 Mirian Villavicencio-Vásquez1
Mirian Villavicencio-Vásquez1 Fernando Espinoza-Lozano1
Fernando Espinoza-Lozano1 Lisbeth Espinoza-Lozano2
Lisbeth Espinoza-Lozano2 Jonathan Coronel-León1,3,4*
Jonathan Coronel-León1,3,4*- 1Centro de Investigaciones Biotecnológicas del Ecuador (CIBE), Escuela Superior Politécnica del Litoral (ESPOL), Guayaquil, Ecuador
- 2Facultad de Ciencias de la Vida, Escuela Superior Politécnica del Litoral, Escuela Superior Politécnica del Litoral (ESPOL), Guayaquil, Ecuador
- 3Facultad de Ingeniería Mecánica y Ciencias de la Producción, Escuela Superior Politécnica del Litoral, Escuela Superior Politécnica del Litoral (ESPOL), Guayaquil, Ecuador
- 4Institute of Agrifood Research and Technology (IRTA), Food Safety and Functionality Programme, Monells, Spain
The increasing global population has intensified the demand for food production, both in quantity and quality. To meet this challenge and prevent crop diseases, chemical pesticides have been widely used. However, their negative effects on human health and the environment have driven the search for sustainable alternatives. In this context, microbial-based biopesticides, particularly Trichoderma and Bacillus, have emerged as key biocontrol agents in agroecosystems. This review provides a comprehensive overview of their agricultural significance, focusing on their mechanisms of action, efficacy, and biotechnological advancements aimed at optimizing their performance. Trichoderma spp. act primarily through mycoparasitism, nutrient competition, and antibiosis, while Bacillus spp. employ mechanisms such as lipopeptide production, lytic enzymes, siderophores, and the activation of induced systemic resistance (ISR). Unlike traditional reviews that primarily describe their biological functions, this work offers a structured framework for their application, addressing key factors such as selection criteria for effective biocontrol agents, formulation strategies, product stability, viability, and regulatory challenges in commercial agriculture. Additionally, recent studies on genetic enhancement, including CRISPR-based modifications, are examined to improve their adaptability, metabolic activity, and effectiveness in pathogen control across diverse agricultural systems. By integrating biological, technological, and practical perspectives, this review aims to bridge the gap between scientific advancements and real-world agricultural applications, contributing to the development of scalable and sustainable disease management strategies.
1 Introduction
Agriculture has been a fundamental pillar in the social and economic transformation of countries, particularly in developing nations, where it plays a crucial role in ensuring food security and contributing significantly to the economy. However, the prolonged use of agrochemicals has led to serious consequences for the environment, soil health, and human well-being, making the need for more sustainable alternatives ever more pressing (Zhao et al., 2020). According to the World Health Organization (WHO, 2022), over 1,000 pesticides are used worldwide, many of which accumulate in the adipose tissue of organisms, posing risks to both health and the environment (Souza, 2016; Zhang et al., 2019). Although pesticides are necessary for crop protection and food security, their inappropriate or excessive use can lead to serious consequences (Beyuo et al., 2024). These include the negative impact on non-target organisms, such as pollinators and beneficial species that naturally control pests (Elhamalawy et al., 2024), as well as the promotion of pest resistance, which complicates pest control and increases production cost (Kumar Chaube and Pandey, 2022).
In the face of these challenges, it is fundamental to explore alternatives such as Integrated Pest Management (IPM), which integrates biological control agents (BCAs) such as bacteria, fungi, and viruses (Thomine et al., 2022). Recent advancements in pesticide application technologies, such as precision application systems, allow for more accurate dosing, reducing pesticide use and minimizing environmental impact (Anastasiou et al., 2023). Furthermore, the increasing pressure from consumers and international organizations for clean food production has spurred the search for innovative solutions that minimize the harmful effects of agrochemicals (Thilagam et al., 2023). An example is the global commitment to achieving sustainable agriculture by 2050, aligned with the Sustainable Development Goals (SDGs) (Blazhevska, 2019).
In this context, BCAs have emerged as a promising solution to reduce dependence on synthetic pesticides and minimize their environmental impact (Damalas and Koutroubas, 2018). Notably, the genera Trichoderma and Bacillus have gained considerable attention due to their ability to suppress pathogens and promote plant growth naturally (Tyagi et al., 2024). These microorganisms employ several mechanisms to protect plants, including nutrient competition, production of antibiotic metabolites, induction of systemic resistance (ISR) in plants, and competition for space in the soil (Poveda, 2021; Prasad et al., 2023).
This article provides a comprehensive review of the potential of Trichoderma spp. and Bacillus spp. as BCAs, analyzing their mechanisms of action, selection processes, and the challenges associated with their formulation and large-scale implementation. The effectiveness of these microorganisms is influenced by various factors, including environmental conditions, proper formulation, and the selection of specific strains with desirable characteristics (Naamala and Smith, 2020). By reviewing the latest scientific literature, this paper aims to provide a holistic view of how these microorganism function in modern agriculture, highlighting their successes and the challenges that must still be overcome to ensure their widespread adoption in sustainable agricultural systems. Moreover, this review compiles a list of biological products currently available on the Ecuadorian market (Supplementary Materials), offering valuable insights into the practical application of BCAs in deifferent agricultural contexts. This information can help guide future efforts in integrating these biocontrol solutions more broadly into farming practices, contributing to a shift toward more environmentally sustainable agriculture.
2 Trichoderma and Bacillus in agriculture
The use of microorganisms in agriculture has gained significant attention as environmentally friendly alternatives to chemical pesticides. These microorganisms include bacteria, fungi, viruses, and protozoa, that enhancing plant health and performance. Various studies have highlighted the effectiveness of bacteria such as Rhizobium, Azospirillum, Azotobacter, Burkholderia, Klebsiella, Bacillus, and Pseudomonas in agricultural applications (Alizadeh et al., 2020). Also, endophytic microorganisms, which live inside plants without causing harm, are also part of the plant growth-promoting (PGP) group. Among endophytic fungi, species like Penicillium sp., Guignardia mangiferae, Hypocrea sp., Neurospora sp., Eupenicillium javanicum, Lasiodiplodia theobromae, and Trichoderma sp. have proven effective in inhibiting Fusarium oxysporum f.sp. cucumerinum, the principal causal agent of stem rot in cucumber under greenhouse conditions (Abro et al., 2019). Among these agents, fungi such as Trichoderma spp. and bacteria such as Bacillus spp. enhancing soil health, promoting plant growth, and controlling plant pathogens (Alizadeh et al., 2020).
Trichoderma is a filamentous fungus present in the rhizosphere and roots (Ghorbanpour et al., 2018; Jaroszuk-ściseł et al., 2019). According to the MycoBank classification, the Hypocrea/Trichoderma includes more than 300 species characterized both molecularly and morphologically (Tamandegani et al., 2020). These fungi colonize plant roots and produce metabolites with antimicrobial properties, contributing to plant health (Tyśkiewicz et al., 2022). Trichoderma spp. can degrade toxic contaminants like herbicides and fungicides through enzymatic processes that break down cellulose and lignin (Escudero-Leyva et al., 2022). Trichoderma species are renowned for their antagonistic activities against various fungal pathogens, including F. oxysporum, Botrytis cinerea, Rhizoctonia solani, and Alternaria alternata (Es-Soufi et al., 2020). T. asperellum and T. harzianum are frequently noted for their ability to inhibit the growth of Fusarium spp. through the production of hydrolytic enzymes and growth inhibition mechanisms (Vargas-Hoyos and Gilchrist-Ramelli, 2015; Miguel-Ferrer et al., 2021).The wide array of enzymes, including exo- and endo-β-glucanases and proteases, which degrade the structural components of the pathogen’s cell walls, further facilitating colonization (Alizadeh et al., 2020). In comparison, T. spirale and T. atroviride excel in antibiosis and the production of extracellular metabolites, which suppress pathogens like Corynespora cassiicola and Spodoptera frugiperda, respectively (Contreras-Cornejo et al., 2018; Baiyee et al., 2019).
Economically, applying Trichoderma has led to reduced pesticide usage, decreased crop losses, and potentially higher yields, offering a cost-effective solution for sustainable farming, especially in developing countries (Morales-Mora et al., 2020; Martínez-Salgado et al., 2021). The data presented in Table 1 illustrates Trichoderma’s significant role in controlling numerous economically important pathogens across various crops, including tomatoes, wheat, maize, and avocados.
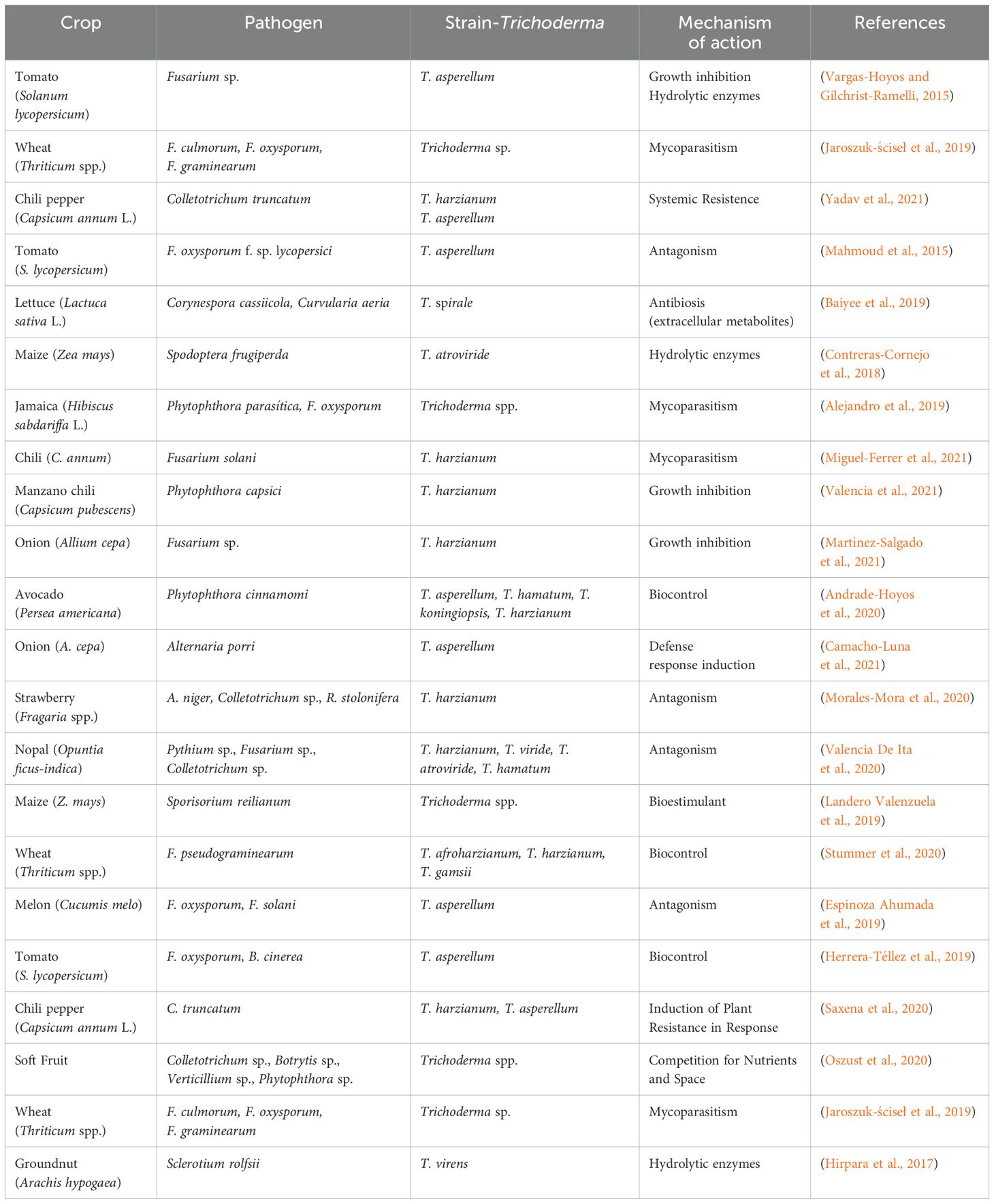
Table 1. Plant pathogenic fungi controlled by different strains of Trichoderma.
On the other hand, Bacillus spp. is recognized for promoting plant growth, enhancing soil fertility, and serving as a biocontrol agent. These Gram-positive, aerobic bacteria produce bioactive compounds that support plant health and reduce pathogen pressure (Etesami et al., 2023). Bacillus subtilis exhibits antifungal properties attributed to metabolites like surfactin, iturin, and fengycin (Méndez-Úbeda et al., 2018; Chen et al., 2021). They also act as biofertilizers or biostimulators by facilitating nutrient uptake and producing plant hormones (Dimopoulou et al., 2021). Integrating Bacillus strains into agricultural practices has led to reduced reliance on chemical pesticides and supported sustainable farming methods.
The selection of Trichoderma and Bacillus species for agricultural use requires careful consideration of factors such as their ability to produce effective metabolites, efficiency in nutrient acquisition, and stability under varying environmental conditions (Silva et al., 2019). Both genera offer distinct advantages for different agricultural systems, with Trichoderma providing broad-spectrum biocontrol against fungal pathogens and Bacillus contributing to disease management and plant growth promotion. Their integration into farming practices can lower production costs, enhance soil health, and improve overall crop productivity (Martínez, 2021). Table 2 highlights the effectiveness of different Bacillus spp. in managing plant diseases across various crops, underscoring their importance in sustainable agriculture.
