 Alba González-Rodríguez1F. Javier De Toro2,3,4Alberto Jorge-Mora5Pablo Fernandez-Pernas6Carlota Probaos Rivadulla2,3,4María Fraga7
Alba González-Rodríguez1F. Javier De Toro2,3,4Alberto Jorge-Mora5Pablo Fernandez-Pernas6Carlota Probaos Rivadulla2,3,4María Fraga7 Juan A. Fafián-Labora2,3,4*
Juan A. Fafián-Labora2,3,4* María C. Arufe2,3,4*
María C. Arufe2,3,4*- 1Servicio de Cirugía Plástica, Complexo Hospitalario Universitario de A Coruña (CHUAC), A Coruña, Spain
- 2Grupo de Terapia Celular Medicina, Instituto de Investigación Biomédica de A Coruña (INIBIC), Complexo Hospitalario Universitario de A Coruña (CHUAC), Sergas/ Universidade da Coruña (UDC), As Xubías, A Coruña, Spain
- 3CICA - Centro Interdisciplinar de Química e Bioloxía e, Departamento de Bioloxía, Facultade de Ciencias, Universidade da Coruña (UDC), Campus de Elviña, CICA, A Coruña, Spain
- 4Departamento de Fisioterapia, Medicina y Ciencias Biomédicas, Facultad de Ciencias de la Salud, Universidade da Coruña (UDC), Campus de Elviña, CICA, A Coruña, Spain
- 5Cirugíía Ortopédica y Traumatología / Complejo Hospitalario de A Coruña, A Coruña, Spain
- 6Centre of Biological Sciences, University of Veterinary Medicine of Vienna, Vienna, Austria
- 7Department of Anatomical Pathology, University Hospital Complex A Coruña, A Coruña, Spain
Osteoarthritis (OA) is a degenerative joint disease marked by inflammation, cartilage degradation, and pain, leading to a significant decline in quality of life. Recent advancements in extracellular vesicle (EV) research have introduced new therapeutic possibilities, with small extracellular vesicles (sEV) emerging as a promising strategy for OA treatment. sEV, particularly those derived from mesenchymal stem cells (MSCs), synoviocytes, chondrocytes, and induced pluripotent stem cells (iPSCs), demonstrate substantial anti-inflammatory and regenerative properties. These nanosized vesicles facilitate intercellular communication, delivering bioactive molecules that can modulate the joint microenvironment, promote chondrogenesis, and alleviate pain. Preclinical and early clinical studies indicate that sEV-based therapies may slow disease progression and enhance cartilage repair in OA patients. Despite the promising potential, challenges remain, including standardizing isolation techniques, understanding underlying mechanisms, and navigating regulatory pathways. This systematic review analyzes relevant publications published between 2019 and 2025, highlighting the therapeutic and biomarker potential of sEV in OA. Although there is substantial ongoing research into sEV and biomarkers, the fundamental understanding of OA pathogenesis remains largely unchanged, with most studies continuing to focus on established mechanisms of cartilage degradation, inflammation, and subchondral bone changes. The findings suggest that while therapeutic research into sEV is progressing, advancements in unraveling new pathophysiological mechanisms of OA are more limited. Further research is essential to optimize therapeutic protocols and establish clinical efficacy, marking sEV-based therapies as a promising but evolving approach for OA treatment.
1 Introduction
Osteoarthritis (OA) is one of the most common degenerative joint disorders and a major contributor to global, particularly among the aged population. It is a complex, multifactorial disease characterized by the progressive degeneration of articular cartilage, Changes in subchondral bone, formation of osteophytes, and inflammation of the synovial membrane (Xu et al., 2025). These alterations lead to chronic pain, joint stiffness, functional impairment, and a reduced quality of life. Despite the high prevalence and socioeconomic burden of OA, effective disease-modifying treatments remain elusive.
Currently, OA management relies heavily on conservative strategies such as non-steroidal anti-inflammatory drugs (NSAIDs), corticosteroid injections, physical therapy, and lifestyle modifications, with joint replacement surgery rather than targeting the underlying biological mechanisms driving cartilage degradation and joint dysfunction (Figueroa-Valdés et al., 2025). This therapeutic gap underscores the urgent need for innovative, disease-modifying interventions that can halt or even reverse OA progression.
Extracellular vesicles (EV) are membrane-bound particles released by cells that play essential roles in intercellular communication, particularly in the context of osteoarthritis (OA), where they influence inflammation, cartilage degradation, and tissue regeneration. EV are broadly categorized into small EVs (sEV), typically under 200 nm, encompassing exosomes (30–150 nm) that originate from the endosomal system and large EV (lEV), which include microvesicles (100–1,000 nm) shed directly from the plasma membrane, as well as apoptotic bodies (500–5,000 nm) released during programmed cell death. Various isolation techniques are employed to obtain EV from biological fluids or cell culture media, each with distinct benefits and limitations (Gupta et al., 2021). Differential ultracentrifugation is widely used to separate EVs based on size and density, while density gradient centrifugation improves purity. Size exclusion chromatography allows for the separation of EVs from soluble proteins and smaller particles, and polymer-based precipitation (e.g., with polyethylene glycol) offers a rapid, scalable option (Yuan et al., 2024). More advanced methods, such as immunoaffinity capture and microfluidic platforms, enable greater specificity and are increasingly adopted for isolating defined EV subpopulations, crucial for investigating their diverse roles in OA pathology and therapeutic potential.
In recent years, regenerative medicine has emerged as a promising avenue for OA treatment. Among the various regenerative strategies under investigation, sEV have gained significant attention due to their intrinsic capacity for intercellular communication and their ability to modulate a variety of biological processes. sEV are nanoscale membrane-bound vesicles released by almost all cell types and contain a rich cargo of proteins (Ruiz et al., 2020; Zhang et al., 2021; Li et al., 2022; Nguyen et al., 2022; Ragni et al., 2022b; Genestra et al., 2023; Cheng et al., 2024; Forteza-Genestra et al., 2024; Mohd Noor et al., 2024; Scalzone et al., 2024; Wu R. et al., 2024), lipids (Kolhe et al., 2020; Mustonen et al., 2022; Zhang et al., 2023), mRNAs (Li et al., 2024c), and microRNAs (LIN et al., 2021). These vesicles play critical roles in maintaining tissue homeostasis and orchestrating cellular responses in both physiological and pathological conditions.
Preclinical studies have demonstrated that sEV derived from mesenchymal stem cells (MSCs), chondrocytes, and synoviocytes exert anti-inflammatory, anti-apoptotic, and pro-regenerative effects in the context of OA. For instance, MSC-derived sEV have been shown to promote cartilage repair, modulate immune responses, and inhibit matrix-degrading enzymes (Tang et al., 2021; Morente-López et al., 2022; Warmink et al., 2023). Likewise, chondrocyte-derived sEV can enhance extracellular matrix synthesis (Shang et al., 2021; Hosseinzadeh et al., 2023), while synoviocyte-derived sEV may influence synovial homeostasis and reduce joint inflammation (Chen et al., 2022b; Asghar et al., 2024). These findings suggest that sEV-based therapies hold substantial potential as minimally invasive, cell-free approaches for OA treatment.
However, despite this promising landscape, several critical gaps in the literature remain. Firstly, the precise molecular mechanisms through which sEV exert their therapeutic effects in OA are not yet fully elucidated, with much of the existing evidence limited to in vitro or small-animal models. Secondly, there is a lack of standardized protocols for the isolation, purification, and characterization of sEV, leading to variability in experimental outcomes and complicating cross-study comparisons. Thirdly, the comparative efficacy of sEV derived from different cell sources remains poorly defined, as does the optimal route of administration, dosage, and treatment frequency. Moreover, while some early-phase clinical trials are underway, robust clinical evidence supporting the safety, efficacy, and long-term outcomes of sEV-based therapies in human OA patients is still lacking.
This review seeks to address these deficiencies by providing a comprehensive and critical analysis of current research on the application of sEV in OA therapy. Specifically, it aims to 1) examine the therapeutic mechanisms of sEV in cartilage regeneration, immune modulation, and inflammation resolution; 2) compare the regenerative potential of sEV derived from various cell sources; and 3) discuss the key translational challenges and considerations for clinical application, including regulatory hurdles, manufacturing scalability, and safety concerns.
By integrating and synthesizing the latest findings in this rapidly evolving field, this manuscript contributes to a more nuanced understanding of sEV as emerging therapeutic agents for OA and highlights avenues for future research and clinical translation. Addressing these critical questions is essential for advancing sEV-based therapies from experimental models toward safe and effective clinical applications that could transform the management of osteoarthritis.
2 Methodology
2.1 Search strategy
A systematic literature search was conducted using the PubMed and Web of Science databases. The search strategy combined keywords and Medical Subject Headings (MeSH) relevant to osteoarthritis and small extracellular vesicles (sEV). Boolean operators (AND, OR) were used to refine search results.
Search terms included: “osteoarthritis,” “small extracellular vesicles,” “exosomes,” “extracellular vesicles,” “cartilage breakdown,” “matrix metalloproteinases (MMPs),” “aggrecanases,” “subchondral bone changes,” “sclerosis formation,” “osteophyte formation,” “synovial inflammation,” and “pro-inflammatory cytokines.”
The search was limited to articles published between January 2019 and March 2025.
2.2 Eligibility criteria
Studies were selected using the PICO framework:
• Population: Human patients, animal models, or in vitro systems affected by osteoarthritis.
• Intervention: Administration or analysis of sEV.
• Comparator(s): Placebo, untreated controls, or standard interventions (if reported).
• Outcomes: Biomarker changes and therapeutic efficacy in OA models.
Inclusion Criteria:
• Original research on the role of sEV in OA pathophysiology, diagnosis, or treatment.
• Articles published in English in peer-reviewed journals.
• Studies employing human, animal, or in vitro models.
Exclusion Criteria:
• Review articles, meta-analyses, editorials, or abstracts without original data.
• Studies not directly related to OA or sEV
• Articles focusing solely on large extracellular vesicles or unrelated vesicles subtypes.
• Non-English publications.
2.3 Data sources and study selection
Two independent reviewers (Reviewer A and Reviewer B) screened all retrieved titles and abstracts for relevance. Full-text articles were then assessed based on the predefined inclusion and exclusion criteria. Disagreements were resolved through discussion or by a third reviewer (Reviewer C). No automation tools were used in this stage.
A total of 162 studies were ultimately included in the qualitative synthesis. These were grouped into three main thematic areas based on the primary focus of each publication:
39 studies focused on OA pathogenesis, including cartilage degradation, synovial inflammation, and subchondral bone changes.
122 studies investigated the therapeutic potential of sEV in OA, primarily using in vitro or in vivo preclinical models.
22 studies evaluated sEV as potential biomarkers for OA diagnosis, prognosis, or treatment response.
2.4 Data extraction
Data from the included studies were extracted using a standardized data extraction form by two reviewers independently. Information collected included:
• Study characteristics (author, publication year, journal, study design)
• Type of model used (in vivo or in vitro)
• Outcomes measured, including:
Biomarkers in OA: inflammatory cytokines (IL-1β, TNF-α), matrix metalloproteinases (MMP-13), aggrecanases, cartilage-specific proteins (collagen II, aggrecan); measured by ELISA, RT-PCR, or immunohistochemistry.
Therapeutic efficacy: improvements in joint structure or function, evaluated via WOMAC scores, MRI/CT imaging, or histological scoring systems.
All extracted data were cross-verified, and disagreements were resolved by consensus. No data automation tools were employed, and no study authors were contacted for additional data.
• Key findings and conclusions.
2.5 Quality assessment and risk of bias
Quality and risk of bias were assessed independently by two reviewers using appropriate tools:
• For in vivo studies: ARRIVE (Animal Research: Reporting of in vivo Experiments) guidelines.
• For in vitro studies: Internal quality assessment criteria were developed based on reproducibility, experimental controls, and reporting clarity.
• For all studies: Risk of bias was evaluated using a modified version of the SYRCLE Risk of Bias tool for preclinical studies, and criteria included randomization, blinding, and outcome reporting.
Each study was assessed by two reviewers working independently. Discrepancies were resolved via discussion or consultation with a third reviewer.
2.6 Data synthesis
All included studies were synthesized qualitatively. Narrative summaries were used to describe key findings and thematic patterns. Where appropriate, data were tabulated to compare models, outcomes, and results across studies.
The 39 studies related to OA pathogenesis were further divided into three subcategories: Cartilage Breakdown (n = 23); synovial Inflammation (n = 7); subchondral bone changes (n = 9).
The 122 therapeutic studies assessed outcomes such as cartilage repair, inflammation reduction, and functional improvements using sEV derived from various cell sources, most commonly MSCs. The 22 biomarker studies explored the diagnostic potential of sEV cargo.
No meta-analysis was conducted due to heterogeneity in study designs and outcome measures.
2.7 Ethical considerations
This systematic review adhered to the PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analyses) guidelines. As the review did not involve primary data collection, ethical approval was not required.
3 Results
A total of 354 records were retrieved from the Web of Science and PubMed databases after duplicate removal. After screening the titles and abstracts, 53 records were excluded, leaving 301 articles to be assessed after reading the abstract. Following full-text screening, 281 scientific papers meeting these criteria were selected. After excluding studies that did not meet the specific selection criteria, in vitro, in vivo and clinical studies, out of a total of 162 scientific papers, were included in the qualitative synthesis. The PRISMA flow diagram is shown in Figure 1.
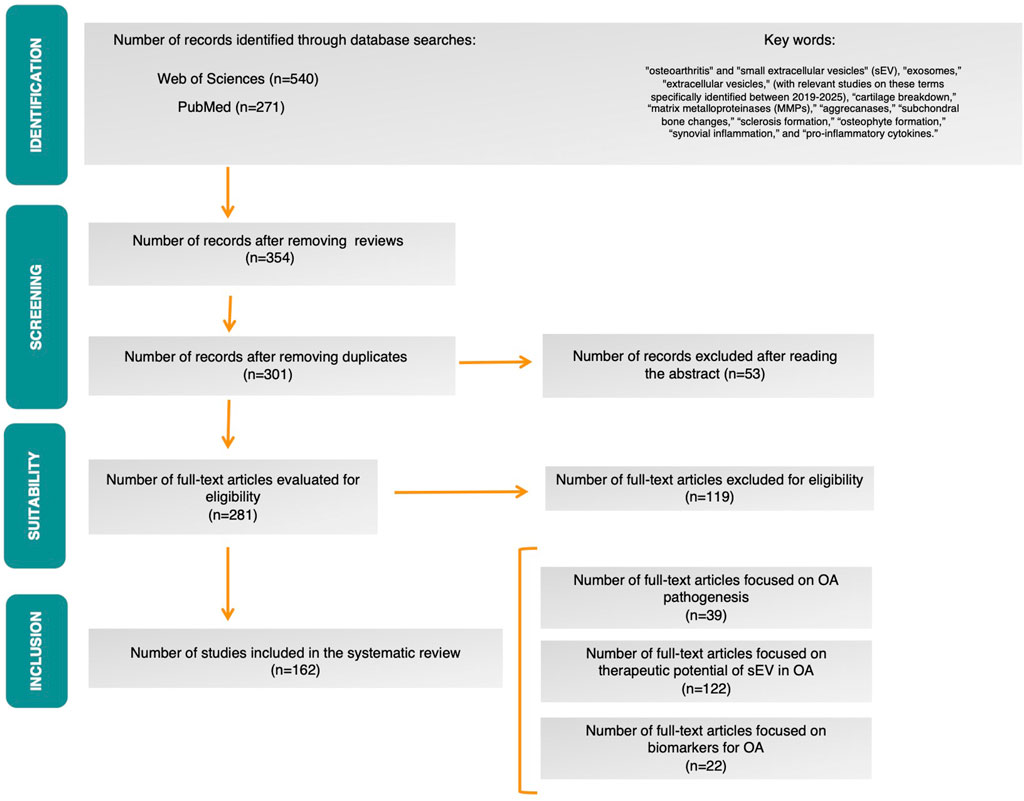
Figure 1. PRISMA flow diagram.
In our systematic review, we analyzed a total of 39 relevant publications published between 2019 and 2025, focusing on key aspects of osteoarthritis (OA) pathogenesis. However, to provide a comprehensive state of the art and due to the lack of significant advances in recent years, we also included studies prior to 2019, as earlier works remain crucial for understanding the fundamental basis of the disease. These publications were grouped into three main categories:
i) Cartilage Breakdown: 23 publications discussed cartilage degradation mediated by factors such as MMPs and aggrecanases.
ii) Synovial Inflammation: 7 publications addressed the role of pro-inflammatory cytokines, such as IL-1β and TNF-α, in synovial inflammation.
iii) Subchondral Bone Changes: 9 publications discussed changes in the subchondral bone, including sclerosis and osteophyte formation.
The inclusion of studies prior to 2019 highlights that, the foundational knowledge of OA pathogenesis remains relatively stable. Most of the relevant studies build upon established concepts such as cartilage degradation mechanisms and inflammatory processes, which have been extensively studied in previous years. This suggests that while therapeutic research is advancing, progress in uncovering new aspects of OA pathogenesis has been limited. The current focus appears to be more on applying existing knowledge to explore new therapeutic avenues rather than discovering new pathophysiological mechanisms (Figure 2).

