fertilizer and application timing in corn. Nitrogen Cycling Soil Health 15. doi: 10.1007/978-981-96-1132-4_2
Móring A., Hooda S., Raghuram N., Adhya T. K., Ahmad A., Bandyopadhyay S. K., et al. (2021). Nitrogen challenges and opportunities for agricultural and environmental science in India. Front. Sustain. Food Syst. 5, 505347.
Nacry P., Bouguyon E., and Gojon A.. (2013). Nitrogen acquisition by roots: physiological and developmental mechanisms ensuring plant adaptation to a fluctuating resource. Plant and Soil 370, 1-29.
Olsen S. R. (1954). Estimation of available phosphorus in soils by extraction with sodium bicarbonate (No. 939) (US Department of Agriculture).
Omara P., Aula L., Oyebiyi F., and Raun W. R. (2019). World cereal nitrogen use efficiency trends: review and current knowledge. Agrosystems Geosciences Environ. 2, 1–8. doi: 10.2134/age2018.10.0045
Raghuram N. and Sharma N. (2019). Improving crop Nitrogen use efficiency. Comprehensive Biotechnology. 3rd ed Vol. 4 (ElsevierPergamon), 2011–2220.
Ramachandra C., Anwarulla M. S., Janardhan G., and Murthy P. (2007). Production potential and economics of rice-based cropping systems in hill zone of Karnataka. Indian J. Agric. Sci. 3, 127–129.
Rao U. A., Sridhar T. V., Lakshmi D. A., and Krishnam R. S. (2014). Identification of viable rice-based cropping system for double cropped delta areas of Andhra Pradesh. Int. J. Natural Sci. 5, 512–514.
Reganold J. (1995). Soil quality and proftability of biodynamic and conventional farming systems: A review. Am. J. Altern. Agric. 10, 36–45. doi: 10.1017/S088918930000610X
Rengel Z. and Marschner P. (2005). Nutrient availability and management in the rhizosphere: exploiting genotypic differences. New Phytol. 168, 305–312. doi: 10.1111/j.1469-8137.2005.01558.x
Roy R. N., Finck A., Blair G. J., and Tandon H. L. S. (2006). “Nutrient management guidelines for some major field crops,” in Plant nutrition for food security. A guide for integrated nutrient management(FAO, Rome), 235–349. doi: 10.1017/S0014479706394537
Sapkota T. B. and Takele R. (2023). Improving nitrogen use efficiency and reducing nitrogen surplus through best fertilizer nitrogen management in cereal production: The case of India and China. Adv. Agron. 178, 233–294. doi: 10.1016/bs.agron.2022.11.006
Sarmiento G., Silva M. P., Naranjo M. E., Naranjo M. E., and Pinillos M. (2006). Nitrogen and phosphorus as limiting factors for growth and primary production in a flooded savanna in the Venezuelan Llanos. J. Trop. Ecol. 22, 203–212. doi: 10.1017/S0266467405003068
Schleuss P. M., Widdig M., Buschart A. H., and Kirkman K and Spohn M. (2020). Interactions of nitrogen and phosphorus cycling promote P acquisition and explain synergistic plant-growth responses. Ecology 101, e03003. doi: 10.1002/ecy.3003
Sendhil R., Kumari B., Khandoker S., Jalali S., Acharya K. K., Gopalareddy K., et al. (2022). “Wheat in Asia: trends, challenges and research priorities,” in New horizons in wheat and barley research: global trends, breeding and quality enhancement (Springer Singapore, Singapore), 33–61.
Sherman J., Fernandez I. J., Norton S. A., and Ohno T and Rustad L. E. (2006). Soil aluminium, iron, and phosphorus dynamics in response to long-term experimental nitrogen and sulphur additions at the Bear Brook watershed in Maine, USA. Environ. Monitoring Assess. 121, 421–429. doi: 10.1007/s10661-005-9140-2
Singh B. (2022). Nitrogen use efficiency in crop production in India: Trends, issues, and challenges. Agric. Res. 12, 32–44.
Srivastava R. K., Purohit S., Alam E., and Islam M. K. (2024). Advancements in soil management: optimizing crop production through interdisciplinary approaches. J. Agric. Food Res. 101528. doi: 10.1016/j.jafr.2024.101528
Stringer L. C., Fraser E. D., Harris D., Lyon C., Pereira L., Ward C. F., et al. (2020). Adaptation and development pathways for different types of farmers. Environ. Sci. Policy 104, 174–189. doi: 10.1016/j.envsci.2019.10.007
Su R., Wu J., Hu J., Ma L., Ahmed S., Zhang Y., et al. (2022). Minimalizing non-point source pollution using a cooperative ion-selective electrode system for estimating nitrate nitrogen in soil. Front. Plant Sci. 12, 810214. doi: 10.3389/fpls.2021.810214
Subbiah B. V. and Asija G. L. (1956). A rapid method for the estimation of nitrogen in soil. Curr. Sci. 26, 259–260.