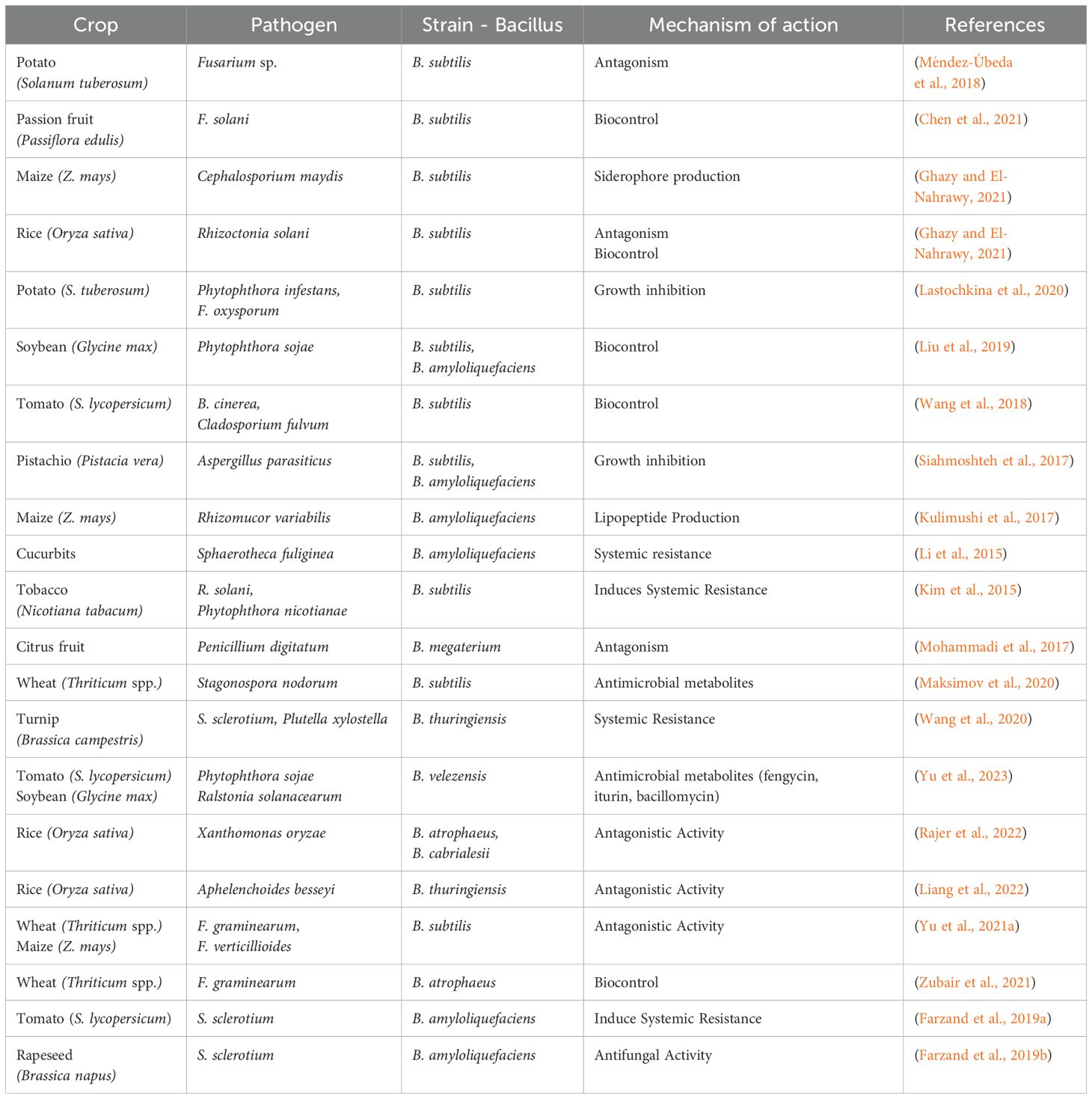
Table 2. Plant pathogenic fungi controlled by different strains of Bacillus.
3 Mechanism of action of Trichoderma spp.
As mentioned, BCAs offer significant benefits to the agricultural sector. However, their effective application depends on a thorough understanding of each microorganism’s specific mechanism of action. This lack of clarity can raise concerns among farmers about whether the selected microorganism is truly appropriate for their needs. To address this issue, this section provides farmers and policymakers with a detailed description of the mechanisms of action of Trichoderma spp. The key biocontrol strategies that Trichoderma spp. develops in direct conflict with fungal pathogens, such as mycoparasitism, competition, and antibiosis (Figure 1).
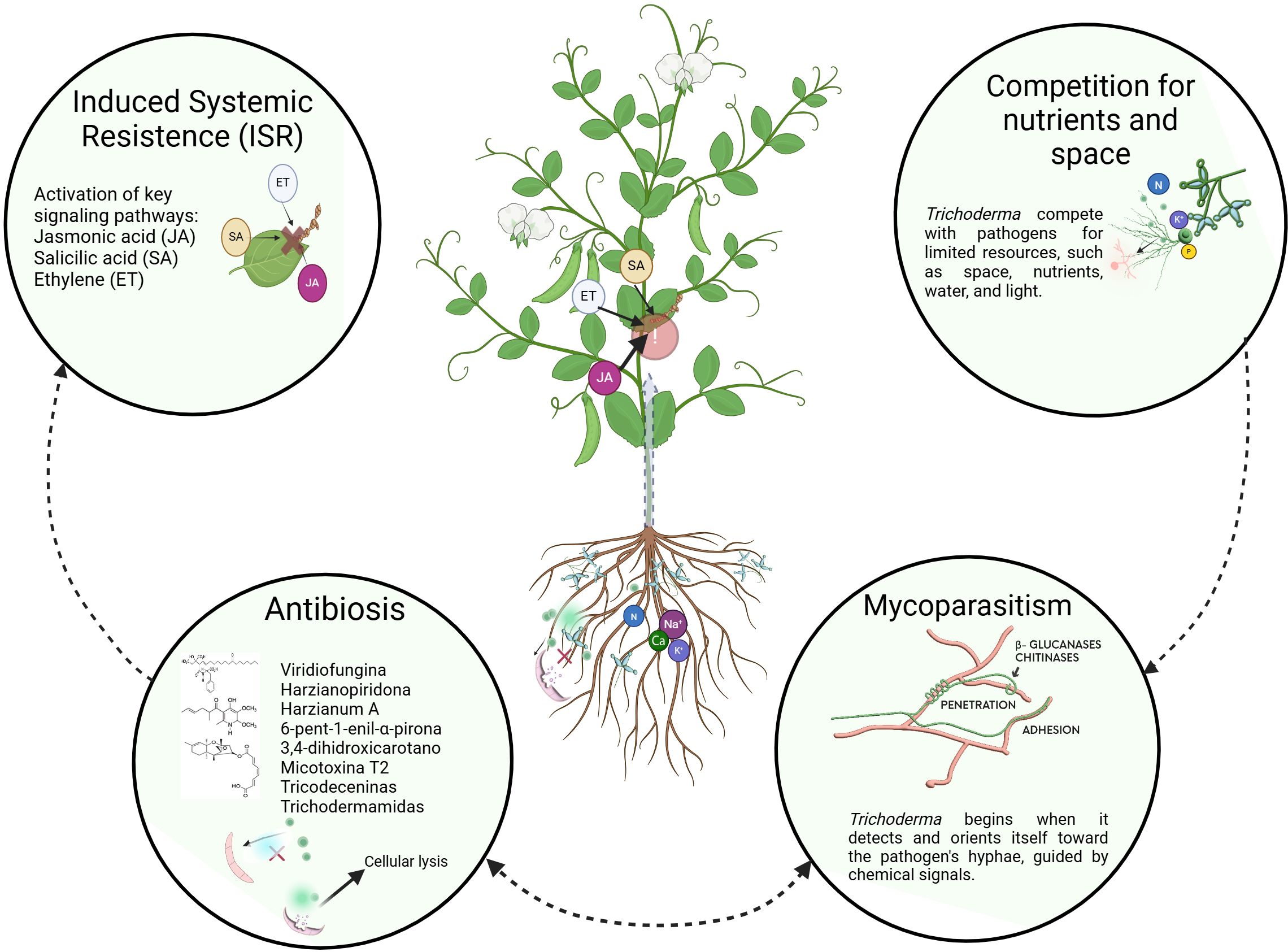
Figure 1. Biocontrol mechanisms used by Trichoderma spp. against plant pathogenic microorganisms.
3.1 Mycoparasitism
Trichoderma spp. employs a dual strategy to degrade the pathogen’s cell wall: enzymatic degradation and secondary metabolite production. It secretes extracellular enzymes such as chitinases, glucanases, and proteases (Cortés Hernández et al., 2023), breaking down structural components of the pathogen’s hyphae. In addition, bioactive compounds like peptaibols and gliotoxins further weaken the pathogen, enhancing penetration and colonization (Leiva et al., 2020; Pimentel et al., 2020). For example, T. asperellum produces chitinases and cellulases that significantly affect F. oxysporum in Stevia rebaudiana (Díaz-Gutiérrez et al., 2021). Similarly, T. virens has demonstrated effectiveness against R. solani, with electron microscopy revealing structural changes such as protuberances, coiling, and cell wall lysis (Inayati et al., 2020). Additionally, hyphal interactions between T. virens and Sclerotium rolfsii have been documented where T. virens acts targets sclerotia and conidia within the host fungal tissue (Mukherjee et al., 2022).
3.2 Competition for nutrients and space
Trichoderma spp. exerts biocontrol through competition by effectively competing with pathogens for limited resources such as space, nutrients, water, and light (Khan et al., 2020). Its rapid growth allows it to efficiently consume nutrients, such as carbohydrates, nitrogen, and oxygen, reducing their availability for pathogens (Hariharan et al., 2022). This ability to quickly colonize decreases the amount of nutrients accessible to phytopathogenic fungi, contributing to their control (Cortés Hernández et al., 2023).
Additionally, Trichoderma spp. colonizes the plant rhizosphere, forming a protective layer around the roots and preventing pathogen establishment (Basińska-Barczak et al., 2020; Oszust et al., 2020). T. harzianum has a growth rate 2.0 to 4.2 times faster than B. cinerea (Risoli et al., 2022), T. viride competes with Fusarium graminearum, the causative agent of fusariosis in cereals, through mechanisms such as adhesion, entanglement, and interpenetration of its hyphae, damaging the structures of F. graminearum with enzymes like chitinases and glucanases (Dugassa et al., 2021).
The competition for nutrients between Trichoderma spp. and plants can be significant, particularly in soils with low levels of nitrogen, phosphorus, or potassium. Under these conditions, Trichoderma spp. may deplete essential resources before plants can absorb them, which is especially concerning in degraded soils or those with poor fertilization management (Pedraza et al., 2020). Additionally, excessive application of Trichoderma spp. can lead to high concentrations in the soil, intensifying competition with plant roots and potentially hindering early growth (Sood et al., 2020). Environmental stress factors such as drought or salinity further influence this competition, limiting nutrient availability and reducing the efficiency of beneficial symbioses, such as those with mycorrhizal fungi (Xiao et al., 2023). Moreover, in soils with high microbial activity, Trichoderma may compete with other beneficial microorganisms for space and resources, potentially disrupting the soil microbiota and indirectly affecting plant health.
To mitigate these challenges, several strategies have been proposed. First, co-applying Trichoderma spp. with plant growth-promoting bacteria (PGPB) helps balance nutrient competition while maintaining its biocontrol efficacy (Pedraza et al., 2020). Second, using controlled-release formulations, such as encapsulated systems, enables a gradual release of Trichoderma spp., preventing excessive competition in the early stages of plant development and promoting a more stable soil establishment (Mukherjee et al., 2022). Additionally, optimizing fertilization practices ensures adequate levels of essential nutrients, reducing competition and allowing Trichoderma spp. to function effectively as a biocontrol agent without compromising plant growth (Xiao et al., 2023). Finally, selecting Trichoderma strains adapted to specific soil conditions and crop nutritional needs enhances pathogen suppression while minimizing negative effects on plant nutrient uptake (Baker et al., 2020).
3.3 Antibiosis
Trichoderma spp. produce a wide range of secondary metabolites with antimicrobial properties that inhibit the growth of pathogens (Masi et al., 2018). These compounds include peptaibols, polyketides, and terpenes (Cortés Hernández et al., 2023). T. virens produces trichodermamides, while T. koningii synthesizes koniginins, both of which exhibit antimicrobial activity against pathogens such as R. solani, F. oxysporum, Verticillium dahliae, and B. cinerea (Manganiello et al., 2018). T. atroviride produces the volatile compound 6-pentyl-2H-pyran-2-one (6-PP), which not only promotes plant growth but also regulates sugar transport in the roots of Arabidopsis (Esparza-Reynoso et al., 2021) and T. viride synthesizes trichotoxins A and B, tricodecenins, tricorovins, and tricocelins (Khan et al., 2020).
Among the bioactive compounds, Trichoderma also produces growth hormones such as auxins, cytokinins, and gibberellins, which stimulate plant development and overall growth (Estrada-Rivera et al., 2019). It activates ISR, enhancing the plant’s defense mechanisms against pathogens. Trichoderma’s ability to induce ISR in plants such as chili peppers and tomatoes against Colletotrichum truncatum and F. oxysporum (Yadav et al., 2021) strengthens the plant’s defense system, providing long-term protection beyond direct antagonism. T. asperellum is particularly effective in controlling Fusarium spp. and inducing ISR, especially in tomatoes and onions (Camacho-Luna et al., 2021). Strains like T. asperellum are known to improve nutrient uptake, particularly phosphorus, by solubilizing ions that are typically inaccessible in many soils (Bononi et al., 2020). This nutrient solubilization, combined with ISR, enhances the plant’s overall health, making it more resistant to both biotic and abiotic stresses. Another mechanism associated with the use of Trichoderma is its ability to produce antimicrobial compounds, lignify cell walls, and activate signaling pathways such as jasmonic acid (JA) and ethylene (ET), which are essential for the plant’s response to biotic stress (Alizadeh et al., 2020).
Together, the multiple mechanisms of action of Trichoderma from the production of antimicrobial compounds to the stimulation of induced defenses and the optimization of plant nutrition not only make it a highly effective biocontrol agent but also reduce reliance on synthetic agrochemicals. This promotes more sustainable agriculture by minimizing environmental impact and enhancing crop resilience against diseases and adverse conditions.
4 Mechanisms of action of Bacillus spp.
The main mechanisms of action of the Bacillus genus are production of lipopeptides, lytic enzymes, siderophores and induced systemic resistence (Figure 2).