Figure 2. Analysis of studies about osteoarthritis pathogenesis. Histogram of number of studies classified by key pathological features. IL-1β (Interleukin-1β); TNF-α (Tumor Necrosis Factor-α); MMPs (matrix metalloproteinases).
In addition to the pathogenic mechanisms, 122 studies investigated the therapeutic potential of sEV in OA, primarily in preclinical models. These studies highlighted the regenerative, anti-inflammatory, and chondroprotective effects of sEV derived from sources such as mesenchymal stem cells (Kouroupis et al., 2023; Warmink et al., 2023), chondrocytes (Wang et al., 2020; Chen et al., 2024; Feng et al., 2024a; Yan et al., 2024), and synoviocytes (Lu et al., 2021; Lai et al., 2023; Wang and Sun, 2023; Cao H. et al., 2024). Furthermore, 22 publications evaluated EVs as potential biomarkers for OA diagnosis, progression monitoring, and treatment response prediction. Together, these findings underscore the growing interest in EVs as both mechanistic contributors to OA pathology and promising tools for therapy and clinical translation.
The consistent upward trend in the number of studies from 2019 to 2025 indicates a growing recognition of sEV as a significant focus in OA research. This growth reflects an expanding understanding of their therapeutic potential and biomarker capabilities in managing OA. A notable increase in publications in 2024 specifically highlights the therapeutic potential and biomarker applications of mesenchymal stem cells (MSCs) and synovial fluid in OA research (Figures 3a, 4a).
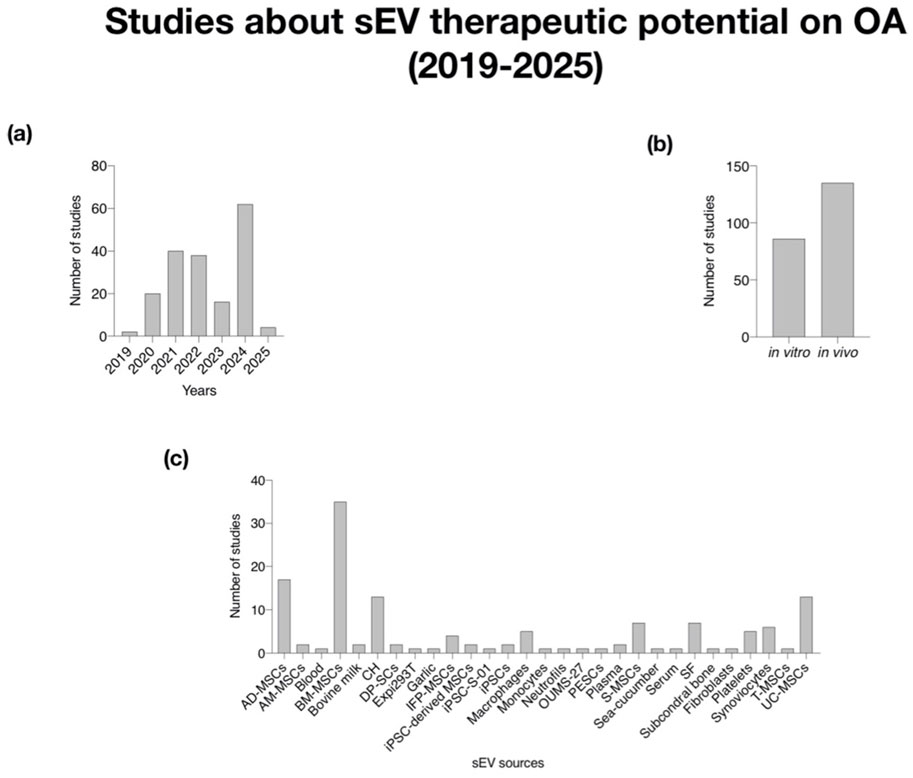
Figure 3. Analysis of studied about sEV therapeutic potential on OA (2019–2025). (a) Histogram of number of studies per year; (b) Histogram of number of studies in vitro and in vivo; (c) Histogram of the number of studies classified by sEV sources. AM-MSCs (Mesenchymal Stem Cells from Ammiotic Membrane); AD-MSCs (Mesenchymal Stem Cells from Adipose Tissue); BM-MSCs (Mesenchymal Stem Cells from Bone Marrow); DP-SCs (Dental Pulp-derived Stem Cells); IFP-MSCs (Infrapatellar Fat Pad-derived Mesenchymal Stem Cells); iPSC-derived MSCs (Mesenchymal Stem Cell-derived iPSCs); iPSCs (induced Pluripotent Stem Cells); S-MSCs (Mesenchymal Stem Cells from synovial membrane); T-MSCs (Tonsil-derived Mesenchymal Stem Cells); SF (Synovial Fluid); UC-MSCs (Umbilical Cord-derived Mesenchymal Stem Cells).
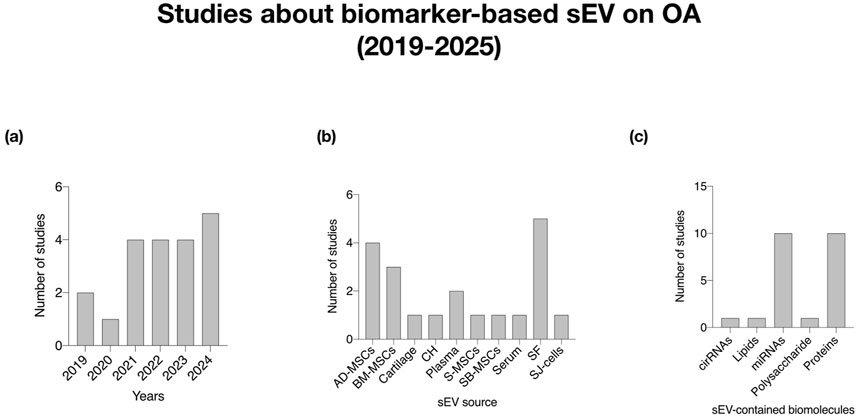
Figure 4. Analysis of studies about sEV cargo as biomarker in OA (2019–2025). (a) Histogram of number of studies per year; (b) Histogram of the number of studies classified by sEV sources; (c) Histogram of number of studies based on sEV-contained biomolecules. CH (Chondrocytes); SJ-cells (Cells from Synovial Joint); AD-MSCs (Mesenchymal Stem Cells from Adipose Tissue); BM-MSCs (Mesenchymal Stem Cells from Bone Marrow).
The rise in studies, particularly in 2024, points to an expanding interest in the therapeutic potential of sEV derived from MSCs and their roles as biomarkers in OA research. This reflects a deeper understanding of how sEV can be leveraged for both treatment and diagnostic purposes.
The distribution of studies between in vitro and in vivo research suggests a systematic approach to validating the effects of sEV before pre-clinical application (Figure 3b). The balance between these research approaches indicates thorough validation, with in vitro studies providing foundational insights on mechanisms, while in vivo studies contribute to understanding the efficacy and safety of sEV therapies in real biological contexts.
The variety of sEV sources identified, including various types of MSCs, plasma, serum, synovial fluid, and others, demonstrates the multifaceted nature of sEV research. The increasing research on MSC-derived sEV, including adipose-derived and bone marrow, highlights their diverse therapeutic potential. Additionally, studies on sEV from synovial fluid emphasize their role in OA treatment, reinforcing the unique properties derived from multiple sources. The inclusion of additional sources such as macrophages and platelets suggest a comprehensive exploration of immune and cellular responses tied to OA pathology (Figure 3c, 4b).
The analysis of sEV-contained biomolecules like miRNAs (10 publications), proteins (9 publications), circRNAs (1 publication), and lipids (1 publication) underscores the relevance of these molecules as potential biomarkers. Notably, miRNAs and proteins emerge as the most extensively studied and utilized biomolecules in sEV research. This highlights the dual role of sEV as both therapeutic agents and diagnostic tools. An increased examination of these biomolecules, particularly miRNAs and proteins from MSCs (10 publications) and synovial fluid (5 publications), suggests that specific sEV-associated biomarkers may lead to personalized treatment strategies (Figure 4c).
The trends observed in both therapeutic and biomarker studies highlight critical avenues for future research, including the need for standardized methodologies and larger clinical trials. The significant volume of research, particularly emphasizing MSCs and synovial fluid, highlights the necessity of refining clinical applications of sEV in OA management. These endeavors could enhance the legitimacy and applicability of sEV therapies in clinical practice for OA.
The insights from this systematic review integrate the intricate relationship between the growing body of literature on sEV, their diverse sources, and their potential roles as both therapeutic tools and biomarkers in OA. This interconnected understanding fosters a holistic approach to addressing OA, paving the way for innovative research and potential clinical advancements in the coming years. The research underscores the interconnectedness of sEV, their diverse sources, and their roles in OA management, supporting a comprehensive approach to addressing this condition while encouraging innovative research and clinical advancements in the field.
This systematic review clarifies the potential impact of sEV, especially those derived from MSCs and synovial fluid, in OA. It also sets the foundation for identifying critical areas for future exploration, particularly given the increase in studies focusing on these topics in 2024.
4 Discussion
4.1 Pathophysiology of OA
OA is a multifactorial disease driven by mechanical stress, inflammation, and metabolic changes. Key pathological features include:
4.1.1 Cartilage breakdown mediated by matrix metalloproteinases (MMPs) and aggrecanases
Cartilage breakdown is a hallmark of OA and other joint degenerative conditions. It is primarily mediated by two enzymatic families: matrix metalloproteinases (MMPs) and aggrecanases. These enzymes degrade the extracellular matrix (ECM), which is crucial for maintaining cartilage integrity and function. While MMPs and aggrecanases target different ECM components, their activities are interrelated. For instance, MMPs can degrade the non-collagenous domains of aggrecan, complementing aggrecanase-mediated cleavage (Madsen et al., 2010). Moreover, MMP activity is essential for aggrecanase activation under certain pathological conditions (Clements et al., 2011). Below, we synthesize findings from key studies to elucidate the roles of MMPs and aggrecanases in cartilage degradation. MMPs are a family of zinc-dependent endopeptidases that degrade various ECM components. Among these, MMP-3 (stromelysin-1) and MMP-13 (collagenase-3) are particularly implicated in cartilage breakdown. MMP-13 degrades type II collagen, a critical structural component of cartilage, while MMP-3 activates other MMPs and contributes to proteoglycan loss (Cawston et al., 1998; Stanton et al., 2002). Elevated MMP activity is observed in inflammatory and degenerative joint diseases, where cytokines such as IL-1β and TNF-α drive their expression (Tsushima et al., 2012).
MMP-mediated cartilage degradation is tightly regulated by tissue inhibitors of metalloproteinases (TIMPs). TIMP-3 (tissue inhibitor of metalloproteinase-3) is particularly significant, as it inhibits both MMPs and aggrecanases, thereby preserving cartilage ECM integrity (Gendron et al., 2003; Li et al., 2004). However, dysregulation of the TIMP/MMP balance contributes to cartilage destruction in OA. MMPs, particularly MMP-3 and MMP-13, degrade cartilage collagens and proteoglycans, while aggrecanases (ADAMTS-4, ADAMTS-5) target aggrecan, a major cartilage proteoglycan, leading cartilage breakdown (Fosang et al., 2000; Stanton et al., 2002; Madsen et al., 2010). Pro-inflammatory cytokines like IL-1β and IL-17 upregulate MMP and aggrecanase expression, intensifying cartilage degradation. IL-17 also stimulates nitric oxide production, suppressing matrix synthesis (Cai et al., 2001; 2002; Huang et al., 2020). TIMP-3 effectively inhibits aggrecanases, while selective metalloproteinase inhibitors reduce cartilage degradation (Gendron et al., 2003).
Aggrecanases, primarily ADAMTS-4 and ADAMTS-5 (a disintegrin and metalloproteinase with thrombospondin motifs), are specialized for degrading aggrecan, the predominant proteoglycan in cartilage. Aggrecan degradation compromises cartilage’s load-bearing properties, exacerbating joint dysfunction (Fosang et al., 2000; Madsen et al., 2010). Aggrecanase activity is often upregulated in response to pro-inflammatory mediators, such as IL-1 and IL-17 (Cai et al., 2001; Wilkinson et al., 2017). Studies have highlighted the molecular characteristics of aggrecan fragments generated during degradation. These fragments serve as biomarkers for cartilage turnover and disease progression (Patel et al., 2007). Importantly, targeted inhibition of aggrecanases has shown promise in preclinical models, reducing cartilage degradation, and improving joint function (Kanakis et al., 2019).
While MMPs and aggrecanases target different ECM components, their activities are interrelated. For instance, MMPs can degrade the non-collagenous domains of aggrecan, complementing aggrecanase-mediated cleavage (Madsen et al., 2010). Moreover, MMP activity is essential for aggrecanase activation under certain pathological conditions (Clements et al., 2011).Nutraceuticals like luteolin and crocin attenuate MMP and aggrecanase activity (Gendron et al., 2003; Moncada-Pazos et al., 2011). Mechanical stress, injury, and oxidative stress induce MMP/aggrecanase expression. Pathways such as ERK, NF-κB, and PKC play critical roles in this regulation (Maeda et al., 2001; Haslauer et al., 2013). Targeted aggrecanase inhibition (e.g., ADAMTS-5) reduces cartilage damage and preserves joint function. Experimental drugs like suramin and recombinant proteins (e.g., TSG-6) show promise in modulating MMP/aggrecanase activity (Kanakis et al., 2019; Drummond et al., 2023).
Cartilage degradation involves both reversible and irreversible processes, depending on the enzymes involved. This study showed that aggrecanase-mediated degradation, especially in early stages, can be reversed with anabolic stimuli like IGF-1, leading to restored proteoglycan content and ECM integrity. In contrast, once MMPs degrade aggrecan and type II collagen, particularly during later stages, the chondrocytes exhibit limited repair capacity. Collagen II formation was only significantly recovered at early time points, suggesting a narrow therapeutic window. Histological analysis confirmed glycosaminoglycan replenishment when intervention occurred early. These findings emphasize the timing and nature of enzymatic activity in OA progression. Therapies aimed at early aggrecanase inhibition may preserve regenerative potential. Meanwhile, MMP-targeted strategies are crucial for halting irreversible damage. The distinction between reversible and irreversible cartilage damage provides a framework for stage-specific OA treatment design (Karsdal et al., 2008). Elevated levels of degradation products (e.g., aggrecan fragments) and inflammatory markers (e.g., S100A8, Tenascin-C) serve as indicators of cartilage breakdown (Van Lent et al., 2008; Sofat et al., 2012). Hyaluronic acid (HA), for example, has been shown to partially inhibit MMP and aggrecanase activity, thereby protecting cartilage (Mladenovic et al., 2014). Similarly, small-molecule inhibitors, such as luteolin and mannosamine, selectively reduce aggrecanase activity, preserving ECM integrity (Patwari et al., 2000; Moncada-Pazos et al., 2011). Modulating the activity of MMPs and aggrecanases represents a therapeutic avenue for preventing cartilage breakdown. Emerging therapies also focus on enhancing endogenous protective mechanisms. For instance, microRNA-17 maintains cartilage homeostasis by suppressing MMP and aggrecanase expression (Zhang et al., 2022). Furthermore, TIMP-3 overexpression and recombinant Link protein supplementation have shown potential in reducing ECM degradation in experimental models (Drummond et al., 2023).
MMPs and aggrecanases are central to the pathogenesis of cartilage degradation, driven by inflammatory and mechanical stressors. Their interplay underscores the complexity of ECM turnover and highlights opportunities for therapeutic intervention. Future research should aim to refine selective inhibitors and explore regenerative strategies to restore cartilage homeostasis.
4.1.2 Synovial inflammation due to the release of pro-inflammatory cytokines such as IL-1β and TNF-α
The release of pro-inflammatory cytokines, particularly IL-1β and TNF-α, plays a pivotal role in initiating and sustaining the inflammatory milieu. These cytokines trigger a cascade of molecular and cellular events that lead to synovial hyperplasia, fibroblast activation, cartilage degradation, and recruitment of immune cells. The interaction between pro-inflammatory cytokines and synovial fibroblasts plays a critical role in the pathogenesis of OA. Farina et al. demonstrated that the activation of synovial fibroblasts by IL-1β and TNF-α is further enhanced by the pro-β-nerve growth factor (proNGF) and its receptor p75NTR (Farina et al., 2022). This novel mechanism amplifies the inflammatory response, making it a promising therapeutic target. Additionally, synovial fibroblasts stimulated with these cytokines release mediators such as Trx80, a truncated form of thioredoxin. According to Lemarechal et al., Trx80 contributes to the perpetuation of inflammation by promoting further cytokine release and oxidative stress, exacerbating joint damage (Lemarechal et al., 2007). The synovial membrane plays a pivotal role in OA progression by responding to mechanical stress and wear particles. Harvanova et al. highlighted how in vitro models of OA can replicate these processes, demonstrating the synovial membrane’s contribution to cartilage and joint degradation (Harvanova et al., 2022). Park et al. further corroborated this, showing that polyethylene wear particles activate synovial inflammation, leading to cartilage damage and meniscus degradation (Park et al., 2013). The pro-inflammatory effects of cytokines extend beyond fibroblast activation. Filková et al. reported that interleukin-35 (IL-35), typically considered anti-inflammatory, can paradoxically exhibit pro-inflammatory effects in rheumatic arthritis (RA) (Filková et al., 2015). This dichotomy underscores the complex cytokine signaling in synovitis. Furthermore, oncostatin M (OSM), another cytokine involved in synovial inflammation, induces pro-inflammatory mediators via the JAK/STAT pathway. Weston et al. demonstrated that JAK inhibitors like baricitinib effectively block this pathway, reducing inflammation in synovial-derived cells. This highlights the therapeutic potential of targeted cytokine pathway inhibition. Targeted drug delivery systems have emerged as promising strategies for treating synovial inflammation (Weston et al., 2022). Hu et al. explored pH-responsive polymeric nanoparticles loaded with rhein, which selectively release the drug in the acidic inflammatory microenvironment. This innovation minimizes systemic side effects while directly addressing synovial inflammation.
