 Mojgan Latifi1
Mojgan Latifi1 Foroozan Jalali Bidgoli1Helia Hajihassani1
Foroozan Jalali Bidgoli1Helia Hajihassani1 Danial Hassani2
Danial Hassani2 Pär K. Ingvarsson3*†
Pär K. Ingvarsson3*† Naser Farrokhi1*†
Naser Farrokhi1*†- 1Department of Cell & Molecular Biology, Faculty of Life Sciences and Biotechnology, Shahid Beheshti University, Tehran, Iran
- 2School of Food and Pharmacy, Institute of Biomaterials and Biomedicine, Shanghai Zhongqiao Vocational and Technical University, Shanghai, China
- 3Department of Plant Biology, Swedish University of Agricultural Sciences, Uppsala, Sweden
Gamma-linolenic acid (GLA) is a biologically active omega-6 fatty acid with anti-inflammatory, immunomodulatory, and cardiovascular protective effects. It is a vital constituent of human health and is finding more widespread applications in nutritional supplements, medications, and functional foods. GLA can be derived from many different natural sources, including plants, fungi, and microorganisms. This review paper presents an overview of the current advances in the discovery, metabolic engineering, and GLA-producing organism optimization. We further present a discussion on new biotechnological approaches—such as culture medium optimization, genetic engineering, and genome editing—that can be employed to enhance GLA production. The paper also presents new trends and directions in the commercial exploitation of GLA-containing products, unveiling new, health-oriented applications.
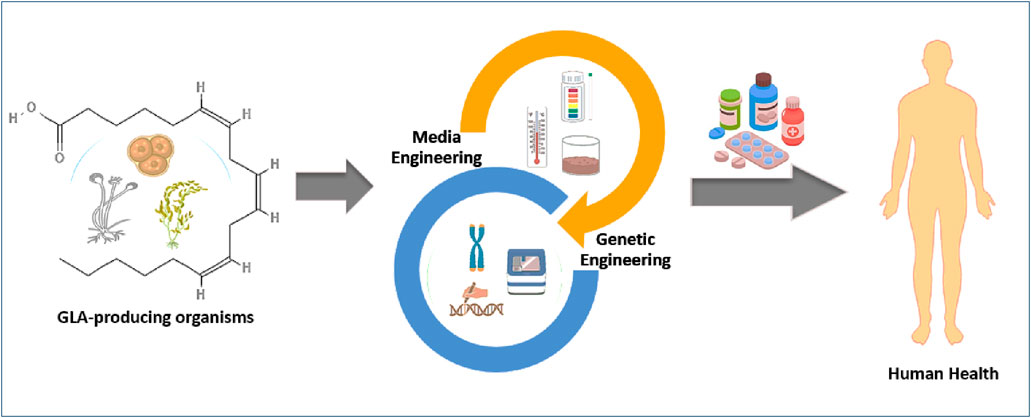
GRAPHICAL ABSTRACT | Organisms that produce GLA can be enhanced through media and genetic engineering techniques to develop better pharmaceuticals and supplements for humans.
1 Introduction
Gamma-linolenic acid (GLA) is a crucial omega-6 polyunsaturated fatty acid (PUFA) with significant implications for human health and nutrition. It is primarily found in seed oils from plants like Ribes nigrum (blackcurrant), Borago officinalis (borage), and Cannabis sativa (hemp). Structurally, GLA consists of an 18-carbon chain with three double bonds at the sixth, ninth, and 12th positions. As a precursor in the biosynthesis of other essential long-chain omega-6 fatty acids, such as dihomo-gamma-linolenic acid (DGLA) and arachidonic acid (ARA), GLA plays a pivotal role in maintaining various physiological processes. However, due to inefficiencies in the conversion of linoleic acid (LA) to GLA in some individuals, it is necessary to obtain GLA from dietary sources (Buist, 2010; Cunnane, 2003).
Beyond its fundamental nutritional value, GLA also possesses bioactive properties. The body can convert GLA into bioactive compounds such as DGLA, which exhibit both anti-inflammatory and potential anticancer effects in vitro (Van Hoorn et al., 2008). This conversion has led to studies exploring GLA supplementation as an alternative to nonsteroidal anti-inflammatory drugs (NSAIDs) for treating conditions like rheumatoid arthritis (Van Hoorn et al., 2008). Additionally, GLA has demonstrated potential benefits in areas such as cardiovascular health, diabetic neuropathy management, treatment of atopic dermatitis, and alleviation of cyclical mastalgia (Belch et al., 1988). Furthermore, GLA is being investigated as a potential therapeutic agent in the treatment of various cancers, including prostate, colorectal and pancreatic cancers (Kairemo et al., 1998; Kalampounias et al., 2024a; Kalampounias et al., 2024b). While most of the evidence remains confined to in vitro studies, these studies have shown promising results, although further clinical trials and epidemiological research are still required. Recent research also highlights that deficiencies in fatty acids, such as GLA, DGLA, ARA, EPA, and ALA may increase susceptibility to diseases like COVID-19. Milder cases of the virus have been observed in children and premenopausal women, who tend to have higher levels of beneficial fatty acids such as GLA and DGLA, which possibly help combat the virus (Das, 2021). This emphasises the unique role of GLA in the immune system, distinguishing it from other fatty acids and showcasing its potential impact on disease prevention.
Given its wide array of health benefits, the commercial market of GLA has experienced significant growth. In 2023, the global market for GLA was valued at US$ 56 million, with projections estimating it will reach US$ 71 million by 2030, reflecting a compound annual growth rate (CAGR) of 3.4% from 2024 to 2030 (Das, 2021). This growth highlights the importance of developing efficient and scalable production techniques, particularly those that incorporate advancements in genetic and metabolic engineering. Overcoming the challenges of scaling up metabolic engineering for GLA biomanufacturing will be essential for meeting future demand and ensuring a reliable, sustainable supply of this valuable compound.
As the scientific understanding of GLA’s health-promoting effects continues to grow, there is an escalating demand for its production, particularly within the pharmaceutical and nutraceutical sectors. Since the human body cannot synthesise GLA, it must be obtained through diet or supplementation (Mudronová et al., 2018). Traditional techniques, such as extracting GLA from plant seeds, often yield limited results and are not always cost-effective. To address these challenges, biomanufacturing approaches, particularly those employing genetic and metabolic engineering, have emerged as more sustainable and efficient alternatives. These biotechnological methods enable higher GLA yields while overcoming the limitations associated with traditional extraction procedures discussed above (Chutrakul et al., 2016).
2 Methodology
This comprehensive review examines the characteristics, biosynthetic pathways, and biotechnological advances of gamma-linolenic acid (GLA)-producing organisms with special reference to microbial and engineered systems. The review followed stringent PRISMA standards of transparency, methodological rigour, and reproducibility in the process of literature screening and analysis.
2.1 Database search and selection strategy
An extensive and systematic search was done across eight major scientific databases: PubMed, Scopus, Web of Science, ScienceDirect, Wiley, Taylor and Francis, SpringerLink, and Google Scholar. Boolean operators were used in combination with suitable keywords such as “gamma-linolenic acid”, “GLA-producing organisms”, “polyunsaturated fatty acid”, “health applications”, and “production methods” to isolate the most relevant articles.
2.2 Inclusion and exclusion criteria
The first search produced 4,493 documents. 1,114 articles were selected for full-text analysis after duplicates were removed and abstracts were screened for relevance. To determine relevance and scientific merit, the following inclusion criteria were applied: original research articles published between 2011 and 2024. English-language papers, and those concerned with GLA-producing organisms (microbial, algal, plant-derived, or engineered systems), direct investigation of biosynthesis pathways, production technology, or therapeutic applications. Exclusion criteria were as follows: non-research content such as news accounts, reviews, and editorials, articles with no full-text access, studies not directly applicable to GLA production or utilisation.
2.3 Data extraction and quality appraisal
Following the full-text screening, 190 articles were screened for thematic synthesis. Studies were thoroughly examined for study design, objectives, method, statistical quality, and direct applicability for GLA production or utilisation. Quality appraisal was conducted using the PRISMA checklist and the Cochrane risk-of-bias tool to select only high-quality evidence-based studies. The retrieved information was organised under the following focus categories: biosynthesis routes and metabolic engineering, genetic engineering platforms for higher GLA yields, extraction and purification technologies for GLA purification, commercial trends, market uses, and scalability.
2.4 Bibliometric analysis and thematic trends
In addition to thematic synthesis, bibliometric analysis was conducted using VOSviewer software (version 1.6.20) to perform keyword co-occurrence analysis of PUFAs in the literature. VOSviewer is a commonly used bibliometric map tool for creating visualisations of the relationships between keywords and, consequently, determining future research directions and lines. At the centre of the analysis was a group of polyunsaturated fatty acids such as GLA, LA, and DGLA, as shown in Figures 1a,b. The labelling of the cluster was based on the frequency and co-occurrence of the corresponding keywords, creating interconnections among the numerous research fields. While VOSviewer enables the visualisation of the scale of relationships, no keyword frequency statistical analysis or trends in cluster emergence were conducted within this study. Full, rather than fractional, counting was employed so that each occurrence of a keyword was counted in its entirety and the landscape of keyword relationships could be defined with a greater degree of granularity, more suitable to the scale of this review.
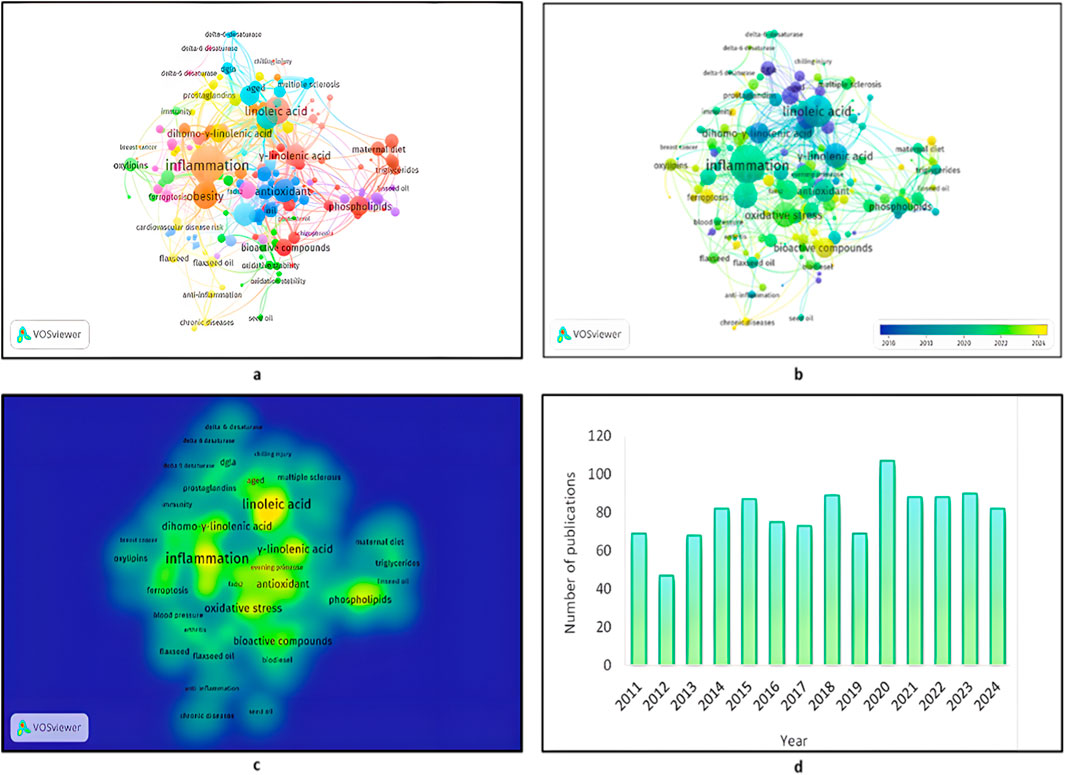
Figure 1. The most frequently occurring keywords in GLA-related research articles. (a) Keyword co-occurrence network of the relationship between GLA, DGLA, LA, and associated biological processes. (b) Clustered visualisation of similar keywords describing research themes such as inflammation, metabolic disorder, and oxidative stress. (c) Frequency of the most frequently used keywords among studies under study, determining research focus. (d) Research article count in relation to different topics in GLA. The keyword analysis was performed employing VOSviewer (version 1.6.20) with a binary counting method and a minimum threshold of two occurrences per keyword.
As compared to fractional counting, this method places equal importance on each instance of a keyword and is not frequency-weighted. This map symbolises the complex web of relationships between GLA, DGLA, and LA, their bioactivities, and their significance in human health. The map also reflected the prime position of these molecules in inflammatory processes, cancer, cardiovascular disorders, and metabolic disorders. Investigating these relationships holds the potential to unveil novel pathways for disease prevention and the development of targeted therapeutic interventions, leading to improved health benefits. Cluster analysis yielded 19 groups, each comprising 8 to 30 keywords, that encompassed different themes related to PUFA studies. The network of keyword co-occurrence positioned GLA, DGLA, and LA at the centre of clusters with connections related to inflammation, obesity, and antioxidants.
Clustering tendency indicated a strong correlation of these fatty acids with inflammatory pathways, as indicated by the dominant orange cluster. GLA and DGLA, as key intermediates of the omega-6 fatty acid pathway, were highly correlated with metabolic and immune responses, indicating their roles in inflammation regulation and lipid metabolism. The blue cluster, rich in antioxidants, is a key area of research emphasis due to its antioxidant activity in preventing oxidative stress induced by lipid peroxidation, which is significantly interconnected with inflammation and obesity. Their interactions with phospholipids and bioactive compounds through LA, GLA, and DGLA also emphasised their role in stabilising cellular membranes and signal transduction. The grouping of the words for phospholipid-related concepts in pink indicates that fatty acids are involved in the function and structure of cell membranes, with the ultimate goal of regulating inflammatory processes. The linking of GLA, DGLA, and LA to the obesity-descriptive words indicates that they are involved in metabolic disturbances and merits further investigation into their use as a drug. Collectively, this visualisation underscores the complex interplay of these fatty acids with basic physiologic processes, affirming their roles in health and disease. The second image is a network visualisation of keyword co-occurrence, where nodes (circles) are terms, and edges (lines) are their co-occurrence in the literature. Larger nodes are more frequent words, and colors mark clusters of relevant terms. At the center of the map are the crucial words such as inflammation, obesity, phospholipids, metabolic syndrome, and chronic diseases, which match their extremely high interconnection with GLA and DGLA.
The blue-to-yellow scale indicates a temporal trend, where blue corresponds to earlier publication dates (2016) and yellow to newer publication dates (2024), and there is an upward trend in interest in subjects such as human cardiotoxicity and neuroprotection. The third image is a density plot illustrating the intensity and frequency of research focus on particular topics. More yellow reflects more research activity, with increasingly blue shadings reflecting relationships with less research. The most researched subjects are GLA, linoleic acid, inflammation, obesity, and phospholipids, which reflect interest in the role that these fatty acids play in inflammation-related diseases and metabolic disorders.
Keywords such as breast cancer, schizophrenia, Alzheimer’s disease, and atopic dermatitis illustrate the development of interest in the therapeutic use of GLA, LA, and DGLA in oncologic, neurological, and inflammatory diseases. The scoping review categorised the current studies into four general themes: the impact of GLA on health, GLA production, manufacturing factors, and directions for future research. These maps were created using VOSviewer (version 1.6.20), employing a complete counting method and a minimum of three mentions per keyword (Figure 1c). A systematic PubMed literature search was conducted to retrieve relevant literature for GLA in biological, chemical, and clinical usage. Literature searching was done with Boolean operators supporting the keywords “gamma-linolenic acid,” “omega-6,” and other similar keywords. Only articles from 2011 to 2024 were chosen, and among these, studies that compared the biochemical, chemical, and clinical significance of GLA were selected. Non-research articles were filtered out to ensure that the dataset consisted only of research studies that report experimental results, therapeutic applications, or biosynthesis pathways of GLA. Out of the total 4,493 documents retrieved from the initial search, these were subsequently screened for inclusion based on specific criteria. Following deduplication and removal of papers unrelated to the research, 1,114 unique studies remained for thematic synthesis. Thematic synthesis revealed trends in research, new themes, and research gaps in the subject area, as well as insights into the evolving understanding of GLA and its potential applications in clinical settings (Figure 1d).
3 Sources of GLA
Research indicates that GLA is present in various plant families, including Boraginaceae, Myricaceae, Onagraceae, Asteliaceae, and Aceraceae (Table 1; Figure 2). GLA is a molecule synthesised by multiple plants, fungi, yeasts, and algae. However, its chemical synthesis is challenging due to the possibility of isomerisation of its double bonds into cis or trans configurations, with only the cis form naturally occurring. Trans isomers of fatty acids can be less biologically active or even harmful compared to their cis counterparts. GLA is naturally found in the oils of several plant seeds, including borage (Borago officinalis), evening primrose (Oenothera biennis), and black currant (Ribes nigrum). Borage oil contains the highest GLA content, ranging from 20%–27%. Although seeds contain more oil than other parts of the plant, different tissues are also recognised as a source of GLA. The Boraginaceae family is particularly noted for its high GLA content due to the presence of specific enzymes that enhance GLA production. Due to the expense and inefficiency of extracting and purifying GLA from natural sources, there is growing interest in using biotechnological methods for GLA production (Pérez-García and Wendisch, 2018). Biotechnological approaches, such as the metabolic engineering of microbes and various fermentation processes, are being explored to offer higher yields, lower costs, and more sustainable alternatives to plant-based extraction.
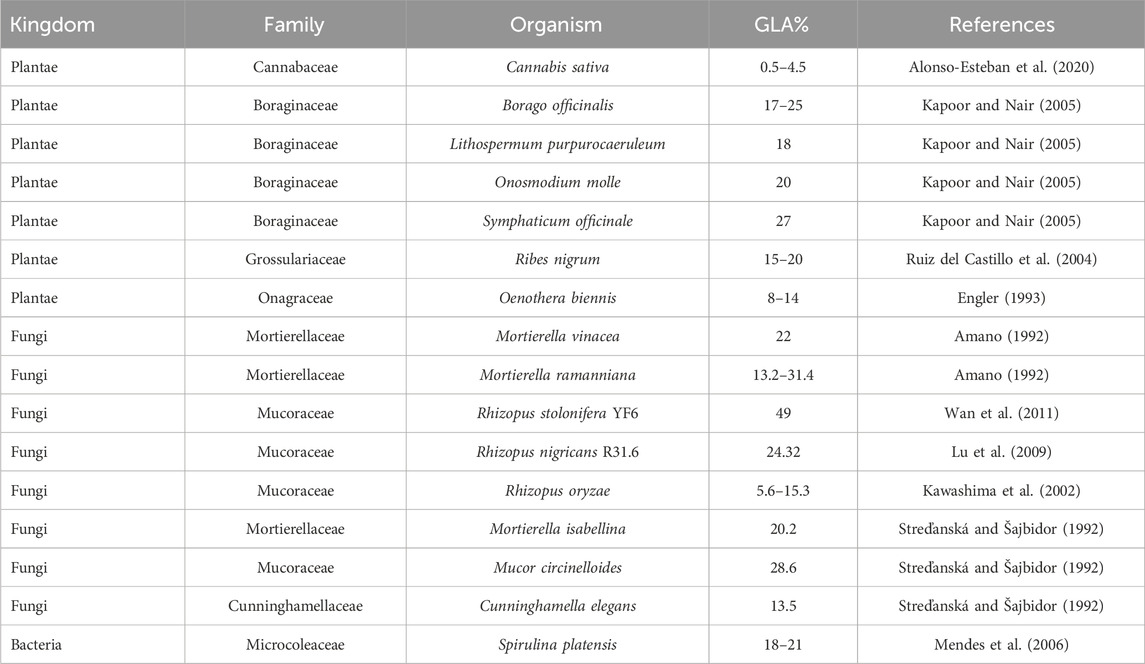
Table 1. GLA-containing organisms.

Figure 2. Plant families containing GLA.
4 Biosynthesis and bioengineering
4.1 GLA biosynthetic pathway
Omega-6 fatty acids, a class of polyunsaturated fatty acids (PUFAs) (Figure 3a), including LA, ARA, GLA, and DGLA, which are primarily found in eggs, nuts, seeds, vegetables, corn oil, and sunflower oil (Czumaj and Śledziński, 2020). These fatty acids are synthesised by the enzymatic conversion of Acetyl-CoA derived from carbohydrates via the glycolytic pathway and NADPH, generated by the malic enzyme in the pentose phosphate pathway, in the cytoplasm. However, the specific pathways for GLA synthesis differ among the organisms producing it. In humans and other animals, GLA is formed following the desaturation of LA as the first step in the synthesis of ARA (Figure 3b) (Kapoor and Huang, 2006; Hernandez, 2016; Simopoulos, 2021). In contrast, this process may differ in microorganisms and plants, reflecting the diversity of enzymatic pathways in various species.
