 Clemens Gögele
Clemens Gögele Girish Pattappa
Girish Pattappa Herbert Tempfer
Herbert Tempfer Denitsa Docheva
Denitsa Docheva Gundula Schulze-Tanzil
Gundula Schulze-Tanzil- 1Institute of Anatomy and Cell Biology, Paracelsus Medical University, Nuremberg, Germany
- 2Department of Musculoskeletal Tissue Regeneration, Orthopaedic Hospital König-Ludwig-Haus, University of Würzburg, Würzburg, Germany
- 3Institute of Tendon and Bone Regeneration, Paracelsus Medical University Salzburg, Salzburg, Austria
- 4Austrian Cluster for Tissue Regeneration, Vienna, Austria
Tendons are often affected by injuries or tendinopathies, resulting in serious and long-lasting impairments. The repair capacity is very low with a high risk of rerupture. Nevertheless, early, moderate and intermittent functional training adapted to the healing process has been shown to support tendon healing. The mechanosensitive tenocytes are responsible for extracellular matrix (ECM) synthesis, a process that is highly dependent on their specific and local mechanotopographical niche. The mechanical stimuli are triggered by the surrounding ECM that are then recognized by the cells via mechanosensation, transduced via activated intracellular molecular cascades to initiate the mechanoresponse, a process known as mechanotransduction. Hereby, the activation of calcium (Ca2+) dependent channels plays an essential role. Moreover, tenocyte primary cilium has been strongly suggested to participate in mechanosensation and -transduction. The cellular mechanoresponse results in processes such as ECM remodeling, collagen fiber alignment, cell proliferation and migration. Diverse approaches have been developed to recapitulate the natural mechanoenvironment and to optimize tenogenesis. It still remains difficult to identify the threshold parameters that determine optimal mechanical stimulation of tenocytes. The diverse effects of mechanical loading on tenocytes are not yet fully understood, as 2D and 3D experiments have not led to consistent conclusions. Further research is needed to fully address the mechanomics of each tendon cell population to gain a more comprehensive picture of cellular mechanoresponses and interdependencies within the tendon tissue that could help to explain possible feedback mechanisms for the regulation of the tendon ECM after mechanical loading. In turn, such efforts and subsequent achievements can help to outlining advanced therapeutic strategies and physiotherapy protocols for tendon health. Future developments in the field of mechanically assisted tendon reconstruction include 4D applications and direct in situ bioprinting.
Native extracellular matrix of tendons
Tendons and ligaments are part of the connective tissues in the body that transmit the forces generated in the muscle to the bone. This is possible due to their abundant and specialized ECM composition (>90% of tissue volume), which predominantly consists of collagens (main component is type I collagen and small levels of type III, V, XI, XII and XIV collagens), small leucine-rich repeat proteoglycans (SLRPs) (e.g., decorin, biglycan, fibromodulin and lumican), large proteoglycans (e.g., versican and small amounts of aggrecan), glycoproteins (lubricin, tenascin C, tenomodulin), adhesion proteins (fibronectin and laminin) and elastic fibers (elastin, fibrillins 1 and 2) alongside the predominant component, water (Varma et al., 2016; Pajala et al., 2009; Smith et al., 2014; Sun et al., 2020; Izu et al., 2021; Ansorge et al., 2009; Yin et al., 2019; Leahy et al., 2023; Eekhoff et al., 2021; Dex et al., 2016; Docheva et al., 2005). Tendons have a hierarchical structure, composed of collagen fibrils, that are grouped in fibers and fiber bundles of different size resulting in the formation of subfascicles and fascicles. The latter and the whole tendon are surrounded by sheathes of loose connective tissue. These layers are called the endotendon that surrounds the subfascicles or peritenon, which encompasses the tendon fascicles and that all together form the interfascicular matrix (IFM), mediating fascicle gliding. A connective tissue sheet named the epitenon surrounds the complete tendon (Schulze-Tanzil et al., 2022; Thorpe et al., 2015; Kannus, 2000; Patel et al., 2021). Due to the parallel collagen fiber alignment in the tendon fascicles that are along to the longitudinal axis of the tissue (in the loading direction), tendons exhibit a high uniaxial mechanical strength (Durgam et al., 2020).
Cellular components of tendons
The described ECM components are synthesized by resident cell types, which are highly specialized fibroblasts, called tenocytes, and other tendon-related cell types such as tendon stem/progenitor cells (TSPCs), myofibroblasts, dendritic cells (DCs), macrophages, as well as so called tenophages (Zhang and Wang, 2010; Szczodry et al., 2009; Lehner et al., 2019). Furthermore, adaptive T cells, endothelial cells and pericytes are also found in tendons (Schulze-Tanzil et al., 2022). Overall, the cellular content is very low in tendons (<10%) (Schneider and Docheva, 2017). The tenocytes as resident cells within the tendon fascicles are longitudinally orientated in rows, whereas those tenocytes in the sheathes formed by the endo-/peri- (IFM tenocytes) and epitenon are less strictly ordered. Within the IFM peri- and epitenon, diverse blood- and lymphatic vessel-derived cell types as well as nerve fibers are located (Zamboulis et al., 2024).
Tendon enthesis
In the gradient attachment zone between tendon and bone, known as the enthesis, there are entheseal progenitor cells that differentiate into fibrochondrocytes in the fibrocartilaginous transition zone and osteoblasts in the bone attachment area (Chen et al., 2023). The enthesis is a mechano-adaptive interface structure that is essential for movement within the joints and provides a firm attachment between tendons and bone, allowing stability and stress reduction between the tissues (Killian, 2022). Moreover, the fibrocartilaginous enthesis is characterized by a spatial regulation of the mineralization front that is mediated by sclerostin (Sost) expression. Yambe et al., have reported fewer sclerostin and fibrochondrocytes in Scleraxis-deficient mice leading to impaired enthesial structure, while Sost knockout mice showed an elevated mineral density and stiffness of their Achilles tendon enthesis (Yambe et al., 2024). During tendon development, cells contain a mechanosensing organelle structure known as the primary cilium that plays an essential role in enthesis formation, via mainly regulation of the hedgehog signaling pathway, although depending upon its mechanical challenges, other pathways are also stimulated. Specifically, mechanical stimulation during in vivo mouse training (a motor-powered treadmill with 10 m/min on a 0° declined lane for 10, 20 or 30 min, 5 days per week) increased the release of transforming growth factor beta 1 (TGF-β1) and promoted tendon healing in the enthesis region via stimulation of TGF-β signaling by the primary cilium (Xiao et al., 2022). The mechanical in vitro stimulation (0%–3% strain; 1 Hz, for up to 24 h) shortened and disassembled the primary cilium of human tenocytes depending upon the TGF-β receptor (Rowson et al., 2018). Other cilium-associated signaling pathways participating in mechanotransduction remain unclear (Fang et al., 2020).
Sensoring of mechanical stimuli by tenocytes
Tenocytes detect mechanical signals (e.g., movements of fibers, or unique biomechanical properties such as stiffness of their environmental substrates) with specific mechanosensitive channels and transmembrane receptors (e.g., connexins, integrins) within their cell membrane (Table 1; Table 2). Many of these receptors are located on the primary cilium (Khayyeri et al., 2015), providing cell surface enlargement and acting as mechanosensory non motile antennae (Lavagnino et al., 2011). As a typical mechanoreceptor of tenocytes, Passini et al., reported that the mechanosensitive ion channel PIEZO 1 underwent conformational changes and allowed a shear-stress induced calcium signaling during mechanical loading of tendons (Table 3) (Passini et al., 2021; Nakamichi et al., 2022). This Ca2+ influx is crucial for initiating downstream signaling pathways that govern tenocyte function and tendon remodeling (Magra et al., 2007). Furthermore, voltage-operated calcium channels (VOCCs), particularly L-type channels like CaV1.2 and TWIK related K+ (TREK)1 channels, are expressed in tenocytes and contribute to calcium signaling upon mechanical loading (Table 3). Activation of these channels enhances intracellular Ca2+ levels, influencing tenocyte behavior (Li H. et al., 2023; Magra et al., 2007). There are further mechanosensitive ion channels like the transient receptor potential (TRP) channels, which form a broad family of non-selective cation channels and especially TRPV4 can reduce mechanically induced Ca2+ signaling (Bian et al., 2024). The stretch-activated channels (SACs), which are also non-specific cation channels that open in response to membrane tension and initiate intracellular signaling cascades such as mitogen activated protein kinase/extracellular signal-regulated kinase (MAPK/ERK) and Wingless (Wnt) via Ca2+ (Millward-Sadler and Salter, 2004) (Table 4). Although not directly mechanosensitive, the P2X, especially P2X7 receptors are activated by ATP release during mechanical strain in tenocytes and contribute to inflammatory signaling and ECM remodeling (Yue et al., 2016) (Table1). This initial process of mechanical signal detection is called mechanosensation (Figure 1).
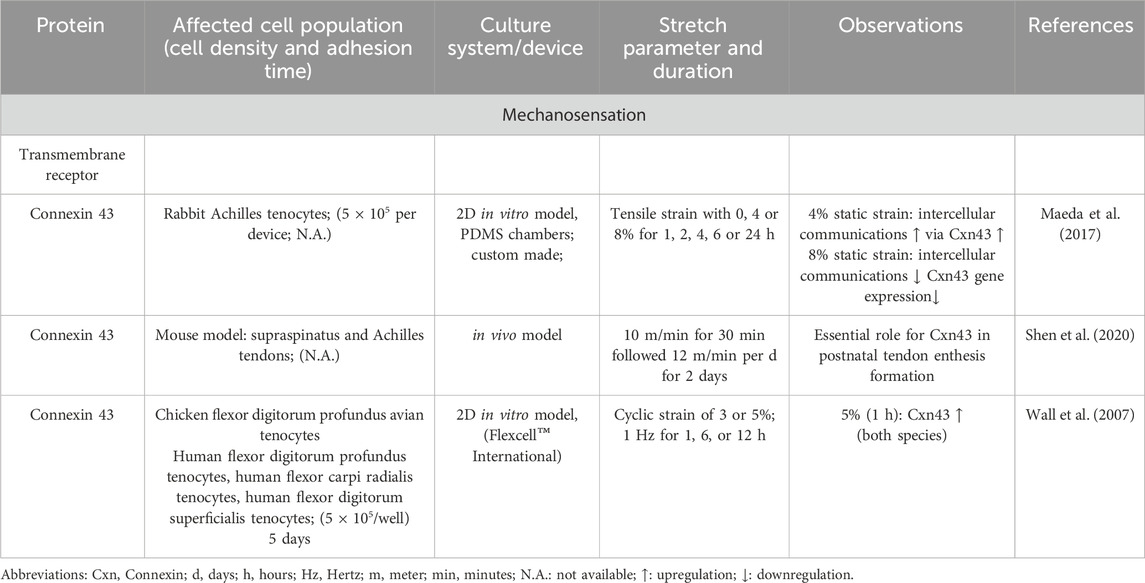
Table 1. The main role of different connexins to sense mechanical stimuli in tenocytes
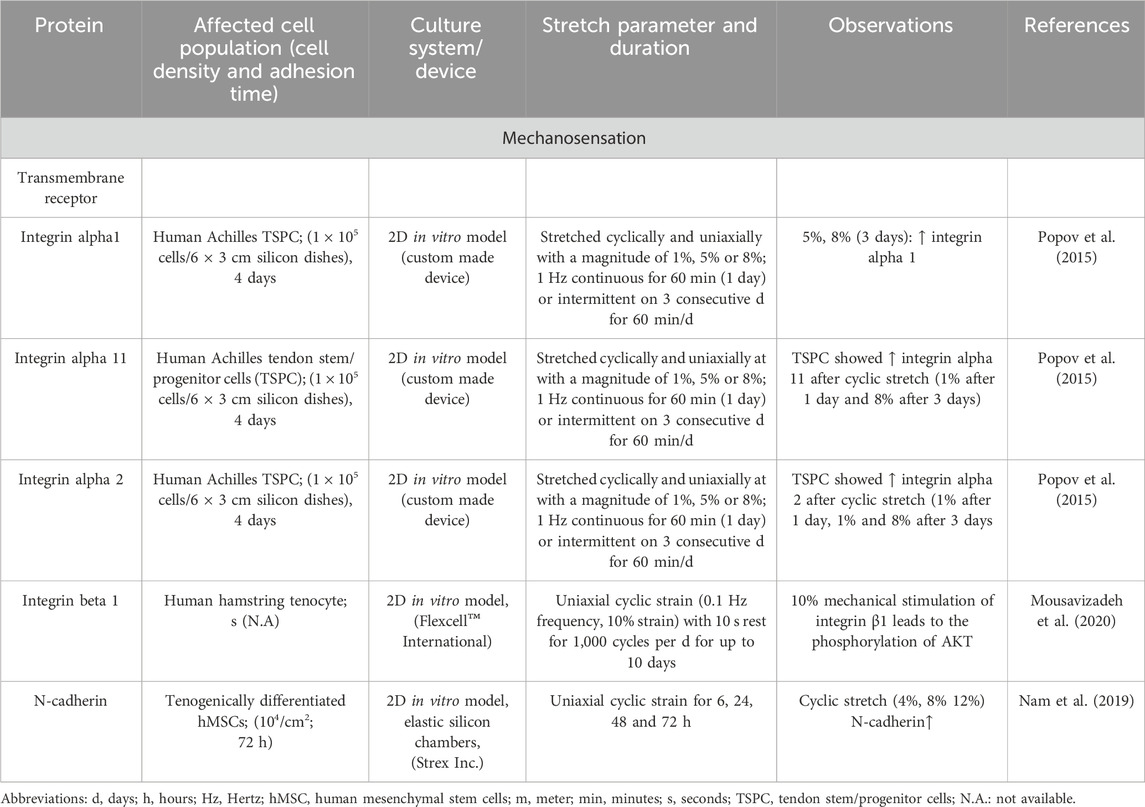
Table 2. The main role of different transmembrane receptors to sense mechanical stimuli in tenocytes
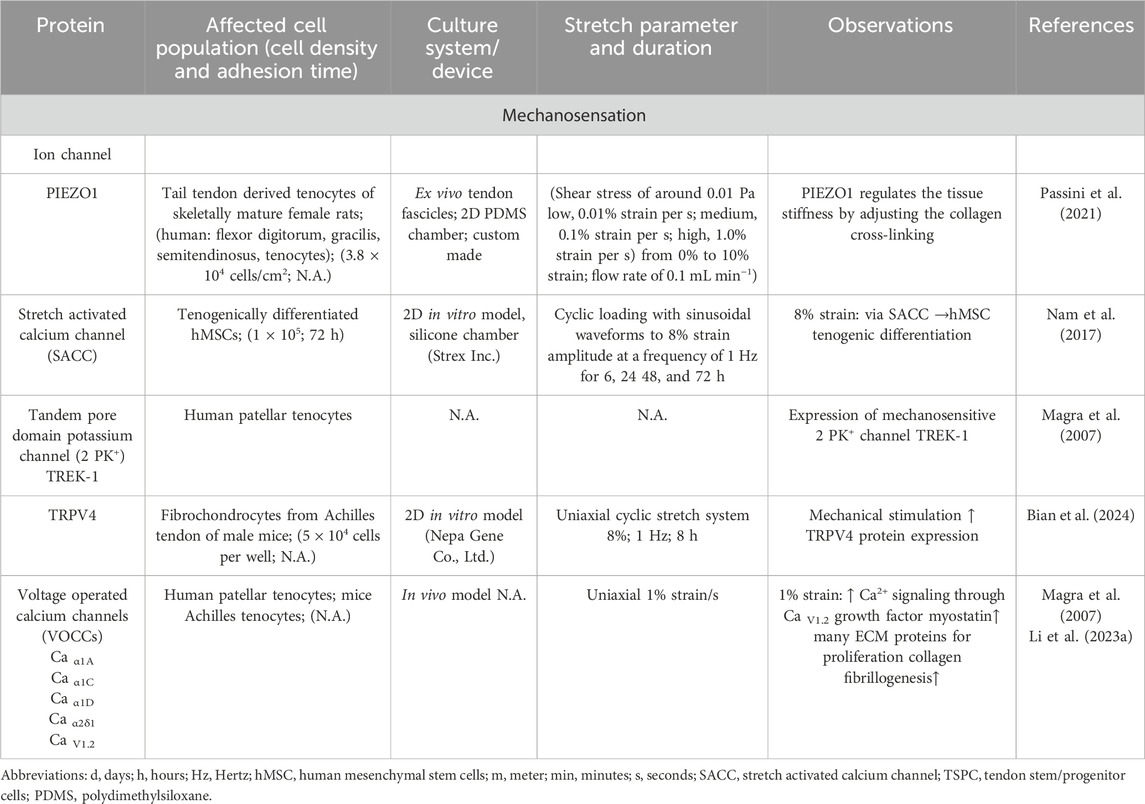
Table 3. The main role of different ion channels to sense mechanical stimuli in tenocytes
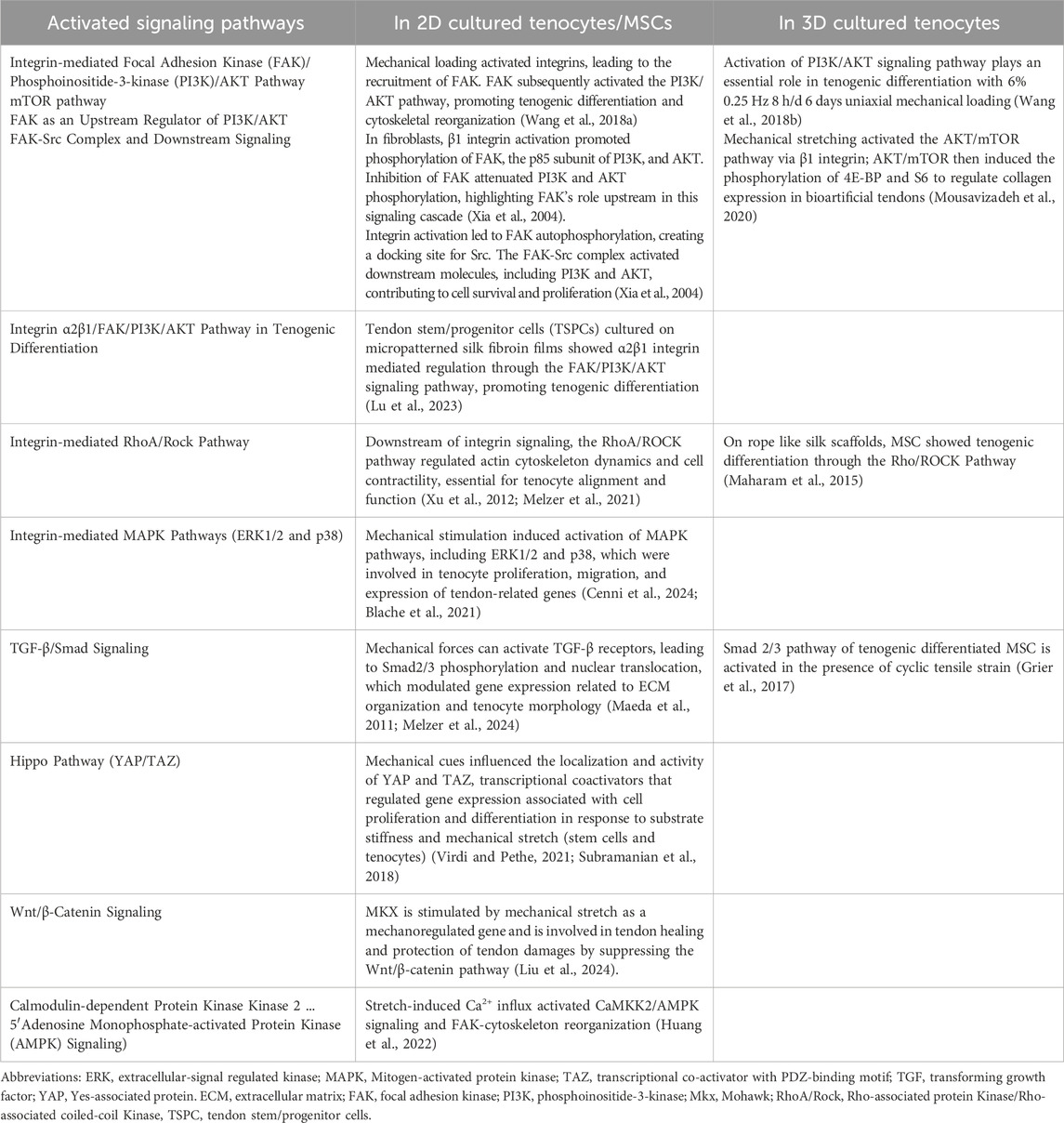
Table 4. Summarizes the activated key signaling pathways of tenocytes or tenogenic cells in 2D and 3D culture.