Pro-inflammatory cytokines IL-1β and TNF-α drive synovial inflammation through multiple mechanisms, including fibroblast activation, cytokine crosstalk, and oxidative stress. Advances in understanding these pathways have unveiled novel therapeutic targets such as proNGF-p75NTR signaling, IL-35 modulation, and JAK/STAT inhibition. Additionally, innovative delivery systems like pH-sensitive nanoparticles present promising strategies to control inflammation locally and effectively (Hu et al., 2020).
Further studies integrating molecular, cellular, and translational approaches are essential for the development of targeted therapies to treat synovial inflammation and its sequelae in OA.
4.1.3 Subchondral bone changes, including sclerosis and osteophyte formation
Changes in subchondral bone, including sclerosis and osteophyte formation, are key features that contribute to joint degeneration. Emerging studies provide detailed insights into the molecular and structural alterations underlying these changes. Subchondral bone sclerosis is a hallmark of OA, characterized by increased bone density and altered architecture. The thickened subchondral plate and trabecular remodeling are thought to result from mechanical stress and aberrant signaling pathways. Ruan et al. highlighted the multifaceted role of ALK3 signaling in OA development, demonstrating its involvement in subchondral bone sclerosis through the regulation of osteoblast activity and bone matrix production (Ruan et al., 2023). The study emphasizes the interplay between cartilage degradation and subchondral bone remodeling, wherein mechanical stress exacerbates both processes. Experimental models, such as the anterior cruciate ligament transection (ACLT) and meniscectomy in rats, have been instrumental in elucidating the mechanisms underlying sclerosis. Pickarski et al. observed significant thickening of the subchondral plate and increased mineralization in these models, accompanied by elevated levels of osteoclast and osteoblast activity markers (Pickarski et al., 2011). Automated micro-CT analysis further supports these findings, as Das Neves Borges et al. demonstrated precise quantification of bone volume and density changes in murine OA models, confirming the progressive nature of subchondral bone sclerosis (Das Neves Borges et al., 2017). Osteophytes, or bone spurs, are another hallmark of OA and represent an adaptive response to joint instability and cartilage damage. Their formation is driven by endochondral ossification and influenced by mechanical stress, inflammation, and signaling pathways. Liao et al. noted that osteophyte development in axial spondyloarthritis shares similarities with OA, particularly in the activation of growth factors like transforming growth factor-β (TGF-β), which stimulates mesenchymal stem cell differentiation into chondrocytes and osteoblasts (Liao et al., 2024). Wang et al. demonstrated the sequential progression of cartilage degradation, bone resorption, and osteophyte formation in a temporomandibular joint OA model (Wang et al., 2012). The study revealed that osteophytes arise in response to altered joint biomechanics and increased expression of pro-osteogenic factors. Additionally, Jimenez et al. observed spontaneous osteophyte formation in Dunkin Hartley guinea pigs, further corroborating the link between mechanical loading and osteophyte development. Increased physical activity, particularly under conditions of compromised cartilage integrity, accelerates osteophyte formation (Jimenez et al., 1997). Siebelt et al. reported severe osteophyte growth in pain-induced OA models subjected to excessive loading. These findings underscore the dual role of mechanical stress in promoting both sclerosis and osteophyte formation, highlighting the importance of joint biomechanics in OA pathophysiology. The interplay between cartilage and subchondral bone is critical in OA progression (Siebelt et al., 2014). Acute synovitis, as described by Liao et al., precedes subchondral bone changes and amplifies joint degeneration through inflammatory mediators that impact both tissues (Liao et al., 2020). Furthermore, Pickarski et al. and Hayami et al. identified coordinated molecular changes in articular cartilage and subchondral bone, suggesting a dynamic feedback loop that perpetuates joint damage. Sclerotic changes are not limited to structural alterations but also involve molecular shifts. Hayami et al. identified upregulation of bone morphogenetic proteins (BMPs) and Wnt signaling in subchondral bone during OA progression, further contributing to increased bone formation and sclerosis (Hayami et al., 2006; Pickarski et al., 2011).
Subchondral bone changes, including sclerosis and osteophyte formation, are integral to OA pathogenesis and progression. Advances in imaging and molecular analysis have deepened our understanding of these processes, offering potential therapeutic targets. Interventions aimed at modulating mechanical stress, inflammation, and aberrant signaling pathways could hold promise in alleviating subchondral bone alterations and mitigating OA severity.
4.2 sEV therapy on OA
sEV have emerged as promising tools in regenerative medicine and disease treatment. These nano-sized particles are released by various cell types and are enriched with bioactive molecules, including proteins, lipids, and nucleic acids. Their ability to deliver these molecules to target cells positions sEV as pivotal agents in intercellular communication, making them a focus of interest in therapeutic development. This review highlights the diverse therapeutic roles of sEV, focusing on their capacity to promote chondrogenesis, modulate inflammation, and stimulate angiogenesis and tissue repair.
4.2.1 Promotion of chondrogenesis and cartilage repair and regeneration
They are critical challenges in orthopedic medicine due to the avascular and aneural nature of cartilage tissue. sEV have demonstrated a remarkable ability to support chondrogenesis, making them a promising therapeutic option for cartilage-related disorders such as OA.
Delivery of chondroprotective molecules in sEV derived from MSCs are particularly notable for their role in cartilage repair. They are enriched with chondroprotective factors, including SOX9 mRNA and specific microRNAs, such as miR-140, which are key regulators of chondrogenesis (LIN et al., 2021). SOX9, a transcription factor, is critical for cartilage formation as it regulates the expression of cartilage-specific extracellular matrix proteins, such as collagen type II and aggrecan. The presence of SOX9 mRNA in sEV ensures the transfer of this essential regulatory molecule to recipient cells, thereby enhancing cartilage repair mechanisms (Li et al., 2024c).
Similarly, miR-140 has been identified as a cartilage-specific microRNA that plays a pivotal role in maintaining cartilage homeostasis. Studies have shown that sEV enriched with miR-140 promote the proliferation and differentiation of chondrocytes while simultaneously inhibiting the expression of matrix-degrading enzymes such MMPs. This dual action ensures the preservation and repair of cartilage tissue, positioning sEV as a potent therapeutic tool (Ragni et al., 2022a).
Additionally, Scalzone et al. have proposed other factors such as BMP-2 and TGF-β3 found in sEV from BM-MSCs, that contribute to cartilage regeneration by enhancing chondrocyte differentiation and extracellular matrix deposition (Scalzone et al., 2024) (Table 1).
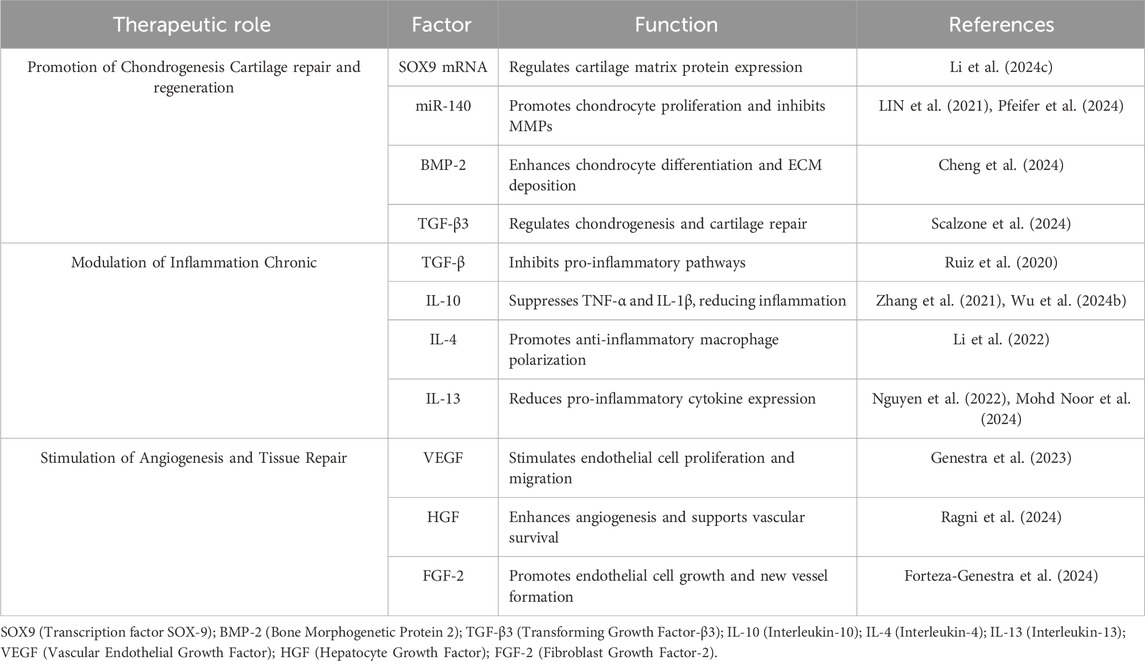
Table 1. Key Factors in sEV with Therapeutic Potential for OA treatment.
4.2.2 Enhancement of extracellular matrix production
A critical mechanism through which sEV promote chondrogenesis is by enhancing the synthesis of ECM components. sEV transport a wide array of bioactive molecules, including growth factors, cytokines, and miRNAs, which modulate cellular behavior and stimulate the production of essential ECM constituents such as type II collagen and glycosaminoglycans (Gendron et al., 2003; Siebelt et al., 2014). Notably, miR-140, a cartilage-specific miRNA frequently enriched in sEV, has been shown to promote chondrocyte proliferation while inhibiting the expression of matrix-degrading enzymes such as MMPs (LIN et al., 2021; Pfeifer et al., 2024). Similarly, bone morphogenetic protein-2 (BMP-2), another cargo molecule identified in therapeutic sEV, plays a pivotal role in enhancing chondrocyte differentiation and ECM deposition (Cheng et al., 2024).
In addition, Otahal et al. demonstrated that this ability to enhance ECM synthesis underscores the therapeutic promise of plasma- and serum-derived sEV in treating cartilage-related disorders (Otahal et al., 2020; 2021).
For example, sEV derived from induced pluripotent stem cells (iPSCs) have been shown to significantly boost ECM production in both in vitro and in vivo cartilage repair models. Hsueh et al. demonstrated that iPSC-derived sEV significantly enhanced ECM production in both in vitro and in vivo OA models. These sEV promoted primary chondrocyte proliferation, reduced premature senescence, and restored collagen II synthesis, while suppressing matrix-degrading enzymes like MMP-13 and ADAMTS-5. In a rabbit anterior cruciate ligament transection (ACLT)-induced OA model, intraarticular injection of iPSC-sEV ameliorated cartilage damage and preserved tissue integrity (Hsueh et al., 2023).
Further advancements have been achieved using MSCs differentiated from iPSCs as a source of sEV. Feng et al. developed engineered sEV derived from iPSC-MSCs, modified with a cartilage-targeting peptide and loaded with siRNA against MDM2. These sEV selectively targeted and eliminated senescent chondrocytes via modulation of the MDM2-p53 pathway, restoring ECM metabolic homeostasis and enhancing collagen II expression. The therapeutic effect was confirmed in both post-traumatic and age-related OA mouse models, as well as in ex vivo cultured human OA cartilage explants (Feng et al., 2025).
To overcome the challenge of limited bioavailability following intraarticular delivery, Yang et al. employed a novel injectable Diels–Alder crosslinked hyaluronic acid/PEG (DAHP) hydrogel to deliver MSC-sEV derived from iPSCs. This hydrogel ensured sustained release and preserved the functional integrity of sEV, resulting in enhanced cartilage repair and ECM regeneration in vivo. The study provided a clinically relevant platform that prolongs joint retention of therapeutic vesicles and supports their gradual release within the synovial environment (Yang et al., 2021).
Collectively, these studies emphasize the capacity of iPSC-derived MSC-sEV to not only stimulate ECM synthesis but also modulate chondrocyte senescence and matrix catabolism. Through innovative delivery systems and bioengineering, sEV emerge as potent, cell-free therapeutic agents with high translational potential in osteoarthritis treatment.
4.2.3 Modulation of inflammation chronic
It is a hallmark of many degenerative diseases, including osteoarthritis and rheumatoid arthritis. The ability of sEV to modulate inflammatory responses makes them an attractive therapeutic candidate for managing these conditions.
sEV carry a diverse array of anti-inflammatory molecules, including cytokines and growth factors such as TGF-β and IL-10. TGF-β is a multifunctional cytokine that plays a critical role in immune regulation and tissue repair. By inhibiting the activation of pro-inflammatory pathways, TGF-β in sEV helps reduce synovial inflammation, a major driver of cartilage degradation in osteoarthritis (Ossendorff et al., 2023).
Similarly, IL-10 is a potent anti-inflammatory cytokine that suppresses the production of pro-inflammatory mediators such as TNF-α and IL-1β. The presence of IL-10 in sEV ensures the inhibition of these harmful inflammatory pathways, creating a more conducive environment for tissue repair and regeneration. In addition, factors such as IL-4 and IL-13 found in sEV contribute to immune modulation by promoting an anti-inflammatory macrophage phenotype and reducing pro-inflammatory cytokine expression (Nakamachi et al., 2023; Mohd Noor et al., 2024) (Table 1).
4.2.4 Stimulation of angiogenesis and tissue repair
The repair and regeneration of damaged tissues frequently requires the re-establishment of vascular networks to ensure a sufficient supply of oxygen, nutrients, and cellular components necessary for repair. In this context, sEV have been shown to play a critical role in stimulating angiogenesis and facilitating tissue repair, making them a valuable therapeutic tool in regenerative medicine.
Angiogenic Factors in sEV are enriched with pro-angiogenic factors such as vascular endothelial growth factor (VEGF) and hepatocyte growth factor (HGF). VEGF is a key regulator of angiogenesis, promoting the proliferation and migration of endothelial cells to form new blood vessels. The presence of VEGF in sEV ensures the stimulation of angiogenic processes, which are critical for tissue repair and regeneration (Shang et al., 2021) (Table 1).
HGF, another important factor found in sEV, plays a dual role in promoting angiogenesis and enhancing tissue repair. HGF stimulates the migration and proliferation of endothelial cells while also promoting the survival of existing vascular structures. Additionally, fibroblast growth factor-2 (FGF-2), found in sEV, further enhances endothelial cell proliferation, and supports the formation of new blood vessels (Thomas et al., 2022) (Table 1).
Although articular cartilage is inherently avascular, a feature that is essential for its unique biomechanical and immune-privileged properties, the role of angiogenesis becomes increasingly relevant in the pathogenesis of OA. During OA progression, aberrant neovascularization has been observed in previously avascular regions such as the deep layers of articular cartilage and the osteochondral junction (Ragni et al., 2022b; Genestra et al., 2023; Forteza-Genestra et al., 2024).
Moreover, vascular remodeling in the subchondral bone and synovial membrane plays a critical role in OA. In the subchondral bone, increased angiogenesis is associated with bone marrow lesions, sclerosis, and osteophyte formation (Ragni et al., 2022b; Forteza-Genestra et al., 2024). Similarly, neovascularization within the synovium can sustain local inflammation and further disrupt joint homeostasis.
Given this context, the modulation of angiogenesis by sEV offers a promising therapeutic avenue in OA. Rather than indiscriminately promoting neovascularization, targeted delivery of sEV could support vascular remodeling in beneficial zones such as the subchondral bone while avoiding detrimental vascular invasion into cartilage.
4.2.5 Enhancement of subchondral bone remodeling
In addition to promoting angiogenesis, sEV also play a role in subchondral bone remodeling. Subchondral bone, located beneath the cartilage, plays a critical role in supporting cartilage health and function. Damage to this structure is a common feature of OA and other joint disorders. sEV derived from MSCs have been shown to promote the repair and remodeling of subchondral bone by delivering bioactive molecules that stimulate osteoblast activity and inhibit osteoclast-mediated bone resorption. This ability to restore subchondral bone integrity further underscores the therapeutic potential of sEV in joint repair (Niedermair et al., 2020).
These findings collectively highlight the significant potential of sEV in regenerative medicine, particularly for cartilage repair, immune modulation, and tissue regeneration.
4.3 OA pre-clinical sEV-based studies
Recent pre-clinical studies have explored the therapeutic potential of sEV derived from various cell sources in OA models. These studies provide compelling evidence for the efficacy of sEV in mitigating OA progression, reducing inflammation, and promoting cartilage regeneration (Table 2).