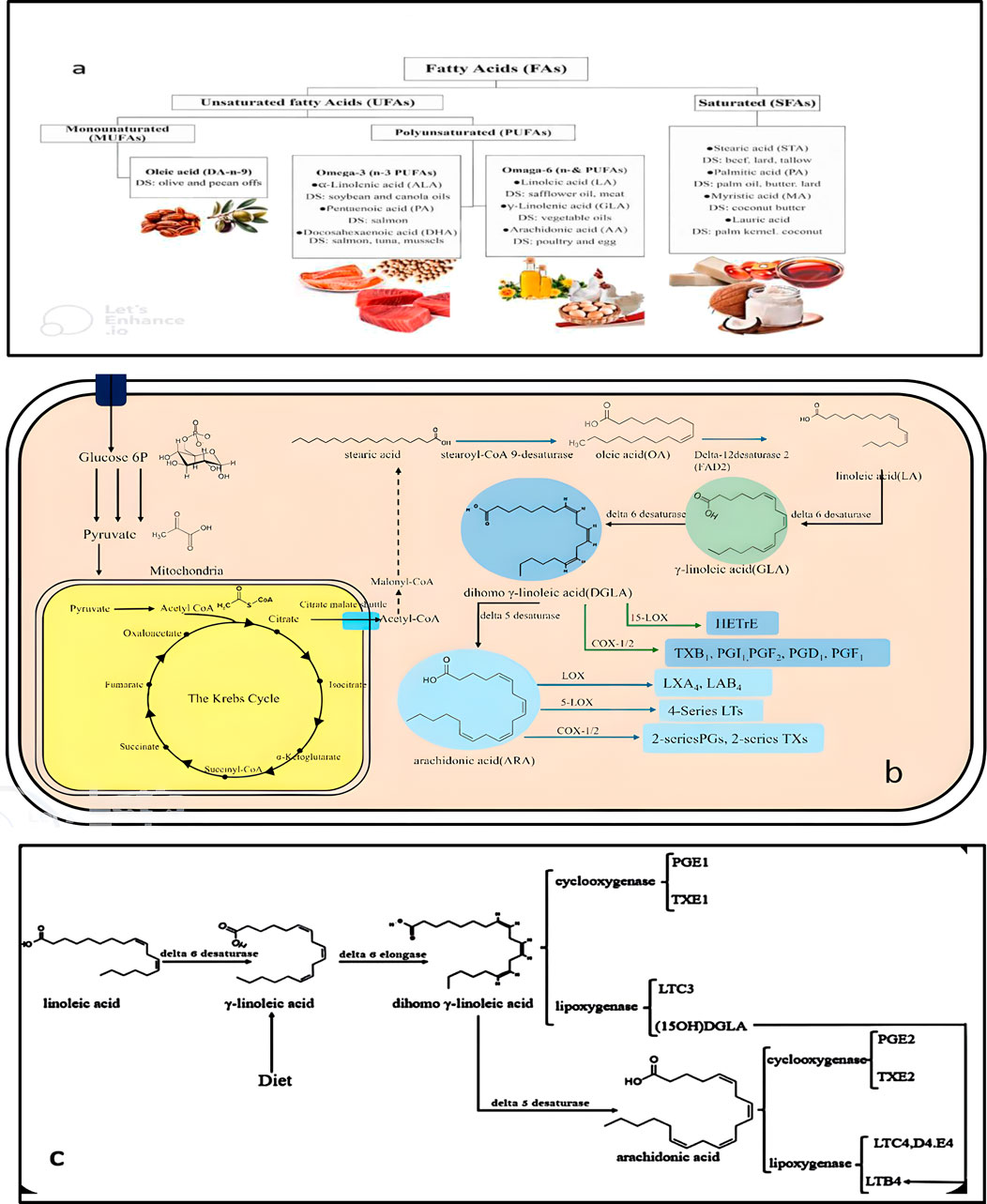
Figure 3. Classification of fatty acids, sources, biosynthesis, and GLA. (a) Saturated (SFAs) and unsaturated (UFAs) are the two categories of fatty acids. Among the unsaturated fatty acids, there are monounsaturated fatty acids (MUFAs) and polyunsaturated fatty acids (PUFAs). Gamma-linolenic acid (GLA) is a polyunsaturated fatty acid (PUFA). DS: Dietary sources. (b) Fatty acid biosynthetic pathway: The biosynthesis of fatty acids in eukaryotic cells generally follows a common scheme initiated with acetyl-CoA and malonyl-CoA, culminating in the formation of fatty acids such as palmitic and stearic acids. Oleic acid (OA), an omega-3 and omega-6 pathway precursor, is formed, and the reaction is initiated when glucose is transported into the cell through specific glucose transporter proteins. The glucose is phosphorylated to glucose-6-phosphate, which is catabolised via glycolysis to yield pyruvate. Pyruvate is fed into the mitochondria and reduced to acetyl-CoA by the complex of pyruvate dehydrogenase. Acetyl-CoA passes into the Krebs cycle and is converted into intermediates like citrate, succinyl-CoA, and malate, before getting converted back to Acetyl-CoA in the cytoplasm. This Acetyl-CoA is carboxylated into malonyl-CoA, which is extended into chains to form stearoyl-CoA, and desaturated into OA. OA is then desaturated to linoleic acid (LA), which is further desaturated to GLA, DGLA, and finally ARA. ARA is metabolised to leukotrienes (LTs), lipoxins (LXs), and cyclooxygenase products such as 2-series prostaglandins (PGs) and thromboxanes (TXs). DGLA is also metabolised via various pathways to yield hydroxy eicosatrienoic acids (HETEs) and other eicosanoids such as TXB1, PGI1, PGF2, PGD1, and PGF1. (c) GLA is converted to DGLA in the liver, and DGLA can be metabolised in three ways: it can be reduced to prostaglandin PGE1 by COX, to 15-hydroxy eicosatrienoic acid (15-HETrE) by lipoxygenase, or to ARA by D5D. The anti-inflammatory activity of DGLA is obtained in three ways: it replaces ARA in inflammatory cells’ membrane, blocks the metabolism of ARA by COX and lipoxygenase by competing with the latter, and forms anti-inflammatory PGE1 that dilates pulmonary vessels. DGLA also forms 15-HETrE and blocks the formation of LTB4 by ARA and is anti-inflammatory.
Functionally, GLA is a precursor to prostaglandins and leukotrienes, which are involved in inflammation and immune responses. The enzyme delta-6 desaturase (D6D) catalyses the desaturation of LA into GLA, and delta-6 elongase (D6E) converts GLA into DGLA. This DGLA subsequently modulates the biosynthesis of various eicosanoids, such as prostaglandins (PGs), leukotrienes (LTs), thromboxanes (TXs), and lipoxins, via cyclooxygenase (COX) and lipoxygenase (LOX) pathways (Kapoor and Huang, 2006). Specific derivatives like PGD2, PGE2, PGI2, PGH3, and LTB4 play vital roles in mediating inflammation, vasodilation, platelet aggregation, and immune signalling (Figure 3c). Importantly, anti-inflammatory mediators such as prostaglandin E1 (PGE1) and 15-HETrE, originating from GLA metabolism, are crucial in regulating fever, smooth muscle contraction, and immune modulation. GLA also influences COX and LOX activity by promoting the synthesis of anti-inflammatory molecules, thereby contributing to immune regulation (Peng et al., 2022; Jang and Park, 2020; Lin et al., 2013; Barry and Dixon, 2021; Gammone et al., 2019; Meechai et al., 2004; Novichkova et al., 2020; Mustonen and Nieminen, 2023; Ikeh et al., 2018; Thurairatnam, 2012). In response to bacterial infections, eicosanoids such as PGE2, PGA2, and series 4 leukotrienes exhibit antibacterial properties and balance inflammatory processes.
Prostaglandins, which are regulated by fatty acids, play essential roles in various bodily functions, including the regulation of organs and immune responses, and contribute to the pathophysiology of cancer and inflammation. Cyclooxygenase enzymes (COX) convert omega-6 fatty acids into prostaglandins, which, in turn, influence cancer cell proliferation. COX inhibitors, such as nonsteroidal anti-inflammatory drugs (NSAIDs), help reduce inflammation and pain (Zarghi and Arfaei, 2011). Gamma-linolenic acid demonstrates promising potential in cancer therapy by selectively inducing apoptosis in cancer cells, while sparing normal cells. It has been shown to disrupt the cancer cell cycle and promote cell death. Moreover, GLA may help prevent inflammatory diseases by converting into dihomo-DGLA, which produces powerful anti-inflammatory agents. Given its anti-inflammatory and immune-modulating effects, GLA holds promise as an adjunct in cancer therapy, although its selective anticancer properties require further investigation (Shi et al., 2023).
4.2 Genetic engineering in GLA-producing organisms
One of the purposes of genetic engineering is to enhance the product yield and efficiency in various systems. Genetic engineering has been utilised in organisms such as Yarrowia lipolytica and Mucor circinelloides to enhance GLA production. In Yarrowia lipolytica, the directed integration of genes encoding delta-12 and delta-6 desaturases increased production. In M. circinelloides, directed evolution techniques were employed to engineer variants with enhanced stability and activity for the treatment of Fabry disease. In plant systems, attempts have been made to produce fatty acids within chloroplasts, but the efficiency and stability of these approaches are still being investigated (Salah et al., 2019).
The selection of host organisms for GLA production must consider multiple factors. The challenges in genetic engineering for production include optimising the expression of critical enzymes involved in the biosynthetic pathway, enhancing production efficiency in host organisms, and ensuring the stability and scalability of production processes (Wu C. et al., 2023). Additionally, challenges may arise in maintaining the integrity of genetic modifications over successive generations of organisms, addressing potential regulatory hurdles related to genetically modified organisms, and optimising fermentation conditions to maximise yield while minimising production costs (Mirbagheri Firoozabad and Akhbariyoon, 2022). These challenges require innovative genetic engineering strategies, robust metabolic engineering approaches, and a thorough understanding of the biochemical pathways involved in GLA biosynthesis to overcome obstacles and achieve efficient and sustainable production. In GLA biosynthesis, three desaturases, delta-9, delta-12, and delta-6, are involved. The genes encoding these desaturases have been cloned from existing prokaryotes to advanced eukaryotes and overexpressed in varieties of hosts, including microalgae, yeast, and plants.
This section examines various host organisms, including plants, fungi, yeasts, algae, and moss, utilised in genetic engineering to enhance GLA production. Each subsection presents valuable insights into the genetic strategies implemented across different systems. The text is rich in detail, offering thorough descriptions of genetic engineering approaches, such as the overexpression of desaturases and gene deletions, which have been shown to improve GLA production in yeasts and fungi. Additionally, comparing the effectiveness of these genetic modifications across various host systems, such as plant-based, microbial, and fungal systems, in terms of yield, scalability, and sustainability would provide a more nuanced perspective.
4.2.1 GLA engineering of plants and mosses
Plants offer advantages such as genetic manipulability, suitability for large-scale cultivation, and are generally regarded as safe (GRAS). However, plants may produce GLA at different levels than other organisms, making them less efficient for production. Currently, GLA sources are limited to a few plant species with weak agronomic properties. Therefore, developing a commercially viable and economical source could be desirable. One approach has been to transfer the D6D gene to convert LA to GLA in oilseed plants (Zhang et al., 2017). Table 2 summarises some plants used as hosts for GLA production (Wu K. et al., 2023). The advantages of plant-based GLA production include cost-effectiveness, compared with traditional chemical synthesis methods, being inherently a renewable resource with sustainable growth, with minimal environmental impact, and efficient in carbon sequestration. Additionally, plant-based GLA can offer nutritional benefits, including omega-3 fatty acids. However, there are challenges and drawbacks, such as limited sources, with only a few plant species currently able to produce significant amounts of GLA, agronomic limitations, as many plants with high GLA content have limited agronomic properties, making large-scale cultivation challenging, and the need for genetic modification, which is often required to enhance GLA production in plants, and can be costly and time-consuming. Recent advancements, such as CRISPR-Cas9 (Kan et al., 2022), have enabled more efficient and targeted genetic modification, leading to the development of crops with enhanced nutritional value, disease resistance, and stress tolerance. These technologies have improved the precision and efficiency of plant genetic modification and have been successfully applied in GLA production. For instance, CRISPR-Cas9 has been used to create plant variants with enhanced GLA content, demonstrating the potential for genetic engineering to optimise GLA yield. In the future, there is likely to be more research focused on optimising plant-based GLA production through genetic engineering and other technologies. Highlighting the potential for developing new plant species or hybrids that could improve GLA yield further reinforces the need for biotechnological advances (Kan et al., 2022).
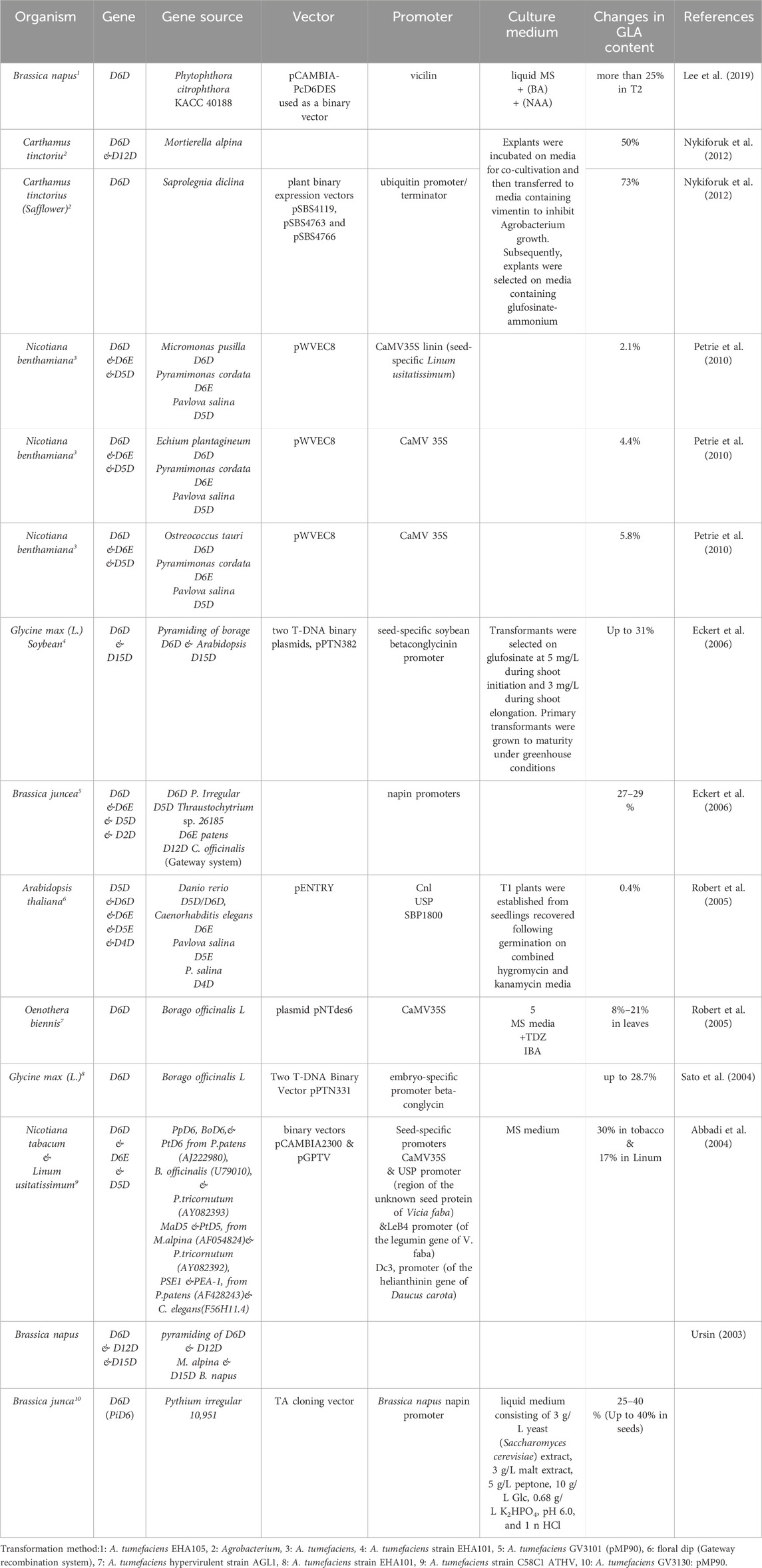
Table 2. Genetic engineering of plants to increase GLA production.
In this context, mosses have emerged as an alternative plant-based platform, particularly in the model organism P. patens, has been significantly advanced through genetic engineering (Beike et al., 2014). Researchers have successfully altered fatty acid biosynthetic pathways by leveraging extensive knowledge of plant oil biosynthesis and employing genetic tools. For instance, the heterologous expression of a D5E gene from the algae Pavlova sp. under the control of a tandemly duplicated 35S promoter enabled the production of ADA in P. patens. This gene facilitated the conversion of endogenous ARA to ADA, achieving a production level of 0.42 mg/L, which was further optimised to 4.51 mg/L using RSM and increased GLA and DGLA content. This study marks a pioneering effort in expressing a PUFA-synthesising enzyme in a non-seed, lower plant without the need for exogenous fatty acid supplementation (Kaewsuwan et al., 2010; Chodok et al., 2012). Moss-based systems, such as P. patens, offer a unique alternative to traditional plant-based systems for GLA production. Unlike seed plants, mosses can be grown in controlled environments without large agricultural spaces. Mosses can be genetically engineered to produce high levels of GLA and other valuable compounds, making them a promising option for researchers exploring alternative non-seed plant hosts for biotechnological applications.
4.2.2 GLA engineering of fungi and yeasts
Fungal genetic engineering can increase GLA production by improving yield, efficiency, and cost-effectiveness compared to traditional chemical synthesis. However, challenges such as scalability issues, high production costs, and potential negative impacts on fungal growth and viability must be addressed. To overcome these challenges, it is essential to optimise genetic engineering techniques, select suitable fungal strains, develop efficient fermentation processes, and explore cost-reduction strategies. The potential of fungal-based GLA production can be effectively realised by addressing these factors.
• Methods to enhance DGLA production include overexpressing the transketolase gene and disrupting a glucan synthase gene.
• Researchers have achieved a 1.9-fold increase in free DGLA yield to 403 mg/L by overexpressing an anticipated transketolase gene in DGLA3.
• Disturbing the α-1,3-glucan synthase gene agsB, involved in cell-wall biosynthesis, has increased the content by 1.3-fold–533 mg/L, resulting in a 2.5% increase in overall yield.
These findings are illustrated in Table 3, which depicts the genetic engineering of fungi to increase the amount of GLA (Tamano et al., 2023).
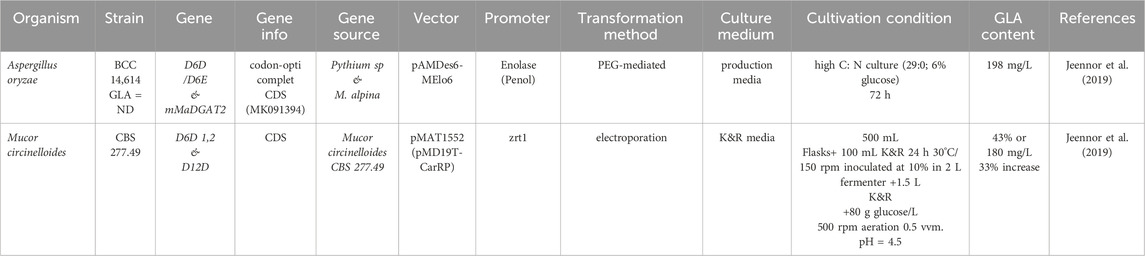
Table 3. Genetic engineering of fungi to increase the amount of GLA.
The D6D gene from Mucor rouxii, involved in GLA formation, was introduced into Hansenula polymorpha yeast. Changes in essential fatty acid synthesis pathways were observed, with GLA production varying based on growth conditions (Laoteng et al., 2005). One study on 28 Zygomycetes fungi revealed that strains exhibiting low oleic acid synthesis due to reduced delta-9 desaturase activity displayed maximal delta-15 desaturase activity. Delta-6 desaturase activities indicated competition among fatty acids in n3, n6, and n9 biosynthetic pathways. Understanding these fatty acid desaturase activities presents new opportunities for optimising the biotechnological production of PUFAs by Zygomycetes fungi (Klempova et al., 2013).
The ease of genetic manipulation and the high efficiency of homologous recombination in yeasts make them ideal platforms for synthetic biology projects and industrial biotechnology applications (Table 4) (Schindler, 2020; Taghon and Strychalski, 2023; Gupta et al., 2024). The codon-optimised D6D from M. alpina was introduced into Yarrowia lipolytic, controlled by the solid hp4d promoter. The engineered strains produced a total fatty acid profile containing 4.6% GLA (Gu et al., 2023).
• In a study on two strains of Y. lipolytica (strain Po1f-Δk and 2Pg2E), researchers produced GLA by eliminating specific genes. They removed the KU70 gene from strain Po1f to create Po1f-Δk and the FAD2 gene to produce strain 2Pg2E. During shake flask fermentation, the synthesised strains produced GLA, which accounted for 22.58% of the total fatty acids (TFAs). Additionally, the corresponding total titers reached 386.59 mg/L.
• Scientists are focusing on modifying the endogenous pathways of yeast, particularly Y. lipolytica, by expressing critical enzymes involved in GLA biosynthesis, such as Acetyl-CoA synthase, Acetyl-CoA carboxylase, and fatty acid synthesis enzymes. These modifications improve the yeast’s ability to utilise acetate during fermentation, increasing lipid production and GLA content (Mirbagheri Firoozabad and Akhbariyoon, 2022).
• Strategies such as inhibiting β-oxidation through genetic deletions and editing the diacylglycerol biosynthesis pathway have been employed to enhance GLA production in yeast cultures further. By optimising these genetic and fermentation processes, researchers are working towards developing sustainable and efficient methods for mass-producing in engineered oleaginous yeasts like Y. lipolytica to meet the increasing demand for this essential fatty acid in various applications (Firoozabad and Akhbariyoon, 2021).
