 Cristina D. Balcón Pacheco1†Carlos A. García Munguía2†
Cristina D. Balcón Pacheco1†Carlos A. García Munguía2† César Ozuna3Román Cardona Herrera1Alberto M. García Munguía4
César Ozuna3Román Cardona Herrera1Alberto M. García Munguía4 Elena Franco Robles2*
Elena Franco Robles2*- 1Division of Life Sciences, University of Guanajuato, Irapuato, Mexico
- 2Department of Veterinary and Animal Science, Division of Life Sciences, University of Guanajuato, Irapuato, Mexico
- 3Department of Food Science, Division of Life Sciences, University of Guanajuato, Irapuato, Mexico
- 4Department of Agronomic Sciences, Center for Agricultural Sciences, Autonomous University of Aguascalientes, Aguascalientes, Mexico
The increasing restrictions on antibiotic use in poultry production have prompted the search for natural alternatives that improve health and productivity in laying hens. This study aimed to evaluate the effects of dietary supplementation with agave fructans (PRE), turmeric powder (AOX), and their combination (PA) on productive performance, egg quality, hematological status, and gastrointestinal traits in early-laying hens. A total of 120 Lohman White hens (20 weeks old) were randomly assigned to five dietary treatments: basal control (CON), antibiotics (ATB), agave fructans (PRE), turmeric powder (AOX), and the combination of both additives (PA). Results showed that agave fructans improved internal egg quality, increasing albumen and yolk height and egg weight without altering feed intake or conversion. Both PRE and AOX enhanced immune status, evidenced by a reduced heterophil-to-lymphocyte ratio and modulation of leukocyte populations. Gastrointestinal traits were minimally affected, with only slight variations in crop and cecal pH and duodenal organ weight. In conclusion, PRE supplementation provided the most consistent physiological benefits, supporting its use as a sustainable strategy to improve egg quality and welfare in laying hens while reducing reliance on antibiotics.
1 Introduction
Over the last three decades, the demand for eggs has increased by 150%, surpassing the demand for meat and milk (FAO, 2023). Due to this, the production of laying hens has intensified, which has implications for bird welfare, including an increased risk of disease transmission. The use of antibiotics—either as growth promoters or for disease prevention and treatment—in large-scale production has led to the emergence of antimicrobial resistance. The selective pressure and horizontal transfer of antibiotic resistance genes among multiple bacterial species that cause human disease are of fundamental importance to the One Health concept and global health security, particularly in low- and middle-income countries (Kim and Cha, 2021; Cella et al., 2023). Furthermore, antibiotics used as feed additives have been detected as residues exceeding maximum residual limits in animal products, including eggs (Owusu-Doubreh et al., 2023). In this context, studies have shown that consumers are more concerned about chemical risks in food, such as antibiotic residues, than about microbiological risks (Meagher, 2019).
As an alternative to the use of antibiotics in poultry production, various compounds, mainly of natural origin, have been evaluated. The popularity of the use of feed additives in the feeding of production animals has increased in recent decades, especially in the production of animal-derived foods intended for consumers who prefer organic, cruelty-free, and safe foods. Accordingly, nutritional supplements such as prebiotics, probiotics, herbal extracts, and others have been investigated for their potential to enhance growth performance and health in livestock and poultry (Lao et al., 2020; Bąkowski and Kiczorowska, 2021; Oluwafemi et al., 2020; Al-Baadani et al., 2021) and are able to replace the use of antibiotics as a feed additive (Balcón-Pacheco et al., 2023; Franco-Robles and Hernández-Granados, 2024).
The inclusion of prebiotics such as fructans in the diet of laying hens has been shown to improve feed conversion, modify intestinal pH, increase calcium absorption and retention, and improve egg production and quality by reducing cholesterol content (H. M. Shang et al., 2010; Świątkiewicz et al., 2010; Gaggìa et al., 2010). Fructans derived from Agave tequilana Weber var. Blue, known as agavins, are prebiotics characterized by a highly branched fructose structure with β(2-1) and β(2-6) glycosidic bonds and terminal glucose units (Lopez et al., 2003).
Turmeric powder from Curcuma longa, which contains high concentrations of polyphenolic compounds, has also been used in poultry feed, often in conjunction with other plant extracts, such as oregano and thyme, among others, and even using supplementation with vitamin E (Rajput et al., 2013). Studies have shown that turmeric and its derivatives improve laying percentage, feed conversion ratio, and egg quality parameters, including shell thickness, yolk and albumen index, Haugh units, and yolk pigmentation, while enhancing antioxidant content and reducing total cholesterol (Rajput et al., 2013; Kermanshahi and Riasi, 2006; Radwan Nadia et al., 2008; Riasi et al., 2012; Khan et al., 2012; Arshami et al., 2013).
Despite extensive research on prebiotics and phytogenic additives, few studies have examined the synergistic potential of agave fructans and turmeric powder as a combined natural alternative to antibiotics in laying hens. This study addresses this gap by evaluating their individual and combined effects on productive performance, physiological parameters, and gastrointestinal characteristics in early-laying hens.
2 Materials and methods
2.1 Ethics statement
The experiment was conducted at the Department of Veterinary and Animal Science, University of Guanajuato, Campus Irapuato–Salamanca, Division of Life Sciences, Mexico. All procedures carried out within the experimental protocol were approved by the Institutional Bioethics Committee of the University of Guanajuato (approval code: CIBIUG-P42-2019; approved on July 5, 2019).
2.2 Animals, management, and experimental design
A total of 120 healthy 18-week-old Lohman White hens were obtained from a commercial hatchery (Jalisco, Mexico) and housed individually in pens (25 cm x 35 cm x 40 cm; floor slope, 12°) under controlled environmental conditions (25-25°C) with 16 h of continuous light and 8 h of darkness per day. Feed and purified water were provided ad libitum. The hens underwent a 2-week adaptation period before the experimental phase.
At 20 weeks of age, hens were randomly assigned to five dietary treatments, with 24 replicates per diet, as follows: CON (basal diet without additives), ATB (antibiotics, basal diet + 0.02% colistin sulfate + 0.05% zinc bacitracin), PRE (basal diet + 0.5% agave fructans), AOX (basal diet + 0.5% turmeric powder), and PA (basal diet + 0.5% agave fructans + 0.5% turmeric powder). The hens were fed the experimental basal diet for 12 weeks (21–32 weeks of age). The composition of the basal diet, formulated according to the nutritional requirements outlined in the Lohmann Laying Hens Management Guide, is presented in Table 1. Colistin sulfate combined with zinc bacitracin was included in the ATB group to represent a conventional antibiotic program historically used in commercial poultry diets in Mexico, enabling comparison with functional feed additives while maintaining alignment with antimicrobial stewardship principles.