Swathy R., Geethalakshmi V., Pazhanivelan S., Kannan P., Annamalai S., and Hwang S. (2024). Real-time nitrogen monitoring and management to augment N use efficiency and ecosystem sustainability–A review. J. Hazardous Materials Adv. 100466. doi: 10.1016/j.hazadv.2024.100466
Touhami D., McDowell R. W., Condron L. M., and Bouray M. (2022). Nitrogen fertilization effects on soil phosphorus dynamics under a grass-pasture system. Nutrient Cycling Agroecosystems 124, 227–246. doi: 10.1007/s10705-021-10191-0
Treseder K. K. (2004). A meta-analysis of mycorrhizal responses to nitrogen, phosphorus, and atmospheric CO2 in field studies. New Phytol. 164, 347–355. doi: 10.1111/j.1469-8137.2004.01159.x
Udvardi M., Below F. E., Castellano M. J., Eagle A. J., Giller K. E., Ladha J. K., et al. (2021). A research road map for responsible use of agricultural nitrogen. Front. Sustain. Food Syst. 5, 660155. doi: 10.3389/fsufs.2021.660155
Uribelarrea M., Moose S. P., and Below F. E. (2007). Divergent selection for grain protein affects nitrogen use in maize hybrids. Field Crops Res. 100, 82–90. doi: 10.1016/j.fcr.2006.05.008
Vargas R., Taboada M., Saynes Santillán V., Cardoso Lisboa C., and Olivera C. (2022). Soils for nutrition: state of the art. Available online at: https://openknowledge.fao.org/items/7ee16f29-37f8-415c-8aa5-002a58da384a (Accessed December 10, 2025).
Velayudhan P. K., Sivalingam N., Jha G. K., Singh A., and Pathak H. (2024). Nitrogen budget of Indian agriculture: trends, determinants and challenges. Environment Dev. Sustainability 26, 10225–10242. doi: 10.1007/s10668-023-03142-y
Walkley A. and Black I. A. (1934). An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 37, 29–38. doi: 10.1097/00010694-193401000-00003
Wen Z., Shen J., Blackwell M., Li H., Zhao B., and Yuan H. (2016). Combined applications of nitrogen and phosphorus fertilizers with manure increase maize yield and nutrient uptake via stimulating root growth in a long-term experiment. Pedosphere 26, 62–73. doi: 10.1016/S1002-0160(15)60023-6
Xia L., Lam S. K., Wolf B., Kiese R., Chen D., and Butterbach-Bahl K. (2018). Trade-offs between soil carbon sequestration and reactive Nitrogen losses under straw return in global agroecosystems. Global Change Biol. 24, 5919–5932. doi: 10.1111/gcb.2018.24.issue-12
Yadav M. R., Kumar S., Lal M. K., Kumar D., Kumar R., Yadav R. K., et al. (2023). Mechanistic understanding of leakage and consequences and recent technological advances in improving nitrogen use efficiency in cereals. Agronomy 13, 527. doi: 10.3390/agronomy13020527
Ye J. Y., Tian W. H., and Jin C. W. (2022). Nitrogen in plants: From nutrition to the modulation of abiotic stress adaptation. Stress Biol. 2 (1), 4.
Yuan Z. Y. and Chen H. Y. H. (2012b). A global analysis of fine root production as affected by soil nitrogen and phosphorus. Proc. R. Soc. B: Biol. Sci. 279, 3796–3802. doi: 10.1098/rspb.2012.0955
Yuan Z. Y. and Chen H. (2015a). Decoupling of nitrogen and phosphorus in terrestrial plants associated with global changes. Nat. Climate Change 5, 465–469. doi: 10.1038/nclimate2549
Zayed O., Hewedy O. A., Abdelmoteleb A., Ali M., Youssef M. S., Roumia A. F., et al. (2023a). Nitrogen journey in plants: From uptake to metabolism, stress response, and microbe interaction. Biomolecules 13, 1443. doi: 10.3390/biom13101443
Zhang W., Awais M., Naqvi S. M. Z. A., Xiong Y., Li L., Zang Y., et al. (2024). Real-time remote corn growth monitoring system using plant wearable fiber Bragg grating sensor. Comput. Electron. Agric. 227, 109538. doi: 10.1016/j.compag.2024.109538
Zhang H., He L., Chen Q., Abdulraheem M. I., Ma G., Zhang Y., et al. (2025). Multi-angular spectroscopic detection of winter wheat nitrogen fertilizer utilization status using integrated feature selection and machine learning. Comput. Electron. Agric. 231, 109916. doi: 10.1016/j.compag.2025.109916
Zhang F., Kang S., Zhang J., and Zhang R and Li F. (2004). Nitrogen fertilization on uptake of soil inorganic phosphorus fractions in the wheat root zone. Soil Sci. Soc. America J. 68, 1890–1895. doi: 10.2136/sssaj2004.1890
Zhang C., Song X., Zhang Y., Wang D., Rees R. M., and Ju X. (2022). Using nitrification inhibitors and deep placement to tackle the trade-offs between NH3 and N2O emissions in global croplands. Global Change Biol. 28, 4409–4422. doi: 10.1111/gcb.v28.14
Zhang H., Khan A., Tan D. K., and Luo H. (2017). Rational water and nitrogen management improves root growth, increases yield and maintains water use efficiency of cotton under mulch drip irrigation. Front. Plant Sci. 8, 912.
Zhao Q., Cao W., Chen X., Stomph T. J., and Zou C. (2022). Global analysis of nitrogen fertilization effects on grain zinc and iron of major cereal crops. Global Food Secur. 33, 100631. doi: 10.1016/j.gfs.2022.100631
Keywords: nitrogen input, macronutrient uptake, system productivity, wheat equivalent yield, wheat cultivars
Citation: Gawdiya S, Kumar D and Sharma RK (2025) Field based evaluation of wheat cultivars under graded nitrogen levels in North-West India. Front. Agron. 7:1558925. doi: 10.3389/fagro.2025.1558925
Received: 11 January 2025; Accepted: 02 April 2025;
Published: 22 May 2025.
Edited by:
Hanuman Singh Jatav, Sri Karan Narendra Agriculture University, IndiaCopyright © 2025 Gawdiya, Kumar and Sharma. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sandeep Gawdiya, c2FuZGVlcGFncm83ODYyNkBnbWFpbC5jb20=; Dinesh Kumar, ZGluZXNoY3R0QHlhaG9vLmNvbQ==