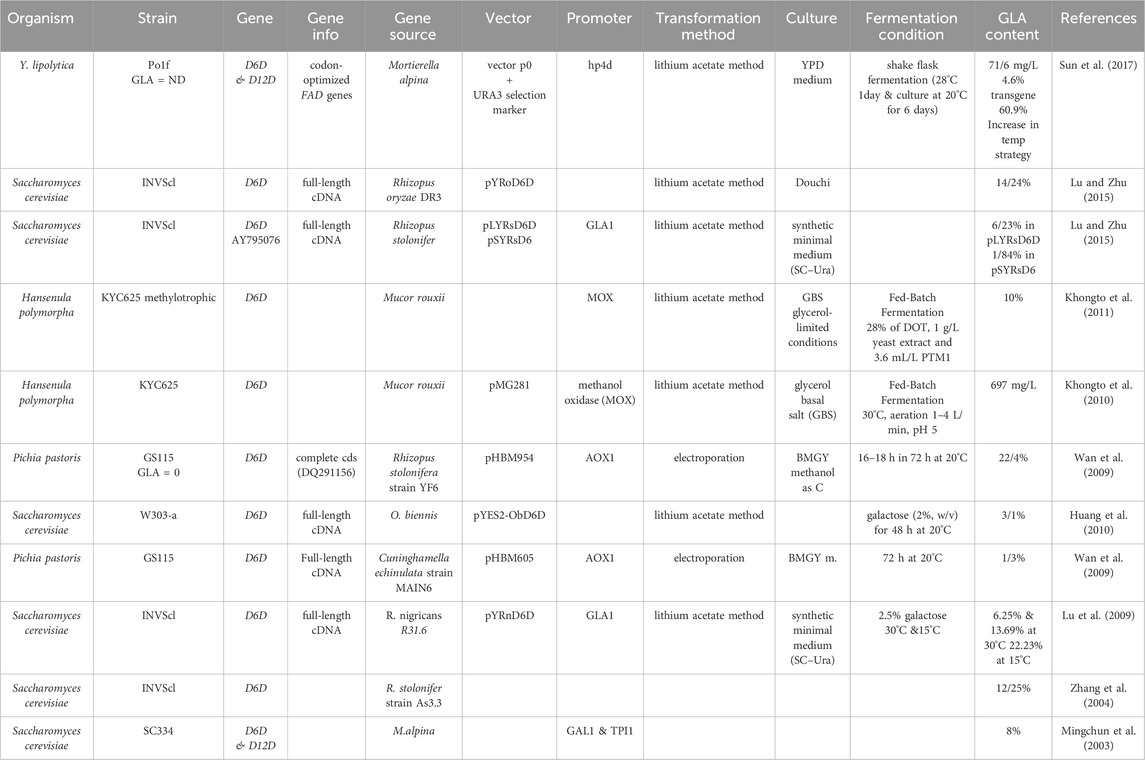
Table 4. Genetic engineering of yeasts to increase GLA production.
Recently, a study was conducted to improve the expression of D6D, a necessary enzyme in the GLA pathway. For the clone M. rouxii delta-6 desaturase, the expression vector pPICZC was used. In Escherichia coli, the engineered vector was first cloned. E. coli DH5α and, after plasmid extraction and sequencing confirmation, was transformed by electroporation into Pichia pastoris GS115. The results showed that in the presence of 0.5% methanol, a genetically modified yeast strain expresses this gene. Lipids and essential fatty acids, particularly GLA, have been obtained to verify the expression. In the studies on the production of lipids and fats, a recombinant strain generated GLA levels up to 19.2% of overall fat content by Sudan black and Nile Red staining, GC, and flow cytology (Mirbagheri Firoozabad and Akhbariyoon, 2022; Chaudhary et al., 2024). The yeast section could benefit from more context regarding the industrial relevance of these modifications. For instance, genetically engineered yeasts provide advantages in terms of scalability and production cost compared to other hosts, making them a promising option for the commercial production of GLA.
Oleaginous yeast technology offers an attractive means of augmenting the yield of lipids and GLA without the challenges of using microalgal sources. Microalgae-derived GDGs and GLA are structurally complex, rare, and difficult to isolate or produce, even with significant demand. Y. lipolytica is a potential alternative, as it is capable of metabolising hydrophobic substrates and has the potential to redirect its metabolic pathway capabilities toward lipid biosynthesis. Its robust lipid accumulation routes and stress tolerance qualify it to be an organism for the large-scale production of GLA. Lipid biosynthesis in Y. lipolytica has been advanced significantly by synthetic biology, gene editing, and metabolic engineering. Advances in biosynthetic genes, metabolic pathways, and fine-tuning of regulation have contributed to enhanced GLA productivity. Moreover, strategies such as adaptive evolution, cocultivation, and bioprocessing have also been optimised for lipid accumulation. Engineering Y. lipolytica for GLA includes redesigning glycosylation pathways and fatty acid biosynthesis to achieve stereoselective synthesis. Subsequent studies will emphasize integrating omics technologies, optimisation of substrate metabolism, and yeast strain engineering towards achieving high-yield, low-cost, and environmentally friendly biosynthesis of GLA (Gu et al., 2023).
Manipulation of Y. lipolytica, both in its genetic background and in culture conditions, has resulted in successful improvement of lipid storage as well as elevating the ω-6 PUFA biosynthesis involving GLA and DGLA. Scientists have optimized the Δ6 and Δ8 biosynthetic pathways of Y. lipolytica, which were initially sourced from M. alpina and Isochrysis galbana, respectively. The Δ6 pathway is more effective for GLA production, while the Δ8 pathway is more effective for the production of DGLA and ARA. Genetic engineering technologies such as the deletion of KU70 to increase homologous recombination and FAD2 to regulate fatty acid desaturation have significantly influenced lipid composition. Additionally, metabolic engineering technologies with strategies to increase precursor supply and suppress substrate competition have also improved ω-6 PUFA production. As a result, engineered strains have exhibited high lipid productivity, with DGLA reaching up to 46.65% of TFAs, confirming the viability of Y. lipolytica as an effective microbial platform in producing industrial ω-6 PUFAs. Optimisation of culture conditions has also played a critical role in favouring lipid and ω-6 PUFA accumulation in genetically modified Y. lipolytica strains. For example, Po1f-Δk and 2Pg2E strains were developed to increase homologous recombination efficiency and initiate OA production, respectively. Genetic engineering led to significant lipid composition alteration, where Po1f-Δk contained 37.52% OA and 8.32% LA of TFAs, and 2Pg2E yielded an OA ratio of 82.31%. The lipid content was 8.50%–14.55% DCW, an indication of the success of selective genetic improvement. Additional optimisations of ω-6 PUFA titers were realised through the alleviation of substrate competition and optimisation of necessary biosynthetic precursors (Wang et al., 2023).
4.2.3 GLA engineering in algae
Algae genetic engineering holds promise for biofuel production, carbon capture, and the synthesis of valuable compounds such as omega-3 fatty acids and pigments (Kumar et al., 2020). Advances in genetic tools and techniques enable the modification of algae to enhance photosynthetic efficiency, increase biomass yield, and produce target metabolites, thereby contributing to sustainable energy solutions and bioproducts (Grama et al., 2022). Algae have been found to contain the necessary genes for GLA production, including the enzymes D6D and D5D. For example, the green algae Chlamydomonas reinhardtii has been discovered to have a D6D enzyme capable of converting LA into GLA. Researchers have tried to modify the genetic makeup of algae to produce GLA. A study successfully introduced a D6D gene from M. alpina into C. reinhardtii, leading to increased GLA production. Another study engineered the green algae Scenedesmus obliquus to produce GLA by overexpressing a D6D gene from Arabidopsis thaliana. Additionally, synthetic biology methods have been explored to create new pathways for GLA production in algae (Campos et al., 2020). Microalgae face several challenges in their use for GLA production. These include slow growth rates, lower GLA yields compared to heterotrophic hosts, high infrastructure, operation, and maintenance costs, as well as difficulties in engineering native host microalgae for GDGs-GLA production. Additionally, seasonality and geographical location can limit algae availability and output, while the lack of regulation for algae as dietary supplements poses further challenges (Bleakley and Hayes, 2017). Algae can be genetically engineered to enhance photosynthetic efficiency, potentially leading to higher biomass yield and target metabolite production. Algae are also capable of carbon capture, contributing to sustainable energy solutions. Like yeast or fungi, algae often face slow growth rates and lower GLA yields. The high infrastructure and operation costs and environmental dependencies, such as seasonality and geographical locatio,n add to the complexity of using algae for GLA production.
4.3 Culture condition engineering
The culture conditions in engineering play a crucial role in enhancing the production of lipids and GLA in microorganisms. Studies on Mucor rouxii and Mucor sp.1b have provided valuable insights into the optimal culture conditions for high accumulation. These strains thrive at low temperatures, around 25°C, and with vigorous light intensity, specifically at 6 kLux. Additionally, adding a primrose oil supplement at a concentration of 0.8% w/v has further enhanced production in these strains. The choice of carbon and nitrogen sources significantly affects lipid and GLA production. Specific carbon sources, such as glucose, sucrose, starch, and lactose, have been identified as influential factors in the growth and metabolism of these microorganisms. Similarly, nitrogen sources like potassium nitrate have been found to impact lipid and GLA production by carefully selecting and optimising these nutrient sources. Scientists can create a favourable environment for the organism, increasing lipid and GLA production. Culture condition engineering is a multifaceted approach that involves manipulating specific cultural parameters to create a favourable environment for the organism to boost GLA production. This can include adjusting temperature, light intensity, and nutrient sources and employing other genetic and metabolic engineering techniques (Table 5) (Hansson and Dostálek, 1988; Certik et al., 2012).
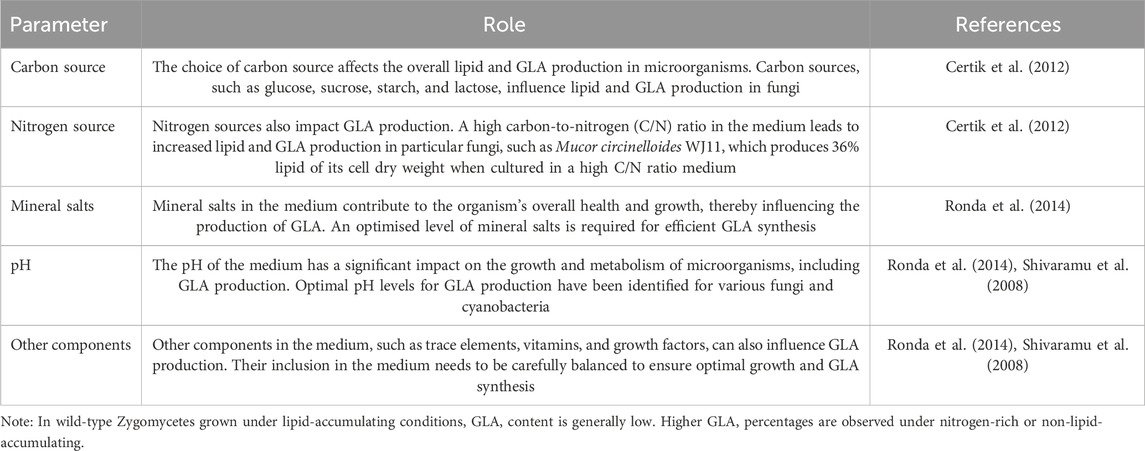
Table 5. Effective parameters on culture optimisation.
Optimising culture conditions, such as temperature and carbon source,s has enhanced GLA production in fungi. The two methods are solid-state fermentation (SSF) and submerged fermentation (SMF). SSF is superior due to its lower water activity, which supports better growth and metabolite production in some fungi.
4.3.1 Culture optimization in plants and mosses
The production of GLA in plants is significantly affected by salinity stress (Urrestarazu et al., 2018). Salinity increases GLA production by activating specific stress-induced pathways that benefit GLA biosynthesis. These pathways enhance the plant’s ability to produce and accumulate GLA in response to stress conditions. Researchers have investigated the impact of salinity on GLA accumulation, discovering that various factors, including light intensity and quality, temperature, carbon dioxide concentration, nutrient availability, water stress, and plant hormones, influence GLA production. For instance, increasing light intensity and using blue or red light can enhance GLA production more effectively than white light. The optimal temperature range for GLA production is typically between 18°C and 22°C, and elevated CO2 concentrations can increase photosynthesis and, consequently, GLA production. Nutrient deficiencies, especially nitrogen and phosphorus, can limit GLA production. At the same time, water stress and plant hormones can affect gene expression and metabolic pathways involved in fatty acid synthesis, thus impacting GLA production. Researchers aim to enhance GLA production in plants for industrial applications by optimising culture conditions through genetic engineering, environmental manipulation, and bioreactor design. Studies have also indicated increased oil content and specific fatty acid compounds, like PA, SA, and GLA, at higher salinity levels. Moreover, a positive correlation exists between D6D gene expression and GLA content, particularly at high salinity levels. Mit Wan utilised saline water as an alternative source for Oenothera biennis production in plant cell culture (Shahbazi et al., 2018).
Similar to higher plants, mosses also respond to environmental and nutritional factors that influence PUFA biosynthesis, including GLA, making them attractive candidates for controlled culture-based production strategies. Researchers have manipulated factors such as light intensity, temperature, CO2 concentration, and nutrient availability to increase GLA production in mosses. Higher light intensity and optimal temperatures can boost GLA production. Elevated CO2 levels can further enhance crop output, and a proper nutrient supply is essential for achieving high yields (Decker et al., 2003). Moss Physcomitrella patens, employing response surface methodology (RSM), in a culture medium containing sucrose (62.92 g/L), potassium nitrate (0.80 g/L), and glutamate (1.42 g/L) showed a significant increase (4.61-fold) in GLA production compared to the standard BCD medium (Chodok, 2022).
In a study, high amounts of very long-chain PUFAs, such as ARA, were reported in 7 moss species (Physcomitrella patens, Encalypta streptocarpa, Pottia lanceolata, Plagiomnium undulatum, Atrichum undulatum, Brachythecium rutabulum, Rhynchostegium (Mural). These species were grown under fully standardised laboratory axenic culture conditions for comparative metabolic studies. The species were modified in Knop medium under light intensity conditions of 55–70 μmol/m/s and a photoperiod of 16 h of light and 8 h of dark at 23°C. In the model organism P. patens, differences in fatty acid composition were observed between filamentous protonema and leaf gametophores, as determined by desaturase obtained from microarray analysis, consistent in both growth stages (Beike et al., 2014). Depending on the growth stage and species, the amount of ARA varied between 6% and 31% of total fatty acids. Spatial investigation of the corresponding FADS revealed the endoplasmic reticulum as the cellular compartment for ARA synthesis. The results showed that very long PUFAs are very abundant metabolites in mosses. Standard cultivation techniques using photobioreactors, combined with the availability of genome sequences and the high rate of homologous recombination in the mitotic cells of P. patens, form the basis for targeted metabolic engineering (Decker and Reski, 2012).
A research study identified and modelled the key factors that significantly affect the production of unsaturated fatty acids and biomass P. patens. The study analysed nine culture variables: temperature, agitation speed, pH, Sucrose, Diammonium Tartrate, CaCl2.2H2O, MgSO4.7H2O, KH2PO4, and KNO3 in a solid BCD environment using the Plackett-Burman statistical design method. The statistical analysis revealed that the moss’s pH and temperature significantly impact biomass production and the production of polyunsaturated fatty acids (PUFAs), such as linoleic acid (LA) and gamma-linolenic acid (GLA). Additionally, the study found that three nutritional variables–sucrose, CaCl2, and MgSO4 – only affected the production of specific PUFAs. Specifically, higher concentrations of sucrose were found to positively influence the production of LA, ARA, and EPA. On the other hand, higher concentrations of metal ions like CaCl2 and MgSO4 were found to hurt the production of ARA and EPA. The study concluded that pH, temperature, sucrose, CaCl2, and MgSO4 are critical parameters for the growth of P. patens and the production of PUFAs by this moss (Chodok et al., 2010). Hence, the P. patens cell suspension culture is regarded as the most suitable medium for PUFA synthesis and could be an appropriate source for producing and commercialising PUFAs. However, the results of this preliminary study on P. patens did not progress to the oil extraction stage (Chodok et al., 2010).
In another study, the D5E gene was transferred from Pavlova sp. algae to P. patens strain Gransden to increase the production of fatty acids. For this purpose, the D5E gene was cloned under the control of the CaMV 35S promoter in the Gateway pMDC43 vector. Then, gene transfer was performed using polyethene glycol (PEG). The concentration of ADA and DPA increased dramatically and increased to 24.3 and 11.7 mg/L, respectively, and accounted for 1.1% and 2.3% of the total fatty acids. The obtained results prove that the pathway of fatty acid biosynthesis with genetic manipulation and food supplements is a promising method for the production of specific PUFAs in a simple seedless plant (Chodok et al., 2012). In another study, two D12Ds associated with the early stages of long-chain PUFA biosynthesis were successfully cloned from P. patens, and their functions were characterised. The open reading frames (ORF) of PpFAD2-1 and PpFAD2-2 consist of 1,112 base pairs and code for 375 amino acids. The polypeptides obtained from them showed 62% and 64% similarity with microsomal D12D from higher plants, and each contained three typical histidine clusters. Examining the expression of these genes in the yeast system indicates the production of a significant amount of PUFA. In addition, decreasing the growth temperature from 30°C to 15°C increased the accumulation of unsaturated fatty acid products (Chodok et al., 2013) (Figure 4). While the genetic manipulation of mosses offers intriguing possibilities, it presents challenges in terms of cost, scalability, and efficiency compared to other host systems. Genetic modifications in mosses are typically more labour-intensive and time-consuming. However, the unique advantages of mosses, such as their high rate of homologous recombination and ability to grow in controlled environments, make them valuable for targeted metabolic engineering and PUFA production.
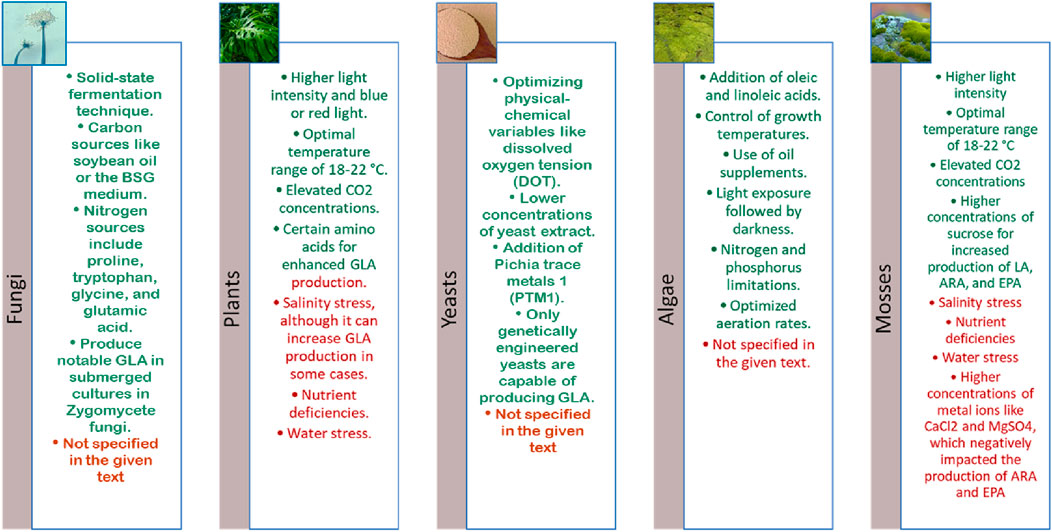
Figure 4. Positive (green) and Negative (red) effects of each organism. This figure summarises the key biological sources and influencing factors on γlinolenic acid (GLA) production. GLA is synthesised by certain Zygomycete fungi such as Mucor spp. and Thamnidium elegans, particularly in submerged culture systems, while edible and medicinal mushrooms do not produce GLA. In plants, environmental factors like light, temperature, CO2 concentration, and specific amino acids (e.g., glutamine, histidine, cysteine) enhance GLA biosynthesis, whereas salinity, water limitation, and nutrient stress may suppress it. GLA production in algae is species-dependent and influenced by factors such as light intensity, nutrient availability, and medium supplementation. Mosses show limited data on GLA biosynthesis, and exposure to stress or heavy metals tends to reduce their polyunsaturated fatty acid levels. Lastly, yeasts require genetic engineering to produce GLA, as wild-type strains lack the biosynthetic pathway. Culture conditions like pH, carbon-to-nitrogen ratio, and aeration predominantly affect lipid accumulation in yeasts.
4.3.2 Culture optimization in fungi and yeasts
More recently, research interests have been oriented toward the feasibility of some fungi species that may be able to produce a desirable PUFA named GLA. There are various fungi species, including Umbelopsis isabellina (Papanikolaou et al., 2004), Cuninghamella echinulata (Fakas et al., 2008a), M. ramanniana (Dyal and Narine, 2005), M. alpina, and M. rouxii (Shivaramu et al., 2008), where it is reported to produce notable levels of GLA under optimised culture conditions.
To improve GLA yield, two standard fermentation methods are utilised: SSF and submerged fermentation (SMF) (Khongto et al., 2011). SSF offers advantages such as high yields of GLA, lower production costs, reduced equipment requirements, and minimal waste generation (Maurice, 2019). It is particularly beneficial for the production of enzymes and other metabolites required for GLA synthesis. On the other hand, SMF allows for greater control over fermentation parameters, such as temperature, pH, and oxygen levels, resulting in more consistent and scalable production. Additionally, SMF facilitates better mass transfer of nutrients, contributing to more efficient fermentation processes (Belorkar and Jogaiah, 2021).
Optimisation of carbon and nitrogen sources is a significant strategy for enhancing GLA production in fungal fermentation. For example, the supplementation of soybean oil (10 mL/L) extensively promoted lipid production in Phaffia rhodozyma, which resulted in high levels of fatty acids like palmitic acid, stearic acid, oleic acid, linoleic acid, alpha-linolenic acid, and GLA (Li et al., 2023). Moreover, M. alpina and M. circinelloides have been reported to be the most efficient fungi for the production of GLA. M. alpina produced 7 g/L biomass and 900 mg/L of fatty acids, with 41% of the fatty acids being polyunsaturated, like ARA and thereby a strain of interest for the production of ARA. Conversely, M. circinelloides produced 6.5 g/L biomass and 600 mg/L of fatty acids, beating Rhizopus stolonifer, which had produced just 250 mg/L of GLA (Serra et al., 2023).