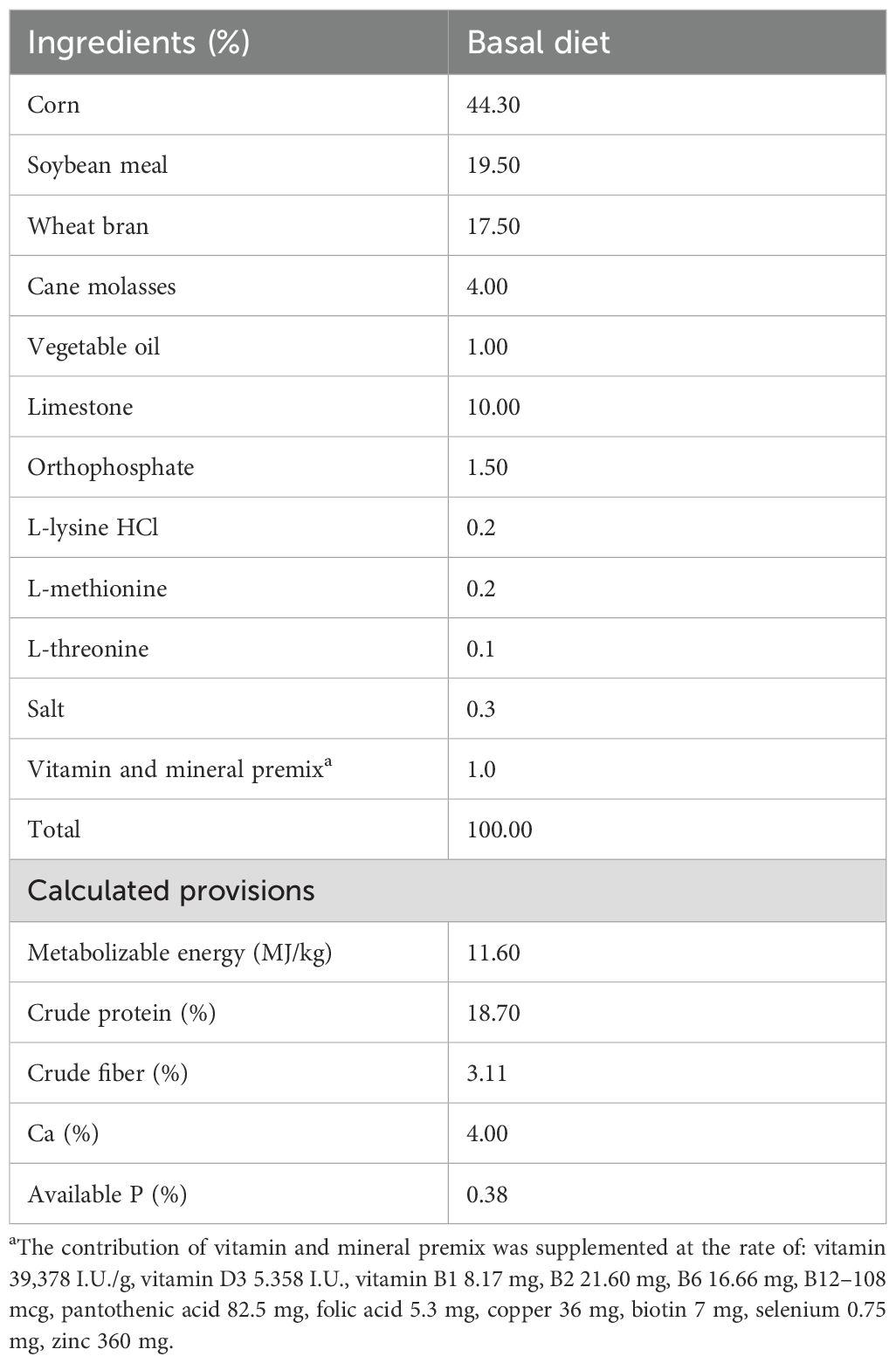
Table 1. Ingredients and calculated nutrient analysis of standard diet.
2.3 Composition of feed functional ingredients
The agave fructans used were a crystalline, water-soluble powder obtained from Agave tequilana Weber var. Azul, containing 90.7% inulin, 5.1% fructose, 2.0% glucose, 1.8% sucrose, and 0.4% other carbohydrates, as determined by high-performance liquid chromatography (HPLC) with refractive index detection (Agilent 1260, Grupo Solalve, Jalisco, Mexico).
The turmeric powder was a commercial yellow–orange powder (>98% curcuminoids) purchased from a commercial supplier (Expo Alimentos, Jalisco, Mexico). To characterize its antioxidant properties, the total phenolic compounds, total flavonoids, carotenoids, chlorophyll content, and in vitro antioxidant capacity were evaluated as described below:
2.3.1 Total phenolic compounds
Extracts used for the determination of total phenolic compounds (TPC), total flavonoids (TF), and antioxidant capacity in vitro were prepared according to (Cardona-Herrera et al., 2025). The TPC was measured as described by (Cardona-Herrera et al., 2025). The reaction was incubated for 1 h in the dark, and absorbance was recorded at 765 nm using a UV–Vis spectrophotometer (Genesys 10S, Thermo Scientific, Waltham, USA). Results were expressed as milligrams of gallic acid equivalents per gram of dry weight (mg GAE/g DW).
2.3.2 Total flavonoid content
The total flavonoid content (TFC) was determined using the aluminum chloride method according to (Castañeda-Salazar et al., 2023). The reaction mixture was read at 415 nm using a UV–Vis spectrophotometer (Genesys 10S, Thermo Scientific, USA). Results were expressed as milligrams of quercetin equivalents per gram of dry weight (mg QE/g DW).
2.3.3 Carotenoids and chlorophyll content
Extracts used for determining carotenoid (CAR) and chlorophyll (CHLO) content were obtained following the procedure of (Samakradhamrongthai et al., 2021). A 2 mL aliquot of the extract was used to determine the CAR and CHLO content. The absorbance was measured with a UV–Vis spectrophotometer (Genesys 10S, Thermo Scientific, Waltham, USA). CARR, and CARY (470 and 508 nm) and CHLO (646 and 663 nm), concentrations were determined according to Equations 1, 2, and 3 proposed by (Lichtenthaler, 1987; Hornero-Méndez and Mínguez-Mosquera, 2001). Results were expressed as micrograms per gram of dry weight (μg/g DW).
2.3.4 Antioxidant capacity in vitro
The Trolox equivalent antioxidant capacity (TEAC) was determined following the method described by (Cardona-Herrera et al., 2025). First, 250 µL of extract was mixed with 2250 µL of 60 µM 2,2-diphenyl-1-picrylhydrazyl (DPPH), and absorbance was measured at 517 nm using a UV–Vis spectrophotometer (Genesys 10S, Thermo Scientific, Waltham, USA). Results were expressed as micromoles of Trolox equivalents per gram of dry weight (µmol TE/g DW).
2.4 Egg sample collection and measurements
Starting at 20 weeks of age, egg production (EP) was recorded daily until the hens reached 32 weeks of age. Body weight was measured weekly. Average daily feed intake (ADFI) was calculated by subtracting the leftover feed weight from the amount provided the previous day.
The egg mass (EM) and feed conversion ratio (FCR) were calculated according to Equations 4 and 5, while the relative organ weight (ROW) was determined as shown in Equation 6.
The feed conversion ratio (FCR) was calculated using (5):
2.5 Egg quality parameters
At three points during the experiment—T0 (week 21), T1 (week 27), and T2 (week 32)—a total of 90 eggs (six eggs per treatment at each time point) were randomly selected to determine egg quality traits. Egg length (EL, cm), egg width (EWD, cm), eggshell thickness (EST, mm), albumen height (AH, mm), thick albumen (TA, mm), thin albumen (tA, mm), yolk height (YH, mm), and yolk width (YW, mm) were measured using a Vernier digital caliper (HER-411, Steren, Mexico). Eggshell weight (ESW, g) was measured using an analytical balance (VE-204, Velab, Mexico). The egg quality index (EQI) was calculated using equations introduced by (Narushin et al., 2021). The color was assessed using the Yolk Color Fan (DSM, USA), while yolk color profile parameters—lightness (L*), redness (a*), and yellowness (b*) values were determined using a Hunterlab Colour Flex (Hunterlab, USA).
2.6 Sampling and hematological and biochemical parameters
At the end of the experiment, 10 birds were selected from each dietary treatment, and blood samples (3 mL) were collected from the wing vein into heparinized tubes (BD Vacutainer®) after a 12-h feed withdrawal. Plasma was obtained by centrifugation at 3000 g for 15 min at 4°C and stored at -20°C for further analysis.