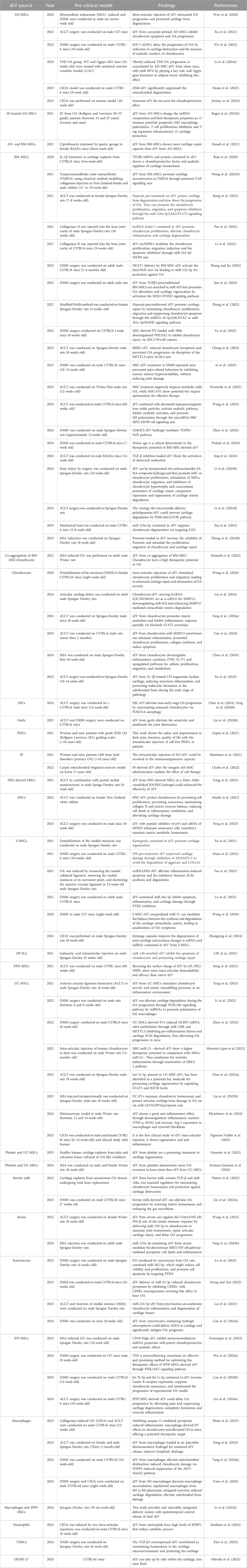
Table 2. The most relevant pre-clinical studies about therapeutic potential of sEV on OA.
Studies utilizing MSC-sEV have demonstrated promising chondroprotective effects. For instance, BM-MSCs sEV have been shown to modulate key signaling pathways such as JAK2/STAT3, SOX11/FOXO, and PI3K/Akt to promote chondrocyte proliferation, migration, and survival, as well as inhibit apoptosis and inflammation (Fazaeli et al., 2021; Hotham et al., 2021; Pan et al., 2021; Ragni et al., 2022b; Li Z. et al., 2024; Scalzone et al., 2024; Tang X. et al., 2024). Notably, pre-conditioning strategies such as hypoxia treatment or cytokine stimulation have enhanced the therapeutic efficacy of these sEV, leading to improved cartilage repair and reduced OA severity (Zhang et al., 2022). Additionally, sEV from BM-MSCs have been found to alleviate oxidative stress, modulate immune cell activity, and regulate the extracellular matrix composition, further contributing to cartilage protection and repair (Hotham et al., 2021; Wang X. et al., 2021; Contentin et al., 2022).
Similarly, AD-MSCs sEV have exhibited significant anti-inflammatory and cartilage-protective properties. Intra-articular injections of sEV from AD-MSCs have resulted in reduced cartilage destruction, increased expression of anabolic markers, and inhibition of inflammatory pathways (Ragni et al., 2020; Cavallo et al., 2022; Hanai et al., 2023). Some studies have further optimized sEV function through priming with bioactive molecules such as curcumin and insulin-like growth factor-1 (IGF-1), leading to enhanced therapeutic effects (Wu et al., 2022). Additionally, AD-MSC-derived sEV have been observed to modulate macrophage polarization, enhancing the regenerative environment within the joint (Ragni et al., 2021b).
In addition to MSC-derived sEV, synoviocyte-derived sEV have shown potential in OA therapy by delivering specific microRNAs that modulate inflammatory and apoptotic pathways (Filali et al., 2022; Wang and Sun, 2023; Ohtsuki et al., 2024). For example, miR-214-3p-enriched sEV from synoviocytes have demonstrated efficacy in reducing chondrocyte inflammation and preventing cartilage degradation (Ragni et al., 2019c). Other studies have indicated that synoviocyte-derived sEV may regulate immune responses by influencing T-cell activity and cytokine release, further contributing to OA mitigation (Farina et al., 2022; Zeng et al., 2022; Kouroupis et al., 2023; Nakamachi et al., 2023; Ossendorff et al., 2023; Mohd Noor et al., 2024).
sEV derived from UC-MSC are the most extensively studied sEV-based therapy for OA, showing greater potential compared to sEV from other sources such as bone marrow or adipose tissue (Tang et al., 2021; Morente-López et al., 2022). A recent pioneering clinical study has demonstrated that UC-MSC-derived sEV are not only safer and more secure than MSCs themselves but also more effective in modulating inflammation (Tang et al., 2021; Li et al., 2022; Nguyen et al., 2022; Zhou et al., 2022; Klyucherev et al., 2024; Figueroa-Valdés et al., 2025). Their therapeutic effect is largely mediated through the ERK signaling pathway, which regulates immune responses and cartilage regeneration (Morente-López et al., 2022).
Chondrocyte-derived sEV have also been extensively studied for their regenerative potential. These sEV have been reported to promote extracellular matrix synthesis and inhibit matrix degradation through mechanisms such as MMP13 modulation and STAT3 pathway inhibition (Yan et al., 2024). Furthermore, their role in maintaining chondrocyte phenotype and preventing hypertrophic differentiation highlights their potential in OA treatment. Research has suggested that chondrocyte-derived sEV may also interact with other joint-resident cells, such as fibroblasts and synoviocytes, to create a more favorable microenvironment for cartilage repair.
Other emerging sources of sEV, including those from iPSCs, macrophages, and neutrophils, have also exhibited promising OA-therapeutic properties. iPSC-derived MSCs sEV, for instance, have been shown to enhance cartilage repair and delay OA progression by maintaining chondrocyte homeostasis and reducing senescence (Hsueh et al., 2023). Moreover, macrophage-derived sEV have been implicated in immunomodulatory functions, promoting M2 macrophage polarization and mitigating synovitis (Song et al., 2024; Wang et al., 2024). Neutrophil-derived sEV, particularly those enriched with anti-inflammatory molecules such as secreted frizzled-related protein 5 (SFRP5) (Kitahara et al., 2024), have demonstrated the ability to counteract catabolic processes in the joint, further supporting their potential use in OA treatment.
There are some alternative sources of sEV offer a complementary approach to cell-derived extracellular vesicle therapies, expanding the possibilities for OA treatment. Bovine milk and garlic-derived sEV provide additional benefits by promoting cartilage homeostasis, regulating the gut microbiota, and reducing pain sensitivity (Pieters et al., 2022; Liu Q. et al., 2023; Liu et al., 2024 Y.).
The delivery of sEV has also been optimized through innovative approaches such as hydrogel encapsulation, microneedle delivery systems, and surface charge modifications, which have improved intra-articular retention and bioavailability (Tao et al., 2021; Yang et al., 2021; Li et al., 2024b; Li et al., 2024 Z.; Song et al., 2024; Zhou et al., 2024). Hydrogel systems, in particular, have been engineered to provide sustained release of sEV, extending their therapeutic effects within the joint. Additionally, researchers have explored the co-administration of sEV with growth factors or nanoparticles to enhance their regenerative properties. These advancements suggest that sEV-based therapies hold great promise for OA treatment by harnessing their regenerative, anti-inflammatory, and chondroprotective capabilities.
Moreover, recent pre-clinical studies have highlighted the importance of donor characteristics and pre-conditioning strategies in determining the efficacy of sEV (Visconte et al., 2024). For example, donor age has been identified as a critical factor influencing the therapeutic potential of BM-MSC-derived sEV, with younger donors exhibiting greater chondroprotective effects. Additionally, inflammatory pre-conditioning or genetic modifications of sEV-producing cells have been explored to enhance their bioactivity, offering new avenues for optimizing sEV-based therapies (Wakale et al., 2024).
Overall, pre-clinical studies indicate that sEV could serve as a viable therapeutic option for OA, paving the way for future clinical trials to further validate their safety and efficacy in human subjects. Given their ability to modulate key signaling pathways, regulate immune responses, and promote cartilage repair, sEV-based interventions represent a promising frontier in regenerative medicine for OA treatment.
4.4 sEV cargo as potential biomarkers in OA
The identification of reliable biomarkers for OA diagnosis, prognosis, and therapeutic monitoring remains a critical challenge. sEV have emerged as promising candidates due to their ability to encapsulate and transport bioactive molecules, including microRNAs, proteins, lipids, and cytokines, which reflect the pathological state of OA.
4.4.1 Nucleic acid cargo
Several studies highlight the role of miRNA profiles within sEV in OA. For instance, miRNA profiling of sEV derived from synovial fluid, cartilage, and BM-MSCs has revealed disease-specific signatures (Ragni et al., 2019b; 2019a; 2021a; Sanjurjo-Rodríguez et al., 2021; Filali et al., 2022; Ning et al., 2023; Asghar et al., 2024; Visconte et al., 2024). Ragni et al. have proposed the most abundant sEV-miRNAs from several OA affected-tissues as potential protective and destructive biomarkers. In cartilage, miR-24-3p and miR-193b-3p as protective and miR-21-5p and miR-16-5p as destructive; in synovium, miR-29a-3p (protective) and miR-34a-5p (destructive) (Ragni et al., 2022b). Sanjurjo-Rodríguez et al. reported that sEV-BM-MSCs derived from OA patients are enriched on miRNA involved in chondrogenesis, including as miR-142-3p, miR-630, miR-29a-3p, and miR-145-5p (Sanjurjo-Rodríguez et al., 2021). In vitro approximation using OA synovial fibroblasts have shown that these cells produce sEV with high content in miRNA-associated with osteoclastogenic process as miR-182, miR-4472-2, miR-1302-3, miR-6720, miR-6087 and miR-4532 (Asghar et al., 2024). The limitation of the miRNA cargo in sEV as a potential biomarker is the inexistence of reference genes in pathological tissues and cells. To solve this weakness, Ragni et al. have reported that miR-103a-3p and miR-22-5p have been identified as stable reference genes in sEV from various joint tissues (Ragni et al., 2021a), suggesting their potential as normalization controls in OA biomarker studies (Ragni et al., 2019b; 2019a). Additionally, integrative analyses of cartilage-derived sEV and single-cell RNA sequencing have uncovered specific miRNAs associated with OA progression, further supporting their diagnostic relevance. Ning et al. have identified miR-7-5p as one of the cartilage-derived sEV, that targets genes involved in the activation of the ferroptosis process and cartilage degradation in OA (Ning et al., 2023). This miRNA could be used as biomarker and potential therapeutic target to avoid the pathology progression.
Expanding on these insights, Liebmann et al. demonstrated that engineering IFP-derived MSCs with an adeno-associated virus (AAV) vector encoding a CGRP antagonist (CGRP8–37) yields sEV (aCGRP IFP-MSC sEV) with potent anti-inflammatory and analgesic properties. These vesicles carried a distinct miRNA and protein cargo involved in immune regulation, pain modulation, and cartilage repair, including miR-21-5p, miR-23a-3p, and proteins like TIMP-2 and IL-10. Functionally, they promoted M2 macrophage polarization and reduced neuroinflammation (Liebmann et al., 2024).
Beyond miRNAs, Circular RNAs (circRNAs), a novel class of endogenous covalently linked RNA molecules, were characterized by a closed continuous loop. CircRNAs are abundant, widespread, and tissue specific. CircRNAs have primary played a role as microRNA sponges. As evolutionarily conserved small noncoding RNAs (ncRNAs), miRNAs could inhibit gene expression by binding the base with the 3′untranslated region (3′-UTR) of mRNAs. Lin et al. have proposed that circPARD3B was down enriched in sEV from OA synovial (Lin et al., 2022).
4.4.2 Protein and lipid cargo
Proteomic analyses of sEV have revealed distinct protein cargo associated with OA severity and progression. iTRAQ proteomic analysis of exosomes derived from synovial fluid has identified disease-specific protein patterns (C3, C4B, APOM, MMP3, DPYSL2) that may serve as potential biomarkers (Wu X. et al., 2024). Similarly, OA modifies fatty acid signatures in synovial fluid from equine OA model and its sEV, indicating a metabolic shift that could be leveraged for early disease detection (Mustonen et al., 2023).
Inflammation plays a central role in OA pathogenesis, and sEV have been shown to carry key inflammatory mediators. Plasma-derived sEV carrying TNF-α have been identified as predictive biomarkers for knee OA progression (Zhang et al., 2021). Furthermore, immune-related plasma pathogenic EV subpopulations have been linked to OA progression, underscoring their potential as non-invasive biomarkers for disease monitoring (Zhang X. et al., 2024).
Surface marker profiling of plasma-derived sEV has provided valuable insights into OA severity. For instance, the expression levels of CD45, CD326, and CD56 on sEV correlate with OA stage, offering a novel diagnostic tool (Matejova et al., 2023). Additionally, tetraspanin profiles of serum sEV have been found to reflect functional limitations and pain perception in knee OA, demonstrating their utility in patient stratification and symptom monitoring (Mustonen et al., 2024).
The emerging evidence underscores the significant potential of sEV as biomarkers in OA. Their molecular cargo—ranging from miRNAs and proteins to lipids and inflammatory mediators—provides a comprehensive reflection of disease pathophysiology (Kolhe et al., 2020; Lin et al., 2022; Mustonen et al., 2022; Zhang et al., 2023). With advancements in high-throughput omics technologies and bioinformatics approaches, sEV-based biomarkers hold promise for improving OA diagnosis, prognostic assessment, and therapeutic response monitoring. Further validation in large-scale clinical studies will be essential to translate these findings into clinical practice.
4.5 Potential of sEV therapy in meniscus, tendons, ligaments, and muscle involvement in OA
Although the current literature on sEV therapy in OA predominantly focuses on cartilage, synovium, and subchondral bone, other periarticular structures such as the meniscus, ligaments, tendons, and muscles also play critical roles in joint homeostasis and OA progression.
The meniscus is essential for load distribution and joint stability. Meniscal damage is a well-established risk factor for OA, and the destabilization of the medial meniscus (DMM) model is widely used in preclinical OA research (Wang et al., 2020; Duan et al., 2021; Tao et al., 2021). While sEV have been tested in DMM-induced OA models, their direct regenerative or protective effects on meniscal tissue remain underexplored. Given the meniscus’s fibrocartilaginous nature, future studies should investigate whether sEVs can modulate meniscal cell metabolism, ECM remodeling, or inflammation.
Similarly, ligamentous structures, particularly the anterior cruciate ligament (ACL), are frequently involved in post-traumatic OA. Several studies in the review utilized the ACLT model (Rong et al., 2021; Cheng et al., 2024), which mimics joint instability and subsequent OA development. However, the potential of sEVs to enhance ligament healing or modulate ligament-derived inflammation has not been directly addressed. Given the regenerative potential of MSC-derived sEVs, their application in ligament repair warrants further investigation.
Tendons, though less frequently studied in OA models, are also susceptible to degeneration and inflammation in the OA joint environment. sEVs have shown promise in tendon healing in other musculoskeletal contexts, suggesting a translational opportunity for OA-related tendon pathologies.
Lastly, muscle weakness and atrophy, particularly of the quadriceps, are common in OA and contribute to joint dysfunction and pain. Although not a primary focus of OA sEV studies, the immunomodulatory and regenerative cargo of sEV, including miRNAs and growth factors, may have potential to support muscle regeneration and neuromuscular function, especially in advanced OA or post-surgical rehabilitation.
Incorporating these tissues into the sEV therapeutic landscape could provide a more holistic and functional approach to OA management, particularly in post-traumatic or multifactorial OA. Future studies should aim to characterize the effects of sEVs on these tissues and explore their potential in multitissue-targeted therapies.
5 Challenges and future perspectives
5.1 Challenges
5.1.1 Isolation and characterization
The standardization of sEV isolation and characterization techniques is a critical challenge. Existing methods such as ultracentrifugation, size-exclusion chromatography, and polymer-based precipitation vary widely in their yield, purity, and reproducibility (Hayami et al., 2006; Otahal et al., 2020; Mustonen et al., 2021; Sanjurjo-Rodríguez et al., 2021; Filali et al., 2022; Zhang et al., 2023). The heterogeneity of sEV populations further complicates their characterization, making it difficult to identify the most therapeutically active subpopulations (Figure 5a). Reliable and scalable methods for isolating specific sEV subsets are urgently needed to ensure consistency in clinical grade sEV preparations.
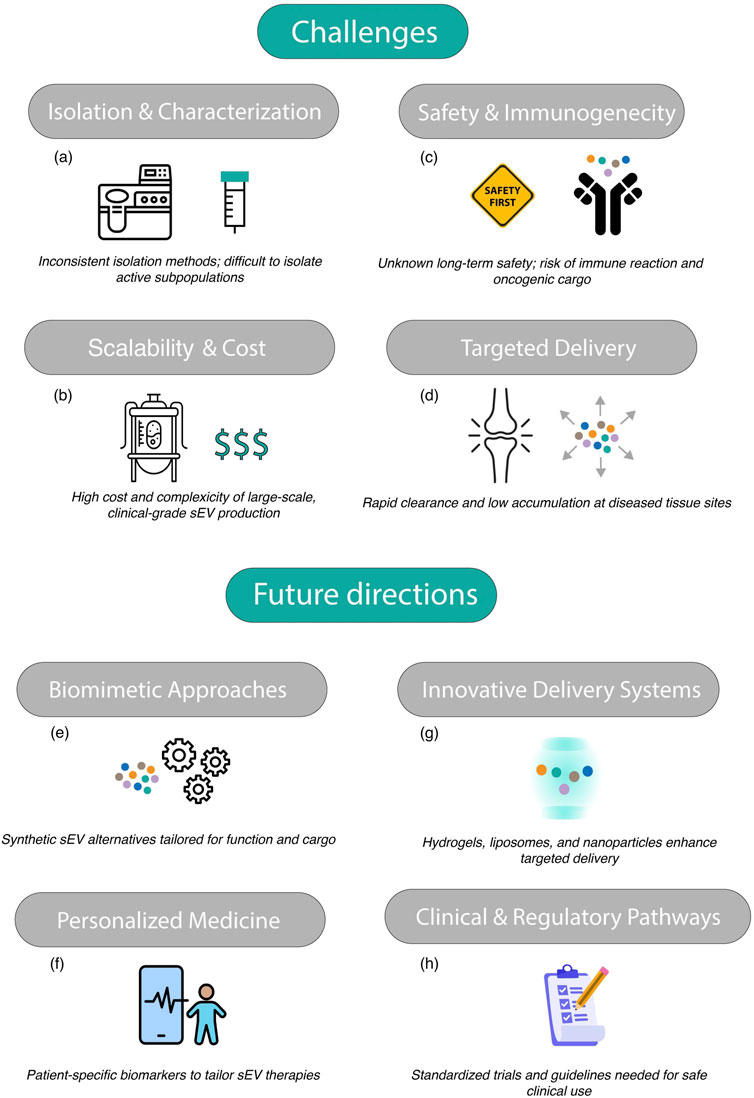
Figure 5. Graphical abstract of challenges and future directions of targeting OA with sEV. Challenges: (a) isolation and characterization, (b) scalability and cost, (c) safety and immunogenecity, (d) targeted delivery; Future directions: (e) Biomimetic approaches, (f) personalized medicine, (g) innovative delivery systems, (h) clinical and regulatory pathways.