Other studies have investigated the impact of various carbon and nitrogen sources on GLA production. For instance, research on Mucor plumbeus with Bacillus subtilis bacteria found that the combination of cellobiose as the carbon source and ammonium sulfate as the nitrogen source maximised biomass, lipid, fatty acid, and GLA yield. The optimal fermentation conditions were a pH of 6.5, a temperature of 30°C, 10% inoculum, 200 rpm agitation, and a 5-day incubation period. Notably, cocultures of M. plumbeus and Bacillus subtilis produced GLA concentrations that were twice as high as those in monocultures. In addition, the role of amino acids in enhancing GLA productivity was also investigated, and results showed that strains like Rhizomucor pusillus AUMC i11616. A and Mucor circinelloides AUMC 6696 had increased GLA production when cultured in proline and tryptophan as the sources of nitrogen, with an increase of 38% and 25%, respectively, compared to an ammonium tartrate control. Glycine and glutamic acid were also shown to have significant functions as precursors of total fatty acid (TFA) synthesis (Azemi et al., 2022).
A novel technique, overexpression of mitochondrial malate transporter proteins, has also hastened lipid accumulation in M. circinelloides. During a 24-h fermentation period that included the addition of 0.8% malic acid (MA), recombinant strain M. circinelloides McMT2 yielded 16 g/L fungal dry biomass, 32% lipid content, and 5.12 g/L yield of lipids. These were 1.6 times greater in biomass and lipid content, and 2.56 times greater in lipid yield compared to the control batches not supplemented with MA.
Apart from malic acid, other carbon sources have been explored for their effectiveness in the yield of fungal lipids. For example, M. isabellina DSM 1414 exhibited the highest biomass and oil production of 10.80 g/L and 5.44 g/L, respectively, under lactose-based culture conditions. The major fatty acids formed under the conditions above were oleic acid (20.42%–42.94%), palmitic acid (14.96%–22.19%), stearic acid (9.00%–26.92%), linoleic acid (11.35%–18.67%), and GLA (3.56%–8.04%) (Zhang Y. et al., 2021).
Furthermore, Rhodosporidium toruloides AS 2.1389 was also studied in lipid production with acetic acid as a cheap carbon source. The organism demonstrated higher lipid content when grown in 4–20 g/L acetic acid than in glucose-based media. Optimal conditions with 20 g/L acetic acid and a C/N ratio = 200 resulted in 4.35 g/L biomass with 48.2% lipid content. Since there is an increased demand for natural sources of beta-carotene and C18 PUFAs, a study was conducted to assess the potential of 28 isolates of Zygomycetes, which were collected from soil. While all the fungi produced C18 PUFAs, nine were capable of producing beta-carotene. The best beta-carotene and GLA producer was Actinomucor elegans CCF 3218, with a yield of 251 mg/L. Conversely, U. isabellina CCF 2412 was the most promising strain because it had the dual potential to produce both GLA (217 mg/L) and beta-carotene (40.7 mg/L) (Huang et al., 2016; Klempova et al., 2013).
Cunninghamella bainieri 2A1 was also explored for its GLA and lipid production under conditions of nitrogen starvation. The highest lipid (6.2 g/L) and GLA (0.4 g/L) content were achieved using 20.2 g/L glucose and 2.12 g/L ammonium tartrate-containing media and were harvested at 48 h (Ganjali Dashti et al., 2014).
Certik et al. (2006) investigated the impact of substrate selectivity on the production of GLA in various strains of fungi that belong to the Zygomycete class. Their findings emphasised the significant role played by carbon sources in the biosynthesis of GLA, as oat flakes were found to be a superior substrate compared to peeled barley. Among the strains that were tested, Thamnidium elegans CCF 1465 produced the highest GLA content, at 4.82 g/kg on oat flakes but only 1.11 g/kg on peeled barley. Additionally, Cunninghamella elegans (CCF103 and CCF1318) and Mucor mucedo CCF1348 showed a tremendous growth enhancement and GLA yield when grown on oat flakes, with yields of 3.58–4.56 g/kg, while only 0.82–1.15 g/kg on peeled barley. This finding strongly suggested that oat flakes are an excellent carbon source, which favours the production of lipids in fungi (Čertik et al., 2006). In comparing the productivity of GLA in different species, Thamnidium elegans CCF 1,465 was the most productive strain, followed by Mucor and Cunninghamella species, which produced moderate levels. M. isabellina produced comparatively low GLA content, reflecting poor biosynthetic efficiency. The Rhizopus species (R. oligosporus, R. microspores, and Rhizopus arrhizus) produced consistently less than 1.0 g/kg of GLA on both substrates.
With optimised conditions, biomass yield, total lipid, and GLA yield were examined in several Zygomycetous isolates. The study highlighted the impact of varied culture conditions—pH, temperature, nitrogen, and carbon sources—on GLA productivity. Cunninghamella blakesleeana JSK2 isolates from Karnataka (India) soil samples were identified as the highest-yielding GLA among the ten fungi isolates evaluated. Under optimum conditions of 28°C, pH 6.0, and glucose and potassium nitrate as carbon and nitrogen sources, C. blakesleeana JSK2 gave 21.09% GLA of total lipids, of which 7.34% was in dry biomass. Culture condition optimisation and genetic manipulation can have beneficial roles in industrial large-scale production (Sukrutha et al., 2014).
Lipid metabolism and GLA production in C. elegans NRRL Y-1392 were investigated with emphasis on the role of the carbon-to-nitrogen (C/N) ratio in enhancing the storage of lipids and biomass formation. Under a high nitrogen level (C/N = 11.0 mol/mol), the fungus had excellent glycerol utilisation and produced high biomass (11.9 g/L), along with the highest GLA and PUFA production (573 mg/L and 224 mg/L, respectively). Under nitrogen starvation (C/N = 110.0 and 220.0 mol/mol), glycerol was utilised at a slower rate, but lipid biosynthesis was increased, with the highest lipid content achieved in C/N = 220.0 mol/mol (59% dry biomass). Temperature was also a determining factor in GLA production since higher temperatures (>25°C) supported growth while diminishing the GLA content. Lowering the temperature to 12°C enhanced GLA accumulation, but it also extended the culture duration and reduced the production rate. The study further identified that pH fluctuations affected fungal metabolism, with nitrogen-excess media causing acid secretion and orange colouration, indicative of an alteration in the fungal metabolic pathway. The research also evaluated the scalability of GLA production in bioreactors, attributing challenges such as biomass heterogeneity and optimisation needs of agitation and aeration parameters. Future research will focus on optimising these parameters to enhance oxygen transfer, nutrient supply, and overall efficiency, paving the way for industrial applications of GLA production (Vasilakis et al., 2024).
The ability of C. elegans NRRL-1393, an oleaginous fungus, to produce unsaturated fatty acids, such as gamma-linolenic acid (GLA) was investigated by Kalampounias et al. (2024a). The fungus was cultivated under nitrogen starvation in shake-flask media containing glycerol or glucose (both at approximately 60 g/L) as the sole carbon source. Both substrates supported equal amounts of biomass, about 13.5 g/L after 330–360 h of growth. However, glycerol synthesised a slightly higher amount of lipids compared to glucose. The latter produced 8.4 g/L of lipids against 7.0 g/L of glucose. The lipids contained significant amounts of unsaturated fatty acids such as oleic acid (C18:1), linoleic acid (C18:2), palmitic acid (C16:0), and GLA (C18:3) that constituted about 9.5% of the total lipids. The lipids were then saponified to yield fatty acid lithium salts (FALS), which exhibited cytotoxicity towards human cancer cell lines, indicating that they have potential as therapeutic drugs. The saponification reaction optimisation for FALS synthesis was also investigated concerning reflux time, hexane volume, and NaCl concentration. All of these parameters were found to influence the fatty acid yield significantly. The study demonstrated that FALS obtained from C. elegans lipids exhibited cytotoxic activity even at low doses, affirming their potential as anticancer agents. The finding suggests that FALS could enhance the efficacy of chemotherapy by inducing oxidative stress in cancer cells while minimising collateral damage to healthy cells. The combined action of oxidative stress induced by GLA and the anticancer effect of lithium holds bright prospects for therapeutic synergies.
Further research into the growth conditions revealed glycerol to be a superior carbon source for lipid storage in C. elegans, and nitrogen starvation to be required for maximising lipid biosynthesis. The 330–360-h growth period allowed maximum biomass and lipid yield, while extended cultivation led to a decrease in GLA productivity. Scale-up operations in bioreactors revealed that optimisation of oxygen supply, pH, and nutrient levels would increase lipid yield even more, making it industrially viable. The research also tested other substrates for fungal growth, including lipid fermentation wastewater (LFW) of M. ramanniana. The novel medium was utilised to carry out green SSF with fungi under nitrogen- and carbon-scarce conditions. The primary carbon source employed was glycerol, which yielded a high level of lipid accumulation coupled with the production of PUFA, particularly GLA. In glycerol-limited cultures, glycerol uptake was rapid, lipid content being at approximately 24% (w/w), and GLA accumulation at levels of up to 430 mg/L. As compared to the above, nitrogen limitation allowed greater lipid material (45% w/w), albeit GLA productivity was lower at 350 mg/L. This observation reflects a general trend in wild-type oleaginous Zygomycetes: when lipid accumulation in dry cell weight exceeds 25% (w/w), the proportion of GLA in total fatty acids tends to decrease. Conversely, under nitrogen-rich or glycerol-limited conditions, where lipid accumulation is restricted, GLA content in total lipids is relatively higher. This inverse relationship between total lipid content and GLA concentration within lipids highlights the importance of tailoring fermentation conditions to balance biomass, total lipid production, and GLA enrichment (Melanouri et al., 2024). This phenomenon may be linked to differential regulation of desaturase enzymes under nutrient-replete vs. starvation conditions.
The study also demonstrated that LFW can serve as an effective substrate for cultivating edible and medicinal mushrooms. The use of LFW promoted fungal biomass development, enhanced the activity of ligninolytic and hydrolytic enzymes, and stimulated exopolysaccharide synthesis. These findings highlight LFW’s potential for application in biotechnological processes aimed at recycling industrial waste and generating value-added byproducts.
In particular, the study explored the utilisation of LFW as maceration water in SSF systems for mushroom cultivation. Although edible and medicinal mushrooms such as Pleurotus ostreatus do not naturally synthesise PUFAs like GLA, ARA, or DHA, LFW proved beneficial in stimulating fungal metabolism and enzymatic activity. Adjusting the carbon-to-nitrogen (C/N) ratio significantly influenced metabolic outcomes: lower C/N ratios enhanced the activity of ligninolytic and hydrolytic enzymes, while higher ratios favoured biomass accumulation. The fruiting of P. ostreatus on LFW-based substrate led to the degradation of total phenolic compounds and a marked increase in laccase activity. Furthermore, high C/N conditions supported carposome formation and contributed to greater biological efficiency and yield.
While these mushrooms are not PUFA-producing organisms, the integration of LFW into their cultivation presents a sustainable and economically viable strategy for managing industrial effluents and producing bioactive fungal compounds. In contrast, microbial lipid biotechnology remains the primary pathway for PUFA production and represents a promising alternative to conventional plant-based oils.
However, the production cost of single-cell oil (SCO) remains a challenge. To mitigate this, researchers have tried to use industrial and agro-industrial waste as low-cost carbon sources. For instance, tomato waste hydrolysate (TWH) and crude glycerol have been discovered to be promising substrates for oleaginous fungi to biosynthesise lipids during nitrogen starvation conditions. Experiments with C. echinulata revealed the effect of limiting nitrogen on lipid content and GLA production. Cultivation of C. echinulata in TWH-supplemented medium resulted in lipid content in biomass of approximately 39.6%, and GLA content was 802 mg/L with 96–120 h of incubation. The study also demonstrated that the use of various carbon sources, such as glucose (30–90 g/L) or glycerol (30–50 g/L), influenced lipid accumulation, highlighting the strain’s versatility in cultivation on diverse substrates for microbial lipid production. Other oleaginous microorganisms, such as Y. lipolytica and M. isabellina, can convert crude glycerol into microbial lipids. Y. lipolytica demonstrated an extremely high lipid content of 51.7% (w/w) in biomass, further confirming its potential for large-scale production of SCO (Fakas et al., 2008b; Fakas et al., 2008a). The accumulation of lipids by the strain has also been studied using various carbon sources, including glucose and glycerol. Hence, it proved to be flexible in terms of strain regarding the application of different substrates for the production of SCO. In a different experiment, T. elegans CCF-1465 was used to grow in submerged shake-flask culture utilising low-cost sugar refinery wastes, i.e., glucose, fructose, and sucrose, as carbon sources. The fungus exhibited high lipid production, with a notable accumulation during the initial phase of sugar consumption. Sugar uptake reduced progressively while lipid accumulation continued to occur. The maximum lipid yield was more than 9 g/L with an optimum 70% (w/w) lipid content in dry biomass, suggesting the suitability of the strain for large-scale lipid production. The lipids produced by the microbes were largely oleic and palmitic acids and seemed suitable as feedstock for biodiesel fuel. GLA was also produced at a level of approximately 510 mg/L, once more highlighting the potential for the utilisation of T. elegans in GLA production (Papanikolaou et al., 2010). Tables 6 and 7 contain additional information regarding the various conditions used in fungi in GLA production.

Table 6. Media engineering and physical conditions in different organisms.
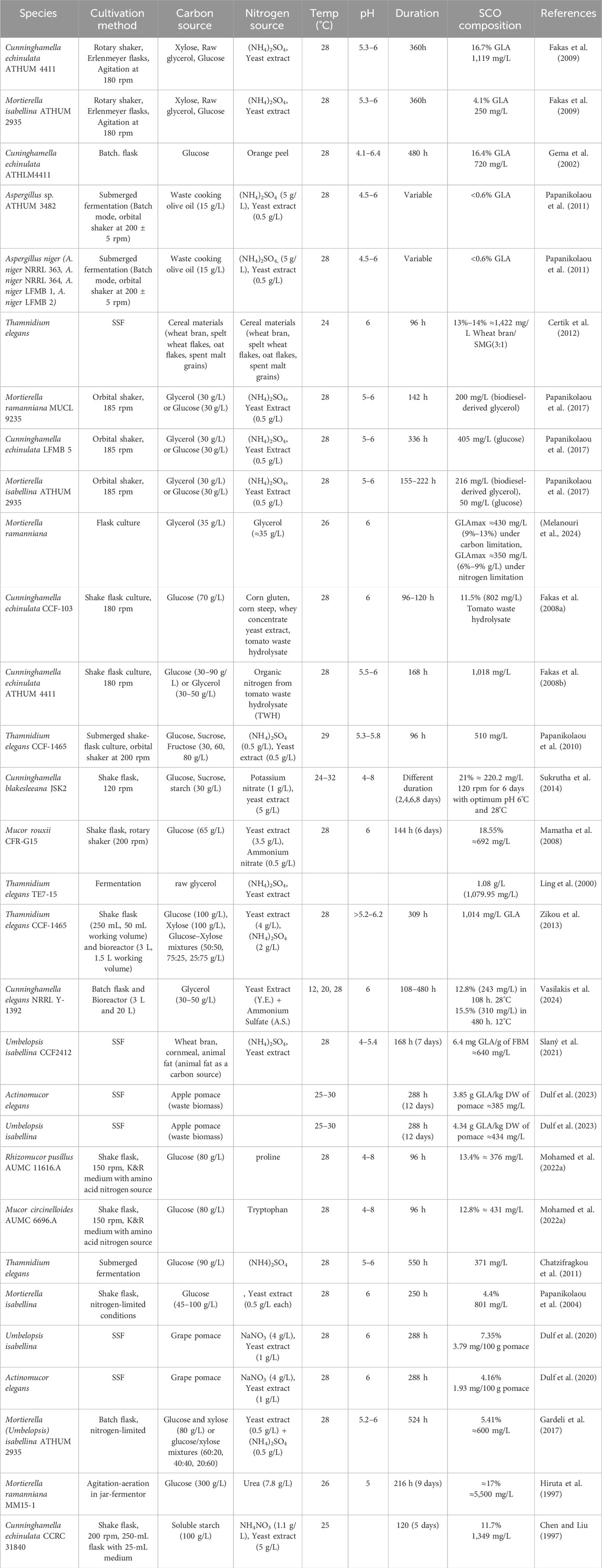
Table 7. GLA production in wild-type and genetically modified fungal systmes
The research on yeast culture engineering for GLA production involves genetic manipulations and optimisation of fermentation processes to enhance GLA yields (Gu et al., 2023). In the study by Khongto et al. (2011), a fed-batch fermentation technique was used to increase the production of GLA in Hansenula polymorpha. After optimising physical-chemical variables during fermentation, the researchers found that the most critical factor influencing GLA output was Dissolved Oxygen Tension (DOT). The ideal parameters for generating GLA were DOT saturation at 28%, yeast extract at 1 g/L, and Pichia trace metals 1 (PTM1) at 3.6 mL/L. GLA content was higher in low quantities of yeast extract and lower in higher concentrations. These optimised conditions improve scalability for industrial production by enhancing yeast growth and lipid accumulation, making large-scale GLA production more feasible (Khongto et al., 2011).
4.3.3 Culture optimisation in algae
The research investigated how Spirulina platensis produces GLA in different stressful conditions. The experiments demonstrated that adding oleic acid and LA at various growth temperatures, along with oil supplements, enhanced the production of fatty acids. Also, when the cultures were exposed to varying light levels, the ratio of GLA to total fatty acids remained constant. The best conditions for producing lipids and GLA in algae depend on various factors such as temperature, light intensity, salinity, and specific oil supplements. Studies suggest controlling these factors is crucial for enhancing GLA production in algae cultures (Wang et al., 2022; Jin et al., 2019). Specific parameters for GLA production in algae, including temperature, light intensity, salinity levels, and oil supplements, have also been identified. Optimising these parameters can improve GLA production in algae, making them suitable for industrial applications (Table 8).
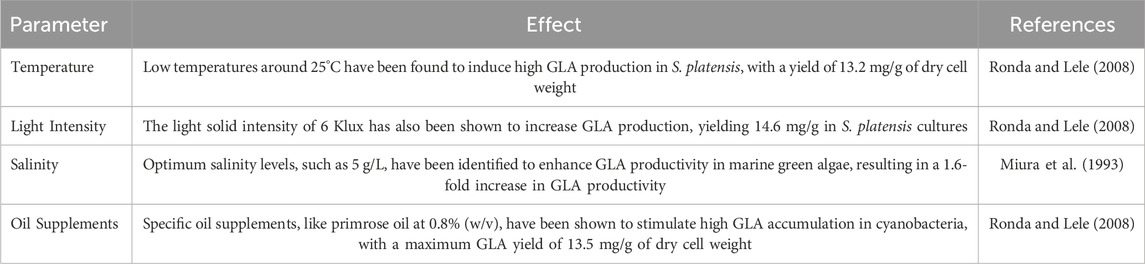
Table 8. Effective parameters in algae culture optimisation.
Spirulina, a commercial cyanobacterium, has been suggested as a more affordable source of GLA, a dietary supplement that is currently expensive. Studies have shown that Spirulina contains PA, saturated fatty acids, LA, and GLA. Spirulina also has the highest percentage of omega-6 fatty acids at 18.8 g. The production of highly pure GLA from microalgal sources could be a promising approach due to their high oil production, fast reproduction, and non-food nature (Choopani et al., 2016).
Higher GLA content can be achieved by manipulating light conditions, such as growing algae under light and then leaving them in the dark. Nitrogen and phosphorus limitation are influential factors for increasing lipid content. Nitrogen increases total fat, while phosphorus limitation significantly affects fat profile, and this type of commercial production of GLA from spirulina can be achieved by shortening the harvest time (6–8 days) and optimising aeration rates. The most straightforward and most efficient method to obtain GLA concentrate is urea accumulation, which does not require any organic solvent except ethanol. The recommended method for lipid extraction from Spirulina on an industrial scale is a three-step process: supercritical extraction, ethanol extraction, and aqueous extraction (Choopani et al., 2016). GLA production is regulated by phosphate and nitrate levels during growth and temperature control. Antarctic cyanobacteria can act as a source of GLA (Verma et al., 2021). Optimising culture conditions (light, salinity and temperature) in algae impacts the cost-effectiveness and sustainability of industrial GLA production.
Algae-based GLA production offers specific advantages over other organisms, like yeast or fungi. Algae are more environmentally sustainable because they can quickly absorb CO2 and produce high amounts of biomass. Additionally, algae do not compete with food crops for arable land, making them an economically viable option for large-scale production. Algae can be cultivated in various environments, including wastewater, reducing production costs and environmental impact.