Hematocrit was measured by the micro method using capillary tubes, centrifuged at 2500 g for 10 min. The counting of total leukocytes (TLC) was performed (Balcón-Pacheco et al., 2023). To calculate total erythrocyte count (TEC), the cells of the five quadrants were added and divided by 100, reporting the number of erythrocytes x 106/µL. A peripheral blood film stained with Hemocrom-Fix (Golden Bell, Mexico) was performed for the differential count of heterophils, lymphocytes, monocytes, eosinophils, and basophils.Plasma glucose, calcium, and phosphorus levels were analyzed using commercial diagnostic kits (Spinreact®, Mexico).
2.7 Relative organ weight
The birds used for blood sampling were euthanized by cervical dislocation. The crop, proventriculus, gizzard, duodenum, jejunum, ileum, large intestine, and ceca were dissected and collected. The contents of each organ were used for pH determination using a potentiometer (Hanna Hi98103, Hanna, USA).
Relative organ weight (ROW) was calculated using the following equation:
The lengths of the small intestine, large intestine, and ceca were measured with a Vernier caliper and expressed in millimeters (mm).
2.8 Statistical analysis
Each replicate was treated as an experimental unit. Data normality was assessed using the Kolmogorov–Smirnov test. A one-way analysis of variance (ANOVA) was performed using Statistica (version 8.0; StatSoft, USA). When significant effects were observed, Bonferroni’s post hoc test was used to adjust for multiple comparisons and to control the overall Type I error rate. A p-value of < 0.05 was considered statistically significant.
3 Results
Table 2 shows the total bioactive compounds and in vitro antioxidant capacity of the turmeric powder (TP) included in the laying hen diet. The total phenolic content (TPC) and total flavonoid content (TF) were 4.76 ± 0.15 mg GAE/g DW and 6.40 ± 0.16 mg QE/g DW, respectively, representing the main contributors to the antioxidant potential of the sample. Yellow carotenoids (CARY) were detected at 3.41 ± 0.23 mg/g DW, whereas red carotenoids (CARR) were not detected. Chlorophyll (CHLO) content reached 30.96 ± 0.91 µg/g DW. Antioxidant capacity assays revealed values of 10.55 ± 0.67 µmol TE/g DW for TEAC and 58.74 ± 0.62 µmol TE/g DW for FRAP, indicating a strong reducing capacity of the turmeric matrix. Overall, these results confirm that the TP used in this study contained considerable amounts of phenolic and flavonoid compounds associated with high antioxidant activity.
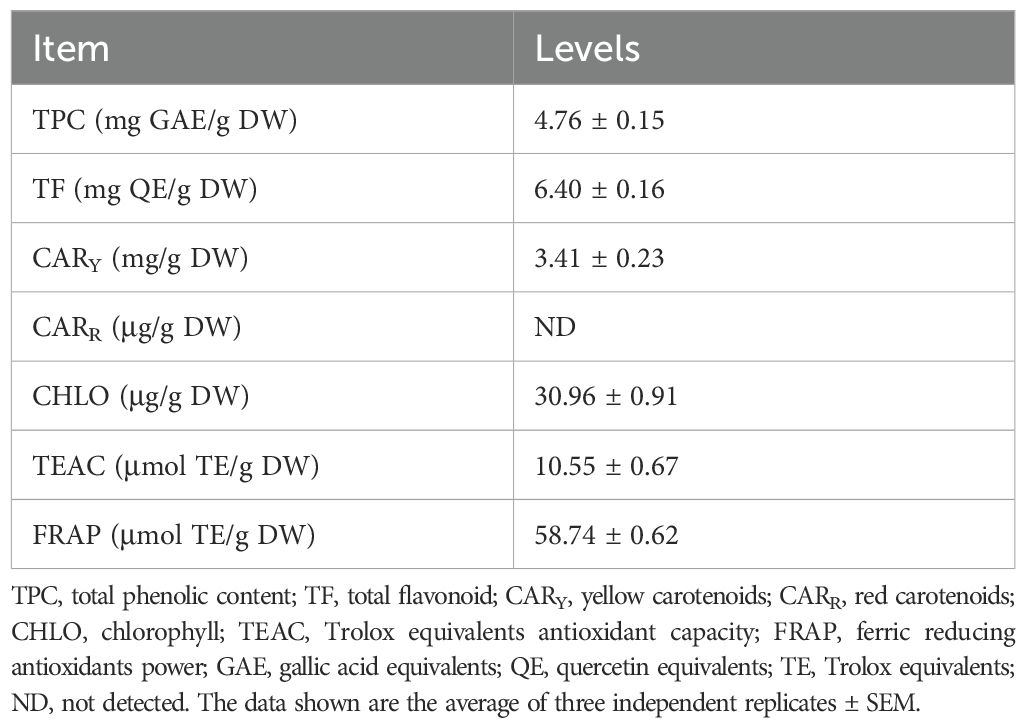
Table 2. Total bioactive compounds and in vitro antioxidants of turmeric powder.
Table 3 summarizes the productive performance of laying hens fed diets containing different additives. No significant differences (p > 0.05) were observed in body weight (BW), average daily feed intake (ADFI), feed conversion ratio (FCR), or egg production (EP) among treatments at any sampling time (T0, T1, or T2). In contrast, egg weight (EW) exhibited significant differences at T1 and T2 according to Bonferroni’s post hoc test (p < 0.05). At T1, hens supplemented with PRE showed higher EW values than those fed ATB diets, while at T2 the PRE group also maintained the highest EW relative to CON and other treatments. Egg mass (EM) increased numerically in the supplemented groups, although differences were not statistically significant (p > 0.05). These results suggest that dietary inclusion of natural additives, particularly PRE supplementation, improved egg weight without affecting feed consumption or conversion efficiency.