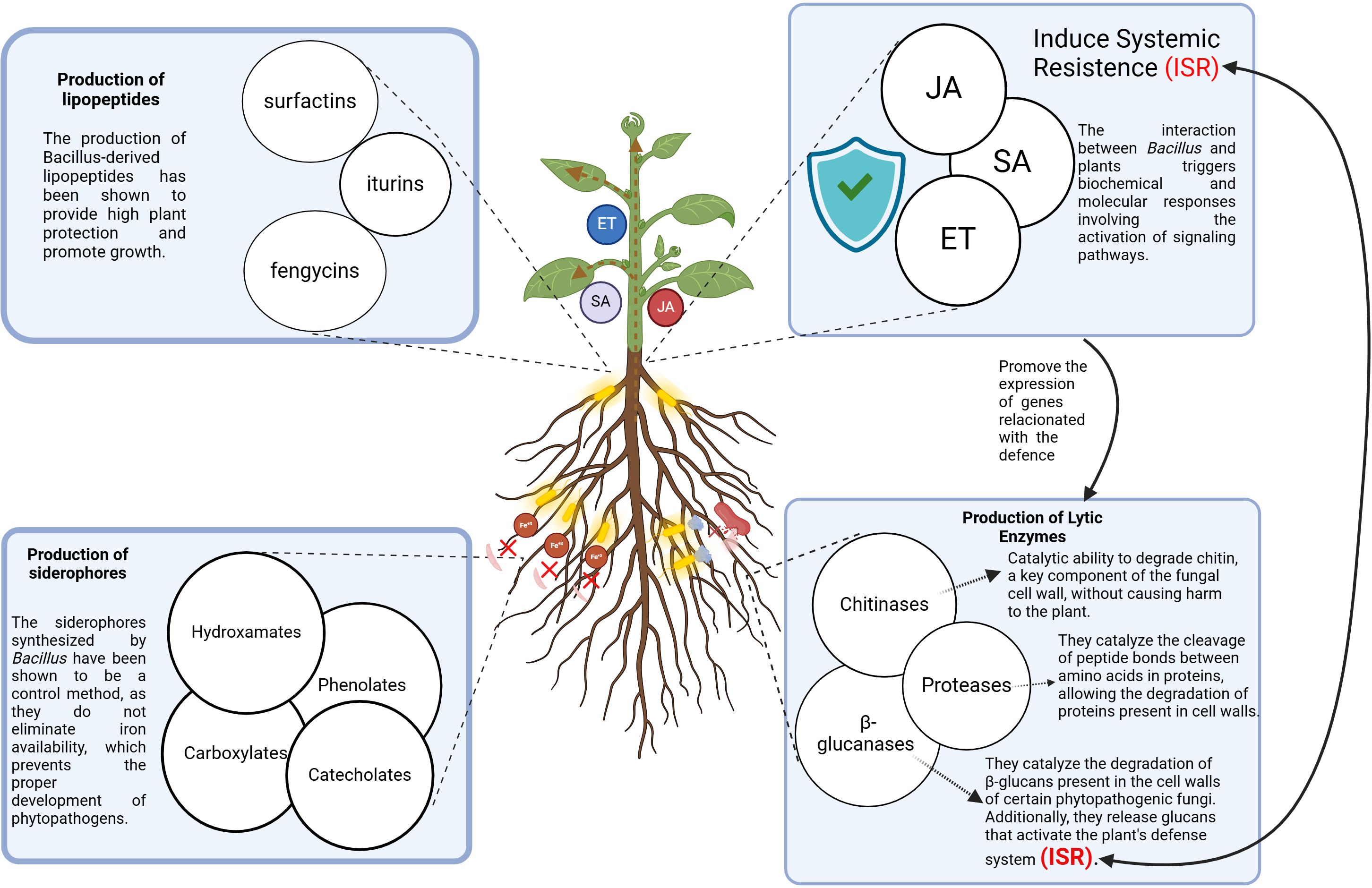
Figure 2. Biocontrol mechanisms used by Bacillus spp. against plant pathogenic microorganisms.
4.1 Production of lipopeptides
Lipopeptides synthesized by species of the genus Bacillus, such as B. subtilis, B. amyloliquefaciens, and B. velezensis, are classified into three main families: surfactins, iturins, and fengycins (Rabbee et al., 2019). These Bacillus-derived lipopeptides play crucial roles in both plant protection and growth promotion (Wang et al., 2024). They exhibit potent antifungal properties, effectively controlling a wide range of plant pathogens, including Monilinia fructicola, Fusarium spp., B. cinerea, Alternaria spp., and Colletotrichum gloeosporioides. They also induce systemic resistance in plants and promote biofilm formation, thereby enhancing rhizosphere antibacterial activity (Huang et al., 2019; Malviya et al., 2020).
The effectiveness of these lipopeptides can be attributed to their unique physicochemical properties. They disrupt lipid bilayers, reduce surface tension, and cause alterations in pathogen cell membranes. This disruption leads to the formation of pores, osmotic imbalance, and, ultimately the death of phytopathogenic microorganisms (Saiyam et al., 2024). Furthermore, Bacillus-derived lipopeptides stimulate plant defense mechanisms by activating systemic acquired resistance (SAR) and ISR, thereby boosting plant immunity (Valenzuela Ruiz et al., 2024).
Among the various lipopeptides, iturins, particularly Iturin A, are known for their ability to disrupt fungal plasma membranes. This action effectively controls plant pathogens and even extends the shelf-life of fruit juices, offering a natural alternative to chemical preservatives (Shi et al., 2018). Surfactins, which are biosurfactants produced by Bacillus spp., help form protective biofilms that inhibit microbial growth, making them valuable for reducing fruit and vegetable decay (Huang et al., 2019). Fengycins, including types A, B, and C, also demonstrate broad-spectrum antimicrobial activiy, inhibiting the growth of both pathogenic bacteria and fungi (Mnif and Ghribi, 2015).
Through the production of antimicrobial compounds, Bacillus spp. exhibit antagonistic mechanisms against various plant pathogens. For instance, B. amyloliquefaciens has shown strong antifungal properties against Rhizomucor variabilis in maize (Kulimushi et al., 2017), Similarly, B. subtilis produces antimicrobial mycosubtilin, which effectively controls F. graminearum in wheat and maize, while B. velezensis produces fengycin, iturin, and bacillomycin, which control Phytophthora sojae and Ralstonia solanacearum in soybean and tomato (Yu et al., 2021a).
In the case of Sclerotinia sclerotiorum, B. amyloliquefaciens suppresses its growth through the production of fengycin, inducing systemic resistance in crops such as rapeseed and tobacco (Farzand et al., 2019b). This highlights the importance of selecting the appropriate Bacillus spp. for specific pathogen control, as different strains produce distinct bioactive compounds with varying efficacy. The diversity of lipopeptide producing Bacillus spp. thus offers a valuable tool for integrated pest management and sustainable agriculture.
4.2 Production of lytic enzymes
Bacillus spp. produce various lytic enzymes, such as chitinases, β-glucanases, and proteases, which contribute to the biological control of phytopathogens. Their mechanism of action involves disrupting the structural integrity of pathogens cell walls by hydrolyzing specific components, ultimately leading to their disintegration (Hakim et al., 2021). These enzymes achieve this by breaking the glycosidic linkages that bind the structural polymers of the cell wall (Santoyo et al., 2021; Ajuna et al., 2023).
Among these enzymes, chitinases have gained significant attention due to their ability to degrade chitin, a component of the fungal cell wall, without harming the host plant (Kumar et al., 2018). Similarly, β-glucanases catalyze the breakdown of β-glucans, polysaccharides present in the cell walls of certain phytopathogenic fungi such as F. oxysporum (Won et al., 2018). Notably, these enzymes not only degrade fungal cells but also enhance Bacillus spp. competitiveness in the soil. Besides, the released glucan fragments can trigger plant defense responses, such as ISR and the production of phytoalexins. For instance, B. amyloliquefaciens FS6 produces a β-1,3-1,4-glucanase with antimicrobial activity against Alternaria panax and B. cinerea (Wang et al., 2021). Likewise, B. velezensis CE 100 synthesizes chitinase and β-1,3-glucanase, which degraded fungal cell walls and effectively inhibit the mycelial growth of M. phaseolina and F. oxysporum f. sp. fragariae, causal agents of charcoal rot and Fusarium wilt in strawberries β-1,3-glucanase (Hong et al., 2022). In addition to chitinases and β-glucanases, Bacillus spp. produces proteases, which catalyze the breakdown of peptide bonds in proteins, facilitating the degradation of fungal cell wall proteins (Rosazza et al., 2023). Beyond their antimicrobial function, these enzymes contribute to plant nutrition and may activate additional defense responses (Gray et al., 2019). An example of this is B. subtilis B315, which effectively reduced bacterial wilt in chili plants by 60.89% (Prihatiningsih et al., 2021).
4.3 Production of siderophores
Bacillus spp. is well known for its ability to produce siderophores, a variety of compounds in their competition for nutrients (Prasad et al., 2023). These molecules sequester ferric iron (Fe³+), giving Bacillus a competitive advantage over phytopathogens that lack siderophore production (Martínez-Canto et al., 2021). Notably, various Bacillus spp., including B. subtilis, B. pumilus, B. cereus, B. licheniformis, B. amyloliquefaciens, B. velezensis, B. thuringiensis, B. halodenitrificans, B. mojavensis, and B. atrophaeus, are known to produce siderophores, making them effective biocontrol agents against multiple phytopathogens (Andrić et al., 2020).
Several studies have demonstrated that siderophore-producing Bacillus strains can reduce the incidence of diseases caused by Fusarium spp. and R. solani in various plant species (Shafi et al., 2017; Dimopoulou et al., 2021). For instance, B. amyloliquefaciens synthesizes the catecholate siderophore bacilibactin under iron-limiting conditions, which effectively inhibits the growth of Pseudomonas syringae pv. tomato both in vitro and in plants (Nikolić et al., 2019). Similarly, B. subtilis MF497446 produces siderophores that not only restrict pathogen growth but also enhance resistance to Cephalosporium maydis in maize (Ghazy and El-Nahrawy, 2021).
4.4 Induced systemic resistence
Refers to a plant defense mechanism activated after interaction with specific microorganisms, such as Bacillus spp. This interaction triggers biochemical and molecular responses, involving the activation of jasmonic acid (JA), salicylic acid (SA), and ethylene (ET) signaling pathways (Shafi et al., 2017). Among the key ISR elicitors produced by Bacillus, cyclic lipopeptides (CLPs), N-alkylated benzylamine derivatives (NABD), and pyoverdines stand out. These compounds enhance plant immunity by activating defense mechanisms, thereby increasing resistance against various pathogens without exerting direct antimicrobial effects.
Several studies highlight the effectiveness of Bacillus in inducing ISR. For instance, B. thuringiensis enhances systemic resistance in soybean plants against Phytophthora sojae, strengthening defenses not only in the roots but throughout the entire plant (Basu et al., 2021). Similarly, B. velezensis PEA1 exhibits both antifungal and antiviral properties, effectively suppressing F. oxysporum and cucumber mosaic virus (CMV) in Datura stramonium leaves (Abdelkhalek et al., 2020). Furthermore, B. subtilis SL18r reduces foliar damage in tomato plants caused by Botrytis cinerea, a benefit attributed to its ability to activate the expression of a long non-coding RNA associated with defense pathways (Zhou et al., 2021).
The versatility of Trichoderma spp. and Bacillus spp. in their mechanisms of action underscores their potential as effective biocontrol agents within sustainable agriculture. Both genera exhibit a multifaceted approach to pathogen suppression, employing strategies such as direct antagonism through the production of antimicrobial compounds, competition for nutrients and space, and the activation of plant defense mechanisms. Notably, Bacillus spp. has shown superior adaptability in iron-limited environments through siderophore production, whereas Trichoderma spp. excels in mycoparasitism through enzymatic degradation of pathogen cell walls. Comparatively, Bacillus spp. tends to provide more immediate and widespread antimicrobial effects due to its production of various bioactive metabolites, including surfactins, iturins, fengycins, and siderophores. These compounds exhibit broad-spectrum activity against bacteria, fungi, and viruses, enhancing their utility across diverse agricultural systems. However, Trichoderma spp. is particularly effective in directly parasitizing phytopathogenic fungi through chitinase and glucanase activity, mechanisms that are often strain-specific but highly efficient when appropriately matched to the target pathogen. In adittion, while both microorganisms induce plant systemic resistance, the pathways activated differ slightly; Trichoderma spp. is more frequently associated with jasmonic acid and ethylene signaling, whereas Bacillus spp. often involves salicylic acid pathways as well.
The successful implementation of Trichoderma spp. and Bacillus spp. in agricultural systems requires careful consideration of their distinct mechanisms of action and environmental compatibility. Various factors, including soil composition, crop type, pathogen species, and climatic conditions, influence the efficacy of Trichoderma spp. and Bacillus spp. Moreover, the interaction between these biocontrol agents and native soil microbiota can significantly impact their performance. Farmers and policymakers must be aware that the indiscriminate or inappropriate application of these agents may yield inconsistent results or even counterproductive outcomes. Therefore, promoting an integrated approach that considers the specific attributes of each microorganism, supported by empirical evidence and tailored to local agricultural conditions, is important for optimizing their biocontrol potential. Ensuring the successful adoption of Trichoderma spp. and Bacillus spp. as biocontrol agents demands comprehensive research, clear guidelines, and educational efforts to inform practitioners about the conditions under which these microorganisms are most effective. Without such a nuanced understanding, their potential benefits may remain underutilized, compromising the progress toward more sustainable agricultural practices.
5 Laboratory scale, industrial scale-up, patents, and regulatory
The development of microbial fungicides follows a structured process to ensure their efficacy and safety in pest or disease control. The first step is identifying the target pathogen, which requires a deep understanding of its biology, life cycle, and environmental conditions that favor its proliferation (Basu et al., 2021). Once identified, a thorough review of previous studies is conducted to select microorganisms with proven effectiveness against the pathogen.
After selecting potential BCAs, they are isolated and cultured in the laboratory, where their efficacy is evaluated under controlled conditions. This evaluation considers various mechanisms of action to determine their potential (Zandi and Basu, 2016). For instance, it is important to assess the specificity of the BCA to ensure it does not affect non-target organisms, especially beneficial ones. Laboratory and field tests further validate their ability to reduce pest populations or disease severity (Jangir et al., 2021). Furthermore, compatibility with the agricultural environment, including climate, crop types, and agronomic practices, must be considered to enhance field performance (Teixidó et al., 2022).