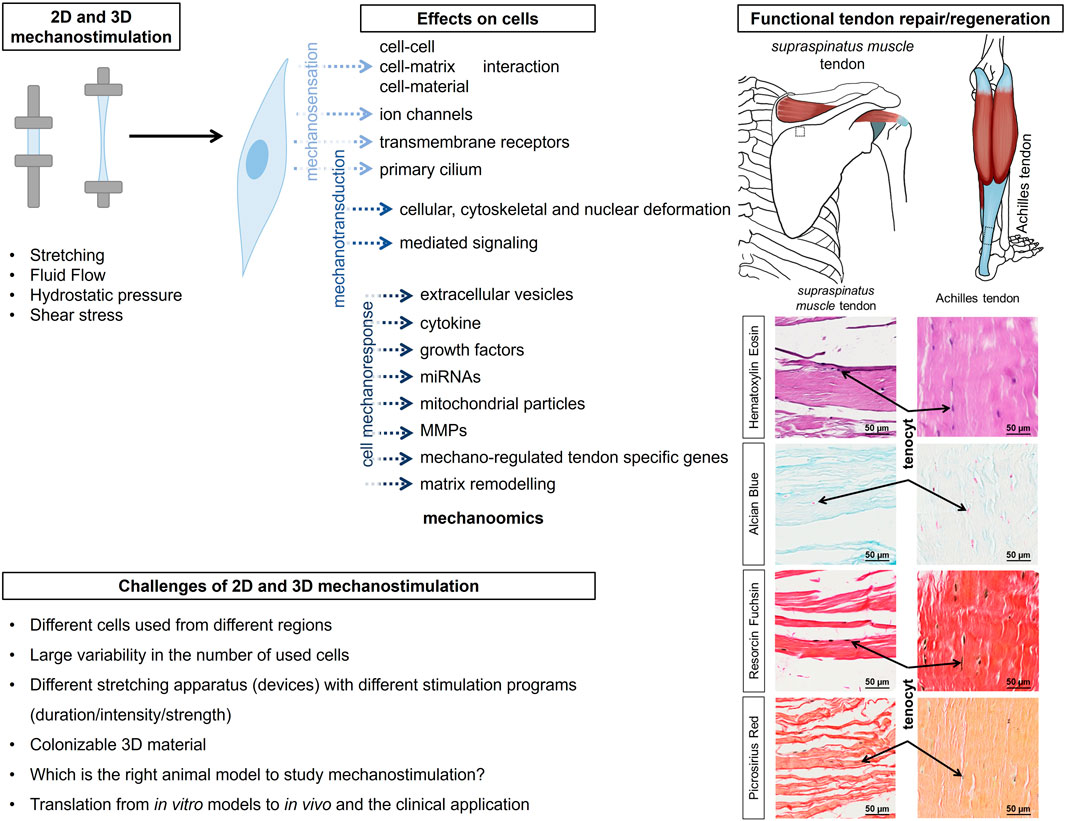
Figure 1. The graphical abstract summarizes the effect of 2D and 3D stimulation on tendon-resident cells for rapid and functional repair/regeneration of tendons, as shown here using the example of the supraspinatus muscle tendon and the Achilles tendon. The inserts show histologically images (Hematoxylin Eosin, Alcian blue, Resorcin Fuchsin and Picrosirius Red staining) and reveal tenocytes lined up like a string of pearls and embedded in or between the collagen fibre bundles. The histological images have been aligned so that they are similar to the anatomical images shown. MMP, matrix metalloproteinases; ECM, extracellular matrix; FAK, focal adhesion kinase; PI3K, phosphoinositide-3-kinase; Mkx, Mohawk; RhoA/Rock, Rho-associated protein Kinase/Rho-associated coiled-coil Kinase, TSPC, tendon stem/progenitor cells.
Mechanotransduction
Then, the mechanical impulse is transduced into biochemical signals leading to a cell response - a process called mechanotransduction. In more detail, van Helvert et al., defined mechanotransduction as a cellular response to tissue organization (ECM stiffness, confinement and topology) and mechanics at subcellular, cellular and multicellular levels generated by the multifaceted interactions between the plasma membrane of the cells and the substrate (van Helvert et al., 2018). Mechanotransduction depends on the interplay between calcium ions (Ca2+) and tenocytes. Mechanosensitive ion channels in tenocytes mediate an influx of Ca2+ in response to mechanical forces such as tensile strain or fluid flow shear stress. This Ca2+ influx activates signaling cascades, which regulate various cellular processes, including cytoskeletal organization, gene expression, and ECM production (Maeda et al., 2013; Huisman et al., 2014). Simultaneous application of cyclic tensile strain and fluid flow shear stress results in a more pronounced increase in intracellular Ca2+ concentration compared to each individual stimulus. This synergistic effect underscores the complexity of mechanical environments in vivo and their impact on tenocyte physiology (Maeda et al., 2013). Furthermore, mechanical stimulation can lead to the propagation of Ca2+ waves between tenocytes via gap junctions. Mechanical stimulation can produce a Ca2+ wave that propagates via the passage of inositol 1,4,5-trisphosphate (IP3) through gap junctions which interconnect neighboring tenocytes. IP3 then acts upon IP3 receptors in the endoplasmic reticulum (ER) to release intracellular Ca2+ into the cytosol. This mechanism highlights the role of gap junctions in facilitating intercellular calcium signaling in response to mechanical stimuli in tenocytes (Lavagnino et al., 2015). Mechanical stimulation of a single cell can initiate intercellular Ca2+ waves that propagate to the tendon tissue network of tenocytes interconnected by gap junctions. These findings suggest that gap junctions play a crucial role in the transmission of Ca2+ signals between cells in response to mechanical stimuli (Iyyathurai et al., 2016; Wall and Banes, 2005). This intercellular communication amplifies the mechanotransduction signal, coordinating a collective cellular response to mechanical cues.
The extracellular Ca2+ ion concentration ([Ca2+]e) modulates tenocyte responsiveness to mechanical stimuli. Alterations in [Ca2+]e can affect the gating of mechanosensitive channels, thereby influencing the magnitude of intracellular Ca2+ signaling and subsequent cellular responses (Wall and Banes, 2005; Chen W. et al., 2015).
Mechanical impulses, registered by integrins as transmembrane receptors, activate downstream signaling cascades, like focal adhesion kinase (FAK), which subsequently trigger the MAPK/ERK and phosphoinositide-3-kinase (PI3K/AKT) pathway. These pathways are crucial for tenocyte migration, proliferation, survival and ECM synthesis (Stanczak et al., 2024). The Yes-associated protein (YAP) and transcriptional coactivator with PDZ-binding motif (TAZ) cascade is a further signaling pathway, which is turned on by mechanical stimulation and influences gene expression related to cell proliferation and ECM production in tendon cells (Randelli et al., 2020) (Figure 1). Mechanical overloading activated the mechanistic target of rapamycin (mTOR) pathway, which can lead to non-tenocyte differentiation and contribute to tendinopathy. Inhibition of mTOR with rapamycin has been shown to mitigate these effects (Nie et al., 2021). An overview of activated key signaling pathways after mechanostimulation was summarized in Table 4.
Effect of mechanical stimuli on cytoskeleton, tendon-specific matrix and cytokines
The mechanotransduction led to nuclear deformation and influenced in the next step the depolymerization of filamentous actin fibers in rat Achilles tenocytes (Chen et al., 2018; Xu P. et al., 2021; Lammerding et al., 2004; Pancheri et al., 2025) (Table 5). This also resulted in a complete reorientation of the cellular structure. Depending on the degree of stretching and strength, this can have different effects on the cytoskeleton (Gögele et al., 2021a; Fleischmann et al., 2024) (Table 6).
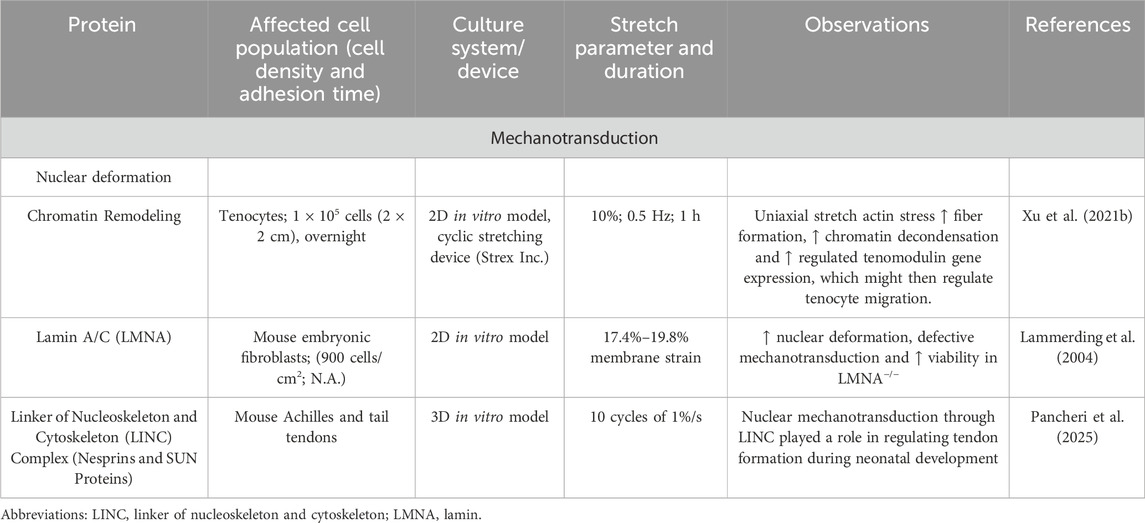
Table 5. Reports on the effect of mechanotransduction on the nuclear deformation.

Table 6. Effect of mechanostimulation on secretion of, growth factors, tendon related transcription factors and cytokines.
Moreover, cell-cell communication between the tenocytes within the longitudinal rows, as well as between the rows, via their plasmalemma extensions passing through the abundant ECM plays a pivotal role to synchronize mechanoresponse via gap junction connections (mainly composed of connexins 32 and 43) at the tissue level (Waggett et al., 2006). There also seem to exist multiple other means of cell-cell communication, facilitating the cell reaction to mechanical stimuli in tendons. Mediators can be released by extracellular vesicles (EVs) in tendon (Song et al., 2023; Zhu et al., 2022). Proliferation, migration and tenogenic differentiation of TSPCs can be induced by small EVs and their local application could be seen as a novel approach to treat tendon injuries (Zhang et al., 2023). It was shown, that rat TSPCs express mechanosensitive miRNA-337-3p upon mechanical loading and the overexpression of this miRNA could effectively rescue in vivo ectopic ossification in rat tendinopathy (Geng et al., 2020). Egerbacher et al., postulated that tenocyte intercellular communication can also occur via ultrathin intercellular structures, called nanotubes (Egerbacher et al., 2020; Waggett et al., 2006). In addition, cell-cell communication in tendons in response to strain-stress can obviously take place through extracellular mitochondrial particles (ExtraMito) from fragmented mitochondria, that are released into the ECM and can regulate the immune microenvironment and recruit macrophages after mechanical stimulation (Chen et al., 2024).
Furthermore, mechanostimulation evokes a release of growth factors including basic fibroblast growth factor (bFGF)2, Early growth response (EGR)1 and 2, Insulin-like growth factor 1 (Scott et al., 2007; Skutek et al., 2001; Herchenhan et al., 2020; Lejard et al., 2011). However, not all of them were upregulated after mechanical stimulation, as TGF-β3 has been shown to be reduced after cyclic stretch (Legerlotz et al., 2013) (Table 7).
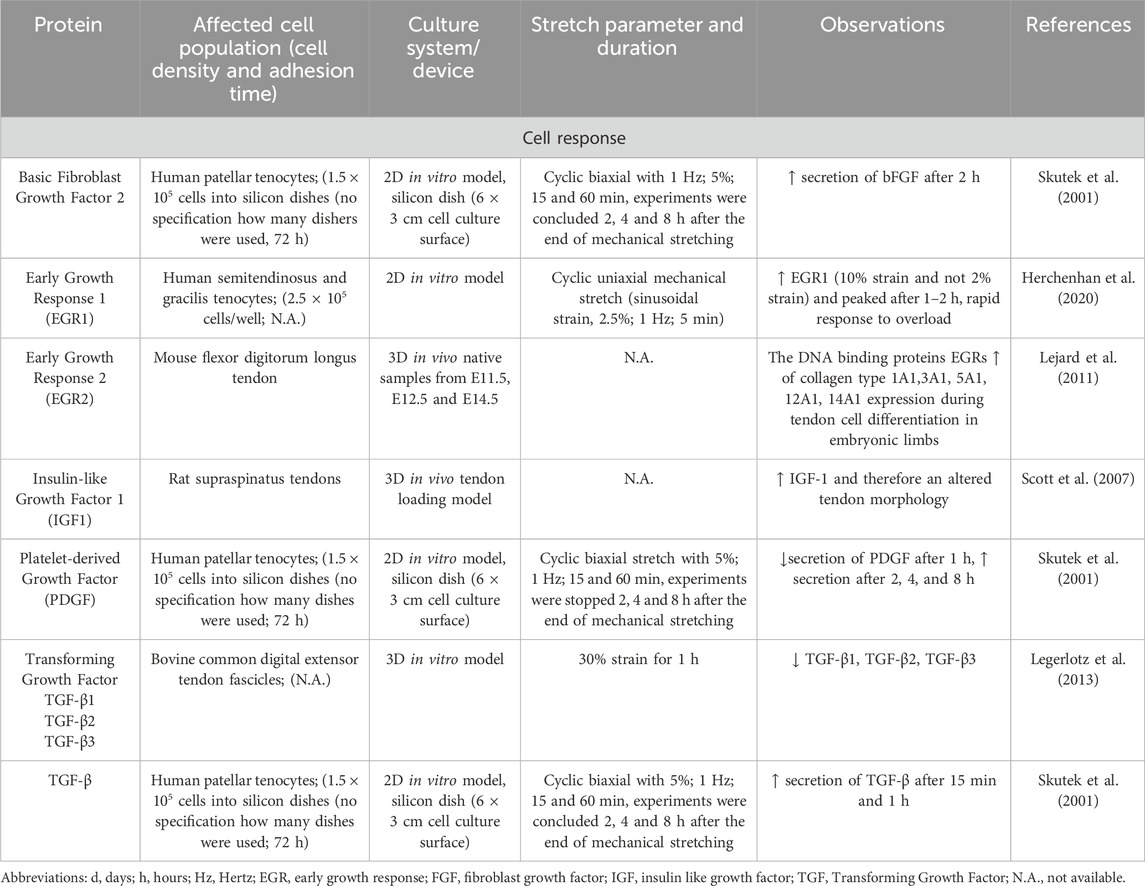
Table 7. Effect of mechanostimulation on secretion of growth factors.
Not only growth factors but also tenogenic transcription factors were upregulated after mechanical loading. In tendon, the mechanical stimulation leads to increased activation of the tenogenic mechanosensitive transcription factor, the Scleraxis basic helix loop helix transcription factor [Scx] (Table 8) (Gumucio et al., 2020; Papalamprou et al., 2023; Kubo et al., 2020), Mohawk [MKX] (Table 9) (Suzuki et al., 2016; Kayama et al., 2016; Liu et al., 2024), EGR1 (Herchenhan et al., 2020), EGR2 (Lejard et al., 2011) and other transcription factors including Runt-related transcription factor 2 [Runx2], sex-determining region Y-box 9 [Sox9]) (Table 10). In addition, mechanical stimulation induced an altered ECM synthesis in tenocytes including tendon-related glycoproteins such as tenomodulin (Dex et al., 2017; Xu P. et al., 2021) (Table 10). Through optimal adaptation to mechanical challenges, a remodeling process can take place (Xu K. et al., 2021; Popov et al., 2015). Maturation and aging trigger tenocytes to shift towards senescence and develop a senescence-associated secretory phenotype (SASP) by activation of the Janus kinase signal transducer and activator of transcription (JAK/STAT) pathway (Tinguely et al., 2023; Yan et al., 2020; Russo et al., 2015; Schulze-Tanzil et al., 2022), leading to dramatic changes in their mechanosensing and mechanotransduction capacity (Zuskov et al., 2020; McCrum et al., 2018; Epro et al., 2017).