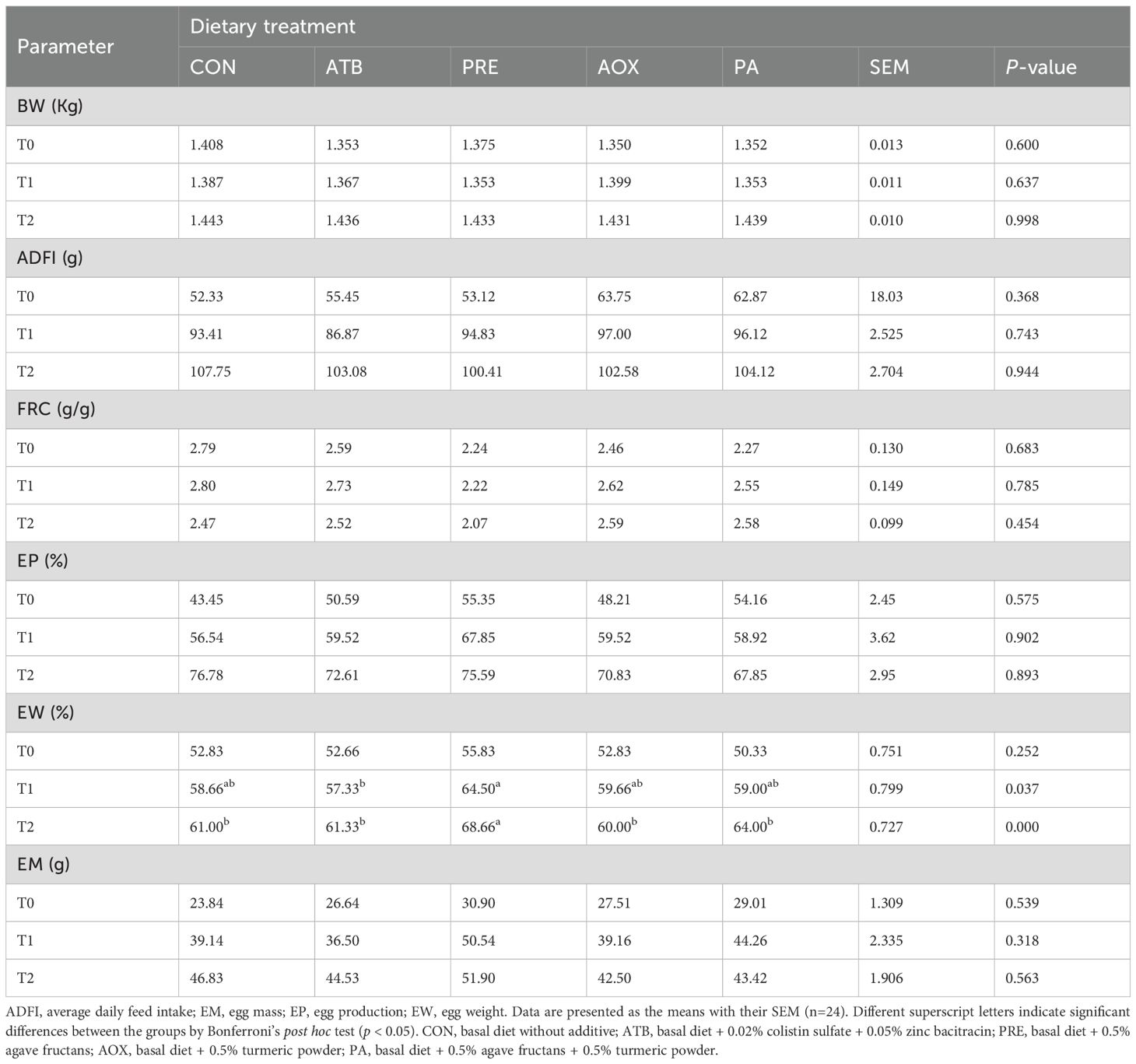
Table 3. Effects of dietary additives on performance of the laying hens over 12 weeks of feeding.
Figure 1 provides a visual comparison of representative eggs obtained from hens under the different dietary treatments.

Figure 1. Representative eggs collected from hens after 12 weeks of dietary supplemented with a basal control diet (CON), antibiotics (ATB), agave fructans (PRE), turmeric powder (AOX), or the combination of both functional additives (PA).
Table 4 presents the effects of the additives on external and internal egg quality traits at T0, T1, and T2. Most external parameters—egg length, egg width, egg shape index, and eggshell thickness—did not differ among treatments (p > 0.05). However, EW differed at T1 (p = 0.037) and T2 (p < 0.001), with PRE hens showing higher values than the remaining treatments.
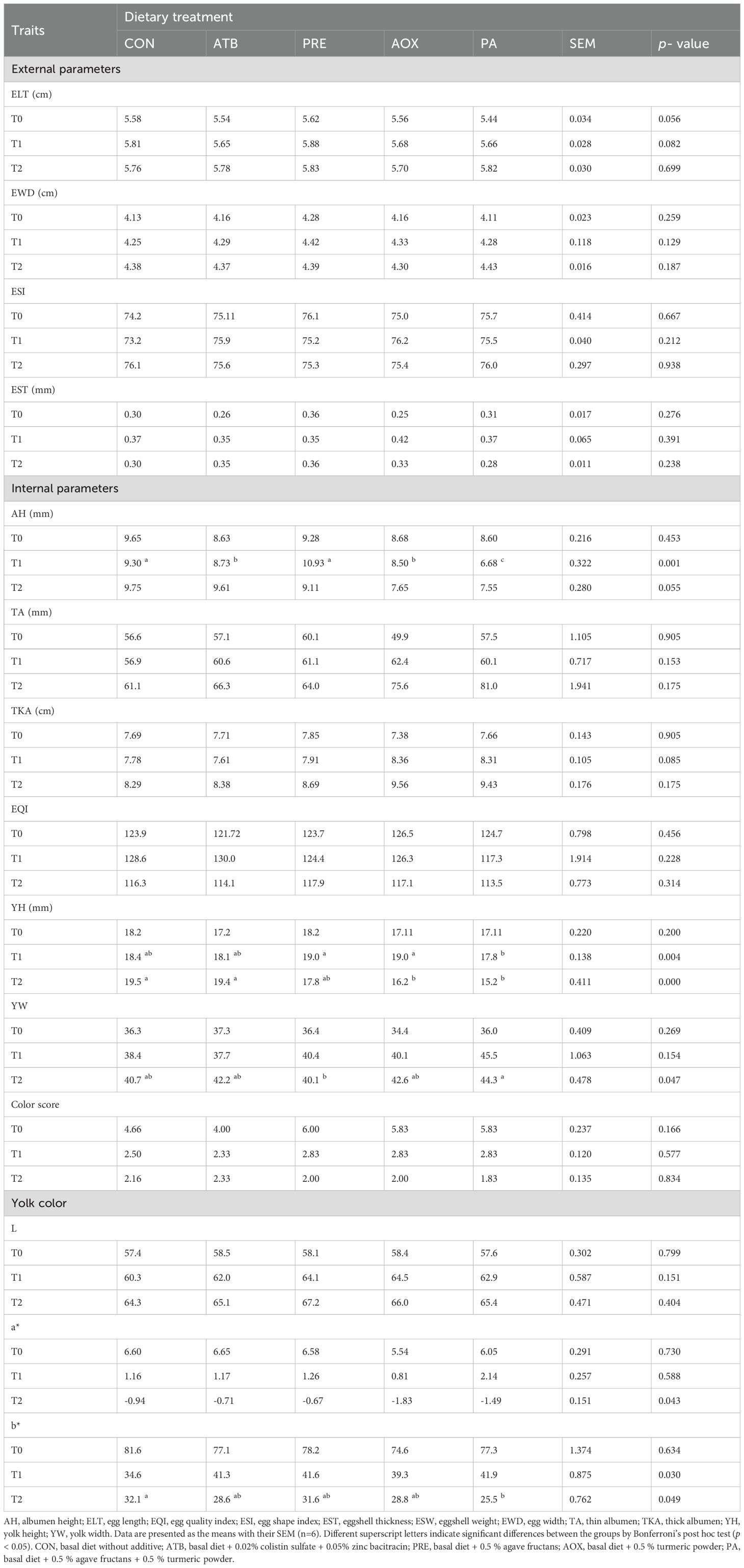
Table 4. Effect of dietary additives on egg quality trails of the laying hens over 12 weeks of feeding.
Regarding internal quality parameters, albumen height (AH) was significantly affected at T1 (p = 0.001): PRE exhibited the highest AH, followed by CON and ATB, while PA showed the lowest. Yolk height (YH) also differed at T1 (p = 0.004) and T2 (p < 0.001). At both times, PRE and AOX showed higher YH than PA, indicating improved yolk structure. In addition, yolk width (YW) at T3 showed a statistically significant difference among treatments (p < 0.05). Thin albumen (tA) at T2 showed a significant reduction in PRE and AOX compared with CON (p = 0.049). No statistical differences were found in the egg quality index (EQI), color score (CS), or L*, a*, b* color coordinates at any sampling point (p > 0.05).