Once a suitable BCA is identified, production begins through fermentation methods, primarily submerged fermentation (SMF) or solid-state fermentation (SSF) (Teixidó et al., 2022). SMF is preferred for bacteria and yeasts due to its high yield, ease of nutrient control, and cost-effectiveness. In contrast, SSF is more suitable for filamentous fungi, which struggles to sporulate in liquid media. To optimize fermentation, key parameters such as temperature, pH, agitation, aeration, inoculum concentration, and duration must be carefully adjusted (Vehapi et al., 2023). However, SSF presents challenges in nutrient addition due to its low water activity, making process control more complex (Fahim et al., 2013). The culture medium composition plays a critical role in microbial growth and viability. It typically includes inorganic (ammonium and nitrate salts) or organic nitrogen sources (amino acids, proteins, or urea), carbon sources such as simple sugars (sucrose, fructose), and complex carbohydrates (starch). Besides that, agro-industrial by-products like molasses, whey, yeast extracts, and peptones are commonly used to enhance growth (Clerici et al., 2021).
Following fermentation, the next step is formulating the BCA into a usable product. Various formulations have been developed for Trichoderma spp., including granules, pellets, wettable powders, capsules, water-dispersible granules, and emulsifiable liquids (Locatelli et al., 2018). However, some of these formulations pose challenges in application, affecting dosing precision and potentially increasing health risks for applicators (Ubando et al., 2020). In this context, Jangir et al., 2021, developed water-dispersible granules of B. subtilis and T. harzianum using response surface methodology (RSM) to control F. oxysporum, providing a more user and environmentally friendly option. Similarly, B. megatherium has been formulated in alginate microcapsules, pellets, or effervescent systems to enhance bacterial release (Wiwattanapatapee et al., 2013). In addition, microbial preparations can be combined with organic amendments to improve soil quality in semi-arid regions (Fendrihan et al., 2016).
Regarding formulation types, liquid formulations are often preferred due to their cost-effectiveness and ease of handling (Palmieri et al., 2022). They typically consist of an active ingredient, either microbial cells or their secondary metabolites combined with a carrier that ensures even distribution and influences release and stability (Berninger et al., 2018). Water is the most used carrier, often supplemented with additives to maintain stability during application (Bashan et al., 2014). Alternatively, oil-based formulations can be developed, provided they are non-toxic to microorganisms, plants, humans, or animals (Palmieri et al., 2022).
On the other hand, solid formulations offer advantages in storage and transport due to their stability and cost-effectiveness. Several dehydration methods are used to preserve microbial viability. Freeze-drying is particularly suitable for heat- and moisture-sensitive materials (Prakash et al., 2013). Spray drying rapidly converts liquid suspensions into powder by atomizing droplets in a hot air chamber (150°C–250°C), offering a fast and scalable process (Teixidó et al., 2022). Also, fluidized bed drying, is employed for granulating and coating, where particles are suspended in hot air to ensure uniform drying, often with cryoprotectants to enhance stability (Chaudhary et al., 2020; Teixidó et al., 2022). A combination of spray drying and fluidized bed drying can further improve uniformity and prevent clumping (Torres et al., 2014).
Scaling up production begins in the laboratory, where Erlenmeyer flasks are used to optimize culture media composition. The process then transitions to small bioreactors (2.0–5.0 liters) for further refinement of growth conditions. At an industrial scale, production is conducted in pilot plant bioreactors with capacities ranging from 100 to 300 liters (Fahim et al., 2013). Once fermentation is complete, the next phase involves recovering the microbial cells, spores, or supernatant containing secreted metabolites, depending on the BCA type. For instance, in B. amyloliquefaciens, it is recommended to recover both cells and metabolites, as both contribute to disease control (Gotor-Vila et al., 2017). Ensuring the viability and stability of the BCA throughout production, storage, and application for its effectiveness. Colony-forming unit (CFU) tests are commonly used to determine viable microorganism counts per milliliter or gram of product (Meyer et al., 2023). A sufficient concentration of viable cells is crucial to achieving the desired biological effects in the field. Furthermore, fungal spores must maintain a high germination rate, as prolonged storage or adverse conditions can reduce their viability and efficacy (Palmieri et al., 2022). Beyond cell viability, metabolic activity must also be assessed to confirm that the microorganisms remain functional. Measuring ATP (adenosine triphosphate) production serves as a reliable indicator of cellular energy and viability (Carrodeguas-González et al., 2022). Ensuring metabolic activity guarantees that BCAs will perform effectively upon application.
After production, proper storage conditions are critical to preserving product shelf life. Most BCAs require refrigeration (4°C–10°C) to maintain microbial viability, though some formulations remain stable at room temperature depending on the organism and formulation used (Alamprese et al., 2017). Humidity control is equally important, as excess moisture can degrade powder or granule formulations, leading to contaminant growth or premature activation.
Finally, the commercialization of a BCA involves regulatory and patenting processes. Patent applications typically grant 20 years of protection to inventions that meet criteria for novelty, non-obviousness, and industrial applicability. A complete description of the BCA, including its production method and biological mechanisms, is required. Additionally, regulatory compliance varies by country; for instance, BCAs must be approved by entities such as the U.S. Environmental Protection Agency (EPA) in the United States or the European Union (EU) regulatory bodies before commercialization (Dourado et al., 2016). Proper labeling is also mandated, providing essential details on composition, application methods, safety precautions, and usage instructions (Palmieri et al., 2022). By following this structured approach from pathogen identification to formulation, production, quality control, storage, and regulatory approval, BCAs can be successfully developed and commercialized as effective alternatives for sustainable disease and pest management. Indeed, the market includes approximately 2,920 bacterial-based biopesticides, 1,658 fungal-based biopesticides, and 234 viral-based biopesticides. Of these, 227 are bacterial and 169 are fungal fungicides (Palmieri et al., 2022). Notably, 77 Trichoderma-based fungicides are available globally, with 7 approved by the European Commission for use in EU member states (Thambugala et al., 2020). These products demonstrate the rigorous regulatory approval processes that often involve high costs and lengthy approval periods. For instance, in countries like the U.S. and EU member states, BCAs must comply with strict manufacturing regulations and are subject to regular audits of production facilities. In contrast, many developing countries may have less stringent manufacturing controls, which raise concerns about product quality and effectiveness. These regulatory processes often include additional testing for safety, efficacy, and product stability. However, in countries with more flexible regulatory frameworks, such as some in Asia and Latin America, the registration process for BCAs may be quicker and less expensive, but this can come at the cost of reduced guarantees of product quality. Regulatory differences across countries and regions further complicate the international marketing and use of BCAs, so these challenges must be addressed carefully (Vekemans and Marchand, 2020).
Undoubtedly, this information offers valuable insights into how the efficacy and safety of these products are ensured within agricultural practices. For farmers, such knowledge is essential, empowering them to make well-informed decisions about selecting microbial fungicides based on crop compatibility, environmental impact, and field performance. Moreover, understanding optimal storage and application conditions allows them to maximize yield and enhance productivity. Beyond practical applications, this information highlights the scientific rigor underpinning the development of biological products, reinforcing that food produced through these methods is both safe and sustainable. It also fosters greater trust in biological alternatives to conventional agrochemicals, promoting a healthier and more environmentally responsible agricultural system.
6 Challenges and future perspectives in the use of BCAs in agriculture
The positive effects of BCAs in the field can vary significantly due to environmental conditions, competition with other microorganisms, and the genetic variability of pathogens. For example, high UV radiation, extreme temperatures, or high humidity can hinder BCA performance after application, limiting their effectiveness in certain regions or for specific crops (Umer et al., 2021). For this, the BCAs should be collected and screened to achieve relevant biocontrol results (Yu et al., 2021b). In addition to environmental limitations, scaling up BCA production presents another challenge. This is primarily due to the costs associated with microorganism propagation under optimal conditions and the need for suitable formulation technologies to ensure their efficacy. Formulations, such as controlled release or encapsulated systems, are necessary for maintaining prolonged stability in the field (Locatelli et al., 2018).
Beyond production challenges, another limitation lies in the interaction of BCAs with other organisms within the agroecosystem, such as pollinators, beneficial insects, and soil microorganisms. Understanding these interactions is vital to minimizing negative impacts and maximizing the benefits of BCAs (Palmieri et al., 2022). However, there is a lack of detailed information on these interactions. The available literature generally focuses on the ecological importance of relationships between plants, microorganisms, and insect communities, with some suggestions that negative interactions may arise when certain microorganisms or insects become parasites affecting plants (Barolia et al., 2023). On the other hand, beneficial microorganisms in the rhizosphere can improve plant health and indirectly support pollinators by enhancing nectar quality. For instance, Lactobacillus kunkeei, found in flowers, inhibits pathogens affecting bees, highlighting a positive interaction. In contrast, soil pathogens can harm plant health and reduce nectar availability for pollinators. This microbial cycle between soil, plants, and pollinators underscores the interconnectedness of ecosystem health (Liu et al., 2019).
Looking to the future of microorganisms such as BCAs offer a promising sustainable alternative to chemical pesticides. These agents address environmental pollution and biodiversity loss, as they do not negatively affect non-target organisms. Unlike chemical pesticides, which can lead to pathogen resistance over time, BCAs have multiple modes of action, reducing the likelihood of resistance development. For example, Trichoderma reduces pathogen resistance by producing secondary metabolites like peptaibols and gliotoxin, which exhibit antifungal activity. Besides, it triggers plant defense mechanisms through elicitors, enhancing systemic resistance and promoting mutualistic associations that improve nutrient uptake and plant health (Orozco-Mosqueda et al., 2023).
Utilizing specific Trichoderma isolates, such as T. afroharzianum TRI07, can effectively inhibit pathogens like A. alternata, enhance tomato plant defense mechanisms, and promote the production of defense-related enzymes and phenolic compounds (Philip et al., 2024). Similarly, B. subtilis, B. mojavensis, and B. velezensis reduce pathogen resistance of F. oxysporum by producing hydrolytic enzymes and secondary metabolites that inhibit pathogen growth. These microorganisms also promote root development and create an unfavorable rhizosphere environment for pathogens, ultimately improving overall plant health and resilience (Auriza Rumandani, 2025).
Considering growing concerns about food safety, there is increasing demand for safer pest control methods that do not leave harmful residues on crops. One of the primary factors influencing consumer acceptance of BCAs is their level of awareness about these agents. Research has shown that individuals who are more informed about BCAs tend to have a more positive perception of their use. Trichoderma has gained acceptance due to its specificity, safety for beneficial organisms, and absence of toxic residues or environmental risks (Sharma and Gothalwal, 2017). This indicates a positive trend toward acceptance among producers and consumers, as sustainable agricultural practices and biological control methods continue to be prioritized within IPM. However, despite these benefits, adoption remains limited in developed countries due to a general lack of awareness (Kumar Chaube and Pandey, 2022).
Biotechnological advances improving the efficacy, stability, and ease of use of BCAs include the development of specialized formulations using protective substances like sugars and polyols, which enhance stability during storage. Techniques such as freeze-drying and fluidized bed-spray-drying are employed to maintain viability (Chaudhary et al., 2020). The optimized packaging conditions, including vacuum packaging and modified atmospheres, help extend shelf life (Kumar et al., 2023). These innovations facilitate the mass production and application of BCAs, making them more viable alternatives to synthetic fungicides (Teixidó et al., 2022).
Moreover, other studies analyze advances in transformation tools for genetic improvement of Trichoderma, enhancing biomass, primary and secondary metabolite production, and enzyme activity, thereby improving efficacy and adaptability in agricultural applications, particularly against challenges posed by climate change and pathogens (Villao-Uzho et al., 2024). One of the most common techniques for genetic transformation is protoplast-mediated transformation using Agrobacterium tumefaciens (Wang et al., 2019). Early methods for transforming Trichoderma involved techniques such as electroporation and biolistics (Schuster et al., 2016). The CRISPR (regularly interspaced clustered short palindromic repeat)-Cas9 (CRISPR-related nuclease 9) system, has many innovative applications in Trichoderma (Wang et al., 2022). CRISPR allows the reduction of undesirable traits and the introduction of new, desirable traits that can target a wide range of pathogens and pests in various cropping systems (Tyagi et al., 2024). Analyzes dedicated to the function of the genes of the beneficial Trichoderma spp. are generally attributed to the construction of recombinant DNA using conventional cloning techniques, which are based on digestion and ligation procedures (Fitz et al., 2018). The genomic organization, together with the regulation and enzymatic hydrolytic expression in the T. harzianum strain IOC3844, are closely related, which helps to increase plant degradation (Ferreira Filho et al., 2020). The layer-by-layer (LbL) encapsulation method using biobased lignin derivatives significantly enhances the stability and efficacy of Trichoderma spores, protecting them from extreme temperatures and UV exposure, thus improving their performance as biocontrol agents in sustainable agriculture (Løvschall et al., 2024). Biopolymer-based emulsions, particularly those using agar and cellulose nanocrystals, enhance the stability and viability of T. atrobrunneum conidia, allowing for prolonged storage and effective encapsulation, thus improving their efficacy (Martínez et al., 2023). Chemical mutagenesis using Ethyl Methyl Sulphonate (EMS) and gamma irradiation have enhanced the biocontrol capabilities of T. viride, resulting in stable mutants with improved antagonistic activities against soil-borne pathogens, thus increasing efficacy and ease of use in agricultural applications (Shewarega Foyate, 2024).
The genetic improvement of Bacillus spp. has enabled the identification of key genes that contribute to plant growth promotion and biopesticide production. Advances in genetic engineering have facilitated the development of strategies to optimize the production of enzymes, chemicals, and antimicrobial compounds, making Bacillus more competitive in the biotechnology industry (Mitra et al., 2021; Muras et al., 2021). One of the most extensively studied cases is B. thuringiensis, widely used in biopesticide development. Recombinant DNA techniques have enhanced its insecticidal potency, expanded its spectrum of action, improved its persistence in crops, and increased its fermentation yield (Ortiz et al., 2024). A notable example is the genetic modification of crops to express cry genes, which have been introduced into potatoes, tomatoes, tobacco, rice, corn, and broccoli to provide resistance against agricultural pests (Abbas, 2018). However, this practice has sparked controversy regarding its safety and potential impacts on human health.