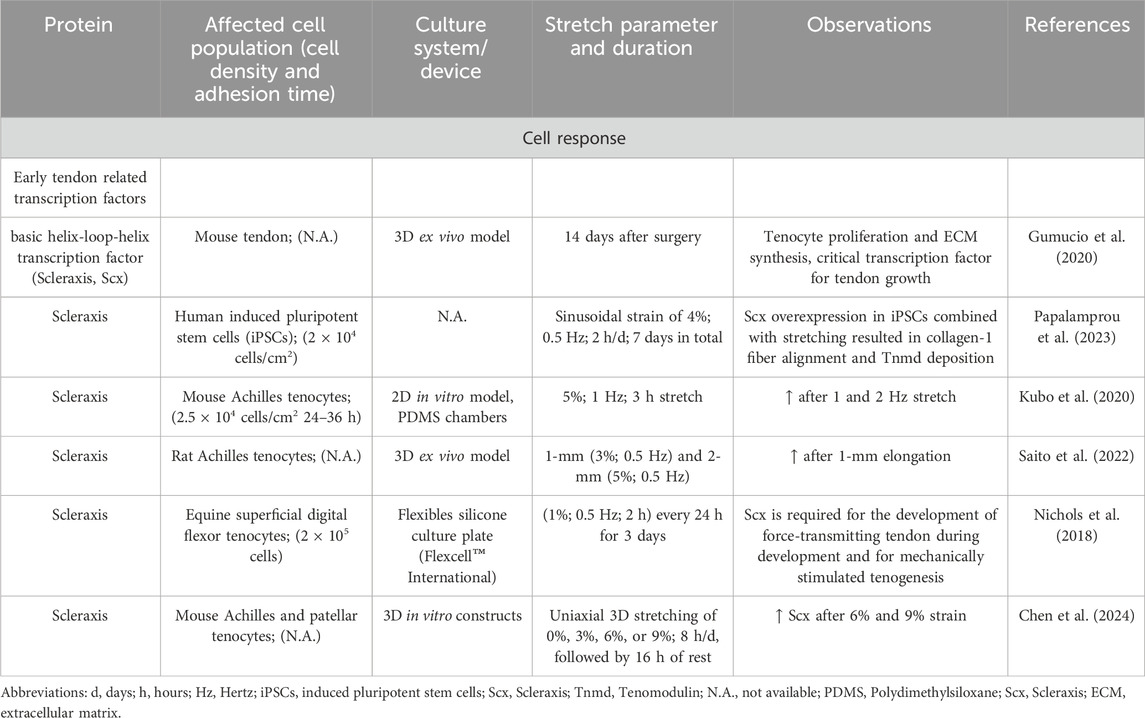
Table 8. Effect of mechanostimulation on secretion of Scleraxis transcription factor.
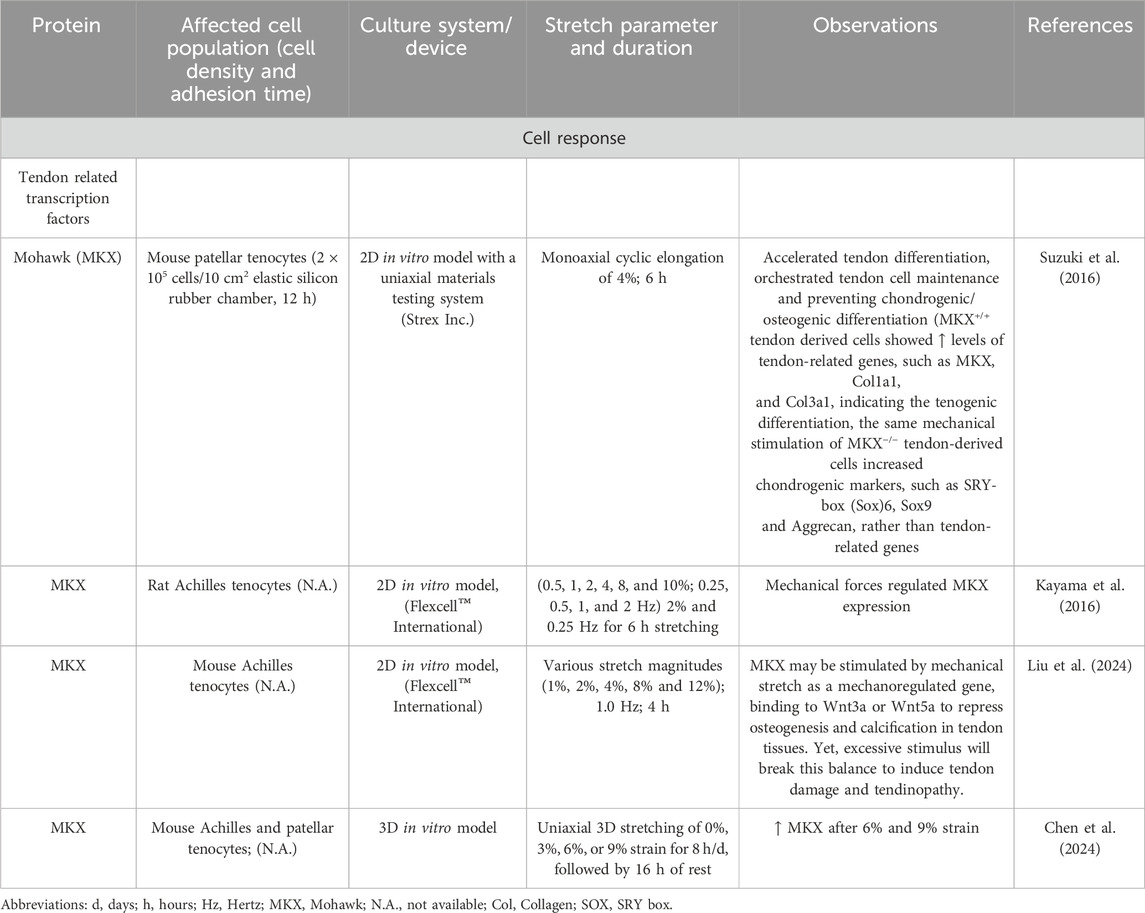
Table 9. Effect of mechanostimulation on secretion of Mohawk tendon related transcription factor.
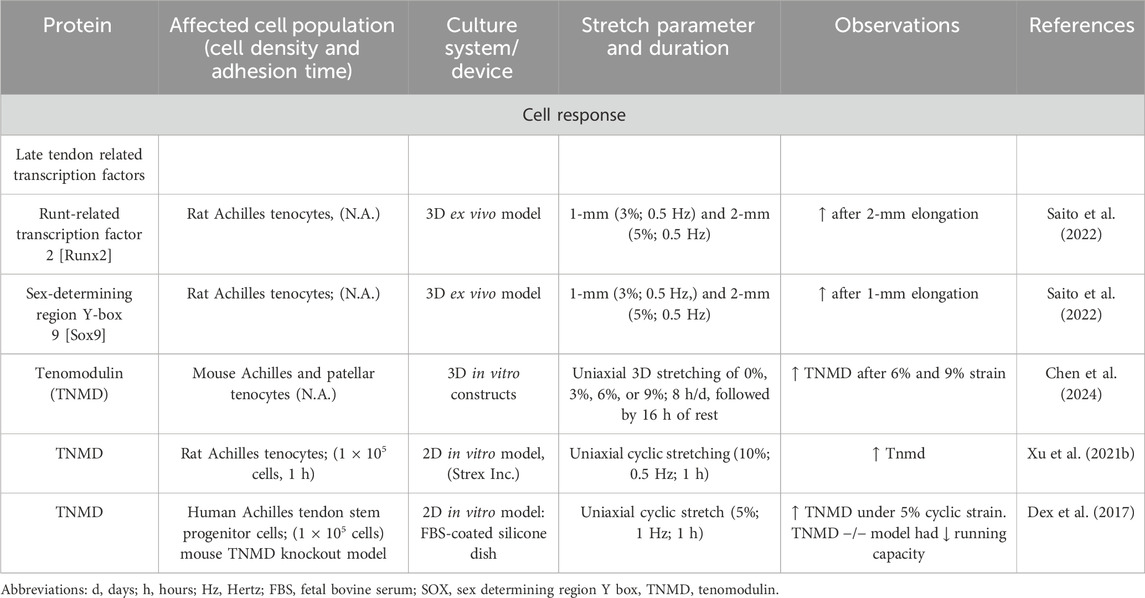
Table 10. Effect of mechanostimulation on secretion of other factors.
The dynamic response of tendons to physical activities is enabled by their biomechanical visco-elastic behavior that is mediated via their unique ECM composition (Notermans et al., 2019). In addition to cyclic stretching, mechanical impulses on tendon cells evoke also repetitive fluid flow of water by binding to and releasing from negatively charged glycosaminoglycans, as well as shear forces caused by fascicle and fiber gliding (Mlyniec et al., 2021) and in some areas, pressure (Figure 2).
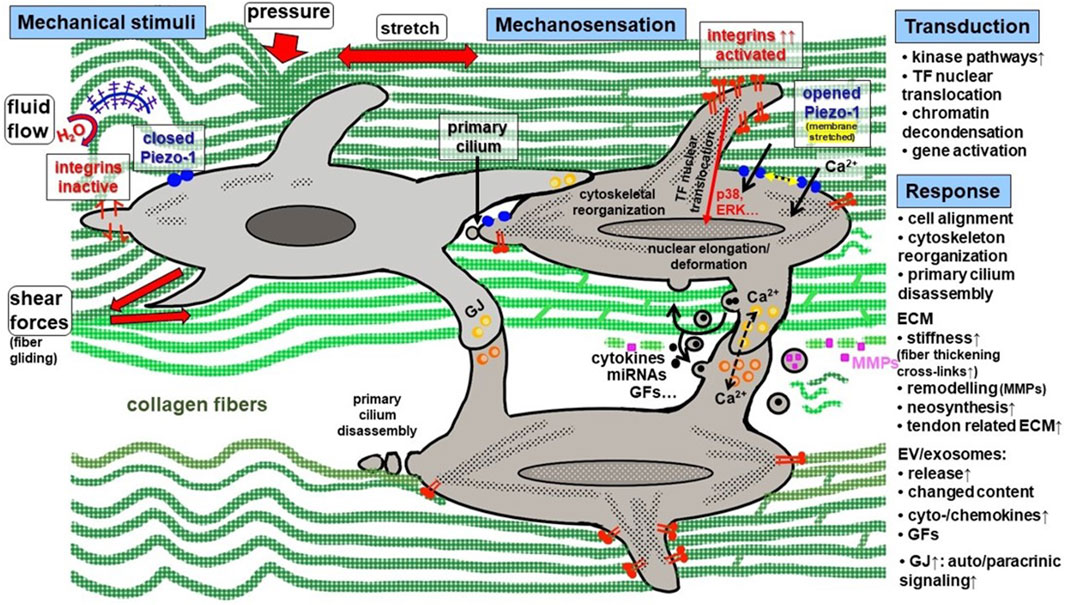
Figure 2. Synopsis of mechanical stimuli and the key steps of mechanosensation, -transduction and–response in tendon. Mechanical stimuli in tendon are: cyclic tension, pressure, fluid flow, shear forces. Mechanosensing is mediated by a group of receptors among them integrins (activated by changes in ECM stiffness) and Piezo-1 channels (activated by membrane stretching). In addition, the primary cilium is a mechanosensing organelle in tenocytes. It goes along with nucleus deformation. Mechanotransduction occurs in response to integrin activation, clustering, focal adhesion component activation, cytoskeleton reorganization and Ca2+ influx, leading to transcription factor nuclear translocation mediated by different activated pathways including typical kinases. Cell response comprises of the release of diverse factors and initiation of auto-/paracrinic response and neomatrix synthesis. Moreover, cells are synchronized by enhanced gap junction expression. ECM, extracellular tendon matrix; ERK, extracellular signal regulated kinase; EV, extracellular vesicles; GF, growth factor; GJ, gap junction; MMP, matrix metalloproteinase; TF, transcription factor.
The remodeling of the ECM is accomplished by matrix metalloproteinases (MMPs), among other things. Mechanical stress has a significant influence on this remodeling through the increased or suppressed release of MMPs, as the following examples show. There was a decrease in MMP1 (Yang et al., 2005) and increase in other types (MMP9, 13, 14) (Saito et al., 2022; Nichols et al., 2018; Lohberger et al., 2016). Patel et al. postulated the upregulation of MMP3 after cyclic loading in a new fiber composite hydrogel may be early responses to changes in the tenocyte environment (Patel et al., 2018) (Table 11).
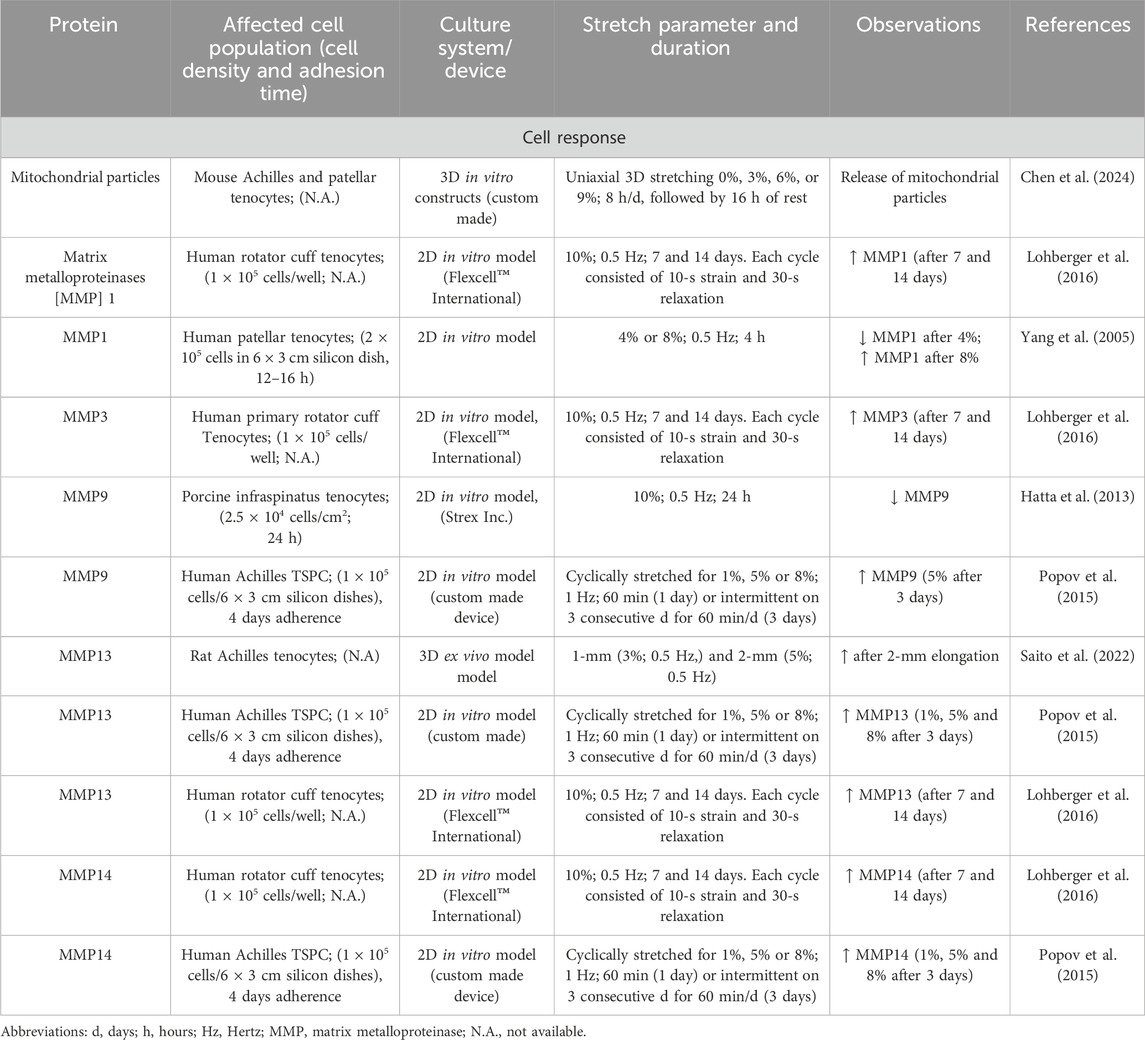
Table 11. Effect of mechanostimulation on secretion of mitochondrial particles and MMPs.
This cell response to mechanosensation orchestrates the release of mediators and the auto-/paracrinic response to them, including not only growth factors, but also cytokines, and neurotransmitters (Tsuzaki et al., 2003; Mousavizadeh et al., 2024; Skutek et al., 2001). Cytokines like interleukin (IL)-1β may act as a defense/survival factor in early cellular responses to mechanical loading (Qi et al., 2006). However, the release of IL-1β also depends strongly on the environment (2D or 3D) and the strain parameters (Yang et al., 2005). Especially, IL-6, that was already significantly upregulated after 1 h and 30% strain in native bovine foot extensor tendons and seemed to play a role in tendon adaptation by regulating healing processes (Legerlotz et al., 2013). This upregulation of IL-6 after cyclic loading could be seen too in a new fiber composite hydrogel (Patel et al., 2018). Skutek et al. investigated the effects of cyclic biaxial mechanical stretching (1 Hz frequency and 5% amplitude) on human tendon fibroblasts and found that such mechanical stimulation modulates the secretion patterns of various growth factors, and the cytokine IL-8. Their findings suggest that mechanical stretch can indeed induce IL-8 expression in these cells (Skutek et al., 2001). Mechanical stretching of human patellar tendon fibroblasts controlled the expression of pro-inflammatory markers. To date, however, there is no uniform opinion regarding the connection between increased stretching and inflammatory markers. Specifically, low-magnitude stretching (4%) reduced IL-1β-induced expression of MMP-1, cyclooxygenase-2 (COX-2), and prostaglandin E2 (PGE2) production, whereas high-magnitude stretching (8%) enhanced their expression. Furthermore, cyclic mechanical stretching of human tendon fibroblasts increased PGE2 production in a magnitude-dependent manner. Stretching at 8% and 12% of silicon membranes for 24 h resulted in 1.7-fold and 2.2-fold increases in PGE2 production, respectively, compared to non-stretched controls. The study also reported increased expression of COX-1 and COX-2, indicating that the enhanced PGE2 production was associated with elevated COX expression (Wang et al., 2003). This indicated that the magnitude of mechanical stretch influences the inflammatory response in tendon cells (Yang et al., 2005; Chen W. et al., 2015).