Table 5 shows the hematological and biochemical parameters of laying hens at the end of the feeding trial (T2). Erythrocyte counts were significantly higher in PRE compared with ATB and PA, while CON and AOX showed intermediate values (p < 0.05). No significant differences were observed in hematocrit (p > 0.05). Total leukocytes were significantly lower in AOX and PA compared with CON and PRE (p < 0.05). The leukocyte profile was modified by dietary treatments: heterophils were significantly lower and lymphocytes significantly higher in PRE compared with CON and ATB (p < 0.05). Consequently, the H:L ratio was significantly lower in PRE than in CON and ATB (p < 0.0001). Monocyte percentages were higher in ATB than in AOX and PA (p < 0.05). No significant differences were observed in eosinophils or basophils (p > 0.05).
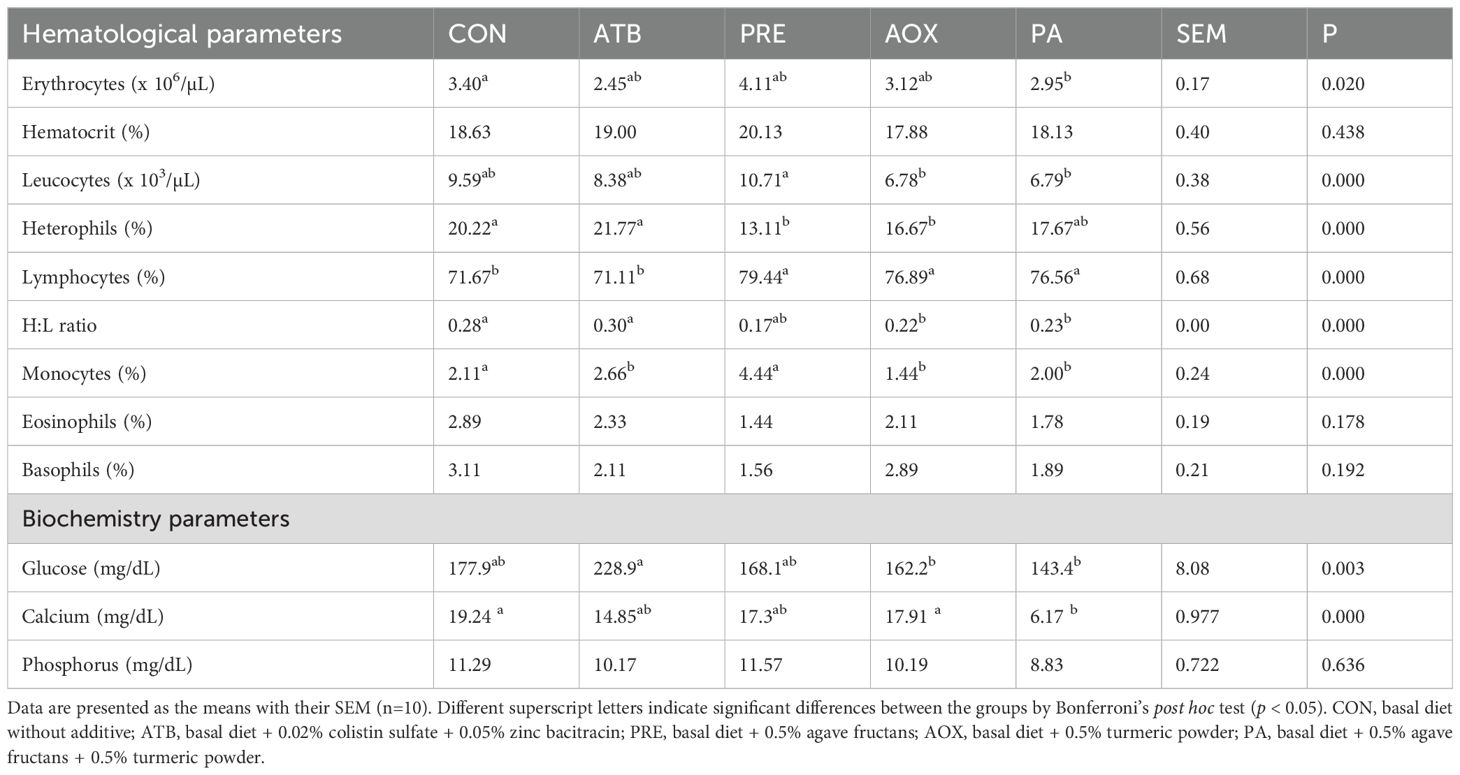
Table 5. Effect of dietary additives on hematological parameters of the laying hens at the end of 12 weeks of feeding.
Regarding serum metabolites, glucose levels were lower in PA compared with CON and ATB (p < 0.05). Calcium levels were also lower in PA compared with CON, ATB, and PRE (p < 0.05). No significant differences were detected in phosphorus levels (p > 0.05).
Supplementary Table S1 summarizes pH values, relative organ weights (ROW), and gastrointestinal tract lengths of hens at T2. In the crop, pH was significantly lower in ATB and PA than in CON (p < 0.05), while ROW did not differ among treatments. In the gizzard, hens fed PRE had lower pH than those in ATB and AOX (p < 0.05), although gizzard ROW did not differ. Duodenal ROW increased in AOX compared with ATB and PA (p < 0.05), although pH remained unchanged. No significant differences were observed in pH or ROW in the proventriculus, jejunum, ileum, or large intestine. In the cecum, PRE hens exhibited higher pH values than ATB (p < 0.05), while ATB showed reduced cecal ROW compared with CON, PRE, and AOX (p < 0.05). The lengths of the small intestine, large intestine, and ceca were not affected by dietary treatments.
4 Discussion
In this study, we evaluated the combination of the antibiotics zinc bacitracin and colistin sulfate, commonly used in commercial poultry feed, particularly for laying hens and broilers. It is important to clarify that this antibiotic treatment was included solely as a conventional production benchmark, not as a recommended practice, in alignment with antimicrobial stewardship principles. The inappropriate and widespread use of antibiotics poses risks to consumer health, as residues may remain in animal-derived foods (Feng et al., 2016). These two antibiotics have been detected in milk and animal tissues in cattle (Wan et al., 2006), posing a risk of antimicrobial resistance in enterococcal infections in humans (Singer and Johnson, 2024). In this regard, recent research efforts to improve production parameters and egg quality in laying hens have focused on replacing antibiotics in animal feed. Diets enriched with functional ingredients improve production parameters and egg quality in laying hens (Vlaicu et al., 2021). These ingredients include antioxidants, organic acids, plant extracts, and oligosaccharides, among others (Goliomytis et al., 2019; Dilawar et al., 2021; Zhang et al., 2024, 2021). Moreover, there is evidence of the use of turmeric powder has been evaluated as a dietary additive for laying hens, both alone and in combination with prebiotics or other functional ingredients (Bozorgkhoo and Kuhi, 2025). In the present study, we proposed two natural dietary additives—agave fructans and turmeric powder—with well-documented functional properties demonstrated in in vitro assays and animal models as potential alternatives to antibiotics. The selected doses were based on recent work by our research group (Franco-Robles and Hernández-Granados, 2024; Hernández-Granados et al., 2022; Guzmán-Rodríguez et al., 2024) to evaluate whether these additives, individually or in combination, could improve egg quality. The characterization of total bioactive compounds and in vitro antioxidant capacity in turmeric was consistent with values previously reported by (Yang et al., 2020; Pal et al., 2020). Regarding photosynthetic pigments, no CARR were detected; thus, total CAR consisted primarily of CARY pigments, mainly β-carotene, β-cryptoxanthin, and zeaxanthin (Hornero-Méndez and Mínguez-Mosquera, 2001). Similarly (Vasanthkumar et al., 2024), reported CAR values ranging from 2.78 to 17.32 mg/g and CHLO levels between 10 and 100 µg/g in Curcuma longa.