Advancements in genetic engineering have also benefited other Bacillus species. For instance, in B. amyloliquefaciens, the manipulation of regulatory genes such as codY, comA, degU, and spo0A has proven to be an effective strategy for enhancing the production of antimicrobial lipopeptides (bacillomycin D, fengycin, and surfactin), resulting in strains with high potential for industrial applications (Sun et al., 2021). The application of advanced technologies, such as CRISPR, has revolutionized the genetic modification of Bacillus spp. It has been used in B. subtilis to enhance the production of the terpenoid artemisinic acid (Song et al., 2021) and in B. licheniformis to improve its production capacity (Zhou et al., 2019). Furthermore, genetic engineering allows not only gene modification but also the manipulation of their products, as seen in the case of B. thuringiensis Cry toxins, whose optimization has increased their insecticidal efficiency.
Biological products are increasingly integrated into existing agricultural systems as part of IMP strategies. This integration is particularly effective when combined with other sustainable agricultural practices, such as crop rotation, the use of resistant crops, or the application of organic fertilizers. These practices not only enhance the efficiency of biological control but also reduce reliance on chemical products (Baker et al., 2020). Moreover, when BCAs are integrated with other products like bactericides and organic fertilizers, their effectiveness is strengthened, contributing to crop sustainability and boosting agricultural productivity. By carefully considering interactions with the existing biota, the ecological functions of the system are optimized, leading to more efficient and environmentally friendly pest and disease control (Vasquez-Lopez et al., 2021). In the case of Trichoderma, studies have shown that it is not only safe for the soil ecosystem but also helps maintain its balance and sustainability (Li et al., 2024). From an ecological perspective, Trichoderma is not considered an aggressive soil colonizer that disrupts existing microbiota. Instead, its mode of action is based on competition, which does not harm other beneficial microorganisms (Zin and Badaluddin, 2020). Given that Trichoderma is naturally found in soils, it serves to strengthen, rather than alter, the biological mechanisms in the soil. Importantly, it does not leave toxic residues or affect soil pH, making it an ecologically sustainable alternative (Zafra et al., 2015).
Based on the aforementioned, despite the ecological benefits of BCAs, their commercialization can be more costly compared to conventional chemical pesticides, which limit their adoption, particularly in resource-limited countries (Droby et al., 2016). These high production costs are attributed to the controlled and specific conditions required for multiplying microorganisms like Trichoderma and Bacillus. Such processes involve specialized facilities, expensive growth media, and highly skilled personnel (Palmieri et al., 2022). For this reason, there is a need to assess the impact of these organisms in a holistic manner, given the increasing demand for sustainable alternatives that has driven significant growth in the biofungicide market in recent years. This approach is supported by ongoing research demonstrating the effectiveness of microbial biocontrol agents in diverse crops. For instance, a study evaluated the effect of Bacillus and Trichoderma species in managing bacterial wilt in tomatoes (Lycopersicon esculentum). The Bacillus CB64 and Trichoderma T1 isolates achieved significant reductions in disease incidence and severity in the field, as well as a decrease of over 90% in the soil pathogen population (Kariuki et al., 2020). Additionally, research on bananas has shown that strains of B. licheniformis CSR-D4 significantly reduce the incidence of wilt caused by F. oxysporum f. sp. cubense in banana plants. The bioactive metabolites produced by this strain inhibited pathogen invasion, demonstrating its potential as an effective biocontrol agent (Izquierdo-García et al., 2024). Therefore, improving the connection and relationship between these findings highlights the importance of adopting integrated biocontrol strategies to enhance agricultural sustainability.
7 Conclusion
The agri-food industry bears a significant responsibility to meet global food demand with products that are not only nutritious and of high quality but also safe. As highlighted throughout this review, BCAs such as Trichoderma and Bacillus offer a viable and sustainable alternative to conventional chemical pesticides. These beneficial microorganisms have demonstrated their ability to enhance crop productivity, promote soil health, and improve resilience against various diseases, thereby contributing to eco-friendly agricultural systems. Understanding the mechanisms of action, selection criteria, formulation processes, and challenges associated with BCAs is essential for optimizing their efficacy in diverse agricultural environments. This knowledge empowers farmers to select appropriate formulations that align with specific environmental and soil conditions, enhancing their productivity while reducing dependency on synthetic agrochemicals. Additionally, integrating BCAs with other sustainable practices can further bolster agricultural resilience and contribute to more environmentally responsible food production. Moreover, from a consumer perspective, BCAs present a safer alternative by reducing the presence of toxic residues in food products. Raising awareness and providing education about these biocontrol agents are essential steps in fostering their acceptance and encouraging wider adoption. Informed consumers are more likely to support sustainable agricultural solutions, which can drive market growth and innovation in this field. On a broader scale, challenges such as regulatory disparities between countries, economic barriers to production, and commercialization hurdles remain significant. Addressing these obstacles through collaborative efforts between researchers, industry, and policymakers is crucial to achieving the large-scale adoption of BCAs. Thus, this review has underscored the importance of developing strategies that harmonize regulatory frameworks and enhance economic feasibility across different agricultural contexts. While the positive impacts of BCAs on productivity and sustainability are evident, further research is needed to optimize formulations, improve application techniques, and assess long-term efficacy under various agroecological conditions. Continued efforts in these areas will be essential to unlocking the full potential of biological control agents and ensuring their successful integration into mainstream agricultural practices.
Author contributions
MV: Data curation, Formal Analysis, Investigation, Writing – original draft. FE: Data curation, Formal Analysis, Writing – original draft. LE: Supervision, Validation, Writing – review & editing. JC: Conceptualization, Data curation, Resources, Supervision, Validation, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The authors acknowledge the Biotechnology Research Center of Ecuador (CIBE) for supporting founding, Project CIBE-5-2023.
Conflict of interest
The authors declare that the review was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fagro.2025.1578915/full#supplementary-material
References
Abbas M. S. T. (2018). Genetically engineered (Modified) crops (bacillus thuringiensis crops) and the world controversy on their safety. Egyptian J. Biol. Pest Control 28, 1–12. doi: 10.1186/s41938-018-0051-2
Abdelkhalek A., Behiry S. I., Al-Askar A. A. (2020). Bacillus velezensispea1 inhibits fusarium oxysporum growth and induces systemic resistance to cucumber mosaic virus. Agronomy 10, 1312. doi: 10.3390/agronomy10091312
Abro M. A., Sun X., Li X., Jatoi G. H., Guo L. D. (2019). Biocontrol potential of fungal endophytes against Fusarium oxysporum f. Sp. cucumerinum causing wilt in cucumber. Plant Pathol. J. 35, 598–608. doi: 10.5423/PPJ.OA.05.2019.0129
Ajuna H. B., Lim H., Moon J., Won S., Choub V., Choi S., et al. (2023). The prospect of hydrolytic enzymes from Bacillus species in the biological control of pests and diseases in forest and fruit tree production. Int. J. Mol. Sci. 24, 16889. doi: 10.3390/ijms242316889
Alamprese C., Cappa C., Ratti S., Limbo S., Signorelli M., Fessas D., et al. (2017). Shelf life extension of whole-wheat breadsticks: Formulation and packaging strategies. Food Chem. 230, 532–539. doi: 10.1016/j.foodchem.2017.03.092
Alejandro C. M., Hernandez-Morales J., Toledo-Aguilar R., Sabino Lopez J. E., Romero-Rosales T. (2019). Capacidad Antagónica de Trichoderma spp. Nativa contra Phytophthora parasitica y Fusarium oxysporum Aislados De Cultivos De Jamaica. Rev. Fitotec. Mex. 42, 235–241.
Alizadeh M., Vasebi Y., Safaie N. (2020). Microbial antagonists against plant pathogens in Iran: A review. Open Agric. 5, 404–440. doi: 10.1515/opag-2020-0031
Anastasiou E., Fountas S., Voulgaraki M., Psiroukis M., Koutsiaras M., Kriezi O., et al. (2023). Precision farming technologies for crop protection: A meta-analysis. Smart Agric. Technol. 5, 100323. doi: 10.1016/j.atech.2023.100323
Andrade-Hoyos P., Silva-Rojas H. V., Romero-Arenas O. (2020). Endophytic Trichoderma species isolated from Persea americana and Cinnamomum verum roots reduce symptoms caused by Phytophthora cinnamomi in avocado. Plants 9, 1–17. doi: 10.3390/plants9091220
Andrić S., Meyer T., Ongena M. (2020). Bacillus responses to plant-associated fungal and bacterial communities. Front. Microbiol. 11. doi: 10.3389/fmicb.2020.01350
Auriza Rumandani N. (2025). The Utilization of Bacillus spp. As endhophytes In enhancing plant resistance and health against Fusarium oxysporum: a mini review. Syntax Literate 10 (1), 490–503. doi: 10.36418/syntax-literate.v10i1.56149
Baiyee B., Pornsuriya C., Ito S. I., Sunpapao A. (2019). Trichoderma spirale T76–1 displays biocontrol activity against leaf spot on lettuce (Lactuca sativa L.) caused by Corynespora cassiicola or Curvularia aeria. Biol. Control 129, 195–200. doi: 10.1016/j.biocontrol.2018.10.018
Baker B. P., Green T. A., Loker A. J. (2020). Biological control and integrated pest management in organic and conventional systems. Biol. Control 140, 104095. doi: 10.1016/j.biocontrol.2019.104095
Barolia S. K., Sharma P., Ansari Z. A., Kumar S. (2023). “Three-way interactions between plants, microbes and insects communities,” in Advanced Research in Biological Science Vol. Vol. 4. (India: B P International (a part of SCIENCEDOMAIN International), 144–157. doi: 10.9734/bpi/arbs/v4/7040a
Bashan Y., de-Bashan L. E., Prabhu S. R., Hernandez J. P. (2014). Advances in plant growth-promoting bacterial inoculant technology: Formulations and practical perspectives, (1998-2013). Plant Soil 378, 1–33. doi: 10.1007/s11104-013-1956-x
Basińska-Barczak A., Błaszczyk L., Szentner K. (2020). Plant cell wall changes in common wheat roots as a result of their interaction with beneficial fungi of trichoderma. Cells 9, 1–28. doi: 10.3390/cells9102319
Basu A., Prasad P., Das S. N., Kalam S., Sayyed R. Z., Reddy M. S., et al. (2021). Plant growth promoting rhizobacteria (Pgpr) as green bioinoculants: Recent developments, constraints, and prospects. Sustain. (Switzerland) 13, 1–20. doi: 10.3390/su13031140
Berninger T., González López Ó., Bejarano A., Preininger C., Sessitsch A. (2018). Maintenance and assessment of cell viability in formulation of non-sporulating bacterial inoculants. Microb. Biotechnol. 11, 277–301. doi: 10.1111/1751-7915.12880
Beyuo J., Sackey L. N. A., Yeboah C., Kayoung P. Y., Koudadje D. (2024). The implications of pesticide residue in food crops on human health: a critical review. Discov. Agric. 2, 123. doi: 10.1007/s44279-024-00141-z
Blazhevska V. (2019). Growing at a slower pace, world population is expected to reach 9.7 billion in 2050 and could peak at nearly 11 billion around 2100. Available online at: https://www.un.org/sustainabledevelopment/blog/2019/06/growing-at-a-slower-pace-world-population-is-expected-to-reach-9-7-billion-in-2050-and-could-peak-at-nearly-11-billion-around-2100-un-report/ (Accessed June 11, 2024).
Bononi L., Chiaramonte J. B., Pansa C. C., Moitinho M. A., Melo I. S. (2020). Phosphorus-solubilizing Trichoderma spp. from Amazon soils improve soybean plant growth. Sci. Rep. 10, 2858. doi: 10.1038/s41598-020-59793-8
Camacho-Luna V., Flores-Moctezuma H. E., Rodríguez-Monroy M., Montes-Belmont R., Sepúlveda-Jiménez G. (2021). Induction of the defense response of onion plants in interaction with Trichoderma asperellum and Alternaria porri. Rev. Mexicana Cienc. Agrícolas 12, 685–698. doi: 10.29312/remexca.v12i4.2683
Carrodeguas-González A., Zúñiga-Orozco A., Ortíz-Cruz M. V. (2022). Ensayos in vitro para cuantificar la actividad biológica de citocinas. Rev. Tecnol. en Marcha. doi: 10.18845/tm.v35i2.5638
Chaudhary T., Dixit M., Gera R., Shukla A. K., Prakash A., Gupta G., et al. (2020). Techniques for improving formulations of bioinoculants. 3 Biotech. 10, 199. doi: 10.1007/s13205-020-02182-9
Chen Y. H., Lee P. C., Huang T. P. (2021). Biological control of collar rot on passion fruits via induction of apoptosis in the collar rot pathogen by Bacillus subtilis. Phytopathology 111, 627–638. doi: 10.1094/PHYTO-02-20-0044-R
Clerici N. J., Lermen A. M., Daroit D. J. (2021). Agro-industrial by-products as substrates for the production of bacterial protease and antioxidant hydrolysates. Biocatal. Agric. Biotechnol. 37, 102174. doi: 10.1016/j.bcab.2021.102174
Contreras-Cornejo H. A., Macías-Rodríguez L., del-Val E., Larsen J. (2018). The root endophytic fungus Trichoderma atroviride induces foliar herbivory resistance in maize plants. Appl. Soil Ecol. 124, 45–53. doi: 10.1016/j.apsoil.2017.10.004
Cortés Hernández F., del C., Alvarado Castillo G., Sanchez Viveros G. (2023). Trichoderma spp., una alternativa para la agricultura sostenible: una revisión. Rev. Colombiana Biotecnol. 25, 62–76. Available at: https://revistas.unal.edu.co/index.php/biotecnologia/article/view/111384 ( Accessed March 28, 2025).