Mechanical loading significantly influences the expression of various neuropeptides (neuropeptide Y [NPY], Substance P, Calcitonin gene-related peptide [CGRP]) and neurotransmitters (Catecholamines) in tendons, affecting both their healing and pathological processes (Table 12). An augmented expression of NPY involved in various physiological processes, including pain modulation and delayed tissue remodeling could be observed after mechanical stimulation in tendons (Bjur et al., 2009; Chen et al., 2021). Another neuropeptide is Substance P, which is a mechanoinduced regulator of human Achilles tenocyte proliferation (10% strain with a frequency of 1 Hz for a total of 120 min) and creates therefore probably a healing effect (Backman et al., 2011).
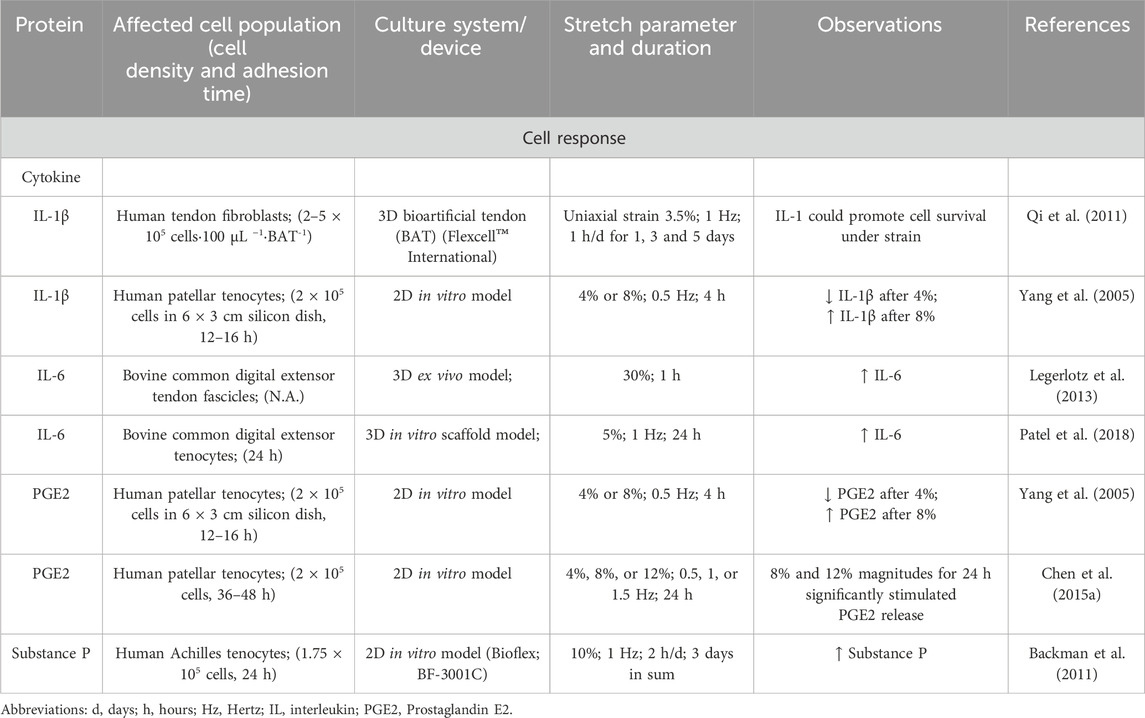
Table 12. Effect of mechanostimulation on secretion of cytokines.
Nevertheless, the question arises as to how the mechanoresponse of tendon cells is influenced during healing processes.
Mechanoimpulses and tendon healing response
The ability of tendons to regenerate is largely restricted by their hypocellularity, limited blood supply, low proliferation rate and the post-traumatic inflammation (Nichols et al., 2019). In principle, the healing process can be divided into four overlapping phases (hemorrhagic, inflammatory, proliferation, remodeling phases) (Schulze-Tanzil et al., 2022; Schneider et al., 2018). Directly after tendon injury, the hemorrhagic phase leads to bleeding and also to a release of platelets and immune cells, such as neutrophils, lymphocytes and other leukocytes. The latter initiate an inflammatory response (inflammatory phase) (Rosenbaum et al., 2010). No studies were found, that investigated the effect of mechanical loading on tendons during the hemorrhagic phase. The inflammatory phase is triggered in the first 72 h after an injury via autocrine and paracrine signals. During this inflammatory phase, macrophages migrate into the ruptured structure and the adjacent tissue and phagocytize tissue fragments and necrotizing material (Manning et al., 2014). Despite so far not investigated for tendon healing, macrophage`s proinflammatory M1 polarization was enhanced by 3% strain (Babaniamansour et al., 2024). Growth factors (e.g., platelet derived growth factor [PDGF], epidermal growth factor [EGF], IGF, vascular endothelial growth factor [VEGF] and TGF) are released, which promote granulation and neovascularization due to a chemotactic effect on endothelial cells and tenocytes (Molloy et al., 2003). This stimulation causes proliferation, synthesis of collagen types I, III, V and non-collagenous proteins and angiogenesis (proliferation/granulation phase). As a follow up to this process of new ECM synthesis, the orientation of the cells in longitudinal rows and the subsequent predominately parallel orientation of the collagen fibers play an important role in order to regain the structural and functional tissue features (remodeling phase). This phase is characterized by decreased cellularity, reduced collagen and glycosaminoglycan synthesis, and the transition of repair tissue into a reorganized functional tissue. The ECM and cell realignment represent processes strongly guided by mechanical strain. Introducing mechanical stimuli shortly after tendon injury can modulate the healing environment. However, excessive loading during the initial inflammatory phase may disrupt the delicate balance required for effective healing (Hammerman et al., 2024). Mechanical loading during the proliferative phase (5 days post-injury) of tendon healing has been shown to increase the quality and efficiency of rat Achilles tendon repair by increasing tendon-related gene expression (Eliasson et al., 2009). Specifically, loading increased the expression of inflammation-related genes such as IL-1β, inducible nitric oxide synthase (iNOS), prostaglandin E synthase (PGES), IL-6, and Chemokine CC motif ligand 7 (CCL7). These findings suggest that mechanical loading during the proliferation phase can modulate both inflammatory responses and tissue remodeling processes (Hammerman et al., 2018; Hammerman et al., 2017).
Mechanical loading during the remodeling phase of tendon healing (typically commencing around 4–6 weeks post-injury) is pivotal in enhancing tissue organization, strength, and function. Appropriate mechanical stimuli during this period can significantly influence the quality of tendon repair and overcome a frequent complication that is post-traumatic tendon heterotopic ossification (HO). Interestingly, certain tendon-specific factors, such as tenomodulin are involved in prevention of tendon HO. Delgado et al. have shown that loss of tenomodulin results in significantly increased HO during the tendon remodeling phase in the mouse Achilles tendon, which was concomitant with compromised biomechanical properties (Delgado Caceres et al., 2021). Mechanical loading facilitates the reorganization of collagen fibers, promoting their alignment along the direction of tensile forces. This alignment enhances the mechanical properties of the tendon, increasing its stiffness and tensile strength. Additionally, loading stimulates the formation of covalent bonds between collagen fibers, further strengthening the tissue. These adaptations contribute to the restoration of tendon function and reduce the risk of re-injury. Hence, appropriate mechanical stimuli are essential to promote healing, maintain tendon homeostasis and prevent the development of tendinopathy (Stanczak et al., 2024).
However, neither the morphological structure nor the biomechanical properties can be fully restored to their original state after a tendon rupture. This is due to the abnormal concentration, organization and cross-linking of collagen type I fibrils (crosslinks) (Frank et al., 1992; Frank et al., 1995). Furthermore, the ratio of collagen types I and III is unfavorable and even if there is an increase in collagen type III fibers during the initial repair phases, these are thinner than those in healthy tendons (Eriksen et al., 2002). The stability of repaired tendons is reduced by altered properties such as the degree of cross-linking, the fibril thickness, density and composite formations with other ECM components. The healed tissue is characterized by the variant composition of the tendon neo-ECM with other ratios of collagen and proteoglycan types. In addition to local molecular factors, differing from the non-injured tendon ECM, novel heterogeneous cell populations, either immigrated or amplified during healing could be detected (Freedman et al., 2018; Andarawis-Puri and Flatow, 2018; Maffulli et al., 2002; Chamberlain et al., 2011). Furthermore, the intimate interaction between resident (“intrinsic”) cell populations and extrinsic cells, including immune cells, might substantially change the overall mechanoresponse in a healing tendon, unless they decline over time.
Cells respond intimately to their biomechanical microenvironment, such as ECM stiffness. Therefore, this complex healing scenario gives rise to a multitude of questions. For example, how could the synchronized mechano-related cell-cell communication being reestablished in the damaged tendon during healing, or how to coordinate the mechanoresponse in a functional manner again? The posttraumatric inflammatory microenvironment might strongly influence cellular mechanoresponse but it still remains unclear how this microenvironment is exactly changed. An issue that demands further research.
Insufficient tendon healing has been recently considered as a result of a lack of vascularization, cell rounding, disorganization of the collagens in the ECM, neuronal ingrowth, persistence of inflammation and a change in metabolic processes (Wunderli et al., 2020). All these parameters might change the mechanical niche in healing tendons. It remains questionable whether and if so, how these unfavorable processes impact and lead to aberrant tendon healing and how could they be counteracted by a properly adapted exercise protocol. Since tendon healing partially recapitulates steps of tendon development, a more detailed understanding of the role of mechanobiology in developmental tenogenesis would be helpful in the future. Meanwhile, it is well known that mechanical stimulation is crucial in the early intrauterine development of tendons (Okech and Kuo, 2016). This observation supports the conclusion that mechanical stimuli have a significant influence on the tendon ECM synthesis and maturation, which is helpful to properly guide healing processes biomechanically in the future. Moreover, exercise protocols might also be adapted to individual healing scenarios leading to personalized mechanostimulation.
In summary, it has clearly been shown that mechanostimulation has an effect on the healing process of tendon and ligament tissues (Young et al., 1998; Awad et al., 2000; Juncosa et al., 2003). However, it is still unclear which parameters (amplitude, frequency, direction and duration) represent an optimal condition for functional healing. The influence of distinct protocols of mechanostimulation on -sensation, -transduction and -response (e.g., change in cellular function), still needs to be clarified, since the mechanical stimulus can be exerted on the cells via or as a combination of stretching, fluid flow, hydrostatic pressure or shear stress.
The effect of mechostimulation in 2D culture
Diverse devices for 2D stimulation of tenocytes and TSPCs could be used to enhance the proliferation rate, metabolism and directional deposition of ECM components (Gögele et al., 2021a; Chernak Slane and Thelen, 2014; Xu P. et al., 2021; Passini et al., 2021; Rowson et al., 2018). Mechanical stimuli in 2D are one reason for cell proliferation, migration, tissue remodeling, and cellular processes including metabolism, gene transcription and cell differentiation (Papalamprou et al., 2023; Morita et al., 2019; Huang et al., 2023). During 2D mechanotransduction numerous mechanosensitive ion channels become activated, chromatin conformation changes and activation of transcription factors, as well as the release of various growth factors (TGF-β, VEGF, PDGF, bFGF) takes place (Subramanian et al., 2018; Petersen et al., 2004; Skutek et al., 2001). The transmission of mechanical signals, sensed at the plasmalemma, transduced via actin stress fibers to the nucleus causes chromatin decondensation and thus, promotes the expression of tenomodulin, whereby tenocyte migration was increased (Xu P. et al., 2021) (Figure 2). In 2D, it could already be shown that Ca2+ mediated mechanosensing occurring via cyclic stretch activated the calmodulin-dependent protein kinase kinase 2 (CaMKK2)/5′adenosine monophosphate-activated protein kinase (AMPK) signaling and the (FAK)-cytoskeleton reorganization, and thereby, helps to maintain tenocyte phenotype and homeostasis (Huang et al., 2022). In another study, the authors demonstrated that TSPCs, mechanostimulated in 2D, responded to complex changes in gene and protein expression of integrins, MMPs and ECM proteins that is mediated via well-known mechano-regulated kinases, namely, p38 and ERK1/2 (Popov et al., 2015). We also showed that at 14% stretch in 2D culture, anterior cruciate ligament-derived fibroblasts aligned against the direction of stretch, thereby altering their cytoskeleton by becoming even more spindle-shaped and slender than in the unstimulated control (Gögele et al., 2021a). This was confirmed in a recent study using rat Achilles tenocytes in the same in vitro model (Fleischmann et al., 2024). This phenomenon is contrary to the situation in 3D tissues, whereby there is a parallel cell alignment in the direction of axial extension. However, 2D stretch is limited by the fact that cells are not surrounded by their own ECM providing their natural mechanotopographical niche (Leal-Egana et al., 2020). Moreover, differentiated tenocytes devoid of their ECM usually dedifferentiate in a 2D environment that might also affect their mechanosensing and -transducing pathways. At this point, it should be mentioned that most 2D mechanostimulation experiments are carried out by means of cyclic stretching via stretched silicone membrane of different stiffness. There is a great need to study more thoroughly the effects of hydrostatic pressure or shear stress on tendon-relevant cell types.
3D stimulated cell-material constructs
3D stimulation of tendon-derived cells requires a 3D cell carrier to optimize tendon biofabrication applications. There are numerous biomaterials (e.g., diverse synthetic and natural polymers including collagen, silk forming scaffolds (Shiroud Heidari et al., 2023; Xue et al., 2022) or hydrogels (Wan et al., 2024; Bs et al., 2024; Li S. et al., 2023) that are used in the tendon biofabrication field based on the fact that they try to mimic the native tendon ECM as closely as possible (Gehwolf et al., 2019; Stoll et al., 2011; Hahn et al., 2019). These scaffolds alone, already have some capacity to guide regeneration due to similar mechanical strength compared to the native tendon/ligament (Hahn et al., 2019; Miescher et al., 2024). It has been shown that precolonized scaffolds with primary human adipose-derived mesenchymal stem cells (hADMSCs) or induced pluripotent stem cell-derived mesenchymal stem cells (iPSCs) can support defect healing in vivo even more efficiently compared to uncolonized scaffolds (Jiang et al., 2020; Kaneda et al., 2023). The interplay between scaffold, cells and mechanical loading is not yet fully understood. The phenotypical switch of macrophages towards M2 that is potentially beneficial for tendon healing, should be investigated in more detail in the future (Schoenenberger et al., 2020). Bioreactors are required to cultivate cell-loaded scaffolds in vitro in order to be able to properly simulate the natural microenvironment (Gögele et al., 2021b). The use of force-transmitting 3D bioreactors allows us to understand the mechanobiology in more detail, thereby reducing the number of in vivo experiments. Like in 2D, ECM synthesis and cell differentiation are supported by cyclic stretch (Burk et al., 2016; Zhang et al., 2018). In contrast to 2D, cells start alignment parallel to the direction of stretch, very similar to their orientation in native tendon. However, similar to 2D culture, there is still no uniform consensus on the duration, amplitude and frequency of stimulation under 3D conditions to promote tenogenesis by using different tenocyte subpopulations or stem cells (TSPCs or mesenchymal stromal cells [MSCs] from bone marrow or fat tissue) (Chen et al., 2022; Wang et al., 2018b; Rinoldi et al., 2019; Russo et al., 2022; Xu et al., 2015). In most 3D studies, stimulation is exerted by cyclic stretch. Fluid flow and pressure are also expected to stimulate tenogenic cells, but should be validated more extensively in the coming years. Moreover, the stimulation protocols must be properly adapted to the respective cell types. However, for this reason, the comparability of the different studies on the tendon mechanotransduction under 3D conditions is limited and the interpretation of results remains controversial, due to the specific advantages and disadvantages of different systems.
Advantages and disadvantages of different 2D and 3D stretch devices
Depending upon the research question addressed, there is now a whole range of different devices for 2D and 3D stimulation of tendons or tenogenically differentiated cells. A brief overview of the advantages and disadvantages of 2D (Table 13) and 3D (Table 14) stimulators is given.
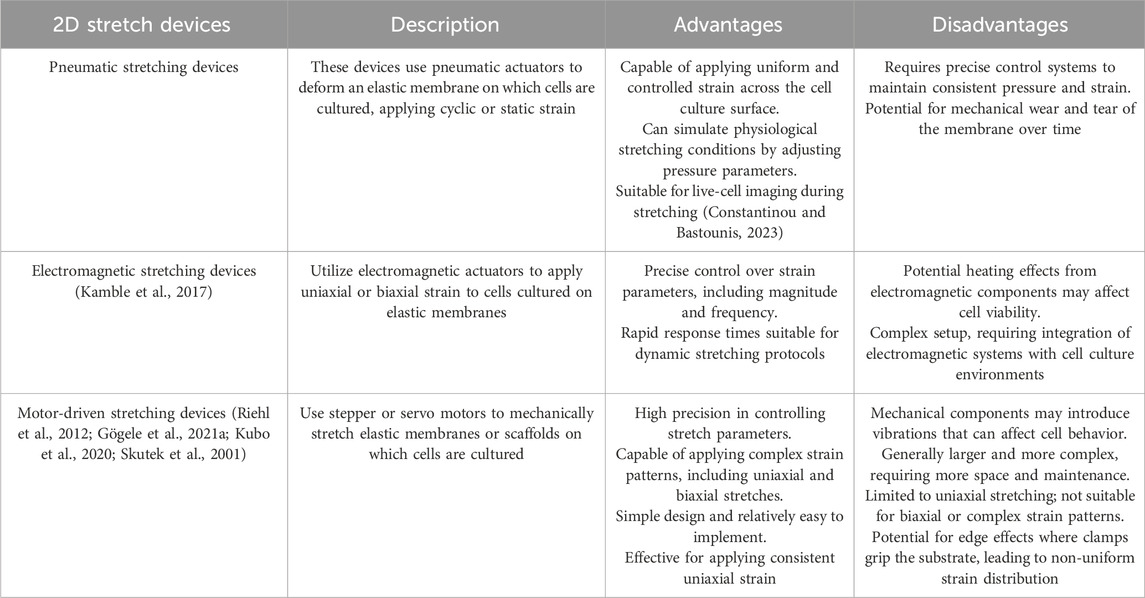
Table 13. The advantages and disadvantages of different 2D stretch devices for tenocyte/tendon stimulation.