5.1.2 Scalability and cost
Producing sEV on a large scale while maintaining their functional integrity is a significant barrier to their widespread adoption (Wang et al., 2020; Duan et al., 2021; Liu W. et al., 2023; Warmink et al., 2023; Asghar et al., 2024; Mustonen et al., 2024; Yan et al., 2024). The culture systems required for generating clinical-grade sEV are resource-intensive, and the downstream purification processes are time-consuming and costly (Figure 5b). For example, the use of bioreactors for large-scale sEV production can introduce additional complexity and variability. Achieving cost-effective scalability is critical for translating preclinical successes into accessible clinical therapies.
5.1.3 Safety and immunogenicity
While sEV are generally considered biocompatible, their long-term safety profiles in human applications remain unclear (Gupta et al., 2021; Ragni et al., 2022b; Kouroupis et al., 2023; Ossendorff et al., 2023). Concerns include potential off-target effects, such as unintended interactions with non-target tissues, and the risk of eliciting immune responses (Figure 5c). Additionally, the possibility of transferring pathogenic molecules, such as oncogenic miRNAs or viral particles, underscores the need for rigorous quality control and safety testing.
5.1.4 Delivery to target tissues
Efficiently delivering sEV to specific tissues, such as OA joints, remains challenging. After systemic administration, sEV are often rapidly cleared by the reticuloendothelial system, reducing their bioavailability at target sites (Fosang et al., 2000; Hayami et al., 2006; Otahal et al., 2020; Mustonen et al., 2021; Sanjurjo-Rodríguez et al., 2021; Xu et al., 2021; Filali et al., 2022; Wang et al., 2022; Zhang et al., 2023; Ohtsuki et al., 2024) (Figure 5d). Strategies to enhance the stability and retention of sEV in vivo are required to improve their therapeutic efficacy.
5.2 Future directions
5.2.1 Biomimetic approaches
One promising strategy to address scalability and production challenges is the development of biomimetic alternatives, such as artificial sEV or synthetic nanoparticles designed to replicate the functional properties of natural sEV (Wu et al., 2022; Jing et al., 2024; Kim et al., 2024; Yan et al., 2024) (Figure 5e). These engineered vesicles can be tailored to carry specific cargos, such as chondroprotective molecules or anti-inflammatory factors, while avoiding the complexities associated with isolating natural sEV.
5.2.2 Personalized medicine
Advances in omics technologies are paving the way for personalized sEV-based therapies. By identifying patient-specific biomarkers, it may be possible to design tailored sEV treatments that address the unique molecular and cellular characteristics of each patient’s condition (Madsen et al., 2010; Matejova et al., 2023; Wu X. et al., 2024) (Figure 5f). For example, sEV could be engineered to carry specific miRNAs or proteins that counteract disease-specific pathways, enhancing their therapeutic efficacy.
5.2.3 Innovative delivery systems
To improve the targeted delivery and bioavailability of sEV, innovative delivery systems such as hydrogels (Zhou et al., 2024), liposomes, or nanoparticle-based carriers are being explored (Hu et al., 2020; Kim et al., 2024). For instance, hydrogels can provide sustained release of sEV within osteoarthritic joints, enhancing their therapeutic effects (Figure 5g). Nanoparticle systems can also be functionalized with surface ligands to target specific cell types, further improving delivery precision.
5.2.4 Clinical trials and regulatory frameworks
The successful translation of sEV therapies into clinical practice requires robust clinical trials to establish their safety and efficacy (Figueroa-Valdés et al., 2025). Large-scale, randomized trials with standardized protocols are needed to generate high-quality evidence (Figure 5h). Additionally, the development of clear regulatory guidelines for the production, characterization, and clinical use of sEV will be critical for their widespread adoption.
6 Conclusion
sEV have emerged as a transformative tool in regenerative medicine, offering a multifaceted approach to addressing the challenges of OA and other degenerative diseases. By delivering bioactive molecules that promote chondrogenesis, modulate inflammation, and stimulate angiogenesis and tissue repair, sEV hold great promise for advancing the field of therapeutic interventions.
Despite these advancements, significant challenges remain on the path to clinical translation. Standardizing isolation techniques, achieving scalable production, ensuring safety, and developing efficient delivery systems are essential steps toward realizing the full therapeutic potential of sEV. Biomimetic technologies, personalized medicine, and advanced delivery strategies represent exciting avenues for overcoming these hurdles.
Looking forward, rigorous clinical trials and collaborative efforts among researchers, clinicians, and industry stakeholders will be instrumental in establishing sEV therapies as a cornerstone of modern medicine. As research continues to evolve, sEV are poised to revolutionize the treatment of osteoarthritis and other chronic conditions, offering hope for improved patient outcomes and quality of life.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
AG-R: Writing – review and editing, Writing – original draft. FD: Writing – review and editing. AJ-M: Writing – review and editing. PF-P: Writing – review and editing. CR: Writing – review and editing. MF: Writing – review and editing. JF-L: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review and editing. MA: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. JF-L was funded by Xunta de Galicia, grant number ED481D-2021-020, Ministerio de Ciencia e Innovación (RYC 2021-032567-I), funded by MCIN/AEI/10.13039/501100011033 and from the European Union «NextGenerationEU»/PRTR», InTalent program from UDC-Inditex for the research grant and Project PI23/01347, funded by Instituto de Salud Carlos III (ISCIII) and co-funded by the European Union. MCA has been funded by the project “PI20/00497,” funded by Instituto de Salud Carlos III (ISCIII) and co-funded by the European Union. JF-L and MA are funded by Xunta de Galicia (ED431F 2023/30). JF-L and MA were funded by Transference Project financed FINIBIC (RESISFERRO and ENDOPROT, respectively). We also thank Fundación Pública Galega de Investigación Biomédica INIBIC (A Coruña, Spain) for granting support for the publication of open-access articles (2024/2025).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ai, M., Hotham, W. E., Pattison, L. A., Ma, Q., Henson, F. M. D., and Smith, E. S. J. (2023). Role of human mesenchymal stem cells and derived extracellular vesicles in reducing sensory Neuron Hyperexcitability and pain behaviors in murine osteoarthritis. Arthritis Rheumatology 75, 352–363. doi:10.1002/art.42353
Asghar, S., Litherland, G. J., Cole, J. J., McInnes, I. B., Meek, R. M. D., Lockhart, J. C., et al. (2024). Small extracellular vesicles derived from synovial fibroblasts contain distinct miRNA profiles and contribute to chondrocyte damage in osteoarthritis. Arthritis Res. Ther. 26, 167. doi:10.1186/s13075-024-03398-3
Cai, L., Suboc, P., Hogue, D. A., Fei, D. T. W., and Filvaroff, E. H. (2002). Interleukin 17 induced nitric oxide suppresses matrix synthesis and protects cartilage from matrix breakdown. J. Rheumatology 29, 1725–1736.
Cai, L., Yin, J., Starovasnik, M. A., Hogue, D. A., Hillan, K. J., Mort, J. S., et al. (2001). Pathways by which interleukin 17 induces articular cartilage breakdown in vitro and in vivo. Cytokine 16, 10–21. doi:10.1006/cyto.2001.0939
Cao, H., Li, W., Zhang, H., Hong, L., Feng, X., Gao, X., et al. (2024a). Bio-nanoparticles loaded with synovial-derived exosomes ameliorate osteoarthritis progression by modifying the oxidative microenvironment. J. Nanobiotechnology 22, 271–15. doi:10.1186/s12951-024-02538-w
Cao, Y., Ruan, J., Kang, J., Nie, X., Lan, W., Ruan, G., et al. (2024b). Extracellular vesicles in Infrapatellar Fat Pad from osteoarthritis patients Impair cartilage metabolism and induce senescence. Adv. Sci. 11, e2303614–e2303615. doi:10.1002/advs.202303614
Cavallo, C., Merli, G., Zini, N., D’Adamo, S., Cattini, L., Guescini, M., et al. (2022). Small extracellular vesicles from inflamed adipose derived stromal cells enhance the NF- κ B-dependent inflammatory/catabolic environment of osteoarthritis. Stem Cells Int. 2022, 1–19. doi:10.1155/2022/9376338
Cawston, T. E., Curry, V. A., Summers, C. A., Clark, I. M., Riley, G. P., Life, P. F., et al. (1998). The role of oncostatin M in animal and human connective tissue collagen turnover and its localization within the rheumatoid joint. Arthritis Rheum. 41, 1760–1771. doi:10.1002/1529-0131(199810)41:10<1760::AID-ART8>3.0.CO;2-M
Chen, J., Tan, Y., Chen, Z., Yang, H., Li, X., Long, X., et al. (2024). Exosomes derived from primary cartilage stem/progenitor cells promote the repair of osteoarthritic chondrocytes by modulating immune responses. Int. Immunopharmacol. 143, 113397. doi:10.1016/j.intimp.2024.113397
Chen, P., Tang, S., Gao, H., Zhang, H., Chen, C., Fang, Z., et al. (2022a). Wharton’s jelly mesenchymal stem cell-derived small extracellular vesicles as natural nanoparticles to attenuate cartilage injury via microRNA regulation. Int. J. Pharm. 623, 121952. doi:10.1016/j.ijpharm.2022.121952
Chen, P., Zhou, J., Ruan, A., Guan, H., Xie, J., Zeng, L., et al. (2022b). Synovial tissue-derived extracellular vesicles induce chondrocyte inflammation and degradation via NF-κB signalling pathway: an in vitro study. J. Cell. Mol. Med. 26, 2038–2048. doi:10.1111/jcmm.17227
Cheng, S., Xu, X., Wang, R., Chen, W., Qin, K., and Yan, J. (2024). Chondroprotective effects of bone marrow mesenchymal stem cell-derived exosomes in osteoarthritis. J. Bioenerg. Biomembr. 56, 31–44. doi:10.1007/s10863-023-09991-6
Clarke, E. J., Johnson, E., Caamaño Gutierrez, E., Andersen, C., Berg, L. C., Jenkins, R. E., et al. (2022). Temporal extracellular vesicle protein changes following intraarticular treatment with integrin α10β1-selected mesenchymal stem cells in equine osteoarthritis. Front. Vet. Sci. 9, 1057667. doi:10.3389/fvets.2022.1057667
Clements, K. M., Flannelly, J. K., Tart, J., Brockbank, S. M. V., Wardale, J., Freeth, J., et al. (2011). Matrix metalloproteinase 17 is necessary for cartilage aggrecan degradation in an inflammatory environment. Ann. Rheum. Dis. 70, 683–689. doi:10.1136/ard.2010.130757
Contentin, R., Jammes, M., Bourdon, B., Cassé, F., Bianchi, A., Audigié, F., et al. (2022). Bone marrow MSC secretome increases equine articular chondrocyte collagen Accumulation and their migratory Capacities. Int. J. Mol. Sci. 23, 5795. doi:10.3390/ijms23105795
Das Neves Borges, P., Vincent, T. L., and Marenzana, M. (2017). Automated assessment of bone changes in cross-sectional micro-CT studies of murine experimental osteoarthritis. PLoS One 12, e0174294–22. doi:10.1371/journal.pone.0174294
Drummond, S. P., Bartnik, E., Kouvatsos, N., Scott, J. L., Dyer, D. P., Thomson, J. M., et al. (2023). The recombinant Link module of human TSG-6 suppresses cartilage damage in models of osteoarthritis: a potential disease-modifying OA drug. Osteoarthr. Cartil. 31, 1353–1364. doi:10.1016/j.joca.2023.05.013
Duan, A., Shen, K., Li, B., Li, C., Zhou, H., Kong, R., et al. (2021). Extracellular vesicles derived from LPS-preconditioned human synovial mesenchymal stem cells inhibit extracellular matrix degradation and prevent osteoarthritis of the knee in a mouse model. Stem Cell. Res. Ther. 12, 427–20. doi:10.1186/s13287-021-02507-2
Ebata, T., Terkawi, M. A., Kitahara, K., Yokota, S., Shiota, J., Nishida, Y., et al. (2023). Noncanonical pyroptosis triggered by macrophage-derived extracellular vesicles in chondrocytes leading to cartilage catabolism in osteoarthritis. Arthritis Rheumatology 75, 1358–1369. doi:10.1002/art.42505
Esmaeili, A., Hosseini, S., Kamali, A., Hosseinzadeh, M., Shekari, F., and Baghaban Eslaminejad, M. (2022). Co-aggregation of MSC/chondrocyte in a dynamic 3D culture elevates the therapeutic effect of secreted extracellular vesicles on osteoarthritis in a rat model. Sci. Rep. 12, 19827–15. doi:10.1038/s41598-022-22592-4
Farina, L., Minnone, G., Alivernini, S., Caiello, I., MacDonald, L., Soligo, M., et al. (2022). Pro nerve growth factor and its receptor p75NTR activate inflammatory responses in synovial fibroblasts: a novel targetable mechanism in arthritis. Front. Immunol. 13, 818630–13. doi:10.3389/fimmu.2022.818630
Fazaeli, H., Kalhor, N., Naserpour, L., Davoodi, F., Sheykhhasan, M., Hosseini, S. K. E., et al. (2021). A comparative study on the effect of exosomes secreted by mesenchymal stem cells derived from adipose and bone marrow tissues in the treatment of osteoarthritis-induced mouse model. Biomed. Res. Int. 2021, 9688138. doi:10.1155/2021/9688138
Feng, K., Liu, J., Gong, L., Ye, T., Chen, Z., Wang, Y., et al. (2025). Engineered MSC-sEVs as a Versatile Nanoplatform for enhanced osteoarthritis treatment via targeted elimination of senescent chondrocytes and Maintenance of cartilage matrix metabolic homeostasis. Adv. Sci. 12, e2413759–18. doi:10.1002/advs.202413759
Feng, K., Wang, F., Chen, H., Zhang, R., Liu, J., Li, X., et al. (2024a). Cartilage progenitor cells derived extracellular vesicles-based cell-free strategy for osteoarthritis treatment by efficient inflammation inhibition and extracellular matrix homeostasis restoration. J. Nanobiotechnology 22, 345–18. doi:10.1186/s12951-024-02632-z
Feng, K., Xie, X., Yuan, J., Gong, L., Zhu, Z., Zhang, J., et al. (2021). Reversing the surface charge of MSC-derived small extracellular vesicles by εPL-PEG-DSPE for enhanced osteoarthritis treatment. J. Extracell. Vesicles 10, e12160. doi:10.1002/jev2.12160
Feng, K., Ye, T., Xie, X., Liu, J., Gong, L., Chen, Z., et al. (2024b). ESC-sEVs alleviate non-early-stage osteoarthritis progression by rejuvenating senescent chondrocytes via FOXO1A-autophagy axis but not inducing apoptosis. Pharmacol. Res. 209, 107474. doi:10.1016/j.phrs.2024.107474
Figueroa-Valdés, A. I., Luz-Crawford, P., Herrera-Luna, Y., Georges-Calderón, N., García, C., Tobar, H. E., et al. (2025). Clinical-grade extracellular vesicles derived from umbilical cord mesenchymal stromal cells: preclinical development and first-in-human intra-articular validation as therapeutics for knee osteoarthritis. J. Nanobiotechnology 23, 13. doi:10.1186/s12951-024-03088-x
Filali, S., Darragi-Raies, N., Ben-Trad, L., Piednoir, A., Hong, S. S., Pirot, F., et al. (2022). Morphological and mechanical characterization of extracellular vesicles and parent human synoviocytes under physiological and inflammatory conditions. Int. J. Mol. Sci. 23, 13201–13222. doi:10.3390/ijms232113201
Filková, M., Vernerová, Z., Hulejová, H., Prajzlerová, K., Veigl, D., Pavelka, K., et al. (2015). Pro-inflammatory effects of interleukin-35 in rheumatoid arthritis. Cytokine 73, 36–43. doi:10.1016/j.cyto.2015.01.019
Forteza-Genestra, M. A., Antich-Rosselló, M., Ráez-Meseguer, C., Sangenís, A. T., Calvo, J., Gayà, A., et al. (2024). Intra-articular injection of platelet lysate-derived extracellular vesicles recovers from knee osteoarthritis in an in vivo rat model. J. Orthop. Transl. 45, 1–9. doi:10.1016/j.jot.2023.10.005
Fosang, A. J., Last, K., Stanton, H., Weeks, D. B., Campbell, I. K., Hardingham, T. E., et al. (2000). Generation and novel distribution of matrix metalloproteinase-derived aggrecan fragments in porcine cartilage explants. J. Biol. Chem. 275, 33027–33037. doi:10.1074/jbc.M910207199
Gendron, C., Kashiwagi, M., Hughes, C., Caterson, B., and Nagase, H. (2003). TIMP-3 inhibits aggrecanase-mediated glycosaminoglycan release from cartilage explants stimulated by catabolic factors. FEBS Lett. 555, 431–436. doi:10.1016/S0014-5793(03)01295-X
Genestra, M. A. F., Rosselló, M. A., Munar, G. R., Calvo, J., Gayà, A., Monjo, M., et al. (2023). Comparative effect of platelet- and mesenchymal stromal cell-derived extracellular vesicles on human cartilage explants using an ex vivo inflammatory osteoarthritis model. Bone Jt. Res. 12, 667–676. doi:10.1302/2046-3758.1210.BJR-2023-0109.R1
Gupta, A., Maffulli, N., Rodriguez, H. C., Mistovich, R. J., Delfino, K., Cady, C., et al. (2021). Cell-free stem cell-derived extract formulation for treatment of knee osteoarthritis: study protocol for a preliminary non-randomized, open-label, multi-center feasibility and safety study. J. Orthop. Surg. Res. 16, 514–517. doi:10.1186/s13018-021-02672-3
Hanai, H., Hart, D. A., Jacob, G., Shimomura, K., Ando, W., Yoshioka, Y., et al. (2023). Small extracellular vesicles derived from human adipose-derived mesenchymal stromal cells cultured in a new chemically-defined contaminate-free media exhibit enhanced biological and therapeutic effects on human chondrocytes in vitro and in a mouse osteoarthritis model. J. Extracell. Vesicles 12, e12337. doi:10.1002/jev2.12337
Harvanova, D., Matejova, J., Slovinska, L., Lacko, M., Gulova, S., Fecskeova, L. K., et al. (2022). The role of synovial membrane in the development of a potential in vitro model of osteoarthritis. Int. J. Mol. Sci. 23, 2475. doi:10.3390/ijms23052475
Haslauer, C. M., Elsaid, K. A., Fleming, B. C., Proffen, B. L., Johnson, V. M., and Murray, M. M. (2013). Loss of extracellular matrix from articular cartilage is mediated by the synovium and ligament after anterior cruciate ligament injury. Osteoarthr. Cartil. 21, 1950–1957. doi:10.1016/j.joca.2013.09.003
Hayami, T., Pickarski, M., Zhuo, Y., Wesolowski, G. A., Rodan, G. A., and Duong, L. T. (2006). Characterization of articular cartilage and subchondral bone changes in the rat anterior cruciate ligament transection and meniscectomized models of osteoarthritis. Bone 38, 234–243. doi:10.1016/j.bone.2005.08.007
Hosseinzadeh, M., Kamali, A., Baghaban Eslaminejad, M., and Hosseini, S. (2023). Higher ratios of chondrocyte to mesenchymal stem cells elevate the therapeutic effects of extracellular vesicles harvested from chondrocyte/mesenchymal stem cell co-culture on osteoarthritis in a rat model. Cell. Tissue Res. 394, 145–162. doi:10.1007/s00441-023-03819-w
Hotham, W. E., Thompson, C., Szu-Ting, L., and Henson, F. M. D. (2021). The anti-inflammatory effects of equine bone marrow stem cell-derived extracellular vesicles on autologous chondrocytes. Vet. Rec. Open 8, e22. doi:10.1002/vro2.22
Hsueh, Y. H., Buddhakosai, W., Le, P. N., Tu, Y. Y., Huang, H. C., Lu, H. E., et al. (2023). Therapeutic effect of induced pluripotent stem cell -derived extracellular vesicles in an in vitro and in vivo osteoarthritis model. J. Orthop. Transl. 38, 141–155. doi:10.1016/j.jot.2022.10.004
Hu, B., Gao, F., Li, C., Zhang, B., An, M., Lu, M., et al. (2020). Rhein laden pH-responsive polymeric nanoparticles for treatment of osteoarthritis. Amb. Express 10, 158. doi:10.1186/s13568-020-01095-3
Huang, X., You, Y., Xi, Y., Ni, B., Chu, X., Zhang, R., et al. (2020). p-Coumaric acid attenuates IL-1β-induced inflammatory responses and cellular senescence in rat chondrocytes. Inflammation 43, 619–628. doi:10.1007/s10753-019-01142-7
Jérémy, B., Marie, M., Giuliana, B. M., Karine, T., Christian, J., and Danièle, N. (2024). Extracellular vesicles from senescent mesenchymal stromal cells are defective and cannot prevent osteoarthritis. J. Nanobiotechnology 22, 1–18. doi:10.1186/s12951-024-02509-1
Jimenez, P. A., Glasson, S. S., Trubetskly, O. V., and Haimes, H. B. (1997). Spontaneous osteoarthritis in Dunkin Hartley Guinea pigs: histologic, radiologic, and biochemical changes. Lab. Anim. Sci. 47, 590–601.