5 Therapeutic and clinical applications of GLA
The pharmaceutical industry has shown considerable interest in GLA’s therapeutic potential. It has been studied for managing conditions such as diabetic neuropathy, rheumatoid arthritis, and atopic dermatitis (Chang et al., 2017). GLA’s anti-inflammatory and neuroprotective properties have been the basis for developing GLA-based drugs and supplements to treat various health issues (Pagels et al., 2022). GLA is an omega-6 fatty acid with various potential health benefits. GLA has been shown to reverse diabetic neuropathy, a severe complication of diabetes caused by high blood sugar levels that damage the nerves (Pop-Busui et al., 2022). Diabetic animals and humans cannot convert dietary LA to GLA, which is necessary for standard neuronal structure and function (Ramli et al., 2017). Supplementing helps correct biochemical defects and restores GLA metabolite levels, significantly improving clinical symptoms and neurophysiological function in diabetic neuropathy (Shi and Zhao, 2017). In a controlled clinical trial involving 111 patients with mild diabetic neuropathy, supplementation with 480 mg of GLA per day for 1 year resulted in notable improvements in 13 out of 16 evaluated neuropathy parameters. The beneficial effects were observed regardless of factors such as gender, age, type, or duration of diabetes. However, the treatment had a more significant impact on patients whose diabetes was relatively well-controlled (Horrobin, 1992). Another study involving 22 patients with diabetic distal polyneuropathy found that daily intake of 360 mg of GLA for 6 months led to significant improvements in the signs of neuropathy compared to the placebo group (Peng et al., 2022). These findings suggest that supplementation can effectively improve symptoms, including nerve conduction velocity, sensory nerve function, motor activities, tendon reflexes, and sensory functions in the hands and feet of patients with diabetic neuropathy (Hernandez, 2016). At the same time, GLA may control blood lipids by reducing triglyceride levels, total cholesterol, and LDL cholesterol while increasing HDL cholesterol. In a clinical trial involving 12 patients with hyperlipidemia, the daily intake of 3 gr of GLA for 4 months resulted in a 48% reduction in plasma triglyceride levels, a 22% increase in HDL cholesterol levels, and a significant decrease in total cholesterol and LDL cholesterol (Sun et al., 2017). These findings suggest that GLA supplementation has a positive impact on lipid profiles and improves cardiovascular health. However, further research is necessary to fully understand the effects on blood lipids and their clinical significance (Won et al., 2020). Additionally, GLA supplementation has been found to help improve symptoms of irritable bowel syndrome (IBS), such as abdominal pain, bloating, diarrhoea, constipation, and cramping (Keen et al., 1993). GLA achieves its cardiovascular effects predominantly through its anti-inflammatory action, which is mediated by its beta-oxidation to DGLA and subsequently to bioactive eicosanoids such as PGE1 and 15-HETrE (as outlined in Section 4.1). These cause vasodilatation, inhibit platelet aggregation, stabilise blood pressure, and improve lipid profiles, thereby reducing the risk of arteriosclerosis and vascular dysfunction (Nilsen et al., 2021; Bertoni et al., 2023). Due to its immunomodulatory effects, GLA is also being proposed as an adjunctive treatment for inflammatory disorder-related conditions, such as arthritis and atopic dermatitis (Ruiz-López et al., 2012). By enhancing PGE1 production, it may reduce NSAID reliance by reducing joint stiffness and inflammatory signs (Chen et al., 2010; Tański et al., 2022). The processes mentioned above, already explained in the biosynthesis of GLA section, point towards the systemic benefits of GLA-derived metabolites for cardiovascular and inflammatory regulation.
When used in conjunction with NSAIDs, GLA acts as a synergistic agent and prevents common gastrointestinal side effects associated with NSAIDs (Sohail et al., 2023). Additionally, GLA inhibits leukotriene synthesis and cytokine production, demonstrating benefits in diseases such as asthma and acute respiratory distress syndrome (ARDS) (Su et al., 2023). Rheumatoid arthritis (RA) is an autoimmune disease characterised by immune system attacks on the body’s tissues, resulting in joint inflammation and cartilage destruction. Common symptoms include joint pain, swelling, and stiffness, which can lead to long-term damage, chronic pain, and disability (Leventhal et al., 1993). Clinical trials confirm GLA’s immunoregulatory and anti-inflammatory properties, highlighting its synergistic potential when used alongside standard therapies in inflammatory and autoimmune diseases (Zhang C. et al., 2021; Zurier et al., 1996). Another double-blind, placebo-controlled clinical trial involved 56 patients with active rheumatoid arthritis. They received either 8.2 gr of GLA or a Placebo (sunflower oil) daily for 6 months. Following this phase, all patients received treatment for an additional 6 months in a single-masked manner. The therapy led to a substantial decrease in disease activity (Furse et al., 2001). Atopic dermatitis, a chronic inflammatory skin disorder commonly referred to as eczema, is characterised by symptoms such as dry skin, itching, and redness. It is associated with a D6D deficiency, which converts LA to GLA. Recent studies have shown that supplementation with GLA, found in evening primrose oil, can help alleviate skin inflammation and reduce associated symptoms (Horrobin, 2000). In clinical trials, patients who consumed evening primrose oil experienced a decrease in eczema severity scores. Increasing plasma GLA levels can also be a predictive marker for the response to evening primrose oil treatment. Therefore, evening primrose oil is an effective and safe option for managing atopic dermatitis (Williams, 2003). Multiple Sclerosis (MS) is a neurological condition characterised by inflammation and damage to the central nervous system. Some theoretical evidence suggests that GLA may be beneficial in MS. Animal studies have shown that GLA suppresses the activity of the immune system, particularly by inhibiting the proliferation of T lymphocytes, and reduces the severity of MS-like disease (Kousparou et al., 2023). Researchers from New Zealand have observed that MS patients often experience cold hands and feet, which is typically a sign of impaired peripheral blood flow. An uncontrolled study conducted on 16 patients demonstrated that GLA from evening primrose oil improves peripheral circulation and consequently enhances the strength and function of the hands and feet in these patients. The high content of essential fatty acids also reduces inflammation associated with MS-related neurological damage.
Recent studies have significantly enhanced knowledge about GLA and its anticancer effect, especially when synthesised in microorganisms. Conventionally, research has been focused on “pure” GLA of plant or chemical origin. More and more, however, the focus has shifted to microbial GLA, which is considered “impure” because it is part of the overall lipid in microorganisms. Various studies have linked microbial GLA with diverse cancers, viz., leukaemia, glioblastoma, pancreatic cancer, breast cancer, prostate carcinoma, and papillary thyroid carcinoma. Various journal studies have established that microbial GLA induces oxidative stress, triggers apoptosis and ferroptosis, inhibits cell proliferation, and reduces cell migration and viability, thus indicating its therapeutic value against cancer (Kalampounias et al., 2024a; Kalampounias et al., 2024b; Kairemo et al., 1998).
In addition, recent research has also highlighted that GLA can be obtained from fungi, specifically those of the Zygomycota phylum, which can produce high concentrations of GLA when cultivated on low-cost feedstocks such as glycerol. This production is an economic and environmentally friendly alternative to genetically engineered strains for the mass production of GLA. A specific fungus, Cunninghamella elegans NRRL1393, has been demonstrated to produce high concentrations of lipids when cultured on glycerol, about 8.4 g/L. The GLA-rich lipids from this fungus were then transformed into fatty acid lithium salts (FALSs). The FALSs were found to be cytotoxic with an IC50 value of approximately 60 μg/mL, potently inhibiting cell migration, inducing oxidative stress, and leading to cell death in normal human and cancer cell lines. These findings suggest a general, non-specific mechanism of action, and the high level of oxidative stress may enhance the effectiveness of traditional anticancer therapies (Kalampounias et al., 2024b).
Besides fungi, it was also found to be able to produce PUFAs, including GLA, which are linked with anticancer properties. Such fungi, when cultivated on glucose or glycerol as a carbon source, produce lipids that can be converted into FALS. The produced FALSs have been found to exhibit good potential to inhibit the growth and movement of prostate cancer cell lines, DU145 and PC3, making them more susceptible to lipid derivatives. In contrast to the olive oil-derived FALSs when employed as a control, the microbial FALSs drastically inhibited the proliferation and migration capacity of the cancer cells. These results suggest that microbial lipids, particularly those of fungi such as T. elegans and M. alpina, possess potential anticancer properties in suppressing the growth and motility of cancer cells (Kalampounias et al., 2024a).
Additional therapeutic potential was also demonstrated by an investigation of the polyunsaturated lipids produced by Thamnidium elegans grown on raw glycerol and the microalga Nannochloropsis salina. The fungal lipids, which were GLA-rich, consisted mainly of neutral lipids (82%). These lipids were converted into fatty acid potassium salts (FAPS), which potently inhibited the growth of several bacteria, including Gram-negative bacteria such as Serratia sp. and Neisseria gonorrhoeae, as well as Gram-positive bacteria like Staphylococcus aureus and Enterococcus faecalis. Furthermore, the FAPS were highly toxic against the MCF7 cancer cell line at very low concentrations. These findings suggest that FAPS of microbial polyunsaturated lipids could be developed as novel therapeutic medications (Sayegh et al., 2016).
This building evidence attests to the therapeutic potential of microbial GLA, showing not only that it performs well in isolated forms but also in its derivatives from microbial lipids. Fungal lipids as a source of GLA provide a cost-effective, scalable, and viable approach to cancer drug production.
Evening primrose oil, rich in GLA, is a beneficial natural supplement for women’s health. It relieves premenstrual syndrome (PMS) symptoms by increasing the production of anti-inflammatory prostaglandins, thereby easing muscle cramps and bloating. During menopause, it provides a natural alternative to hormone replacement therapy (HRT), effectively reducing hot flashes, mastalgia, inflammation, and other discomforts by balancing eicosanoids. It also helps manage cyclical breast pain with fewer side effects than medications and significantly reduces hot flash frequency and intensity in postmenopausal women, enhancing their quality of life (Hu et al., 2021). Table 9 illustrates the therapeutic use of GLA in humans.
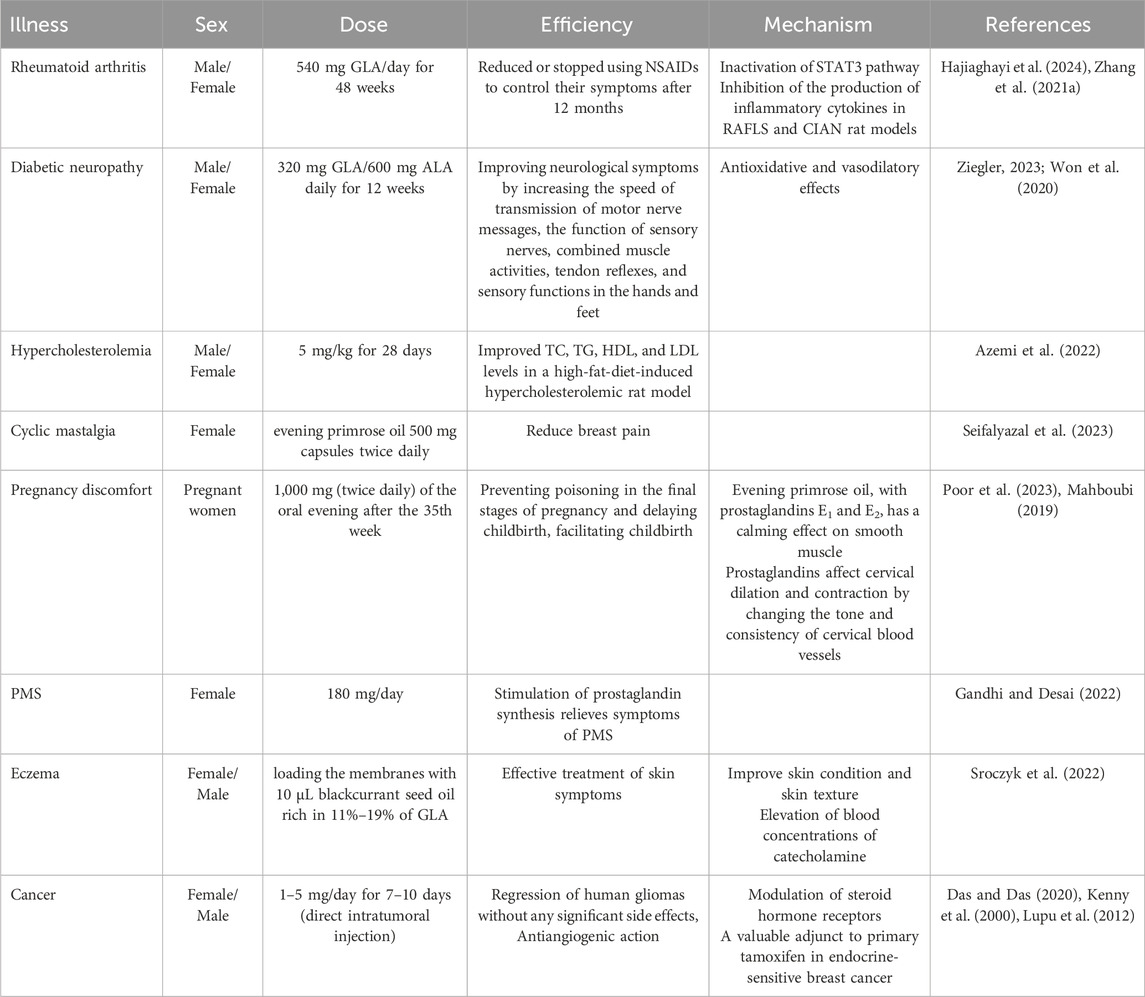
Table 9. Examples of GLA’s therapeutic use in humans.
Current studies suggest that a deficiency in certain fatty acids may make individuals more susceptible to COVID-19. Specifically, low levels of GLA, DGLA, ARA, eicosa pentaenoic acid (EPA), and increased lipid peroxidases and free radicals are associated with severe COVID-19 cases. Higher rates of mortality from COVID-19 are observed in patients with diabetes, hypertension, and coronary artery disease, which may be attributed to their deficiency in GLA, DGLA, ARA, and alpha-linolenic acid (ALA) (Das, 2021). Conversely, the lower prevalence of GLA and milder cases of COVID-19 in children and women may be associated with higher levels of specific fatty acids such as BioActive Lipids (BALs). Children are believed to have a higher capacity for producing fatty acids such as GLA and DGLA, which can be converted into beneficial compounds that combat SARS-CoV-2. Premenopausal women produce significant amounts of LXA4 (lipoxin A4), which confers resistance and reduces the likelihood of severe COVID-19 (Tong et al., 2019). Research indicates that PUFAs, such as GLA and DGLA, act as antibacterial, antifungal, antiviral, and immunomodulatory agents. They can inhibit the entry of coronaviruses and other enveloped viruses into target cells by modulating the fluidity of cell membranes and regulating the ACE-2 receptors responsible for viral entry. PUFAs, including GLA, also modulate the production of M1 and M2 macrophages, reducing the production of pro-inflammatory cytokines like IL-6 and TNF-α. Therefore, they can treat severe cytokine storms in COVID-19 (Matschinske et al., 2020; Baral et al., 2022).
6 Industrial applications
GLA has a range of applications (Figure 5) in the food, cosmetics, livestock, poultry, and pharmaceutical industries (Su et al., 2023). Within the food sector, GLA is valued for its nutritional benefits, particularly its ability to promote heart health and reduce inflammation (Khan et al., 2019). Functional foods enriched with GLA are being developed to enable consumers to incorporate this beneficial ingredient into their everyday diet (Kaur et al., 2014). Methods to enrich food products with GLA and other polyunsaturated fatty acids (PUFAs) include direct addition, microbial supplementation, microencapsulation, or feeding animals PUFA-rich diets (Fan YangYi and Chapkin, 1998). In Japan, Mortierella isabellina is used to produce GLA-rich lipids, which are then utilised as food additives and functional dietary supplements (Demir and Gündes, 2020).
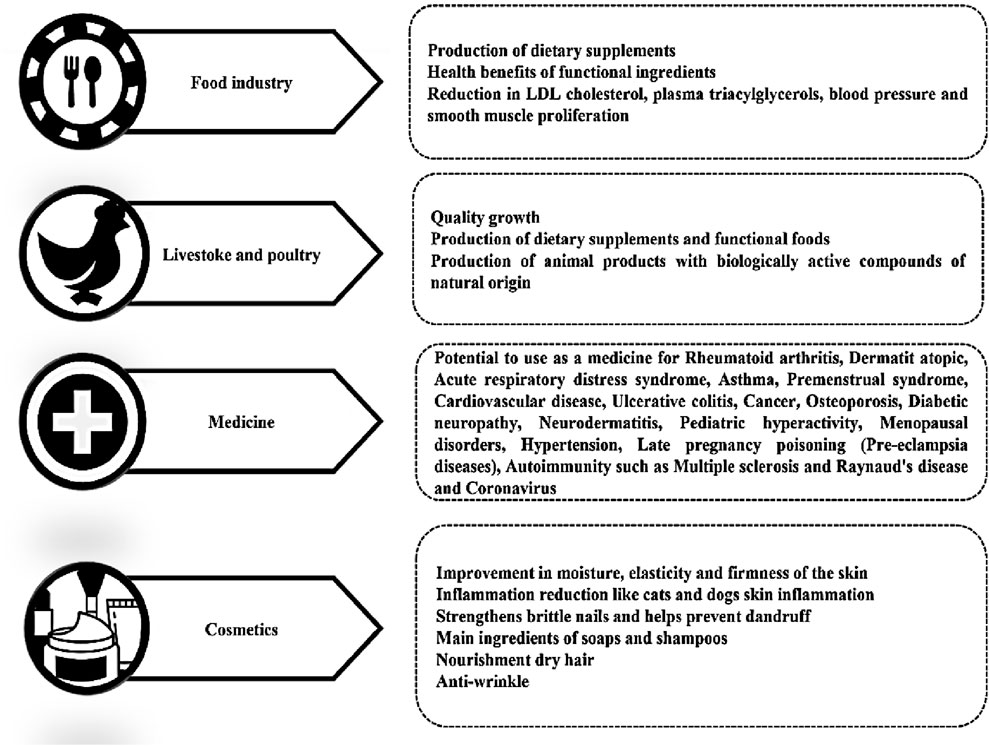
Figure 5. GLA Applications in various industries.
The demand to increase the fatty acid content of animal products, such as meat and eggs, for their improvement in therapeutic and nutritional value for human consumption has accelerated in recent years. Supplementation of animal diets with GLA is one possible method of improving quality of animal products, along with animal health and resistance (Sergeant et al., 2016; Navarette et al., 1992). Due to the limited enzymatic conversion of LA to GLA in animals and humans, dietary supplementation with GLA-enriched feed ingredients has grown as an intervention (Lin et al., 2013).
Various feed supplements containing PUFA, including fish oil, flaxseed, rapeseed, and borage oil, have been tested to determine whether they can raise the levels of GLA in animal tissues. Of these, borage oil has been of therapeutic importance in the management of inflammatory disorders such as ovine chronic interstitial pneumonia and serves as a functional nutrient in equine and ruminant nutrition (Casas-Cardoso et al., 2021). Borage plant supplementation has also been proposed as a stress-mitigating feeding regimen for herbivores during periods of physiological transition (Montaner et al., 2021). Apart from vegetative feedstocks, fungal fermentation through oleaginous Zygomycota fungi is an emerging alternative for the production of GLA-enriched feedstocks, particularly by fermenting crop residues such as grains and glycerol (Chang et al., 2010).
Aside from its applications in animal medicine, GLA has also gained significant interest in the pharmaceutical, cosmetic, and nutraceutical industries. GLA is a highly sought-after ingredient in the cosmetics and personal care industry due to its beneficial effects on skin health. It is known for its moisturising properties, anti-inflammatory effects, and suitability for treating a variety of skin conditions (Gu et al., 2023). GLA-rich oils are commonly incorporated into products such as creams, serums, lotions, soaps, and shampoos to hydrate the skin, delay ageing, and address issues like dry skin, eczema, acne, and skin sensitivity (Kapoor and Nair, 2005). It enhances skin hydration, elasticity, and firmness while combating wrinkles and rough skin texture. Furthermore, its anti-inflammatory properties and ability to regulate sebum production make it effective in treating acne (Ruiz-López et al., 2012). GLA supplements and oils also help alleviate skin problems, allergies, redness, and itching. It plays a crucial role in skin care and promoting healthy skin and hair (Sergeant et al., 2016).
Due to the various applications of GLA, the demand for pure and cost-effective production methods has increased. As a result, various extraction and purification methods have been developed to purify GLA from plant and microbial sources. Traditional solvent extraction methods using hexane or ethanol remain common in the case of seed oils of plant species (e.g., evening primrose, borage), but are normally plagued by poor yield, purity, and environmental issues. To make up for the inadequacies, newer methods such as supercritical CO2 extraction, enzymatic hydrolysis, and urea complexation have been utilized to selectively extract and purify GLA fractions. These technologies are able to increase purity of GLA greater than 80%, and are especially beneficial for downstream applications that require high-grade raw material (Sayegh et al., 2016). Moreover, enzymic interesterification and structured lipid production have been utilized in the incorporation of GLA into omega-3-enriched triglycerides to enhance end product nutritional value and bioavailability. These developments are essential in the design of formulated lipids for clinical nutrition, dermatology, and cardiometabolic therapy. Commerically, the worldwide market of GLA has experienced consistent growth. This increasing trend is the result of increasing consumer awareness of functional fatty acids, along with general industrial requirements for natural, plant, and microbial lipid products. Major players such as Sonova, Efamol, and Nature’s Way have been leaders in the commercialization of GLA-enriched products. Sonova concentrates on high-purity, bioavailable oils for the control of inflammation and immune support. Efamol incorporates GLA with antioxidants and vitamins in its formulations for skin wellness and hormonal stability. Nature’s Way, on the other hand, offers multi-ingredient formulas that are suitable for various nutritional and therapeutic needs, particularly in the women’s health and cardiovascular segments.
Overall, the industrialisation of GLA, including biotechnological production and purification, through to market inclusion, asserts its growing significance as a multifunctional lipid. Continued work to streamline extraction protocols, ensure regulatory compatibility, and integrate GLA into functional ingredients will enhance its position in the global market, opening up opportunities for sustainable and health-enhancing bioactives.
7 Challenges and future directions
Several key challenges persist in hindering the large-scale microbial production of GLA, despite recent advances in metabolic engineering and culture optimisation. These challenges cross biotechnological, genetic, and regulatory fronts. These must be addressed to fully exploit the industrial and therapeutic potential of GLA-producing microbial platforms. A comparative overview of GLA yields in wild-type and genetically modified organisms is presented in Table 10. This table illustrates how significant improvements in GLA yields are achieved through metabolic engineering strategies, including gene overexpression, pathway rerouting, and strain optimisation. These comparisons highlight the inherent significance of genetic modification in overcoming the native constraints of wild-type strains and achieving scalable, commercially viable production outcomes.