Table 14. The advantages and disadvantages of different 3D stretch devices for tenocyte/tendon stimulation.
It should be emphasized at this point that stimulation devices are still needed that can simulate all types of mechanical stimuli in the tendon. With the help of these devices, it may be possible to establish tendon defects/implantation models on chip, in which human cells are also used, so that a regenerative approach can be developed. Another point that has not yet been addressed and should be investigated further in the future, is the fact that tendons and ligaments are usually not stretched in just one direction. For example, in the anterior cruciate ligament, there is both translational and rotational movement of the tissue. To date, there are no commercial devices that can better depict both movement sequences and thus the native state.
Discussion
To summarize, it can be concluded that moderate cyclic mechanical stimuli lead at the cellular level to enhanced cell-ECM interactions and the activation of anabolic molecular processes that are important for tissue homeostasis and supportive for tendon healing. To deepen the understanding of tendon mechanobiology new methods such as RNASeq (transcriptomics) are currently being used to distinguish by single cell analysis multiple novel cell subpopulations in the tendon that react and possibly interact in a diverse manner in response to different mechanical challenges (Marr et al., 2021; De Micheli et al., 2020; Sarmiento and Little, 2021; Ramos-Mucci et al., 2022; Kendal et al., 2020).
The tendon-on-chip technique would be an interesting tool for tissue or single cell analyses (Bakht et al., 2024). The advantage of tendon-on-chip technology compared to others is that a mechanoniche with several cell types could be created in a very small space. It allows us to gain a better understanding of healthy tissue and the causes of tendon fibrosis by investigating the intimate crosstalk between tenocytes and distinct inflammatory mediators (Awad et al., 2025; Ajalik et al., 2025). In order to better characterize the native conditions in vitro, organ-on-chip techniques with additional fluid flow were designed to mimic vascular microenvironments of injured tendons (Linares et al., 2025). Crosstalk analysis between different cell types under biophysiological and biochemical conditions has also been tested with human-adipose tissue derived stem cells to study tendon physiology (Monteiro et al., 2023). The organ-on-chip technique can be used not only for tendons but also to gain research insights into chronic inflammation of the enthesis (Giacomini et al., 2024). Although, it should be added that it has so far only been tested with fluid flow and has not been subjected to dynamic cyclic stretch conditions, which would be very helpful for the future and would allow us to stimulate extensive mechanical stress or overload that can also induce tendon pathologies. In addition, tendons compromised by background diseases (e.g., diabetes) or aging might respond in a different manner to mechanical challenges, and this could be investigated in tendon-on-chip disease models.
Age-related changes in tendon mechanosensitivity involve alterations at cellular, structural and functional levels, impacting the tendon’s ability to respond and withstand to mechanical stimuli (Grote et al., 2019; Thampatty and Wang, 2018). Aged tendons exhibit changes in mechanotransduction pathways, leading to diminished responses to mechanical loading (Aggouras et al., 2024; Dudhia et al., 2007). This includes alterations in focal adhesion components and cytoskeletal organization that are critical for mechanosensing (Arnesen and Lawson, 2006; Kwan et al., 2023; Leahy et al., 2024).
Such changes can impair the tendon’s adaptive responses to mechanical stress (Epro et al., 2017; McCarthy and Hannafin, 2014; Kwan et al., 2023). Furthermore, aging decreased cellularity in tendons and tenocyte`s proliferation and metabolic activity. This decline in cellular functions affected the tendon’s capacity for repair and adaptation to mechanical stimuli (Lui and Wong, 2019; Svensson et al., 2016).
Tenocyte senescence in aged tendons impairs the capacity for ECM remodelling (Stowe et al., 2024). Hence with aging, tendons undergo structural modifications, such as disorganized collagen fiber arrangement and increased presence of non-tendinous tissue components, such as fatty deposits and calcifications. These changes can compromise the tendon mechanical integrity and its ability to transmit forces effectively to fulfil its key function (Lui and Wong, 2019; Goh et al., 2018).
The discussion of age-related changes in mechanosensitivity can only be dealt with briefly here, since as there are now many comprehensive studies on this topic (Epro et al., 2017; Korcari et al., 2023; Carroll et al., 2008). The study by Aggouras et al. (2024) confirmed that young and aged tendons display different mechanisms of ECM remodeling in response to tensile loading, supporting the notion that tendon pathologies require age-specific clinical interventions and therapies (Aggouras et al., 2024). In addition, Kwan and colleagues (2023) have already published a review on this topic and no more recent literature could be found that investigated the difference in the effects comparing 2D and 3D mechanostimulation of aged tenocytes in vitro or in vivo (Kwan et al., 2023).
A better understanding of mechanotransduction is also needed, in order to treat tendon injuries with the help of pre-stimulated cell-seeded 3D scaffolds. Transferring mechanobiological principles into the design of biomaterial scaffolds is pivotal for enhancing tissue regeneration by amplified cell proliferation and ECM synthesis. Mechanomimetic scaffolds can be engineered to optimally transduce mechanical cues that guide tissue formation. The key strategies for designing these scaffolds are to provide an optimized scaffold architecture (e.g., by adapting porosity, pore size, fiber alignment), avoiding the risk of cell squeezing under strain, the utilization of mechanically instructive and sufficiently resilient materials (with appropriate stiffness, elasticity and surface topology) (Zhang et al., 2012; Li et al., 2024; Huang et al., 2023).
Another important aspect for studying tendon mechanobiology in vivo is to take different species (horses, cow, pigs, sheep, rabbits, dogs, rats and mice) in account with respect to animal models with significant importance for translational applications. These differences manifest in tendon structure, cellularity, cellular composition, mechanical properties and mechanoresponses. Therefore, it is important to choose the appropriate animal model, closest reflecting human conditions and species to harvest tenocytes for tendon research (Burgio et al., 2022; Oreff et al., 2021).
A comprehensive review highlights that many animal models, particularly rodents, do not fully replicate human tendon anatomy and loading conditions (Hast et al., 2014; Aykora et al., 2025). Mechanical loading induced different cellular and molecular responses in animal compared to human tendon. Cyclic loading in rodent models may lead to different adaptations than observed in human tendons (Stanczak et al., 2024; Magnusson and Kjaer, 2019). Quadrupedal animals distribute mechanical loads differently than bipedal humans, affecting tendon adaptation and therefore, the healing process (O'Neill et al., 2022; Roberts et al., 1998). In this context, it should also be mentioned that, depending upon topography and type, not all tendons are the same in their mechanoresponse, even in the same species.
In our view, no 2D or 3D stretch protocol investigated in vitro can yet be directly transferred to in vivo conditions due to complexity of the in vivo mechanical environment (Wang et al., 2018a), the differences in mechanical tendon properties (Bossuyt et al., 2024), the variations in cellular responses, the limitations in replicating physiological conditions (Escriche-Escuder et al., 2022; Stirling and McEachan, 2023) and the challenges in measuring and applying appropriate loads. In other words, a large-scale study would be needed to demonstrate direct transferability. An interdisciplinary team has also been recommended to work in multidisciplinary collaboration of doctors, scientists and therapists to use primary patient cells for the 2D and 3D stretcher and to establish a treatment protocol for patients in parallel (Wang et al., 2018a). Furthermore, this should be carried out with humans and not with animal models, as there is only limited direct transferability. Due to the different localizations (Achilles tendon, rotator cuff tendons, cruciate ligament), a direct transfer from an in vitro stretching protocol to the rehabilitation protocol in humans cannot be carried out, so the authors did not want to make any statement within the present review. The current treatment protocols for patients after Achilles tendon ruptures are also very different, depending on whether surgery was performed or not and especially in terms of timing weight bearing, immobilization strategies and rehab milestones (Saxena et al., 2022; Cohen et al., 2023).
Further perspectives for targeted tendon repair by mechanical loading
Looking at the mechanomics of each tendon cell population promises a great potential for future discoveries into the mechanobiology of tendons and mechano-induced regeneration approaches. In addition, the large number of different stretching protocols, the wide range of different stretching devices and the high costs of 3D bioreactors mean that standardized parameters for biofabrication are still in demand. Further developments in the field of tendon reconstruction in the future will include 4D applications and direct in situ bioprinting using robotics (Patrawalla et al., 2024).
Conclusion
Tenocyte subpopulations, TSPCs, myofibroblasts, and MSCs provide a cellular basis for tendon tissue engineering due to their mechanosensitivity. Meanwhile, numerous 2D and 3D in vitro and in vivo studies have shown that mechanical stimulation is primarily necessary to maintain tendon homeostasis and to stimulate tissue formation. However, there are still challenges in translating findings from mechanical stimulation to clinical applications. Mechanocompetent and tenoconductive 3D biomaterials serving as a cell carrier with sufficient cell adhesion motifs could be used in conjunction with appropriate mechanical stimulation of these cells, as an implant to treat tendon defects. These tissue engineered constructs could accelerate tendon regeneration in the future. To achieve this vision, first, uniform and sophisticated 3D models are required, reflecting the complex loading conditions (cyclic stretch, pressure, through fiber torsion, fluid flow and shear forces) of the particular tendon being investigated. These models should also rebuild the 3D mechanotopographical niche of the respective investigated tendon with its resident cell populations. These tools will be helpful to elucidate the mechanomics of tendons in more detail. Nowadays, the emerging tendon on chip platforms have started to address many of these requirements. Furthermore, identification of pharmaceutically addressable mechanosensitive ion channels or receptors, as a target to accelerate tendon regeneration is needed. Altogether, such cumulative efforts could lead to major steps towards more efficient and satisfactory management of tendon pathologies.
Author contributions
CG: Conceptualization, Formal Analysis, Funding acquisition, Investigation, Methodology, Visualization, Writing – original draft, Writing – review and editing. GP: Formal Analysis, Writing – original draft, Writing – review and editing. HT: Writing – original draft, Writing – review and editing. DD: Conceptualization, Funding acquisition, Supervision, Writing – review and editing. GS-T: Conceptualization, Formal Analysis, Funding acquisition, Methodology, Project administration, Validation, Visualization, Writing – original draft, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The authors acknowledge the COST Action TENET (TEndon Regeneration NETwork) grant (Proposal Nr. CA22170) as well as Nürnberger Krankenversicherung and the Kerscher Foundation. DD acknowledges the BMBF Grant “CellWiTaL: Reproducible cell systems for drug research—transfer layer-free laser printing of highly specific single cells in three-dimensional cellular structures” Proposal Nr. 13N15874. “Good Fibration—Targeting Fibrosis to Enhance Regeneration” (&20102/F2400948-FPR; Land Salzburg/WISS2025) and the Bridge Project of the Paracelsus Medical University “Inflammatory response in diabetic tendons—a potential role of tenophages”.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be constructed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Aggouras, A. N., Stowe, E. J., Mlawer, S. J., and Connizzo, B. K. (2024). Aged tendons exhibit altered mechanisms of strain-dependent extracellular matrix remodeling. J. Biomech. Eng. 146, 071009. doi:10.1115/1.4065270
Ajalik, R. E., Linares, I., Alenchery, R. G., Zhang, V. Z., Wright, T. W., Miller, B. L., et al. (2025). Human tendon-on-a-chip for modeling the myofibroblast microenvironment in peritendinous fibrosis. Adv. Healthc. Mater 14, e2403116. doi:10.1002/adhm.202403116
Andarawis-Puri, N., and Flatow, E. L. (2018). Promoting effective tendon healing and remodeling. J. Orthop. Res. 36, 3115–3124. doi:10.1002/jor.24133
Ansorge, H. L., Meng, X., Zhang, G., Veit, G., Sun, M., Klement, J. F., et al. (2009). Type XIV collagen regulates fibrillogenesis: premature collagen fibril growth and tissue dysfunction in null mice. J. Biol. Chem. 284, 8427–8438. doi:10.1074/jbc.M805582200
Arnesen, S. M., and Lawson, M. A. (2006). Age-related changes in focal adhesions lead to altered cell behavior in tendon fibroblasts. Mech. Ageing Dev. 127, 726–732. doi:10.1016/j.mad.2006.05.003
Awad, H., Ajalik, R., Alenchery, R., Linares, I., Wright, T., Miller, B., et al. (2025). Human tendon-on-a-chip for modeling vascular inflammatory fibrosis. Adv Healthc Mater. 14 (4), e2403116. doi:10.1002/adhm.202403116
Awad, H. A., Butler, D. L., Harris, M. T., Ibrahim, R. E., Wu, Y., Young, R. G., et al. (2000). In vitro characterization of mesenchymal stem cell-seeded collagen scaffolds for tendon repair: effects of initial seeding density on contraction kinetics. J. Biomed. Mater Res. 51, 233–240. doi:10.1002/(sici)1097-4636(200008)51:2<233::aid-jbm12>3.0.co;2-b
Aykora, D., Tasci, B., Sahin, M. Z., Tekeoglu, I., Uzun, M., Sarafian, V., et al. (2025). Tendon regeneration deserves better: focused review on in vivo models, artificial intelligence and 3D bioprinting approaches. Front. Bioeng. Biotechnol. 13, 1580490. doi:10.3389/fbioe.2025.1580490
Babaniamansour, P., Jacho, D., Rabino, A., Garcia-Mata, R., and Yildirim-Ayan, E. (2024). Synergetic role of TRPV4 inhibitor and mechanical loading on reducing inflammation. Front. Immunol. 15, 1456042. doi:10.3389/fimmu.2024.1456042
Backman, L. J., Fong, G., Andersson, G., Scott, A., and Danielson, P. (2011). Substance P is a mechanoresponsive, autocrine regulator of human tenocyte proliferation. PLoS One 6, e27209. doi:10.1371/journal.pone.0027209
Bakht, S. M., Pardo, A., Gomez-Florit, M., Caballero, D., Kundu, S. C., Reis, R. L., et al. (2024). Human tendon-on-chip: unveiling the effect of core compartment-T cell spatiotemporal crosstalk at the onset of tendon inflammation. Adv. Sci. (Weinh) 11, e2401170. doi:10.1002/advs.202401170
Bian, X., Liu, X., Zhou, M., Tang, H., Wang, R., Ma, L., et al. (2024). Mechanical stimulation promotes fibrochondrocyte proliferation by activating the TRPV4 signaling pathway during tendon-bone insertion healing: CCN2 plays an important regulatory role. Burns Trauma 12, tkae028. doi:10.1093/burnst/tkae028
Bjur, D., Alfredson, H., and Forsgren, S. (2009). Presence of the neuropeptide Y1 receptor in tenocytes and blood vessel walls in the human Achilles tendon. Br. J. Sports Med. 43, 1136–1142. doi:10.1136/bjsm.2008.055780