Productive performance was not affected by the treatments, which aligns with previous studies where inulin-type fructans and turmeric supplementation do not compromise feed intake or hen productivity (Shang et al., 2010; Mosayyeb Zadeh et al., 2023). However, other reports have shown improvements in weight gain and feed consumption when agave fructans are supplemented for longer periods or at different physiological stages. For instance, Sánchez-Chiprés et al. (2021) observed enhanced weight gain from weeks 5 to 18 and increased feed consumption at weeks 15 and 20 in 36-week-old hens fed 0.1% of 0.2% agave fructans (Sánchez-Chiprés et al., 2021). Moreover (Shang et al., 2020), observed a 3.09% increase in laying rate and a 3.61% decrease in feed conversion ratio with inulin supplementation during feeding weeks 1 to 8. The contrast between these results may be due to differences in the degree of polymerization or botanical source of the fructans used (Mueller et al., 2016), as our formulation used a polydisperse mixture, in addition to the fact that the age of the hens was different (Gu et al., 2021). Moreover, this variability may depend on dosage, laying phase, and duration of supplementation.
In the present study, supplementation with 0.5% agave fructans improved egg weight and internal quality traits such as albumen and yolk height throughout the experimental period. These findings are consistent with previous reports indicating that inulin-type fructans can enhance nutrient utilization and egg mass. Shang et al., 2020 observed a 2.54% increase in egg weight with dietary inclusion of 15 g/kg inulin. Moreover, Obianwuna et al. (2022) demonstrated improvements in egg parameters after 84 days of supplementation with 0.3%–0.6% FOS, and Alsherify and Hassanabadi (2024) reported that 3.0 g/kg FOS improved the shape index and Haugh unit without affecting overall performance. Collectively, these results suggest that fructan-based prebiotics help preserve internal egg composition and structural integrity.
In contrast, the effects of dietary turmeric powder were less consistent, as the 0.5% inclusion did not produce quality improvements comparable to PRE. However, the relatively small number of eggs analyzed per treatment may have limited statistical power for detecting subtle effects in some internal quality traits. Future studies with larger sample sizes per replicate are therefore recommended. Similar dose-dependent responses have been reported previously, where turmeric levels below 2% reduced feed efficiency and negatively affected egg production and quality traits (Dalal and Kosti, 2018). Some studies report improvements at 0.5-1% inclusion or turmeric powder while other studies found that the inclusion of 10 or 30 g/kg does not influence egg weight (Kosti et al., 2020). This variability indicates that higher doses or improved bioavailability of curcuminoids may be necessary to maximize the functional benefits of turmeric powder on egg quality.
To analyze the health status of the hens during supplementation with functional ingredients, we determined the hematological parameters. Bioactive dietary components are known to exert immunomodulatory effects that enhance resistance to pathogenic challenges in poultry (Obianwuna et al., 2022; Youssef et al., 2023). Our data showed that 0.5% agave fructans exhibited and increased total leucocyte count, accompanied by a reduction in heterophils and a higher proportion of lymphocytes resulting in a lower H:L ratio, an accepted indicator of animal stress (Franco-Robles and Hernández-Granados, 2024). These results suggest that agave fructans improve immune readiness and stress resilience in early-laying hens. Importantly, unlike mannanoligosaccharides (MOS), which have shown no changes in hematological profiles at dietary levels of 0.1-0.5 g/kg (Youssef et al., 2023), agave fructans elicited a clear immunomodulatory response. The effects of turmeric powder were more modest but still measurable, as reflected by a reduced H:L ratio compared with the control group. This finding is consistent with the known anti-inflammatory and antioxidant benefits of turmeric in laying hens (Kosti et al., 2020). Together, these results indicate that both agave fructans and turmeric enhance immune status, with the former displaying a more pronounced effect under the experimental conditions.
In this study, serum glucose levels were significantly higher in hens fed the antibiotic diet compared with all other treatments. Although further metabolic indicators were not assessed, this response may reflect metabolic stress associated with bacitracin exposure, as previously reported (Dowling, 2024). Regarding serum calcium, overall concentrations remained stable across diets except for the combination treatment (PA), which showed a significant decrease compared with the other groups. Prebiotics have been reported to improve calcium absorption and mineral homeostasis (Khan et al., 2020); therefore, the unexpected reduction observed in the PA group may indicate an interaction between fructans and turmeric that interferes with mineral uptake, warranting further investigation.
Finally, only limited gastrointestinal responses were observed following supplementation with 0.5% agave fructans and 0.5% turmeric powder. Diets containing PRE altered pH levels in specific intestinal segments, while AOX increased duodenal relative weight, indicating localized physiological responses in the upper digestive tract. These findings are consistent with previous reports suggesting that functional ingredients such as turmeric can influence digestive physiology and intestinal morphology in laying hens (Kosti et al., 2020). However, because changes were not consistent across all intestinal regions, the biological relevance of these effects remains uncertain. It is possible that the observed modifications are linked to microbial activity or fermentation processes in the gut, although this mechanism could not be confirmed in the present study. Thus, interpretations involving microbiota-driven effects should remain cautious until supported by microbial evidence in future investigations.
5 Conclusion
In summary, dietary supplementation with 0.5% agave fructans provided consistent physiological benefits in early-laying hens, improving egg weight and internal egg quality while favorably modulating hematological indicators of immune status, without compromising productive performance. Turmeric powder also contributed to a healthier immune profile, although its effects on egg quality were less evident at the evaluated dose. The combination of both additives did not outperform agave fructans alone.
Overall, these findings support agave fructans as a promising functional ingredient for improving egg quality and bird welfare, offering a viable alternative to conventional antibiotic growth promoters. Further research—including gut microbiota characterization and optimization of turmeric dosage and bioavailability—will be essential to fully elucidate the mechanisms involved and to refine combination strategies aligned with sustainable poultry production and One Health objectives.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by Comité de Ética para la Investigación de la Universidad de Guanajuato. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
CBP: Conceptualization, Investigation, Methodology, Writing – review & editing. CGM: Conceptualization, Formal analysis, Investigation, Methodology, Supervision, Writing – review & editing. RCH: Conceptualization, Investigation, Methodology, Writing – review & editing. CO: Writing – review & editing. AGM: Formal analysis, Supervision, Writing – review & editing. EFR: Conceptualization, Investigation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research, and/or publication of this article.
Acknowledgments
Cristina D. Balcón-Pacheco thanks SECIHTI for scholarship no. 907241 for their postgraduate studies and the Interinstitutional Master’s Degree in Livestock Production. Project carried out by Elena Franco-Robles, member of the Mexican Thematic network Sustainable integral use and biotechnology of agaves (AGARED) SECIHTI, Mexico.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fanim.2025.1688496/full#supplementary-material
References
Świątkiewicz S., Koreleski J., and Arczewska-Włosek A. (2010). Effect of prebiotic fructans and organic acids on mineral retention in laying hens. Acta Agriculturae Scand. Section A 60, 125–128. doi: 10.1080/09064702.2010.482593
Al-Baadani H. H., Al-Mufarrej S. I., Al-Garadi M. A., Alhidary I. A., Al-Sagan A. A., and Azzam M. M. (2021). The use of gum arabic as a natural prebiotic in animals: A review. Anim. Feed Sci. Technol. 274, 114894. doi: 10.1016/j.anifeedsci.2021.114894
Alsherify S. M. and Hassanabadi A. (2024). Effects of adding different levels of fructooligosaccharide to diet on productive performance, egg quality traits, immune response and blood metabolites in commercial laying hens. Veterinary Med. Sci. 10, e15505. doi: 10.1002/vms3.1550
Arshami J., Pilevar M., Azghadi M. A., and Raji A. R. (2013). Hypolipidemic and antioxidative effects of curcumin on blood parameters, humoral immunity, and jejunum histology in hy-line hens. Avicenna J. Phytomedicine 3, 1785.