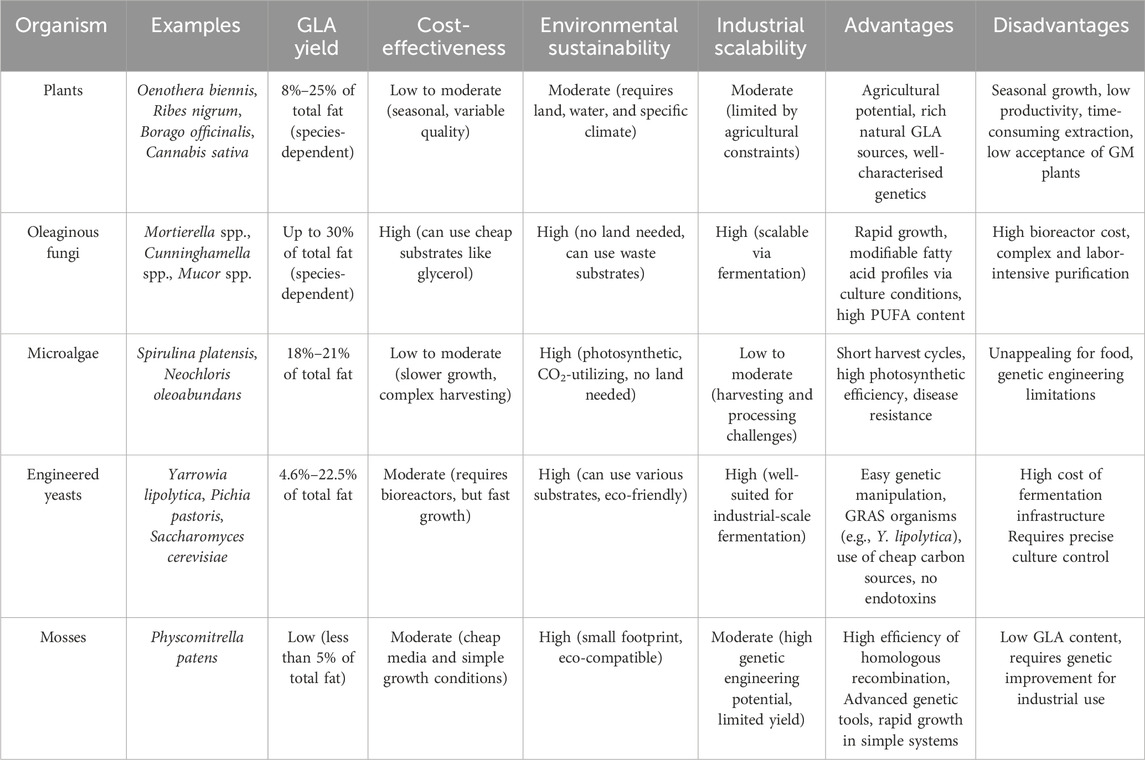
Table 10. Comparison of GLA production in wild-type and genetically engineered organisms.
7.1 Technical and bioprocess bottlenecks
Microorganisms, particularly oleaginous fungi and yeasts, offer a promising and scalable system for GLA production. However, large-scale production of the fatty acid faces several bioprocess, genetic, and engineering challenges that limit the full exploitation of its therapeutic and commercial potential.
The first significant challenge is the limitations of scalability in fermentation processes. For instance, in the 1980s, large-scale cultivation of Mucor circinelloides for GLA-rich single-cell oil (SCO) production was successfully demonstrated in large stirred-tank bioreactors at an industrial scale. This resulted in biomass with 25% total lipids, of which up to 19% was GLA. Yet, difficulties related to bioreactor design, oxygenation, and even nutrient supply in dense cultures led to inefficiencies on an industrial scale (Ratledge, 2013). The other significant challenge in microbial GLA production is the inefficiency of genetic tools. Unlike Y. lipolytica, which facilitates genetic modification with ease, the majority of filamentous fungi lack strong promoters, selectable markers, or stable gene delivery systems. This significantly limits the metabolic engineering of these organisms, resulting in a reliance on model strains (Sun et al., 2017).
From a bioprocess perspective, process maintenance of fermentation and real-time process monitoring optimisation at the industrial level represents another challenge. More specifically, monitoring of parameters such as lipid accumulation, pH, and dissolved oxygen remains technically challenging and typically requires expensive equipment (Belorkar and Jogaiah, 2021). In addition, substrate inconsistency represents another challenge; although the use of inexpensive substrates, such as glycerol or agricultural waste, can yield economic benefits, their unstable composition can undermine reproducibility.
Another key limitation is the formation of unwanted by-products such as ethanol, organic acids, or surplus biomass, which lowers overall GLA production efficiency. Additionally, genetic stability of the engineered strains is a concern because some strains lose their desirable phenotype after several generations, necessitating continuous monitoring and re-engineering (Gu et al., 2023).
Briefly, to surmount these issues, interdisciplinary solutions incorporating genetic engineering, next-generation bioreactor design, the development of real-time monitoring systems, and the use of optimised and homogeneous feedstocks are required. By using technologies such as synthetic biology, intelligent control systems, and GRAS-status microbial platforms, GLA biosynthesis productivity can be transformed.
7.2 Regulatory and social constraints of GMOs
Genetically modified organisms (GMOs) are increasingly employed for GLA production due to their capacity to express desaturase genes, optimise metabolic fluxes, and improve lipid yields. For instance, transgenic Yarrowia lipolytica strains have reached a maximum of 22.5% GLA content in total fatty acids (Sun et al., 2017), while transgenic safflower seeds, which were transformed with delta-6 desaturase, have attained over 40% GLA content (Nykiforuk et al., 2012). However, despite such advancements, the commercial application of GMOs is controlled rigorously.
Other nations, such as the European Union, Japan, and those in South America, have strict regulatory frameworks that include severe biosafety, allergenicity, and gene flow tests. These long approval processes result in marketplace delays and increased production costs. Furthermore, the public’s lack of confidence in products produced from GMOs—particularly in the nutraceutical and food sectors—continues to be a deterrent, with labelling regulations and consumer sentiment often discouraging market adoption (Huesing et al., 2016).
7.3 Agronomic challenges in plant cultivation
Borage, one of the most traditional sources of GLA, is also affected by several agronomic challenges that keep it small-scale as well as economically unviable (Montaner et al., 2021). Although over 95% of global borage production is concentrated in five countries—the United Kingdom, Canada, the Netherlands, New Zealand, and Poland—the crop is extremely labour-intensive to cultivate. Additionally, seed yields are variable, primarily due to premature seed fall and non-synchronous flowering, ranging from 100 to 500 kg/ha, depending on the location (El Hafid et al., 2002). Another major challenge is the poor competitiveness of borage with weeds, especially dicotyledonous weeds. This is exacerbated by a lack of herbicide-resistant crop varieties and the phytotoxicity of current herbicides, such as yellowing and stunting under changing environmental conditions. These constraints pose a major impediment to achieving uniform yields. Despite all this, recent progress in plant biotechnology has come as some relief. Tissue culture, somatic embryogenesis, and liquid-based systems of in vitro propagation have been explored to produce GLA from borage somatic embryos. These present novel avenues for biotechnological production of GLA, particularly by alleviating field-level variability and genetic control over oil traits.
7.4 Future directions and opportunities
To address these limitations and improve GLA productivity, the following strategies should be undertaken: integrating synthetic biology, integrating synbiotics such as CRISPR/Cas9, utilising modular cloning vectors, and employing biosensors to control pathways, which can significantly enhance production (Kan et al., 2022). Host Optimisation: the construction of GRAS-status hosts, such as Y. lipolytica, P. patens, or algae with improved stress tolerance and enhanced precursor flux, is a viable option (Gu et al., 2023). Fermentation process development, which involves improving scalability and consistency, can be achieved through the development of advanced fed-batch or continuous fermentation systems with real-time control and sensor-based monitoring (Belorkar and Jogaiah, 2021). Regulatory harmonisation and effective communication of GMOs at the global level, along with improved public communication, are necessary to ensure consumer confidence and streamline approval processes. Overcoming these technical and regulatory challenges is key to unlocking the full biotechnological and commercial value of microbial GLA production. Multidisciplinary solutions that bring together strain engineering, process optimisation, and regulatory change must be mobilised to bring sustainable, high-productive, and socially acceptable GLA biofactories to market.
8 Conclusion and discussion
Gamma-linolenic acid (GLA) is a valuable dietary polyunsaturated fatty acid with demonstrated health virtues in humans, particularly for the management of inflammation, cardiovascular disease, and metabolic disorders. Due to the continuously increasing demand worldwide for GLA, scientists have been attempting to discover novel methods for enhancing production, maximising extraction methods, and determining alternative sources apart from traditional plant oils such as evening primrose and borage. One of the most promising areas for the production of GLA is microbial fermentation using oleaginous fungi such as Mortierella ramanniana, Cunninghamella elegans, Cunninghamella echinulata, and T. elegans. These accumulate lipids in high amounts, and some strains can produce over 1,000 mg/L of GLA when subjected to optimised fermentation conditions. Among the fermentation processes, submerged fermentation (SmF) is the most effective microbial system for GLA production, as it allows controlled conditions of aeration, pH, and nutrient feeding. The use of glycerol as a carbon source in the glycolytic pathway under SmF has resulted in maximised lipid production. Additionally, agricultural and food processing industry wastes are being studied as low-cost and environmentally friendly substrates for fermentation. Next to oleaginous fungi, microalgae like Spirulina platensis have been recognised as sustainable natural sources of GLA, although less productive than fungi and consequently less efficient for industrial cultivation. Oleaginous yeasts like Y. lipolytica have also been recognised as novel platforms for GLA production. The organism enjoys faster growth rates compared to filamentous fungi and can be easily genetically transformed for enhanced lipid accumulation. As a result of metabolic engineering, Y. lipolytica can produce more GLA today and is a potential candidate for industrial fermentation. In parallel, genetic engineering plays a crucial role in enhancing the yield of GLA. Researchers have managed to increase yield by overexpressing key enzymes such as delta-6 desaturase, which is responsible for catalysing the transformation of LA into GLA. For example, GLA content has been attained at 40% of total lipids in genetically engineered M. alpina. Similarly, C. echinulata strains have been optimised to produce more lipid yield and enzymatic activity. Besides microorganisms, some plants have also been genetically transformed to synthesise GLA. The D6D gene has been expressed in plants such as tomato, tobacco, and canola. High GLA-content seed oil transgenic lines, such as safflower (Carthamus tinctorius, up to 40% GLA) were developed.
A second significant field in GLA manufacturing involves the maximisation of extraction and purification processes. Reactions such as hydrolysis and urea complexation can increase the concentration of GLA in oils by up to 40%–80%. Enzymatic and chemical modifications, such as interesterification, are also used to alter the fatty acid composition of GLA-laden oils according to their intended end-use functions. Another method is the manufacturing of structured lipids by adding GLA to omega-3-rich triacylglycerols, thereby upgrading the nutritional and functional qualities of these oils. As the commercialisation of GLA continues to rise, rigorous quality control and safety regulations have become mandatory. Sophisticated spectroscopic, chromatographic, and molecular techniques such as PCR are employed to analyse the safety and purity of GLA-rich oils. New high-throughput screening technologies, automation of bioprocessing, and lipidomics tools will significantly enhance both the efficiency of production and the quality of the product.
In the years to come, the future of GLA production will depend on the convergence of new biotechnological tools, optimisation of industrial processes, and adoption of economically beneficial and environmentally friendly production systems. The principal research fields include optimising fermentation conditions, breakthroughs in genetic engineering, and improving microbial strains through synthetic biology. Emerging technologies such as CRISPR gene editing, novel bioreactor design, and high-accuracy lipidomics profiling will transform the efficacy of GLA production and recovery. Automation systems for high-throughput screening are important in identifying top-performing strains for industrial applications, esp. in engineered fungi and yeast. Furthermore, the combination of microbial fermentation, plant biotechnology, and advanced extraction and purification technology will reduce production costs and increase product purity. This interdisciplinarity option not only ensures sustainable and scalable GLA availability but also opens new frontiers for its clinical, nutritional, and industrial applications. Pursuing these directions, GLA can become a key compound in future worldwide efforts at health, prevention, and nutrition.
Author contributions
ML: Investigation, Writing – original draft. FJ: Formal Analysis, Investigation, Writing – review and editing. HH: Formal Analysis, Investigation, Writing – review and editing. DH: Writing – review and editing. PI: Resources, Writing – review and editing. NF: Conceptualization, Project administration, Writing – review and editing.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Correction note
This article has been corrected with minor changes. These changes do not impact the scientific content of the article.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
ALA, Alpha-linolenic acid; EPA, Eicosapentaenoic acid; MS, Multiple Sclerosis; NSAIDs, Non-steroidal anti-inflammatory drugs; FAPS, Fatty acid potassium salts; FALS, Fatty acid lithium salts; D12D, Delta-12 desaturase; D5D, Delta-5 desaturase; D6D, Delta-6 desaturase; PUFA, Polyunsaturated fatty acid; DCW, Dry cell weight; OA, Oleic acid; ARA, Arachidonic acid; DGLA, Dihomo-gamma-linolenic acid; LA, Linoleic acid; GLA, Gamma-linolenic acid; SSF, Solid-state fermentation; SMF, Submerged fermentation.
References
Abbadi, A., Domergue, F., Bauer, J., Napier, J. A., Welti, R., Zahringer, U., et al. (2004). Biosynthesis of very-long-chain polyunsaturated fatty acids in transgenic oilseeds: constraints on their accumulationw⃞. Plant Cell 16 (10), 2734–2748. doi:10.1105/tpc.104.026070
Alonso-Esteban, J. I., González-Fernández, M. J., Fabrikov, D., Torija-Isasa, E., Sánchez-Mata, M. D. C., and Guil-Guerrero, J. L. (2020). Hemp (Cannabis sativa L.) varieties: fatty acid profiles and upgrading of γ-linolenic acid–containing hemp seed oils. Eur. J. Lipid Sci. Technol. 122 (7), 1900445. doi:10.1002/ejlt.201900445
Amano, N. (1992). Chemotaxonomic significance of fatty acid composition in the genus mortierella (zygomycetes, mortierellaceae). Mycotaxon 45, 257–265.
Azemi, N. A., Azemi, A. K., Abu-Bakar, L., Sevakumaran, V., Muhammad, T. S. T., and Ismail, N. (2022). Effect of linoleic acid on cholesterol levels in a high-fat diet-induced hypercholesterolemia rat model. Metabolites 13 (1), 53. doi:10.3390/metabo13010053
Baral, P. K., Amin, M. T., Rashid, M. M. O., and Hossain, M. S. (2022). Assessment of polyunsaturated fatty acids on COVID-19-associated risk reduction. Rev. Bras. Farmacogn. 32, 50–64. doi:10.1007/s43450-021-00213-x
Barry, A. R., and Dixon, D. L. (2021). Omega-3 fatty acids for the prevention of atherosclerotic cardiovascular disease. Pharmacotherapy 41 (12), 1056–1065. doi:10.1002/phar.2615
Beike, A. K., Jaeger, C., Zink, F., Decker, E. L., and Reski, R. (2014). High contents of very long-chain polyunsaturated fatty acids in different moss species. Plant Cell Rep. 33, 245–254. doi:10.1007/s00299-013-1525-z
Belch, J. J., Ansell, D., Madhok, R., O'dowd, A., and Sturrock, R. D. (1988). Effects of altering dietary essential fatty acids on requirements for non-steroidal anti-inflammatory drugs in patients with rheumatoid arthritis: a double blind placebo controlled study. Ann. Rheum. Dis. 47 (2), 96–104. doi:10.1136/ard.47.2.96
Belorkar, S. A., and Jogaiah, S. (2021). Protocols appl. Enzymol. Academic Press. doi:10.1016/B978-0-12-823859-8.00024-1
Bertoni, C., Abodi, M., D’Oria, V., Milani, G. P., Agostoni, C., and Mazzocchi, A. (2023). Alpha-linolenic acid and cardiovascular events: a narrative review. Int. J. Mol. Sci. 24 (18), 14319. doi:10.3390/ijms241814319
Bleakley, S., and Hayes, M. (2017). Algal proteins: extraction, application, and challenges concerning production. Foods 6 (5), 33. doi:10.3390/foods6050033
Buist, P. H. (2010). “1.02 - unsaturated fatty acids,” in Compreh. Nat. Prod. II. Editors H.-W. LIU, and L. MANDER (Oxford: Elsevier). doi:10.1016/b978-008045382-8.00720-6
Campos, M. L., Prado, G. S., Dos Santos, V. O., Nascimento, L. C., Dohms, S. M., da Cunha, N. B., et al. (2020). Mosses: versatile plants for biotechnological applications. Biotechnol. Adv. 41, 107533. doi:10.1016/j.biotechadv.2020.107533
Casas-Cardoso, L., Mantell, C., Obregón, S., Cejudo-Bastante, C., Alonso-Moraga, Á., de la Ossa, E. J. M., et al. (2021). Health-promoting properties of borage seed oil fractionated by supercritical carbon dioxide extraction. Foods 10 (10), 2471. doi:10.3390/foods10102471
Čertík, M., Adamechová, Z., and Laoteng, K. (2012). Microbial production of γ-linolenic acid: submerged versus solid-state fermentations. Food Sci. Biotechnol. 21, 921–926. doi:10.1007/s10068-012-0121-2
Čertik, M., Slavikova, L., Masrnova, S., and Šjbidor, J. (2006). Enhancement of nutritional value of cereals with γ-linolenic acid by fungal solid-state fermentations. Food Technol. Biotechnol. 44.
Chang, C. S., Sun, H. L., Lii, C. K., Chen, H. W., Chen, P. Y., and Liu, K. L. (2010). Gamma-linolenic acid inhibits inflammatory responses by regulating NF-κB and AP-1 activation in lipopolysaccharide-induced RAW 264.7 macrophages. Inflammation 33, 46–57. doi:10.1007/s10753-009-9157-8
Chatzifragkou, A., Makri, A., Belka, A., Bellou, S., Mavrou, M., Mastoridou, M., et al. (2011). Biotechnological conversions of biodiesel derived waste glycerol by yeast and fungal species. Energy 36 (2), 1097–1108. doi:10.1016/j.energy.2010.11.040
Chaudhary, R., Nawaz, A., Fouillaud, M., Dufossé, L., Haq, I. U., and Mukhtar, H. (2024). Microbial cell factories: biodiversity, pathway construction, robustness, and industrial applicability. Microbiol. Res. 15 (1), 247–272. doi:10.3390/microbiolres15010018
Chen, H. C., and Liu, T. M. (1997). Inoculum effects on the production of γ-linolenic acid by the shake culture of Cunninghamella echinulata CCRC 31840. Enzyme Microb. Technol. 21 (2), 137–142. doi:10.1016/S0141-0229(96)00262-1
Chen, L., Deng, H., Cui, H., Fang, J., Zuo, Z., Deng, J., et al. (2017). Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 9 (6), 7204–7218. doi:10.18632/oncotarget.23208
Chodok, P. (2022). Response surface modeling and optimization of media nutrients for enhanced production of y-linolenic acid in the moss Physcomitrella Patens using response surface methodology. RMUTP Res. J. Sci. Technol. 15 (1), 51–65. doi:10.14456/jrmutp.2021.5
Chodok, P., Cove, D. J., Quatrano, R. S., Kanjana-Opas, A., and Kaewsuwan, S. (2012). Metabolic engineering and oil supplementation of physcomitrella Patens for activation of C 22 polyunsaturated fatty acid production. J. Am. Oil Chem. Soc. 89, 465–476. doi:10.1007/s11746-011-1927-4
Chodok, P., Eiamsa-ard, P., Cove, D. J., Quatrano, R. S., and Kaewsuwan, S. (2013). Identification and functional characterization of two Δ12-fatty acid desaturases associated with essential linoleic acid biosynthesis in physcomitrella Patens. J. Ind. Microbiol. Biotechnol. 40 (8), 901–913. doi:10.1007/s10295-013-1285-3
Chodok, P., Kanjana-Opas, A., and Kaewsuwan, S. (2010). The plackett–burman design for evaluating the production of polyunsaturated fatty acids by physcomitrella Patens. J. Am. Oil Chem. Soc. 87, 521–529. doi:10.1007/s11746-009-1532-y
Choopani, A., Poorsoltan, M., Fazilati, M., Latifi, A. M., and Salavati, H. (2016). Spirulina: a source of gamma-linoleic acid and its applications. J. Appl. Biotechnol. Rep. 3 (4), 483–488.
Chutrakul, C., Jeennor, S., Panchanawaporn, S., Cheawchanlertfa, P., Suttiwattanakul, S., Veerana, M., et al. (2016). Metabolic engineering of long chain-polyunsaturated fatty acid biosynthetic pathway in oleaginous fungus for dihomo-gamma linolenic acid production. J. Biotechnol. 218, 85–93. doi:10.1016/j.jbiotec.2015.12.003
Cunnane, S. C. (2003). Problems with essential fatty acids: time for a new paradigm? Prog. Lipid Res. 42 (6), 544–568. doi:10.1016/s0163-7827(03)00038-9
Czumaj, A., and Śledziński, T. (2020). Biological role of unsaturated fatty acid desaturases in health and disease. Nutrients 12 (2), 356. doi:10.3390/nu12020356
Das, U. N. (2021). Bioactive lipid-based therapeutic approach to COVID-19 and other similar infections. Arch. Med. Sci. AMS 19 (5), 1327–1359. doi:10.5114/aoms/135703
Das, U. N., and Das, U. N. (2020). Molecular mechanism of anti-cancer action of PUFAs with particular reference to GLA in glioma. Mol. Biochem. Asp. Cancer, 181–206. doi:10.1007/978-1-0716-0741-1_5
Decker, E. L., Gorr, G., and Reski, R. (2003). Moss–an innovative tool for protein production. BIOforum Eur. 2, 96–97.