Blache, U., Wunderli, S. L., Hussien, A. A., Stauber, T., Fluckiger, G., Bollhalder, M., et al. (2021). Inhibition of ERK 1/2 kinases prevents tendon matrix breakdown. Sci. Rep. 11, 6838. doi:10.1038/s41598-021-85331-1
Bossuyt, F. M., Leonard, T. R., Scott, W. M., Taylor, W. R., and Herzog, W. (2024). In-vivo and in-vitro environments affect the storage and release of energy in tendons. Front. Physiol. 15, 1443675. doi:10.3389/fphys.2024.1443675
Bs, A., Ganesh, S. B., Eswaramoorthy, R., and G, K. (2024). Hyaluronic acid hydrogel combined with Ocimum sanctum and Tendon-derived extracellular matrix enhances tenogenesis in periodontal ligament stem cells. Cureus 16, e54060. doi:10.7759/cureus.54060
Burgio, V., Civera, M., Rodriguez Reinoso, M., Pizzolante, E., Prezioso, S., Bertuglia, A., et al. (2022). Mechanical properties of animal tendons: a review and comparative study for the identification of the most suitable human tendon surrogates. Processes 10, 485. doi:10.3390/pr10030485
Burk, J., Plenge, A., Brehm, W., Heller, S., Pfeiffer, B., and Kasper, C. (2016). Induction of tenogenic differentiation mediated by extracellular Tendon matrix and short-term cyclic stretching. Stem Cells Int. 2016, 7342379. doi:10.1155/2016/7342379
Carroll, C. C., Dickinson, J. M., Haus, J. M., Lee, G. A., Hollon, C. J., Aagaard, P., et al. (2008). Influence of aging on the in vivo properties of human patellar tendon. J. Appl. Physiol. 105, 1907–1915. doi:10.1152/japplphysiol.00059.2008
Cenni, V., Sabatelli, P., Di Martino, A., Merlini, L., Antoniel, M., Squarzoni, S., et al. (2024). Collagen VI deficiency impairs tendon fibroblasts mechanoresponse in ullrich congenital muscular dystrophy. Cells 13, 378. doi:10.3390/cells13050378
Chamberlain, C. S., Crowley, E. M., Kobayashi, H., Eliceiri, K. W., and Vanderby, R. (2011). Quantification of collagen organization and extracellular matrix factors within the healing ligament. Microsc. Microanal. 17, 779–787. doi:10.1017/S1431927611011925
Chen, H., Chen, L., Cheng, B., and Jiang, C. (2015a). Cyclic mechanical stretching induces autophagic cell death in tenofibroblasts through activation of prostaglandin E2 production. Cell Physiol. Biochem. 36, 24–33. doi:10.1159/000374050
Chen, Q., Zhou, J., Zhang, B., Chen, Z., Luo, Q., and Song, G. (2018). Cyclic stretching exacerbates tendinitis by enhancing NLRP3 inflammasome activity via F-Actin depolymerization. Inflammation 41, 1731–1743. doi:10.1007/s10753-018-0816-5
Chen, W., Deng, Y., Zhang, J., and Tang, K. (2015b). Uniaxial repetitive mechanical overloading induces influx of extracellular calcium and cytoskeleton disruption in human tenocytes. Cell Tissue Res. 359, 577–587. doi:10.1007/s00441-014-2018-2
Chen, Y., Zhang, T., Wan, L., Wang, Z., Li, S., Hu, J., et al. (2021). Early treadmill running delays rotator cuff healing via neuropeptide Y mediated inactivation of the Wnt/β-catenin signaling. J. Orthop. Transl. 30, 103–111. doi:10.1016/j.jot.2021.08.004
Chen, Z., Chen, P., Ruan, R., and Zheng, M. (2022). In vitro 3D mechanical stimulation to tendon-derived stem cells by bioreactor. Methods Mol. Biol. 2436, 135–144. doi:10.1007/7651_2021_432
Chen, Z., Jin, M., He, H., Dong, J., Li, J., Nie, J., et al. (2023). Mesenchymal stem cells and macrophages and their interactions in tendon-bone healing. J. Orthop. Transl. 39, 63–73. doi:10.1016/j.jot.2022.12.005
Chen, Z., Li, M., Chen, P., Tai, A., Li, J., Bassonga, E. L., et al. (2024). Mechanical overload-induced release of extracellular mitochondrial particles from tendon cells leads to inflammation in tendinopathy. Exp. Mol. Med. 56, 583–599. doi:10.1038/s12276-024-01183-5
Chernak Slane, L., and Thelen, D. G. (2014). The use of 2D ultrasound elastography for measuring tendon motion and strain. J. Biomech. 47, 750–754. doi:10.1016/j.jbiomech.2013.11.023
Cohen, D., Sandman, E., Saran, N., Petrisor, B., Bhandari, M., Veljkovic, A., et al. (2023). Evidence-based treatment of achilles tendon rupture. Can. J. Surg. 66, E356–E357. doi:10.1503/cjs.008321
Constantinou, I., and Bastounis, E. E. (2023). Cell-stretching devices: advances and challenges in biomedical research and live-cell imaging. Trends Biotechnol. 41, 939–950. doi:10.1016/j.tibtech.2022.12.009
Delgado Caceres, M., Angerpointner, K., Galler, M., Lin, D., Michel, P. A., Brochhausen, C., et al. (2021). Tenomodulin knockout mice exhibit worse late healing outcomes with augmented trauma-induced heterotopic ossification of achilles tendon. Cell Death Dis. 12, 1049. doi:10.1038/s41419-021-04298-z
De Micheli, A. J., Swanson, J. B., Disser, N. P., Martinez, L. M., Walker, N. R., Oliver, D. J., et al. (2020). Single-cell transcriptomic analysis identifies extensive heterogeneity in the cellular composition of mouse achilles tendons. Am. J. Physiol. Cell Physiol. 319, C885–C894. doi:10.1152/ajpcell.00372.2020
Dex, S., Alberton, P., Willkomm, L., Sollradl, T., Bago, S., Milz, S., et al. (2017). Tenomodulin is required for Tendon endurance running and collagen I fibril adaptation to mechanical load. EBioMedicine 20, 240–254. doi:10.1016/j.ebiom.2017.05.003
Dex, S., Lin, D., Shukunami, C., and Docheva, D. (2016). Tenogenic modulating insider factor: systematic assessment on the functions of tenomodulin gene. Gene 587, 1–17. doi:10.1016/j.gene.2016.04.051
Docheva, D., Hunziker, E. B., Fassler, R., and Brandau, O. (2005). Tenomodulin is necessary for tenocyte proliferation and tendon maturation. Mol. Cell Biol. 25, 699–705. doi:10.1128/MCB.25.2.699-705.2005
Dudhia, J., Scott, C. M., Draper, E. R., Heinegard, D., Pitsillides, A. A., and Smith, R. K. (2007). Aging enhances a mechanically-induced reduction in tendon strength by an active process involving matrix metalloproteinase activity. Aging Cell 6, 547–556. doi:10.1111/j.1474-9726.2007.00307.x
Durgam, S., Singh, B., Cole, S. L., Brokken, M. T., and Stewart, M. (2020). Quantitative assessment of Tendon hierarchical structure by combined second harmonic generation and immunofluorescence microscopy. Tissue Eng. Part C Methods 26, 253–262. doi:10.1089/ten.TEC.2020.0032
Eekhoff, J. D., Steenbock, H., Berke, I. M., Brinckmann, J., Yanagisawa, H., Wagenseil, J. E., et al. (2021). Dysregulated assembly of elastic fibers in fibulin-5 knockout mice results in a tendon-specific increase in elastic modulus. J. Mech. Behav. Biomed. Mater 113, 104134. doi:10.1016/j.jmbbm.2020.104134
Egerbacher, M., Gabner, S., Battisti, S., and Handschuh, S. (2020). Tenocytes form a 3-D network and are connected via nanotubes. J. Anat. 236, 165–170. doi:10.1111/joa.13089
Eliasson, P., Andersson, T., and Aspenberg, P. (2009). Rat Achilles tendon healing: mechanical loading and gene expression. J. Appl. Physiol. 107, 399–407. doi:10.1152/japplphysiol.91563.2008
Epro, G., Mierau, A., Doerner, J., Luetkens, J. A., Scheef, L., Kukuk, G. M., et al. (2017). The Achilles tendon is mechanosensitive in older adults: adaptations following 14 weeks versus 1.5 years of cyclic strain exercise. J. Exp. Biol. 220, 1008–1018. doi:10.1242/jeb.146407
Eriksen, H. A., Pajala, A., Leppilahti, J., and Risteli, J. (2002). Increased content of type III collagen at the rupture site of human Achilles tendon. J. Orthop. Res. 20, 1352–1357. doi:10.1016/S0736-0266(02)00064-5
Escriche-Escuder, A., Cuesta-Vargas, A. I., and Casana, J. (2022). Modelling and in vivo evaluation of tendon forces and strain in dynamic rehabilitation exercises: a scoping review. BMJ Open 12, e057605. doi:10.1136/bmjopen-2021-057605
Fang, F., Schwartz, A. G., Moore, E. R., Sup, M. E., and Thomopoulos, S. (2020). Primary cilia as the nexus of biophysical and hedgehog signaling at the tendon enthesis. Sci. Adv. 6, eabc1799. doi:10.1126/sciadv.abc1799
Fleischmann, N., Hofmann, S., Gögele, C., Frank, E., Werner, C., Kokozidou, M., et al. (2024). Achilles tenocytes from diabetic and non diabetic donors exposed to hyperglycemia respond differentially to inflammatory stimuli and stretch. J. Anat. doi:10.1111/joa.14207
Frank, C., Mcdonald, D., Bray, D., Bray, R., Rangayyan, R., Chimich, D., et al. (1992). Collagen fibril diameters in the healing adult rabbit medial collateral ligament. Connect. Tissue Res. 27, 251–263. doi:10.3109/03008209209007000
Frank, C., Mcdonald, D., Wilson, J., Eyre, D., and Shrive, N. (1995). Rabbit medial collateral ligament scar weakness is associated with decreased collagen pyridinoline crosslink density. J. Orthop. Res. 13, 157–165. doi:10.1002/jor.1100130203
Freedman, B. R., Rodriguez, A. B., Leiphart, R. J., Newton, J. B., Ban, E., Sarver, J. J., et al. (2018). Dynamic loading and tendon healing affect multiscale tendon properties and ECM stress transmission. Sci. Rep. 8, 10854. doi:10.1038/s41598-018-29060-y
Gehwolf, R., Spitzer, G., Wagner, A., Lehner, C., Weissenbacher, N., Tempfer, H., et al. (2019). 3D-Embedded cell cultures to study tendon biology. Methods Mol. Biol. 2045, 155–165. doi:10.1007/7651_2019_208
Geng, Y., Zhao, X., Xu, J., Zhang, X., Hu, G., Fu, S. C., et al. (2020). Overexpression of mechanical sensitive miR-337-3p alleviates ectopic ossification in rat tendinopathy model via targeting IRS1 and Nox4 of tendon-derived stem cells. J. Mol. Cell Biol. 12, 305–317. doi:10.1093/jmcb/mjz030
Giacomini, F., Rho, H. S., Eischen-Loges, M., Tahmasebi Birgani, Z., Van Blitterswijk, C., Van Griensven, M., et al. (2024). Enthesitis on chip - a model for studying acute and chronic inflammation of the enthesis and its pharmacological treatment. Adv. Healthc. Mater 13, e2401815. doi:10.1002/adhm.202401815
Gögele, C., Hoffmann, C., Konrad, J., Merkel, R., Schwarz, S., Tohidnezhad, M., et al. (2021a). Cyclically stretched ACL fibroblasts emigrating from spheroids adapt their cytoskeleton and ligament-related expression profile. Cell Tissue Res. 384, 675–690. doi:10.1007/s00441-021-03416-9
Gögele, C., Konrad, J., Hahn, J., Breier, A., Schroepfer, M., Meyer, M., et al. (2021b). Maintenance of ligament homeostasis of spheroid-colonized embroidered and functionalized scaffolds after 3D stretch. Int. J. Mol. Sci. 22, 8204. doi:10.3390/ijms22158204
Goh, K. L., Holmes, D. F., Lu, Y. H., Kadler, K. E., and Purslow, P. P. (2018). Age-related dataset on the mechanical properties and collagen fibril structure of tendons from a murine model. Sci. Data 5, 180140. doi:10.1038/sdata.2018.140
Grier, W. G., Moy, A. S., and Harley, B. A. (2017). Cyclic tensile strain enhances human mesenchymal stem cell smad 2/3 activation and tenogenic differentiation in anisotropic collagen-glycosaminoglycan scaffolds. Eur. Cell Mater 33, 227–239. doi:10.22203/eCM.v033a14
Grote, C., Reinhardt, D., Zhang, M., and Wang, J. (2019). Regulatory mechanisms and clinical manifestations of musculoskeletal aging. J. Orthop. Res. 37, 1475–1488. doi:10.1002/jor.24292
Gumucio, J. P., Schonk, M. M., Kharaz, Y. A., Comerford, E., and Mendias, C. L. (2020). Scleraxis is required for the growth of adult tendons in response to mechanical loading. JCI Insight 5, e138295. doi:10.1172/jci.insight.138295
Hahn, J., Schulze-Tanzil, G., Schröpfer, M., Meyer, M., Gögele, C., Hoyer, M., et al. (2019). Viscoelastic behavior of embroidered scaffolds for ACL tissue engineering made of PLA and P(LA-CL) after in vitro degradation. Int. J. Mol. Sci. 20, 4655. doi:10.3390/ijms20184655
Hammerman, M., Blomgran, P., Dansac, A., Eliasson, P., and Aspenberg, P. (2017). Different gene response to mechanical loading during early and late phases of rat achilles tendon healing. J. Appl. Physiol. 123, 800–815. doi:10.1152/japplphysiol.00323.2017
Hammerman, M., Dietrich-Zagonel, F., Blomgran, P., Eliasson, P., and Aspenberg, P. (2018). Different mechanisms activated by mild versus strong loading in rat Achilles tendon healing. PLoS One 13, e0201211. doi:10.1371/journal.pone.0201211
Hammerman, M., Pierantoni, M., Isaksson, H., and Eliasson, P. (2024). Deprivation of loading during rat Achilles tendon healing affects extracellular matrix composition and structure, and reduces cell density and alignment. Sci. Rep. 14, 23380. doi:10.1038/s41598-024-74783-w
Hast, M. W., Zuskov, A., and Soslowsky, L. J. (2014). The role of animal models in tendon research. Bone Jt. Res. 3, 193–202. doi:10.1302/2046-3758.36.2000281
Hatta, T., Sano, H., Sakamoto, N., Kishimoto, K. N., Sato, M., and Itoi, E. (2013). Nicotine reduced MMP-9 expression in the primary porcine tenocytes exposed to cyclic stretch. J. Orthop. Res. 31, 645–650. doi:10.1002/jor.22259
Herchenhan, A., Dietrich-Zagonel, F., Schjerling, P., Kjaer, M., and Eliasson, P. (2020). Early growth response genes increases rapidly after mechanical overloading and unloading in Tendon constructs. J. Orthop. Res. 38, 173–181. doi:10.1002/jor.24513
Huang, L., Chen, L., Chen, H., Wang, M., Jin, L., Zhou, S., et al. (2023). Biomimetic scaffolds for Tendon tissue regeneration. Biomimetics (Basel) 8, 246. doi:10.3390/biomimetics8020246
Huang, Y. T., Wu, Y. F., Wang, H. K., Yao, C. J., Chiu, Y. H., Sun, J. S., et al. (2022). Cyclic mechanical stretch regulates the AMPK/Egr1 pathway in tenocytes via Ca2+-mediated mechanosensing. Connect. Tissue Res. 63, 590–602. doi:10.1080/03008207.2022.2044321
Huisman, E., Lu, A., Mccormack, R. G., and Scott, A. (2014). Enhanced collagen type I synthesis by human tenocytes subjected to periodic in vitro mechanical stimulation. BMC Musculoskelet. Disord. 15, 386. doi:10.1186/1471-2474-15-386
Iyyathurai, J., Himpens, B., Bultynck, G., and D’hondt, C. (2016). Calcium wave propagation triggered by local mechanical stimulation as a method for studying gap junctions and hemichannels. Methods Mol. Biol. 1437, 203–211. doi:10.1007/978-1-4939-3664-9_15
Izu, Y., Adams, S. M., Connizzo, B. K., Beason, D. P., Soslowsky, L. J., Koch, M., et al. (2021). Collagen XII mediated cellular and extracellular mechanisms regulate establishment of tendon structure and function. Matrix Biol. 95, 52–67. doi:10.1016/j.matbio.2020.10.004
Jiang, X., Wu, S., Kuss, M., Kong, Y., Shi, W., Streubel, P. N., et al. (2020). 3D printing of multilayered scaffolds for rotator cuff tendon regeneration. Bioact. Mater 5, 636–643. doi:10.1016/j.bioactmat.2020.04.017
Juncosa, N., West, J. R., Galloway, M. T., Boivin, G. P., and Butler, D. L. (2003). In vivo forces used to develop design parameters for tissue engineered implants for rabbit patellar tendon repair. J. Biomech. 36, 483–488. doi:10.1016/s0021-9290(02)00459-1
Kamble, H., Vadivelu, R., Barton, M., Boriachek, K., Munaz, A., Park, S., et al. (2017). An electromagnetically actuated double-sided cell-stretching device for mechanobiology research. Micromachines (Basel) 8, 256. doi:10.3390/mi8080256
Kaneda, G., Chan, J. L., Castaneda, C. M., Papalamprou, A., Sheyn, J., Shelest, O., et al. (2023). iPSC-derived tenocytes seeded on microgrooved 3D printed scaffolds for Achilles tendon regeneration. J. Orthop. Res. 41, 2205–2220. doi:10.1002/jor.25554
Kannus, P. (2000). Structure of the tendon connective tissue. Scand. J. Med. Sci. Sports 10, 312–320. doi:10.1034/j.1600-0838.2000.010006312.x
Kayama, T., Mori, M., Ito, Y., Matsushima, T., Nakamichi, R., Suzuki, H., et al. (2016). Gtf2ird1-dependent Mohawk expression regulates mechanosensing properties of the Tendon. Mol. Cell Biol. 36, 1297–1309. doi:10.1128/MCB.00950-15
Kendal, A. R., Layton, T., Al-Mossawi, H., Appleton, L., Dakin, S., Brown, R., et al. (2020). Multi-omic single cell analysis resolves novel stromal cell populations in healthy and diseased human tendon. Sci. Rep. 10, 13939. doi:10.1038/s41598-020-70786-5
Khayyeri, H., Barreto, S., and Lacroix, D. (2015). Primary cilia mechanics affects cell mechanosensation: a computational study. J. Theor. Biol. 379, 38–46. doi:10.1016/j.jtbi.2015.04.034
Killian, M. L. (2022). Growth and mechanobiology of the tendon-bone enthesis. Semin. Cell Dev. Biol. 123, 64–73. doi:10.1016/j.semcdb.2021.07.015
Korcari, A., Przybelski, S. J., Gingery, A., and Loiselle, A. E. (2023). Impact of aging on tendon homeostasis, tendinopathy development, and impaired healing. Connect. Tissue Res. 64, 1–13. doi:10.1080/03008207.2022.2102004
Kubo, Y., Hoffmann, B., Goltz, K., Schnakenberg, U., Jahr, H., Merkel, R., et al. (2020). Different frequency of cyclic Tensile strain relates to Anabolic/Catabolic conditions consistent with immunohistochemical staining intensity in tenocytes. Int. J. Mol. Sci. 21, 1082. doi:10.3390/ijms21031082