Damalas C. A., Koutroubas S. D. (2018). Current status and recent developments in biopesticide use. Agric. (Switzerland) 8, 13. doi: 10.3390/agriculture8010013
Díaz-Gutiérrez C., Arroyave C., Llugany M., Poschenrieder C., Martos S., Peláez C. (2021). Trichoderma asperellum as a preventive and curative agent to control Fusarium wilt in Stevia rebaudiana. Biol. Control 155, 104537. doi: 10.1016/j.biocontrol.2021.104537
Dimopoulou A., Theologidis I., Benaki D., Koukounia M., Zervakou A., Diallinas G., et al. (2021). Direct antibiotic activity of bacillibactin broadens the biocontrol range of Bacillus amyloliquefaciens MBI600. mSphere 6 (4), 37621. doi: 10.1128/msphere.00376-21
Dourado F., van den Berg C., Gama M. (2016). “European regulatory framework on novel foods and novel food additives,” in Bacterial Nanocellulose: From Biotechnology to Bio-Economy (Luxembourg: Elsevier Inc), 135–144. doi: 10.1016/B978-0-444-63458-0.00008-1
Droby S., Wisniewski M., Teixidó N., Spadaro D., Jijakli M. H. (2016). The science, development, and commercialization of postharvest biocontrol products. Postharvest Biol. Technol. 122, 22–29. doi: 10.1016/j.postharvbio.2016.04.006
Dugassa A., Alemu T., Woldehawariat Y. (2021). In-vitro compatibility assay of indigenous Trichoderma and Pseudomonas species and their antagonistic activities against black root rot disease (Fusarium solani) of faba bean (Vicia faba L.). BMC Microbiol. 21, 115. doi: 10.1186/s12866-021-02181-7
Elhamalawy O., Bakr A., Eissa F. (2024). Impact of pesticides on non-target invertebrates in agricultural ecosystems. Pesticide Biochem. Physiol. 202, 105974. doi: 10.1016/j.pestbp.2024.105974
Escudero-Leyva E., Alfaro-Vargas P., Muñoz-Arrieta R., Charpentier-Alfaro C., Granados-Montero M., Valverde-Madrigal K., et al. (2022). Tolerance and biological removal of fungicides by Trichoderma species isolated from the endosphere of wild rubiaceae plants. Front. Agron. 3, 772170. doi: 10.3389/fagro.2021.772170
Esparza-Reynoso S., Ruíz-Herrera L., Pelagio-Flores R., Macías-Rodríguez L., Martínez-Trujillo M., López-Coria M., et al. (2021). Trichoderma atroviride-emitted volatiles improve growth of Arabidopsis seedlings through modulation of sucrose transport and metabolism. Plant Cell Environ. 44, 1961–1976. doi: 10.1111/pce.14014
Espinoza Ahumada C. A., Gallegos Morales G., Hernández Castillo F. D., Ochoa Fuentes Y. M., Cepeda Siller M., Castillo Reyes F. (2019). Antagonistas microbianos a Fusarium spp., como agente causal de pudrición de raíces y tallo en melón. Ecosistemas y Recursos Agropecuarios 6, 45–55. doi: 10.19136/era.a6n16.1843
Es-Soufi R., Tahiri H., Azaroual L., Oualkadi A., Martin P., Badoc A., et al. (2020). In Vitro Antagonistic Activity of Trichoderma harzianum and Bacillus amyloliquefaciens against Colletotrichum acutatum. Adv. Microbiol. 10, 82–94. doi: 10.4236/aim.2020.103008
Estrada-Rivera M., Rebolledo-Prudencio O. G., Pérez-Robles D. A., Rocha-Medina M. D. C., González-López M. D. C., Casas-Flores S. (2019). Trichoderma histone deacetylase HDA-2 modulates multiple responses in arabidopsis. Plant Physiol. 179, 1343–1361. doi: 10.1104/pp.18.01092
Etesami H., Jeong B. R., Glick B. R. (2023). Potential use of Bacillus spp. as an effective biostimulant against abiotic stresses in crops—A review. Curr. Res. Biotechnol. 5, 100128. doi: 10.1016/j.crbiot.2023.100128
Fahim S., Dimitrov K., Vauchel P., Gancel F., Delaplace G., Jacques P., et al. (2013). Oxygen transfer in three phase inverse fluidized bed bioreactor during biosurfactant production by Bacillus subtilis. Biochem. Eng. J. 76, 70–76. doi: 10.1016/j.bej.2013.04.004
Farzand A., Moosa A., Zubair M., Khan A., Massawe V., Tahir H., et al. (2019a). Suppression of sclerotinia sclerotiorum by the induction of systemic resistance and regulation of antioxidant pathways in tomato using fengycin produced by Bacillus amyloliquefaciens FZB42. Biomolecules 9, 613. doi: 10.3390/biom9100613
Farzand A., Moosa A., Zubair M., Rashid Khan A., Hanif A., Tahir H. A. S., et al. (2019b). Marker assisted detection and LC-MS analysis of antimicrobial compounds in different Bacillus strains and their antifungal effect on Sclerotinia sclerotiorum. Biol. Control 133, 91–102. doi: 10.1016/j.biocontrol.2019.03.014
Fendrihan S., Constantinescu F., Sicuia O.-A., Dinu S. (2016). Beneficial Bacillus strains improve plant resistance to phytopathogens: A review. Int. J. Environ. Agric. Biotechnol. (IJEAB) 1, 2456–1878. Available at: www.ijeab.com ( Accessed March 28, 2025).
Ferreira Filho J. A., Horta M., dos Santos C., Almeida D., Murad N., Mendes J., et al. (2020). Integrative genomic analysis of the bioprospection of regulators and accessory enzymes associated with cellulose degradation in a filamentous fungus (Trichoderma harzianum). BMC Genomics 21 (1), 757. doi: 10.1186/s12864-020-07158-w
Fitz E., Wanka F., Seiboth B. (2018). The promoter toolbox for recombinant gene expression in Trichoderma reseii. Front. Bioeng. Biotechnol. 6. doi: 10.3389/fbioe.2018.00135
Ghazy N., El-Nahrawy S. (2021). Siderophore production by Bacillus subtilis MF497446 and Pseudomonas koreensis MG209738 and their efficacy in controlling Cephalosporium maydis in maize plant. Arch. Microbiol. 203, 1195–1209. doi: 10.1007/s00203-020-02113-5
Ghorbanpour M., Omidvari M., Abbaszadeh-Dahaji P., Omidvar R., Kariman K. (2018). Mechanisms underlying the protective effects of beneficial fungi against plant diseases. Biol. Control 117, 147–157. doi: 10.1016/j.biocontrol.2017.11.006
Gotor-Vila A., Usall J., Torres R., Abadias M., Teixidó N. (2017). Formulation of the biocontrol agent Bacillus amyloliquefaciens CPA-8 using different approaches: liquid, freeze-drying and fluid-bed spray-drying. BioControl 62, 545–555. doi: 10.1007/s10526-017-9802-3
Gray D. A., Dugar G., Gamba P., Strahl H., Jonker M. J., Hamoen L. W. (2019). Extreme slow growth as alternative strategy to survive deep starvation in bacteria. Nat. Commun. 10, 890. doi: 10.1038/s41467-019-08719-8
Hakim S., Naqqash T., Nawaz M., Laraib I., Siddique M., Zia R., et al. (2021). Rhizosphere engineering with plant growth-promoting microorganisms for agriculture and ecological sustainability. Front. Sustain. Food Syst. 5, 617157. doi: 10.3389/fsufs.2021.617157
Hariharan G., Rifnas L. M., Prasannath K. (2022). “Role of Trichoderma spp. in biocontrol of plant diseases,” in Microbial Biocontrol: Food Security and Post Harvest Management: Volume 2 (Switzerland: Springer International Publishing), 39–78. doi: 10.1007/978-3-030-87289-2_3
Herrera-Téllez V. I., Cruz-Olmedo A. K., Plasencia J., Gavilanes-Ruíz M., Arce-Cervantes O., Hernández-León S., et al. (2019). The protective effect of Trichoderma asperellum on tomato plants against Fusarium oxysporum and botrytis cinerea diseases involves inhibition of reactive oxygen species production. Int. J. Mol. Sci. 20, 2007. doi: 10.3390/ijms20082007
Hirpara D. G., Gajera H. P., Hirpara H. Z., Golakiya B. A. (2017). Antipathy of Trichoderma against Sclerotium rolfsii Sacc.: Evaluation of Cell Wall-Degrading Enzymatic Activities and Molecular Diversity Analysis of Antagonists. J. Mol. Microbiol. Biotechnol. 27, 22–28. doi: 10.1159/000452997
Hong S., Kim T. Y., Won S. J., Moon J. H., Ajuna H. B., Kim K. Y., et al. (2022). Control of fungal diseases and fruit yield improvement of strawberry using Bacillus velezensis CE 100. Microorganisms 10, 365. doi: 10.3390/microorganisms10020365
Huang J., Liu S., Zhang C., Wang X., Pu J., Ba F., et al. (2019). Programmable and printable Bacillus subtilis biofilms as engineered living materials. Nat. Chem. Biol. 15, 34–41. doi: 10.1038/s41589-018-0169-2
Inayati A., Sulistyowati L., Qurata L., Yusnawan E. (2020). Mycoparasitic Activity of Indigenous Trichoderma virens Strains Against Mungbean Soil Borne Pathogen Rhizoctonia solani: Hyperparasite and Hydrolytic Enzyme Production. AGRIVITA J. Agric. Sci. 42, 229–242. doi: 10.17503/agrivita.v0i0.2514
Izquierdo-García L. F., Carmona-Gutiérrez S., Moreno-Velandia C., Villareal-Navarrete A., Burbano-David D., Quiroga-Mateus R., et al. (2024). Microbial-Based Biofungicides Mitigate the Damage Caused by Fusarium oxysporum f. sp. cubense Race 1 and Improve the Physiological Performance in Banana. J. Fungi 10 (6), 419. doi: 10.3390/jof10060419
Jangir M., Sharma S., Sharma S. (2021). Development of next-generation formulation against Fusarium oxysporum and unraveling bioactive antifungal metabolites of biocontrol agents. Sci. Rep. 11, 22895. doi: 10.1038/s41598-021-02284-1
Jaroszuk-ściseł J., Tyskiewicz R., Nowak A., Ozimek E., Majewska M., Hanaka A., et al. (2019). Phytohormones (Auxin, gibberellin) and ACC deaminase in vitro synthesized by the mycoparasitic trichoderma DEMTKZ3A0 strain and changes in the level of auxin and plant resistance markers in wheat seedlings inoculated with this strain conidia. Int. J. Mol. Sci. 20 (19), 4923. doi: 10.3390/ijms20194923
Kariuki C. K., Mutitu E. W., Muiru W. M. (2020). Effect of Bacillus and Trichoderma species in the management of the bacterial wilt of tomato (Lycopersicum esculentum) in the field. Egyptian J. Biol. Pest Control 30, 109. doi: 10.1186/s41938-020-00310-4
Khan R. A. A., Najeeb S., Hussain S., Xie B., Li Y. (2020). Bioactive secondary metabolites from Trichoderma spp. Against phytopathogenic fungi. Microorganisms 8, 817. doi: 10.3390/microorganisms8060817
Kim J., Lee J., Lee C. H., Woo S. Y., Kang H., Seo S. G., et al. (2015). Activation of pathogenesis-related genes by the Rhizobacterium, Bacillus sp. JS, which induces systemic resistance in Tobacco plants. Plant Pathol. J. 31, 195–201. doi: 10.5423/PPJ.NT.11.2014.0122
Kulimushi P. Z., Arias A. A., Franzil L., Steels S., Ongena M. (2017). Stimulation of fengycin-type antifungal lipopeptides in Bacillus amyloliquefaciens in the presence of the maize fungal pathogen Rhizomucor variabilis. Front. Microbiol. 8. doi: 10.3389/fmicb.2017.00850
Kumar, Brar A., Yadav M., Chawade A., Vivekanand V., Pareek N. (2018). Chitinases—Potential candidates for enhanced plant resistance towards fungal pathogens. Agric. (Switzerland) 8, 88. doi: 10.3390/agriculture8070088
Kumar V., Koul B., Taak P., Yadav D., Song M. (2023). Journey of Trichoderma from pilot scale to mass production: A review. Agric. (Switzerland) 13, 2022. doi: 10.3390/agriculture13102022
Kumar Chaube S., Pandey S. (2022). TRICHODERMA: A valuable multipurpose fungus for sustainable agriculture. Malaysian J. Sustain. Agric. 6, 97–100. Available at: www.myjsustainagri.com ( Accessed April 22, 2025).