Bąkowski M. and Kiczorowska B. (2021). Probiotic microorganisms and herbs in ruminant nutrition as natural modulators of health and production efficiency–a review. Ann. Anim. Sci. 21, 3–285. doi: 10.2478/aoas-2020-0081
Balcón-Pacheco C. D., Coronel-Gómez F. D., Ozuna C., Jiménez-Fernández M., Angel-Sahagún C. A., and Franco-Robles E. (2023). Effect of the probiotic bifidobacterium animalis as a substitute to growth-promoting antibiotics on performance and egg quality in laying hens. Iranian J. Appl. Anim. Sci. 13, 357–365. Available online at: https://dorl.net/dor/20.1001.1.2251628.2023.13.2.16.4.
Bozorgkhoo S. and Kuhi H. D. (2025). Effect of dietary supplementation with garlic powder, turmeric powder, saccharomyces cerevisiae yeast extract and commercial probiotics on growth performance, intestinal morphology and microflora and immune response of broiler chickens. Iranian J. Anim. Sci. 56, 71–945. doi: 10.22059/ijas.2024.371066.653991
Cardona-Herrera R., Pérez-Carrillo E., Quinones-Munoz T. A., and Ozuna C. (2025). Nixtamalization and its impact on the physicochemical, bioactive, and antioxidant properties of cucurbita pepo, cucurbita argyrosperma, and cucurbita maxima. LWT 223, 117760. doi: 10.1016/j.lwt.2025.117760
Castañeda-Salazar A., de Figueroa-Cárdenas J. D., López M. G., and Mendoza S. (2023). Physicochemical and functional characterization of agave fructans modified by cationization and carboxymethylation. Carbohydr. Polymer Technol. Appl. 5, 100284. doi: 10.1016/j.carpta.2023.100284
Cella E., Giovanetti M., Benedetti F., Scarpa F., Johnston C., Borsetti A., et al. (2023). Joining forces against antibiotic resistance: the one health solution. Pathogens 12, 1074. doi: 10.3390/pathogens12091074
Dalal R. and Kosti D. (2018). Turmeric powder as feed additive in laying hen—A review. J. Pharmacognosy Phytochem. 7, 2686–2895.
Dilawar M. A., Mun H. S., Rathnayake D., Yang E. J., Seo Y. S., Park H. S., et al. (2021). Egg quality parameters, production performance and immunity of laying hens supplemented with plant extracts. Animals 11, 975. doi: 10.3390/ani11040975
Dowling P. M. (2024). Peptide antibiotics: polymyxins, glycopeptides, bacitracin, and fosfomycin. Antimicrobial Ther. Veterinary Med., 187–201. doi: 10.1002/9781119654629
FAO (2023). “The State of Food and Agriculture 2023,” in Revealing the True Cost of Food to Transform Agrifood Systems (Rome: Food and agriculture Organization of the United Nations (FAO)).
Feng M. X., Wang G. N., Yang K., Liu H. Z., and Wang J. P. (2016). Molecularly imprinted polymer-high performance liquid chromatography for the determination of tetracycline drugs in animal derived foods. Food Control 69, 171–176. doi: 10.1016/j.foodcont.2016.04.050
Franco-Robles E. and Hernández-Granados M. J. (2024). Effects of dietary supplementation of cobiotic based on agave fructans on growth performance, blood parameters, oxidative damage and immune status of broiler. Trop. Anim. Health Production 56, 2155. doi: 10.1007/s11250-024-04059-y
Gaggìa F., Mattarelli P., and Biavati B. (2010). Probiotics and prebiotics in animal feeding for safe food production. Int. J. Food Microbiol. 141, S15–S28. doi: 10.1016/j.ijfoodmicro.2010.02.031
Goliomytis M., Simitzis P., Papalexi A., Veneti N., Hager-Theodorides A. L., Charismiadou M. A., et al. (2019). Influence of citrus flavonoids on laying hen performance, inflammatory immune response, egg quality and yolk oxidative stability. Br. Poultry Sci. 60, 272–278. doi: 10.1080/00071668.2019.1587150
Gu Y. F., Chen Y. P., Jin R., Wang C., Wen C., and Zhou Y. M. (2021). A comparison of intestinal integrity, digestive function, and egg quality in laying hens with different ages. Poultry Sci. 100, 100949. doi: 10.1016/j.psj.2020.12.046
Guzmán-Rodríguez J. J., Gutiérrez-Chávez A. J., Meléndez-Soto R. M., Amador-Sánchez M. A., and Franco-Robles E. (2024). Antimicrobial activities of agave fructans against multi-resistant and biofilm-producing staphylococcus aureus isolated from bovine mastitis. Veterinary Res. Commun. 48, 61–675. doi: 10.1007/s11259-023-10180-4
Hernández-Granados M.J., Ortiz-Basurto R. I., Jiménez-Fernández M., García-Munguía C. A., and Franco-Robles E. (2022). Dietary encapsulated bifidobacterium animalis and agave fructans improve growth performance, health parameters, and immune response in broiler chickens. Anim. Bioscience 35, 5875. doi: 10.5713/ab.21.0213
Hornero-Méndez D. and Mínguez-Mosquera M.I. (2001). Rapid spectrophotometric determination of red and yellow isochromic carotenoid fractions in paprika and red pepper oleoresins. J. Agric. Food Chem. 49, 3584–3885. doi: 10.1021/jf010400l
Kermanshahi H. and Riasi A. (2006). Effect of turmeric rhizome powder (Curcuma longa) and soluble NSP degrading enzyme on some blood parameters of laying hens. Int. J. Poultry Sci. 5, 494–498.
Khan S., Moore R. J., Stanley D., and Chousalkar K. K. (2020). The gut microbiota of laying hens and its manipulation with prebiotics and probiotics to enhance gut health and food safety. Appl. Environ. Microbiol. 86, e00600–e00205. doi: 10.1128/AEM.00600-20
Khan R. U., Naz S., Javdani M., Nikousefat Z., Selvaggi M., Tufarelli V., et al. (2012). The use of turmeric (Curcuma longa) in poultry feed. World’s Poultry Sci. J. 68, 97–103. doi: 10.1017/S0043933912000104
Kim D.-W. and Cha C.-J. (2021). Antibiotic resistome from the one-health perspective: understanding and controlling antimicrobial resistance transmission. Exp. Mol. Med. 53, 301–395. doi: 10.1038/s12276-021-00569-z
Kosti D., Dahiya D. S., Dalal R., Tewatia B. S., and Vijayalakshmy K. (2020). Role of turmeric supplementation on production, physical and biochemical parameters in laying hens. World’s Poultry Sci. J. 76, 625–375. doi: 10.1080/00439339.2020.1764460
Lao E. J., Dimoso N., Raymond J., and Mbega E. R. (2020). The prebiotic potential of brewers’ Spent grain on livestock’s health: A review. Trop. Anim. Health Production 52, 461–472. doi: 10.1007/s11250-019-02120-9
Lichtenthaler H. K. (1987). “[34] Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes,” in Methods in Enzymology, vol. 148. (San Diego, CA, USA: Elsevier).