Kwan, K. Y. C., Ng, K. W. K., Rao, Y., Zhu, C., Qi, S., Tuan, R. S., et al. (2023). Effect of aging on Tendon biology, biomechanics and implications for treatment approaches. Int. J. Mol. Sci. 24, 15183. doi:10.3390/ijms242015183
Lammerding, J., Schulze, P. C., Takahashi, T., Kozlov, S., Sullivan, T., Kamm, R. D., et al. (2004). Lamin A/C deficiency causes defective nuclear mechanics and mechanotransduction. J. Clin. Invest. 113, 370–378. doi:10.1172/JCI19670
Lavagnino, M., Arnoczky, S. P., and Gardner, K. (2011). In situ deflection of tendon cell-cilia in response to tensile loading: an in vitro study. J. Orthop. Res. 29, 925–930. doi:10.1002/jor.21337
Lavagnino, M., Wall, M. E., Little, D., Banes, A. J., Guilak, F., and Arnoczky, S. P. (2015). Tendon mechanobiology: current knowledge and future research opportunities. J. Orthop. Res. 33, 813–822. doi:10.1002/jor.22871
Leahy, T. P., Chenna, S. S., Soslowsky, L. J., and Dyment, N. A. (2024). Focal adhesion kinase regulates tendon cell mechanoresponse and physiological tendon development. FASEB J. 38, e70050. doi:10.1096/fj.202400151R
Leahy, T. P., Fung, A. K., Weiss, S. N., Dyment, N. A., and Soslowsky, L. J. (2023). Investigating the temporal roles of decorin and biglycan in tendon healing. J. Orthop. Res. 41, 2238–2249. doi:10.1002/jor.25590
Leal-Egana, A., Balland, M., and Boccaccini, A. R. (2020). Re-engineering artificial neoplastic milieus: taking lessons from Mechano- and topobiology. Trends Biotechnol. 38, 142–153. doi:10.1016/j.tibtech.2019.08.003
Legerlotz, K., Jones, G. C., Screen, H. R., and Riley, G. P. (2013). Cyclic loading of tendon fascicles using a novel fatigue loading system increases interleukin-6 expression by tenocytes. Scand. J. Med. Sci. Sports 23, 31–37. doi:10.1111/j.1600-0838.2011.01410.x
Lehner, C., Spitzer, G., Gehwolf, R., Wagner, A., Weissenbacher, N., Deininger, C., et al. (2019). Tenophages: a novel macrophage-like tendon cell population expressing CX3CL1 and CX3CR1. Dis. Model Mech. 12, dmm041384. doi:10.1242/dmm.041384
Lejard, V., Blais, F., Guerquin, M. J., Bonnet, A., Bonnin, M. A., Havis, E., et al. (2011). EGR1 and EGR2 involvement in vertebrate tendon differentiation. J. Biol. Chem. 286, 5855–5867. doi:10.1074/jbc.M110.153106
Li, H., Korcari, A., Ciufo, D., Mendias, C. L., Rodeo, S. A., Buckley, M. R., et al. (2023a). Increased Ca(2+) signaling through Ca(V) 1.2 induces tendon hypertrophy with increased collagen fibrillogenesis and biomechanical properties. FASEB J. 37, e23007. doi:10.1096/fj.202300607R
Li, S., Sun, Y., Chen, Y., Lu, J., Jiang, G., Yu, K., et al. (2023b). Sandwich biomimetic scaffold based Tendon stem/progenitor cell alignment in a 3D microenvironment for functional tendon regeneration. ACS Appl. Mater Interfaces 15, 4652–4667. doi:10.1021/acsami.2c16584
Li, Y., Ge, Z., Liu, Z., Li, L., Song, J., Wang, H., et al. (2024). Integrating electrospun aligned fiber scaffolds with bovine serum albumin-basic fibroblast growth factor nanoparticles to promote tendon regeneration. J. Nanobiotechnol. 22, 799. doi:10.1186/s12951-024-03022-1
Linares, I., Chen, K., Saffren, A., Mansouri, M., Abhyankar, V. V., Miller, B. L., et al. (2025). Fluid flow impacts endothelial-monocyte interactions in a model of vascular inflammatory fibrosis. Sci. Rep. 15, 3227. doi:10.1038/s41598-025-85987-z
Liu, Z., Han, W., Meng, J., Pi, Y., Wu, T., Fan, Y., et al. (2024). Mohawk protects against tendon damage via suppressing Wnt/β-catenin pathway. Heliyon 10, e25658. doi:10.1016/j.heliyon.2024.e25658
Lohberger, B., Kaltenegger, H., Stuendl, N., Rinner, B., Leithner, A., and Sadoghi, P. (2016). Impact of cyclic mechanical stimulation on the expression of extracellular matrix proteins in human primary rotator cuff fibroblasts. Knee Surg. Sports Traumatol. Arthrosc. 24, 3884–3891. doi:10.1007/s00167-015-3790-6
Lu, K., Tang, H., Wang, Y., Wang, L., Zhou, M., He, G., et al. (2023). Micropattern silk fibroin film facilitates Tendon repair in vivo and promotes tenogenic differentiation of Tendon stem/progenitor cells through the alpha2beta1/FAK/PI3K/AKT signaling pathway in vitro. Stem Cells Int. 2023, 2915826. doi:10.1155/2023/2915826
Lui, P. P. Y., and Wong, C. M. (2019). Biology of tendon stem cells and tendon in aging. Front. Genet. 10, 1338. doi:10.3389/fgene.2019.01338
Maeda, E., Hagiwara, Y., Wang, J. H., and Ohashi, T. (2013). A new experimental system for simultaneous application of cyclic tensile strain and fluid shear stress to tenocytes in vitro. Biomed. Microdevices 15, 1067–1075. doi:10.1007/s10544-013-9798-0
Maeda, E., Pian, H., and Ohashi, T. (2017). Temporal regulation of gap junctional communication between tenocytes subjected to static tensile strain with physiological and non-physiological amplitudes. Biochem. Biophys. Res. Commun. 482, 1170–1175. doi:10.1016/j.bbrc.2016.12.007
Maeda, T., Sakabe, T., Sunaga, A., Sakai, K., Rivera, A. L., Keene, D. R., et al. (2011). Conversion of mechanical force into TGF-beta-mediated biochemical signals. Curr. Biol. 21, 933–941. doi:10.1016/j.cub.2011.04.007
Maffulli, N., Moller, H., and Evans, C. (2002). Tendon healing: can it be optimised? Br. J. sports Med. 36, 315–316. doi:10.1136/bjsm.36.5.315
Magnusson, S. P., and Kjaer, M. (2019). The impact of loading, unloading, ageing and injury on the human tendon. J. Physiol. 597, 1283–1298. doi:10.1113/JP275450
Magra, M., Hughes, S., El Haj, A. J., and Maffulli, N. (2007). VOCCs and TREK-1 ion channel expression in human tenocytes. Am. J. Physiol. Cell Physiol. 292, C1053–C1060. doi:10.1152/ajpcell.00053.2006
Maharam, E., Yaport, M., Villanueva, N. L., Akinyibi, T., Laudier, D., He, Z., et al. (2015). Rho/rock signal transduction pathway is required for MSC tenogenic differentiation. Bone Res. 3, 15015. doi:10.1038/boneres.2015.15
Manning, C. N., Havlioglu, N., Knutsen, E., Sakiyama-Elbert, S. E., Silva, M. J., Thomopoulos, S., et al. (2014). The early inflammatory response after flexor tendon healing: a gene expression and histological analysis. J. Orthop. Res. 32, 645–652. doi:10.1002/jor.22575
Marr, N., Meeson, R., Kelly, E. F., Fang, Y., Peffers, M. J., Pitsillides, A. A., et al. (2021). CD146 delineates an interfascicular cell sub-population in tendon that is recruited during injury through its ligand Laminin-α4. Int. J. Mol. Sci. 22, 9729. doi:10.3390/ijms22189729
Mccarthy, M. M., and Hannafin, J. A. (2014). The mature athlete: aging tendon and ligament. Sports Health 6, 41–48. doi:10.1177/1941738113485691
Mccrum, C., Leow, P., Epro, G., Konig, M., Meijer, K., and Karamanidis, K. (2018). Alterations in leg extensor muscle-tendon unit biomechanical properties with ageing and mechanical loading. Front. Physiol. 9, 150. doi:10.3389/fphys.2018.00150
Melzer, M., Niebert, S., Heimann, M., Ullm, F., Pompe, T., Scheiner-Bobis, G., et al. (2024). Differential Smad2/3 linker phosphorylation is a crosstalk mechanism of Rho/ROCK and canonical TGF-β3 signaling in tenogenic differentiation. Sci. Rep. 14, 10393. doi:10.1038/s41598-024-60717-z
Melzer, M., Schubert, S., Muller, S. F., Geyer, J., Hagen, A., Niebert, S., et al. (2021). Rho/ROCK inhibition promotes TGF-beta3-induced tenogenic differentiation in mesenchymal stromal cells. Stem Cells Int. 2021, 8284690. doi:10.1155/2021/8284690
Miescher, I., Schaffner, N., Rieber, J., Burgisser, G. M., Ongini, E., Yang, Y., et al. (2024). Hyaluronic acid/PEO electrospun tube reduces tendon adhesion to levels comparable to native tendons - an in vitro and in vivo study. Int. J. Biol. Macromol. 273, 133193. doi:10.1016/j.ijbiomac.2024.133193
Millward-Sadler, S. J., and Salter, D. M. (2004). Integrin-dependent signal cascades in chondrocyte mechanotransduction. Ann. Biomed. Eng. 32, 435–446. doi:10.1023/b:abme.0000017538.72511.48
Mlyniec, A., Dabrowska, S., Heljak, M., Weglarz, W. P., Wojcik, K., Ekiert-Radecka, M., et al. (2021). The dispersion of viscoelastic properties of fascicle bundles within the tendon results from the presence of interfascicular matrix and flow of body fluids. Mater Sci. Eng. C Mater Biol. Appl. 130, 112435. doi:10.1016/j.msec.2021.112435
Molloy, T., Wang, Y., and Murrell, G. (2003). The roles of growth factors in tendon and ligament healing. Sports Med. 33, 381–394. doi:10.2165/00007256-200333050-00004
Monteiro, R. F., Bakht, S. M., Gomez-Florit, M., Stievani, F. C., Alves, A. L. G., Reis, R. L., et al. (2023). Writing 3D in vitro models of human Tendon within a biomimetic fibrillar support platform. ACS Appl. Mater Interfaces 15, 50598–50611. doi:10.1021/acsami.2c22371
Morita, Y., Sato, T., Higashiura, K., Hirano, Y., Matsubara, F., Oshima, K., et al. (2019). The optimal mechanical condition in stem cell-to-tenocyte differentiation determined with the homogeneous strain distributions and the cellular orientation control. Biol. Open 8, bio039164. doi:10.1242/bio.039164
Mousavizadeh, R., Hojabrpour, P., Eltit, F., Mcdonald, P. C., Dedhar, S., Mccormack, R. G., et al. (2020). β1 integrin, ILK and mTOR regulate collagen synthesis in mechanically loaded tendon cells. Sci. Rep. 10, 12644. doi:10.1038/s41598-020-69267-6
Mousavizadeh, R., Waugh, C. M., Mccormack, R. G., Cairns, B. E., and Scott, A. (2024). MRGPRX2-mediated mast cell activation by substance P from overloaded human tenocytes induces inflammatory and degenerative responses in tendons. Sci. Rep. 14, 13540. doi:10.1038/s41598-024-64222-1
Nakamichi, R., Ma, S., Nonoyama, T., Chiba, T., Kurimoto, R., Ohzono, H., et al. (2022). The mechanosensitive ion channel PIEZO1 is expressed in tendons and regulates physical performance. Sci. Transl. Med. 14, eabj5557. doi:10.1126/scitranslmed.abj5557
Nam, H. Y., Balaji Raghavendran, H. R., Pingguan-Murphy, B., Abbas, A. A., Merican, A. M., and Kamarul, T. (2017). Fate of tenogenic differentiation potential of human bone marrow stromal cells by uniaxial stretching affected by stretch-activated calcium channel agonist gadolinium. PLoS One 12, e0178117. doi:10.1371/journal.pone.0178117
Nam, H. Y., Pingguan-Murphy, B., Abbas, A. A., Merican, A. M., and Kamarul, T. (2019). Uniaxial cyclic tensile stretching at 8% strain exclusively promotes tenogenic differentiation of human bone marrow-derived mesenchymal stromal cells. Stem Cells Int. 2019, 1–16. doi:10.1155/2019/9723025
Nichols, A. E. C., Best, K. T., and Loiselle, A. E. (2019). The cellular basis of fibrotic tendon healing: challenges and opportunities. Transl. Res. 209, 156–168. doi:10.1016/j.trsl.2019.02.002
Nichols, A. E. C., Settlage, R. E., Werre, S. R., and Dahlgren, L. A. (2018). Novel roles for scleraxis in regulating adult tenocyte function. BMC Cell Biol. 19, 14. doi:10.1186/s12860-018-0166-z
Nie, D., Zhou, Y., Wang, W., Zhang, J., and Wang, J. H. (2021). Mechanical overloading induced-activation of mTOR signaling in tendon stem/progenitor cells contributes to tendinopathy development. Front. Cell Dev. Biol. 9, 687856. doi:10.3389/fcell.2021.687856
Notermans, T., Khayyeri, H., and Isaksson, H. (2019). Understanding how reduced loading affects achilles tendon mechanical properties using a fibre-reinforced poro-visco-hyper-elastic model. J. Mech. Behav. Biomed. Mater 96, 301–309. doi:10.1016/j.jmbbm.2019.04.041
Okech, W., and Kuo, C. K. (2016). Informing stem cell-based tendon tissue engineering approaches with embryonic tendon development. Adv. Exp. Med. Biol. 920, 63–77. doi:10.1007/978-3-319-33943-6_6
O’neill, M. C., Demes, B., Thompson, N. E., Larson, S. G., Stern, J. T., and Umberger, B. R. (2022). Adaptations for bipedal walking: musculoskeletal structure and three-dimensional joint mechanics of humans and bipedal chimpanzees (pan troglodytes). J. Hum. Evol. 168, 103195. doi:10.1016/j.jhevol.2022.103195
Oreff, G. L., Fenu, M., Vogl, C., Ribitsch, I., and Jenner, F. (2021). Species variations in tenocytes’ response to inflammation require careful selection of animal models for tendon research. Sci. Rep. 11, 12451. doi:10.1038/s41598-021-91914-9
Pajala, A., Melkko, J., Leppilahti, J., Ohtonen, P., Soini, Y., and Risteli, J. (2009). Tenascin-C and type I and III collagen expression in total achilles tendon rupture. An immunohistochemical study. Histol. Histopathol. 24, 1207–1211. doi:10.14670/HH-24.1207
Pancheri, N. M., Daw, J. T., Ditton, D., Schiele, N. R., Birks, S., Uzer, G., et al. (2023). The LINC complex regulates achilles tendon elastic Modulus, achilles and tail tendon collagen crimp, and Achilles and tail tendon lateral expansion during early postnatal development. J. Orthop. Res. 43 (6), 1090–1100. doi:10.1002/jor.26069
Papalamprou, A., Yu, V., Chen, A., Stefanovic, T., Kaneda, G., Salehi, K., et al. (2023). Directing iPSC differentiation into iTenocytes using combined scleraxis overexpression and cyclic loading. J. Orthop. Res. 41, 1148–1161. doi:10.1002/jor.25459
Passini, F. S., Jaeger, P. K., Saab, A. S., Hanlon, S., Chittim, N. A., Arlt, M. J., et al. (2021). Shear-stress sensing by PIEZO1 regulates tendon stiffness in rodents and influences jumping performance in humans. Nat. Biomed. Eng. 5, 1457–1471. doi:10.1038/s41551-021-00716-x
Patel, D., Sharma, S., Screen, H. R. C., and Bryant, S. J. (2018). Effects of cell adhesion motif, fiber stiffness, and cyclic strain on tenocyte gene expression in a tendon mimetic fiber composite hydrogel. Biochem. Biophys. Res. Commun. 499, 642–647. doi:10.1016/j.bbrc.2018.03.203
Patel, D., Zamboulis, D. E., Spiesz, E. M., Birch, H. L., Clegg, P. D., Thorpe, C. T., et al. (2021). Structure-function specialisation of the interfascicular matrix in the human achilles tendon. Acta Biomater. 131, 381–390. doi:10.1016/j.actbio.2021.07.019
Patrawalla, N. Y., Liebendorfer, K., and Kishore, V. (2024). An innovative 4D printing approach for fabrication of anisotropic collagen scaffolds. Biofabrication 17, 015002. doi:10.1088/1758-5090/ad7f8f
Petersen, W., Varoga, D., Zantop, T., Hassenpflug, J., Mentlein, R., and Pufe, T. (2004). Cyclic strain influences the expression of the vascular endothelial growth factor (VEGF) and the hypoxia inducible factor 1 alpha (HIF-1α) in tendon fibroblasts. J. Orthop. Res. 22, 847–853. doi:10.1016/j.orthres.2003.11.009
Popov, C., Burggraf, M., Kreja, L., Ignatius, A., Schieker, M., and Docheva, D. (2015). Mechanical stimulation of human tendon stem/progenitor cells results in upregulation of matrix proteins, integrins and MMPs, and activation of p38 and ERK1/2 kinases. BMC Mol. Biol. 16, 6. doi:10.1186/s12867-015-0036-6
Qi, J., Chi, L., Bynum, D., and Banes, A. J. (2011). Gap junctions in IL-1β-mediated cell survival response to strain. J. Appl. Physiol. 110, 1425–1431. doi:10.1152/japplphysiol.00477.2010
Qi, J., Chi, L., Maloney, M., Yang, X., Bynum, D., and Banes, A. J. (2006). Interleukin-1β increases elasticity of human bioartificial tendons. Tissue Eng. 12, 2913–2925. doi:10.1089/ten.2006.12.2913
Ramos-Mucci, L., Sarmiento, P., Little, D., and Snelling, S. (2022). Research perspectives-pipelines to human tendon transcriptomics. J. Orthop. Res. 40, 993–1005. doi:10.1002/jor.25315
Randelli, F., Sartori, P., Carlomagno, C., Bedoni, M., Menon, A., Vezzoli, E., et al. (2020). The collagen-based medical device MD-Tissue acts as a mechanical scaffold influencing morpho-functional properties of cultured human tenocytes. Cells 9, 2641. doi:10.3390/cells9122641
Riehl, B. D., Park, J. H., Kwon, I. K., and Lim, J. Y. (2012). Mechanical stretching for tissue engineering: two-dimensional and three-dimensional constructs. Tissue Eng. Part B Rev. 18, 288–300. doi:10.1089/ten.TEB.2011.0465
Rinoldi, C., Fallahi, A., Yazdi, I. K., Campos Paras, J., Kijenska-Gawronska, E., Trujillo-De Santiago, G., et al. (2019). Mechanical and biochemical stimulation of 3D multilayered scaffolds for tendon tissue engineering. ACS Biomater. Sci. Eng. 5, 2953–2964. doi:10.1021/acsbiomaterials.8b01647
Roberts, T. J., Chen, M. S., and Taylor, C. R. (1998). Energetics of bipedal running. II. Limb design and running mechanics. J. Exp. Biol. 201, 2753–2762. doi:10.1242/jeb.201.19.2753
Rosenbaum, A. J., Wicker, J. F., Dines, J. S., Bonasser, L., Razzano, P., Dines, D. M., et al. (2010). Histologic stages of healing correlate with restoration of tensile strength in a model of experimental tendon repair. HSS J. 6, 164–170. doi:10.1007/s11420-009-9152-5