Jing, Z., Zhang, G., Cai, Y., Liang, J., Lv, L., and Dang, X. (2024). Engineered extracellular vesicle-delivered TGF-β inhibitor for attenuating osteoarthritis by targeting subchondral bone. J. Tissue Eng. 15, 20417314241257781. doi:10.1177/20417314241257781
Kanakis, I., Liu, K., Poulet, B., Javaheri, B., van ’t Hof, R. J., Pitsillides, A. A., et al. (2019). Targeted inhibition of aggrecanases Prevents articular cartilage degradation and Augments bone Mass in the STR/Ort mouse model of spontaneous osteoarthritis. Arthritis Rheumatology 71, 571–582. doi:10.1002/art.40765
Karsdal, M. A., Madsen, S. H., Christiansen, C., Henriksen, K., Fosang, A. J., and Sondergaard, B. C. (2008). Cartilage degradation is fully reversible in the presence of aggrecanase but not matrix metalloproteinase activity. Arthritis Res. Ther. 10, R63–12. doi:10.1186/ar2434
Kim, J. Y., Rhim, W. K., Lee, S. Y., Park, J. M., Song, D. H., Cha, S. G., et al. (2024). Hybrid nanoparticle engineered with transforming growth factor -β1-Overexpressed extracellular vesicle and cartilage-targeted anti-inflammatory liposome for osteoarthritis. ACS Nano 18, 33937–33952. doi:10.1021/acsnano.4c07992
Kitahara, K., Ebata, T., Liyile, C., Nishida, Y., Ogawa, Y., Tokuhiro, T., et al. (2024). Chondroprotective functions of neutrophil-derived extracellular vesicles by promoting the production of secreted frizzled-related protein 5 in cartilage. Cell. Commun. Signal. 22, 569. doi:10.1186/s12964-024-01953-8
Klyucherev, T. O., Peshkova, M. A., Revokatova, D. P., Serejnikova, N. B., Fayzullina, N. M., Fayzullin, A. L., et al. (2024). The therapeutic potential of exosomes vs. Matrix-bound Nanovesicles from human umbilical cord mesenchymal stromal cells in osteoarthritis treatment. Int. J. Mol. Sci. 25, 11564. doi:10.3390/ijms252111564
Kolhe, R., Owens, V., Sharma, A., Lee, T. J., Zhi, W., Ghilzai, U., et al. (2020). Sex-specific differences in extracellular vesicle protein cargo in synovial fluid of patients with osteoarthritis. Life 10, 337–13. doi:10.3390/life10120337
Kouroupis, D., Kaplan, L. D., Huard, J., and Best, T. M. (2023). CD10-Bound human mesenchymal stem/stromal cell-derived small extracellular vesicles possess immunomodulatory cargo and maintain cartilage homeostasis under inflammatory conditions. Cells 12, 1824–23. doi:10.3390/cells12141824
Lai, C., Cheng, X., Yuan, T., Fang, P., Qian, H., Jiang, H., et al. (2024). A novel mechanism behind irreversible development of cartilage degradation driven articular cartilage defects revealed by rat model: the chain reaction initiated by extracellular vesicles delivered LOC102546541. Int. Immunopharmacol. 137, 112467. doi:10.1016/j.intimp.2024.112467
Lai, C., Liao, B., Peng, S., Fang, P., Bao, N., and Zhang, L. (2023). Synovial fibroblast-miR-214-3p-derived exosomes inhibit inflammation and degeneration of cartilage tissues of osteoarthritis rats. Mol. Cell. Biochem. 478, 637–649. doi:10.1007/s11010-022-04535-9
Lemarechal, H., Anract, P., Beadudex, J. L., Bonnefont-Rousselot, D., Ekindjian, O. G., and Borderie, D. (2007). Expression and extracellular release of Trx80, the truncated form of thioredoxin, by TNF-alpha- and IL-1beta-stimulated human synoviocytes from patients with rheumatoid arthritis. Clin. Sci. 113, 149–155. doi:10.1042/CS20070067
Li, B., Zhang, J., Zhang, B., Wang, Y., Lu, T., Li, J., et al. (2024a). Adipose tissue-derived extracellular vesicles aggravate temporomandibular joint osteoarthritis associated with obesity. Clin. Transl. Med. 14, e70029. doi:10.1002/ctm2.70029
Li, K., Yan, G., Huang, H., Zheng, M., Ma, K., Cui, X., et al. (2022). Anti-inflammatory and immunomodulatory effects of the extracellular vesicles derived from human umbilical cord mesenchymal stem cells on osteoarthritis via M2 macrophages. J. Nanobiotechnology 20, 38–20. doi:10.1186/s12951-021-01236-1
Li, S., Liu, J., Liu, S., Jiao, W., and Wang, X. (2021). Mesenchymal stem cell-derived extracellular vesicles prevent the development of osteoarthritis via the circHIPK3/miR-124-3p/MYH9 axis. J. Nanobiotechnology 19, 194–20. doi:10.1186/s12951-021-00940-2
Li, S., Yuan, Q., Yang, M., Long, X., Sun, J., Yuan, X., et al. (2024b). Enhanced cartilage regeneration by icariin and mesenchymal stem cell-derived extracellular vesicles combined in alginate-hyaluronic acid hydrogel. Nanomedicine 55, 102723. doi:10.1016/j.nano.2023.102723
Li, S., Zheng, W., Deng, W., Li, Z., Yang, J., Zhang, H., et al. (2024c). Logic-based strategy for Spatiotemporal release of dual extracellular vesicles in osteoarthritis treatment. Adv. Sci. 11, e2403227–17. doi:10.1002/advs.202403227
Li, W. Q., Qureshi, H. Y., Liacini, A., Dehnade, F., and Zafarullah, M. (2004). Transforming growth factor β1 induction of Tissue Inhibitor of MetalloProteinases 3 in articular chondrocytes is mediated by reactive oxygen species. Free Radic. Biol. Med. 37, 196–207. doi:10.1016/j.freeradbiomed.2004.04.028
Li, Z., Lu, H., Fan, L., Ma, X., Duan, Z., Zhang, Y., et al. (2024d). Microneedle-Delivered PDA@Exo for multifaceted osteoarthritis treatment via PI3K-Akt-mTOR pathway. Adv. Sci. 11, e2406942–12. doi:10.1002/advs.202406942
Liao, L., Zhang, S., Zhao, L., Chang, X., Han, L., Huang, J., et al. (2020). Acute synovitis after trauma precedes and is associated with osteoarthritis onset and progression. Int. J. Biol. Sci. 16, 970–980. doi:10.7150/ijbs.39015
Liao, S., Cheng, L., Zhao, Z., Zhy, J., and Huang, F. (2024). Distinct characteristics and progression patterns of facet joint structural lesions in radiographic axial spondyloarthritis. Ther. Adv. Musculoskelet. Dis. 9, 1759720X241281201–10. doi:10.1177/1759720x241281201
Liebmann, K., Castillo, M. A., Jergova, S., Best, T. M., Sagen, J., and Kouroupis, D. (2024). Modification of mesenchymal stem/stromal cell-derived small extracellular vesicles by Calcitonin gene related peptide (CGRP) antagonist: potential Implications for inflammation and pain reversal. Cells 13, 484. doi:10.3390/cells13060484
Lin, Z., Ma, Y., Zhu, X., Dai, S., Sun, W., Li, W., et al. (2022). Potential predictive and therapeutic applications of small extracellular vesicles-derived circPARD3B in osteoarthritis. Front. Pharmacol. 13, 968776–16. doi:10.3389/fphar.2022.968776
Lin, T., Wu, N., Wang, L., Zhang, R., Pan, R., and Chen, Y. F. (2021). Inhibition of chondrocyte apoptosis in a rat model of osteoarthritis by exosomes derived from miR-140-5p-overexpressing human dental pulp stem cells. Int. J. Mol. Med. 47, 7–10. doi:10.3892/ijmm.2020.4840
Liu, Q., Hao, H., Li, J., Zheng, T., Yao, Y., Tian, X., et al. (2023a). Oral administration of Bovine milk-derived extracellular vesicles attenuates cartilage degeneration via modulating gut microbiota in DMM-induced mice. Nutrients 15, 747. doi:10.3390/nu15030747
Liu, Q., Wu, J., Wang, H., Jia, Z., and Li, G. (2024a). Human Infrapatellar Fat Pad mesenchymal stem cell-derived extracellular vesicles purified by anion Exchange chromatography suppress osteoarthritis progression in a mouse model. Clin. Orthop. Relat. Res. 482, 1246–1262. doi:10.1097/CORR.0000000000003067
Liu, W., Liu, A., Li, X., Sun, Z., Sun, Z., Liu, Y., et al. (2023b). Dual-engineered cartilage-targeting extracellular vesicles derived from mesenchymal stem cells enhance osteoarthritis treatment via miR-223/NLRP3/pyroptosis axis: toward a precision therapy. Bioact. Mater 30, 169–183. doi:10.1016/j.bioactmat.2023.06.012
Liu, Y., Nie, M., Li, X., Wang, H., Ren, S., Zou, D., et al. (2024b). Garlic-derived exosomes alleviate osteoarthritis through inhibiting the MAPK signaling pathway. Appl. Biochem. Biotechnol. 197, 518–533. doi:10.1007/s12010-024-05047-6
Lu, H., Yang, Y., Ou, S., Qi, Y., Li, G., He, H., et al. (2022). miRNA-382-5p carried by extracellular vesicles in osteoarthritis reduces cell Viability and proliferation, and promotes cell apoptosis by targeting PTEN. DNA Cell. Biol. 41, 1012–1025. doi:10.1089/dna.2021.0726
Lu, L., Wang, J., Fan, A., Wang, P., Chen, R., Lu, L., et al. (2021). Synovial mesenchymal stem cell-derived extracellular vesicles containing microRN555A-26a-5p ameliorate cartilage damage of osteoarthritis. J. Gene Med. 23, e3379–14. doi:10.1002/jgm.3379
Madsen, S. H., Sumer, E. U., Bay-Jensen, A. C., Sondergaard, B. C., Qvist, P., and Karsdal, M. A. (2010). Aggrecanase- and matrix metalloproteinase-mediated aggrecan degradation is associated with different molecular characteristics of aggrecan and separated in time ex vivo. Biomarkers 15, 266–276. doi:10.3109/13547500903521810
Maeda, S., Dean, D. D., Sylvia, V. L., Boyan, B. D., and Schwartz, Z. (2001). Metalloproteinase activity in growth plate chondrocyte cultures is regulated by 1,25-(OH)2D3 and 24,25-(OH)2D3 and mediated through protein kinase C. Matrix Biol. 20, 87–97. doi:10.1016/S0945-053X(01)00123-8
Matejova, J., Fecskeova, L. K., Slovinska, L., Harvanova, D., Spakova, T., and Bzdilova, J. (2023). Plasma-derived extracellular vesicle surface markers CD45, CD326 and CD56 correlate with the stage of osteoarthritis: a primary study of a novel and promising diagnostic tool of the disease. Sci. Rep. 13, 20071–14. doi:10.1038/s41598-023-47074-z
Mladenovic, Z., Saurel, A. S., Berenbaum, F., and Jacques, C. (2014). Potential role of hyaluronic acid on bone in osteoarthritis: matrix metalloproteinases, aggrecanases, and RANKL expression are partially prevented by hyaluronic acid in interleukin 1-stimulated osteoblasts. J. Rheumatology 41, 945–954. doi:10.3899/jrheum.130378
Mohd Noor, N. A., Quan, N. J., Adibah Mazlan, N. A., Abdullah Nurul, A., Mohd Zain, M. R. A., and Azlan, M. (2024). Synovial fluid-derived exosomes from osteoarthritis patients modulate cell surface phenotypes of Monocytes and cytokine Secretions. Immunol. Investig. 19, 1–16. doi:10.1080/08820139.2024.2443244
Moncada-Pazos, A., Obaya, A. J., Viloria, C. G., López-Otín, C., and Cal, S. (2011). The nutraceutical flavonoid luteolin inhibits ADAMTS-4 and ADAMTS-5 aggrecanase activities. J. Mol. Med. 89, 611–619. doi:10.1007/s00109-011-0741-7
Morente-López, M., Mato-Basalo, R., Lucio-Gallego, S., Silva-Fernández, L., González-Rodríguez, A., De Toro, F. J., et al. (2022). Therapy free of cells vs human mesenchymal stem cells from umbilical cord stroma to treat the inflammation in OA. Cell. Mol. Life Sci. 79, 557. doi:10.1007/s00018-022-04580-z
Mustonen, A. M., Capra, J., Rilla, K., Lehenkari, P., Oikari, S., Kääriäinen, T., et al. (2021). Characterization of hyaluronan-coated extracellular vesicles in synovial fluid of patients with osteoarthritis and rheumatoid arthritis. BMC Musculoskelet. Disord. 22, 247–11. doi:10.1186/s12891-021-04115-w
Mustonen, A. M., Lehmonen, N., Oikari, S., Capra, J., Raekallio, M., Mykkänen, A., et al. (2022). Counts of hyaluronic acid-containing extracellular vesicles decrease in naturally occurring equine osteoarthritis. Sci. Rep. 12, 17550–12. doi:10.1038/s41598-022-21398-8
Mustonen, A. M., Lehmonen, N., Paakkonen, T., Raekallio, M., Käkelä, R., Niemelä, T., et al. (2023). Equine osteoarthritis modifies fatty acid signatures in synovial fluid and its extracellular vesicles. Arthritis Res. Ther. 25, 39–11. doi:10.1186/s13075-023-02998-9
Mustonen, A. M., Palviainen, M., Säisänen, L., Karttunen, L., Tollis, S., Esrafilian, A., et al. (2024). Tetraspanin profiles of serum extracellular vesicles reflect functional limitations and pain perception in knee osteoarthritis. Arthritis Res. Ther. 26, 33–16. doi:10.1186/s13075-023-03234-0
Nakamachi, Y., Uto, K., Hayashi, S., Okano, T., Morinobu, A., Kuroda, R., et al. (2023). Exosomes derived from synovial fibroblasts from patients with rheumatoid arthritis promote macrophage migration that can be suppressed by miR-124-3p. Heliyon 9, e14986. doi:10.1016/j.heliyon.2023.e14986
Nguyen, T. H., Dao, H. H., Duong, C. M., Nguyen, X. H., Hoang, D. H., Do, X. H., et al. (2022). Cytokine-primed umbilical cord mesenchymal stem cells enhanced therapeutic effects of extracellular vesicles on osteoarthritic chondrocytes. Front. Immunol. 13, 1041592–12. doi:10.3389/fimmu.2022.1041592
Niedermair, T., Lukas, C., Li, S., Stöckl, S., Craiovan, B., Brochhausen, C., et al. (2020). Influence of extracellular vesicles isolated from osteoblasts of patients with Cox-Arthrosis and/or Osteoporosis on metabolism and osteogenic differentiation of BMSCs. Front. Bioeng. Biotechnol. 8, 615520–15. doi:10.3389/fbioe.2020.615520
Ning, Y., Zhang, F., Li, S., Wang, C., Wu, Y., Chen, S., et al. (2023). Integrative analysis of miRNA in cartilage-derived extracellular vesicles and single-cell RNA-seq profiles in knee osteoarthritis. Arch. Biochem. Biophys. 748, 109785. doi:10.1016/j.abb.2023.109785
Ohtsuki, T., Sato, I., Takashita, R., Kodama, S., Ikemura, K., Opoku, G., et al. (2024). Distribution and Incorporation of extracellular vesicles into chondrocytes and synoviocytes. Int. J. Mol. Sci. 25, 11942–12. doi:10.3390/ijms252211942