Decker, E. L., and Reski, R. (2012). Glycoprotein production in moss bioreactors. Plant Cell Rep. 31 (3), 453–460. doi:10.1007/s00299-011-1152-5
Demir, M., and Gündes, A. G. (2020). Single-cell oil production by Mortierella isabellina DSM 1414 using different sugars as carbon source. Biotechnol. Prog. 36 (6), e3050. doi:10.1002/btpr.3050
Dulf, F. V., Vodnar, D. C., and Dulf, E. H. (2023). Solid-state fermentation with Zygomycetes fungi as a tool for biofortification of Apple pomace with γ-linolenic acid, carotenoid pigments and phenolic antioxidants. Food Res. Int. 173, 113448. doi:10.1016/j.foodres.2023.113448
Dulf, F. V., Vodnar, D. C., Toşa, M. I., and Dulf, E. H. (2020). Simultaneous enrichment of grape pomace with γ-linolenic acid and carotenoids by solid-state fermentation with Zygomycetes fungi and antioxidant potential of the bioprocessed substrates. Food Chem. 310, 125927. doi:10.1016/j.foodchem.2019.125927
Dyal, S. D., and Narine, S. S. (2005). Implications for the use of mortierella fungi in the industrial production of essential fatty acids. Food Res. Int. 38 (4), 445–467. doi:10.1016/j.foodres.2004.11.002
Eckert, H., LaVallee, B., Schweiger, B. J., Kinney, A. J., Cahoon, E. B., and Clemente, T. (2006). Co-expression of the borage Δ 6 desaturase and the arabidopsis Δ 15 desaturase results in high accumulation of stearidonic acid in the seeds of transgenic soybean. Planta 224, 1050–1057. doi:10.1007/s00425-006-0291-3
El Hafid, R., Blade, S. F., and Hoyano, Y. (2002). Seeding date and nitrogen fertilization effects on the performance of borage (Borago officinalis L.). Industrial Crops Prod. 16 (3), 193–199. doi:10.1016/S0926-6690(02)00047-X
Engler, M. M. (1993). Comparative study of diets enriched with evening primrose, Black currant, borage or fungal oils on blood pressure and pressor responses in spontaneously hypertensive rats. Prostagl. Leukot. Essent. Fat. Acids 49 (4), 809–814. doi:10.1016/0952-3278(93)90030-Z
Fakas, S., Čertik, M., Papanikolaou, S., Aggelis, G., Komaitis, M., and Galiotou-Panayotou, M. (2008a). γ-Linolenic acid production by Cunninghamella echinulata growing on complex organic nitrogen sources. Bioresour. Technol. 99 (13), 5986–5990. doi:10.1016/j.biortech.2007.10.016
Fakas, S., Papanikolaou, S., Batsos, A., Galiotou-Panayotou, M., Mallouchos, A., and Aggelis, G. (2009). Evaluating renewable carbon sources as substrates for single cell oil production by Cunninghamella echinulata and Mortierella isabellina. Biomass Bioenergy 33 (4), 573–580. doi:10.1016/j.biombioe.2008.09.006
Fakas, S., Papanikolaou, S., Galiotou-Panayotou, M., Komaitis, M., and Aggelis, G. (2008b). Organic nitrogen of tomato waste hydrolysate enhances glucose uptake and lipid accumulation in Cunninghamella echinulata. J. Appl. Microbiol. 105 (4), 1062–1070. doi:10.1111/j.1365-2672.2008.03839.x
Fan YangYi, F. Y., and Chapkin, R. S. (1998). Importance of dietary γ-linolenic acid in human health and nutrition. J. Nutr. 128 (9), 1411–1414. doi:10.1093/jn/128.9.1411
Firoozabad, M. S. M., and Akhbariyoon, H. (2021). An integrated approach to produce a recombinant yeast Pichia pastoris for GLA production. doi:10.21203/rs.3.rs-706359/v1
Flider, F. J. (2013). Development and commercialization of GLA safflower oil. Lipid Technol. 25 (10), 227–229. doi:10.1002/lite.201300302
Furse, R. K., Rossetti, R. G., and Zurier, R. B. (2001). Gamma-linolenic acid, an unsaturated fatty acid with anti-inflammatory properties, blocks amplification of IL-1β production by human monocytes. J. Immunol. 167 (1), 490–496. doi:10.4049/jimmunol.167.1.490
Gammone, M. A., Riccioni, G., Parrinello, G., and D’Orazio, N. (2019). Omega-3 polyunsaturated fatty acids: benefits and endpoints in sport. Nutrients 11 (1), 46. doi:10.3390/nu11010046
Gandhi, A. B., and Desai, A. (2022). Drug corner pharmacological basis of evening primrose oil in premenstrual Syndrome—An evidenced based approach. J. Indian Med. Assoc. 120 (7), 75–79.
Ganjali Dashti, M., Abdeshahian, P., Wan Yusoff, W. M., Kalil, M. S., and Abdul Hamid, A. (2014). Repeated batch fermentation biotechnology for the biosynthesis of lipid and gamma-linolenic acid by Cunninghamella bainieri 2A1. Biol. Med. Res. Int. 2014 (1), 831783. doi:10.1155/2014/831783
Gardeli, C., Athenaki, M., Xenopoulos, E., Mallouchos, A. K. A. A., Koutinas, A. A., Aggelis, G., et al. (2017). Lipid production and characterization by mortierella (umbelopsis) isabellina cultivated on lignocellulosic sugars. J. Appl. Microbiol. 123 (6), 1461–1477. doi:10.1111/jam.13587
Gema, H., Kavadia, A., Dimou, D., Tsagou, V., Komaitis, M., and Aggelis, G. (2002). Production of γ-linolenic acid by Cunninghamella echinulata cultivated on glucose and Orange peel. Appl. Microbiol. Biotechnol. 58, 303–307. doi:10.1007/s00253-001-0910-7
Grama, S. B., Liu, Z., and Li, J. (2022). Emerging trends in genetic engineering of microalgae for commercial applications. Mar. Drugs 20 (5), 285. doi:10.3390/md20050285
Gu, X., Huang, L., and Lian, J. (2023). Biomanufacturing of γ-linolenic acid-enriched galactosyldiacylglycerols: challenges in microalgae and potential in oleaginous yeasts. Synth. Syst. Biotechnol. 8 (3), 469–478. doi:10.1016/j.synbio.2023.06.007
Gupta, A., Kang, K., Pathania, R., Saxton, L., Saucedo, B., Malik, A., et al. (2024). Harnessing genetic engineering to drive economic bioproduct production in algae. Front. Bioeng. Biotechnol. 12, 1350722. doi:10.3389/fbioe.2024.1350722
Hajiaghayi, M., Dolati-Somarin, A. H., and Gholizadeh, F. (2024). The efficacy of evening primrose in rheumatoid arthritis: a systematic review. Res. J. Pharmacogn. 11 (2), 71–79. doi:10.22127/rjp.2023.414090.2212
Hansson, L., and Dostálek, M. (1988). Effect of culture conditions on mycelial growth and production of γ-linolenic acid by the fungus Mortierella ramanniana. Appl. Microbiol. Biotechnol. 28, 240–246. doi:10.1007/BF00250448
Hernandez, E. M. (2016). Specialty oils: functional and nutraceutical properties. Funct. Diet. Lipids, 69–101. doi:10.1016/B978-1-78242-247-1.00004-1
Hiruta, O., Yamamura, K., Takebe, H., Futamura, T., Iinuma, K., and Tanaka, H. (1997). Application of maxblend fermentor® for microbial processes. J. Ferment. Bioeng. 83 (1), 79–86. doi:10.1016/S0922-338X(97)87331-X
Horrobin, D. F. (2000). Essential fatty acid metabolism and its modification in atopic eczema. Am. J. Clin. Nutr. 71 (1), 367S–372S. doi:10.1093/ajcn/71.1.367S
Horrobin, D. F. (1992). The use of gamma-linolenic acid in diabetic neuropathy. Agents Actions Suppl. 37, 120–144. doi:10.1007/978-3-0348-7262-1_18
Hu, D., Cui, Y., and Zhang, J. (2021). Nervonic acid ameliorates motor disorder in mice with Parkinson’s disease. Neurochem. J. 15, 317–324. doi:10.1134/S1819712421030065
Huang, S., Liu, R., Niu, Y., and Hasi, A. (2010). Cloning and functional characterization of a fatty acid Δ6-desaturase from oenothera Biennis: production of γ-linolenic acid by heterologous expression in Saccharomyces cerevisiae. Russ. J. Plant Physiol. 57, 568–573. doi:10.1134/S1021443710040163
Huang, X. F., Liu, J. N., Lu, L. J., Peng, K. M., Yang, G. X., and Liu, J. (2016). Culture strategies for lipid production using acetic acid as sole carbon source by Rhodosporidium toruloides. Bioresour. Technol. 206, 141–149. doi:10.1016/j.biortech.2016.01.073
Huesing, J. E., Andres, D., Braverman, M. P., Burns, A., Felsot, A. S., Harrigan, G. G., et al. (2016). Global adoption of genetically modified (GM) crops: challenges for the public sector. J. Agric. food Chem. 64 (2), 394–402. doi:10.1021/acs.jafc.5b05116
Ikeh, M. A., Fidel Jr, P. L., and Noverr, M. C. (2018). Prostaglandin E2 receptor antagonist with antimicrobial activity against methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 62 (3), e01920-17–1128. doi:10.1128/aac.01920-17
Jang, H., and Park, K. (2020). Omega-3 and omega-6 polyunsaturated fatty acids and metabolic syndrome: a systematic review and meta-analysis. Clin. Nutr. 39 (3), 765–773. doi:10.1016/j.clnu.2019.03.032
Jeennor, S., Anantayanon, J., Panchanawaporn, S., Chutrakul, C., and Laoteng, K. (2019). Morphologically engineered strain of Aspergillus oryzae as a cell chassis for production development of functional lipids. Gene 718, 144073. doi:10.1016/j.gene.2019.144073
Jin, M., Zhai, R., Xu, Z., and Wen, Z. (2019). Production of high-value polyunsaturated fatty acids using microbial cultures. Microb. Lipid Prod. Methods Protoc. 1995, 229–248. doi:10.1007/978-1-4939-9484-7_15
Kaewsuwan, S., Bunyapraphatsara, N., Cove, D. J., Quatrano, R. S., and Chodok, P. (2010). High level production of adrenic acid in physcomitrella Patens using the algae pavlova sp. Δ5-elongase gene. Bioresour. Technol. 101 (11), 4081–4088. doi:10.1016/j.biortech.2009.12.138
Kairemo, K. J., Jekunen, A. P., Korppi-Tommola, E. T., and Pyrhönen, S. O. (1998). The effect of lithium γ-linolenate therapy of pancreatic cancer on perfusion in liver and pancreatic tissues. Pancreas 16 (1), 105–106. doi:10.1097/00006676-199801000-00017
Kalampounias, G., Dritsas, P., Karayannis, D., Androutsopoulou, T., Gardeli, C., Papanikolaou, S., et al. (2024a). Polyunsaturated fatty acids from Thamnidium elegans and Mortierella alpina suppress prostate cancer cell proliferation and migration. Fermentation 10 (11), 578. doi:10.3390/fermentation10110578
Kalampounias, G., Gardeli, C., Alexis, S., Anagnostopoulou, E., Androutsopoulou, T., Dritsas, P., et al. (2024b). Poly-unsaturated fatty acids (PUFAs) from Cunninghamella elegans grown on glycerol induce cell death and increase intracellular reactive oxygen species. J. Fungi 10 (2), 130. doi:10.3390/jof10020130
Kan, M., Huang, T., and Zhao, P. (2022). Artificial chromosome technology and its potential application in plants. Front. Plant Sci. 13, 970943. doi:10.3389/fpls.2022.970943
Kapoor, R., and Huang, Y. S. (2006). Gamma linolenic acid: an antiinflammatory omega-6 fatty acid. Curr. Pharm. Biotechnol. 7 (6), 531–534. doi:10.2174/138920106779116874
Kapoor, R., and Nair, H. (2005). “Gamma linolenic acid: sources and functions,” in Bailey's industrial oil and fat products, 1–45. doi:10.1002/047167849X.bio026.pub2
Kaur, N., Chugh, V., and Gupta, A. K. (2014). Essential fatty acids as functional components of foods-a review. J. Food Sci. Technol. 51, 2289–2303. doi:10.1007/s13197-012-0677-0
Kawashima, A., Shimada, Y., Nagao, T., Ohara, A., Matsuhisa, T., Sugihara, A., et al. (2002). Production of structured TAG rich in 1, 3-dicapryloyl-2-γ-linolenoyl glycerol from borage oil. J. Am. Oil Chem. Soc. 79, 871–877. doi:10.1007/s11746-002-0572-9
Keen, H., Payan, J., Allawi, J., Walker, J., Jamal, G. A., Weir, A. I., et al. (1993). Treatment of diabetic neuropathy with γ-linolenic acid. Diabetes Care 16 (1), 8–15. doi:10.2337/diacare.16.1.8
Kenny, F. S., Pinder, S. E., Ellis, I. O., Gee, J. M., Nicholson, R. I., Bryce, R. P., et al. (2000). Gamma linolenic acid with tamoxifen as primary therapy in breast cancer. Int. J. Cancer 85 (5), 643–648. doi:10.1002/(SICI)1097-0215(20000301)85:5<643::AID-IJC8>3.0.CO;2-Z
Khan, M. A. K., Yang, J., Hussain, S. A., Zhang, H., Liang, L., Garre, V., et al. (2019). Construction of DGLA producing cell factory by genetic modification of mucor Circinelloides. Microb. Cell Fact. 18, 64–68. doi:10.1186/s12934-019-1110-4
Khongto, B., Laoteng, K., and Tongta, A. (2010). Fermentation process development of recombinant Hansenula polymorpha for gamma-linolenic acid production. J. Microbiol. Biotechnol. 20 (11), 1555–1562. doi:10.4014/jmb.1003.03004
Khongto, B., Laoteng, K., and Tongta, A. (2011). Enhancing the production of gamma-linolenic acid in Hansenula polymorpha by fed-batch fermentation using response surface methodology. Chem. Pap. 65, 124–131. doi:10.2478/s11696-010-0099-2
Klempova, T., Basil, E., Kubatova, A., and Certik, M. (2013). Biosynthesis of gamma-linolenic acid and beta-carotene by Zygomycetes fungi. Biotechnol. J. 8 (7), 794–800. doi:10.1002/biot.201200099
Kousparou, C., Fyrilla, M., Stephanou, A., and Patrikios, I. (2023). DHA/EPA (Omega-3) and LA/GLA (Omega-6) as bioactive molecules in neurodegenerative diseases. Int. J. Mol. Sci. 24 (13), 10717. doi:10.3390/ijms241310717
Kumar, G., Shekh, A., Jakhu, S., Sharma, Y., Kapoor, R., and Sharma, T. R. (2020). Bioengineering of microalgae: recent advances, perspectives, and regulatory challenges for industrial application. Front. Bioeng. Biotechnol. 8, 914. doi:10.3389/fbioe.2020.00914
Laoteng, K., Ruenwai, R., Tanticharoen, M., and Cheevadhanarak, S. (2005). Genetic modification of essential fatty acids biosynthesis in Hansenula polymorpha. FEMS Microbiol. Lett. 245 (1), 169–178. doi:10.1016/j.femsle.2005.03.006
Lee, K. R., Yu, H., Jeon, I., Kim, K. H., Park, J. S., Lee, J., et al. (2019). Accumulation of γ-linolenic acid and stearidonic acid in rapeseeds that heterologously express the Phytophthora citrophthora Δ6 desaturase gene. Plant Biotechnol. Rep. 13, 399–407. doi:10.1007/s11816-019-00547-y
Leventhal, L. J., Boyce, E. G., and Zurier, R. B. (1993). Treatment of rheumatoid arthritis with gammalinolenic acid. Ann. Intern. Med. 119 (9), 867–873. doi:10.7326/0003-4819-119-9-199311010-00001
Li, Q., Zhang, J., Guan, X., Lu, Y., Liu, Y., Liu, J., et al. (2023). Metabolite analysis of soybean oil on promoting astaxanthin production of Phaffia rhodozyma. J. Sci. Food Agric. 103 (6), 2997–3005. doi:10.1002/jsfa.12365
Lin, F., Chen, Y., Levine, R., Lee, K., Yuan, Y., and Lin, X. N. (2013). Improving fatty acid availability for bio-hydrocarbon production in Escherichia coli by metabolic engineering. PloS one 8 (10), e78595. doi:10.1371/journal.pone.0078595
Ling, Z., Zhifeng, L., Bingsen, L., Xiaojing, S., Yafang, T., and Shuqin, S. (2000). Experiment study on the fermentation production of gamma-linolenic acid by thamnidium elegans TE7-15. Shengwu Jishu= Biotechnol. 10 (5), 25–27.
Lu, H., Li, J. N., Chai, Y. R., and Zhang, X. K. (2009). Identification and characterization of a novel ∆ 6-fatty acid desaturase gene from Rhizopus nigricans. Mol. Biol. Rep. 36, 2291–2297. doi:10.1007/s11033-009-9447-0
Lu, H., and Zhu, Y. (2015). Screening and molecular identification of overproducing γ-linolenic acid fungi and cloning the Delta 6-desaturase gene. Biotechnol. Appl. Biochem. 62 (3), 316–322. doi:10.1002/bab.1281
Lupu, R., Busby, R., Cheol, P. H., and Espinoza, I. (2012). Abstract 3854: gamma-Linolenic acid (GLA) sensitizes pancreatic cancer cells to gemcitabine. Cancer Res. 72 (8_Suppl. ment), 3854. doi:10.1158/1538-7445.AM2012-3854
Mahboubi, M. (2019). Evening primrose (Oenothera biennis) oil in management of female ailments. J. Menopausal Med. 25 (2), 74–82. doi:10.6118/jmm.18190
Mamatha, S. S., Ravi, R., and Venkateswaran, G. (2008). Medium optimization of gamma linolenic acid production in mucor Rouxii CFR-G15 using RSM. Food Bioprocess Technol. 1, 405–409. doi:10.1007/s11947-008-0103-9
Matschinske, J., Salgado-Albarrán, M., Sadegh, S., Bongiovanni, D., Baumbach, J., and Blumenthal, D. B. (2020). Individuating possibly repurposable drugs and drug targets for COVID-19 treatment through hypothesis-driven systems medicine using CoVex. Assay. Drug Dev. Technol. 18 (8), 348–355. doi:10.1089/adt.2020.1010
Maurice, N. (2019). “Role of solid-state fermentation to enhance cellulase production,” in New future dev. Microbial biotechnol. Bioeng. (Elsevier), 127–153. doi:10.1016/B978-0-444-64223-3.00009-6
Meechai, A., Pongakarakun, S., Deshnium, P., Cheevadhanarak, S., and Bhumiratana, S. (2004). Metabolic flux distribution for γ-linolenic acid synthetic pathways in Spirulina platensis. Biotechnol. Bioprocess Eng. 9, 506–513. doi:10.1007/BF02933494
Melanouri, E. M., Papanikolaou, S., and Diamantopoulou, P. (2024). Mortierella ramanniana lipid fermentation wastewater as an innovative maceration liquid medium for sustainable solid-state cultivation of higher fungi. Waste Biomass Valor 15 (12), 6903–6925. doi:10.1007/s12649-024-02575-4
Mendes, R. L., Reis, A. D., and Palavra, A. F. (2006). Supercritical CO2 extraction of γ-linolenic acid and other lipids from arthrospira (spirulina) maxima: Comparison with organic solvent extraction. Food Chem. 99 (1), 57–63. doi:10.1016/j.foodchem.2005.07.019
Mingchun, L., Li, L., Guowu, H., and Laijun, R. (2003). Studies on the cloning, structure and function of the Δ∼(6)-fatty acid desaturase genes from Mortierella Aplina. Wei Sheng wu xue bao= Acta Microbiol. Sin. 43 (2), 220–227.