Landero Valenzuela N., Lara Viveros F. M., Rodríguez Ortega A., Pérez Vite A., Ortíz Hernández A. (2019). Trichoderma posible micoparásito de Sporisorium reilianum y su influencia en el rendimiento de maíz. Entreciencias: Diálogos en la Sociedad del Conocimiento 7, 13–23. doi: 10.22201/enesl.20078064e.2019.20.67345
Lastochkina O., Baymiev A., Shayahmetova A., Garshina D., Koryakov I., Shpirnaya I., et al. (2020). Effects of endophytic bacillus subtilis and salicylic acid on postharvest diseases (Phytophthora infestans, Fusarium oxysporum) development in stored potato tubers. Plants 9 (1), 76. doi: 10.3390/plants9010076
Leiva S., Oliva M., Hernández E., Chuquibala B., Rubio K., García F., et al. (2020). Assessment of the potential of Trichoderma spp. strains native to bagua (Amazonas, Peru) in the biocontrol of frosty pod rot (Moniliophthora roreri). Agronomy 10, 1376. doi: 10.3390/agronomy10091376
Li Y., Cui J., Kang J., Zhao W., Yang K., Fu J. (2024). Trichoderma rhizosphere soil improvement: regulation of nitrogen fertilizer in saline–alkali soil in semi-arid region and its effect on the microbial community structure of maize roots. Agronomy 14, 2340. doi: 10.3390/agronomy14102340
Li Y., Gu Y., Li J., Xu M., Wei Q., Wang Y. (2015). Biocontrol agent Bacillus amyloliquefaciens LJ02 induces systemic resistance against cucurbits powdery mildew. Front. Microbiol. 6. doi: 10.3389/fmicb.2015.00883
Liang Z., Ali Q., Wang Y., Mu G., Kan X., Ren Y., et al. (2022). Toxicity of Bacillus thuringiensis Strains Derived from the Novel Crystal Protein Cry31Aa with High Nematicidal Activity against Rice Parasitic Nematode Aphelenchoides besseyi. Int. J. Mol. Sci. 23 (15), 8189. doi: 10.3390/ijms23158189
Liu D., Li K., Hu J., Wang W., Liu X., Gao Z. (2019). Biocontrol and action mechanism of Bacillus amyloliquefaciens and Bacillus subtilis in soybean phytophthora blight. Int. J. Mol. Sci. 20, 2908. doi: 10.3390/ijms20122908
Locatelli G. O., dos Santos G. F., Botelho P. S., Finkler C. L. L., Bueno L. A. (2018). Development of Trichoderma sp. formulations in encapsulated granules (CG) and evaluation of conidia shelf-life. Biol. Control 117, 21–29. doi: 10.1016/j.biocontrol.2017.08.020
Løvschall K. B., Velasquez S. T. R., Kowalska B., Ptaszek M., Jarecka A., Szczech M., et al. (2024). Enhancing stability and efficacy of Trichoderma bio-control agents through layer-by-layer encapsulation for sustainable plant protection. Adva. Sustain. Syst. 8, 2300409. doi: 10.1002/adsu.202300409
Mahmoud E., Saleh A. A., Eranthodi A., Molan Y. Y. (2015). Characterization of novel Trichoderma asperellum isolates to select effective biocontrol agents against tomato fusarium wilt. Plant Pathol. J. 31, 50–60. doi: 10.5423/PPJ.OA.09.2014.0087pISSN1598-2254eISSN2093–9280
Maksimov I. V., Blagova D., Veselova S., Sorokan A., Burkhanova G., Cherepanova E., et al. (2020). Recombinant Bacillus subtilis 26DCryChS line with gene Btcry1Ia encoding Cry1Ia toxin from Bacillus thuringiensis promotes integrated wheat defense against pathogen Stagonospora nodorum Berk. and greenbug Schizaphis graminum Rond. Biol. Control 144, 104242. doi: 10.1016/j.biocontrol.2020.104242
Malviya D., Sahu P., Singh U., Paul S., Gupta A., Gupta A., et al. (2020). Lesson from ecotoxicity: Revisiting the microbial lipopeptides for the management of emerging diseases for crop protection. Int. J. Environ. Res. Public Health 17 (4), 1434. doi: 10.3390/ijerph17041434
Manganiello G., Sacco A., Ercolano M., Vinale F., Lanzuise S., Pascale A., et al. (2018). Modulation of tomato response to rhizoctonia solani by Trichoderma harzianum and its secondary metabolite harzianic acid. Front. Microbiol. 9, 1966. doi: 10.3389/fmicb.2018.01966
Martínez L. (2021). La bacteria Bacillusen el Manejo Integrado de Plagas: Una amiga para el agricultor moderno. TecnoVita.
Martínez, Heeb M., Kalač T., Gholam Z., Schwarze F. W. M. R., Nyström G., et al. (2023). Biopolymer-based emulsions for the stabilization of Trichoderma atrobrunneum conidia for biological control. Appl. Microbiol. Biotechnol. 107, 1465–1476. doi: 10.1007/s00253-023-12381-y
Martínez-Canto O. J., Cristóbal-Alejo J., Tun-Suárez J. M., Reyes-Ramírez A. (2021). Detección de genes Epl1 y Sm1 en Trichoderma spp. antagonistas contra hongos fitopatógenos. Ecosistemas y Recursos Agropecuarios 8, e2791. doi: 10.19136/era.a8n2.2791
Martínez-Salgado S., Andrade-Hoyos P., Romero-Arenas O., Villa-Ruano N., Landeta-Cortés G., Rivera-Tapia J. A. (2021). Control in vitro de Fusarium sp. asociado al cultivo de cebolla mediante Trichoderma harzianum. Rev. Mexicana Fitopatología Mexican J. Phytopathol. 39, 314–328. doi: 10.18781/r.mex.fit.2101-4
Masi M., Nocera P., Reveglia P., Cimmino A., Evidente A. (2018). Fungal metabolites antagonists towards plant pests and human pathogens: Structure-activity relationship studies. Molecules 23, 834. doi: 10.3390/molecules23040834
Méndez-Úbeda J. M., Flores Hernández M. S., Páramo-Aguilera L. A. (2018). Aislamiento e Identificación de Bacillus subtilis y Evaluación del antagonismo in vitro frente hongos fitopatógenos. Nexo Rev. Científica 30, 96–110. doi: 10.5377/nexo.v30i2.5530
Meyer C. T., Lynch G. K., Stamo D. F., Miller E. J., Chatterjee A., Kralj J. M. (2023). A high-throughput and low-waste viability assay for microbes. Nat. Microbiol. 8, 2304–2314. doi: 10.1038/s41564-023-01513-9
Miguel-Ferrer L., Romero-Arenas O., Andrade-Hoyos P., Sánchez-Morales P., Rivera-Tapia J. A., Fernández-Pavía S. P. (2021). Antifungal activity of Trichoderma harzianum and T. koningiopsis against Fusarium solani in seed germination and vigor of Miahuateco chili seedlings. Rev. Mexicana Fitopatología Mexican J. Phytopathol. 39, 228–247. doi: 10.18781/r.mex.fit.2101-5
Mitra D., Chaudhary P., Verma D., Khoshru B., Senapati A., Mahakur B., et al. (2021). “Bioinformatics’ role in studying microbe-mediated biotic and abiotic stress tolerance,” in Microbial Management of Plant Stresses: Current Trends, Application and Challenges (Hooghly, India: Elsevier), 203–219. doi: 10.1016/B978-0-323-85193-0.00005-X
Mnif I., Ghribi D. (2015). Lipopeptides biosurfactants: Mean classes and new insights for industrial, biomedical, and environmental applications. Biopolymers 104, 129–147. doi: 10.1002/bip.22630
Mohammadi P., Tozlu E., Kotan R., Şenol Kotan M. (2017). Potential of some bacteria for biological control of postharvest citrus green mould caused by Penicillium digitatum. Plant Prot. Sci. 53, 134–143. doi: 10.17221/55/2016-PPS
Morales-Mora L. A., Andrade-Hoyos P., Valencia-de Ita M. A., Romero-Arenas O., Silva-Rojas H. V., Contreras-Paredes C. A. (2020). Characterization of strawberry associated fungi and in vitro antagonistic effect of Trichoderma harzianum. Rev. Mexicana Fitopatología Mexican J. Phytopathol. 38, 434–449. doi: 10.18781/r.mex.fit.2005-7
Mukherjee P. K., Mendoza-Mendoza A., Zeilinger S., Horwitz B. A. (2022). Mycoparasitism as a mechanism of Trichoderma-mediated suppression of plant diseases. Fungal Biol. Rev. 39, 15–33. doi: 10.1016/j.fbr.2021.11.004
Muras A., Romero M., Mayer C., Otero A. (2021). Biotechnological applications of Bacillus licheniformis. Crit. Rev. Biotechnol. 41, 609–627. doi: 10.1080/07388551.2021.1873239
Naamala J., Smith D. L. (2020). Relevance of plant growth promoting microorganisms and their derived compounds, in the face of climate change. Agronomy 10, 1179. doi: 10.3390/agronomy10081179
Nikolić I., Berić T., Dimkić I., Popović T., Lozo J., Fira D., et al. (2019). Biological control of Pseudomonas syringae pv. aptata on sugar beet with Bacillus pumilus SS-10.7 and Bacillus amyloliquefaciens (SS-12.6 and SS-38.4) strains. J. Appl. Microbiol. 126 (1), 165–176. doi: 10.1111/jam.14070
Orozco-Mosqueda M., Santoyo G., Glick B. (2023). Recent advances in the bacterial phytohormone modulation of plant growth. Plants 12, 606. doi: 10.3390/plants12030606
Ortiz A., Sansinenea E., Keswani C., Minkina T., Singh S., Rekadwad B., et al. (2024). Bioengineering Bacillus spp. for sustainable crop production: recent advances and resources for biotechnological applications. J. Plant Growth Regul. doi: 10.1007/s00344-024-11553-y
Oszust K., Cybulska J., Frąc M. (2020). How do Trichoderma genus fungi win a nutritional competition battle against soft fruit pathogens? A report on niche overlap nutritional potentiates. Int. J. Mol. Sci. 21, 1–19. doi: 10.3390/ijms21124235
Palmieri D., Ianiri G., Del Grosso C., Barone G., De Curtis F., Castoria R., et al. (2022). Advances and perspectives in the use of biocontrol agents against fungal plant diseases. Horticulturae 8, 577. doi: 10.3390/horticulturae8070577
Pedraza L. A., López C. E., Uribe-Vélez D. (2020). Mechanisms of action of bacillus spp. (bacillaceae) against phytopathogenic microorganisms during their interaction with plants. Acta Biol. Colombiana 25, 112–125. doi: 10.15446/abc.v25n1.75045
Philip B., Behiry S. I., Salem M. Z. M., Amer M. A., El-Samra I. A., Abdelkhalek A., et al. (2024). Trichoderma afroharzianum TRI07 metabolites inhibit Alternaria alternata growth and induce tomato defense-related enzymes. Sci. Rep. 14, 1874. doi: 10.1038/s41598-024-52301-2
Pimentel M. F., Arnão E., Warner A., Subedi A., Rocha L., Srour A., et al. (2020). Trichoderma isolates inhibit Fusarium virguliforme growth, reduce root rot, and induce defense-related genes on soybean seedlings. Plant Dis. 104, 1949–1959. doi: 10.1094/PDIS-08-19-1676-RE
Poveda J. (2021). Trichoderma as biocontrol agent against pests: New uses for a mycoparasite. Biol. Control 159, 104634. doi: 10.1016/j.biocontrol.2021.104634
Prakash O., Nimonkar Y., Shouche Y. S. (2013). Practice and prospects of microbial preservation. FEMS Microbiol. Lett. 339, 1–9. doi: 10.1111/1574-6968.12034
Prasad B., Sharma D., Kumar P., Chandra Dubey R. (2023). Biocontrol potential of Bacillus spp. for resilient and sustainable agricultural systems. Physiol. Mol. Plant Pathol. 128, 102173. doi: 10.1016/j.pmpp.2023.102173
Prihatiningsih N., Asnani A., Djatmiko H. A. (2021). Extracellular protease from Bacillus subtilis b315 with antagonistic activity against bacterial wilt pathogen (Ralstonia solanacearum) of chili. Biodiversitas 22, 1291–1295. doi: 10.13057/biodiv/d220327
Rabbee M. F., Sarafat Ali M., Choi J., Hwang B. S., Jeong S. C., Baek K. H. (2019). Bacillus velezensis: A valuable member of bioactive molecules within plant microbiomes. Molecules 24. doi: 10.3390/molecules24061046
Rajer F. U., Samma M., Ali Q., Rajar W., Wu H., Raza W., et al. (2022). Bacillus spp.-Mediated Growth Promotion of Rice Seedlings and Suppression of Bacterial Blight Disease under Greenhouse Conditions. Pathogens 11 (11), 1251. doi: 10.3390/pathogens11111251
Risoli S., Cotrozzi L., Sarrocco S., Nuzzaci M., Pellegrini E., Vitti A. (2022). Trichoderma-induced resistance to Botrytis cinerea in Solanum species: A meta-analysis. Plants 11, 180. doi: 10.3390/plants11020180
Rosazza T., Eigentler L., Earl C., Davidson F. A., Stanley-Wall N. R. (2023). Bacillus subtilis extracellular protease production incurs a context-dependent cost. Mol. Microbiol. 120, 105–121. doi: 10.1111/mmi.15110
Saiyam D., Dubey A., Malla M. A., Kumar A. (2024). Lipopeptides from Bacillus: unveiling biotechnological prospects—sources, properties, and diverse applications. Braz. J. Microbiol. 55, 281–295. doi: 10.1007/s42770-023-01228-3
Santoyo G., Guzmán-Guzmán P., Parra-Cota F. I., de los Santos-Villalobos S., Orozco-Mosqueda M. D. C., Glick B. R. (2021). Plant growth stimulation by microbial consortia. Agronomy 11, 219. doi: 10.3390/agronomy11020219
Saxena A., Mishra S., Ray S., Raghuwanshi R., Singh H. B. (2020). Differential Reprogramming of Defense Network in Capsicum annum L. Plants Against Colletotrichum truncatum Infection by Phyllospheric and Rhizospheric Trichoderma Strains. J. Plant Growth Regul. 39, 751–763. doi: 10.1007/s00344-019-10017-y
Schuster M., Schweizer G., Reissmann S., Kahmann R. (2016). Genome editing in Ustilago maydis using the CRISPR-Cas system. Fungal Genet. Biol. 89, 3–9. doi: 10.1016/j.fgb.2015.09.001
Shafi J., Tian H., Ji M. (2017). Bacillus species as versatile weapons for plant pathogens: a review. Biotechnol. Biotechnol. Equip. 31, 446–459. doi: 10.1080/13102818.2017.1286950
Sharma P. K., Gothalwal R. (2017). “Trichoderma: A potent fungus as biological control agent,” in Agro-Environmental Sustainability (Hauppauge, NY: Springer International Publishing), 113–125. doi: 10.1007/978-3-319-49724-2_6