Ossendorff, R., Grad, S., Tertel, T., Wirtz, D. C., Giebel, B., Börger, V., et al. (2023). Immunomodulatory potential of mesenchymal stromal cell-derived extracellular vesicles in chondrocyte inflammation. Front. Immunol. 14, 1198198–12. doi:10.3389/fimmu.2023.1198198
Otahal, A., Kramer, K., Kuten-pella, O., Moser, L. B., Neubauer, M., Lacza, Z., et al. (2021). Effects of extracellular vesicles from blood-derived products on osteoarthritic chondrocytes within an inflammation model. Int. J. Mol. Sci. 22, 1–15. doi:10.3390/ijms22137224
Otahal, A., Kramer, K., Kuten-Pella, O., Weiss, R., Stotter, C., Lacza, Z., et al. (2020). Characterization and chondroprotective effects of extracellular vesicles from plasma- and serum-based autologous blood-derived products for osteoarthritis therapy. Front. Bioeng. Biotechnol. 8, 584050–13. doi:10.3389/fbioe.2020.584050
Pan, C., Huang, W., Chen, Q., Xu, J., Yao, G., Li, B., et al. (2021). LncRNA Malat-1 from MSCs-derived extracellular vesicles suppresses inflammation and cartilage degradation in osteoarthritis. Front. Bioeng. Biotechnol. 9, 772002–772012. doi:10.3389/fbioe.2021.772002
Park, D. Y., Min, B. H., Kim, D. W., Song, B. R., Kim, M., and Kim, Y. J. (2013). Polyethylene wear particles play a role in development of osteoarthritis via detrimental effects on cartilage, meniscus, and synovium. Osteoarthr. Cartil. 21, 2021–2029. doi:10.1016/j.joca.2013.09.013
Patel, K. P., Sandy, J. D., Akeda, K., Miyamoto, K., Chujo, T., Howard, S. A., et al. (2007). Aggrecanases and aggrecanase-generated fragments in the human intervertebral disc at early and advanced stages of disc degeneration. Spine (Phila Pa 1976) 32, 2596–2603. doi:10.1097/BRS.0b013e318158cb85
Patwari, P., Kurz, B., Sandy, J. D., and Grodzinsky, A. J. (2000). Mannosamine inhibits aggrecanase-mediated changes in the physical properties and biochemical composition of articular cartilage. Arch. Biochem. Biophys. 374, 79–85. doi:10.1006/abbi.1999.1538
Pfeifer, J. P. H., Stievani, F. de C., Fernandes, C. J. da C., Rosa, G., dos, S., Apolonio, E. V. P., et al. (2024). Influence of inflammation on the expression of microRNA-140 in extracellular vesicles from 2D and 3D culture models of synovial-membrane-derived stem cells. Front. Bioeng. Biotechnol. 12, 1416694–16. doi:10.3389/fbioe.2024.1416694
Pickarski, M., Hayami, T., Zhuo, Y., and Duong, L. T. (2011). Molecular changes in articular cartilage and subchondral bone in the rat anterior cruciate ligament transection and meniscectomized models of osteoarthritis. BMC Musculoskelet. Disord. 12, 197. doi:10.1186/1471-2474-12-197
Pieters, B. C. H., Arntz, O. J., Aarts, J., Feitsma, A. L., van Neerven, R. J. J., van der Kraan, P. M., et al. (2022). Bovine milk-derived extracellular vesicles inhibit catabolic and inflammatory processes in cartilage from osteoarthritis patients. Mol. Nutr. Food Res. 66, e2100764–12. doi:10.1002/mnfr.202100764
Ragni, E., Colombini, A., De Luca, P., Libonati, F., Viganò, M., Perucca Orfei, C., et al. (2021a). miR-103a-3p and miR-22-5p are reliable reference genes in extracellular vesicles from cartilage, adipose tissue, and bone marrow cells. Front. Bioeng. Biotechnol. 9, 632440–15. doi:10.3389/fbioe.2021.632440
Ragni, E., Colombini, A., Viganò, M., Libonati, F., Perucca Orfei, C., Zagra, L., et al. (2021b). Cartilage protective and immunomodulatory features of osteoarthritis synovial fluid-treated adipose-derived mesenchymal stem cells secreted factors and extracellular vesicles-embedded mirnas. Cells 10, 1072. doi:10.3390/cells10051072
Ragni, E., De Luca, P., Landoni, S., Valli, F., Mortati, L., Palombella, S., et al. (2024). High efficiency protocol for platelet derived fibrin gel loaded with mesenchymal stromal cells extracellular vesicles. Regen. Ther. 26, 442–457. doi:10.1016/j.reth.2024.06.020
Ragni, E., De Luca, P., Orfei, C. P., Colombini, A., Viganò, M., Lugano, G., et al. (2019a). Insights into inflammatory priming of adipose-derived mesenchymal stem cells: validation of extracellular vesicles-embedded miRNA reference genes as a crucial step for donor selection. Cells 8, 1–17. doi:10.3390/cells8040369
Ragni, E., Orfei, C. P., De Luca, P., Colombini, A., Viganò, M., and de Girolamo, L. (2020). Secreted factors and ev-mirnas orchestrate the healing capacity of adipose mesenchymal stem cells for the treatment of knee osteoarthritis. Int. J. Mol. Sci. 21, 1582. doi:10.3390/ijms21051582
Ragni, E., Orfei, C. P., De Luca, P., Colombini, A., Viganò, M., Lugano, G., et al. (2019b). Identification of miRNA reference genes in extracellular vesicles from adipose derived mesenchymal stem cells for studying osteoarthritis. Int. J. Mol. Sci. 20, 1108. doi:10.3390/ijms20051108
Ragni, E., Orfei, C. P., Sinigaglia, F., and de Girolamo, L. (2022a). Joint tissue protective and immune-modulating miRNA landscape of mesenchymal stromal cell-derived extracellular vesicles under different osteoarthritis-Mimicking conditions. Pharmaceutics 14, 1400. doi:10.3390/pharmaceutics14071400
Ragni, E., Perucca Orfei, C., and de Girolamo, L. (2022b). Secreted factors and extracellular vesicles Account for the immunomodulatory and tissue regenerative properties of bone-marrow-derived mesenchymal stromal cells for osteoarthritis. Cells 11, 3501. doi:10.3390/cells11213501
Ragni, E., Perucca Orfei, C., De Luca, P., Lugano, G., Viganò, M., Colombini, A., et al. (2019c). Interaction with hyaluronan matrix and miRNA cargo as contributors for in vitro potential of mesenchymal stem cell-derived extracellular vesicles in a model of human osteoarthritic synoviocytes. Stem Cell. Res. Ther. 10, 109–117. doi:10.1186/s13287-019-1215-z
Rong, Y., Zhang, J., Jiang, D., Ji, C., liu, W., Wang, J., et al. (2021). Hypoxic pretreatment of small extracellular vesicles mediates cartilage repair in osteoarthritis by delivering miR-216a-5p. Acta Biomater. 122, 325–342. doi:10.1016/j.actbio.2020.12.034
Ruan, X., Gu, J., Chen, M., Zhao, F., Aili, M., and Zhang, D. (2023). Multiple roles of ALK3 in osteoarthritis A NARRATIVE REVIEW. Bone Jt. Res. 12, 397–411. doi:10.1302/2046-3758.127.BJR-2022-0310.R1
Ruiz, M., Toupet, K., Maumus, M., Rozier, P., Jorgensen, C., and Noël, D. (2020). TGFBI secreted by mesenchymal stromal cells ameliorates osteoarthritis and is detected in extracellular vesicles. Biomaterials 226, 119544. doi:10.1016/j.biomaterials.2019.119544
Sanjurjo-Rodríguez, C., Crossland, R. E., Reis, M., Pandit, H., Wang, X. N., and Jones, E. (2021). Characterization and miRNA profiling of extracellular vesicles from human osteoarthritic subchondral bone Multipotential stromal cells (MSCs). Stem Cells Int. 2021, 1–16. doi:10.1155/2021/7232773
Scalzone, A., Sanjurjo-Rodríguez, C., Berlinguer-Palmini, R., Dickinson, A. M., Jones, E., Wang, X. N., et al. (2024). Functional and molecular analysis of human osteoarthritic chondrocytes treated with bone marrow-derived MSC-EVs. Bioengineering 11, 1–22. doi:10.3390/bioengineering11040388
Shang, X., Böker, K. O., Taheri, S., Lehmann, W., and Schilling, A. F. (2021). Extracellular vesicles allow epigenetic mechanotransduction between chondrocytes and osteoblasts. Int. J. Mol. Sci. 22, 13282. doi:10.3390/ijms222413282
Siebelt, M., Groen, H. C., Koelewijn, S. J., de Blois, E., Sandker, M., Waarsing, J. H., et al. (2014). Increased physical activity severely induces osteoarthritic changes in knee joints with papain induced sulfate-glycosaminoglycan depleted cartilage. Arthritis Res. Ther. 16, R32–12. doi:10.1186/ar4461
Sofat, N., Robertson, S. D., Hermansson, M., Jones, J., Mitchell, P., and Wait, R. (2012). Tenascin-C fragments are endogenous inducers of cartilage matrix degradation. Rheumatol. Int. 32, 2809–2817. doi:10.1007/s00296-011-2067-8
Song, X., Xiao, J., Ai, X., Li, Y., Sun, L., and Chen, L. (2024). An injectable thermosensitive hydrogel delivering M2 macrophage-derived exosomes alleviates osteoarthritis by promoting synovial lymphangiogenesis. Acta Biomater. 189, 130–142. doi:10.1016/j.actbio.2024.09.034
Stanton, H., Ung, L., and Fosang, A. J. (2002). The 45 kDa collagen-binding fragment of fibronectin induces matrix metalloproteinase-13 synthesis by chondrocytes and aggrecan degradation by aggrecanases. Biochem. J. 364, 181–190. doi:10.1042/bj3640181
Sun, Y., Zhao, J., Wu, Q., Zhang, Y., You, Y., Jiang, W., et al. (2022). Chondrogenic primed extracellular vesicles activate miR-455/SOX11/FOXO axis for cartilage regeneration and osteoarthritis treatment. NPJ Regen. Med. 7, 53. doi:10.1038/s41536-022-00250-7
Tang, S., Chen, P., Zhang, H., Weng, H., Fang, Z., Chen, C., et al. (2021). Comparison of curative effect of human umbilical cord-derived mesenchymal stem cells and their small extracellular vesicles in treating osteoarthritis. Int. J. Nanomedicine 16, 8185–8202. doi:10.2147/IJN.S336062
Tang, X., He, J., and Hao, Y. (2024a). Extracellular vesicles derived Ectonucleoside Triphosphate Diphosphohydrolase 3 alleviates Mitochondrial dysfunction of osteoarthritis chondrocytes via Ectonucleotide pyrophosphatase/phosphodiesterase 1-induced suppression of the AKT/Notch2 pathway. J. Biochem. Mol. Toxicol. 38, e70064–15. doi:10.1002/jbt.70064
Tang, Z., Shu, L., Cao, Z., Xu, Y., and Li, C. (2024b). Osteoarthritis rat serum-derived extracellular vesicles aggravate osteoarthritis development by inducing NLRP3-mediated pyroptotic cell death and cellular inflammation. Hum. Cell. 37, 1624–1637. doi:10.1007/s13577-024-01119-1
Tao, S. C., Huang, J. Y., Gao, Y., Li, Z. X., Wei, Z. Y., Dawes, H., et al. (2021). Small extracellular vesicles in combination with sleep-related circRNA3503: a targeted therapeutic agent with injectable thermosensitive hydrogel to prevent osteoarthritis. Bioact. Mater 6, 4455–4469. doi:10.1016/j.bioactmat.2021.04.031
Thomas, M. A., Fahey, M. J., Pugliese, B. R., Irwin, R. M., Antonyak, M. A., and Delco, M. L. (2022). Human mesenchymal stromal cells release functional mitochondria in extracellular vesicles. Front. Bioeng. Biotechnol. 10, 870193–12. doi:10.3389/fbioe.2022.870193
Tsushima, H., Okazaki, K., Hayashida, M., Ushijima, T., and Iwamoto, Y. (2012). CCAAT/enhancer binding protein β regulates expression of matrix metalloproteinase-3 in arthritis. Ann. Rheum. Dis. 71, 99–107. doi:10.1136/annrheumdis-2011-200061
Van Lent, P. L. E. M., Grevers, L. C., Blom, A. B., Arntz, O. J., Van De Loo, F. A. J., Van Der Kraan, P., et al. (2008). Stimulation of chondrocyte-mediated cartilage destruction by S100A8 in experimental murine arthritis. Arthritis Rheum. 58, 3776–3787. doi:10.1002/art.24074
Visconte, C., Taiana, M. M., Colombini, A., De Luca, P., Ragni, E., and de Girolamo, L. (2024). Donor sites and Harvesting techniques Affect miRNA cargos of extracellular vesicles released by human adipose-derived mesenchymal stromal cells. Int. J. Mol. Sci. 25, 6450. doi:10.3390/ijms25126450
Wakale, S., Chen, Y., Sun, A. R., Liyanage, C., Gunter, J., Batra, J., et al. (2024). Comparative analysis of the therapeutic potential of extracellular vesicles secreted by aged and Young bone marrow-derived mesenchymal stem cells in osteoarthritis pathogenesis. Cell. Prolif. 58, e13776–18. doi:10.1111/cpr.13776
Wang, H., Shu, J., Zhang, C., Wang, Y., Shi, R., Yang, F., et al. (2022). Extracellular vesicle-mediated miR-150-3p delivery in joint homeostasis: a potential treatment for osteoarthritis? Cells 11, 2766. doi:10.3390/cells11172766
Wang, J., and Sun, T. (2023). Mir-25-3p in extracellular vesicles from fibroblast-like synoviocytes alleviates pyroptosis of chondrocytes in knee osteoarthritis. J. Bioenerg. Biomembr. 55, 365–380. doi:10.1007/s10863-023-09964-9
Wang, R., Jiang, W., Zhang, L., Xie, S., Zhang, S., Yuan, S., et al. (2020). Intra-articular delivery of extracellular vesicles secreted by chondrogenic progenitor cells from MRL/MpJ superhealer mice enhances articular cartilage repair in a mouse injury model. Stem Cell. Res. Ther. 11, 93–14. doi:10.1186/s13287-020-01594-x
Wang, T., Zhao, H., Zhang, Y., Liu, Y., Liu, J., Chen, G., et al. (2024). A novel extracellular vesicles production system harnessing matrix homeostasis and macrophage reprogramming mitigates osteoarthritis. J. Nanobiotechnology 22, 79–17. doi:10.1186/s12951-024-02324-8
Wang, X., Li, Z., Cui, Y., Cui, X., Chen, C., and Wang, Z. (2021a). Exosomes isolated from bone marrow mesenchymal stem cells exert a protective effect on osteoarthritis via lncRNA LYRM4-AS1-GRPR-miR-6515-5p. Front. Cell. Dev. Biol. 9, 644380–15. doi:10.3389/fcell.2021.644380
Wang, X. D., Kou, X. X., He, D. Q., Zeng, M. M., Meng, Z., Bi, R. Y., et al. (2012). Progression of cartilage degradation, bone resorption and pain in rat temporomandibular joint osteoarthritis induced by injection of Iodoacetate. PLoS One 7, e45036–12. doi:10.1371/journal.pone.0045036