Lopez M. G., Mancilla-Margalli N. A., and Mendoza-Díaz G. (2003). Molecular structures of fructans from agave tequilana weber var. Azul. J. Agric. Food Chem. 51, 7835–7405. doi: 10.1021/jf030383v
Meagher K. D. (2019). Public perceptions of food-related risks: A cross-national investigation of individual and contextual influences. J. Risk Res. 22, 919–935. doi: 10.1080/13669877.2017.1422789
Mosayyeb Zadeh A., Mirghelenj S. A., Hasanlou P., and Alishah H. S. (2023). Effects of turmeric (Curcuma longa) powder supplementation in laying hens’ Diet on production performance, blood biochemical parameters and egg quality traits. J. Anim. Physiol. Anim. Nutr. 107, 691–7025. doi: 10.1111/jpn.13746
Mueller M., Reiner J., Fleischhacker L., Viernstein H., Loeppert R., and Praznik W. (2016). Growth of selected probiotic strains with fructans from different sources relating to degree of polymerization and structure. J. Funct. Foods 24, 264–275. doi: 10.1016/j.jff.2016.04.010
Narushin V. G., Romanov M. N., and Griffin D. K. (2021). A novel egg quality index as an alternative to haugh unit score. J. Food Eng. 289, 110176. doi: 10.1016/j.jfoodeng.2020.110176
Obianwuna U. E., Chang X.-Y., Wang J., Zhang H. J., Qi G. H., Qiu K., et al. (2022). Dietary fructooligosaccharides effectively facilitate the production of high-quality eggs via improving the physiological status of laying hens. Foods 11, 1828. doi: 10.3390/foods11131828
Oluwafemi R. A., Olawale I., and Alagbe J. O. (2020). Recent trends in the utilization of medicinal plants as growth promoters in poultry nutrition-A review. Res.: Agric. Veterinary Sci. 4, 5–115.
Owusu-Doubreh B., Appaw W. O., and Abe-Inge V. (2023). Antibiotic residues in poultry eggs and its implications on public health: A review. Sci. Afr. 19, e01456. doi: 10.1016/j.sciaf.2022.e01456
Pal K., Chowdhury S., Dutta S. K., Chakraborty G. K., Pandit G. K., Dutta S., et al. (2020). Analysis of rhizome colour content, bioactive compound profiling and ex-situ conservation of turmeric genotypes (Curcuma longa L.) from sub-himalayan terai region of India. Ind. Crops Products 150, 112401. doi: 10.1016/j.indcrop.2020.112401
Radwan Nadia L., Hassan R. A., Qota E. M., and Fayek H. M. (2008). Effect of natural antioxidant on oxidative stability of eggs and productive and reproductive performance of laying hens. Int. J. Poultry Sci. 7, 134–150. doi: 10.3923/ijps.2008.134.150
Rajput N., Muhammad N., Yan R., Zhong X., and Wang T. (2013). Effect of dietary supplementation of curcumin on growth performance, intestinal morphology and nutrients utilization of broiler chicks. J. Poultry Sci. 50, 44–525. doi: 10.2141/jpsa.0120065
Riasi A., Kermanshahi H., and Mahdavi A. H. (2012). Production performance, egg quality and some serum metabolites of older commercial laying hens fed different levels of turmeric rhizome (Curcuma longa) powder. J. Medicinal Plants Res. 6, 2141–2455. doi: 10.5897/JMPR11.1316
Samakradhamrongthai R. S., Jannu T., and Renaldi G. (2021). Physicochemical properties and sensory evaluation of high energy cereal bar and its consumer acceptability. Heliyon 7, e07776. doi: 10.1016/j.heliyon.2021.e07776
Sánchez-Chiprés D. R., Chávez-Mora I. Y., Reynoso-Orozco R., and Noa-Pérez M. (2021). Levels of polyamines in feces of laying hens fed with agave fructans (Agave tequilana, weber) in association with the quality and production of egg. Braz. J. Poultry Sci. 23, eRBCA–2020-1253. doi: 10.1590/1806-9061-2020-1253
Shang H. M., Hu T. M., Lu Y. J., and Wu H. X. (2010). Effects of inulin on performance, egg quality, gut microflora and serum and yolk cholesterol in laying hens. Br. Poultry Sci. 51, 791–796. doi: 10.1080/00071668.2010.531005
Shang H., Zhao J., Dong X., Guo Y., Zhang H., Cheng J., et al. (2020). Inulin improves the egg production performance and affects the cecum microbiota of laying hens. Int. J. Biol. Macromolecules 155, 1599–1609. doi: 10.1016/j.ijbiomac.2019.11.137
Singer R. S. and Johnson T. J. (2024). Assessing the risk of antimicrobial resistant enterococcal infections in humans due to bacitracin usage in poultry. J. Food Prot. 87, 1002675. doi: 10.1016/j.jfp.2024.100267
Vasanthkumar R., Baskar V., Vinoth S., Roshna K., Mary T. N., Alagupandi R., et al. (2024). Biogenic carbon quantum dots from marine endophytic fungi (Aspergillus flavus) to enhance the curcumin production and growth in curcuma longa L. Plant Physiol. Biochem. 211, 108644. doi: 10.1016/j.plaphy.2024.108644
Vlaicu P. A., Panaite T. D., and Turcu R. P. (2021). Enriching laying hens eggs by feeding diets with different fatty acid composition and antioxidants. Sci. Rep. 11, 207075. doi: 10.1038/s41598-021-00343-1
Wan E. C.-h., Ho C., Sin D. W.-m., and Wong Y.-c. (2006). Detection of residual bacitracin A, colistin A, and colistin B in milk and animal tissues by liquid chromatography tandem mass spectrometry. Analytical Bioanalytical Chem. 385, 181–885. doi: 10.1007/s00216-006-0325-5
Yang Q.-Q., Cheng L.-Z., Zhang T., Yaron S., Jiang H. X., Sui Z. Q., et al. (2020). Phenolic profiles, antioxidant, and antiproliferative activities of turmeric (Curcuma longa). Ind. Crops Products 152, 112561. doi: 10.1016/j.indcrop.2020.112561
Youssef I. M., Khalil H. A., Jaber F. A., Alhazzaa R. A., Alkholy S. O., Almehmadi A. M., et al. (2023). Influence of dietary mannan-oligosaccharides supplementation on hematological characteristics, blood biochemical parameters, immune response and histological state of laying hens. Poultry Sci. 102, 103071. doi: 10.1016/j.psj.2023.103071
Zhang L., Ge J., Gao F., Yang M., Li H., Xia F., et al. (2024). Rosemary leaf powder improves egg quality, antioxidant status, gut barrier function, and cecal microbiota and metabolites of late-phase laying hens. Anim. Nutr. 17, 325–334. doi: 10.1016/j.aninu.2024.02.003
Keywords: bioactive-enriched feed, agave fructans, turmeric, keyword, egg quality, poultry nutrition, antibiotic alternatives, sustainable livestock production
Citation: Balcón Pacheco CD, García Munguía CA, Ozuna C, Cardona Herrera R, García Munguía AM and Franco Robles E (2025) Bioactive-enriched feed containing agave fructans and turmeric powder as an alternative to antibiotics in early-laying hens. Front. Anim. Sci. 6:1688496. doi: 10.3389/fanim.2025.1688496
Received: 19 August 2025; Accepted: 30 October 2025;
Published: 27 November 2025.
Edited by:
Arabela Untea, National Research Development Institute for Animal Biology and Nutrition, RomaniaReviewed by:
Igor Popov, Don State Technical University, RussiaMD. Shariful Islam, University of Rajshahi, Bangladesh
Copyright © 2025 Balcón Pacheco, García Munguía, Ozuna, Cardona Herrera, García Munguía and Franco Robles. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Elena Franco Robles, ZS5mcmFuY29AdWd0by5teA==
†These authors have contributed equally to this work and share first authorship