Shewarega Foyate M. (2024). Improvement in biocontrol ability of Trichoderma through chemical mutation and gamma irradiation for the control of soil borne pathogens. J. Clin. Res. Rep. 15, 01–09. doi: 10.31579/2690-1919/347
Shi J., Zhu X., Lu Y., Zhao H., Lu F., Lu Z. (2018). Improving iturin A production of Bacillus amyloliquefaciens by genome shuffling and its inhibition against Saccharomyces cerevisiae in orange juice. Front. Microbiol. 9. doi: 10.3389/fmicb.2018.02683
Siahmoshteh F., Siciliano I., Banani H., Hamidi-Esfahani Z., Razzaghi-Abyaneh M., Gullino M. L., et al. (2017). Efficacy of Bacillus subtilis and Bacillus amyloliquefaciens in the control of Aspergillus parasiticus growth and aflatoxins production on pistachio. Int. J. Food Microbiol. 254, 47–53. doi: 10.1016/j.ijfoodmicro.2017.05.011
Silva R. N., Monteiro V. N., Steindorff A. S., Gomes E. V., Noronha E. F., Ulhoa C. J. (2019). Trichoderma/pathogen/plant interaction in pre-harvest food security. Fungal Biol. 123, 565–583. doi: 10.1016/j.funbio.2019.06.010
Song Y., He S., Abdallah I., Jopkiewicz A., Setroikromo R., Van Meerkerk R., et al. (2021). Engineering of multiple modules to improve amorphadiene production in Bacillus subtilis using CRISPR-Cas9. J. Agric. Food Chem. 69, 4785–4794. doi: 10.1021/acs.jafc.1c00498
Sood M., Kapoor D., Kumar V., Sheteiwy M., Ramakrishnan M., Landi M., et al. (2020). Trichoderma: The “secrets” of a multitalented biocontrol agent. Plants 9, 1–25. doi: 10.3390/plants9060762
Souza E. (2016). The effects of sublethal doses of essential oils and their constituents on antimicrobial susceptibility and antibiotic resistance among food-related bacteria: A review. Trends Food Sci. Technol. 56, 1–12. doi: 10.1016/j.tifs.2016.07.012
Stummer B. E., Zhang Q., Zhang X., Warren R. A., Harvey P. R. (2020). Quantification of Trichoderma afroharzianum, Trichoderma harzianum and Trichoderma gamsii inoculants in soil, the wheat rhizosphere and in planta suppression of the crown rot pathogen Fusarium pseudograminearum. J. Appl. Microbiol. 129, 971–990. doi: 10.1111/jam.14670
Sun J., Liu Y., Lin F., Lu Z., Lu Y. (2021). CodY, ComA, DegU and Spo0A controlling lipopeptides biosynthesis in Bacillus amyloliquefaciens fmbJ. J. Appl. Microbiol. 131, 1289–1304. doi: 10.1111/jam.15007
Tamandegani P. R., Marik T., Zafari D., Balázs D., Vágvölgyi C., Szekeres A., et al. (2020). Changes in peptaibol production of Trichoderma species during in vitro antagonistic interactions with fungal plant pathogens. Biomolecules 10, 730. doi: 10.3390/biom10050730
Teixidó N., Usall J., Torres R. (2022). Insight into a successful development of biocontrol agents: production, formulation, packaging, and shelf life as key aspects. Horticulturae 8, 305. doi: 10.3390/horticulturae8040305
Thambugala K. M., Daranagama D. A., Phillips A. J. L., Kannangara S. D., Promputtha I. (2020). Fungi vs. Fungi in biocontrol: an overview of fungal antagonists applied against fungal plant pathogens. Front. Cell. Infect. Microbiol. 10. doi: 10.3389/fcimb.2020.604923
Thilagam P., Sharanappa C. H., Roy S., Deb L., Padhan S., Srividhya S., et al. (2023). A review on advances in biocontrol techniques for managing insect pests in sustainable agriculture. Int. J. Environ. Climate Change 13, 2114–2125. doi: 10.9734/ijecc/2023/v13i113373
Thomine E., Mumford J., Rusch A., Desneux N. (2022). Using crop diversity to lower pesticide use: Socio-ecological approaches. Sci. Total Environ. 804, 150156. doi: 10.1016/j.scitotenv.2021.150156
Torres R., Solsona C., Viñas I., Usall J., Plaza P., Teixidó N. (2014). Optimization of packaging and storage conditions of a freeze-dried Pantoea agglomerans formulation for controlling postharvest diseases in fruit. J. Appl. Microbiol. 117, 173–184. doi: 10.1111/jam.12511
Tyśkiewicz R., Nowak A., Ozimek E., Jaroszuk-ściseł J. (2022). Trichoderma: the current status of its application in agriculture for the biocontrol of fungal phytopathogens and stimulation of plant growth. Int. J. Mol. Sci. 23, 2329. doi: 10.3390/ijms23042329
Tyagi A., Lama Tamang T., Kashtoh H., Mir R., Mir Z., Manzoor S., et al. (2024). A review on biocontrol agents as sustainable approach for crop disease management: applications, production, and future perspectives. Horticulturae 10 (8), 805. doi: 10.3390/horticulturae10080805
Ubando A. T., Felix C. B., Chen W. H. (2020). Biorefineries in circular bioeconomy: A comprehensive review. Bioresour. Technol. 299, 122585. doi: 10.1016/j.biortech.2019.122585
Umer M., Mubeen M., Iftikhar Y., Ali Shad M., Usman H., Sohail M., et al. (2021). Role of rhizobacteria on plants growth and biological control of plant diseases: a review. Plant Prot. 5, 59–73. doi: 10.33804/pp.005.01.3565
Valencia M., Fátima J. H., Lezama C. P., Simón A. B., Cortés G. L., Romero-Arenas O. (2021). Bio-controller effect of four native strains of Trichoderma spp., on Phytophthora capsici in Manzano Chili (Capsicum pubescens) in Puebla-Mexico. J. Pure Appl. Microbiol. 15, 998–1005. doi: 10.22207/JPAM.15.2.58
Valencia De Ita M. A., Lezama C. P., Simon A. B., Morales Mora L. A., Lara M. H., Garcia B. L., et al. (2020). In vitro antagonism of strains of Trichoderma spp., on pathogenic fungi of nopal vegetable. J. Pure Appl. Microbiol. 14, 1345–1352. doi: 10.22207/JPAM.14.2.31
Valenzuela Ruiz V., Gándara-Ledezma A., Villareal-Delgado M., Villa-Rodriguez E., Parra-Cota F.I., Santoyo G., et al. (2020). Regulation, biosynthesis, and extraction of Bacillus-derived lipopeptides and its implications in biological control of phytopathogens. Stresses 2024 (4), 107–132. doi: 10.3390/stresses4010007
Vargas-Hoyos H., Gilchrist-Ramelli E. (2015). Producción de enzimas hidrolíticas y actividad antagónica de Trichoderma asperellum sobre dos cepas de Fusarium aisladas de cultivos de tomate (Solanum lycopersicum). Rev. Mexicana Micol. 42, 9–16.
Vasquez-Lopez A., Gomez-Jaimes R., Villarreal-Barajas T. (2021). Effectiveness of neutral electrolyzed water and copper oxychloride on fungi spores isolated from tropical fruits. Heliyon 7, e07935. doi: 10.1016/j.heliyon.2021.e07935
Vehapi M., İnan B., Kayacan-Cakmakoglu S., Sagdic O., Özçimen D. (2023). Production of Bacillus subtilis soil isolate as biocontrol agent under bioreactor conditions. Arch. Microbiol. 205, 52. doi: 10.1007/s00203-022-03381-z
Vekemans M.-C., Marchand P. A. (2020). The fate of biocontrol agents under the European phytopharmaceutical regulation: how this regulation hinders the approval of botanicals as new active substances. Environ. Sci. Pollut. Res. 27, 39879–39887. doi: 10.1007/s11356-020-10114-6
Villao-Uzho L., Espinoza-Lozano F., Galarza-Romero L., Santos-Ordóñez E. (2024). Biotechnological tools for genetic improvement of Trichoderma. Scientia Agropecuaria 15, 213–223. doi: 10.17268/sci.agropecu.2024.016
Wang, Chen H., Ma L., Gong M., Wu Y., Bao D., et al. (2022). Use of CRISPR-Cas tools to engineer Trichodermaspecies. Microb. Biotechnol. 15, 2521–2532. doi: 10.1111/1751-7915.14126
Wang H., Shi Y., Wang D., Yao Z., Wang Y., Liu J., et al. (2018). A biocontrol strain of Bacillus subtilis WXCDD105 used to control tomato Botrytis cinerea and Cladosporium fulvum cooke and promote the growth of seedlings. Int. J. Mol. Sci. 19 (5), 1371. doi: 10.3390/ijms19051371
Wang R., Long Z., Liang X., Guo S., Ning N., Yang L., et al. (2021). The role of a β-1,3-1,4-glucanase derived from Bacillus amyloliquefaciens FS6 in the protection of ginseng against Botrytis cinerea and Alternaria panax. Biol. Control 164, 104765. doi: 10.1016/j.biocontrol.2021.104765
Wang Q., Chen L., Fang C., Wang H., Shi Y., Zhao Y., et al. (2019). The overexpression of one single cbh gene making Trichoderma asperellum T-1 a better cellulase producer. Ann. Microbiol. 69 (7), 673–683. doi: 10.1007/s13213-019-01458-7
Wang Z., et al. (2024). Classification, application, multifarious activities and production improvement of lipopeptides produced by Bacillus. Crit. Rev. Food Sci. Nutr. 64, 7451–7464. doi: 10.1080/10408398.2023.2185588
Wang M., Geng L., Sun X., Shu C., Song F., Zhang J. (2020). Screening of Bacillus thuringiensis strains to identify new potential biocontrol agents against Sclerotinia sclerotiorum and Plutella xylostella in Brassica campestris L. Biol. Control 145, 104262. doi: 10.1016/j.biocontrol.2020.104262
WHO (2022). Pesticide residues in food. Available online at: https://www.who.int/news-room/fact-sheets/detail/pesticide-residues-in-food (Accessed February 11, 2025).
Wiwattanapatapee R., Chumthong A., Pengnoo A., Kanjanamaneesathian M. (2013). Preparation and evaluation of Bacillus megaterium-alginate microcapsules for control of rice sheath blight disease. World J. Microbiol. Biotechnol. 29, 1487–1497. doi: 10.1007/s11274-013-1314-4
Won S. J., Choub V., Kwon J. H., Kim D. H., Ahn Y. S. (2018). The control of fusarium root rot and development of coastal pine (Pinus thunbergii Parl.) Seedlings in a container nursery by use of bacillus licheniformis MH48. Forests 10, 6. doi: 10.3390/f10010006
Xiao Z., Zhao Q., Li W., Gao L., Liu G. (2023). Strain improvement of Trichoderma harzianum for enhanced biocontrol capacity: Strategies and prospects. Front. Microbiol. 14. doi: 10.3389/fmicb.2023.1146210
Yadav M., Dubey M. K., Upadhyay R. S. (2021). Systemic resistance in chilli pepper against anthracnose (Caused by Colletotrichum truncatum) induced by Trichoderma harzianum, Trichoderma asperellum and Paenibacillus dendritiformis. J. Fungi 7, 307. doi: 10.3390/jof7040307
Yu C., Liu X., Zhang X., Zhang M., Gu Y., Ali Q., et al. (2021a). Mycosubtilin produced by Bacillus subtilis ATCC6633 inhibits growth and mycotoxin biosynthesis of Fusarium graminearum and Fusarium verticillioides. Toxins 13 (11), 791. doi: 10.3390/toxins13110791
Yu C., Chen H., Zhu L., Song Y., Jiang Q., Zhang Y., et al. (2023). Profiling of antimicrobial metabolites synthesized by the endophytic and genetically amenable biocontrol strain Bacillus velezensis DMW1. Microbiol. Spectr. 11 (2), e0003823. doi: 10.1128/spectrum.00038-23
Yu, Wang Z., Zhang Y., Wang Y., Liu Z. (2021b). Biocontrol and growth-promoting effect of Trichoderma asperellum TaspHu1 isolate from Juglans mandshurica rhizosphere soil. Microbiol. Res. 242, 126596. doi: 10.1016/j.micres.2020.126596
Zafra G., Moreno-Montaño A., Absalón Á. E., Cortés-Espinosa D. V. (2015). Degradation of polycyclic aromatic hydrocarbons in soil by a tolerant strain of Trichoderma asperellum. Environ. Sci. Pollut. Res. 22, 1034–1042. doi: 10.1007/s11356-014-3357-y
Zandi P., Basu S. (2016). Role of plant growth-promoting rhizobacteria (PGPR) as BioFertilizers in stabilizing agricultural ecosystems 71–87. doi: 10.1007/978-3-319-26803-3_3
Zhang S., Zhang S., Zhang C., Wang X., Shi Y. (2019). Cucumber leaf disease identification with global pooling dilated convolutional neural network. Comput. Electron. Agric. 162, 422–430. doi: 10.1016/j.compag.2019.03.012
Zhao, Lyu N., Wang D., Wu X., Zhao Y., Zhang L., et al. (2020). PhlG mediates the conversion of DAPG to MAPG in Pseudomonas fluorescens 2P24. Sci. Rep. 10, 4296. doi: 10.1038/s41598-020-60555-9
Zhou C., Li P., Wu S., Zhao P., Gao H. (2019). Bacillus subtilis CF-3 volatile organic compounds inhibit Monilinia fructicola growth in peach fruit. Int. J. Biol. Macromol. 10, 1804. doi: 10.3389/fmicb.2019.01804
Zhou C., Zhu J., Qian N., Guo J., Yan C. (2021). Bacillus subtilis SL18r Induces Tomato Resistance Against Botrytis cinerea, Involving Activation of Long Non-coding RNA, MSTRG18363, to Decoy miR1918. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.634819
Zin N. A., Badaluddin N. A. (2020). Biological functions of Trichoderma spp. for agriculture applications. Ann. Agric. Sci. 65, 168–178. doi: 10.1016/j.aoas.2020.09.003
Keywords: beneficial microorganisms, biological control agents, biopesticides, crop protection, sustainability
Citation: Villavicencio-Vásquez M, Espinoza-Lozano F, Espinoza-Lozano L and Coronel-León J (2025) Biological control agents: mechanisms of action, selection, formulation and challenges in agriculture. Front. Agron. 7:1578915. doi: 10.3389/fagro.2025.1578915
Received: 18 February 2025; Accepted: 08 April 2025;
Published: 07 May 2025.
Edited by:
Hakim Manghwar, Lushan Botanical Garden (CAS), ChinaReviewed by:
Pankaj Singh, Rammanohar Lohia Avadh University, IndiaKahsay Mawcha, Aksum University, Ethiopia
Copyright © 2025 Villavicencio-Vásquez, Espinoza-Lozano, Espinoza-Lozano and Coronel-León. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jonathan Coronel-León, anJjb3JvbmVAZXNwb2wuZWR1LmVj