Wang, Y., Zhao, M., Li, W., Yang, Y., Zhang, Z., Ma, R., et al. (2021b). BMSC-derived small extracellular vesicles induce cartilage Reconstruction of temporomandibular joint osteoarthritis via Autotaxin–YAP signaling Axis. Front. Cell. Dev. Biol. 9, 656153–16. doi:10.3389/fcell.2021.656153
Warmink, K., Rios, J. L., Varderidou-Minasian, S., Torres-Torrillas, M., van Valkengoed, D. R., Versteeg, S., et al. (2023). Mesenchymal stem/stromal cells-derived extracellular vesicles as a potentially more beneficial therapeutic strategy than MSC-based treatment in a mild metabolic osteoarthritis model. Stem Cell. Res. Ther. 14, 137–15. doi:10.1186/s13287-023-03368-7
Weston, S., Macdonald, J. L., Williams, L. M., Roussou, E., Kang, N. V., Kiriakidis, S., et al. (2022). The JAK inhibitor baricitinib inhibits oncostatin M induction of proinflammatory mediators in ex-vivo synovial derived cells. Clin. Exp. Rheumatol. 40, 1620–1628. doi:10.55563/clinexprheumatol/cfsajk
Wilkinson, D. J., Habgood, A., Lamb, H. K., Thompson, P., Hawkins, A. R., Désilets, A., et al. (2017). Matriptase induction of metalloproteinase-dependent Aggrecanolysis in vitro and in vivo: Promotion of osteoarthritic cartilage damage by multiple mechanisms. Arthritis Rheumatology 69, 1601–1611. doi:10.1002/art.40133
Woo, C. H., Kim, H. K., Jung, G. Y., Jung, Y. J., Lee, K. S., Yun, Y. E., et al. (2020). Small extracellular vesicles from human adipose-derived stem cells attenuate cartilage degeneration. J. Extracell. Vesicles 9, 1735249. doi:10.1080/20013078.2020.1735249
Wu, H., Peng, Z., Xu, Y., Sheng, Z., Liu, Y., Liao, Y., et al. (2022). Engineered adipose-derived stem cells with IGF-1-modified mRNA ameliorates osteoarthritis development. Stem Cell. Res. Ther. 13, 19–15. doi:10.1186/s13287-021-02695-x
Wu, J., Wu, J., Xiang, W., Gong, Y., Feng, D., Fang, S., et al. (2024a). Engineering exosomes derived from TNF-α preconditioned IPFP-MSCs enhance both yield and therapeutic efficacy for osteoarthritis. J. Nanobiotechnology 22, 555. doi:10.1186/s12951-024-02795-9
Wu, R., Xie, Y., Peng, Y., Wu, X., Ma, Y., Lyu, F. J., et al. (2024b). Young human plasma-derived extracellular vesicles rescue and reactivate IL-1β and TNF-α treated chondrocytes. Exp. Cell. Res. 437, 114009. doi:10.1016/j.yexcr.2024.114009
Wu, X., Li, H., Meng, F., Lui, T. H., and Pan, X. (2024c). iTRAQ proteomic analysis of exosomes derived from synovial fluid reveals disease patterns and potential biomarkers of Osteoarthritis. J. Orthop. Surg. Res. 19, 849. doi:10.1186/s13018-024-05336-0
Xia, Q., Wang, Q., Lin, F., and Wang, J. (2021). miR-125a-5p-abundant exosomes derived from mesenchymal stem cells suppress chondrocyte degeneration via targeting E2F2 in traumatic osteoarthritis. Bioengineered 12, 11225–11238. doi:10.1080/21655979.2021.1995580
Xu, C., Zhai, Z., Ying, H., Lu, L., Zhang, J., and Zeng, Y. (2022). Curcumin primed ADMSCs derived small extracellular vesicle exert enhanced protective effects on osteoarthritis by inhibiting oxidative stress and chondrocyte apoptosis. J. Nanobiotechnology 20, 123–16. doi:10.1186/s12951-022-01339-3
Xu, X., Liang, Y., Li, X., Ouyang, K., Wang, M., Cao, T., et al. (2021). Exosome-mediated delivery of kartogenin for chondrogenesis of synovial fluid-derived mesenchymal stem cells and cartilage regeneration. Biomaterials 269, 120539. doi:10.1016/j.biomaterials.2020.120539
Xu, Y., Ye, Z., Wang, Y., Ma, Y., Chen, X., Wang, S., et al. (2025). Alleviating osteoarthritis-induced damage through extracellular vesicles derived from inflammatory chondrocytes. Int. Immunopharmacol. 146, 113829. doi:10.1016/j.intimp.2024.113829
Yan, W., Li, Y., Xie, S., Tao, W. A., Hu, J., Liu, H., et al. (2024). Chondrocyte-targeted delivery system of Sortase A-engineered extracellular vesicles Silencing MMP13 for osteoarthritis therapy. Adv. Healthc. Mater 13, e2303510–e2303514. doi:10.1002/adhm.202303510
Yang, Y., Zhu, Z., Gao, R., Yuan, J., Zhang, J., Li, H., et al. (2021). Controlled release of MSC-derived small extracellular vesicles by an injectable Diels-Alder crosslinked hyaluronic acid/PEG hydrogel for osteoarthritis improvement. Acta Biomater. 128, 163–174. doi:10.1016/j.actbio.2021.04.003
Ye, P., Mi, Z., Wei, D., Gao, P., Ma, M., and Yang, H. (2022). MiR-3960 from mesenchymal stem cell-derived extracellular vesicles Inactivates SDC1/Wnt/β -catenin Axis to Relieve chondrocyte injury in osteoarthritis by targeting PHLDA2. Stem Cells Int. 2022, 1–18. doi:10.1155/2022/9455152
Yuan, Q., Yang, M., Zheng, H., Cai, Y., Luo, P., Wang, X., et al. (2024). M2 macrophage-derived extracellular vesicles encapsulated in hyaluronic acid alleviate osteoarthritis by modulating macrophage polarization. ACS Biomater. Sci. Eng. 10, 3355–3377. doi:10.1021/acsbiomaterials.3c01833
Zeng, Z., Dai, Y., Deng, S., Zou, S., Dou, T., and Wei, F. (2022). Synovial mesenchymal stem cell-derived extracellular vesicles alleviate chondrocyte damage during osteoarthritis through microRNA-130b-3p-mediated inhibition of the LRP12/AKT/β-catenin axis. Immunopharmacol. Immunotoxicol. 44, 247–260. doi:10.1080/08923973.2022.2038192
Zhang, B., Tian, X., Qu, Z., Hao, J., and Zhang, W. (2022). Hypoxia-Preconditioned extracellular vesicles from mesenchymal stem cells improve cartilage repair in osteoarthritis. Membr. (Basel) 12, 225–14. doi:10.3390/membranes12020225
Zhang, S., and Jin, Z. (2022). Bone mesenchymal stem cell-derived extracellular vesicles containing long noncoding RNA NEAT1 Relieve osteoarthritis. Oxid. Med. Cell. Longev. 2022, 5517648. doi:10.1155/2022/5517648
Zhang, X., Hsueh, M. F., Huebner, J. L., and Kraus, V. B. (2021). TNF-Α carried by plasma extracellular vesicles Predicts knee osteoarthritis progression. Front. Immunol. 12, 758386–10. doi:10.3389/fimmu.2021.758386
Zhang, X., Ma, S., Naz, S. I., Jain, V., Soderblom, E. J., Aliferis, C., et al. (2023). Comprehensive characterization of pathogenic synovial fluid extracellular vesicles from knee osteoarthritis. Clin. Immunol. 257, 109812. doi:10.1016/j.clim.2023.109812
Zhang, X., Ma, S., Naz, S. I., Soderblom, E., Jain, V., Aliferis, C., et al. (2024a). 165 plasma pathogenic extracellular vesicle subpopulations Predict progression of knee radiographic osteoarthritis. Int. J. Mol. Sci. 32, S124. doi:10.1016/j.joca.2024.02.176
Zhang, Y., Liu, K., Ma, X., Su, X., Zhao, L., Wu, Y., et al. (2024b). Therapeutic effects of Puerarin loaded bone marrow mesenchymal stem cell-derived exosomes in a rat model of osteoarthritis. Chem. Biodivers. 202402095 22, e202402095. doi:10.1002/cbdv.202402095
Zhongying, C., Xue, Z., Xiaofei, Z., Junbo, Z., Puwei, Y., and Yajun, S. (2024). Transcriptome sequencing-based study on the mechanism of action of Jintiange capsules in regulating synovial mesenchymal stem cells exosomal miRNA and articular chondrocytes mRNA for the treatment of osteoarthritis. J. Tradit. Chin. Med. 44, 1153–1167. doi:10.19852/j.cnki.jtcm.20240927.004
Zhou, H., Shen, X., Yan, C., Xiong, W., Ma, Z., Tan, Z., et al. (2022). Extracellular vesicles derived from human umbilical cord mesenchymal stem cells alleviate osteoarthritis of the knee in mice model by interacting with METTL3 to reduce m6A of NLRP3 in macrophage. Stem Cell. Res. Ther. 13, 322–21. doi:10.1186/s13287-022-03005-9
Zhou, R., Guo, J., and Jin, Z. (2024). Advancing osteoarthritis therapy with GMOCS hydrogel-loaded BMSCs-exos. J. Nanobiotechnology 22, 493–24. doi:10.1186/s12951-024-02713-z
Glossary
3′-UTR 3′untranslated region
ACLT Anterior Cruciate Ligament Transection
AD-MSCs Mesenchymal Stem Cells from Adipose Tissue
ADAMTS-4 DAM metallopeptidase with thrombospondin type 1 motif 4
ADAMTS-5 DAM metallopeptidase with thrombospondin type 1 motif 5
ALK3 Bone morphogenetic protein receptor type-1A
AM-MSCs Mesenchymal Stem Cells from Ammiotic Membrane
APOM Apolipoprotein M
ARRIVE Animal Research: Reporting of in vivo Experiments
AVV Adeno-Associated Virus
BM-MSCs Mesenchymal Stem Cells from Bone Marrow
BMP-2 Bone Morphogenetic Protein 2
BMPs Bone Morphogenetic Proteins
C3 General transcription factor 3C polypeptide 3
C4B C4b-binding protein β chain
CD10 Neprilysin
CD326 Epithelial cell adhesion molecule
CD45 Receptor-type tyrosine-protein phosphatase C
CD56 Neural cell adhesion molecule 1
CH Chondrocytes
CIOA Collagenase-Induced OA
circRNAs Circular RNAs
COL2A Collagen α-1(II) chain
CPEB1 Cytoplasmic polyadenylation element-binding protein 1
DM4 Glyceraldehyde-3-phosphate dehydrogenase
DMM Destabilization of the meniscus
DP-SCs Dental Pulp-derived Stem Cells
DPYSL2 Dihydropyrimidinase-related protein 2
E2F2 Transcription factor E2F2
ECM Extracellular Matrix
ENPP1 Ectonucleotide pyrophosphatase/phosphodiesterase family member 1
ERK Mitogen-activated protein kinase 4
ESCs Embryponic Stem Cells
EV Extracellular Vesicle
FGF-2 Fibroblast Growth Factor-2
Ha Hyaluronic Acid
HGF Hepatocyte Growth Factor
IGF-1 Insulin-like Growth Factor-1
IGF1R Insulin-like growth factor 1 receptor
IL-1 Interleukin-1
IL-10 Interleukin-10
IL-13 Interleukin-13
IL-17 Interleukin-17
IL-1β Interleukin-1β
IL-35 Interleukin-35
IL-4 Interleukin-4
iNOS Nitric oxide synthase, inducible
IPFP/IFP-MSCs Infrapatellar Fat Pad-derived Mesenchymal Stem Cells
iPSC-derived MSCs Mesenchymal Stem Cell-derived iPSCs
iPSCs induced Pluripotent Stem Cells
iTRAQ Isobaric Tag for Relative and Absolute Quantitation
JAK/STAT Tyrosine-protein kinase JAK/Signal transducer and transcription activator
JAK2/STAT3 Tyrosine-protein kinase JAK2/Signal transducer and transcription activator-3
KL Kellgren Lawrence
lncRNA long non-coding RNAs
MAPK MAP kinase-activated protein kinase 3
MDM2 E3 ubiquitin-protein ligase Mdm2
METTL3 N(6)-adenosine-methyltransferase catalytic subunit METTL3
MFG-E8 Lactadherin
MIA Monosodium Iodoacetate
miRNAs/miR microRNAs
MMP-13 collagenase-3
MMP-3 stromelysin-1
MMPs matrix metalloproteinases
MMx Resection of Medial Menisci
MSCs Mesenchumal Stem Cells
mTOR Serine/threonine-protein kinase mTOR
MYH9 Myosin-9
ncRNAs noncoding RNAs
NEAT1 Protein HEXIM1
NF-κB Nuclear factor NF-kappa-B
NLRP3 NACHT, LRR and PYD domains-containing protein 3
Nrf2 Nuclear factor erythroid 2-related factor 2
OA Osteoarthritis
OSM Oncostatin M
p53 Cellular tumor antigen p53
p75NTR p75 Neurotrophin Receptor
PESCs Progenitor Endothelial Stem Cells
PHLDA2 Pleckstrin homology-like domain family A member 2
PI3K/Akt Phosphatidylinositol 3-kinase catalytic subunit type 3/RAC-β serine/threonine-protein kinase
PKC Protein kinase C α type
PPD-MSCs Positively charged MSC-sEV
PRISMA Preferred Reporting Items for Systematic Reviews and Meta-Analyses
proNGF pro-β -nerve growth factor
RA Rheumatic Arthritis
S-MSCs Mesenchymal Stem Cells from Synovial membrane
S100A8 Protein S100-A8
sEV small Extracellular Vesicles
SF Synovial Fluid
SFRP5 Secreted Frizzled-Related Protein 5
SJ-cells Cells from Synovial Joint
SOD3 Extracellular superoxide dismutase [Cu-Zn]
SOX11/FOXO Transcription factor SOX-11/Forkhead box protein O
SOX9 Transcription factor SOX-9
STAT3 Signal transducer and transcription activator-3
T-MSCs Tonsil-derived Mesenchymal Stem Cells
TGF-β Transforming Growth Factor-β
TGF-β3 Transforming Growth Factor-β3
TIMP-3 Tissue Inhibitors of Metalloproteinases-3
TIMPs Tissue Inhibitors of Metalloproteinases
TMJOA Temporomandibular Joint Osteoarthritis
TNF-α Tumor Necrosis Factor-α
Trim14 Tripartite motif-containing protein 14
Trx80 Recombinant human Thioredoxin 80
TSG-6 Tumor necrosis factor-inducible gene 6 protein
UAC Anterior Crossbite Models
UC-MSC Mesenchymal Stem Cells from Umbilical Cord
VEGF Vascular Endothelial Growth Factor
Wnt Protein Wnt
YAP Transcriptional coactivator YAP1
εPL-PEG-DSPE ε-polylysine-polyethylene-distearyl phosphatidylethanolamine
Keywords: small extracelllular vesicles, osteoarthiritis, therapy, biomarker, personalized medicine
Citation: González-Rodríguez A, De Toro FJ, Jorge-Mora A, Fernandez-Pernas P, Rivadulla CP, Fraga M, Fafián-Labora JA and Arufe MC (2025) Targeting osteoarthritis with small extracellular vesicle therapy: potential and perspectives. Front. Bioeng. Biotechnol. 13:1570526. doi: 10.3389/fbioe.2025.1570526
Received: 05 February 2025; Accepted: 30 May 2025;
Published: 20 June 2025.
Edited by:
Serena Duchi, The University of Melbourne @ St. Vincent’s Hospital, AustraliaReviewed by:
Magali Cucchiarini, Saarland University Medical Center, GermanyAnna Trengove, The University of Melbourne, Australia
Copyright © 2025 González-Rodríguez, De Toro, Jorge-Mora, Fernandez-Pernas, Rivadulla, Fraga, Fafián-Labora and Arufe. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: María C. Arufe, bWFyaWEuYXJ1ZmVAdWRjLmVz; Juan A. Fafián-Labora, anVhbi5sYWJvcmFAdWRjLmVz