Rowson, D. T., Shelton, J. C., Screen, H. R. C., and Knight, M. M. (2018). Mechanical loading induces primary cilia disassembly in tendon cells via TGFβ and HDAC6. Sci. Rep. 8, 11107. doi:10.1038/s41598-018-29502-7
Russo, V., El Khatib, M., Prencipe, G., Mauro, A., Di Giacinto, O., Haidar-Montes, A. A., et al. (2022). Tendon 3D scaffolds establish a tailored microenvironment instructing paracrine mediated regenerative amniotic epithelial stem cells potential. Biomedicines 10, 2578. doi:10.3390/biomedicines10102578
Russo, V., Mauro, A., Martelli, A., Di Giacinto, O., Di Marcantonio, L., Nardinocchi, D., et al. (2015). Cellular and molecular maturation in fetal and adult ovine calcaneal tendons. J. Anat. 226, 126–142. doi:10.1111/joa.12269
Saito, T., Nakamichi, R., Yoshida, A., Hiranaka, T., Okazaki, Y., Nezu, S., et al. (2022). The effect of mechanical stress on enthesis homeostasis in a rat achilles enthesis organ culture model. J. Orthop. Res. 40, 1872–1882. doi:10.1002/jor.25210
Sarmiento, P., and Little, D. (2021). Tendon and multiomics: advantages, advances, and opportunities. NPJ Regen. Med. 6, 61. doi:10.1038/s41536-021-00168-6
Saxena, A., Giai Via, A., Gravare Silbernagel, K., Walther, M., Anderson, R., Gerdesmeyer, L., et al. (2022). Current consensus for rehabilitation protocols of the surgically repaired acute mid-substance achilles rupture: a systematic review and recommendations from the “GAIT” study group. J. Foot Ankle Surg. 61, 855–861. doi:10.1053/j.jfas.2021.12.008
Schneider, M., Angele, P., Jarvinen, T. a. H., and Docheva, D. (2018). Rescue plan for Achilles: therapeutics steering the fate and functions of stem cells in tendon wound healing. Adv. Drug Deliv. Rev. 129, 352–375. doi:10.1016/j.addr.2017.12.016
Schneider, M., and Docheva, D. (2017). Mysteries behind the cellular content of tendon tissues. J. Am. Acad. Orthop. Surg. 25, e289–e290. doi:10.5435/JAAOS-D-17-00520
Schoenenberger, A. D., Tempfer, H., Lehner, C., Egloff, J., Mauracher, M., Bird, A., et al. (2020). Macromechanics and polycaprolactone fiber organization drive macrophage polarization and regulate inflammatory activation of tendon in vitro and in vivo. Biomaterials 249, 120034. doi:10.1016/j.biomaterials.2020.120034
Schulze-Tanzil, G. G., Delgado-Calcares, M., Stange, R., Wildemann, B., and Docheva, D. (2022). Tendon healing: a concise review on cellular and molecular mechanisms with a particular focus on the achilles tendon. Bone Jt. Res. 11, 561–574. doi:10.1302/2046-3758.118.BJR-2021-0576.R1
Scott, A., Cook, J. L., Hart, D. A., Walker, D. C., Duronio, V., and Khan, K. M. (2007). Tenocyte responses to mechanical loading in vivo: a role for local insulin-like growth factor 1 signaling in early tendinosis in rats. Arthritis Rheum. 56, 871–881. doi:10.1002/art.22426
Shen, H., Schwartz, A. G., Civitelli, R., and Thomopoulos, S. (2020). Connexin 43 is necessary for murine tendon enthesis formation and response to loading. J. Bone Min. Res. 35, 1494–1503. doi:10.1002/jbmr.4018
Sheng, R., Jiang, Y., Backman, L. J., Zhang, W., and Chen, J. (2020). The application of mechanical stimulations in tendon tissue engineering. Stem Cells Int. 2020, 1–14. doi:10.1155/2020/8824783
Shiroud Heidari, B., Muinos Lopez, E., Harrington, E., Ruan, R., Chen, P., Davachi, S. M., et al. (2023). Novel hybrid biocomposites for tendon grafts: the addition of silk to polydioxanone and poly(lactide-co-caprolactone) enhances material properties, in vitro and in vivo biocompatibility. Bioact. Mater 25, 291–306. doi:10.1016/j.bioactmat.2023.02.003
Skutek, M., Van Griensven, M., Zeichen, J., Brauer, N., and Bosch, U. (2001). Cyclic mechanical stretching modulates secretion pattern of growth factors in human tendon fibroblasts. Eur. J. Appl. Physiol. 86, 48–52. doi:10.1007/s004210100502
Smith, S. M., Zhang, G., and Birk, D. E. (2014). Collagen V localizes to pericellular sites during tendon collagen fibrillogenesis. Matrix Biol. 33, 47–53. doi:10.1016/j.matbio.2013.08.003
Song, W., Ma, Z., Wang, X., Wang, Y., Wu, D., Wang, C., et al. (2023). Macroporous granular hydrogels functionalized with aligned architecture and small extracellular vesicles stimulate osteoporotic tendon-to-bone healing. Adv. Sci. (Weinh) 10, e2304090. doi:10.1002/advs.202304090
Stanczak, M., Kacprzak, B., and Gawda, P. (2024). Tendon cell biology: effect of mechanical loading. Cell Physiol. Biochem. 58, 677–701. doi:10.33594/000000743
Stirling, P. H. C., and Mceachan, J. E. (2023). In vitro testing of flexor tendon repair models: variability in the current literature. J. Hand Surg. Eur. Vol. 48, 162–168. doi:10.1177/17531934221139068
Stoll, C., John, T., Conrad, C., Lohan, A., Hondke, S., Ertel, W., et al. (2011). Healing parameters in a rabbit partial tendon defect following tenocyte/biomaterial implantation. Biomaterials 32, 4806–4815. doi:10.1016/j.biomaterials.2011.03.026
Stowe, E. J., Keller, M. R., and Connizzo, B. K. (2024). Cellular senescence impairs tendon extracellular matrix remodeling in response to mechanical unloading. Aging Cell 23, e14278. doi:10.1111/acel.14278
Subramanian, A., Kanzaki, L. F., Galloway, J. L., and Schilling, T. F. (2018). Mechanical force regulates tendon extracellular matrix organization and tenocyte morphogenesis through TGFbeta signaling. Elife 7, e38069. doi:10.7554/eLife.38069
Sun, M., Luo, E. Y., Adams, S. M., Adams, T., Ye, Y., Shetye, S. S., et al. (2020). Collagen XI regulates the acquisition of collagen fibril structure, organization and functional properties in tendon. Matrix Biol. 94, 77–94. doi:10.1016/j.matbio.2020.09.001
Suzuki, H., Ito, Y., Shinohara, M., Yamashita, S., Ichinose, S., Kishida, A., et al. (2016). Gene targeting of the transcription factor Mohawk in rats causes heterotopic ossification of achilles tendon via failed tenogenesis. Proc. Natl. Acad. Sci. U. S. A. 113, 7840–7845. doi:10.1073/pnas.1522054113
Svensson, R. B., Heinemeier, K. M., Couppe, C., Kjaer, M., and Magnusson, S. P. (2016). Effect of aging and exercise on the tendon. J. Appl. Physiol. 121, 1353–1362. doi:10.1152/japplphysiol.00328.2016
Szczodry, M., Zhang, J., Lim, C., Davitt, H. L., Yeager, T., Fu, F. H., et al. (2009). Treadmill running exercise results in the presence of numerous myofibroblasts in mouse patellar tendons. J. Orthop. Res. 27, 1373–1378. doi:10.1002/jor.20878
Thampatty, B. P., and Wang, J. H. (2018). Mechanobiology of young and aging tendons: in vivo studies with treadmill running. J. Orthop. Res. 36, 557–565. doi:10.1002/jor.23761
Thorpe, C. T., Godinho, M. S. C., Riley, G. P., Birch, H. L., Clegg, P. D., and Screen, H. R. C. (2015). The interfascicular matrix enables fascicle sliding and recovery in tendon, and behaves more elastically in energy storing tendons. J. Mech. Behav. Biomed. Mater 52, 85–94. doi:10.1016/j.jmbbm.2015.04.009
Tinguely, Y., Shi, V., Klatte-Schulz, F., Duda, G. N., Freedman, B. R., and Mooney, D. J. (2023). Aging and injury affect nuclear shape heterogeneity in tendon. J. Orthop. Res. 41, 2186–2194. doi:10.1002/jor.25649
Tsuzaki, M., Bynum, D., Almekinders, L., Yang, X., Faber, J., and Banes, A. J. (2003). ATP modulates load-inducible IL-1β, COX 2, and MMP-3 gene expression in human tendon cells. J. Cell Biochem. 89, 556–562. doi:10.1002/jcb.10534
Van Helvert, S., Storm, C., and Friedl, P. (2018). Mechanoreciprocity in cell migration. Nat. Cell Biol. 20, 8–20. doi:10.1038/s41556-017-0012-0
Varma, S., Orgel, J. P., and Schieber, J. D. (2016). Nanomechanics of type I collagen. Biophys. J. 111, 50–56. doi:10.1016/j.bpj.2016.05.038
Virdi, J. K., and Pethe, P. (2021). Biomaterials regulate mechanosensors YAP/TAZ in stem cell growth and differentiation. Tissue Eng. Regen. Med. 18, 199–215. doi:10.1007/s13770-020-00301-4
Waggett, A. D., Benjamin, M., and Ralphs, J. R. (2006). Connexin 32 and 43 gap junctions differentially modulate tenocyte response to cyclic mechanical load. Eur. J. Cell Biol. 85, 1145–1154. doi:10.1016/j.ejcb.2006.06.002
Wall, M. E., and Banes, A. J. (2005). Early responses to mechanical load in tendon: role for calcium signaling, gap junctions and intercellular communication. J. Musculoskelet. Neuronal Interact. 5 (1), 70–84.
Wall, M. E., Otey, C., Qi, J., and Banes, A. J. (2007). Connexin 43 is localized with actin in tenocytes. Cell Motil. Cytoskelet. 64, 121–130. doi:10.1002/cm.20170
Wan, R., Luo, Z., Nie, X., Feng, X., He, Y., Li, F., et al. (2024). A mesoporous silica-loaded multi-functional hydrogel enhanced tendon healing via immunomodulatory and pro-regenerative effects. Adv. Healthc. Mater 13, e2400968. doi:10.1002/adhm.202400968
Wang, J. H., Jia, F., Yang, G., Yang, S., Campbell, B. H., Stone, D., et al. (2003). Cyclic mechanical stretching of human tendon fibroblasts increases the production of prostaglandin E2 and levels of cyclooxygenase expression: a novel in vitro model study. Connect. Tissue Res. 44, 128–133. doi:10.1080/03008200390223909
Wang, T., Chen, P., Zheng, M., Wang, A., Lloyd, D., Leys, T., et al. (2018a). In vitro loading models for tendon mechanobiology. J. Orthop. Res. 36, 566–575. doi:10.1002/jor.23752
Wang, T., Thien, C., Wang, C., Ni, M., Gao, J., Wang, A., et al. (2018b). 3D uniaxial mechanical stimulation induces tenogenic differentiation of tendon-derived stem cells through a PI3K/AKT signaling pathway. FASEB J. 32, 4804–4814. doi:10.1096/fj.201701384R
Wu, S., Wang, Y., Streubel, P. N., and Duan, B. (2017a). Living nanofiber yarn-based woven biotextiles for tendon tissue engineering using cell tri-culture and mechanical stimulation. Acta Biomater. 62, 102–115. doi:10.1016/j.actbio.2017.08.043
Wu, Y., Wang, Z., Fuh, J. Y., Wong, Y. S., Wang, W., and Thian, E. S. (2016). Mechanically-enhanced three-dimensional scaffold with anisotropic morphology for tendon regeneration. J. Mater Sci. Mater Med. 27, 115. doi:10.1007/s10856-016-5728-z
Wu, Y., Wong, Y. S., and Fuh, J. Y. (2017b). Degradation behaviors of geometric cues and mechanical properties in a 3D scaffold for tendon repair. J. Biomed. Mater Res. A 105, 1138–1149. doi:10.1002/jbm.a.35966
Wunderli, S. L., Blache, U., Beretta Piccoli, A., Niederost, B., Holenstein, C. N., Passini, F. S., et al. (2020). Tendon response to matrix unloading is determined by the patho-physiological niche. Matrix Biol. 89, 11–26. doi:10.1016/j.matbio.2019.12.003
Xia, H., Nho, R. S., Kahm, J., Kleidon, J., and Henke, C. A. (2004). Focal adhesion kinase is upstream of phosphatidylinositol 3-kinase/Akt in regulating fibroblast survival in response to contraction of type I collagen matrices via a beta 1 integrin viability signaling pathway. J. Biol. Chem. 279, 33024–33034. doi:10.1074/jbc.M313265200
Xiao, H., Zhang, T., Li, C., Cao, Y., Wang, L., Chen, H., et al. (2022). Mechanical stimulation promotes enthesis injury repair by mobilizing Prrx1(+) cells via ciliary TGF-beta signaling. Elife 11, e73614. doi:10.7554/eLife.73614
Xu, B., Song, G., Ju, Y., Li, X., Song, Y., and Watanabe, S. (2012). RhoA/ROCK, cytoskeletal dynamics, and focal adhesion kinase are required for mechanical stretch-induced tenogenic differentiation of human mesenchymal stem cells. J. Cell Physiol. 227, 2722–2729. doi:10.1002/jcp.23016
Xu, K., Shao, Y., Xia, Y., Qian, Y., Jiang, N., Liu, X., et al. (2021a). Tenascin-C regulates migration of SOX10 tendon stem cells via integrin-α9 for promoting patellar tendon remodeling. Biofactors 47, 768–777. doi:10.1002/biof.1759
Xu, P., Deng, B., Zhang, B., Luo, Q., and Song, G. (2021b). Stretch-Induced tenomodulin expression promotes tenocyte migration via F-actin and chromatin remodeling. Int. J. Mol. Sci. 22, 4928. doi:10.3390/ijms22094928
Xu, Y., Wang, Q., Li, Y., Gan, Y., Li, P., Li, S., et al. (2015). Cyclic tensile strain induces tenogenic differentiation of tendon-derived stem cells in bioreactor culture. Biomed. Res. Int. 2015, 1–13. doi:10.1155/2015/790804
Xue, Y., Kim, H. J., Lee, J., Liu, Y., Hoffman, T., Chen, Y., et al. (2022). Co-electrospun silk fibroin and gelatin methacryloyl sheet seeded with mesenchymal stem cells for Tendon regeneration. Small 18, e2107714. doi:10.1002/smll.202107714
Yambe, S., Yoshimoto, Y., Ikeda, K., Maki, K., Takimoto, A., Tokuyama, A., et al. (2024). Sclerostin modulates mineralization degree and stiffness profile in the fibrocartilaginous enthesis for mechanical tissue integrity. Front. Cell Dev. Biol. 12, 1360041. doi:10.3389/fcell.2024.1360041
Yan, Z., Yin, H., Brochhausen, C., Pfeifer, C. G., Alt, V., and Docheva, D. (2020). Aged tendon stem/progenitor cells are less competent to form 3D tendon organoids due to cell autonomous and matrix production deficits. Front. Bioeng. Biotechnol. 8, 406. doi:10.3389/fbioe.2020.00406
Yang, G., Im, H. J., and Wang, J. H. (2005). Repetitive mechanical stretching modulates IL-1β induced COX-2, MMP-1 expression, and PGE2 production in human patellar tendon fibroblasts. Gene 363, 166–172. doi:10.1016/j.gene.2005.08.006
Yin, H., Caceres, M. D., Yan, Z., Schieker, M., Nerlich, M., and Docheva, D. (2019). Tenomodulin regulates matrix remodeling of mouse tendon stem/progenitor cells in an ex vivo collagen I gel model. Biochem. Biophys. Res. Commun. 512, 691–697. doi:10.1016/j.bbrc.2019.03.063
Young, R. G., Butler, D. L., Weber, W., Caplan, A. I., Gordon, S. L., and Fink, D. J. (1998). Use of mesenchymal stem cells in a collagen matrix for Achilles tendon repair. J. Orthop. Res. 16, 406–413. doi:10.1002/jor.1100160403
Yue, J., Jin, S., Li, Y., Zhang, L., Jiang, W., Yang, C., et al. (2016). Magnesium inhibits the calcification of the extracellular matrix in tendon-derived stem cells via the ATP-P2R and mitochondrial pathways. Biochem. Biophys. Res. Commun. 478, 314–322. doi:10.1016/j.bbrc.2016.06.108
Zamboulis, D. E., Marr, N., Lenzi, L., Birch, H. L., Screen, H. R. C., Clegg, P. D., et al. (2024). The interfascicular matrix of energy storing tendons houses heterogenous cell populations disproportionately affected by aging. Aging Dis. 15, 295–310. doi:10.14336/AD.2023.0425-1
Zhang, B., Luo, Q., Deng, B., Morita, Y., Ju, Y., and Song, G. (2018). Construction of tendon replacement tissue based on collagen sponge and mesenchymal stem cells by coupled mechano-chemical induction and evaluation of its tendon repair abilities. Acta Biomater. 74, 247–259. doi:10.1016/j.actbio.2018.04.047
Zhang, J., and Wang, J. H. (2010). Characterization of differential properties of rabbit tendon stem cells and tenocytes. BMC Musculoskelet. Disord. 11, 10. doi:10.1186/1471-2474-11-10
Zhang, T., Wu, Y., Li, X., Zhang, A., Liu, H., Shi, M., et al. (2023). Small extracellular vesicles derived from tendon stem cells promote the healing of injured Achilles tendons by regulating miR-145-3p. Acta Biomater. 172, 280–296. doi:10.1016/j.actbio.2023.10.004
Zhang, X., Bogdanowicz, D., Erisken, C., Lee, N. M., and Lu, H. H. (2012). Biomimetic scaffold design for functional and integrative tendon repair. J. Shoulder Elb. Surg. 21, 266–277. doi:10.1016/j.jse.2011.11.016
Zhu, Z., Gao, R., Ye, T., Feng, K., Zhang, J., Chen, Y., et al. (2022). The therapeutic effect of iMSC-Derived small extracellular vesicles on tendinopathy related pain through alleviating inflammation: an in vivo and in vitro Study. J. Inflamm. Res. 15, 1421–1436. doi:10.2147/JIR.S345517
Keywords: tendon, tenocytes, mechanostimulation, mechanosensation, mechanotransduction, mechanoresponse, cyclic stretch, biofabrication
Citation: Gögele C, Pattappa G, Tempfer H, Docheva D and Schulze-Tanzil G (2025) Tendon mechanobiology in the context of tendon biofabrication. Front. Bioeng. Biotechnol. 13:1560025. doi: 10.3389/fbioe.2025.1560025
Received: 09 July 2025; Accepted: 16 July 2025;
Published: 28 August 2025.
Edited by:
Kalpana S. Katti, North Dakota State University, United StatesReviewed by:
Mohammad El Khatib, University of Teramo, ItalyBiaggio Uricoli, Emory University, United States
Damla Aykora, Bitlis Eren University, Türkiye
Copyright © 2025 Gögele, Pattappa, Tempfer, Docheva and Schulze-Tanzil. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Clemens Gögele, Q2xlbWVucy5nb2VnZWxlQHBtdS5hYy5hdA==