Mirbagheri Firoozabad, M. S., and Akhbariyoon, H. (2022). Engineering and fermenter production of fungi GLA in Pichia pastoris GS115 using oil waste. Arch. Microbiol. 204 (10), 635. doi:10.1007/s00203-022-03182-4
Miura, Y., Sode, K., Nakamura, N., Matsunaga, N., and Matsunaga, T. (1993). Production of γ-linolenic acid from the marine green alga chlorella sp. NKG 042401. FEMS Microbiol. Lett. 107 (2-3), 163–167. doi:10.1111/j.1574-6968.1993.tb06024.x
Mohamed, H., Awad, M. F., Shah, A. M., Nazir, Y., Naz, T., Hassane, A., et al. (2022a). Evaluation of different standard amino acids to enhance the biomass, lipid, fatty acid, and γ-linolenic acid production in Rhizomucor pusillus and mucor Circinelloides. Front. Nutr. 9, 876817. doi:10.3389/fnut.2022.876817
Mohamed, H., Awad, M. F., Shah, A. M., Sadaqat, B., Nazir, Y., Naz, T., et al. (2022b). Coculturing of mucor Plumbeus and Bacillus subtilis bacterium as an efficient fermentation strategy to enhance fungal lipid and gamma-linolenic acid (GLA) production. Sci. Rep. 12 (1), 13111. doi:10.1038/s41598-022-17442-2
Montaner, C., Zufiaurre, R., Movila, M., and Mallor, C. (2021). Evaluation of borage (Borago officinalis L.) genotypes for nutraceutical value based on leaves fatty acids composition. Foods 11 (1), 16. doi:10.3390/foods11010016
Mudroňová, D., Karaffová, V., Koščová, J., Bartkovský, M., Marcinčáková, D., Popelka, P., et al. (2018). Effect of fungal gamma-linolenic acid and beta-carotene containing prefermented feed on immunity and gut of broiler chicken. Poult. Sci. 97 (12), 4211–4218. doi:10.3382/ps/pey306
Mustonen, A. M., and Nieminen, P. (2023). Dihomo-γ-linolenic acid (20: 3N-6)—metabolism, derivatives, and potential significance in chronic inflammation. Int. J. Mol. Sci. 24 (3), 2116. doi:10.3390/ijms24032116
Navarette, R., Tang, W., and Ziboh, V. A. (1992). Dietary intake of concentrated gamma-linolenic acid (GLA)-Enriched oil suppresses cutaneous level of dihomo-gamma-linolenic acid (DGLA): possible in vivo inhibition of microsomal elongation of GLA to DGLA. Prostagl. Leukot. Essent. Fat. Acids 46 (2), 139–144. doi:10.1016/0952-3278(92)90220-D
Nilsen, D. W. T., Myhre, P. L., Kalstad, A., Schmidt, E. B., Arnesen, H., and Seljeflot, I. (2021). Serum levels of dihomo-gamma (γ)-linolenic acid (DGLA) are inversely associated with linoleic acid and total death in elderly patients with a recent myocardial infarction. Nutrients 13 (10), 3475. doi:10.3390/nu13103475
Novichkova, E., Chumin, K., Eretz-Kdosha, N., Boussiba, S., Gopas, J., Cohen, G., et al. (2020). DGLA from the microalga lobosphaera incsa P127 modulates inflammatory response, inhibits iNOS expression and alleviates NO secretion in RAW264. 7 murine macrophages. Nutrients 12 (9), 2892. doi:10.3390/nu12092892
Nykiforuk, C. L., Shewmaker, C., Harry, I., Yurchenko, O. P., Zhang, M., Reed, C., et al. (2012). High level accumulation of gamma linolenic acid (C18: 3Δ6. 9, 12 cis) in transgenic safflower (Carthamus tinctorius) seeds. Transgenic Res. 21, 367–381. doi:10.1007/s11248-011-9543-5
Pagels, F., Guedes, A. C., Vasconcelos, V., and Lopes, G. (2022). “Anti-inflammatory compounds from Cyanobacteria,” in Pharmacol. Potent. Cyanobact. (Academic Press), 81–105. doi:10.1016/B978-0-12-821491-6.00004-1
Papanikolaou, S., Diamantopoulou, P., Chatzifragkou, A., Philippoussis, A., and Aggelis, G. (2010). Suitability of low-cost sugars as substrates for lipid production by the fungus Thamnidium elegans. Energy fuels. 24 (7), 4078–4086. doi:10.1021/ef1004804
Papanikolaou, S., Dimou, A., Fakas, S., Diamantopoulou, P., Philippoussis, A., Galiotou-Panayotou, M., et al. (2011). Biotechnological conversion of waste cooking olive oil into lipid-rich biomass using aspergillus and penicillium strains. J. Appl. Microbiol. 110 (5), 1138–1150. doi:10.1111/j.1365-2672.2011.04961.x
Papanikolaou, S., Komaitis, M., and Aggelis, G. (2004). Single cell oil (SCO) production by Mortierella isabellina grown on high-sugar content media. Bioresour. Technol. 95 (3), 287–291. doi:10.1016/j.biortech.2004.02.016
Papanikolaou, S., Rontou, M., Belka, A., Athenaki, M., Gardeli, C., Mallouchos, A., et al. (2017). Conversion of biodiesel-derived glycerol into biotechnological products of industrial significance by yeast and fungal strains. Eng. Life Sci. 17 (3), 262–281. doi:10.1002/elsc.201500191
Peng, B., Bandari, N. C., Lu, Z., Howard, C. B., Scott, C., Trau, M., et al. (2022). Engineering eukaryote-like regulatory circuits to expand artificial control mechanisms for metabolic engineering in Saccharomyces cerevisiae. Commun. Biol. 5 (1), 135. doi:10.1038/s42003-022-03070-z
Pérez-García, F., and Wendisch, V. F. (2018). Transport and metabolic engineering of the cell factory Corynebacterium glutamicum. FEMS Microbiol. Lett. 365 (16), fny166. doi:10.1093/femsle/fny166
Petrie, J. R., Shrestha, P., Mansour, M. P., Nichols, P. D., Liu, Q., and Singh, S. P. (2010). Metabolic engineering of omega-3 long-chain polyunsaturated fatty acids in plants using an acyl-CoA Δ6-desaturase with ω3-preference from the marine microalga Micromonas pusilla. Metab. Eng. 12 (3), 233–240. doi:10.1016/j.ymben.2009.12.001
Poor, M. H., Saghafi, Z., and Ghaedamini, H. (2023). Comparing the effects of oral and vaginal evening primrose capsules with concurrent labor induction and exclusive labor induction on bishop score and some labor outcomes: a triple-blind clinical trial. J. Nurs. Midwifery Sci. 10 (4). doi:10.5812/jnms-140350
Pop-Busui, R., Ang, L., Boulton, A. J., Feldman, E. L., Marcus, R. L., Mizokami-Stout, K., et al. (2022). Diagnosis and treatment of painful diabetic peripheral neuropathy. Compendia 2022, 1–32. doi:10.2337/db2022-01
Ramli, D., Aziz, I., Mohamad, M., Abdulahi, D., and Sanusi, J. (2017). The changes in rats with sciatic nerve crush injury supplemented with evening primrose oil: behavioural, morphologic, and morphometric analysis. Evid. Based Complement. Altern. Med. 2017 (1), 3476407. doi:10.1155/2017/3476407
Ratledge, C. (2013). Microbial production of polyunsaturated fatty acids as nutraceuticals. in Microbial production of food ingredients, enzymes and nutraceuticals (Cambridge, United Kingdom: Woodhead Publishing), 531–558. doi:10.1533/9780857093547.2.531
Robert, S. S., Singh, S. P., Zhou, X. R., Petrie, J. R., Blackburn, S. I., Mansour, P. M., et al. (2005). Metabolic engineering of arabidopsis to produce nutritionally important DHA in seed oil. Funct. Plant Biol. 32 (6), 473–479. doi:10.1071/FP05084
Ronda, S. R., and Lele, S. S. (2008). Culture conditions stimulating high γ-Linolenic acid accumulation by Spirulina platensis. Braz. J. Microbiol. 39, 693–697. doi:10.1590/S1517-83822008000400018
Ronda, S. R., Parupudi, P. L. C., Vemula, S., Tumma, S., Botlagunta, M., Settaluri, V. S., et al. (2014). Optimization of medium components using orthogonal arrays for γ-Linolenic acid production by Spirulina platensis. Korean J. Chem. Eng. 31, 1839–1844. doi:10.1007/s11814-014-0082-7
Ruiz del Castillo, M. L., Dobson, G., Brennan, R., and Gordon, S. (2004). Fatty acid content and juice characteristics in Black currant (Ribes nigrum L.) genotypes. J. Agric. Food Chem. 52 (4), 948–952. doi:10.1021/jf034950q
Ruiz-López, N., Sayanova, O., Napier, J. A., and Haslam, R. P. (2012). Metabolic engineering of the omega-3 long chain polyunsaturated fatty acid biosynthetic pathway into transgenic plants. J. Exp. Bot. 63 (7), 2397–2410. doi:10.1093/jxb/err454
Salah, A., Zhan, M., Cao, C., Han, Y., Ling, L., Liu, Z., et al. (2019). γ-Aminobutyric acid promotes chloroplast ultrastructure, antioxidant capacity, and growth of waterlogged maize seedlings. Sci. Rep. 9 (1), 484. doi:10.1038/s41598-018-36334-y
Sato, S., Xing, A., Ye, X., Schweiger, B., Kinney, A., Graef, G., et al. (2004). Production of γ-Linolenic acid and stearidonic acid in seeds of marker-free transgenic soybean 1. Crop Sci. 44 (2), 646–652. doi:10.2135/cropsci2004.6460
Sayegh, F., Elazzazy, A., Bellou, S., Moustogianni, A., Elkady, A. I., Baeshen, M. N., et al. (2016). Production of polyunsaturated single cell oils possessing antimicrobial and anticancer properties. Ann. Microbiol. 66, 937–948. doi:10.1007/s13213-015-1176-0
Schindler, D. (2020). Genetic engineering and synthetic genomics in yeast to understand life and boost biotechnology. Bioengineering 7 (4), 137. doi:10.3390/bioengineering7040137
Seifalyazal, M. E., Albalkiny, S., Fawzy, G., and Mortada, A. E. (2023). Vitamin E versus evening primrose oil versus placebo for the treatment of cyclic mastalgia: a double-blind randomized controlled trial. Egypt. J. Hosp. Med. 91 (1), 3870–3876. doi:10.21608/ejhm.2023.293472
Sergeant, S., Rahbar, E., and Chilton, F. H. (2016). Gamma-linolenic acid, dihommo-gamma linolenic, eicosanoids and inflammatory processes. Eur. J. Pharmacol. 785, 77–86. doi:10.1016/j.ejphar.2016.04.020
Serra, S., D’Arrigo, P., Rossato, L. A. M., and Ruffini, E. (2023). Microbial fermentation of the water-soluble fraction of brewers’ spent grain for the production of high-value fatty acids. Fermentation 9 (12), 1008. doi:10.3390/fermentation9121008
Shahbazi, M., Nematzadeh-Gharakhili, G., Yamchi, A., and Bagherani, N. (2018). Effect of salinity stress on yield, oil quality and expression of Delta 6-fatty acid desatutase (D6DES) gene in evening primrose (Oenothera biennis L.). Plant Cell Biotechnol. Mol. Biol. 19, 148–154.
Shi, A., Liu, Y., Jia, B., Zheng, G., and Yao, Y. (2023). Metabolic engineering of microorganisms to produce L-Aspartate and its derivatives. Fermentation 9 (8), 737. doi:10.3390/fermentation9080737
Shi, S., and Zhao, H. (2017). Metabolic engineering of oleaginous yeasts for production of fuels and chemicals. Front. Microbiol. 8, 2185. doi:10.3389/fmicb.2017.02185
Shivaramu, M., Ravi, R., and Venkateswaran, G. (2008). Medium Optimization of Gamma Linolenic Acid Production in Mucor rouxii CFR-G15 using RSM. Food and Bioprocess Technol, 1, 405–409. doi:10.1007/s11947-007-0017-9
Simopoulos, A. P. (2021). Genetic variation, diet, inflammation, and the risk for COVID-19. Lifest. Genomics 14 (2), 37–42. doi:10.1159/000513886
Slaný, O., Klempová, T., Marcinčák, S., and Čertík, M. (2020). Production of high-value bioproducts enriched with γ-linolenic acid and β-carotene by filamentous fungi Umbelopsis isabellina using solid-state fermentations. Ann. Microbiol. 70, 1–11. doi:10.1186/s13213-020-01545-0
Slaný, O., Klempová, T., Shapaval, V., Zimmermann, B., Kohler, A., and Čertík, M. (2021). Animal fat as a substrate for production of n-6 fatty acids by fungal solid-state fermentation. Microorganisms 9 (1), 170. doi:10.3390/microorganisms9010170
Sohail, R., Mathew, M., Patel, K. K., Reddy, S. A., Haider, Z., Naria, M., et al. (2023). Effects of non-steroidal anti-inflammatory drugs (NSAIDs) and gastroprotective NSAIDs on the gastrointestinal tract: a narrative review. Cureus 15 (4), e37080. doi:10.7759/cureus.37080
Somashekar, D., Venkateshwaran, G., Sambaiah, K., and Lokesh, B. R. (2003). Effect of culture conditions on lipid and gamma-linolenic acid production by mucoraceous fungi. Process Biochem. 38 (12), 1719–1724. doi:10.1016/S0032-9592(02)00258-3
Song, Z., Rogers, A., Cho, B. A., Jing, K., and Zhang, D. (2022). “Kinetic modelling of γ-linolenic acid production by Cunninghamella echinulata,”, 51. Comput. Aided Chem. Eng. 109–114. doi:10.1016/B978-0-323-95879-0.50019-9
Sroczyk, E. A., Berniak, K., Jaszczur, M., and Stachewicz, U. (2022). Topical electrospun patches loaded with oil for effective gamma linoleic acid transport and skin hydration towards atopic dermatitis skincare. Chem. Eng. J. 429, 132256. doi:10.1016/j.cej.2021.132256
Streďanská, S., and Šajbidor, J. (1992). Oligounsaturated fatty acid production by selected strains of micromycetes. Folia Microbiol. 37, 357–359. doi:10.1007/BF02815662
Su, H., Shi, P., Shen, Z., Meng, H., Meng, Z., Han, X., et al. (2023). High-level production of nervonic acid in the oleaginous yeast Yarrowia lipolytica by systematic metabolic engineering. Commun. Biol. 6 (1), 1125. doi:10.1038/s42003-023-05502-w
Sukrutha, S. K., Adamechova, Z., Rachana, K., Savitha, J., and Certik, M. (2014). Optimization of physiological growth conditions for maximal gamma-linolenic acid production by cunninghamella blakesleeana-JSK2. J. Am. Oil Chem. Soc. 91 (9), 1507–1513. doi:10.1007/s11746-014-2507-1
Sun, M. L., Madzak, C., Liu, H. H., Song, P., Ren, L. J., Huang, H., et al. (2017). Engineering Yarrowia lipolytica for efficient γ-linolenic acid production. Biochem. Eng. J. 117, 172–180. doi:10.1016/j.bej.2016.10.014
Taghon, G. J., and Strychalski, E. A. (2023). Rise of synthetic yeast: charting courses to new applications. Cell Genomics 3 (11), 100438. doi:10.1016/j.xgen.2023.100438
Tamano, K., Nakai, S., Takayama, H., and Imai, Y. (2023). Overexpression of a predicted transketolase gene and disruption of an α-1, 3-glucan synthase gene in Aspergillus oryzae DGLA3 strain enhances the yield of free dihomo-γ-linolenic acid. Biosci. Biotechnol. Biochem. 87 (4), 448–457. doi:10.1093/bbb/zbad003
Tański, W., Świątoniowska-Lonc, N., Tabin, M., and Jankowska-Polańska, B. (2022). The relationship between fatty acids and the development, course and treatment of rheumatoid arthritis. Nutrients 14 (5), 1030. doi:10.3390/nu14051030
Thurairatnam, S. (2012). Hematopoietic prostaglandin D synthase inhibitors. Prog. Med. Chem. 51, 97–133. doi:10.1016/B978-0-12-396493-9.00004-2
Tong, Z., Zheng, X., Tong, Y., Shi, Y. C., and Sun, J. (2019). Systems metabolic engineering for citric acid production by Aspergillus Niger in the post-genomic era. Microb. Cell Fact. 18, 28–15. doi:10.1186/s12934-019-1064-6
Urrestarazu, M., Manuel Gallegos-Cedillo, V., Ferrón-Carrillo, F., Luis Guil-Guerrero, J., Lao, T., and Eugenio Álvaro, J. (2018). Effects of the salinity under soilless culture systems on gamma linolenic acid levels in borage seed oil. bioRxiv, 454405. doi:10.1371/journal.pone.0207106
Ursin, V. M. (2003). Modification of plant lipids for human health: development of functional land-based omega-3 fatty acids. J. Nutr. 133 (12), 4271–4274. doi:10.1093/jn/133.12.4271
Van Hoorn, R., Kapoor, R., and Kamphuis, J. (2008). A short review on sources and health benefits of GLA, the GOOD omega-6. Oléagineux, Corps gras, Lipides 15 (4), 262–264. doi:10.1051/ocl.2008.0207
Vasilakis, G., Roidouli, C., Karayannis, D., Giannakis, N., Rondags, E., Chevalot, I., et al. (2024). Study of different parameters affecting production and productivity of polyunsaturated fatty acids (PUFAs) and γ-linolenic acid (GLA) by Cunninghamella elegans through glycerol conversion in shake flasks and bioreactors. Microorganisms 12 (10), 2097. doi:10.3390/microorganisms12102097
Verma, D., Asthana, R. K., and Nath, G. (2021). Antarctic cyanobacterium nostoc CCC537, a new source of Î3-linolenic acid and its antibacterial potential. Chem. Biol. Lett. 8 (2), 50–58.
Wan, X., Zhang, Y., Wang, P., Huang, F., Chen, H., and Jiang, M. (2009). Production of gamma-linolenic acid in Pichia pastoris by expression of a delta-6 Desaturase gene from Cunninghamella echinulata. J. Microbiol. Biotechnol. 19 (10), 1098–1102. doi:10.4014/jmb.0902.0071
Wan, X., Zhang, Y., Wang, P., and Jiang, M. (2011). Molecular cloning and expression analysis of a Delta 6-fatty acid desaturase gene from Rhizopus stolonifer strain YF6 which can accumulate high levels of gamma-linolenic acid. J. Microbiol. 49, 151–154. doi:10.1007/s12275-011-0254-7
Wang, J., Yu, X., Wang, K., Lin, L., Liu, H. H., Ledesma-Amaro, R., et al. (2023). Reprogramming the fatty acid metabolism of Yarrowia lipolytica to produce the customized omega-6 polyunsaturated fatty acids. Bioresour. Technol. 383, 129231. doi:10.1016/j.biortech.2023.129231
Wang, X., Yang, J., Mohamed, H., Shah, A. M., Li, S., Pang, S., et al. (2022). Simultaneous overexpression of ∆ 6-∆ 12-and∆ 9-desaturases enhanced the production of γ-linolenic acid in mucor Circinelloides WJ11. Front. Microbiol. 13, 1078157. doi:10.3389/fmicb.2022.1078157
Williams, H. C. (2003). Evening primrose oil for atopic dermatitis. BMJ 327 (7428), 1358–1359. doi:10.1136/bmj.327.7428.1358
Won, J. C., Kwon, H. S., Moon, S. S., Chun, S. W., Kim, C. H., Park, I. B., et al. (2020). γ-Linolenic acid versus α-lipoic acid for treating painful diabetic neuropathy in adults: a 12-week, double-placebo, randomized, noninferiority trial. Diabetes Metab. J. 44 (4), 542–554. doi:10.4093/dmj.2019.0099
Wu, C., Yang, J., Li, S., Shi, W., Xue, F., Liu, Q., et al. (2023a). Construction of eicosatetraenoic acid producing cell factory by genetic engineering of mucor circinelloides. Fermentation 9 (7), 653. doi:10.3390/fermentation9070653
Wu, K., Xu, C., Li, T., Ma, H., Gong, J., Li, X., et al. (2023b). Application of nanotechnology in plant genetic engineering. Int. J. Mol. Sci. 24 (19), 14836. doi:10.3390/ijms241914836
Zarghi, A., and Arfaei, S. (2011). Selective COX-2 inhibitors: a review of their structure-activity relationships. Iran. J. Pharm. Res. IJPR 10 (4), 655–683.
Zhang, C., Ma, K., Yang, Y., Wang, F., and Li, W. (2021a). Glaucocalyxin A suppresses inflammatory responses and induces apoptosis in TNF-a-induced human rheumatoid arthritis via modulation of the STAT3 pathway. Chem.-Biol. Interact. 341, 109451. doi:10.1016/j.cbi.2021.109451
Zhang, Q., Li, M., Ma, H., Sun, Y., and Xing, L. (2004). Identification and characterization of a novel Δ6-fatty acid desaturase gene from Rhizopus arrhizus. FEBS Lett. 556 (1-3), 81–85. doi:10.1016/S0014-5793(03)01380-2
Zhang, Y., Liu, Q., Li, P., Wang, Y., Li, S., Gao, M., et al. (2021b). Enhanced lipid production by addition of malic acid in fermentation of recombinant mucor Circinelloides Mc-MT-2. Sci. Rep. 11 (1), 12674. doi:10.1038/s41598-021-92324-7
Zhang, Y., Luan, X., Zhang, H., Garre, V., Song, Y., and Ratledge, C. (2017). Improved γ-linolenic acid production in mucor Circinelloides by homologous overexpressing of delta-12 and delta-6 Desaturases. Microb. Cell Fact. 16, 113–119. doi:10.1186/s12934-017-0723-8
Ziegler, D. (2023). Pathogenetic treatments for diabetic peripheral neuropathy. Diabetes Res. Clin. Pract. 206, 110764. doi:10.1016/j.diabres.2023.110764
Zikou, E., Chatzifragkou, A., Koutinas, A. A., and Papanikolaou, S. (2013). Evaluating glucose and xylose as cosubstrates for lipid accumulation and γ-linolenic acid biosynthesis of Thamnidium elegans. J. Appl. Microbiol. 114 (4), 1020–1032. doi:10.1111/jam.12116
Keywords: gamma-linolenic acid, GLA production enhancem, metabolic engineering, genetic modification, genome editing technologies, GLA-producing microorganisms, omega-6 polyunsaturated fatty acids (PUFAs)
Citation: Latifi M, Jalali Bidgoli F, Hajihassani H, Hassani D, Ingvarsson PK and Farrokhi N (2025) Recent advances and future directions on GLA-producing organisms. Front. Bioeng. Biotechnol. 13:1567840. doi: 10.3389/fbioe.2025.1567840
Received: 28 January 2025; Accepted: 12 June 2025;
Published: 09 July 2025; Corrected: 08 September 2025.
Edited by:
Alok Patel, Luleå University of Technology, SwedenReviewed by:
Carla C. C. R. de Carvalho, Universidade de Lisboa, PortugalIqra Mariam, Luleå University of Technology, Sweden
Copyright © 2025 Latifi, Jalali Bidgoli, Hajihassani, Hassani, Ingvarsson and Farrokhi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Naser Farrokhi, bl9mYXJyb2toaUBzYnUuYWMuaXI=; Pär K. Ingvarsson, cGFyLmluZ3ZhcnNzb25Ac2x1LnNl
†ORCID: Naser Farrokhi, orcid.org/0000-0003-4676-1545; Pär K. Ingvarsson, orcid.org/0000-0001-9225-7521